
Submitted:
03 May 2024
Posted:
07 May 2024
You are already at the latest version
Abstract
1) Background: carbapenem-resistant Acinetobacter baumannii (CRAB) is an opportunistic Gram-negative pathogen that has a significant role in healthcare-associated infections. Unlike several studies on the antibiotic-resistant epidemiology of A. baumanni, virulence molecular epidemiology was less studied. This study aimed to investigate CRAB virulence genes and their ability to form biofilms, and to correlate their biofilm formation ability with both; biofilm-encoding virulence genes and carbapenemase-encoding resistance genes. 2) Methods: 107 CRAB clinical isolates were collected from two hospitals in Jordan between 2018 and 2019 and were screened for virulence genes using PCR. In addition, biofilm formation ability was assessed using the microtiter plate method. 3) Results: the frequencies of the bap, OmpA, surA, PLD, paaE, basD, and traT virulence genes were 99.10%, 98.20%, 98.20%, 95.50%, 89.10%, 86.40%, and 8.20%, respectively. Overall, 86.4% of the tested isolates were biofilm formers with varying degrees; weak (28.2%), moderate (36.4%) and strong (21.8%). A significant relationship was found between the carbapenemase-encoding gene (OXA-23 gene) and biofilm production. 4) Conclusion: to the best of our knowledge, this is the first study in Jordan that inspected CRAB virulence genes and highlighted the importance of improving infection control measures to avoid CRAB outbreaks.
Keywords:
Acinetobacter
; carbapenem
; virulence
; PCR
; biofilm
; gene
; microtiter
; resistance
Introduction
Acinetobacter baumannii is ubiquitous bacteria that is frequently recovered from soil, water, animals, and humans [1]. They are part of the normal flora found on the skin [2], mucosal membranes, and the respiratory tract [3,4]. In addition, these opportunistic bacteria inhabit hospitals’ environment and are recognized as the leading cause of healthcare associated infections [5,6]. A. baumannii is a Gram-negative, non-lactose fermenter, aerobic, non-fastidious, non-motile, oxidase-negative, indole-negative, citrate-positive, and catalase-positive bacterium [7].
Biofilm formation, resistance to a wide spectrum of antibiotics and other virulence factors enable A. baumannii to resist desiccation and disinfectants and to colonize biotic and abiotic surfaces, and thus explain its significance as a nosocomial pathogen [8,9]. A. baumannii may contaminate medical tools and equipment's, such as ventilators, arterial pressure monitors, humidifiers, washbasins, respirometers and dialysis machines and thus are considered as reservoirs for hospital acquired pneumonia, urinary tract infections, bacteremia, wound infection and meningitis [10].
Acinetobacter baumannii pathogenesis is associated with numerous virulence factors, encompassing outer membrane proteins, lipopolysaccharide, capsule, phospholipase, nutrient-acquisition systems, efflux pumps, protein secretion systems, quorum sensing, and biofilm formation. Altogether these virulence factors play significant roles during bacterial pathogenesis stages, including transmission, adhesion, colonization, invasion and evasion of host’s defenses [11]. Table 1 describes important A. baumannii virulence factors and their roles in pathogenesis.
Acinetobacter baumannii was originally extrinsically sensitive to all antibiotics until the early 1970s [18]. However, the extrinsic resistance of A. baumannii was increased to include multiple antibiotics with alarming rate, especially during 1980s and 1990s [19]. The emergence of carbapenem-resistant A. baumannii (CRAB) is associated with high rates of morbidity, mortality, and nosocomial outbreaks worldwide [20]. The main mechanism of carbapenem resistance is the production of beta-lactamase enzymes belonging to oxacillinases, metallo-β-lactamases (MBLs), or Amber class A carbapenemases that can be intrinsic or acquired [19]. Several different classes of carbapenemases are released by CRAB, including class D beta-lactamases (OXA-23, OXA-51, and OXA-53 enzymes), and class B metallo-beta lactamases (IMP and VIM enzymes) [21]. Limited treatment options for CRAB infections poses a significant challenge in providing effective therapy to affected individuals [17].
Acinetobacter baumannii was categorized as a member of ESKAPE pathogens family, which consist of six nosocomial antibiotic resistant bacteria; Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter species [22]. The World Health Organization (WHO), and Centers for Disease Control and Prevention (CDC) have identified CRAB as a critical pathogen requiring extensive research, and drug development [8,23].
CRAB is rapidly increasing globally with a significant rise in morbidity and mortality rates among hospitalized patients. CRAB is endemic in the Mediterranean countries with resistance to many antibiotic family types [24], for example, in Jordan, A. baumannii isolates exhibit significant resistance to cephalosporins, penicillins, imipenem, and quinolones [25,26].
In Jordan, several projects extensively studied the clinical epidemiology and antibiotic resistance patterns for A. baumannii, but on the other hand no virulence epidemiology was investigated. This study inspected the prevalence of virulence genes in CRAB and its ability to form biofilms to understand its pathogenic potential, improve infection control measures, and disease management.
Materials and Methods
Sample Collection
In the present study, a total of 110 clinical isolates of carbapenemase-producing A. baumannii were provided by the faculty of medicine at Hashemite University. These isolates had been previously collected from Prince Hamzah Hospital and the Islamic Hospital from January 2018 to December 2019 and were stored in deep freeze at -70° C.
Demographic Data Recruitment
Patient’s data were collected from the archive department using patients’ identification codes.
Characteristics of Isolates
These clinical isolates were published previously by Al-Tamimi et al [25]. They were obtained from various sample sources, including sputum, wounds, blood, urine, cerebrospinal fluid and bronchial lavage. The main collection department was the intensive care unit (ICU). They were identified by routine bacteriological methods and were confirmed by polymerase chain reaction (PCR). They exhibited high levels of resistance to a broad range of antibiotics except for colistin.
Bacterial Reactivation
All CRAB isolates were reactivated by culturing the preserved cells into nutrient broth media (Himedia, India) and incubating for 18-24 hours at 37°C. Then, dense bacterial suspension was centrifuged for 10 minutes at 5000g, the supernatant was removed, and the bacterial pellet was sub-cultured on MacConkey agar. Pure colonies on MacConkey agar were observed next day and were introduced for biofilm production test and DNA extraction.
Detection of Biofilm Production
Biofilm production test was performed according to microtiter plate (MTP) method that was published previously [10], with minor modifications. In this protocol, bacterial colonies were transferred into 10ml of tryptic soya broth (TSB) and the tube turbidity was adjusted with a final concentration of 106 CFU/ml using turbidimeter. Aliquots of 200µL fresh bacterial suspensions were then dispensed into a 96-well flat-bottom polystyrene microtiter plate, and then incubated at 37°C for 24 hours. After that, the contents of wells were discarded and were washed three times with 200μl of sterile phosphate buffered saline (PBS) and then followed by adding 200μl of 99% methanol for 15 minutes to ensure fixation of any adherent bacteria, after that the wells were emptied, and dried. Afterward, the plates were stained with (1%) Hucker crystal violet for 20 minutes. The excess stain was washed out with running tap water.
The plates were then air-dried, and 160μl of 33% (v/v) glacial acetic acid was added to each well to resolubilize the dye bound to the adhered bacterial cells. The mean optical density at 570 nm of the non-biofilm producer A. baumannii was used as the OD cut-off value (ODc) with an automated Biotic Synergy HTX Multi-Mode Reader [27]. The quantitative biofilm assay was conducted in triplicate for all isolates. A sterile TSB was utilized as a negative control. The OD results of all tests were divided into four groups: (1) OD ≤ ODc : non biofilm producer; (2) ODc < OD ≤ 2× ODc : weak biofilm producer; (3) 2× ODc < OD ≤ 4 × ODc : moderate biofilm producer; and (4) 4× ODc < OD = strong biofilm producer [10].
Polymerase Chain Reaction for Seven Virulence Genes
DNA Extraction
The DNA was extracted from CRAB broth preparations using a commercial extraction DNeasy tissue kit (Qiagen, Holden, Germany) according to manufacturer instructions and the final DNA yield (at least 50ng/µl) for every sample was introduced for PCR.
Primers
All primers for the seven virulence genes (bap, surA1, basD, PLD, OmpA, paaE, and traT) were published previously (Table 2). They were ordered commercially (Macrogen Inc., Rockville, MD) and were purchased in a lyophilized form and were diluted with the desired volume of nuclease-free water to prepare a final concentration of 10 pmol/µL for working primer according to the manufacturer’s instructions.
PCR Assay
The optimized PCR mixture for every gene contained 2.0 μL of extracted DNA in 25 μL final reaction volume consisting of 12.5μL 2x one Taq quick load PCR mix containing reaction buffer, comprising 5 mM dNTPs, 15mM MgCl2, stabilizers, enhancers, and DNA polymerase (New England Biolabs/USA). In addition to 1.0μL of 1.0μM of each forward and reverse primers, and 8.5μL nuclease free PCR water (Avantor/USA). Amplification was performed with PCR thermocycler (Qiagen, Germany) using standard settings. The optimized thermal profile was initial denaturation at 94ºC for 5 minutes, followed by 30 cycles of denaturation step at 94ºC for 45 seconds, annealing step for 15 seconds at different temperatures according to every gene (Table 1) and extension step at 72 ºC for 1 minute followed by final extension step at 72 ºC for 10 minutes. No-template (PCR water) and positive controls were included in each run to rule out amplification failure or possible contamination.
For all seven virulence genes, internal positive controls were applied. Moreover, virulence genes were amplified by PCR using primer set in this study. After that PCR products were sent for purification and sequencing via commercial service (Macrogen, Korea). The results of Sanger sequencing were received, and the raw sequences, when required, were trimmed by removing the unwanted noise nucleotides using bioedit version 7 software. The purified sequences were submitted to Pubmed website and were aligned by blastn feature and finally confirmed the gene identity, data not shown.
Agarose gel Electrophoresis
Using 1.5 % agarose gel loaded with ethidium bromide stain, PCR products along with 100-base-pairs standard DNA ladder (New England Biolabs/USA) electrophoresis was performed and the separated amplicons were visualized by UV-based gel documentation system (Alpha Innotech/USA).
Statistical Analysis
Descriptive and analytical analysis was done using Statistical Package for Social Science (SPSS) version 25 software. The correlation between variables analyzed by the Pearson Chi-Square test. p value ≤ 0.05 was considered statically significant.
Results
Demographic Data
In this study, CRAB isolates were collected from two major hospitals in Jordan. Out of 110 patients, 64% were males and the mean age of the participents was 59.7 years (SD±21.6). The highest precentage of CRAB clinical isolates was observed in patients aged above 60 years (57%), Figure 1A. The study encompassed various departments, with most cases observed in the intensive care unit (ICU) at 91%, Figure 1B. Various types of samples were collected for analysis, with the highest percentage being sputum samples (69%), Figure 1C.
Distribution of Virulence Genes among CRAB Isolates
In this study, the frequencies of the inspected virulence genes in CRAB isolates ranged from 8.2% for traT gene to 99.1% for bap gene. Figure 2. All genes were amplified successfully for positive isolates.
Biofilm Detection by Microtiter Plate Method (MTP) in CRAB
Statistical Associations
After the investigation of the relationship between biofilm production, and biofilm-related genes, the results showed that Bap, and OmpA, genes were found in 98.9% (94/95), and 97.9% (93/95) of the biofilm producers, respectively with insignificant associations. On the other hand, the results showed that VIM gene was found in 28.4% (27out of 95) of the biofilm producers with no positive association, whereas OXA-23 gene was found in 100% of the biofilm producers with a positive association (p value <0.05) as shown in Table 4.
Discussion
A. baumannii is an opportunistic pathogen that thrives in hospital environments, resisting desiccation, and surviving on inanimate surfaces for years [27]. Multi-drug-resistant strains of this pathogen are mostly treated with carbapenems. Recently, carbapenemase-producing A. baumannii (CRAB) is increasing at an alarming rate causing global medical challenges [28], posing a significant threat to public health and leaving limited options for treatment, which in turn leads to significantly higher morbidity and mortality rates particularly in immune-compromised patients [29]. CRAB outbreaks have been reported frequently in occupied ICU units [24,30,31].
In recent years A. baumannii research in Jordan has focused on its phenotypic and genotypic resistance profiles, the mechanism of drug resistance and clinical epidemiology with little attention on the mechanism of virulence and pathogenesis [25,32,33]. In this research, molecular and conventional assays were conducted to inspect the virulence genes harbored in CRAB strains isolated from clinical samples in two hospitals in Jordan.
This study involved 110 CRAB isolates, most samples were collected mainly from the ICU accounting for 91% among other hospitals departments, which concurs with previous literature [34,35]. The high prevalence of CRAB in ICU may be attributed to the increased exposure to several risk factors that increase the chance of acquiring the infection, such as immunocompromisation, using catheters, mechanical ventilators, ventriculoperitoneal shunts, and central lines, in addition to the widespread administration of antibiotics of variant activity spectrum [9].
Since pneumonia and other pulmonary infections and complications are commonly seen in the ICU department that enforce toward applying mechanical ventilators and other respiratory procedures, which in turn increases the risk of acquiring the infection [28], hence more than two-thirds (69%) of the isolates in this study were collected from respiratory samples, which agrees with previous studies [36,37].
The proportion of CRAB in males was higher than in females, which might be related to the behavior, lifestyle, and the increased likelihood of male for developing chronic obstructive pulmonary diseases (COPD) that increase the chance of CRAB infection [38]. In terms of age groups, children under 18 years have a relatively low risk of infection, accounting for only 2%, which might be related to their less exposure to infection predisposing factors than adults [39]. Same CRAB frequencies were reported in both genders and age groups in previous literatures [33,40].
In this study, all of the virulence genes were reported in high percentages ranging from 86.4% to 99.1%, except for traT gene (8.2%). Many studies have reported either similar or less percentages of the same virulence genes which is attributed to their endemicity, as well as prevalence variation worldwide [41], Table 5.
In the current study, the presence of high frequency of Bap and OmpA genes predicts bacterial biofilm formation ability and thus expecting prolong strain persistence in the hospital and resistance to a wide range of antibiotics [41].
Biofilm formation among CRAB isolates was determined in vitro using the microtiter plate assay. Biofilm formation was observed in 86.4% of the isolates, which concurs with the fact that A. baumannii is a significant biofilm former, where biofilms enable prolonged survival in harsh environments and resistance to antibiotics [27]. The high prevalence of biofilm formation aligns with other studies published worldwide [27,44,45,46].
In this study, out of 110 tested CRAB isolates, 86.4% (n=95) were biofilm producers, among which 31 (32.6%) were week biofilm producers, 40 (42.1%) were moderate biofilm producers, and 24 (25.3%) were strong biofilm producers. Similar results were reported previously [47], but still disagreement present with other study that reported different percentages [48]. This variation might be attributed with the widespread prevalence of biofilm-encoding genes among A. baumannii strains, variations of the biofilm-formation assay principles, or environmental factors [49]. Abdi-Ali et al. [50] has applied test tube and microtiter plate methods for evaluating biofilm formation. The findings of the microtiter plate method were as follows: 25% negative, 41% weak, 10% moderate, and 18% strong. The test tube method resulted in the following results: 18% negative, 42% weak, 18% moderate, and 22% strong. In addition, it is crucial to emphasize that the method of assessing biofilm production in vitro may not adequately reflect the complexity of biofilm formation in clinical settings [49]. Finally, all of CRAB in this study were isolated from clinical samples; it was reported that clinical CRAB isolates exhibited a higher ability to form strong biofilm than environmental isolates [51].
In this study, the correlation between the formation of biofilm and its corresponding genes (bap and OmpA) was investigated and reported (97.9%, and 98.9%, respectively). No significant correlation reported when comparing biofilm-positive and biofilm-negative CRAB with the presence of bap and OmpA genes (p value > 0.05), [10]. This insignificant correlation might be attributed to the complexity of the biofilm formation process, which is regulated by a variety of genetic and environmental factors. Therefore, further research is required to investigate the role of other genes, regulatory mechanisms, and environmental variables in CRAB biofilm development [10,49].
On the other hand, the relationship between the presence of biofilm formation and carbapenem resistance genes (VIM and OXA-23) was studied and reported both insignificant and significant correlations of VIM and OXA-23, respectively, (p value< 0.05), which was consistent with an earlier study in Turkey [52]. However, the type of statistical associations between biofilm formation and antibiotic resistance remains controversial [53]. The type of resistance determinants harbored by A. baumannii can influence its ability to form biofilm. Moreover, biofilm formation is more strongly associated with CRAB strains than with the susceptible strains [53].
Conclusion
Our data showed a high prevalence of six virulence genes except the traT gene, which reflect their virulence potential. Virulence genes are correlated with various mechanisms that help CRAB to adapt and thrive in different environments. Our results also showed high prevalence of biofilm production among CRAB isolates, which may have contributed to their hospitals persistence and treatment resistance and thus being a challenge to control and manage infections.
Author’s Contributions
Conceptualization, A.M.Z.; Methodology, D.A.B.; Software, D.A.B.; Validation, A.M.Z, H.M.S, R.B.S, and H.A.B.; Formal Analysis, A.M.Z and D.A.B; Investigation, D.A.B.; Resources, M.A.T.; Data Curation, D.A.B.; Writing Original Draft Preparation, A.M.Z.; Writing – Review & Editing, H.M.S, R.B.S and T.M.; Visualization, D.S.H.; Supervision, A.M.Z.; Project Administration, A.M.Z.; Funding Acquisition, A.M.Z and R.B.S.
Funding
This research was funded by the Hashemite university, Zarqa, Jordan under full research grant scheme number 50/2022; and Princess Nourah bint Abdulrahman, Riyadh, Saudi Arabia under Grant University Researchers Supporting Project number (PNURSP2024R304).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the ethical committee of the Hashemite University (P. O box 330127/13133) Institutional Review Board (IRB) under the number (No.16/3/2022/2023) on 23/1/2022.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author due to privacy reasons.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baumann, P. Isolation of Acinetobacter from Soil and Water. J. Bacteriol. 1968, 96, 39–42. [CrossRef]
- Towner A, Dworkin M, Falkow S, Rosenberg E, Schleifer K, Stackebrandt E. The Prokaryotes. Springer; 2006.
- Munoz-Price LS, Weinstein RA. Acinetobacter infection. New England Journal of Medicine. 2008;358(12):1271-81.
- Seifert, H.; Dijkshoorn, L.; Gerner-Smidt, P.; Pelzer, N.; Tjernberg, I.; Vaneechoutte, M. Distribution of Acinetobacter species on human skin: comparison of phenotypic and genotypic identification methods. J. Clin. Microbiol. 1997, 35, 2819–2825. [CrossRef]
- Lee, C.-R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.-J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, Antibiotic Resistance Mechanisms, and Prospective Treatment Options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [CrossRef]
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [CrossRef]
- Jung, J.; Park, W. Acinetobacter species as model microorganisms in environmental microbiology: current state and perspectives. Appl. Microbiol. Biotechnol. 2015, 99, 2533–2548. [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [CrossRef]
- Kyriakidis, I.; Vasileiou, E.; Pana, Z.D.; Tragiannidis, A. Acinetobacter baumannii Antibiotic Resistance Mechanisms. Pathogens 2021, 10, 373. [CrossRef]
- Thummeepak R, Kongthai P, Leungtongkam U, Sitthisak SJIM. Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. International microbiology. 2016;19(2):121-9.
- Dehbanipour, R.; Ghalavand, Z. Acinetobacter baumannii : Pathogenesis, virulence factors, novel therapeutic options and mechanisms of resistance to antimicrobial agents with emphasis on tigecycline. J. Clin. Pharm. Ther. 2022, 47, 1875–1884. [CrossRef]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii Biofilm-Associated Protein Plays a Role in Adherence to Human Epithelial Cells. Infect. Immun. 2012, 80, 228–233. [CrossRef]
- Loehfelm, T.W.; Luke, N.R.; Campagnari, A.A. Identification and Characterization of an Acinetobacter baumannii Biofilm-Associated Protein. J. Bacteriol. 2008, 190, 1036–1044. [CrossRef]
- Conde-Pérez, K.; Vázquez-Ucha, J.C.; Álvarez-Fraga, L.; Ageitos, L.; Rumbo-Feal, S.; Martínez-Guitián, M.; Trigo-Tasende, N.; Rodríguez, J.; Bou, G.; Jiménez, C.; et al. In-Depth Analysis of the Role of the Acinetobactin Cluster in the Virulence of Acinetobacter baumannii. Front. Microbiol. 2021, 12. [CrossRef]
- Depka, D.; Bogiel, T.; Rzepka, M.; Gospodarek-Komkowska, E. The Prevalence of Virulence Factor Genes among Carbapenem-Non-Susceptible Acinetobacter baumannii Clinical Strains and Their Usefulness as Potential Molecular Biomarkers of Infection. Diagnostics 2023, 13, 1036. [CrossRef]
- Mozafari, H.; Mirkalantari, S.; Kalani, B.S.; Amirmozafari, N. Prevalence Determination of Virulence Related and Biofilm Formation Genes in Acinetobacter baumannii Isolates from Clinical Respiratory Samples in Imam Khomeini Hospital, Tehran, Iran in 2018. Iran. J. Med Microbiol. 2021, 15, 266–280. [CrossRef]
- Liu, C.; Chang, Y.; Xu, Y.; Luo, Y.; Wu, L.; Mei, Z.; Li, S.; Wang, R.; Jia, X. Distribution of virulence-associated genes and antimicrobial susceptibility in clinical Acinetobacter baumannii isolates. Oncotarget 2018, 9, 21663–21673. [CrossRef]
- Howard A, O’Donoghue M, Feeney A, Sleator RD. Acinetobacter baumannii: an emerging opportunistic pathogen. Virulence. 2012;3(3):243-50.
- Doi, Y.; Murray, G.L.; Peleg, A.Y. Acinetobacter baumannii: Evolution of Antimicrobial Resistance-treatment Options. Semin. Respir. Crit. Care Med. 2015, 36, 85–98. [CrossRef]
- Joly-Guillou, M.-L. Clinical impact and pathogenicity of Acinetobacter. Clin. Microbiol. Infect. 2005, 11, 868–873. [CrossRef]
- Poirel L, Naas T, Nordmann P. Diversity, epidemiology, and genetics of class D β-lactamases. Antimicrobial agents and chemotherapy. 2010;54(1):24-38.
- Morris, S.; Cerceo, E. Trends, Epidemiology, and Management of Multi-Drug Resistant Gram-Negative Bacterial Infections in the Hospitalized Setting. Antibiotics 2020, 9, 196. [CrossRef]
- Abadi, A.T.B.; Rizvanov, A.A.; Haertle, T.; Blatt, N.L. World Health Organization Report: Current Crisis of Antibiotic Resistance. BioNanoScience 2019, 9, 778–788. [CrossRef]
- Medioli F, Bacca E, Faltoni M, Burastero GJ, Volpi S, Menozzi M, et al. Is It Possible to Eradicate Carbapenem-Resistant Acinetobacter baumannii (CRAB) from Endemic Hospitals? Antibiotics. 2022;11(8):1015.
- Al-Tamimi, M.; Albalawi, H.; Alkhawaldeh, M.; Alazzam, A.; Ramadan, H.; Talalwih, M.; Alma’aitah, A.; Al Balawi, D.; Shalabi, S.; Abu-Raideh, J.; et al. Multidrug-Resistant Acinetobacter baumannii in Jordan. Microorganisms 2022, 10, 849. [CrossRef]
- Batarseh, A.; Al-Sarhan, A.; Maayteh, M.; Al-Khatirei, S.; Alarmouti, M. Antibiogram of multidrug resistant Acinetobacter baumannii isolated from clinical specimens at King Hussein Medical Centre, Jordan: a retrospective analysis.. East. Mediterr. Heal. J. 2015, 21, 828–834. [CrossRef]
- Smitran A, Lukovic B, Bozic L, Jelic D, Jovicevic M, Kabic J, et al. Carbapenem-Resistant Acinetobacter baumannii: Biofilm-Associated Genes, Biofilm-Eradication Potential of Disinfectants, and Biofilm-Inhibitory Effects of Selenium Nanoparticles. Microorganisms. 2023;11(1).
- Nguyen M, Joshi SG. Carbapenem resistance in Acinetobacter baumannii, and their importance in hospital-acquired infections: a scientific review. Journal of applied microbiology. 2021 Dec 1;131(6):2715-38.
- Harding, C.M.; Hennon, S.W.; Feldman, M.F. Uncovering the mechanisms of Acinetobacter baumannii virulence. Nat. Rev. Microbiol. 2018, 16, 91–102. [CrossRef]
- Obenhuber, T.; Scheier, T.; Stutz, T.; Hug, M.; Fontein, D.; Kaiser, A.; Schoene, S.; Steiger, P.; Brugger, S.; Zingg, W.; et al. An outbreak of multi-drug-resistant Acinetobacter baumannii on a burns ICU and its control with multi-faceted containment measures. J. Hosp. Infect. 2024, 146, 102–108. [CrossRef]
- Raddaoui A, Mabrouk A, Chebbi Y, Frigui S, Al-Gallas N, Abbassi MS, et al. Outbreak caused by pandrug-resistant blaOXA-69/blaOXA-23/blaGES harboring Acinetobacter baumannii ST2 in an intensive care unit. Acta Microbiologica et Immunologica Hungarica. 2024.
- Al-Sheboul, S.A.; Al-Moghrabi, S.Z.; Shboul, Y.; Atawneh, F.; Sharie, A.H.; Nimri, L.F. Molecular Characterization of Carbapenem-Resistant Acinetobacter baumannii Isolated from Intensive Care Unit Patients in Jordanian Hospitals. Antibiotics 2022, 11, 835. [CrossRef]
- Gharaibeh, M.H.; Abandeh, Y.M.; Elnasser, Z.A.; Lafi, S.Q.; Obeidat, H.M.; Khanfar, M.A. Multi-drug Resistant Acinetobacter baumannii: Phenotypic and Genotypic Resistance Profiles and the Associated Risk Factors in Teaching Hospital in Jordan. J. Infect. Public Heal. 2024, 17, 543–550. [CrossRef]
- Ibrahim, S.; Al-Saryi, N.; Al-Kadmy, I.M.S.; Aziz, S.N. Multidrug-resistant Acinetobacter baumannii as an emerging concern in hospitals. Mol. Biol. Rep. 2021, 48, 6987–6998. [CrossRef]
- Zhang, X.; Li, F.; Awan, F.; Jiang, H.; Zeng, Z.; Lv, W. Molecular Epidemiology and Clone Transmission of Carbapenem-Resistant Acinetobacter baumannii in ICU Rooms. Front. Cell. Infect. Microbiol. 2021, 11. [CrossRef]
- Bostanghadiri, N.; Narimisa, N.; Mirshekar, M.; Dadgar-Zankbar, L.; Taki, E.; Navidifar, T.; Darban-Sarokhalil, D. Prevalence of colistin resistance in clinical isolates of Acinetobacter baumannii: a systematic review and meta-analysis. Antimicrob. Resist. Infect. Control. 2024, 13, 1–17. [CrossRef]
- Karakonstantis, S.; Rousaki, M.; Vassilopoulou, L.; Kritsotakis, E.I. Global prevalence of cefiderocol non-susceptibility in Enterobacterales, Pseudomonas aeruginosa, Acinetobacter baumannii, and Stenotrophomonas maltophilia: a systematic review and meta-analysis. Clin. Microbiol. Infect. 2024, 30, 178–188. [CrossRef]
- Adeloye, D.; Song, P.; Zhu, Y.; Campbell, H.; Sheikh, A.; Rudan, I. Global, regional, and national prevalence of, and risk factors for, chronic obstructive pulmonary disease (COPD) in 2019: a systematic review and modelling analysis. Lancet Respir. Med. 2022, 10, 447–458. [CrossRef]
- Wang, M.; Ge, L.; Chen, L.; Komarow, L.; Hanson, B.; Reyes, J.; Cober, E.; Alenazi, T.; Zong, Z.; Xie, Q.; et al. Clinical Outcomes and Bacterial Characteristics of Carbapenem-resistant Acinetobacter baumannii Among Patients From Different Global Regions. Clin. Infect. Dis. 2023, 78, 248–258. [CrossRef]
- Abarca-Coloma, L.; Puga-Tejada, M.; Nuñez-Quezada, T.; Gómez-Cruz, O.; Mawyin-Muñoz, C.; Barungi, S.; Perán, M. Risk Factors Associated with Mortality in Acinetobacter baumannii Infections: Results of a Prospective Cohort Study in a Tertiary Public Hospital in Guayaquil, Ecuador. Antibiotics 2024, 13, 213. [CrossRef]
- Park SM, Suh JW, Ju YK, Kim JY, Kim SB, Sohn JW, et al. Molecular and virulence characteristics of carbapenem-resistant Acinetobacter baumannii isolates: a prospective cohort study. Scientific Reports. 2023;13(1):19536.
- Sung, J.Y.; Koo, S.H.; Kim, S.; Kwon, G.C. Persistence of Multidrug-Resistant Acinetobacter baumannii Isolates Harboring blaOXA-23 and bap for 5 Years. J. Microbiol. Biotechnol. 2016, 26, 1481–1489. [CrossRef]
- Fallah, A.; Rezaee, M.A.; Hasani, A.; Barhaghi, M.H.S.; Kafil, H.S. Frequency of bap and cpaA virulence genes in drug resistant clinical isolates of Acinetobacter baumannii and their role in biofilm formation. Iranian journal of basic medical sciences 2017, 20, 849–855. [CrossRef]
- Altınok Ö, Boral B, Ergin MU, Eser ÖZ. Existence of biofilm and biofilm-associated virulence genes in multi-drug resistant invasive acinetobacter baumannii isolates Çok İlaca dirençli İnvaziv acinetobacter baumannii İzolatlarında biyofilm ve biyofilm İlişkili virülans genlerinin varlığı. Mikrobiyoloji Bulteni. 2020;54(1).
- Khalil, M.A.F.; Ahmed, F.A.; Elkhateeb, A.F.; Mahmoud, E.E.; Ahmed, M.I.; Ahmed, R.I.; Hosni, A.; Alghamdi, S.; Kabrah, A.; Dablool, A.S.; et al. Virulence Characteristics of Biofilm-Forming Acinetobacter baumannii in Clinical Isolates Using a Galleria mellonella Model. Microorganisms 2021, 9, 2365. [CrossRef]
- Santajit, S.; Bhoopong, P.; Kong-Ngoen, T.; Tunyong, W.; Horpet, D.; Paehoh-Ele, W.; Zahedeng, T.; Pumirat, P.; Sookrung, N.; Hinthong, W.; et al. Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand. Antibiotics 2023, 12, 580. [CrossRef]
- Hazhirkamal, M.; Zarei, O.; Movahedi, M.; Karami, P.; Shokoohizadeh, L.; Taheri, M. Molecular typing, biofilm production, and detection of carbapenemase genes in multidrug-resistant Acinetobacter baumannii isolated from different infection sites using ERIC-PCR in Hamadan, west of Iran. BMC Pharmacol. Toxicol. 2021, 22, 1–7. [CrossRef]
- Khoshnood S, Savari M, Abbasi Montazeri E, Farajzadeh Sheikh A. Survey on Genetic Diversity, Biofilm Formation, and Detection of Colistin Resistance Genes in Clinical Isolates of Acinetobacter baumannii. Infection and Drug Resistance. 2020;13:1547-58.
- Longo, F.; Vuotto, C.; Donelli, G. Biofilm formation in Acinetobacter baumannii. New Microbiology 2014, 37, 119–27.
- Abdi-Ali, A.; Hendiani, S.; Mohammadi, P.; Gharavi, S. Assessment of Biofilm Formation and Resistance to Imipenem and Ciprofloxacin among Clinical isolates of Acinetobacter baumannii in Tehran. Jundishapur J. Microbiol. 2014, 7, e8606. [CrossRef]
- Bardbari, A.M.; Arabestani, M.R.; Karami, M.; Keramat, F.; Alikhani, M.Y.; Bagheri, K.P. Correlation between ability of biofilm formation with their responsible genes and MDR patterns in clinical and environmental Acinetobacter baumannii isolates. Microb. Pathog. 2017, 108, 122–128. [CrossRef]
- zkul C, Hazırolan G. Oxacillinase gene distribution, antibiotic resistance, and their correlation with biofilm formation in Acinetobacter baumannii bloodstream isolates. Microbial Drug Resistance. 2021;27(5):637-46.
- Eze, E.C.; Chenia, H.Y.; El Zowalaty, M. Acinetobacter baumannii biofilms: effects of physicochemical factors, virulence, antibiotic resistance determinants, gene regulation, and future antimicrobial treatments. Infect. Drug Resist. 2018, ume 11, 2277–2299. [CrossRef]
Figure 1.
Distribution of a: age interval, b: department source and c: sample source among 110 CRAB isolates. CAN: central admission nursery, MCU: medical care unit, D.F: diabetic foot, U.D: urethral discharge, B.L: bronchial lavage.
Figure 1.
Distribution of a: age interval, b: department source and c: sample source among 110 CRAB isolates. CAN: central admission nursery, MCU: medical care unit, D.F: diabetic foot, U.D: urethral discharge, B.L: bronchial lavage.
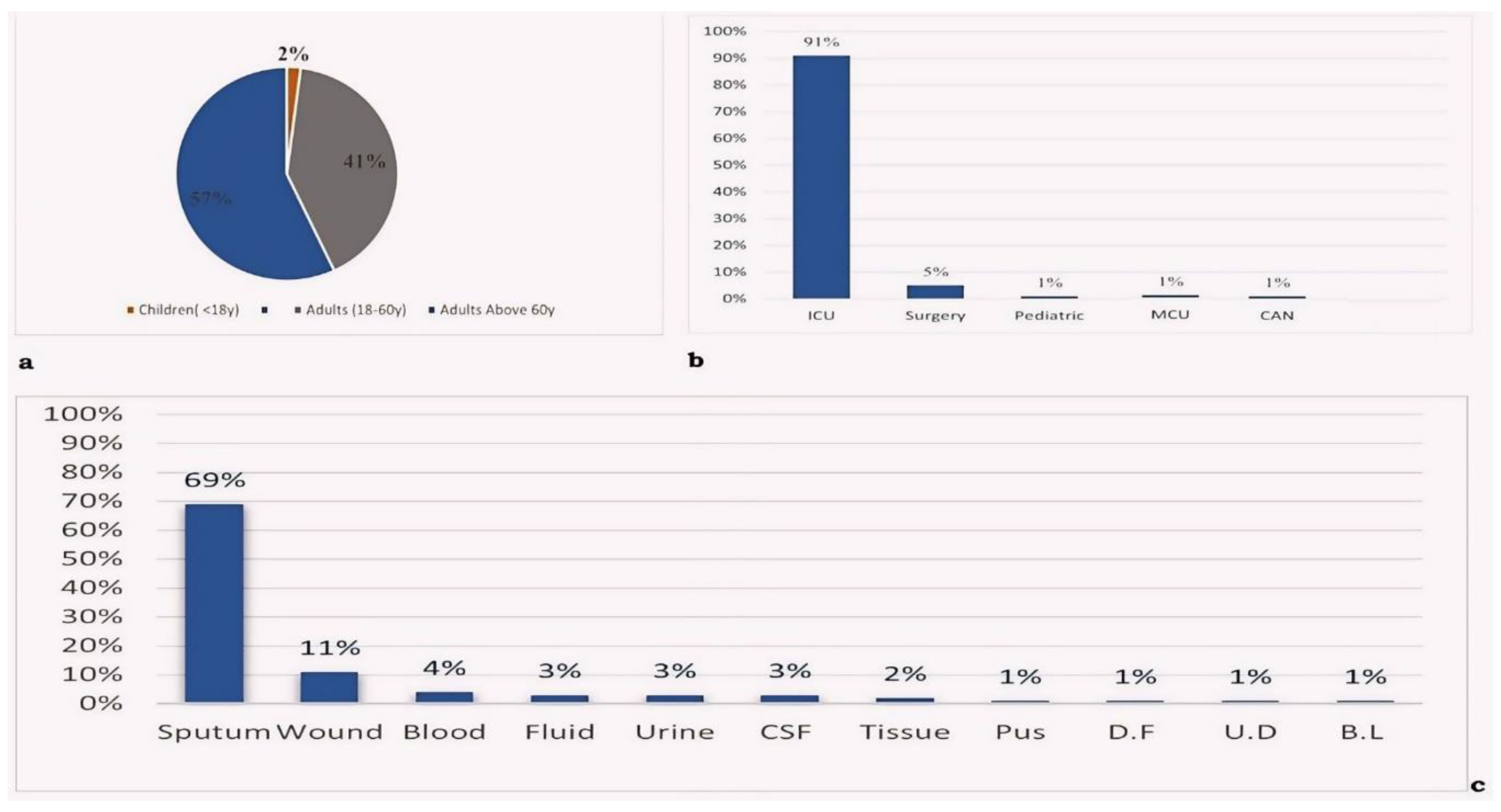
Figure 2.
Percentages of the virulence genes in A.baumannii clinical isolates.
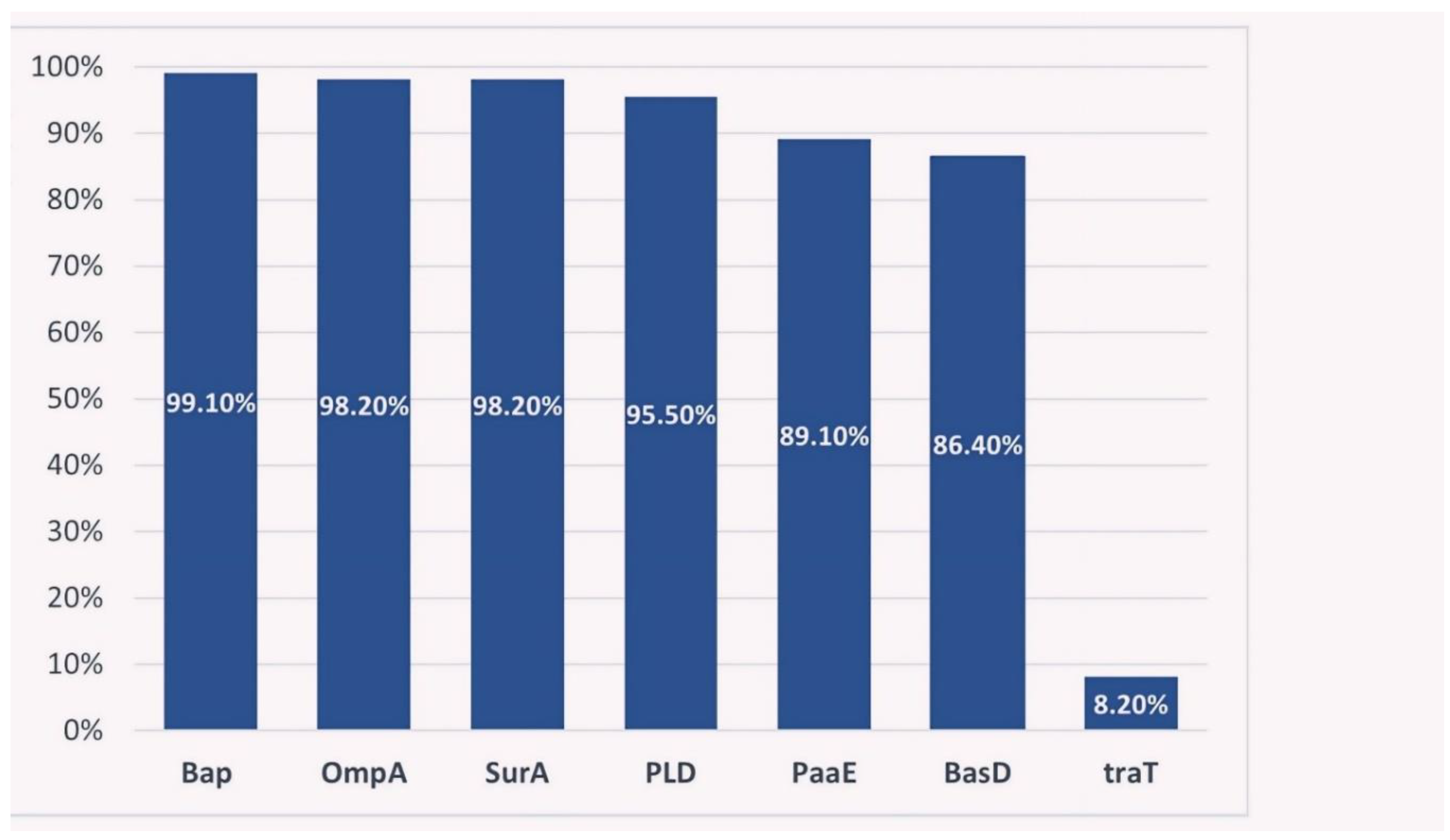
Figure 3.
Microtiter plate method for detection biofilm production ( A: negative control, B: strong producer, C: moderate producer, D: weak producer, E: non producer).
Figure 3.
Microtiter plate method for detection biofilm production ( A: negative control, B: strong producer, C: moderate producer, D: weak producer, E: non producer).
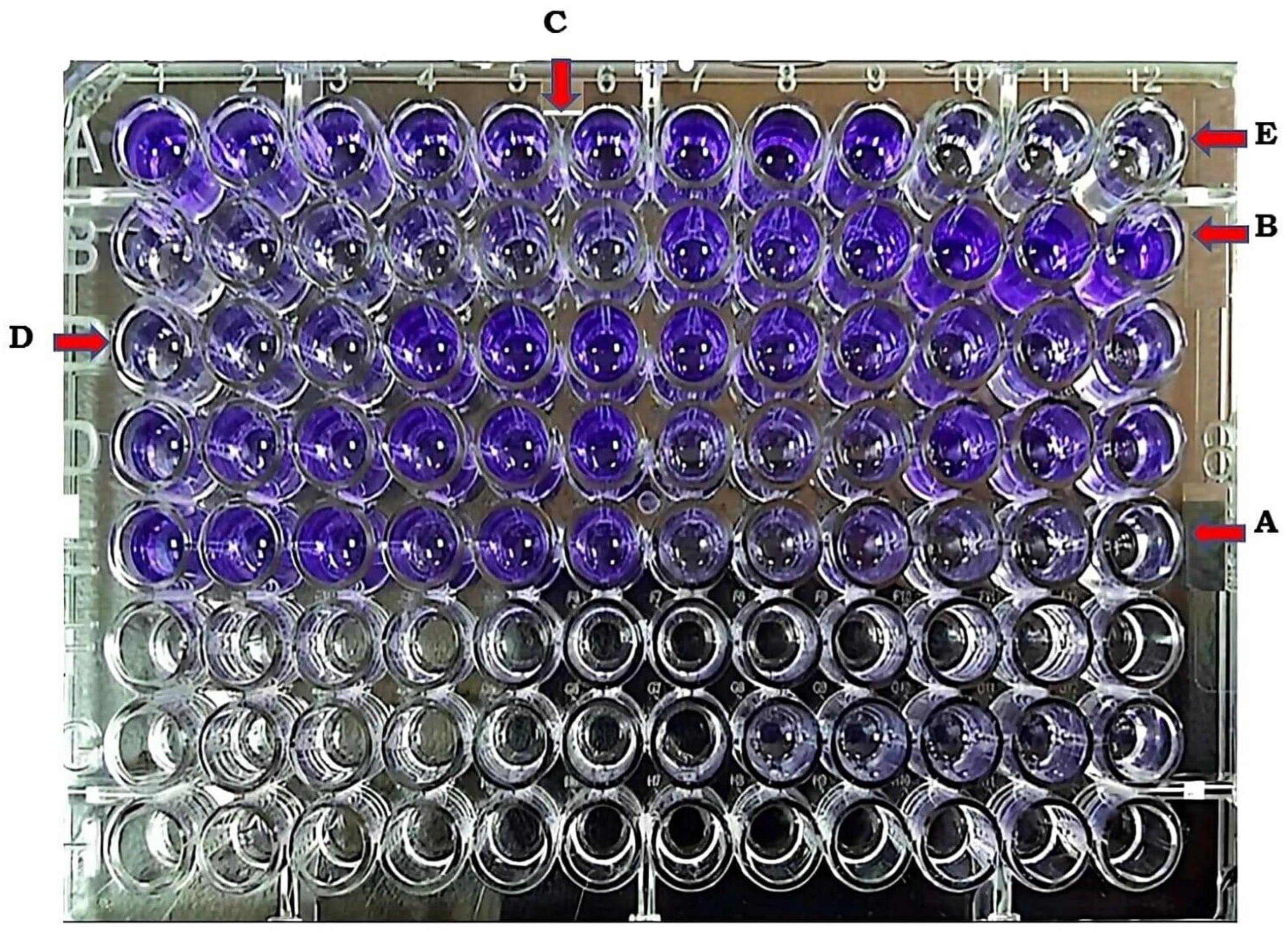

Table 1.
Selected A. baumannii virulence factors.
| Virulence factor | Gene | Role during pathogenesis | Ref. |
|---|---|---|---|
| biofilm-associated protein | Bap | Have a key role in the development, thickening of mature biofilm structure, and its intercellular adhesion. It enhances the persistence of A. baumannii in the hospital environment, and promotes antibiotic resistance and tolerance to other inhibitors | [12,13] |
| Surface antigen protein | surA1 | Surface antigen protein is a periplasmic chaperone protein necessary for cellular invasion. | |
| Iron acquisition system | BasD | Iron acquisition system is essential for bacterial survival and growth in a host under iron-limited conditions | [14] |
| Phospholipase D | Pld | Phospholipase D enhances A. baumannii ability to thrive in serum and invade epithelial cells | [15] |
| Outer membrane protein A | OmpA | Outer membrane protein A (OmpA) is an adhesion protein enhances bacterial attachment to eukaryotic epithelial cells and formation of biofilms | [16] |
| Phenylalanine catabolic pathway | PaaE | A. baumannii exhibits a unique enzymatic pathway encoded by PaaE gene for degrading phenylacetic acid and a carbon source to enhance its adaptability and survival in various environments | [17] |
| Surface-exposed lipoprotein | traT | A surface-exposed lipoprotein linked with serum resistance and evasion of the host immune response and may contribute to the ability to survive and infect the host | [17] |
Table 2.
Primers details for all seven virulence genes.
| Gene | Sequence | Annealing temp (ºC) | Ref. |
|---|---|---|---|
| Bap | F:AGTTAAAGAAGGGCAAGAAG | 50 | [17] |
| R:GGAGCACCACCTAACTGA | |||
| surA1 | F:CAATTGGTAGCTGGCGATCA | 55 | [17] |
| R:TTAGGCGGGACTCAGCTTTT | |||
| BasD | F: CTCTTGCATGGCAACACCAC | 65 | [17] |
| R:CCAACGAGACCGCTTATGGT | |||
| Pld | F:CGTCAATTACGCCAAGCTG | 64.7 | [17] |
| R:CTGACGCTACCTGACGGTTT | |||
| OmpA | F:CGCTTCTGCTGGTGCTGAAT | 50 | [10] |
| R:CGTGCAGTAGCGTTAGGGTA | |||
| PaaE | F:CTATTTAGGCGTTGCTGCGG | 64.5 | [17] |
| R:CCTTACAACGACAGGTCGCA | |||
| traT | F:GGTGTGGTGCGATGAGCACAG | 67.9 | [17] |
| R:CACGGTTCAGCCATCCCTGAG |
Table 3.
Biofilm production (strong, moderate, weak, and non) among carbapenem-resistant Acinetobacter baumannii isolates.
Table 3.
Biofilm production (strong, moderate, weak, and non) among carbapenem-resistant Acinetobacter baumannii isolates.
| Biofilm production | |||||
|---|---|---|---|---|---|
| Strong | Moderate | Weak | Non | Total | |
| Frequency | 24 | 40 | 31 | 15 | 110 |
| Percent | 21.8% | 36.4% | 28.2% | 13.6% | 100% |
Table 4.
Correlation between biofilm and presence of carbapenem resistance gene (VIM, OXA-23) genes.
Table 4.
Correlation between biofilm and presence of carbapenem resistance gene (VIM, OXA-23) genes.
| Carbapenem resistance gene | Biofilm production | p-value | ||
|---|---|---|---|---|
| Negative | Positive | |||
| VIM | Negative | 13 | 68 | 0.218 |
| Positive | 2 | 27 | ||
| OXA-23 | Negative | 1 | 0 | 0.011 |
| Positive | 14 | 95 | ||
Table 5.
A summary for the prevalence of CRAB virulence genes.
| Virulence gene | Frequency in Jordan (this study) | Study country | Frequency range | Ref |
|---|---|---|---|---|
| Bap | 99.1% | S. Korea, Iran Serbia, Thailand | 48-100% | [10,27,42,43] |
| OmpA | 98.2% | Iran, S. Korea, | 77.1-100% | [16,41] |
| surA | 98.2% | Iran, China | 95-98% | [16,17] |
| PLD | 95.5% | China, Poland | 87.5-99% | [15,17] |
| PaaE | 89.1% | China | 88.6% | [17] |
| basD | 86.4% | China, Poland | 92-95% | [17,15] |
| traT | 8.2% | China, Iran | 0-80% | [17,43] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



