
Submitted:
08 May 2024
Posted:
09 May 2024
You are already at the latest version
Abstract
The gut microbiota (GM) communicates with the brain via biochemical signaling constituting the gut-brain axis, which significantly regulates the body's physiological processes. The GM dysbiosis can impact the digestive system and the functioning of the central nervous system (CNS) linked to the onset of neurodegenerative diseases. In this review, the scientific data compiled from diverse sources primarily emphasizes the neuropathological characteristics linked to the accumulation of modified insoluble proteins (such as β-Amyloid peptides and hyperphosphorylated tau proteins) in Alzheimer’s Disease (AD) and the potential impact of gut microbiota (GM) on AD susceptibility or resilience. The specific GM profile of human beings may serve as an essential tool for preventing or progressing neurodegenerative diseases like AD. This review focuses mainly on the effect of gut microfauna on the gut-brain axis in the onset and progression of AD. The GM produces various bioactive molecules that may serve as proinflammatory or anti-inflammatory signaling, contributing directly or indirectly to the repression or progression of neurodegenerative disorders by modulating the response of the brain axis. Human studies must focus on further understanding the gut-brain axis and venture to clarify microbiota-based therapeutic strategies for AD.
Keywords:
Alzheimer’s Disease
; Neurodegenerative disorders
; β amyloid
; tau protein
; Gut microbiota brain axis
1. Introduction
The term "human microbiota" refers to the collection of tiny creatures inhabiting aspecific habitat, including protists, bacteria, fungi, archaea, and viruses. Specific microbiomes in the body are found in organs like the lungs, skin, and gastrointestinal system. The gastrointestinal tract (GIT's) microbiota, or bacteria, has been the subject of the most scientific research among these ecosystems. The microbiota, which consists of commensal (or potentially pathogenic) bacteria, is found on the surfaces of the mucosal membranes and in the lumen of GIT. Over 90% of the species of bacteria in the GIT belong to the Firmicutes and phyla Bacteroidetes, with very few amounts of Proteobacteria, Actinobacteria, and Verrucomicrobia [1].
The gut-brain axis is a bidirectional, biochemical signaling interaction between the brain and gastrointestinal system. The GM plays a role in physio-biochemical signaling events involving the duo's participation. Hence, it is also referred to as the Microbiota-Gut-Brain (MGB) or Brain-Gut-Microbiota (BGM) axis [2]. The integrative physiology of the MGB axis involves the participation of the central nervous system (CNS), autonomous nervous system, hypothalamic pituitary adrenal axis, enteric nervous system, intestines, and other organs to orchestrate response that influences the functioning of gut cells such as epithelial, smooth muscle, immune, enterochromaffin and other cells (Figure 1) [3]. Gut and brain work in close coordination, influencing each other’s functioning due to their common origin from the tissue- the neural crest [4]. They perform abundant physiological functions, including food intake, satiety, bone metabolism, insulin secretion, responsiveness, glucose, and fat metabolism [5]. The mucosal barrier of the intestine comprises the intestinal epithelial layer, mucosal layer, and microorganisms. The microorganisms constituting the GM influence the proper functioning of the intestinal mucosa and epithelial cells. However, GM dysbiosis has an impact on the digestive system and the functioning of the CNS. The microorganisms that help compensate the intestinal flora may otherwise have a significant capacity for secreting amyloid and lipopolysaccharides (LPS), which may affect signaling pathways as well as the generation of pro-inflammatory mediators connected to the initiation of AD [6].
AD pathogenesis can be explained by different hypotheses, including the amyloid cascade hypothesis (ACH), tau hyperphosphorylation or accumulation of neurofibrillary tangles, and oxidative stress. The development of Alzheimer's disease (AD) involves complex molecular and cellular mechanisms. Extracellular amyloid β (Aβ) protein, which forms senile plaques (SP), and intracellular neurofibrillary tangles (NFTs) made up of phosphorylated tau (p-tau) protein, are well-known pathological features of AD. Additionally, neuroinflammation has been shown to contribute significantly to the disease progression of AD. Other contributing factors include cerebrovascular amyloidosis and alterations in synaptic function. Collectively, these pathological changes disrupt the delicate balance of cellular pathways and ultimately lead to cognitive decline and other symptoms of AD. Other factors, such as aging, genetics, lifestyle, viral and bacterial infections, etc., may also affect AD pathogenesis. [7,8,9,10]. The gut-brain axis is a bidirectional communication path between the CNS and gastrointestinal tract. Moreover, the LPS produced and secreted by pathogenic GM is regarded as one of the factors of gut dysbiosis characterized by an imbalance between eco-friendly and pathogenic GM. LPS from dead bacteria are far more ubiquitous in the microbiota-associated composition with AD. LPS stimulates enteric signaling pathways producing the proinflammatory cytokine-nuclear factor kappa-B (NFk-B). NFk-B acts as a precursor for the production of interleukin-1 (IL-1), interleukin-6 (IL-6), as well as tumor necrosis factor-α (TNF-α). Studies have shown that LPS has the ability to enter the bloodstream from the digestive tract and activate astrocytes and microglia in the brain, which promotes neuroinflammation in brain tissues.
The blood-brain barrier (BBB) is formed by endothelial cells, pericytes, and mural cells from the perivascular region. It controls molecular exchange in the intricate networks of the brain's microvasculature. Larger molecules, like proteins, peptides, and highly harmful blood-related substances, are blocked from entering the brain by the BBB, but nutrients like sugars and proteins can be transported from blood to the brain and back and is managed by carrier-derived bidirectional trans-endothelial transfer. [11]. The interaction of several neurovascular unit cell types regulates the integrity of the BBB, cerebral blood flow, and several other CNS activities. The normal integrity of BBB plays a major role in preventing the entry of bioactive harmful substances released from GM, which will otherwise lead to CNS damage.
With the advancement of scientific research in neuroinflammation, a wide contribution of GM has been found to be associated with the interference of the brain's physiological processes. Any disturbance in the gut-brain axis negatively impacts the pathogenesis of neurodegenerative diseases. Shifting of the balanced and stable state of GM composition directly affects the permeability of the blood-gut barrier, which leads to the passage of GM-released bioactive substances and, subsequently, activation of the enteric immune system [12]. In aged individuals, an activated systemic immune system significantly promotes neural injury and, finally, neuroinflammation, also known as “inflammaging” [13]. In germ-free mice (GFM), it has been found that the gut microbe composition significantly influences early CNS development and neurogenesis [14]. Reports from various scientific sources prove that the typical pathognomic lesions, like accumulation of p-tau proteins intracellularly and amyloid β fibrils, are associated with the change in the constitution of gut microfauna [15]. Therefore, there is a need for an hour to have a comprehensive knowledge of GM and its role in the development of brain inflammation.
Diet is an essential determinant of the type and role of GM in human beings. Diet plan and composition are closely related to developing AD [16]. Dietary pattern is an adaptable factor for the amelioration of microbiota. Neuroprotection is linked with the Mediterranean diet and the Ketogenic diet (KD). Probiotics, prebiotics, and fecal microbiota transplantation (FMT) may be potential therapeutics for AD and other neuroinflammatory conditions [17]. However, to draw a conclusive statement concerning a certain form of dietary pattern as an alternative to therapeutics, further studies are to be carried out for absolute validation. AD is the most common neurodegenerative disorder seen in age-dependent dementia and is characterized by progressive cognitive impairment. GM of the gastrointestinal ecosystem possesses a diverse population of microbial species whose modifications can have a role in the progression of gastric and CNS disorders like AD [18]. A comprehensive update on GM-associated neuroprotection or neurodegenerative disorders associated with dysbiosis is relevant to explore the mechanistic approach and potential targets in preventing and managing neurodegenerative disorders.
The current research review article exclusively emphasizes the bidirectional communications of faunal life residing in the gastrointestinal tract with the brain and the effects of their shifts toward the development of AD in humans, which is exclusively associated with neurodegeneration.
2. Gut-Brain axis Dysbiosis in AD development
The role of distinct GM serves as one of the essential innovative ways of studying the functioning of the gut-brain axis (GBA) besides, this bidirectional connection with the brain is termed the MGB axis” (Figure 1). MGB refers to a bidirectional communication between the brain and the gut involving multiple overlapping pathways, including the autonomic, neuroendocrine, vagus nerve, the immune system, or the metabolic processes of GM and immune system in addition to bacterial metabolites and neuromodulatory molecules [19,20].
Biological research output related to AD and the role of GM with the onset of AD are still in their initial stages. The last few decades, neuroscientists have been working tirelessly to find the exact cause of this neurodegenerative disease, which remains a puzzle, but recent scientific data reveals that its occurrence begins ten years before the appearance of clinical symptoms [21]. Metchnikoff, a noble laureate in immunology, hypothesized in the early 21st century that some distinct bacteria (commensals) residing in the colon of human beings have beneficial effects on human health while some other bacteria have negative effects (pathogenic). Under normal physiological condition, the gut microbiota (GM) plays a role in promoting proper immune maturation and serve as a source of essential nutrients for the host. However, dysbiosis, which refers to imbalance in the composition of beneficial and pathogenic GM, has been associated with development of various diseases, including AD. Spychala et al., (2018) have found a balance of gut microbiota may be important for overall health, including the prevemntion of certain diseases.
Recent studies suggest a significant linkage between the gastrointestinal tract, its GM and the brain via bidirectional communications. The signaling mechanisms include top-down signaling (brain→gut) and bottom-up signaling (gut →brain). The top-down signaling proceeds directly via sympathetic and parasympathetic neurons or indirectly by stimulating the enteric nervous system located in the sub-mucosa and myenteric plexi in the gut wall [22]. Alteration in the interaction between gut dysbiosis and host microbiota leads to neuroinflammation in the brain closely associated with the development of AD, which follows the progressive degenerative pathways leading to alteration/increase in permeability of the gut barrier and activation of active cellular components of the immune system that harm blood-brain functions, promote neuronal injury, neuroinflammation and finally development of AD [13]. The GM is well known as the origin of an abundant amyloid, LPS, and other toxic chemicals; therefore, microbiota predisposes animals to systemic inflammation and disruption of physiological barriers [13]. Microbiota inhabited in the human gut not only assist gut-specific activities like the fermentation of carbohydrates for byproduct formation, vitamin synthesis, and detoxification of xenobiotics [14,15] but also act as a protective tool against the pathogenic bacteria in the gut [23] and maintain the neuronal health of an individual.
Based on clinical and preclinical studies, the production of some biomolecules like neuromodulators and neurotransmitters, from GM has a direct impact on gut-brain communication, behavioral changes and brain activities [24,25,26,27]. The released bioactive chemicals/transmitters of GM can intrude from GIT to the brain or spinal cord, especially during old age, which can activate abnormal formation of amyloid protein with enhanced aggregation [13]. The GM products can activate immune cells, boosting inflammatory response in brain, which in turn influences immune cells like microglial cells function. Activated microglial cells start neuroinflammation in CNS, resulting in neuron damage, which is a major factor contributing to AD development. The two characteristic pathological lesions of AD i.e. extracellular amyloid plaques and neurofibrillary tangles composed of mainly Aβand p-tau proteins respectively [28].
A rat model study based on Bifidobacterium infantis, one of the essential microorganism, indicated that the gut microbe has a significant connection to immune response in the brain [24]. Increase in the number of Lactobacillus casei, Bacteroides fragilis, and Streptococcus thermophiles in the rodent intestine revealed a positive effect on brain activity and performance[24,29,30]. On the other hand, Eubacterium rectale, Porphyromonas gingivalis, and Lactobacillus rhamnosus can play a dynamic part in the start of AD [31,32,33,34,35,36]
GM like Escherichia, Lactobacillus, Saccharomyces, and Bacillus are actively involved in the synthesis of gamma-amino butyric acid (GABA), 5-hydroxytryptamine, dopamine, butyrate, histamine, and serotonin, which play an important contribution in regulating the brain activity of an individual after entering the mucosal layer of intestine and successively are capable of entering the blood stream (Table 1) [37,38,39](22, 23, 24]. Decreased population of microbiota in aged individuals with brain pathology has been associated with lower levels of butyrate which in turn predisposes other brain injuries and progressive cognitive loss [40]. Recently it was revealed that AD could be initiated even in the gut and progress towards the brain due to translocation of αβ oligomers [41].
2.1. Bridge between Neuroinflammation and Microbiota Dysbiosis
The research suggests that there is a supplementary relationship between immune system and GM. The immune system plays a role in regulating the composition and structre of the GM, while GM influences the maturation and function of the immune system [46,47]. Numerous studies shown that antibiotic-treated mice or germ-free mice (GFM) possess lower ratios and impaired differentiation capabilities of myeloid cell progenitors, indicating that the GM influences immune system development [48]. Additionally, the absence of of GM in mice or germ-free rodents can lead to altred gene expression in immune cells like microglia, resulting in dysregulation of immune system related defecnce activities such as pathogen recognition and cell activation.
[49].
Tau pathway:
AD is included in one of the tautopathies characterized by the formation of neurofibrillary tangles composed of p-tau proteins [50]. Tau, is a water soluble protein associated with the stability of axonal microtubules [51] if detached from these microtubules results in the loss of synapsis and neuronal death [50]. According to tau hypothesis, altered tau proteins deposits stimulate the neurodegenerative disease as found in AD [50]. On a molecular level, tau proteins are microtubule-associated proteins that assist in axonal transport and microtubular stability [52]. Tau’s inflection of tubulin assembly and stability is regulated by its degree of phosphorylation. In neuroinflammatory processes, such as AD, Tau protein undergoes hyperphosphorylation, leading to its conformational change over into β-sheet rich structures, forming large and insoluble tangles [53]. During pathological conditions, there is depolymerisation of microtubules due to hyperphosphorylation of tau proteins, which disintegrates them from each other leading to the p-tau protein aggregations and, characteristically the formation of neurofibrillary tangles (NFTs) inside the neurons [54]. By the age of 40 years, there is increase in concentration of p-tau and total tau (t-tau) proteins inside neurons[55], which block the transport of nutrients into the neurons leading to the cell death Besides hampering nutritional transport in neurons, the altered tau proteins are responsible for activation of microglial cells; hyperphosphorylated tau proteins also spread in a prion-like fashion, resulting in neuronal and synaptic anomalies [56]. Recent studies have shown that disturbances in insulin-mediated molecular pathways are also associated with AD pathogenesis [57]. The binding of insulin to insulin receptors leads to the activation of various insulin receptor substances (IRS), which activate downstream signaling pathways, including phosphatidylinositol 3-kinase (PI3K) and extracellular signal-regulated kinase (ERK) cascades that regulate various physiological processes [58]. While progressing the PI3K pathway, there is phosphorylation of glycogen synthase kinase (GSK) 3β, which deactivates it. GSK 3β regulates tau phosphorylation, which contributes to the formation of NFTs. Activation of GSK 3β could thus induce hyperphosphorylation of tau proteins, thus contributing to the development of AD [59].
GM secretes an abundant number of metabolites which mainly include valeric acid, isovaleric acid, isobutyric acid, butyric acid, propionic acid, acetic acid and formic acid and their effects on the pathogenesis of AD have been explored by disturbing the activations of microglia and astrocyte, help to reduce inflammation, and aggregations of Aβ and tau proteins[60]. After proving by the meta-analysis, various studies have indicated that the bioactive chemicals released from GM can increase the degree of inflammation, tautopathies and Aβ aggregation in CNS[61].
Moreover, tau hyperphosphorylation has been found to be facilitated by IL-1 via MAPK pathway, IL-6 via cyclin-dependent kinase 5 pathway and NO compound [62,63]. Data related to microbiome effects on gut-brain health implies decrease in the levels of pro-inflammatory cytokines such as IL-1β, IL-6, TNF-α, TNF-β[64], whose increased levels have been found to be associated with neurological disorders [65]. Thus, these results suggest a close association between the immunomodulatory action of GM-released pro-inflammatory cytokines and neuroinflammation [66]. Clinton et al., 2010) studied the cross-interaction between the different proteins of amyloid plaques in AD and reported a mutual interaction of tau protein with Aβ proteins [67]. Microglia maturation plays a crucial role in limiting the concentration of p-tau proteins, thereby reducing the chances of AD. Erny et al. (2015) studied the inflammation of the brain in GFM where they found an increase in microglia number; however, after treatment of these GFM with lipopolysaccharide (LPS), an agonist of transmembrane immune system associated receptor, TLR4, they observed an increased microglia maturation with reduced levels of chemokines and cytokines[68].
In their stdy, Zhao and Lukiw investigated the role of microglia activation and reduction in p-tau proteins under physiological condition. They suggested that bacterial amyloids facilitate the clearance of Aβ aggregates and hyperphosphorylated tau proteins by microglia cells through the receptor of microglia/myeloid-2 cells (TREM-2). This suggest that bacterial amyloids may play a role in the regulation of protein aggregation in the brain, potentially influencing neurodegenerative process.
(TREM-2) [69].
2.2. Blood Brain Axis Alteration by GM
The term "gut microbiota-blood brain axis" describes a two-way network of information that consistently exchanges information between the neurological and digestive systems [70]. It is widely acknowledged that the intestinal microbiota has an impact on the physiological, behavioral, and cognitive processes of the brain (Table 2). The parasympathetic and sympathetic-autonomic neurological networks, the nervous system, metabolic waste GM, the neuroendocrine system, the immune system of the nervous system, and the CNS may be included in this. Other potential communication channels in the GM and brain, including the BBB and the intestinal mucosal barrier, neuroendocrine adrenal axis, pituitary and hypothalamic, immune system of the gut, neurotransmitters, and regulators of neural activity, are produced by the bacteria of the gut [70]. Our metabolism, digestion, nutrition, growth, immunology, and defense against infections all depend majorly on the human microbiota [71,72,73]. The easiest way to understand how microbiota affects immunity is from the perspective of evolution, as it serves the microorganism’s adaptive interest to keep the host in a state allowing for colonization. The complex molecular progression has preserved host to preserve immunological tolerance due to utmost importance of microorganisms for human existence. The term "brain-gut-microbiota axis" was expanded due to the significance of the microbiota's impact on the brain [74].
Under normal circumstances, low-density lipoproteins of receptor-derived protein 1 transport dissolved amyloid beta (A) in the reverse direction from the circulatory system to the nervous system[80]. Post-mortem investigations done on AD patients revealed BBB damage as well as an accumulation of capillary substances in the nervous system [81]. This process was supported by MRI scans of the human brain, which revealed age-dependent blood-brain protective narrowing in the hippocampus, a region responsible for memory and learning. The breakdown that Cerebrospinal fluid (CSF) analysis linked to pericyte damage was more severe in mild cognitive impairment. Pericyte destruction is accelerated by the E gene 4 apolipoprotein alleles, which are the primary genetic susceptibility factors for AD [82].
According to certain studies, stress during a child's formative years may change the microbiota, which raises the risk of stress-related disorders later in life[83,84]. Maternal separation following three days after birth caused a reduction in lactobacillus levels in the baby rats that had a large effect on the gut microbiota [83]. Prenatal stress altered the microbiome of rhesus monkeys by decreasing the overall concentration of Lactobacillus and Bifidobacterium [85]. The incidence of experimental colitis in children was also increased by prenatal antibiotic use [86]. These results revealed stress might alter the flora in the gut. A neurovascular unit is made up of glial cells (microglia and astrocytes), vascular endothelial cells, neurons, and pericytes [11].
Permeability of the BBB and gut blood barrier is highly influenced by microbial dysbiosis, which serves as a predisposing factor for neurological disturbances like progression of AD pathogenesis as well as other neurodegenerative diseases, especially those linked to aging. The integrity of the BBB is inadequate, enabling LPS to transfer through it and leading to inflammation in the brain. As a result, a thorough understanding of GM and their role in the development of brain inflammation is required (Figure 2). The endothelial cells of the brain and pericytes create the BBB, which safeguards the brain and nervous system from blood-derived chemicals, infections, and cells [80]. Modern research on transgenic mouse models has demonstrated the importance of pericytes in regulating the BBB integrity and its loss can contribute in the genesis of small vessels diseases of long-term BBB collapse, and neurodegenerative alterations [87,88,89].. Multiple investigations have shown that the apolipoprotein E4 (apoE4) genotype accelerates the loss of integrity of the cerebrovascular system and BBB impairment in AD [90,91,92]. Three main apoE isoforms exist in humans: E2, E3 and E4. Genetic evidence, particularly new genomic sequence linkage analyses, indicate apoE4 as the most important and increasingly recurrent genetic potential risk factor for AD. A single apoE4 copy confers approximately a 3.7-fold increased risk of AD compared to two copies, which confers a 12-fold increased risk compared to carriers of the apoE3/E3 genotype. Research utilising transgenic mice has shown that apoE4 enhances BBB sensitivity to injury, induces BBB breakdown, and decreases micro vascular concentration compared to apoE3 mice [93].
The exact etiology involved in the breakdown of BBB in AD, especially in apoE4 individuals, is still unknown. Inhibition of the cyclophilin A proinflammatory (CypA)-metalloproteinase-9 (MMP-9) mechanism throughout pericytes through reduced lipid receptor-derived peptides-1 (LRP1), a crucial apoE binding site, astrocyte-released apoE2 as well as apoE3 were shown to preserve the integrity of the blood-brain protective layer in recent research utilizing transgenic apoE2 as well as apoE3 models of mice [94,95]. The CypA-MMP-9 system is ineffectually inhibited by astrocyte-released apoE4 in the APOE4 mouse models, which leads to MMP-9-derived degradation of the BBB firm connection, causing basal lamina proteins as well as BBB breakdown [96]. Additionally, a recent study contrasting cognitively normal healthy apoE4 transport to apoE3 providers found a dependence of age and an active MMP-9 concentration in the CSF, suggesting activation of the CypA-MMP-9 route. This raised ratio of CSF to plasma albumin indicates BBB collapse and is found to correlate with the CypA-MMP-9 signalling pathway [97]. Here, researchers use post-mortem human brain tissue analysis as indicators of BBB collapse to evaluate that apoE4 compared to apoE3 increases pericyte losses as well as micro vascular abnormalities in the disease of AD, and correlates with the amount of BBB disintegration to molecules like immunoglobulin G as well as fibrin. apoE4 generates a larger aggregation of CypA and MMP-9 in endothelial cells and pericytes in AD relative to apoE3, indicating a higher stimulation of the LRP1-dependent CypA-MMP-9 BBB-mortifying mechanisms. The loss of pericytes may also contribute to apoE4's elevated CypA as well as MMP-9 build-up, which may hasten BBB rupturing in AD apoE4.
3. Mechanism of Brain and Intestinal Blood Barriers Interaction in Alzheimer's Disease
The GIB constantly contains the microbiota, sometimes called innocuous microbial flora and food antigens, which are essential for digestion and act as the first line of defence in the event of an attack by invasive infections [98]. The mucosal barrier of the intestine is made up of the intestinal epithelial layer, mucosal layer, and microorganisms. The mucous barrier, which is composed of mucin molecules secreted by goblet cells, protects the epithelial layer from infections by acting as a barrier. Proteins of tight-junction like occludin, claudins, and adhesion of junctional cells or kitricellulin join epithelial tissues. The microbiota improves the viability of the intestinal mucosa as they indirectly produce SCFAs and stimulate the proliferation of epithelial cells. The ability of intestinal epithelial cell monolayers to become more permeable by the pro-inflammatory cytokines IL13, TNF, IL6, IL4, and IL13 has been associated with higher production of claudins [99].
Specific microbiomes in the body can be discovered in organs like the lungs, skin, and gastrointestinal system. The GIT microbiota, or bacteria, has been the subject of the most research among these ecosystems. The microbiota, which consists of commensal (or potentially pathogenic) bacteria, is found on the surfaces of the mucosa and in the lumen of the GIT lumen. Over 90% of the species of bacteria in the GIT belong to the Firmicutes and phyla Bacteroidetes, with very small amounts of Proteobacteria, Actinobacteria, and Verrucomicrobia[1].
Our gastrointestinal ecosystem is a diverse population of microbial species called GM, whose modifications can impact both digestive illness and CNS diseases like AD. The permeability of the BBB and gut caused by microbial dysbiosis also acts as a mediator or impacts the pathogenesis of AD and other neurodegenerative diseases, especially those linked to aging. The microorganisms that help compensate the intestinal flora also have a significant capacity for secreting amyloid and lipopolysaccharides, which may affect signalling pathways and the generation of mediators of pro-inflammatory response connected to the initiation of AD [6].
LPS models for neuropathology in AD Bactericides that create LPS are far more ubiquitous in the microbiota composition associated with AD. When LPS stimulates enteric signaling pathways to produce IL-1, IL-6, as well as tumour necrosis (TNF), the nuclear factor kappa-B (NF-B) pathway is activated, which increases the production of proinflammatory cytokines. Similarly, LPS activates microglia in the brain by enhancing the non-selective nerve cell that promotes neuroinflammation. LPS seems abled to enter the bloodstream from the digestive tract and activate astrocytes. The integrity of the BBB is inadequate, enabling LPS to transfer through it and leading to inflammation in the brain.
3.1. The Cerebral Blood Barrier's Involvement in the Brain Microbiota Intestines Axis in Alzheimer's Disease
The IEB is established by the mucosal barrier, which divides the host and microbes and permits only a small number of specific, extremely tiny molecules to pass through. Monolayer epithelial cells comprise this interface and its IEB consists of diverse immunological cellular immunity that assists in the host's defense and physiology. The functions and permeability of the IEB are primarily regulated by chemicals found in the body, including inflammatory agents, immune cells, bacteria, and metabolites. The vascular barrier of the gut (GVB), an acellular membrane underneath the epithelium, has just recently been established. GVB, the second line of defense, controls antigen translocation as well as prevents the entrance of intestinal microbiota.
The BBB consists of the CNS, segmental circulation of blood, pericytes, and brain endothelium. The condition, also known as "leaky gut syndrome," affects IEB permeability and causes peripheral circulatory system inflammation and the expulsion of microorganism secretions (including endotoxins such as lipopolysaccharide). Peripheral monocytes activate during inflammation, penetrate the BBB, activate microglia, release inflammatory cytokines, and ultimately lead to neuronal inflammation in AD disease. Additionally, the development of neuroinflammation depends on the infiltration of peripheral lymphocytes, including 17 T-helper cells (4a).
3.2. Intestinal Disorders
Because the GIB has an important role in preserving gut homeostasis, problems related to any of its parts may exacerbate irritable bowel conditions. In UC, IBD, and Crohn's disease (CD) featured as ongoing inflammation that may harm the colon and terminal ileum. The illness may affect all mucosal surfaces, and a histological examination will reveal granulomas in 50% of cases. Contrarily, with UC, the colon is the organ affected and the inflammation only affects the surface mucosa. These illnesses are complicated and have both genetic and environmental components.
Evidence that the bacteria has an important role in IBD development includes the ability of antibiotics to lessen the severity of the condition [100], the genetic link between IBD and mutations within genes encrypting bacterial detectors TLR4, CARD4 (NOD1), CARD15 (NOD2), or TLR4 [101,102], or genes mutations related to cellular metabolism, which are essential for innate defense [103,104]. Whilst sought after, there is no concrete evidence that any pathogenic microorganisms cause IBD. Contrarily, it has been shown that IBD individuals also have innate immunity to their own flora, however, healthy individuals exclusively do so in exposure to exogenous flora [105].
It is still unclear where these reactions start and whether there is a fundamental issue with intestinal absorption that allows germs that are typically excluded to enter. Furthermore, since the surrounding microenvironment gives DC’s and macrophages a non-inflammatory appearance, changes in this compartment affect the gut's capability to sustain homoeostasis by reducing immune cells' capacity to control their pro-inflammatory potential [106,107,108,109]. Another inflammatory gastrointestinal condition that may be related to GIB impairment are IBS, which is a complex, multifaceted disorder. IBS is associated with psychological stress, most likely due to the delivery of corticotrophin hormone release (CRH) [110]. CRH can stimulate the immune cells, degranulate mast cells, and enhance intestinal absorption [111]. The IBS Individuals' rectal biopsies, independent of their IBS subtypes, contain an overwhelming number of mast cells, pointing to a link between mast cell activity and IBS [112]. It would be fascinating to learn if problems with BBB permeability could affect gastrointestinal disorders or vice versa.
Various investigations are conducted on animals. GF animals live in a sterile environment and are microbially lacking. These animals provide exceptional opportunities to investigate the role of the intestinal microbiota in the etiology of AD. The GF mice displayed abnormalities in the object recognition test, a non-spatial task, and had decreased CNS neurotropic factor levels in their hippocampi. [33]. On the other hand, the GF mice showed a deficiency in the innate immune system, which was shown by immaturity in the microglia. According to research by Fujii et al., mice that had the GM of an AD patient transplanted showed abnormal behavioral cognition.[113].
The blood vessels in the CNS are notable for providing the BBB. Contrary to blood vessels in most tissues, which are porous and allow a relatively unrestricted stream of capillary ions, molecules, and cells through into tissue, arteries in the CNS detrimentally restrict the flow of ionic species, blood-related molecules, as well as cells in nerve tissue [114].
The BBB is an essential secondary barrier since the CNS tissue cannot regenerate after illness or injury. Stroke, trauma, and multiple sclerosis are a few neurological disorders that cause the barrier to collapse, and they plays a significant role in the pathophysiology of these illnesses[11].
In disease, the BBB collapses. AD leads to the deposition of insoluble proteins, which causes microbleeds and hemorrhages in the brain. A plaque develops along the blood vessel walls. [115]. The brains of AD patients have been found to have extracellular clumps of the possibly toxic Aβ protein and intracellular p-tau. [116]. As a result, a lot of evidence points to the importance of these proteins in the pathogenesis of AD.
Numerous epidemiological studies have also demonstrated a high linkage between AD and cardiovascular risk factors. [117]. It has long been established that the BBB plays an important role in the pathophysiology of AD through brain clearance A, lymphocyte recruitment further into the brain, and inflammatory response regulation. [80]. It has been shown that the BBB leaks in people before the hippocampal shrinkage, which is frequently noticed in the initial stages of AD. [118]. This shows that vascular abnormalities precede neurodegeneration. The extravasation of the blood-related chemicals in the brain, which causes inflammation and immunological reactions, also frequently occurs in conjunction with AD. [119]. The BBB protects the brain from blood-related derived substances, infections, and cells, which is required for proper information processing and neural activity. The post-mortem and scanning of brain research have shown a disruption of the BBB and blood-related accumulation of molecules in the cerebrospinal fluid, hippocampus, and cortex. Additionally, research using murine models has demonstrated that blood-derived chemicals, such as thrombin, immunoglobulin, albumin, fibrinogen, prothrombin kinase-2, and thrombin that are typically penetrated in the brain. However, damage to the neurons with enhanced BBB membrane permeability in the hippocampus accumulates in tissue as a result of BBB disruptions. Both barriers are essential as they restrict the admission of dangerous molecules and possess a significant impact on immunological recognition as well as homoeostasis preservation. Additionally, the BBB and IEB are essential for controlling the microbiota-intestine brain-gut axis [120].
4. Gut Microbiota Models of Animals in Alzheimer’s Disease
Microbiota Gut Animal models of AD give a real image of the GM, contributing to the disease's etiology [121,122]. In an AD mouse model, neuroinflammation and amyloidosis had an impact on the variety of the GM [121]. Some studies included extracting 16S rRNA from bacteria (fecal APP transgenic mice samples) and showed that the microbiota of the guts of these animals differed from that of wild-type control mice. [123]. Rikenellaceae increased due to these modifications, but Allobacillum and Akkermansia decreased. Obesity and type 2 diabetes have already been linked to a decreased abundance of Akkermansia, which are both known to increase the risk of dementia. Additionally, there was a bad connection between the abundance and presence of A42 in the brain [124,123] Cerebral A was considerably decreased in transgenic APP transgenic mice that were germ-free. Additionally, reduced microgliosis and modifications to the cytokine profile were seen. Recolonizing the mice with germ-free APP transgenic mice microbiota increased cerebral pathology, whereas using wild mouse microbiota had less effect[123]. Similarly, PD-modelling germ-free animals overexpressing -syn showed lower -synmicroglial inclusions and activation compared to the controls. Reintroducing microbiota, particularly those from patients of PD donors, or adding SCFAs—by-products of microbial metabolism—restored these characteristics [125]. It's an intriguing observation that recolonization with germs from patients with PD worsens physical limitations.
5. Biomarkers: A Novel Diagnostic Approach for AD
The updated version of the 2011 guidelines by National Institute of Aging and Alzheimer’s in association with the Alzheimer’s Association, has introduced a new research framework for AD. This framework is based on observational and international research and provides separate diagnostic recommendation for the preclinical, mild congnitive impairment, and dementia stages of AD. The focus of this framwowk is on the use of biomarkers rather than solely relying on clinical symptoms for the diagnosis of AD in living individuals.
The AD associated biomarkers are categorized into three groups: (a) β Amyloid protein, b) Pathologiacal Tau protein, and C) Neurogeneration [AT(N)]. It is important to note that this classification may evolve as new biomarkers are discovered. Insome cases, biomarkers may not be available or their application may hinder specific research objectives (Jack et al., 2018).
The discovery of reliable biomarkwers for early detection and assessment of AD severity remains a critical need. It is essential to avoid considering biomarker-based research as a onesize fits all approach for studing age ralted cognitive impairment and dementia. Instead, the utilization of biomarkers should be based on their compatibility with the specific research objectives of each study.
A Recent study by Intili et al. (2023) has demonstrated a link betwwen gut dysbiosis and changes in the intestinal epithelium, which may contribute to the development of degenerative diseases in the brain. Furthermore, there is a growing intrest in exploring the potential of certain bacterial strains as biomarkers for AD, These newly discovered biomarkers, as outlined in Table 3, offer the possibility of transforming the diagnostic landscape by proving cost effective and accessible screening methods. Integrating these biomarkers into diagnostic protocol has the potential to detect AD at an early stage, allowing for more effective intervention and treatment. The affordability and accessibility of these biomarkers could have a significant impact on public health by facilitating widespread screening initiatives and promoting early intervention strategies. Ongling research aims to explore these biomarkers in biomarkers in diverse populations and incorporate them into routine clinical practice, potentially reshaping the approach to AD diagnosis and management.
AD, Alzheimer disease; MCI, mild cognitive impaired; aMCI, amnestic mild cognitive impairment; SMC, significant memory concern; EMCI, early mild cognitive impairment; LMCI, late mild cognitive impairment; NC, normal controls; CN, cognitively normal; CN−, Aβ-negative cognitively normal; CN+, Aβ-positive cognitively normal; CI, cognitive impairment; GM, gut microbiome; CSF, cerebrospinal fluid; GC, gas chromatography; LC, liquid chromatography; MS, mass spectrometry; BA, Bile acid; LPS, Lipopolysaccharides; SCFAs, short chain fatty acids; TMAO, Trimethylamine N-oxide; ADNI, Alzheimer’s Disease Neuroimaging Initiative; ADMC, Alzheimer Disease Metabolomics Consortium; Aβ, amyloid-β; CA, cholic acid; CDCA, chenodeoxycholic acid; GCDCA, glycochenodeoxycholic acid; GDCA, glycodeoxycholic acid; GLCA, glycolithocholic acid; TLCA, taurolithocholic acid; CCK, Cholecystokinin; DAO, diaminooxidase; sCD14, soluble CD 14; 16SrDNA, 16Sribosomal DNA; 16SrRNA, 16Sribosomal RNA; ELISA, Enzyme linked immunosorbent assays; CLIA, chemiluminescence immunoassay; DHEAS, Dehydroepiandrosterone sulfate; MSD, meso scale discovery; USA, United States of America; p-tau, phosphorylated tau.
6. Targeted Therapy for AD: Amelioration of Microbiota
A major concern of antibiotic use is the long-term alteration of the normal healthy GM and horizontal transfer of resistance genes that could result in a reservoir of organisms with a multidrug-resistant gene pool.
GM regulation is fundamental for the well-being of human health; its deregulation in AD patients is established. For the maintenance of healthy GM, diet is an essential factor. The onset of AD and dietary pattern is linked as proposed by many studies, conferring diet as an adaptable risk element [140];[141]. The contributing factors suggested in the pathogenesis of AD reflecting dysbiosis are poor dietary habits and inflammatory response. Well preserved cognitive functions with anti-inflammatory properties have been associated with Mediterranean diet, which is characterized by consumption of mainly vegetables, fruits, cereals and lean meat with reduced high fat diet intake [142]. The MedDiet is endowed with fibers, minerals, and vitamins (especially B complex) that influence the microbiota. Ghosh et al.'s study of the effect of one year MedDiet on the GM in 612 non-frail subjects from five European countries demonstrated alteration in gut microbiome in elderly subjects with an improvement in their health status [143]. Similar studies by Hoscheidt et al provided evidence that the MedDiet encourages metabolic and brain health during mid-life as compared to western diet. The diet response during middle age is important to take into account as this phase may involve early pathophysiological processes that increase AD risk [144].
Another dietary pattern supporting the neuro-protective behavior is believed to be the KD. It is a very high-fat, low-carbohydrate diet that brings the body into a state of ketosis by implicating a fasting-like effect [145]. Neuronal loss by apoptosis is reduced by low serum glucose and high serum fat levels. This diet attunes the upregulation of hippocampal genes coding for mitochondrial enzymes, thus providing alternative energy substrates [146]. Moreover, KD reduces oxidative stress, inflmmatory response and encourages neurotrophic factors besides raising the GABA-to-glutamate ratio and activation of ATP-sensitive potassium channels. Such changes contribute to the improvement of brain metabolism and KD is gaining attention as a probable therapy for neurodegenerative disorders like AD [147]. Ketones have been considered as endogenous factors protecting AD. The neuroprotective mechanism of ketones in AD is attributed to the blockage of intracellular Aβ -42 buildups in mouse models [148]. The involvement of Aβ-42 in the pathogenesis of AD has paved the way to screen those formulations, which will selectively reduce the Aβ 42 either by reducing its formation or increasing clearance without affecting the normal protein processing and formation.
Early interventions with KD can increase useful microbiota, and improve brain vascular functions besides, intermittent fasting (IF) reduces insulin resistance neuroinflammation and increases metabolic regulation and Brain-derived Neurotrophic Factor (BDNF) levels [149]. Dietary interventions are usually more beneficial and safer than pharmaceutical therapies due to their economical and easy-handling properties.
6.1. Probiotics
Probiotics are a combination of live microorganisms (bacteria and yeast), inhabitants of our body, used to improve the healthy microbiota, thereby improving the life of the host when ingested in adequate amounts [150]. They have shown wider effects on human health, correcting immunity, hormone functions, metabolic functions, regulating pH levels, preserving the integrity of intestinal lining, delaying aging and enriching the brain-derived neurotrophic factor [151]. These neurotrophic factors are protein factors that find important role in neurology by making the survival and differentiation of neurons possible. The most fascinating effect of probiotics on the host is in improving brain health, behavior and functioning. The clinical trials have demonstrated restoration of GM, restore the integrity of BBB, diminished oxidative stress, improved cognitive decline, lowered neuroinflammation and improved insulin sensitivity by supplementing microbiota. The probiotics used to rectify the misbalance of the microbiota-regulated gut-brain axis are termed psychobiotics [152]. Probiotic alteration influences the onset of AD due to the deregulation of microbial picture. This imbalance in AD has shown a reduction in Verrucomicrobia, Proteobacteria, Actinobacteria, Firmicutes, Bifidobacterium bacteria and increase in Bacteroides and Tenericutes. It is speculated that change in GM can cause colonization of internal pathogens leading to increased gut permeability. A study of Wu et al 2017 on Drosophila AD model established that enterobacteria infection aggravates AD progression by influencing immune hemocyte recruitment to the brain[153]. There is evidence supporting probiotic administration with its superior effects on gut brain axis. In a randomized controlled trial, a mixture of probiotic consumption in health volunteers for 4 weeks significantly lowered sensitivity to sad mood [154]. Benton et al 2007 reported improved mood upon yogurt consumption for 3 weeks in healthy subjects [155]. Probiotics also exhibit immunological benefits that seem to be strain-specific, while strain specificity is well recognized in AD. Therefore all probiotics do not have psychobiotic potential, unlike few Bifidobacterium and Lactobacillus is frequently used as psychobiotics[156] . More research is needed to screen the effective probiotic strains in the management of AD.
6.2. Prebiotics
Prebiotics are nutrients (plant fibers) degraded by GM in the lower part of the GI tract. Intestinal bacteria feed on them and the degradation products are short-chain fatty acids. The short-chain fatty acids (SCFA) enter the systemic circulation and can affect other organs besides GI tract. Fructo and galacto-oligosaccharides are notable prebiotics used alone or in combination with probiotics that have beneficial effects on human health [157]. Prebiotics act as substrates and are fermented by gut bacteria like Bifidobacteria and Lactobacill, increasing anti-inflammatory metabolite machinery. Evidence suggests prebiotics' role in improving neurological disorders in rat models by reducing the raised cortisol response and altered sentimental bias in normal volunteers[158]. Amelioration of Cognitive deficits and neurodegeneration in AD transgenic mice model was reported by the administration of fructooligosaccharides (FOS). There was an upregulation of synapsin 1, PSD-95 with a reduced phosphorylated level of JNK, reduced GLP-1R and increased levels of GLP-1, signifying the favorable effects of FOS against AD by governing the gut microbiota-GLP-1/GLP-1R pathways [159]. Recently, lactulose, commonly used as a food additive given to AD mice, utilized by GM, has shown neuroprotective effects and increased the levels of the autophagic pathways with improvement in synaptic protein expression[160]. Sodium oligomannate (GV-971) derived from marine sources was tested in a 24-week double blind, randomized, placebo-controlled phase II clinical trial in China. There was a cognitive improvement as attributed to the decline of ADAS-cog12 (Alzheimer's Disease Assessment Scale-cognitive subscale 12-item) and improvement in ADCS-ADL (Alzheimer's Disease Cooperative Study – Activities of Daily Living) and NPI (neuropsychiatric inventory) scales [161]. The study shows GV-971 as a novel agent that may enhance cognition in AD by reconstituting the dysbiosis of GM. Overall, the literature suggests personalized prebiotic administration, which acts by modulation of GM, may speak for a new therapeutic approach for AD.
6.3. Fecal Microbiota Transplantation (FMT)
FMT is one of the therapeutic procedures in the alleviation of AD with minor precautions, as the entry of live microorganisms or metabolites is to be taken care of. Sun et al. (2019) used the AAPswe/PS1dE9 Tg mouse model to testify to the action of FMT for the treatment of AD. GM and SCFA analyzed by 16S rRNA sequencing and NMR depicted improved cognitive deficits, reduced Aβ deposition, and increased synaptic plasticity and reduced neuroinflammation. A study on 82 years AD patients with recurrent Clostridium difficile infection intervened by FMT infusion showed improvement in symptoms of AD[162]. Zhan et al 2018 observed antibiotic treatment diminished cognitive function in wild mice and fecal intake after improving memory and spatial learning[163]. These studies suggest cognitive decline is associated with antibiotics, which can be reversed with FMT. Conversely, few studies suggest a reduction in Aβ deposition after antibiotic administration. When administered with antibiotic cocktail, Dodiya et al., 2019, observed disturbed microbiome with lowered Aβ plaque pathology and astrogliosis in male mice [164]. Hence FMT may act as a potential therapeutics for AD and other neuroinflammatory conditions. However, the available literature is still meager and conflicting studies are to be taken into account. Nevertheless, further studies involving double blinded randomized controlled trials are to be carried out to further cross-validate the action of FMT in neurodegenerative disorders.
7. Conclusions
The gut-brain axis plays a significant role in regulating the body's physiological processes, including food intake, glucose and fat metabolism, insulin secretion, and responsiveness. The gut microbiota (GM) communicates with the brain via biochemical signaling, and GM dysbiosis can impact the digestive system and the functioning of the central nervous system (CNS). Furthermore, GM dysbiosis can lead to the generation of mediators of pro-inflammatory connected to the initiation of AD. Maintaining the normal integrity of the BBB is crucial in preventing harmful substances released from the GM from entering the CNS and causing damage.
The reduction in the activity of choline acetyl-transferase in the cerebral cortex and hippocampus is the most consistent molecular insult that leads to AD changes. Glial cells are also found to play an important role in reacting with tau and amyloid pathology in AD patients. The presence of plaques and tangles in the brain meets the histopathologic diagnostic standards for AD in 90% of AD-diagnosed cases, as indicated by anatomo-clinical studies. However, some non-demented elderly patients may also qualify for a pathologic diagnosis of AD. These findings highlight the complex and multifactorial nature of AD, with implications for developing more effective diagnostic and therapeutic strategies. There is strong evidence supporting the role of Aβ in contributing to AD. Aβ multimeric aggregates are suspected of contributing to the disease, forming long fibrils and aggregates known as plaques, the neuropathological hallmark of AD. The amyloid cascade hypothesis plays an important role in explaining the pathogenesis of AD, Mutations associated with the familial type of AD increase Aβ production or modify its production rate. Aβ has diverse toxic mechanisms that contribute to its toxicity, including oxidative stress, mitochondrial diffusion, membrane permeability, inflammation, synaptic changes, and excite-toxicity. β-sheet-rich fibrils formed by aggregation of synthetic Aβ have been hypothesized to cause neurodegeneration in AD, and soluble Aβ oligomers are more cytotoxic than insoluble fibrillary aggregates. Overall, the evidence supports the contribution of Aβ to AD pathogenesis, and targeting Aβ production or accumulation is a promising approach for treating AD. Furthermore, research is needed for better understanding of the signaling pathways and underlying mechanisms that will open a way for identification of new targets and holistic intervention of AD.
Alternative conclusion: The gut-brain axis significantly regulates physiological processes, and GM dysbiosis can impact CNS functioning, contributing to pro-inflammatory mediators associated with AD initiation. Maintaining the BBB's normal integrity is crucial in preventing harmful substances from the GM entering the CNS. Molecular insults, such as reduced choline acetyl-transferase activity and glial cell reactions, play pivotal roles in AD pathogenesis. Plaques and tangles meet diagnostic standards in most AD cases, emphasizing the complex and multifactorial nature of AD. Strong evidence supports β-amyloid's contribution to AD, with diverse toxic mechanisms. Targeting Aβ production or accumulation shows promise in AD treatment. However, further research is crucial for a holistic understanding and identification of new therapeutic targets for AD intervention.
Conflicts of Interest
The authors declare no competing interests
References
- Scheff, S.W.; Price, D.A. Alzheimer’s disease-related alterations in synaptic density: Neocortex and hippocampus. J. Alzheimer’s Dis. 2006, 9, 101–115. [Google Scholar] [CrossRef] [PubMed]
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209. [Google Scholar] [PubMed]
- Beopoulos, A.; Gea, M.; Fasano, A.; Iris, F. Autonomic Nervous System Neuroanatomical Alterations Could Provoke and Maintain Gastrointestinal Dysbiosis in Autism Spectrum Disorder (ASD): A Novel Microbiome-Host Interaction Mechanistic Hypothesis. Nutrients 2021, 14. [Google Scholar] [CrossRef] [PubMed]
- Breit, S.; Kupferberg, A.; Rogler, G.; Hasler, G. Vagus Nerve as Modulator of the Brain–Gut Axis in Psychiatric and Inflammatory Disorders. Front. Psychiatry 2018, 9. Available from: https://www.frontiersin.org/articles/10.3389/fpsyt.2018.00044. [CrossRef] [PubMed]
- Thakur, A.K.; Shakya, A.; Husain, G.M.; Emerald, M.; Kumar, V. Gut-Microbiota and Mental Health: Current and Future Perspectives. J. Pharmacol. Clin. Toxicol. 2014, 2, 1016. [Google Scholar]
- Jiang, C.; Li, G.; Huang, P.; Liu, Z.; Zhao, B. The Gut Microbiota and Alzheimer’s Disease. J. Alzheimers. Dis. 2017, 58, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Karaman, R. Comprehensive Review on Alzheimer’s Disease: Causes and Treatment. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- González, A.; Singh, S.K.; Churruca, M.; Maccioni, R.B. Alzheimer’s Disease and Tau Self-Assembly: In the Search of the Missing Link. Int. J. Mol. Sci. 2022, 23. Available from: https://www.mdpi.com/1422-0067/23/8/. [CrossRef]
- Lloret, A.; Fuchsberger, T.; Giraldo, E.; Viña, J. Molecular mechanisms linking amyloid beta toxicity and Tau hyperphosphorylation in Alzheimer’s Disease. Free Radic. Biol. Med. 2015, 83. [Google Scholar] [CrossRef]
- Rajmohan, R.; Reddy, P.H. Amyloid-Beta and Phosphorylated Tau Accumulations Cause Abnormalities at Synapses of Alzheimer’s disease Neurons. J. Alzheimers. Dis. 2017, 57, 975–999. [Google Scholar] [CrossRef]
- Zlokovic, B. V The blood-brain barrier in health and chronic neurodegenerative disorders. Neuron 2008, 57, 178–201. [Google Scholar] [CrossRef] [PubMed]
- Ballan, R.; Battistini, C.; Xavier-Santos, D.; Saad, S.M.I. Interactions of probiotics and prebiotics with the gut microbiota. Prog. Mol. Biol. Transl. Sci. 2020, 171, 265–300. [Google Scholar] [PubMed]
- Megur, A.; Baltriukienė, D.; Bukelskienė, V.; Burokas, A. The Microbiota-Gut-Brain Axis and Alzheimer’s Disease: Neuroinflammation Is to Blame? Nutrients 2020, 13. [Google Scholar] [CrossRef]
- Salminen, S.; Bouley, C.; Boutron-Ruault, M.C.; Cummings, J.H.; Franck, A.; Gibson, G.R.; Isolauri, E.; Moreau, M.C.; Roberfroid, M.; Rowland, I. Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 1998, 80 Suppl 1, S147–71. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, T.S.B.; Raes, J.; Bork, P. The Human Gut Microbiome: From Association to Modulation. Cell 2018, 172, 1198–1215. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? A changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Nandwana, V.; Debbarma, S. Fecal Microbiota Transplantation: A Microbiome Modulation Technique for Alzheimer’s Disease. Cureus 2021, 13. [Google Scholar] [CrossRef]
- Mancuso, C.; Santangelo, R. Alzheimer’s disease and gut microbiota modifications: the long way between preclinical studies and clinical evidence. Pharmacol. Res. 2018, 129, 329–336. [Google Scholar] [CrossRef]
- Luca, M.; Di Mauro, M.; Di Mauro, M.; Luca, A. Gut Microbiota in Alzheimer’s Disease, Depression, and Type 2 Diabetes Mellitus: The Role of Oxidative Stress. Oxid. Med. Cell. Longev. 2019, 2019, 4730539. [Google Scholar] [CrossRef]
- Zhu, X.; Han, Y.; Du, J.; Liu, R.; Jin, K.; Yi, W. Microbiota-gut-brain axis and the central nervous system. Oncotarget 2017, 8, 53829–53838. [Google Scholar] [CrossRef]
- Long, J.M.; Holtzman, D.M. Alzheimer Disease: An Update on Pathobiology and Treatment Strategies. Cell 2019, 179, 312–339. [Google Scholar] [CrossRef] [PubMed]
- Durgan, D.J.; Lee, J.; McCullough, L.D.; Bryan, R.M.J. Examining the Role of the Microbiota-Gut-Brain Axis in Stroke. Stroke 2019, 50, 2270–2277. [Google Scholar] [CrossRef] [PubMed]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar Reddy, D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Desbonnet, L.; Garrett, L.; Clarke, G.; Kiely, B.; Cryan, J.F.; Dinan, T.G. Effects of the probiotic Bifidobacterium infantis in the maternal separation model of depression. Neuroscience 2010, 170, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- van de Wouw, M.; Boehme, M.; Lyte, J.M.; Wiley, N.; Strain, C.; O’Sullivan, O.; Clarke, G.; Stanton, C.; Dinan, T.G.; Cryan, J.F. Short-chain fatty acids: microbial metabolites that alleviate stress-induced brain-gut axis alterations. J. Physiol. 2018, 596, 4923–4944. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.; Loreth, D.; Pöttker, B.; Hefti, K.; Innos, J.; Schwald, K.; Hengstler, H.; Menzel, L.; Sommer, C.J.; Radyushkin, K.; et al. Neuronal Growth and Behavioral Alterations in Mice Deficient for the Psychiatric Disease-Associated Negr1 Gene. Front. Mol. Neurosci. 2018, 11, 30. [Google Scholar] [CrossRef] [PubMed]
- Webster, S.J.; Bachstetter, A.D.; Nelson, P.T.; Schmitt, F.A.; Van Eldik, L.J. Using mice to model Alzheimer’s dementia: an overview of the clinical disease and the preclinical behavioral changes in 10 mouse models. Front. Genet. 2014, 5, 88. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.-P.; Arai, T.; Miklossy, J.; McGeer, P.L. Abeta and tau form soluble complexes that may promote self aggregation of both into the insoluble forms observed in Alzheimer’s disease. Proc. Natl. Acad. Sci. U. S. A. 2006, 103, 1953–1958. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Repáraz, J.; Kasper, L.H. The Microbiome and Neurologic Disease: Past and Future of a 2-Way Interaction. Neurother. J. Am. Soc. Exp. Neurother. 2018, 15, 1–4. [Google Scholar] [CrossRef]
- Colpitts, S.L.; Kasper, E.J.; Keever, A.; Liljenberg, C.; Kirby, T.; Magori, K.; Kasper, L.H.; Ochoa-Repáraz, J. A bidirectional association between the gut microbiota and CNS disease in a biphasic murine model of multiple sclerosis. Gut Microbes 2017, 8, 561–573. [Google Scholar] [CrossRef]
- Mehrabadi, S.; Sadr, S.S. Assessment of Probiotics Mixture on Memory Function, Inflammation Markers, and Oxidative Stress in an Alzheimer’s Disease Model of Rats. Iran. Biomed. J. 2020, 24, 220–228. [Google Scholar] [CrossRef]
- Chang, C.-H.; Lin, C.-H.; Lane, H.-Y. d-glutamate and Gut Microbiota in Alzheimer’s Disease. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Gareau, M.G.; Wine, E.; Rodrigues, D.M.; Cho, J.H.; Whary, M.T.; Philpott, D.J.; MacQueen, G.; Sherman, P.M. Bacterial infection causes stress-induced memory dysfunction in mice. Gut 2011, 60, 307–317. [Google Scholar] [CrossRef]
- Singhrao, S.K.; Harding, A.; Poole, S.; Kesavalu, L.; Crean, S. Porphyromonas gingivalis Periodontal Infection and Its Putative Links with Alzheimer’s Disease. Mediators Inflamm. 2015, 2015, 137357. [Google Scholar] [CrossRef]
- Cattaneo, A.; Cattane, N.; Galluzzi, S.; Provasi, S.; Lopizzo, N.; Festari, C.; Ferrari, C.; Guerra, U.P.; Paghera, B.; Muscio, C.; et al. Association of brain amyloidosis with pro-inflammatory gut bacterial taxa and peripheral inflammation markers in cognitively impaired elderly. Neurobiol. Aging 2017, 49, 60–68. [Google Scholar] [CrossRef]
- Farzi, A.; Fröhlich, E.E.; Holzer, P. Gut Microbiota and the Neuroendocrine System. Neurother. J. Am. Soc. Exp. Neurother. 2018, 15, 5–22. [Google Scholar] [CrossRef]
- Baj, A.; Moro, E.; Bistoletti, M.; Orlandi, V.; Crema, F.; Giaroni, C. Glutamatergic Signaling Along The Microbiota-Gut-Brain Axis. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef]
- Jameson, K.G.; Hsiao, E.Y. Linking the Gut Microbiota to a Brain Neurotransmitter. Trends Neurosci. 2018, 41, 413–414. [Google Scholar] [CrossRef]
- Franceschi, F.; Ojetti, V.; Candelli, M.; Covino, M.; Cardone, S.; Potenza, A.; Simeoni, B.; Gabrielli, M.; Sabia, L.; Gasbarrini, G.; et al. Microbes and Alzheimer’ disease: lessons from H. pylori and GUT microbiota. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 426–430. [Google Scholar] [CrossRef]
- Naveed, M.; Mubeen, S.; Khan, A.; Ibrahim, S.; Meer, B. Plasma Biomarkers: Potent Screeners of Alzheimer’s Disease. Am. J. Alzheimers. Dis. Other Demen. 2019, 34, 290–301. [Google Scholar] [CrossRef]
- Sun, Y.; Sommerville, N.R.; Liu, J.Y.H.; Ngan, M.P.; Poon, D.; Ponomarev, E.D.; Lu, Z.; Kung, J.S.C.; Rudd, J.A. Intra-gastrointestinal amyloid-β1-42 oligomers perturb enteric function and induce Alzheimer’s disease pathology. J. Physiol. 2020, 598, 4209–4223. [Google Scholar] [CrossRef]
- Cerovic, M.; Forloni, G.; Balducci, C. Neuroinflammation and the Gut Microbiota: Possible Alternative Therapeutic Targets to Counteract Alzheimer’s Disease? Front. Aging Neurosci. 2019, 11, 284. [Google Scholar] [CrossRef]
- Deng, H.; Yang, S.; Zhang, Y.; Qian, K.; Zhang, Z.; Liu, Y.; Wang, Y.; Bai, Y.; Fan, H.; Zhao, X.; et al. Bacteroides fragilis Prevents Clostridium difficile Infection in a Mouse Model by Restoring Gut Barrier and Microbiome Regulation. Front. Microbiol. 2018, 9, 2976. [Google Scholar] [CrossRef]
- Tanida, M.; Yamano, T.; Maeda, K.; Okumura, N.; Fukushima, Y.; Nagai, K. Effects of intraduodenal injection of Lactobacillus johnsonii La1 on renal sympathetic nerve activity and blood pressure in urethane-anesthetized rats. Neurosci. Lett. 2005, 389, 109–114. [Google Scholar] [CrossRef]
- Yang, X.; Yu, D.; Xue, L.; Li, H.; Du, J. Probiotics modulate the microbiota-gut-brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm. Sin. B 2020, 10, 475–487. [Google Scholar] [CrossRef]
- Sigal, M.; Meyer, T.F. Coevolution between the Human Microbiota and the Epithelial Immune System. Dig. Dis. 2016, 34, 190–193. [Google Scholar] [CrossRef]
- Schluter, J.; Peled, J.U.; Taylor, B.P.; Markey, K.A.; Smith, M.; Taur, Y.; Niehus, R.; Staffas, A.; Dai, A.; Fontana, E.; et al. The gut microbiota is associated with immune cell dynamics in humans. Nature 2020, 588, 303–307. [Google Scholar] [CrossRef]
- Khosravi, A.; Yáñez, A.; Price, J.G.; Chow, A.; Merad, M.; Goodridge, H.S.; Mazmanian, S.K. Gut microbiota promote hematopoiesis to control bacterial infection. Cell Host Microbe 2014, 15, 374–381. [Google Scholar] [CrossRef]
- Kierdorf, K.; Prinz, M. Factors regulating microglia activation. Front. Cell. Neurosci. 2013, 7. Available from: https://www.frontiersin.org/articles/. [CrossRef]
- Jouanne, M.; Rault, S.; Voisin-Chiret, A.-S. Tau protein aggregation in Alzheimer’s disease: An attractive target for the development of novel therapeutic agents. Eur. J. Med. Chem. 2017, 139, 153–167. [Google Scholar] [CrossRef]
- Köhler, C.A.; Maes, M.; Slyepchenko, A.; Berk, M.; Solmi, M.; Lanctôt, K.L.; Carvalho, A.F. The Gut-Brain Axis, Including the Microbiome, Leaky Gut and Bacterial Translocation: Mechanisms and Pathophysiological Role in Alzheimer’s Disease. Curr. Pharm. Des. 2016, 22, 6152–6166. [Google Scholar] [CrossRef] [PubMed]
- Goedert, M.; Eisenberg, D.S.; Crowther, R.A. Propagation of Tau Aggregates and Neurodegeneration. Annu. Rev. Neurosci. 2017, 40, 189–210. Available from: https://www.frontiersin.org/articles/. [CrossRef]
- Avila, J.; Jiménez, J.S.; Sayas, C.L.; Bolós, M.; Zabala, J.C.; Rivas, G.; Hernández, F. Tau Structures. Front. Aging Neurosci. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Alonso, A. del C.; Grundke-Iqbal, I. Cytosolic abnormally hyperphosphorylated tau but not paired helical filaments sequester normal MAPs and inhibit microtubule assembly. J. Alzheimers. Dis. 2008, 14, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Lott, I.T.; Dierssen, M. Cognitive deficits and associated neurological complications in individuals with Down’s syndrome. Lancet. Neurol. 2010, 9, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Hoover, B.R.; Reed, M.N.; Su, J.; Penrod, R.D.; Kotilinek, L.A.; Grant, M.K.; Pitstick, R.; Carlson, G.A.; Lanier, L.M.; Yuan, L.-L.; et al. Tau mislocalization to dendritic spines mediates synaptic dysfunction independently of neurodegeneration. Neuron 2010, 68, 1067–1081. [Google Scholar] [CrossRef] [PubMed]
- Sato, N.; Takeda, S.; Uchio-Yamada, K.; Ueda, H.; Fujisawa, T.; Rakugi, H.; Morishita, R. Role of insulin signaling in the interaction between Alzheimer disease and diabetes mellitus: a missing link to therapeutic potential. Curr. Aging Sci. 2011, 4, 118–127. [Google Scholar] [CrossRef] [PubMed]
- Takeda, S.; Sato, N.; Rakugi, H.; Morishita, R. Molecular mechanisms linking diabetes mellitus and Alzheimer disease: beta-amyloid peptide, insulin signaling, and neuronal function. Mol. Biosyst. 2011, 7, 1822–1827. [Google Scholar] [CrossRef]
- Rayasam, G.V.; Tulasi, V.K.; Sodhi, R.; Davis, J.A.; Ray, A. Glycogen synthase kinase 3: more than a namesake. Br. J. Pharmacol. 2009, 156, 885–898. [Google Scholar] [CrossRef]
- Macfarlane, G.T.; Macfarlane, S. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 2012, 95, 50–60. [Google Scholar] [CrossRef]
- Xu, R.; Wang, Q. Towards understanding brain-gut-microbiome connections in Alzheimer’s disease. BMC Syst. Biol. 2016, 10, 63. [Google Scholar] [CrossRef]
- Lyman, M.; Lloyd, D.G.; Ji, X.; Vizcaychipi, M.P.; Ma, D. Neuroinflammation: the role and consequences. Neurosci. Res. 2014, 79, 1–12. [Google Scholar] [CrossRef]
- Morales, I.; Guzmán-Martínez, L.; Cerda-Troncoso, C.; Farías, G.A.; Maccioni, R.B. Neuroinflammation in the pathogenesis of Alzheimer’s disease. A rational framework for the search of novel therapeutic approaches. Front. Cell. Neurosci. 2014, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Avila, J.; Jiménez, J.S.; Sayas, C.L.; Bolós, M.; Zabala, J.C.; Rivas, G.; Hernández, F. Tau Structures. Front. Aging Neurosci. 2016; 8. Available from: https://www.frontiersin.org/articles/10.3389/fnagi.2016.00262.
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, prebiotics and immunomodulation of gut mucosal defences: homeostasis and immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef] [PubMed]
- Savignac, H.M.; Couch, Y.; Stratford, M.; Bannerman, D.M.; Tzortzis, G.; Anthony, D.C.; Burnet, P.W.J. Prebiotic administration normalizes lipopolysaccharide (LPS)-induced anxiety and cortical 5-HT2A receptor and IL1-β levels in male mice. Brain. Behav. Immun. 2016, 52, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Clinton, L.K.; Blurton-Jones, M.; Myczek, K.; Trojanowski, J.Q.; LaFerla, F.M. Synergistic Interactions between Abeta, tau, and alpha-synuclein: acceleration of neuropathology and cognitive decline. J. Neurosci. Off. J. Soc. Neurosci. 2010, 30, 7281–7289. [Google Scholar] [CrossRef] [PubMed]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lukiw, W.J. Microbiome-generated amyloid and potential impact on amyloidogenesis in Alzheimer’s disease (AD). J. Nat. Sci. 2015, 1. [Google Scholar]
- Dinan, T.G.; Cryan, J.F. Gut instincts: microbiota as a key regulator of brain development, ageing and neurodegeneration. J. Physiol. 2017, 595, 489–503. [Google Scholar] [CrossRef]
- Mary, A.; Dayan, J.; Leone, G.; Postel, C.; Fraisse, F.; Malle, C.; Vallée, T.; Klein-Peschanski, C.; Viader, F.; De la Sayette, V. Resilience after trauma: The role of memory suppression. Science (80-. ). 2020, 367, eaay8477. [Google Scholar] [CrossRef]
- Ubeda, C.; Djukovic, A.; Isaac, S. Roles of the intestinal microbiota in pathogen protection. Clin. Transl. Immunol. 2017, 6, e128. [Google Scholar] [CrossRef] [PubMed]
- Zeng, M.Y.; Cisalpino, D.; Varadarajan, S.; Hellman, J.; Warren, H.S.; Cascalho, M.; Inohara, N.; Núñez, G. Gut microbiota-induced immunoglobulin G controls systemic infection by symbiotic bacteria and pathogens. Immunity 2016, 44, 647–658. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Microbiota-brain-gut axis and neurodegenerative diseases. Curr. Neurol. Neurosci. Rep. 2017, 17, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harach, T.; Marungruang, N.; Duthilleul, N.; Cheatham, V.; Mc Coy, K.D.; Frisoni, G.; Neher, J.J.; Fåk, F.; Jucker, M.; Lasser, T. Reduction of Abeta amyloid pathology in APPPS1 transgenic mice in the absence of gut microbiota. Sci. Rep. 2017, 7, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Minter, M.R.; Zhang, C.; Leone, V.; Ringus, D.L.; Zhang, X.; Oyler-Castrillo, P.; Musch, M.W.; Liao, F.; Ward, J.F.; Holtzman, D.M. Antibiotic-induced perturbations in gut microbial diversity influences neuro-inflammation and amyloidosis in a murine model of Alzheimer’s disease. Sci. Rep. 2016, 6, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Main, B.S.; Minter, M.R. Microbial immuno-communication in neurodegenerative diseases. Front. Neurosci. 2017, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Fröhlich, E.E.; Farzi, A.; Mayerhofer, R.; Reichmann, F.; Jačan, A.; Wagner, B.; Zinser, E.; Bordag, N.; Magnes, C.; Fröhlich, E. Cognitive impairment by antibiotic-induced gut dysbiosis: analysis of gut microbiota-brain communication. Brain. Behav. Immun. 2016, 56, 140–155. [Google Scholar] [CrossRef] [PubMed]
- Nie, H.; Song, C.; Wang, D.; Cui, S.; Ren, T.; Cao, Z.; Liu, Q.; Chen, Z.; Chen, X.; Zhou, Y. MicroRNA-194 inhibition improves dietary-induced non-alcoholic fatty liver disease in mice through targeting on FXR. Biochim. Biophys. Acta (BBA)-Molecular Basis Dis. 2017, 1863, 3087–3094. [Google Scholar] [CrossRef] [PubMed]
- Montagne, A.; Barnes, S.R.; Sweeney, M.D.; Halliday, M.R.; Sagare, A.P.; Zhao, Z.; Toga, A.W.; Jacobs, R.E.; Liu, C.Y.; Amezcua, L.; et al. Blood-Brain Barrier Breakdown in the Aging Human Hippocampus. Neuron 2015, 85, 296–302. [Google Scholar] [CrossRef]
- Welling, M.M.; Nabuurs, R.J.A.; van der Weerd, L. Potential role of antimicrobial peptides in the early onset of Alzheimer’s disease. Alzheimer’s Dement. 2015, 11, 51–57. [Google Scholar] [CrossRef]
- Halliday, M.R.; Rege, S. V; Ma, Q.; Zhao, Z.; Miller, C.A.; Winkler, E.A.; Zlokovic, B. V Accelerated pericyte degeneration and blood–brain barrier breakdown in apolipoprotein E4 carriers with Alzheimer’s disease. J. Cereb. Blood Flow Metab. 2016, 36, 216–227. [Google Scholar] [CrossRef] [PubMed]
- O’Mahony, S.M.; Marchesi, J.R.; Scully, P.; Codling, C.; Ceolho, A.-M.; Quigley, E.M.M.; Cryan, J.F.; Dinan, T.G. Early life stress alters behavior, immunity, and microbiota in rats: implications for irritable bowel syndrome and psychiatric illnesses. Biol. Psychiatry 2009, 65, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Sudo, N. Stress and gut microbiota: does postnatal microbial colonization program the hypothalamic–pituitary–adrenal system for stress response. Int Congr Ser 2006, 1287, 350–354. [Google Scholar] [CrossRef]
- Bailey, M.T.; Lubach, G.R.; Coe, C.L. Prenatal stress alters bacterial colonization of the gut in infant monkeys. J. Pediatr. Gastroenterol. Nutr. 2004, 38, 414–421. [Google Scholar] [PubMed]
- Munyaka, P.M.; Eissa, N.; Bernstein, C.N.; Khafipour, E.; Ghia, J.-E. Antepartum antibiotic treatment increases offspring susceptibility to experimental colitis: a role of the gut microbiota. PLoS One 2015, 10, e0142536. [Google Scholar] [CrossRef] [PubMed]
- Armulik, A.; Genové, G.; Mäe, M.; Nisancioglu, M.H.; Wallgard, E.; Niaudet, C.; He, L.; Norlin, J.; Lindblom, P.; Strittmatter, K. Pericytes regulate the blood–brain barrier. Nature 2010, 468, 557–561. [Google Scholar] [CrossRef] [PubMed]
- Daneman, R.; Zhou, L.; Kebede, A.A.; Barres, B.A. Pericytes are required for blood–brain barrier integrity during embryogenesis. Nature 2010, 468, 562–566. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.D.; Winkler, E.A.; Sagare, A.P.; Singh, I.; LaRue, B.; Deane, R.; Zlokovic, B. V Pericytes control key neurovascular functions and neuronal phenotype in the adult brain and during brain aging. Neuron 2010, 68, 409–427. [Google Scholar] [CrossRef]
- Sengillo, J.D.; Winkler, E.A.; Walker, C.T.; Sullivan, J.S.; Johnson, M.; Zlokovic, B. V Deficiency in Mural Vascular Cells Coincides with Blood–Brain Barrier Disruption in A lzheimer’s Disease. Brain Pathol. 2013, 23, 303–310. [Google Scholar] [CrossRef]
- Blennow, K.; Wallin, A.; Fredman, P.; Karlsson, I.; Gottfries, C.G.; Svennerholm, L. Blood-brain barrier disturbance in patients with Alzheimer’s disease is related to vascular factors. Acta Neurol. Scand. 1990, 81, 323–326. [Google Scholar] [CrossRef]
- Fiala, M.; Liu, Q.N.; Sayre, J.; Pop, V.; Brahmandam, V.; Graves, M.C.; Vinters, H. V Cyclooxygenase-2-positive macrophages infiltrate the Alzheimer’s disease brain and damage the blood–brain barrier. Eur. J. Clin. Invest. 2002, 32, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Alata, W.; Ye, Y.; St-Amour, I.; Vandal, M.; Calon, F. Human apolipoprotein e ε4 expression impairs cerebral vascularization and blood—brain barrier function in mice. J. Cereb. Blood Flow Metab. 2015, 35, 86–94. [Google Scholar] [CrossRef] [PubMed]
- Verghese, P.B.; Castellano, J.M.; Holtzman, D.M. Apolipoprotein E in Alzheimer’s disease and other neurological disorders. Lancet Neurol. 2011, 10, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Nishitsuji, K.; Hosono, T.; Nakamura, T.; Bu, G.; Michikawa, M. Apolipoprotein E regulates the integrity of tight junctions in an isoform-dependent manner in an in vitro blood-brain barrier model. J. Biol. Chem. 2011, 286, 17536–17542. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.D.; Winkler, E.A.; Singh, I.; Sagare, A.P.; Deane, R.; Wu, Z.; Holtzman, D.M.; Betsholtz, C.; Armulik, A.; Sallstrom, J. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature 2012, 485, 512–516. [Google Scholar] [CrossRef] [PubMed]
- Halliday, M.R.; Pomara, N.; Sagare, A.P.; Mack, W.J.; Frangione, B.; Zlokovic, B. V Relationship between cyclophilin a levels and matrix metalloproteinase 9 activity in cerebrospinal fluid of cognitively normal apolipoprotein e4 carriers and blood-brain barrier breakdown. JAMA Neurol. 2013, 70, 1198–1200. [Google Scholar] [CrossRef]
- Ley, R.E.; Peterson, D.A.; Gordon, J.I. Ecological and Evolutionary Forces Shaping Microbial Diversity in the Human Intestine. Cell 2006, 124, 837–848. Available from: https://www.sciencedirect.com/science/article/pii/S0092867406001929. [CrossRef] [PubMed]
- Sánchez de Medina, F.; Romero-Calvo, I.; Mascaraque, C.; Martínez-Augustin, O. Intestinal Inflammation and Mucosal Barrier Function. Inflamm. Bowel Dis. 2014, 20, 2394–2404. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Tejpar, Q.Z.; Soo, I.; Madsen, K.L.; Fedorak, R.N. The role of antibiotic and probiotic therapies in current and future management of inflammatory bowel disease. Curr. Gastroenterol. Rep. 2006, 8, 486–498. [Google Scholar] [CrossRef]
- De Jager, P.L.; Franchimont, D.; Waliszewska, A.; Bitton, A.; Cohen, A.; Langelier, D.; Belaiche, J.; Vermeire, S.; Farwell, L.; Goris, A. The role of the Toll receptor pathway in susceptibility to inflammatory bowel diseases. Genes Immun. 2007, 8, 387–397. [Google Scholar] [CrossRef]
- Franchimont, D.; Vermeire, S.; El Housni, H.; Pierik, M.; Van Steen, K.; Gustot, T.; Quertinmont, E.; Abramowicz, M.; Van Gossum, A.; Devière, J. Deficient host-bacteria interactions in inflammatory bowel disease? The toll-like receptor (TLR)-4 Asp299gly polymorphism is associated with Crohn’s disease and ulcerative colitis. Gut 2004, 53, 987–992. [Google Scholar] [CrossRef] [PubMed]
- Hampe, J.; Franke, A.; Rosenstiel, P.; Till, A.; Teuber, M.; Huse, K.; Albrecht, M.; Mayr, G.; De La Vega, F.M.; Briggs, J. A genome-wide association scan of nonsynonymous SNPs identifies a susceptibility variant for Crohn disease in ATG16L1. Nat. Genet. 2007, 39, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Fujita, N.; Jang, M.H.; Uematsu, S.; Yang, B.-G.; Satoh, T.; Omori, H.; Noda, T.; Yamamoto, N.; Komatsu, M.; et al. Loss of the autophagy protein Atg16L1 enhances endotoxin-induced IL-1β production. Nature 2008, 456, 264–268. [Google Scholar] [CrossRef]
- Duchmann, R.; Kaiser, I.; Hermann, E.; Mayet, W.; Ewe, K.; BÜSCHENFELDE, K.M.Z.U.M. Tolerance exists towards resident intestinal flora but is broken in active inflammatory bowel disease (IBD). Clin. Exp. Immunol. 1995, 102, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Mileti, E.; Matteoli, G.; Chieppa, M.; Rescigno, M. Intestinal epithelial cells promote colitis-protective regulatory T-cell differentiation through dendritic cell conditioning. Mucosal Immunol. 2009, 2, 340–350. [Google Scholar] [CrossRef] [PubMed]
- Rimoldi, M.; Chieppa, M.; Salucci, V.; Avogadri, F.; Sonzogni, A.; Sampietro, G.M.; Nespoli, A.; Viale, G.; Allavena, P.; Rescigno, M. Intestinal immune homeostasis is regulated by the crosstalk between epithelial cells and dendritic cells. Nat. Immunol. 2005, 6, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Iliev, I.D.; Spadoni, I.; Mileti, E.; Matteoli, G.; Sonzogni, A.; Sampietro, G.M.; Foschi, D.; Caprioli, F.; Viale, G.; Rescigno, M. Human intestinal epithelial cells promote the differentiation of tolerogenic dendritic cells. Gut 2009, 58, 1481–1489. [Google Scholar] [CrossRef] [PubMed]
- Smythies, L.E.; Sellers, M.; Clements, R.H.; Mosteller-Barnum, M.; Meng, G.; Benjamin, W.H.; Orenstein, J.M.; Smith, P.D. Human intestinal macrophages display profound inflammatory anergy despite avid phagocytic and bacteriocidal activity. J. Clin. Invest. 2005, 115, 66–75. [Google Scholar] [CrossRef] [PubMed]
- Fukudo, S. Role of corticotropin-releasing hormone in irritable bowel syndrome and intestinal inflammation. J. Gastroenterol. 2007, 42, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Wallon, C.; Yang, P.-C.; Keita, Å. V; Ericson, A.-C.; McKay, D.M.; Sherman, P.M.; Perdue, M.H.; Söderholm, J.D. Corticotropin-releasing hormone (CRH) regulates macromolecular permeability via mast cells in normal human colonic biopsies in vitro. Gut 2008, 57, 50–58. [Google Scholar] [CrossRef]
- Lee, K.J.; Kim, Y.B.; Kim, J.H.; Kwon, H.C.; Kim, D.K.; Cho, S.W. The alteration of enterochromaffin cell, mast cell, and lamina propria T lymphocyte numbers in irritable bowel syndrome and its relationship with psychological factors. J. Gastroenterol. Hepatol. 2008, 23, 1689–1694. [Google Scholar] [CrossRef] [PubMed]
- Fujii, Y.; Nguyen, T.T.T.; Fujimura, Y.; Kameya, N.; Nakamura, S.; Arakawa, K.; Morita, H. Fecal metabolite of a gnotobiotic mouse transplanted with gut microbiota from a patient with Alzheimer’s disease. Biosci. Biotechnol. Biochem. 2019, 83, 2144–2152. [Google Scholar] [CrossRef] [PubMed]
- Rubin, L.L. Staddon. The cell biology of the blood-brain barrier. Ann Rev Neurosci 1999, 22, 11–28. [Google Scholar] [CrossRef] [PubMed]
- Bösel, J.; Schiller, P.; Hook, Y.; Andes, M.; Neumann, J.-O.; Poli, S.; Amiri, H.; Schönenberger, S.; Peng, Z.; Unterberg, A. Stroke-related early tracheostomy versus prolonged orotracheal intubation in neurocritical care trial (SETPOINT) a randomized pilot trial. Stroke 2013, 44, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.J. Clearing the brain’s amyloid cobwebs. Neuron 2001, 32, 177–180. [Google Scholar] [CrossRef] [PubMed]
- Zlokovic, B. V Cerebrovascular disease in Alzheimer’s disease. Trends Neurosci. 2005, 4, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Cullen, K.M.; Kócsi, Z.; Stone, J. Pericapillary haem-rich deposits: evidence for microhaemorrhages in aging human cerebral cortex. J. Cereb. Blood Flow Metab. 2005, 25, 1656–1667. [Google Scholar] [CrossRef] [PubMed]
- Ryu, J.K.; McLarnon, J.G. A leaky blood–brain barrier, fibrinogen infiltration and microglial reactivity in inflamed Alzheimer’s disease brain. J. Cell. Mol. Med. 2009, 13, 2911–2925. [Google Scholar] [CrossRef] [PubMed]
- Raven, E.P.; Lu, P.H.; Tishler, T.A.; Heydari, P.; Bartzokis, G. Increased iron levels and decreased tissue integrity in hippocampus of Alzheimer’s disease detected in vivo with magnetic resonance imaging. J. Alzheimer’s Dis. 2013, 37, 127–136. [Google Scholar] [CrossRef]
- Singh, N.; Thangaraju, M.; Prasad, P.D.; Martin, P.M.; Lambert, N.A.; Boettger, T.; Offermanns, S.; Ganapathy, V. Blockade of dendritic cell development by bacterial fermentation products butyrate and propionate through a transporter (Slc5a8)-dependent inhibition of histone deacetylases. J. Biol. Chem. 2010, 285, 27601–27608. [Google Scholar] [CrossRef]
- Wu, J.; Liu, C.; Zhang, L.; Qu, C.; Sui, X.; Zhu, H.; Huang, L.; Xu, Y.; Han, Y.; Qin, C. Histone deacetylase-2 is involved in stress-induced cognitive impairment via histone deacetylation and PI3K/AKT signaling pathway modification. Mol. Med. Rep. 2017, 16, 1846–1854. [Google Scholar] [CrossRef]
- Frost, G.; Sleeth, M.L.; Sahuri-Arisoylu, M.; Lizarbe, B.; Cerdan, S.; Brody, L.; Anastasovska, J.; Ghourab, S.; Hankir, M.; Zhang, S. The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 2014, 5, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cong, L.; Lukiw, W.J. Lipopolysaccharide (LPS) Accumulates in Neocortical Neurons of Alzheimer’s Disease (AD) Brain and Impairs Transcription in Human Neuronal-Glial Primary Co-cultures. Front. Aging Neurosci. 2017, 9, 407. [Google Scholar] [CrossRef] [PubMed]
- Lien, E.; Means, T.K.; Heine, H.; Yoshimura, A.; Kusumoto, S.; Fukase, K.; Fenton, M.J.; Oikawa, M.; Qureshi, N.; Monks, B. Toll-like receptor 4 imparts ligand-specific recognition of bacterial lipopolysaccharide. J. Clin. Invest. 2000, 105, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Ouanes, S.; Clark, C.; Richiardi, J.; Maréchal, B.; Lewczuk, P.; Kornhuber, J.; Kirschbaum, C.; Popp, J. Cerebrospinal Fluid Cortisol and Dehydroepiandrosterone Sulfate, Alzheimer’s Disease Pathology, and Cognitive Decline. Front. Aging Neurosci. 2022, 14, 892754. [Google Scholar] [CrossRef] [PubMed]
- Sheng, C.; Yang, K.; He, B.; Du, W.; Cai, Y.; Han, Y. Combination of gut microbiota and plasma amyloid-β as a potential index for identifying preclinical Alzheimer’s disease: a cross-sectional analysis from the SILCODE study. Alzheimers. Res. Ther. 2022, 14, 35. [Google Scholar] [CrossRef] [PubMed]
- Verhaar, B.J.H.; Hendriksen, H.M.A.; de Leeuw, F.A.; Doorduijn, A.S.; van Leeuwenstijn, M.; Teunissen, C.E.; Barkhof, F.; Scheltens, P.; Kraaij, R.; van Duijn, C.M.; et al. Gut Microbiota Composition Is Related to AD Pathology. Front. Immunol. 2021, 12, 794519. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, Y.; Liu, W.; Wang, T.; Wang, L.; Hao, L.; Ju, M.; Xiao, R. Diet quality, gut microbiota, and microRNAs associated with mild cognitive impairment in middle-aged and elderly Chinese population. Am. J. Clin. Nutr. 2021, 114, 429–440. [Google Scholar] [CrossRef]
- Ling, Z.; Zhu, M.; Yan, X.; Cheng, Y.; Shao, L.; Liu, X.; Jiang, R.; Wu, S. Structural and Functional Dysbiosis of Fecal Microbiota in Chinese Patients With Alzheimer’s Disease. Front. cell Dev. Biol. 2020, 8, 634069. [Google Scholar] [CrossRef]
- Wu, L.; Han, Y.; Zheng, Z.; Peng, G.; Liu, P.; Yue, S.; Zhu, S.; Chen, J.; Lv, H.; Shao, L.; et al. Altered Gut Microbial Metabolites in Amnestic Mild Cognitive Impairment and Alzheimer’s Disease: Signals in Host-Microbe Interplay. Nutrients 2021, 13. [Google Scholar] [CrossRef]
- Marizzoni, M.; Cattaneo, A.; Mirabelli, P.; Festari, C.; Lopizzo, N.; Nicolosi, V.; Mombelli, E.; Mazzelli, M.; Luongo, D.; Naviglio, D.; et al. Short-Chain Fatty Acids and Lipopolysaccharide as Mediators Between Gut Dysbiosis and Amyloid Pathology in Alzheimer’s Disease. J. Alzheimers. Dis. 2020, 78, 683–697. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Neth, B.J.; Wang, S.; Mishra, S.P.; Craft, S.; Yadav, H. Gut mycobiome and its interaction with diet, gut bacteria and alzheimer’s disease markers in subjects with mild cognitive impairment: A pilot study. EBioMedicine 2020, 59, 102950. [Google Scholar] [CrossRef] [PubMed]
- Stadlbauer, V.; Engertsberger, L.; Komarova, I.; Feldbacher, N.; Leber, B.; Pichler, G.; Fink, N.; Scarpatetti, M.; Schippinger, W.; Schmidt, R.; et al. Dysbiosis, gut barrier dysfunction and inflammation in dementia: a pilot study. BMC Geriatr. 2020, 20, 248. [Google Scholar] [CrossRef] [PubMed]
- Plagman, A.; Hoscheidt, S.; McLimans, K.E.; Klinedinst, B.; Pappas, C.; Anantharam, V.; Kanthasamy, A.; Willette, A.A. Cholecystokinin and Alzheimer’s disease: a biomarker of metabolic function, neural integrity, and cognitive performance. Neurobiol. Aging 2019, 76, 201–207. [Google Scholar] [CrossRef]
- Nho, K.; Kueider-Paisley, A.; MahmoudianDehkordi, S.; Arnold, M.; Risacher, S.L.; Louie, G.; Blach, C.; Baillie, R.; Han, X.; Kastenmüller, G.; et al. Altered bile acid profile in mild cognitive impairment and Alzheimer’s disease: Relationship to neuroimaging and CSF biomarkers. Alzheimers. Dement. 2019, 15, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; He, Y.; Ma, J.; Huang, P.; Du, J.; Cao, L.; Wang, Y.; Xiao, Q.; Tang, H.; Chen, S. Mild cognitive impairment has similar alterations as Alzheimer’s disease in gut microbiota. Alzheimers. Dement. 2019, 15, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wu, L.; Peng, G.; Han, Y.; Tang, R.; Ge, J.; Zhang, L.; Jia, L.; Yue, S.; Zhou, K.; et al. Altered microbiomes distinguish Alzheimer’s disease from amnestic mild cognitive impairment and health in a Chinese cohort. Brain. Behav. Immun. 2019, 80, 633–643. [Google Scholar] [CrossRef] [PubMed]
- Vogt, N.M.; Romano, K.A.; Darst, B.F.; Engelman, C.D.; Johnson, S.C.; Carlsson, C.M.; Asthana, S.; Blennow, K.; Zetterberg, H.; Bendlin, B.B.; et al. The gut microbiota-derived metabolite trimethylamine N-oxide is elevated in Alzheimer’s disease. Alzheimers. Res. Ther. 2018, 10, 124. [Google Scholar] [CrossRef]
- Wang, Y.; Li, N.; Yang, J.-J.; Zhao, D.-M.; Chen, B.; Zhang, G.-Q.; Chen, S.; Cao, R.-F.; Yu, H.; Zhao, C.-Y.; et al. Probiotics and fructo-oligosaccharide intervention modulate the microbiota-gut brain axis to improve autism spectrum reducing also the hyper-serotonergic state and the dopamine metabolism disorder. Pharmacol. Res. 2020, 157, 104784. [Google Scholar] [CrossRef]
- Berding, K.; Carbia, C.; Cryan, J.F. Going with the grain: Fiber, cognition, and the microbiota-gut-brain-axis. Exp. Biol. Med. 2021, 246, 796–811. [Google Scholar] [CrossRef]
- Yusufov, M.; Weyandt, L.L.; Piryatinsky, I. Alzheimer’s disease and diet: a systematic review. Int. J. Neurosci. 2017, 127, 161–175. Available from: http://gut.bmj.com/content/69/7/1218.abstract. [CrossRef]
- Ghosh, T.S.; Rampelli, S.; Jeffery, I.B.; Santoro, A.; Neto, M.; Capri, M.; Giampieri, E.; Jennings, A.; Candela, M.; Turroni, S.; et al. Mediterranean diet intervention alters the gut microbiome in older people reducing frailty and improving health status: the NU-AGE 1-year dietary intervention across five European countries. Gut 2020, 69, 1218. [Google Scholar] [CrossRef]
- Hoscheidt, S.; Sanderlin, A.H.; Baker, L.D.; Jung, Y.; Lockhart, S.; Kellar, D.; Whitlow, C.T.; Hanson, A.J.; Friedman, S.; Register, T.; et al. Mediterranean and Western diet effects on Alzheimer’s disease biomarkers, cerebral perfusion, and cognition in mid-life: A randomized trial. Alzheimer’s Dement. 2022, 18, 457–468. [Google Scholar] [CrossRef] [PubMed]
- Dowis, K.; Banga, S. The Potential Health Benefits of the Ketogenic Diet: A Narrative Review. Nutrients 2021, 13, 1654. [Google Scholar] [CrossRef]
- Sergi, D.; Naumovski, N.; Heilbronn, L.K.; Abeywardena, M.; O’Callaghan, N.; Lionetti, L.; Luscombe-Marsh, N. Mitochondrial (dys)function and insulin resistance: From pathophysiological molecular mechanisms to the impact of diet. Front. Physiol. 2019, 10, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Rusek, M.; Pluta, R.; Ułamek-Kozioł, M.; Czuczwar, S.J. Ketogenic Diet in Alzheimer’s Disease. Int. J. Mol. Sci. 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.X.; Maalouf, M.; Han, P.; Zhao, M.; Gao, M.; Dharshaun, T.; Ryan, C.; Whitelegge, J.; Wu, J.; Eisenberg, D.; et al. Ketones block amyloid entry and improve cognition in an Alzheimer’s model. Neurobiol. Aging 2016, 39, 25–37. [Google Scholar] [CrossRef]
- Maalouf, M.; Rho, J.M.; Mattson, M.P. The neuroprotective properties of calorie restriction, the ketogenic diet, and ketone bodies. Brain Res. Rev. 2009, 59, 293–315. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, B.; Delgado, S.; Blanco-Míguez, A.; Lourenço, A.; Gueimonde, M.; Margolles, A. Probiotics, gut microbiota, and their influence on host health and disease. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Ziemer, C.J.; Gibson, G.R. An Overview of Probiotics, Prebiotics and Synbiotics in the Functional Food Concept: Perspectives and Future Strategies. Int. Dairy J. 1998, 8, 473–479. [Google Scholar] [CrossRef]
- Shi, H.; Wang, Q.; Zheng, M.; Hao, S.; Lum, J.S.; Chen, X.; Huang, X.-F.; Yu, Y.; Zheng, K. Supplement of microbiota-accessible carbohydrates prevents neuroinflammation and cognitive decline by improving the gut microbiota-brain axis in diet-induced obese mice. J. Neuroinflammation 2020, 17, 77. [Google Scholar] [CrossRef]
- Wu, S.-C.; Cao, Z.-S.; Chang, K.-M.; Juang, J.-L. Intestinal microbial dysbiosis aggravates the progression of Alzheimer’s disease in Drosophila. Nat. Commun. 2017, 8, 24. Available from: https://www.frontiersin.org/articles/10.3389/fnagi.2016.00262. [CrossRef] [PubMed]
- Steenbergen, L.; Sellaro, R.; van Hemert, S.; Bosch, J.A.; Colzato, L.S. A randomized controlled trial to test the effect of multispecies probiotics on cognitive reactivity to sad mood. Brain. Behav. Immun. 2015, 48, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Benton, D.; Williams, C.; Brown, A. Impact of consuming a milk drink containing a probiotic on mood and cognition. Eur. J. Clin. Nutr. 2007, 61, 355–361. [Google Scholar] [CrossRef]
- Burnet, P.W.J.; Cowen, P.J. Psychobiotics highlight the pathways to happiness. Biol. Psychiatry 2013, 74, 708–709. [Google Scholar] [CrossRef]
- Davani-Davari, D.; Negahdaripour, M.; Karimzadeh, I.; Seifan, M.; Mohkam, M.; Masoumi, S.J.; Berenjian, A.; Ghasemi, Y. Prebiotics: Definition, Types, Sources, Mechanisms, and Clinical Applications. Foods (Basel, Switzerland) 2019, 8. [Google Scholar] [CrossRef]
- Schmidt, K.; Cowen, P.J.; Harmer, C.J.; Tzortzis, G.; Errington, S.; Burnet, P.W.J. Prebiotic intake reduces the waking cortisol response and alters emotional bias in healthy volunteers. Psychopharmacology (Berl). 2015, 232, 1793–1801. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Liu, S.; Ling, Z.; Wang, F.; Ling, Y.; Gong, T.; Fang, N.; Ye, S.; Si, J.; Liu, J. Fructooligosaccharides Ameliorating Cognitive Deficits and Neurodegeneration in APP/PS1 Transgenic Mice through Modulating Gut Microbiota. J. Agric. Food Chem. 2019, 67, 3006–3017. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lai, D.-M.; Huang, H.-J.; Lee-Chen, G.-J.; Chang, C.-H.; Hsieh-Li, H.M.; Lee, G.-C. Prebiotic Lactulose Ameliorates the Cognitive Deficit in Alzheimer’s Disease Mouse Model through Macroautophagy and Chaperone-Mediated Autophagy Pathways. J. Agric. Food Chem. 2021, 69, 2422–2437. [Google Scholar] [CrossRef]
- Wang, T.; Kuang, W.; Chen, W.; Xu, W.; Zhang, L.; Li, Y.; Li, H.; Peng, Y.; Chen, Y.; Wang, B.; et al. A phase II randomized trial of sodium oligomannate in Alzheimer’s dementia. Alzheimers. Res. Ther. 2020, 12, 110. [Google Scholar] [CrossRef]
- Hazan, S. Rapid improvement in Alzheimer’s disease symptoms following fecal microbiota transplantation: a case report. J. Int. Med. Res. 2020, 48, 300060520925930. [Google Scholar] [CrossRef] [PubMed]
- Zhan, G.; Yang, N.; Li, S.; Huang, N.; Fang, X.; Zhang, J.; Zhu, B.; Yang, L.; Yang, C.; Luo, A. Abnormal gut microbiota composition contributes to cognitive dysfunction in SAMP8 mice. Aging (Albany. NY). 2018, 10, 1257–1267. [Google Scholar] [CrossRef] [PubMed]
- Dodiya, H.B.; Kuntz, T.; Shaik, S.M.; Baufeld, C.; Leibowitz, J.; Zhang, X.; Gottel, N.; Zhang, X.; Butovsky, O.; Gilbert, J.A.; et al. Sex-specific effects of microbiome perturbations on cerebral Aβ amyloidosis and microglia phenotypes. J. Exp. Med. 2019, 216, 1542–1560. [Google Scholar] [CrossRef]
Figure 1.
Integrative physiology of MGB axis [CNS-central Nervous System, ENS-Enteric Nervous System, HPA-Hypothalamic Pituitary Adrenal Axis).
Figure 1.
Integrative physiology of MGB axis [CNS-central Nervous System, ENS-Enteric Nervous System, HPA-Hypothalamic Pituitary Adrenal Axis).
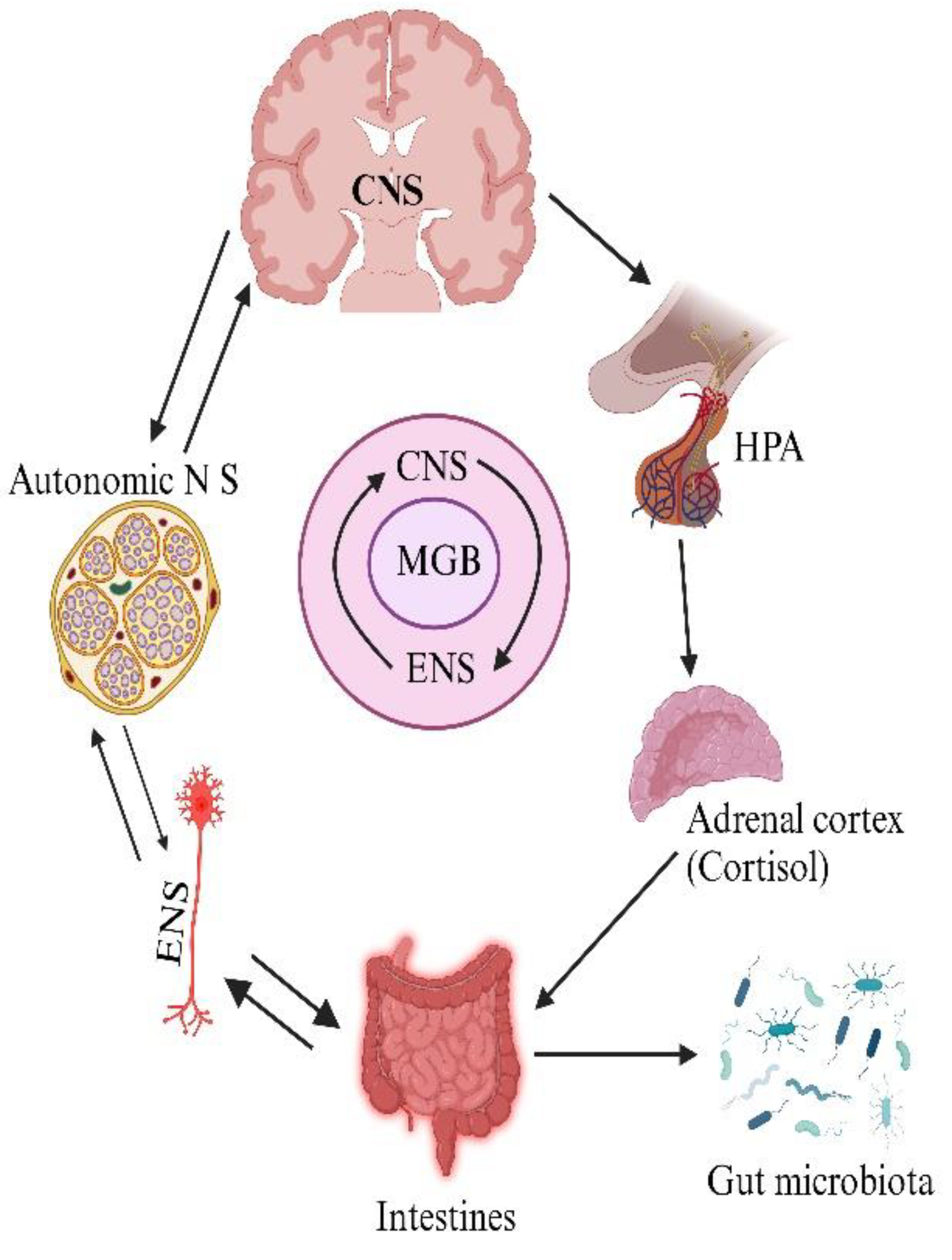
Figure 2.
Illustrative diagram represents the microbial gut dysbiosis leading to production of pro-inflammatory cytokines, nonindispensible harmful metabolites and changes in neurotransmitter production that contributes towards the development of AD. The metabolite diversity in gut dysbiosis increases the intestinal permeability and mucous secretion due to disruption of tight junctions and inflammation of epithelial intestinal cells respectively. Such metabolites may serve as biomarkers of development of AD in serum and faeces. The Ab and tau proteins that travel via vagus nerve and produced from GMD also responsible for development of AD. (AD, Azhiemer`s Disease; GMD, gut microbial dysbiosis; Ab, amyloid beta, PIC, proinfammatory cytokines; SCFA, short chain fatty acids; LPS, lipopolysaccharide; BBB, blood brain Barrier).
Figure 2.
Illustrative diagram represents the microbial gut dysbiosis leading to production of pro-inflammatory cytokines, nonindispensible harmful metabolites and changes in neurotransmitter production that contributes towards the development of AD. The metabolite diversity in gut dysbiosis increases the intestinal permeability and mucous secretion due to disruption of tight junctions and inflammation of epithelial intestinal cells respectively. Such metabolites may serve as biomarkers of development of AD in serum and faeces. The Ab and tau proteins that travel via vagus nerve and produced from GMD also responsible for development of AD. (AD, Azhiemer`s Disease; GMD, gut microbial dysbiosis; Ab, amyloid beta, PIC, proinfammatory cytokines; SCFA, short chain fatty acids; LPS, lipopolysaccharide; BBB, blood brain Barrier).
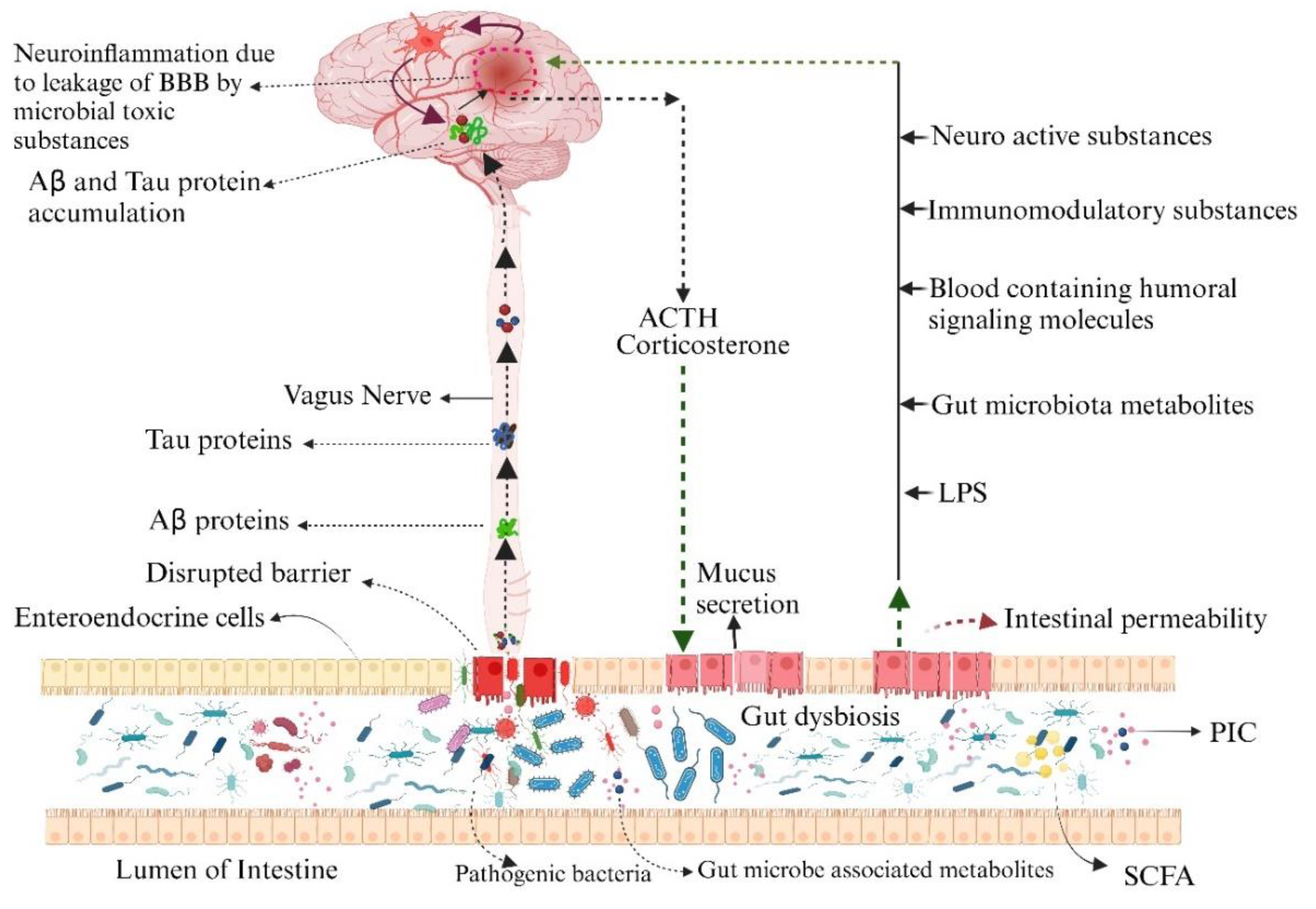

Table 1.
Role of different microorganisms residing in the gut in different subjects.
| Reference | Subject | Bacteria/organism | Effects(+/-) | Role |
|---|---|---|---|---|
| Cerovic et al., 2019 | AD patients | Bacteroides fragilis | + | Protection against CNS demyelination diseases. [42] |
| Deng, H et al., 2018 | C57BL/6 mice | + | In pregnant mice showed an immediate significant diminished autistic behavior [43] | |
| Cattaneo et al 2017 | AD patients | Eubacterium rectale | - | Leads to amyloidosis [35] |
| Tanida et al., 2005 | BB-DR rats Healthy humans |
Lactobacillus johnsonii |
+ | Improved gastric vagus nerve activity. [44] |
| Singhrao et al., 2015 | AD patients |
Porphyromonas gingivalis |
- | Responsible for cirrhosis, which leads to neurodegeneration [34] |
| Yanget al., 2020 | SAMP8 mice |
Lactobacillus acidophilus |
+ | Enhanced the injury in neural proteolysis.[45] |
| Gareau et al., 2005 | C57BL/6 mice |
Campylobacter rodentium |
- | Led to stress and contributed to behavioral abnormalities. [33] |
| Mehrabadi and Sadr,2020 | Wistar rats |
Lactobacillus rhamnosus |
+ | Improved the inflammation level in the brain. [31] |
Table 2.
An overview of animal studies looking at how the host's physiology and cognitive function are affected by induced changes in the GM.
Table 2.
An overview of animal studies looking at how the host's physiology and cognitive function are affected by induced changes in the GM.
| First author journal year | Animal Model | Exposure | Outcome measured | Main results | Proposed mechanism of intestinal microbiota involvement in dementia physiopathology |
|---|---|---|---|---|---|
| [75]Harach,Sci Rep,2017 | APPPSI transgenic mice | Mice that are germ-free thanks to IVF and are raised in a sterile environment | Cerebral amyloid deposition | Germ-free mice have less cerebral amyloid deposition than controls, whereas germ-free mice who received fecal microbiota transplants from controls have more amyloid deposition. | Bacteria in the gastrointestinal tract may cause subclinical brain inflammation that facilitates the deposition of amyloid. |
| [76] Minter,Sci Rep,2016 | APPSWE/PSIΔE9Transgenic mice | Long term broad-Spectrum combinatorial antibiotic therapy | Aβamyloid deposition soluble Aβlevels cerebral glial reactivity Circulating cytokine and chemokine level | Mice given antibiotics had higher levels of circulating cytokines and chemokine’s, less cerebral A-plaque formation, and higher levels of glial activity and circulating A. | Gut dysbiosis may have beneficial effects during the acute phase (reduced A plaque deposition), but negative effects during the long term. |
| [77] Minter, Sci Rep,2017 | APPSWE/PSIΔE9Transgenic mice | Postnatal antibiotic treatment | makeup of the gut microbiome Brain deterioration Foxp3+ Inflammation caused by T-reg cells in the blood, brain serum, and cerebrospinal fluid | Post-antibiotic therapy led to stable changes in GM diversity, including a decrease in species richness and an increase in Lachnospiraceae representation, an up regulation of circulating and brain Foxp3+Treg cells, and a decrease in the breakdown of brain A-amyloid. | Through the synthesis of butyrate and the differentiation of T-cells into Foxp3+Treg cells, the GM may mediate neuroinflammation. |
| [78]Frohlich, Brain Behav Immun, 2016 | Adult mice | Intragastric treatment with multiple antibiotics | colon's metabolic profile circulation microbial metabolites expression of signaling molecules in neurons Cognitive Conduct | Enhanced cytokine representation in the amygdala and hippocampus, decreased neuronal signaling-related chemicals in the brain, decreased colonic and circulating microbial metabolites, and worse novel object recognition memory are all indicators of GM dysbiosis. | Depletion of microbial metabolites in the gut may function as endocrine messengers, affecting the synthesis of neutral signaling molecules, particularly in the amygdala and hippocampus (BDNF, NMDA receptor, serotonin transporter, and NPY). |
| [79]Cui, BBA Mol Basis Dis, 2017 | C57BL/6Jmice | Total abdominal irradiation | circulating and intestinal miRNAs Expression of BDNF in the brain and gut microbiome | Cognitive impairment brought on by radiation, dysbiosis of the gut microbiome, overexpression of the circulating miR-34a-5p gene, and decreased expression of the brain protein BDNF GM eubiosis and BDNF synthesis were restored by injection of miR-34a-5p antagonists. | GM may influence the expression of miRNAs, specifically miR-34a-5p, which may mediate a bidirectional gut-brain interaction. |
Abbreviations: NBDA, N-methyl-D-aspartic acid; Treg, T regulatory; IVF, in vitro fertilization; BDNF, brain-derived neurotropic factor, NPY, neuropeptide Y.
Table 3.
: Microbiome-based biomarkers in humans.
| S.No | Biomarker(s) proposed | Samples and methods | Results | Refrences |
|---|---|---|---|---|
| 1. | Cortisol and DHEAS | ELISA, CLIA | 1.↑ CSF cortisol was associated with↓global cognitive performance and↑disease severity at baseline 2.↑ Cortisol and cortisol/DHEAS ratio were associated with↑tau and p-tau CSF levels and↓ amygdala and insula volumes at baseline. |
[126] |
| 2. | Plasma Aβ + gut microbiota | 16SrRNA Miseq sequencing of fecal microbiome and MSD quantifying the plasma Aβ 40 , Aβ42 , and Aβ 42/Aβ40 | 1.↑↑ The relative abundance of phylum Bacteroidetes in CN+ compared to CN 2.↓↓ Phylum Firmicutes and class Delta proteobacteria in CN+compared to CN 3. The relative abundance of phylum Firmicutes and its corresponding SCFA-producing bacteria in CN−>CN+>CI |
[127] |
| 3. | SCFA-producing microorganisms | 16SrRNA Miseq sequencing of fecal microbiome and CSF Aβ, p-tau | 1.↑ Clostridium leptum,↓ Eubacterium ventriosum group spp., Lachnospiraceae spp., Marvinbryantia spp.,Monoglobus spp., Ruminococcus torques group spp.,Roseburia hominis, and Christensenellaceae R-7 spp. with higher odds of amyloid positivity 2.↓ Lachnospiraceae spp., Lachnoclostridium spp.,Roseburia hominis, and Bilophila wadsworthia with higher odds of positive p-tau status |
[128] |
| 4. | Gut microbiota composition + diet quality scores + serum miRNA | 16SrDNA gene sequencing of fecal microbiome and serum miRNA expression | 1.↓ Microbial diversity, Faecalibacterium, Ruminococcaceae, and Alipstes in MCI compared to controls 2.↑ Proteobacteria and Gammaproteobacteria in MCI compared to controls |
[129] |
| 5. | Faecalibacterium, Bifidobacterium | 16SrRNA Miseq sequencing of fecal microbiome | 1.↓ Microbial diversity in AD compared to controls 2.↓ Butyrate-producing bacteria (Faecalibacterium) in AD 3.↑ Lactate-producing bacteria (Bifidobacterium) in AD |
[130] |
| 6. | Indole-3-pyruvic acid five SCFAs | LC/GC/MS metabolomics profiling of fecal microbiome | 1.↓ Tryptophan metabolites in MCI and, more pronounced, in AD compared to controls 2.↓↓ SCFAs (formic acid, acetic acid, propanoic acid, 2-methylbutyric acid, and isovaleric acid) in AD compared to controls;↓ in MCI compared to controls |
[131] |
| 7. | SCFAs and LPS | ELISA measured blood levels of LPS, and SCFAs by mass spectrometry | 1.↑ Blood LPS, acetate and valerate, pro-inflammatory cytokines, and biomarkers of endothelial dysfunction in dementia 2.↓ Butyrate and the anti-inflammatory cytokine IL10 in dementia |
[132] |
| 8. | Mycobiome signatures | 16SrRNA sequencing and fungal rRNA ITS1 sequencing of fecal bacterial and mycobiome | 1.↑ Genera Botrytis, Kazachstania, Phaeoacremonium, Cladosporium, and families Sclerotiniaceae, Phaffomyceteceae, Trichocomaceae, Cystofilobasidiaceae, and Togniniaceae in MCI compared to CN 2. Meyerozyma in MCI< CN 3. Different correlation patterns with AD markers and GM are displayed by specific fungal species (Debariomyce, Sarocladium, Filobasidium, Candida, and Cladosporium) |
[133] |
| 9. | Lachnospiraceae NK4A136 group, DAO, sCD14 | GM composition, gut barrier dysfunction, bacterial translocation, and inflammation were assessed from stool and serum samples by 16SrRNA sequencing, QIIME 2, Calypso 7.14 tools, and ELISA | 1.↓↓ Lachnospiraceae, genus Lachnospiraceae NK4A136 group in dementia. 2.↓ Eubacterium rectale in dementia. 3.↑ DAO and sCD14 in dementia. 4. Faecalibacterium prausnitzii was associated with mild dementia; Lactobacillus amylovorus was associated with moderate dementia; Clostridium clostridiforme and Streptococcus salivarius were associated with Severe dementia |
[134] |
| 10. | CCK | Targeted quantitation | 1.↑ CCK was a strong relationship associated with↑tau levels 2.↑ CCK was related to↑global and memory scores3.↑ CCK was related to↑gray matter volume |
[135] |
| 11. | BA | targeted metabolomics profiling of the Serum levels of 20 primary and secondary BA metabolite | 1. Three BA (↑ GDCA: CA, TDCA: CA, and GLCA: CDCA)signatures were associated with↓ CSF Aβ1-42 (“A”) 2. Three BA (↑ GCDCA、GLCA and TLCA) with↑ CSFp-tau181 (“T”) 3. The CSF t-tau, glucose metabolism, and atrophy (“N”) were connected to three, twelve, and fourteen BA signatures, respectively |
[136] |
| 12. | Microbial diversity | Analysis of microbiome community in the feces and blood via 16SrRNA sequencing | 1.↓ Microbial diversity in AD and MCI compared to controls 2.11 genera in the feces (↑ Dorea, Lactobacillus, Streptococcus, Bifidobacterium, Blautia, and Escherichia;↓ Alistipes, Bacteroides, Parabacteroides, Sutterella, and Paraprevotella) and in the blood (↑ Propionibacterium, Pseudomonas, Glutamicibacter, Escherichia, and Acidovora;↓ Acinetobacter, Aliihoeflea, Halomonas, Leucobacter, Pannonibacter, and Ochrobactrum) between AD, MCI and controls |
[137] |
| 13. | Enterobacteriaceae | Phylogenetic analysis of communities using 16S rRNA MiSeq sequencing and reconstruction of the feces | 1.↓ Microbial diversity in AD compared to MCI and controls 2.↓ Firmicutes in AD compared to controls 3.↑ Proteobacteria in AD compared to controls 4.↑ Gammaproteobacteria, Enterobacteriales, and Enterobacteriaceae in AD > MCI > controls |
[138] |
| 14. | TMAO | Cerebrospinal TMAO levels measurement and relationships between CSF TMAO and CSF biomarkers of AD | 1.↑ TMAO in AD and MCI compared to controls 2. Elevated CSF TMAO is associated with biomarkers of AD pathology |
[139] |
(↑: increase; ↓: decrease; ↑↑: significant increase; ↓↓: significant decrease). (Zhang et al., 2023).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



