
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
Epilepsy is one of the most prevalent and serious brain disorders, affecting 70 million people worldwide. Antiseizure medications (ASMs) though relieve symptoms and prevent the occurrence of future seizures in epileptic patients have limited effect on epileptogenesis. Addressing the multifaceted nature of epileptogenesis and its association with Nod-like receptor family pyrin domain containing 3 (NLRP3) inflammasome-mediated neuroinflammation requires a comprehensive understanding of the underlying mechanisms for development of targeted therapeutic strategies beyond conventional antiseizure treatments. Several types of NLRP3 inhibitors have been developed and their effect has been validated both in vitro and in vivo models of epileptogenesis. In this review, we discuss the advances in understanding the regulatory mechanisms of NLRP3 activation as well as progress made and challenges facing in the development of NLRP3 inhibitors for the treatment of epilepsy.
Keywords:
epilepsy
; neuroinflammation
; NLRP3 inflammasome
; epileptogenesis
; antiseizure medication
; interleukin-1b
; caspase-1
; ASC
1. Introduction
Epilepsy is a serious chronic brain disorder characterized by spontaneous and recurrent seizures [1,2] that affects over 70 million people worldwide [3]. Approximately 3 million adults and 470,000 children suffer from epilepsy, with an incidence of about 150,000 new cases each year in the USA [4]. According to the World Health Organization, roughly half of epilepsy cases have no known cause, whereas the other half may be due to imbalance of neurotransmitters, brain tumors, strokes, immune disorders, gene mutations, and traumatic brain injury. The majority of epileptic patients also suffer from depression and memory loss, resulting in a poor quality of life and reduced life expectancy [5]. The use of antiseizure medications (ASMs) remains the preferred treatment, and seizures are being effectively controlled in 70% of epileptic patients [6]. Although more than 30 FDA-approved ASMs with diverse molecular targets are available for therapeutic purposes [7], the biggest challenge in the clinical treatment of epilepsy is that about 30% of epileptic patients develop resistance to ASMs. There is an urgent need to develop newer drugs with better efficacy, fewer side effects, and less toxicity. This need is further augmented by the fact that available ASMs do not address epileptogenesis, the disease process that leads to epilepsy [8,9]. Understanding of the mechanisms involved in epileptogenesis will be crucial in the development of novel drugs that target the epileptogenic process.
2. Epileptogenesis
Epileptogenesis is a poorly understood, multifaceted chronic process that alters the normal brain into an epileptic brain capable of generating spontaneous recurrent seizures (SRSs) [10]. Some of the common processes involved in epileptogenesis include neuroinflammation, neuronal cell death in the temporal lobe and hippocampus, bioenergetic dysfunction, altered neuroplasticity, blood–brain barrier (BBB) disruption, astrogliosis, microglial activation, and the infiltration of inflammatory cells into brain parenchyma. Injections of a chemo-convulsant, such as pilocarpine or kainic acid, or repetitive electrical stimulation of the hippocampus or amygdala, can result in a chronic epileptic condition characterized by strong convulsive SRSs in animal models of acquired epilepsy, the use of which will help tremendously in better diagnosis, treatment, and prevention of human epilepsy. Anticonvulsant drugs mostly relieve symptoms and prevent future seizures in patients with epilepsy rather than inhibiting and/or modulating the neuroinflammatory processes of epileptogenesis [9]. The development of targeted therapeutic strategies beyond conventional anticonvulsant/antiseizure treatments is needed to address epileptogenesis and epilepsy.
In general, neuroinflammation is beneficial because it promotes homeostatic neurogenesis [11,12], provides protection against the loss of axons and neurons [13], and promotes axonal regeneration [14,15], but chronic neuroinflammation has been shown to be detrimental in numerous neurological disorders including Alzheimer’s disease, meningitis, Parkinson’s disease, multiple sclerosis, encephalitis [16], and epilepsy [17,18,19]. Despite the absence of pathogens, neuroinflammation in epilepsy is triggered by an abnormal increase in proinflammatory mediators [20] or by endogenous inducers (damage-associated molecular patterns [DAMPs]) produced and secreted by brain cells undergoing stressful events termed as sterile inflammation [21]. Damage-associated molecular patterns such as adenosine triphosphate (ATP), high mobility group box1 (HMGB1), reactive oxygen species (ROSs), K+ efflux, advance glycation end products (AGEs), S1100b (a Ca2+ binding protein), or migration inhibitory factor-related protein 8 (MRP8) are known to induce the formation of inflammasome, a multi-molecular protein complex [22,23,24].
3. Inflammasomes
In 2002, Tschopp and colleagues coined the term “inflammasome” to describe a high-molecular-weight protein complex that mediates the activation of inflammatory caspases [25]. An inflammasome is comprised of a sensor protein (a cytoplasmic pattern-recognition receptor (PRR) classified by its protein domain structures), an adapter protein (apoptosis-associated speck-like protein containing a CARD [ASC]), and pro-caspase-1 as an effector protein [26]. Typically, inflammasomes are named after their cytoplasmic pattern-recognition receptors (PRRs), sensor proteins that are either of a NLR family (a nucleotide binding domain (NBD) combined with a leucine-rich-repeat-containing receptor (LRR-CR) protein) or an ALR family (an absence in melanoma 2 (AIM2)-like receptor (ALR) protein). Most pertinent inflammasomes belong to the NLR family subgroups, containing pyrene domain (NLRP) or CARD domain (NLRC) proteins. The best known inflammasomes are NLRP1; NLRP2; NLRP3; NLRP6, NLRP7; NLRP12; NLR-family apoptosis inhibitory protein (NAIP); NLR-family, CARD-containing 4 (NLRC4); AIM2; and Pyrin inflammasomes. Numerous studies have demonstrated that the central nervous system (CNS) is equipped with different inflammasome subtypes; however, their expression profiles vary among CNS cell types [27,28,29,30,31]. NLRP1 is mostly expressed in neurons and oligodendrocytes [32,33,34,35], NLRP2 in astrocytes [36,37], NLRP3 in microglia [38,39,40,41], NLRC4 in astrocytes [42,43], and AIM2 in neurons [44,45]. NLRP3, also known as cryopyrin, is the most widely studied and best characterized inflammasome. The NLRP3 protein has an N-terminal Pyrin domain (PYD), a central ATPase domain known as NACHT that comprises the NBD, a helical domain 1 (HD1), a winged helix domain (WHD), a helical domain 2 (HD2), and a C-terminal LRR domain [46].
NIMA (never in mitosis A)-related kinase 7 (NEK7) is a serine/threonine kinase that modulates microtubule stability, mitotic spindle formation, and cytokinesis and has now been recognized as a crucial component of the NLRP3 inflammasome [47,48]. NEK7 senses reactive oxygen species (ROSs), K+ efflux, lysosomal destabilization, and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) signaling-mediated activation of NLRP3 [49,50]. A cryo-EM structure of the NLRP3-NEK7 protein complex suggests that the first half of the NEK7 C-terminal interacts with the LRR domain of NLRP3 and the second half with the NBD and HD 2 domains of NACHT [46]. The binding of NEK7 alone is not sufficient to activate NLRP3. In addition, ATP needs to bind to the NBD domain and then be used to phosphorylate S195 in NEK7, prompting NLRP3 to adopt a modified conformation that drives oligomerization and activation of the NLRP3 complex [51]. Thus, NEK7 plays a licensing role in NLRP3 activation rather than serving as the sole activator [46].
4. Activation of the NLRP3 Inflammasome
The physiological activation of NLRP3 is tightly regulated to avoid uncontrolled inflammatory responses and is beneficial against invading pathogens and tissue damage, whereas abnormal or chronic activation plays a detrimental role in cellular and body health. As depicted in Figure 1, NLRP3 inflammasome activation can occur via canonical, non-canonical, and alternative pathways [52]. Optimal activation of NLRP3 via canonical pathway requires priming step/Signal 1, initiated by the detection of extracellular PAMPs and endogenous cytokines by the PRRs, which then upregulates the NF-κB-mediated transcription of NLRP3, pro-IL-1β, and pro-IL-18 [53]. In addition, priming also regulates NLRP3 at the post-transcriptional level through DNA methylation, protein acetylation, and the use of microRNAs. For example, Wei et al. reported that NLRP3 expression was increased through NLRP3 promoter demethylation in response to Mycobacterium tuberculosis infection [54]. It has been revealed that the NLRP3 induction by lipopolysaccharide (LPS) priming is decreased by binding of the RNA-binding protein Tristetraprolin [55] and miRNA-223 [56] to 3’UTR of NLRP3. Moreover, priming also involves various post-translational modifications (PTMs) of the NLRP3 protein, such as ubiquitination and phosphorylation, that silence NLRP3 for subsequent activation by a second signal.
The second activation step, or Signal 2, is the assembly of the inflammasome proteins triggered by sensing intracellular PAMPs/DAMPs through a combination of important and related events such as activation of the purinergic receptor P2X7 by ATP, cathepsin release following lysozyme rupture, the opening of Ca2+ channels to allow ion flux, mitochondrial dysfunction, ROS formation, Golgi apparatus disassembly, and endoplasmic reticulum stress (ERS) [57]. Once activated, oligomerization of the NLRP3 inflammasome involves two homotypic protein–protein interactions. First, the PYD domain of NLRP3 protein interacts with the adaptor protein ASC via PYD–PYD interactions and the CARD domain of ASC interacts with the CARD domain of pro-caspase 1 to recruit it to the NLRP3 inflammasome [58]. The activated NLRP3 inflammasome is thought to induce conformational changes that generate active caspase-1 (Casp1), which converts pro-interleukin-1β (pro-IL-1β) and pro-interleukin-18 (pro-IL-18) to mature bioactive IL-1β and IL-18 [53]. Additionally, Casp1 cleaves the protein gasdermin D (GSDMD) to generate N-terminal GSDMD (N-GSDMD) [59]. After cleavage, N-GSDMD oligomerizes in the cell membrane to form pores, allowing IL-1β and IL-18 to leave the cell and effectively execute a highly inflammatory form of cell death that is termed pyroptosis [25,60].
In the non-canonical pathway for NLRP3 inflammasome activation, intracellular LPS that has been generated by Gram-negative bacteria is directly recognized by the CARD domain of caspase-11 in mice and caspase-4/5 in humans, ultimately leading to IL-1β and IL-18 release through the activation of the NLRP3-ASC-Casp1 pathway [61,62,63,64].
The third or the alternate pathway of NLRP3 inflammasome activation exists in both human and porcine monocytes, but not in murine cells [65]. In this alternate pathway, NLRP3 assembly occurs upon the activation of toll-like receptor 4 (TLR4) in response to LPS and requires the TIR-domain-containing adapter-inducing interferon-β (TRIF)-receptor-interacting serine/threonine-protein kinase 1 (RIPK1)-Fas-associated protein with death domain (FADD)-caspase-8 signaling cascade to activate the NLRP3 inflammasome, leading again to the cleavage of pro-IL-1β to produce mature and active IL-1β [66]. In contrast to classical pathways, the alternative pathway lacks ASC speck formation, pyroptosis induction, or K+ efflux [67].
Figure 1.
Schematic illustration of the mechanism regulating NLRP3 inflammasome activation in canonical, non-canonical, and alternate pathway. Optimal activation of NLRP3 requires two steps. The first step is priming, which is initiated by extracellular PAMPs and endogenous cytokines by the PRRs, which then upregulates the NF-κB-mediated transcription of NLRP3, pro-IL-1β, and pro-IL-18. The second step is activation, which includes canonical and non-canonical pathways. The canonical pathway is triggered by multiple pathogens and inflammatory agents through a combination of important and related events such as activation of the purinergic receptor P2X7 by ATP, cathepsin release following lysozyme rupture, opening of Ca2+ channels to allow ion flux, mitochondrial dysfunction, ROS formation, Golgi apparatus disassembly, and endoplasmic reticulum stress. Once activated, oligomerization of the NLRP3 inflammasome is thought to induce conformational changes that generate active caspase-1, which converts pro-pro-IL-1β and pro-IL-18 to mature bioactive IL-1β and IL-18. Additionally, Casp1 cleaves the protein gasdermin D to generate N-terminal gasdermin D to form pores, allowing IL-1β and IL-18 to leave the cell and effectively execute a highly inflammatory form of cell death that is termed pyroptosis. In the non-canonical pathway, intracellular LPS is directly recognized by the CARD domain of caspase-11 in mice and caspase-4/5 in humans, ultimately leading to IL-1β and IL-18 release through the activation of the NLRP3-ASC-Casp1 pathway. The alternative pathway of activation is caused by TLR4 agonists like LPS, which activates the TLR4-TRIF-RIPK1-FADD-Casp8 signaling pathway. Casp8 activates the NLRP3 inflammasome but lacks ASC speck formation, pyroptosis induction, or K+ efflux.
Figure 1.
Schematic illustration of the mechanism regulating NLRP3 inflammasome activation in canonical, non-canonical, and alternate pathway. Optimal activation of NLRP3 requires two steps. The first step is priming, which is initiated by extracellular PAMPs and endogenous cytokines by the PRRs, which then upregulates the NF-κB-mediated transcription of NLRP3, pro-IL-1β, and pro-IL-18. The second step is activation, which includes canonical and non-canonical pathways. The canonical pathway is triggered by multiple pathogens and inflammatory agents through a combination of important and related events such as activation of the purinergic receptor P2X7 by ATP, cathepsin release following lysozyme rupture, opening of Ca2+ channels to allow ion flux, mitochondrial dysfunction, ROS formation, Golgi apparatus disassembly, and endoplasmic reticulum stress. Once activated, oligomerization of the NLRP3 inflammasome is thought to induce conformational changes that generate active caspase-1, which converts pro-pro-IL-1β and pro-IL-18 to mature bioactive IL-1β and IL-18. Additionally, Casp1 cleaves the protein gasdermin D to generate N-terminal gasdermin D to form pores, allowing IL-1β and IL-18 to leave the cell and effectively execute a highly inflammatory form of cell death that is termed pyroptosis. In the non-canonical pathway, intracellular LPS is directly recognized by the CARD domain of caspase-11 in mice and caspase-4/5 in humans, ultimately leading to IL-1β and IL-18 release through the activation of the NLRP3-ASC-Casp1 pathway. The alternative pathway of activation is caused by TLR4 agonists like LPS, which activates the TLR4-TRIF-RIPK1-FADD-Casp8 signaling pathway. Casp8 activates the NLRP3 inflammasome but lacks ASC speck formation, pyroptosis induction, or K+ efflux.
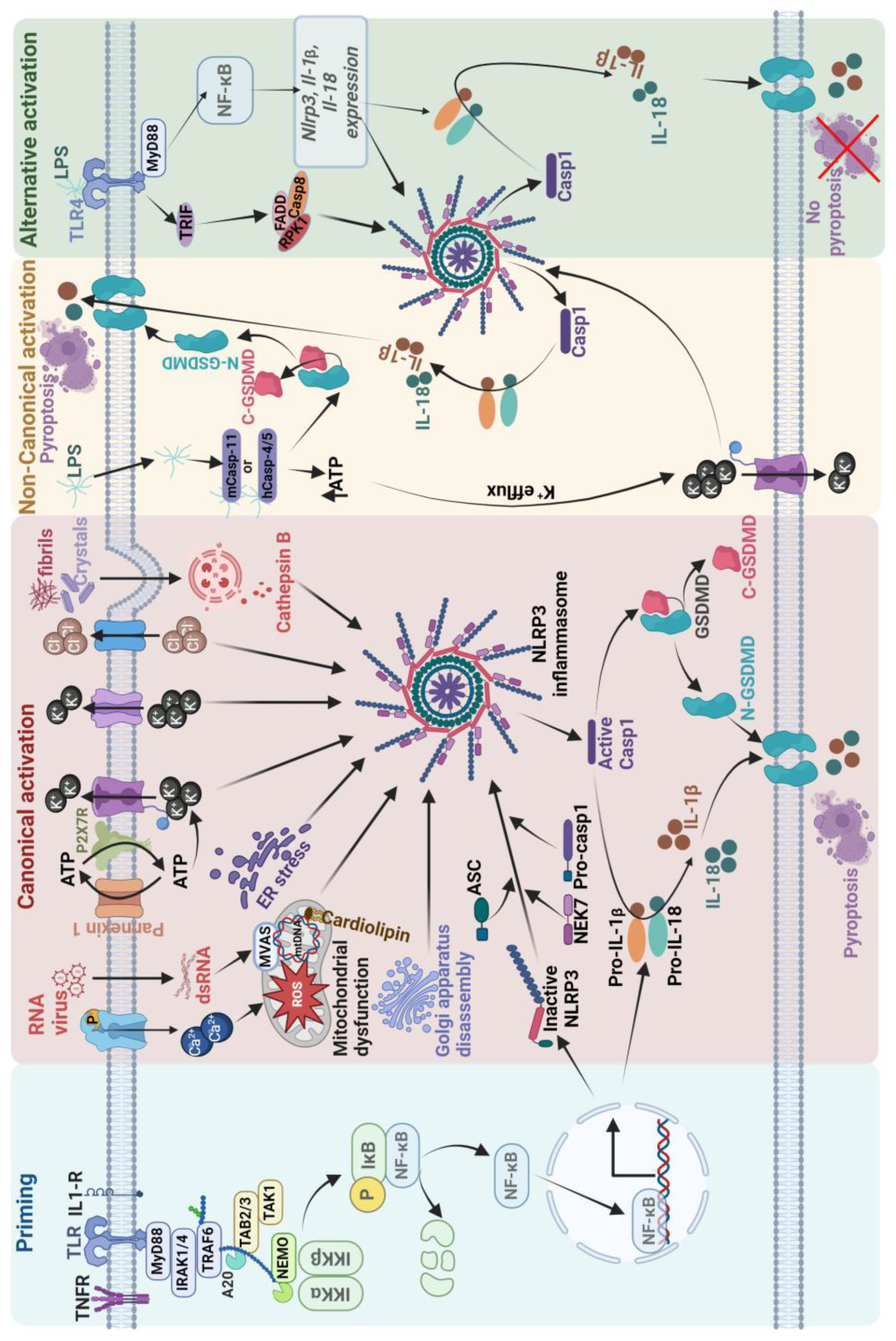
5. NLRP3 Inflammasome Involvement in Epilepsy
An increase in NLRP3 activity has been shown in the brains of individuals with epilepsy and in animal models of epilepsy [68,69,70,71,72,73,74,75]. Meng et al. first documented a significant increase in NLRP3 inflammasome in the amygdala kindling-induced murine model of seizures [70]. NLRP3 inflammasome activation in kainic acid (KA)-induced epileptic rats became evident when significant neuronal loss was noticed along with elevated levels of mature IL-1β, active caspase 1, and expression of the NLRP3 protein [68]. Magalhães et al. documented enhanced expression and activation of the NLRP3 inflammasome in organotypic slices as an ex vivo model of epilepsy [76]. In another in vivo study, NLRP3 inflammasome activation was significantly correlated with epileptic neuronal apoptosis [72]. Jiang and colleagues revealed that an increase in NLRP3 gene transcription is due to binding of Stat3 acetylation of the histone H3K9 site of the NLRP3 promoter resulting in increased NLRP3/caspase-1-mediated hippocampal neuronal cell death in epileptic mice [77]. NLRP3 inflammasome activation has also been reported in the hippocampus of pentylenetetrazole (PTZ)-induced epileptic mice and LPS-induced BV2 microglial cells [78]. Yue and colleagues revealed that the NLRP3 inflammasome components are highly expressed in neurons, microglia, and astrocytes in the epileptogenic zone of patients with temporal lobe epilepsy (TLE), a form of epilepsy associated with hippocampal neuronal atrophy, and in the hippocampi of a mouse model of status epilepticus (SE)-induced by pilocarpine [73]. Furthermore, they also demonstrated a positive correlation between increased levels of NLRP3 inflammasome proteins (NLRP3, ASC, and Caspase1) and ERS-related protein markers (GRP78, PERK, p-PERK, eIF2a, p-eIF2a, ATF4, and CHOP) in patients with TLE. Additional evidence suggests that persistent ERS may induce inflammatory processes, leading to seizures [79]. In another study, they showed that levels of RevErba, a putative down regulator of the NLRP3 inflammasome, is diminished in the leptogenic area of patients with mesial TLE (mTLE) [80]. Wu and colleagues reported that the number of NLRP3+ cells in the temporal lobe cortical tissues of refractory epilepsy patients was significantly higher than the control group [81]. They also demonstrated increased levels of NRLP3 and IL-1β in the blood of patients with refractory TLE. Furthermore, NLRP3 has been reported to be upregulated and expressed by both neuronal and glial cells in the sclerotic hippocampi of mTLE patients, which may contribute to the overexpression of hippocampal caspase -1 and IL-1β [82]. A study form Wu et al. provided the first evidence that autophagy plays a crucial role in NLRP3 inflammasome activation in the development of epilepsy [83]. NLRP3 expression was also found to be increased in children with febrile seizures [84]. A recent report by Zhang et al. confirmed that activation of NLRP3 inflammasome enhances the expression of adenosine kinase to accelerate epilepsy in mice through the CREB/REST/Spa signaling pathway [85]. Pohlentz et al. demonstrated that NLRP3 and its associated signaling molecules are activated in brain tissue samples of patients with TLE and in pilocarpine- and KA-induced SE mouse models [86].
Tan et al. demonstrated, for the first time, the elevated expression of NLRP1 and active caspase-1 in surgically resected hippocampi of patients with mTLE compared to a healthy group [33]. Furthermore, NLRP1-mediated caspase-1-dependent neuronal pyroptosis within the hippocampus was impeded in NLRP1/capase-1-deficient rats. A decreased frequency and severity of seizures was also observed in these rats. Differential gene expression analysis of the RNAseq data demonstrated the upregulation of NLRP1 mRNA in hippocampal tissues resected from patients with mTLE-hippocampal sclerosis [87]. Increased expression of NLRP1 inflammasome was observed in the sclerotic hippocampi of mTLE patients [82]. Gao et al. reported an elevated expression of the NLRP1 inflammasome in hippocampi from the pentylenetetrazol (PTZ) kindling model of epilepsy in rats [88]. Recently, the expression of NLRP1 and NLRP3 mRNAs was significantly increased in the animal model of epilepsy induced by the intrahippocampal injection of KA compared to controls [89].
6. Current Antiseizure Medications for the Treatment of Epilepsy
Since epilepsy is a complex multifaceted disease, no single ASM has emerged over time as the primary treatment. Instead, individualized treatment is given to a patient with a choice of ASM dependent upon the specific epileptic syndrome [6,87[. The majority of the 30 currently existing ASMs are effective and well-tolerated by patients [6], and are known to target either the GABAergic system or voltage-gated channels to curb the abnormal neuronal activity in the brain during seizures [90,91,92]. Some of these drugs are illustrated in Figure 2, emphasizing the diversity in their chemical structures. It is important to note that, even with the use of new ASMs, as many as 30% of epileptic patients still fail to benefit from ASM treatments (5,6, 132–135).
6.1. Currently Preferred ASM Used to Treat Epilepsy
Valproic acid, an 8-carbon 2-chain fatty acid, is the drug of choice for both adults and children suffering from epilepsy [93]. At a therapeutic range (50–100 mg/ml), it attenuates the high frequency firing of cortical and spinal cord neurons via the blockade of Na+, K+, and Ca2+channels [94]. Because of valproate-induced hepatotoxicity, ethosuximide (3-ethyl-3-methyl pyrrolidine-2,5-dione) is considered a safer alternative [95]. This drug acts to disrupt the abnormal electrical activity of the thalamocortical circuitry by blocking T-type Ca2+channels [91]. Clonazepam (1,3-Dihydro-2H-1,4-benzodiazepin-2-one), a long-acting benzodiazepine, can reduce the number of epileptic seizures experienced by epilepsy patients [96] by increasing neurotransmitter γ-aminobutyric acid (GABA) to decrease any abnormal electrical nerve activity in the CNS that might be contributing to seizures [97]. Clonazepam may lose its effectiveness over time due to the development of tolerance. One of the original ASMs, phenobarbital, is still used to prevent all types of seizures, but is not used to treat absence seizures. It exhibits its antiepileptic effect by increasing the inhibitory drive of GABA [98], and despite its sedative side effects, it is still widely used due to its low cost, particularly in developing countries [99]. A recent report suggests that levetiracetam (LEV), a novel ASM, exerts a neuroprotective effect by inhibiting the expression of proinflammatory molecules, such as IL-6, TNF- α, and IL-1β [100,101]. Lamotrigine, a synthetic phenyl triazine, is used alone or with other medications to treat epileptic seizures in children and adults [102]. Although its mechanism of action is not entirely understood, it appears to inhibit the release of excitatory neurotransmitters such as glutamate and aspartate triggered by voltage-sensitive Na+ channels and voltage-gated Ca2+channels in neurons [103]. Results from Marson et al. support the continued use of lamotrigine as the best first-line treatment option for patients newly diagnosed with focal epilepsy [104]. Topiramate, a sulfamate-substituted monosaccharide, reduces the frequency and duration of seizures and is used in the treatment of certain types of epilepsy. Topiramate’s antiepileptic effect is mediated through several mechanisms, including (i) the blockade of voltage-sensitive Na+ channels and/or Ca2+-channels, (ii) the enhancement of GABA-mediated Cl- fluxes into neurons, (iii) increases in K+ conductance, and (iv) the inhibition of glutamate-mediated neurotransmission [105]. Gabapentin, designed as a lipophilic analogue of GABA, can easily cross the BBB to increase brain synaptic GABA [106] and to suppress the influx of Ca2+ ions into neurons via voltage-dependent Ca2+ channels. Phenytoin has been used for several decades in the treatment of children with partial and generalized tonic-clonic seizures via a membrane potential-dependent blockade of Na+ channels. In addition to working through some of the mechanisms described above, zonisamide also reduces the concentration of free radicals, which may be effective against certain primarily generalized seizures, such as absence, tonic-clonic, and tonic seizures. [107]. Oxcarbazepine, a keto analog of carbamazepine, is a safer and more efficacious drug for treating partial onset seizures in both adult and children epileptic patients, and its antiepileptic activity is mediated by the blocking of neuronal ion channels [108]. Eslicarbazepine acetate, a third-generation ASM, has been proven effective when used in combination with other drugs to reduce the number of seizures in drug-resistant focal epilepsy [109].
There are several other drugs available for the treatment of epilepsy, which include lacosamide, brivaracetam, and perampanel [110]. Additionally, new treatments have been developed such as vagal nerve stimulation and ketogenic diets [111,112,113]. These new therapies and drugs have the potential to revolutionize epilepsy treatment, but much research is needed before they can be approved for wider use.
6.2. The Limitations of Current Clinical ASMs and Potential New Avenue for Drug Development in Epilepsy
Despite their effectiveness, ASMs have serious and life-threatening side effects, and a high percentage of epileptic patients develop resistance over time. Some of the generalized side effects of ASMs include depression, suicidal thoughts, mood changes, or hostility, nausea, vomiting, coordination problems, sleepiness, dizziness, and hepatotoxicity. Certain epilepsy medications can also interact with other drugs, such as pain killers or antibiotics, leading to serious consequences. The pharmacodynamic reaction of lamotrigine with carbamazepine can lead to carbamazepine intoxication [114]. Other ASMs such as phenobarbital, carbamazepine, primidone, and phenytoin can cause an imbalance in the patient’s calcium and vitamin D levels, leading to osteoporosis [115]. Due to the restrictive permeability and active efflux of some ASMs, including phenobarbital and phenytoin, the BBB limits the delivery and/or the transport of ASMs to the brain [116]. As the underlying cause(s) of epilepsy are not fully understood, the drugs developed for treating epilepsy primarily aim at controlling seizures and not at addressing the underlying cause of the disease. Given the current situation, there is a need for a more effective therapy for epilepsy.
7. NLRP3 Inflammasome in Epilepsy
Recent research suggest that increased activity of NLRP3 contributes to the development and progression of epilepsy [68,70,71,73,74,75,117]; hence, inhibiting the activity of NLRP3 may reduce inflammation-caused epileptic injuries and potentially improve symptoms [89]. NLRP3 inhibitors, a potential new class of drugs, might have fewer side effects and be more effective in a larger number of patients, and could target the underlying causes of epilepsy. Additionally, NLRP3 inhibitors have the potential to be effective in treating other inflammatory and autoimmune disorders such as gout, Alzheimer's disease, and certain cancers, which makes the development of NLRP3 inhibitors a promising area of research [118,119,120,121].
There are several approaches that can be taken to develop NLRP3 inhibitors, including:
i). Small molecule inhibitors: these are drugs that can bind to specific sites on the NLRP3 protein and prevent it from activating. These can be identified through high-throughput screening of chemical libraries; ii) Peptide inhibitors: these are short chains of amino acids that can bind to the NLRP3 protein and inhibit NLRP3 inflammasome activation. These can be identified through phage display or other peptide-based screening methods; iii). Antibodies: these are proteins that can bind to specific regions of the NLRP3 protein and prevent it from activating. These can be generated through antibody-based screening methods; iv). RNA interference: this is a method for silencing specific genes by targeting their RNA. It can be used to target the NLRP3 gene and prevent it from being expressed.
Once potential NLRP3 inhibitors have been identified, they can be further tested in cell-based assays and animal models to evaluate their efficacy and safety before moving on to clinical trials.
7.1. NLRP3 Inhibitors in Preclinical and Clinical Trial Phase
Since the role of the NLRP3 inflammasome pathway in the pathogenesis and progression of epilepsy has been well documented, the development of NLRP3 inhibitors as a potential therapeutic target for the treatment of seizures and epilepsy is urgently needed. In epileptic animal models, the knock-down of NLRP3 has been shown to reduce neuronal cell death and attenuate the chronic seizure phenotype [70,72,81]. Inhibition of NLRP3 using the pump-mediated in vivo infusion of nonviral siRNA provides neuroprotection in rats following amygdala kindling-induced SE [70]. Recent studies have reported several inhibitors that directly or indirectly target NLRP3 inflammasome and can reduce inflammation, promote neuroprotection, and decrease seizures [122,123,124]. Some of the NLRP3 inhibitors depicted in Figure 3 could help in improving our understanding of the underlying biological mechanisms that contribute to epileptic seizures and, thus, could help in improving our ability to diagnose and treat the condition. Several clinical trials are underway, exploring the efficacy and safety of NLRP3 inhibitors in patients with chronic, drug-resistant epilepsy. Some of NLRP 3 inhibitors with their nature, mechanism of action, and disease models are shown in Table 1.
MCC950 (N-[[(1,2,3,5,6,7-hexahydro-s-indacen-4-yl) amino] carbonyl]-4-(1-hydroxy-1-methylethyl)-2-furansulfonamide), also known as CP-456, 773, or cytokine release inhibitory drugs 3 (CRID3), is best characterized as a potent NLRP3 inhibitor. It blocks both canonical and non-canonical NLRP3 inflammasome activation, but no inhibitory effect has been reported on AIM2, NLRC4, or NLRP1inflammosome activation [125]. Mechanistically, MCC950 does not inhibit the priming step of NLRP3 activation as well as K+ efflux, Ca2+ flux, NLRP3–NLRP3, NEK7–NLRP3, or NLRP3–ASC interactions [125], but it directly interacts with Walker B motif of the NACHT domain with a high-affinity non-covalent interaction, blocking NLRP3 from hydrolyzing ATP to ADP and conformational changes critical for NLRP3 oligomerization and activation [126,127]. Preclinical studies have shown promise in several different types of inflammatory diseases, including Crohn’s disease [128], ulcerative colitis [129], Alzheimer’s disease [130,131], rheumatoid arthritis [132], Huntington’s disease [133], cardiovascular disease [134], and multiple sclerosis [135]. In bone marrow-derived macrophages (BMDMs), MCC950 showed inhibition of IL-1β release at IC50 of 7.5 nM, while in human monocyte-derived macrophages (HMDMs), IC50 is 8.1 nM [125,136]. MCC950 also reduces brain injury and inflammation in a mouse model of traumatic brain injury [137]. Recent evidence showed that, in an in vitro SH SY5Y model and an in vivo model of cerebral trauma induced by PTZ, the administration of MCC950 significantly provided a protective effect, and reduced epileptic neuronal apoptosis by inhibiting NLRP3 inflammasome activation [72]. A positive correlation between NLRP3 and ERS has been observed in several models of epilepsy, including temporal lobe epilepsy, and in human brain tissues from patients with epilepsy [138,139,140,141], suggesting that it may be an underlying mechanism in the development of seizures. Recently, Yue et al. demonstrated that MCC950 significantly reduced the levels of NLRP3 and the expression of ERS related markers in the hippocampi of pilocarpine-induced SE mice [73]. MCC950 has been shown to inhibit the NLRP3 inflammasome activation in KA-induced SE mice and KA-treated astrocytes [85]. Furthermore, clinical trials of MCC950 have also been initiated to evaluate its safety and efficacy [52]. MCC950 was initiated in a phase II clinical trial for rheumatoid arthritis, but it was discontinued due to liver toxicity.

Table 1.
NLRP3 inhibitors as potential epilepsy therapeutics at pre-clinical stages.
| Compound | Nature of Inhibitor | Mechanism of Action | Disease Model | References |
|---|---|---|---|---|
| MCC950 | Sulfonylurea | Directly interacts with Walker B motif of the NACHT domain, changes NLRP3 conformation. Blocks NLRP3-dependent ASC oligomerization and NLRP3 inflammasome activation Blocks ATPase activity |
In vitro SH-SY5Y model and in vivo model of cerebral trauma induced by PTZ Pilocarpine-induced SE mice KA-induced SE mice |
[72,73,85,126,127,131,137] |
| CY-09 | Glitazones | Directly binds to Cys172 residue in the Walker A motif of NLRP3 NACHT domain and inhibits NLRP3 ATPase activity | 0.83 mg/kg LPS-induced mice PTZ induced kindling mouse model |
[144,146,214] |
| Glyburide | Sulfonylurea | Suppresses KATP channels and inhibition of ASC agglomeration Blocks the assembly and activation of NLRP3 inflammasome and IL-1 release by dampening the binding of NEK7 to NLRP3 |
Pilocarpine-induced mouse model of SE Seizures induced by i.v. or i.p. PTZ models |
[149,154,155] |
| BHB | Natural products | Inhibition of K+ efflux and reduced ASC oligomerization and speck formation Inhibit NLRP3 inflammasome activation by modulating the production of ROS and by reducing the levels of ATP in the cell. |
Epileptic Kcna1-null mice 6-Hz induced seizure model of refractory epilepsy |
[161,162,163] |
| Amentoflavone | Naturally occurring bioflavonoid | Exerts neuroprotective effects by inhibiting the NLRP3 inflammasome | The chronic epilepsy model and BV2 microglial cellular inflammation model were established by PTZ kindling or LPS stimulation, respectively. | [78] |
| Semaglutide | Glucagon like peptide-1 | Decreases seizure severity, alleviated hippocampal neuronal apoptosis, ameliorated cognitive dysfunction by blocked ASC oligomerization and NLRP3 inflammasome activation | PTZ-kindled C57/BL6J mouse model and LPS induced inflammation in BV2 cells | [74] |
| Huperzine A | Naturally occurring sesquiterpene | Inhibits activation of NLRP3 inflammasome in a ROS-dependent manner |
Rat KA-induced model of epilepsy | [172] |
| Furosemide | Sulfonamide |
Increases the efficacy of valproic acid by inhibiting NLRP3 inflammasome activation | KA-induced epileptic rats | [89] |
| Ibuprofen | nonsteroidal anti-inflammatory drug (NSAID) | Exhibits antiepileptic and neuroprotective effects via inhibiting NLRP3 inflammasome activation | Rat model of PTZ-induced epilepsy | [173] |
| Rapamycin | Macrolide compound | Inhibits NLRP3inflammasome and ROS production | PTZ-kindled rats | [174] |
| Chaihu-Longgu-Muli decoction | Traditional Chinese medicine | Could significantly suppress the frequency and duration time of epileptic seizures via reducinge the expression of NLRP3, Caspase-1 TNF-α and IL-1β. | Rats with TLE | [175] |
| Parthenolide | Naturally occurring sesquiterpene lactone | Supresses NLRP3 ATPase activity by alkylating cysteine residues in ATPase domain of NLRP3 Inhibits protease activity of caspase 1 |
In vitro LPS and ATP induced NLRP3 stimulation | [177] |
| Bay 11-7082 | Sulfone | Blocks ATPase activity of NLRP3 (Juliana et al., 2010) | In vitro LPS and ATP induced NLRP3 stimulation | [177] |
| Oridonin | Natural terpenoids | Binds to Cys279 of NLRP3 NACHT domain and inhibits the interaction between NLRP3 and NEK7 thereby inhibiting the NLRP3 inflammasome activation | TBI mice | [176,215] |
| Curcumin | Natural polyphenolic compound | Inhibit IL-1b release and prevent inflammation via inhibition of NLRP3 | KA-induced epileptic syndrome in Sprague Dawley rats | [68,184] |
MCC950, 1-(1,2,3,5,6,7-Hexahydro-s-indacen-4-yl)-3-[4-(2-hydroxypropan-2-yl)furan-2-yl]sulfonylurea; NACHT, NAIP (neuronal apoptosis inhibitor protein), C2TA (MHC class 2 transcription activator), HET-E (incompatibility locus protein from Podospora anserina) and TP1 (telomerase-associated protein); NLRP3, NLR family pyrin domain containing 3; ASC, apoptosis-associated speck-like protein containing a CARD; PTZ, pentylenetetrazol; kA, kainic acid; TLE, Temporal lobe epilepsy; SE, status epilepticus, LPS, lipopolysaccharide; CY-09, 4-[[4-Oxo-2-thioxo-3-[[3-(trifluoromethyl) phenyl] methyl] -5-thiazolidinylidene] methyl] benzoic acid; NEK7, NIMA related kinase 7; BHB, beta-hydroxybutyrate; NSAID, nonsteroidal anti-inflammatory drug; ROS, reactive oxygen species; TBI, traumatic brain injury; ATP, adenosine triphosphate; IL-1β; interleukin -1beta.
CY-09 (4-[[4-Oxo-2-thioxo-3-[[3-(trifluoromethyl) phenyl] methyl] -5-thiazolidinylidene] methyl] benzoic acid) is a specific and direct inhibitor of NLRP3 that inhibited its ATPase activity and activation by binding to the Cys172 residue in the Walker A motif of the NACHT domain of NLRP3 [142]. CY-09 demonstrated favorable pharmacokinetic properties for safety, stability, and oral bioavailability. Previous studies have suggested that CY-09 could be used for the treatment of NLRP3 inflammasome-associated diseases, including type 2 diabetes, gout, thrombosis, cryopyrin-associated autoinflammatory syndrome (CAPS) mouse models, and other diseases [142,143]. Shen et al. reported that CY-09 inhibited the NLRP3 driven neuroinflammation in a PTZ-induced kindling mouse model, a chronic model of generalized seizures [144]. CY-09 repressed the expression of NLRP3, IL-1β, and IL-18 in injured brain tissue in the rat TBI models [145]. The findings from Wang et al. showed that CY-09 attenuates depression-like behaviors by inhibiting the NLRP3-mediated neuroinflammation in LPS-induced mice [146]. In a clinical trial, CY-09 was found to be effective in reducing the number and severity of seizures in people with focal epilepsy when compared to placebo. However, confirmatory studies are worth pursuing to broaden its potential in treating epilepsy.
Glyburide, a sulfonylurea also known as glibenclamide, is an FDA-approved ATP-sensitive K+ (KATP) channel inhibitor to treat type 2 diabetes mellitus [147]. In 2001, Perregaux et al. reported for the first time that glyburide inhibits IL-1β release in LPS-activated human monocytes [148]. In another study, glyburide was shown to inhibit IL-1β release during bronchial hypo responsiveness through KATP channels [149]. Glyburide has been reported to exhibit anti-inflammatory effects mainly by the inhibition of microbial ligand-induced NLRP3 inflammasome activation and IL-1β secretion by blocking KATP channels [150]. Glyburide has been shown to block NLRP3 inflammasome activity and IL-1β secretion stimulated by islet amyloid polypeptide, which is associated with type 2 diabetes [151]. In human pancreatic islets, glyburide partially reduced the increased NLRP3 and IL-1β expression induced by LPS and ATP [152]. A recent study has reported that glyburide blocked the assembly and activation of NLRP3 inflammasome and IL-1β release by dampening the binding of NEK7 to NLRP3 in ventilator-induced lung injury [153]. Research has shown that it plays a dual role in attenuating cerebral edema and improving long-term cognitive function in a pilocarpine-induced mouse model of status epilepticus [154]. Acute administration of glyburide, 30 min prior to the PTZ, significantly increased the seizure threshold in an intravenous PTZ model of mice [155].
Beta-hydroxybutyrate (BHB), one of the ketone bodies, has been shown to reduce inflammatory cytokines’ release mediated by NLRP3. It has been studied for its potential therapeutic benefits in various inflammatory diseases, including epilepsy [156,157,158,159,160]. The mechanism of action by which BHB acts as an NLRP3 inhibitor is not fully understood. However, it is thought that BHB may inhibit NLRP3 inflammasome activation by modulating the production of reactive oxygen species (ROSs) and by reducing the levels of ATP in the cell. Beta-hydroxybutyrate also inhibits the NLRP3 inflammasome by preventing K+ efflux and reducing ASC oligomerization and speck formation [161]. Kim et al. reported that BHB reduced the spontaneous recurrent seizures in spontaneously epileptic Kcna1-null mice [162]. Furthermore, BHB decreased the seizure duration and frequency in a 6-Hz-induced seizure model of refractory epilepsy [163]. It is important to note that more research is needed to fully understand the mechanism of action and potential clinical applications of BHB as an NLRP3 inhibitor.
RRx-001 (1-bromoacetyl-3,3-dinitroazetidine) was initially developed as an anticancer agent by the aerospace industry [164], but has been extensively studied in in vitro and in vivo models of several inflammatory diseases including Alzheimer’s disease, stroke, multiple sclerosis, pulmonary fibrosis, and IBD [165]. In a randomized Phase 2 trial called PREVLAR; NCT03699956, RRx-001 administration in 53 first-line head-and-neck cancer patients dramatically improved the incidence, duration, time to onset, and severity of oral mucositis. RRx-001 has been safely evaluated in clinical trials, including in an ongoing phase 3 trial for the treatment of small cell lung cancer (REPLATINUM; NCT03699956) [164]. RRx-001 is a highly selective and the most clinically advanced small molecule NLRP3 inhibitor that has been safely evaluated in over 300 patients [164]. Mechanistically, RRx-001 covalently binds to cysteine 409 of NLRP3 on the central NACHT domain of NLRP3, which inhibits the assembly and activation of the NLRP3 inflammasome [164,165]. The BBB-penetrant nature of RRx-001 inhibitor and the preclinical assessment of this inhibitor in various neurodegenerative diseases [166] advances the possibility of this uncharged small molecule inhibitor being tested soon in epilepsy.
Several other classes of compounds that have been explored for NLRP3 inflammasome inhibition include flavonoids, chalcone, and boron-based compounds. Amentoflavone, a naturally occurring bioflavonoid [78], and semaglutide, a glucagon-like peptide-1 [74], were reported to affect epileptogenesis and reduce seizures via their neuroprotective effects because of NLRP3 inflammasome inhibition in PTZ-kindled mice. Sun et al. revealed that endogenous as well as exogenous IL-10 downregulates IL-1β production in microglia in mice exposed to epileptogenic injury thorough the STAT3-dependent inhibition of NLRP3 inflammasome activity [167].
Licochalcone B, Isoliquiritigenin, and Cardamomin are natural chalcone-based compounds that have shown promising NLRP3 inflammasome inhibitory effects. Licochalcone B binds with NEK7, preventing the interaction with NLRP3 which is important for NLRP3 inflammasome activation [168]. Isoliquiritigenin isolated from Glycyrrhiza uralensis has been reported to activate the Nrf2-mediated antioxidant signaling, preventing the activation of NF-κB and NLRP3 inflammasome [169,170]. Cardamomin reduced the protein levels of NLRP3, Casp1, and IL-1 in 2,4,6-Trinitrobenzenesulfonic acid (TNBS)-induced colitis mice [171]. BC7, BC23, and NBC6 are potent oxanorbornene molecules developed as potent NLRP3 inflammasome inhibitors. Among these three molecules, compound NBC6 showed the most potent inhibition of IL-1β release from THP-1 monocytes with an IC50 of 574 nM. BC7 and BC23 showed IC50 values of 1.16 μM and 2.29 μM, respectively, for the inhibition of IL-1β release [13,14].
Huperzine A, a naturally occurring sesquiterpene alkaloid and valproic acid, and one of the most prescribed medications against epilepsy, has been shown to inhibit activation of the NLRP3 inflammasome in the rat KA-induced model of epilepsy in a ROS-dependent manner [172]. Recently, furosemide (4-chloro-5-sulphonyl-N-furfuryl-anthranilic acid), a diuretic drug, has been shown to decrease the NLRP3 as well as NLRP1 level significantly when treated in combination with valproic acid in KA-induced epileptic rats [89].
Studies from Li and coworkers have reported that ibuprofen may have antiepileptic and neuroprotective effects in the rat model of PTZ-induced epilepsy via inhibiting NLRP3 inflammasome activation [173]. Rapamycin, an inhibitor of mTOR signaling, has been reported to alleviate the symptoms of seizures, anxiety, and depression in PTZ-kindled rats by inhibiting NLRP3 inflammasomes and ROS production [174]. Chaihu-Longgu-Muli decoction, a well-known ancient formula in traditional Chinese medicine, could significantly reduce the frequency and duration time of epileptic seizures, and inhibit the expression of NLRP3, TNF-α, Caspase-1, and IL-1β [175]. Natural products such as parthenolide and oridonin from Rabdosia rubescens were also able to inhibit the NLRP3 inflammasome [176,177]. It was observed that oridonin binds covalently with the Cys279 of the NLRP3 NACHT domain. This binding prevents the interaction between NLRP3 and NEK7, which is essential for NLRP3 inflammasome assembly and activation [176]. Recently, oridonin has been shown to rescue kanamycin-related hearing loss by inhibiting NLRP3 inflammasome activation [153,178]. BAY 11-7082, an NF-κB inhibitor, has also been reported to inhibit the NLRP3 ATPase activity in macrophages, independently of their inhibitory effect on NF-kB activity [177].
Curcumin ((1E,6E)-1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione)), a polyphenolic compound present in turmeric (Curcuma longa), exhibits antioxidant, anti-inflammatory, and neuroprotective properties, and its beneficial effect on epilepsy has been shown in many preclinical studies [179,180,181,182,183]. Curcumin has been reported to inhibit IL-1β release and prevent inflammation via the inhibition of NLRP3 [184] and suppressed KA-induced epileptic syndrome via inhibiting NLRP3 inflammasome activation in Sprague Dawley rats [68].
MicroRNAs (miRNAs) are endogenous ~20–23 nucleotide-long non-coding RNAs that bind to the 3’ untranslated region (3’UTR) of protein-coding mRNAs to regulate their translation and, thus, can have significant impacts on cellular processes. Several miRNAs that can target different components of the NLRP3 inflammasome and modulate its activity have been identified. For example, miR-223 has been shown to target the NLRP3 inflammasome in the brain and reduce neuroinflammation and neuronal damage [185], inhibit NLRP3 expression, and reduce inflammation in various disease models, including arthritis, atherosclerosis, and myocardial infarction [56,186,187]. miR-29c reduces the inflammatory response of microglia by modulating the NLRP3 inflammasome [188], miR-17-5p ameliorated NLRP3 inflammasome activation-mediated hypoxic–ischemic brain injury in rat [189], and miR-138-5p overexpression in epileptic neurons inhibits NLRP3 by directly binding with ubiquitin-specific peptidase 47 (USP47), a positive regulator of NLRP3 [190]. miR-29a-5p mimics protective TBI-induced BBB dysfunction via suppressing NLRP3 inflammasome activation [191]. Other miRNAs, such as miR-23a, miR-let-7e, miR-30e, and miR-223, have also been found to inhibit the NLRP3 inflammasome and reduce inflammation [192]. Overall, the use of miRNAs as NLRP3 inhibitors holds promise as a potential therapeutic approach for treating a range of inflammatory and autoimmune diseases. However, more research is needed to fully understand the mechanisms underlying miRNA regulation of NLRP3 and to develop effective and safe miRNA-based therapies for different diseases.
7.2. NLRP3 Inflammasome Inhibitors and Their Limitations as Remedial Strategies
NLRP3 inhibitors have been investigated as potential drugs for the treatment of various inflammatory and autoimmune diseases. Off-target effects are the significant drawback of using NLRP3 inhibitors. For example, MCC950 at high micromolar concentrations could inhibit carbonic anhydrase 2 [193] and block Cl-1 efflux from nigericin-activated macrophages [142]. CY-09 has been reported to affect cytochrome P450 enzymes [123]. Another inhibitor, oridonin, has several targets, e.g., AKT/2, c-Myc, p39, and MAPK [194]. Since NLRP3 is involved in many important cellular processes, including immune defense and tissue repair, blocking its activity could lead to unintended consequences including impairing pathogen clearance and thus increasing the risk of infection. MCC950 has been examined in a phase II clinical trial for the treatment of rheumatoid arthritis, but the trial was discontinued due to hepatic toxicity [52]. NLRP3 plays a crucial role in the immune system, and blocking its activity may also interfere with the body’s ability to fight infections and heal wounds.
Future studies should take advantage of available cryo-EM and crystal structures of NLRP3 bound to NEK7 [46], and focus on the development of structure-guided direct inhibitors with improved specificity and potency. Recently, Agarwal et al. rationally designed MCC950-derivative compounds [179,195,196]. These compounds were found to be potent and selective NLRP3 inhibitors with a good pharmacokinetic profile and high oral bioavailability in mice. In addition, another NLRP3 inhibitor, NT-0796, boasts innovative chemistry, delivering unparalleled potency and the promise of an extended pharmacodynamic impact. Furthermore, it demonstrates the capability to penetrate the BBB [197]. Since nanoparticle (NP)-based drug delivery is an emerging area of research in the field of nanomedicine and immunotherapy due to their intriguing properties such as target site specificity, systemic stability, and low toxicity [198], Mancuso et al. analyzed the effect of glyburide-loaded nanovesicles (GNVs) on NLRP3 inflammasome activation in a LPS and nigericin-activated THP-1 cell model [199]. Their results confirm that GNVs were able to inhibit IL-1β secretions more efficiently than free glyburide. Recently, Kulkarni and coworkers synthesized and analyzed MCC950-loaded nanoparticles (MCC NPs) and found that MCC NPs showed a significant reduction in IL-1β secretions in vitro and in vivo [200]. Tang et al. engineered a unique delivery system, VHPK-PLGA@COL, incorporating colchicine and demonstrating enhanced biosafety and prolonged drug release. This was validated both in cell culture and in animal models [201]. The inhibitory effect of VHPK-PLGA@COL on NLRP3 and its downstream molecules was more significant than that of free colchicine. Exosome-like nanoparticles from ginger rhizomes strongly inhibited NLRP3 inflammasome activation [202]. Another group prepared garlic chive-derived vesicle-like nanoparticles which exhibit potent anti-NLRP3 inflammasome activity [203]. Moreover, dexamethasone-loaded ROS-responsive polymer nanoparticles prepared by a modified emulsion approach had a strong ability to inhibit the expression of NLRP3, caspase1, and IL-1β [204]. It has been reported that nanoparticles themself trigger NLRP3 inflammasome activation [205,206,207,208]. However, the formation of a protein corona layer on lipid NPs caused a significant reduction in NLRP3 inflammasome activation and controlled the toxicity, biodistribution, and cellular uptake [209]. Chalcones are natural compounds with an α, β unsaturated carbonyl group (Michael acceptor) found in many plants and have gained attention for their medicinal properties [210,211]. We and others investigated some chalcones for their potential as NLRP3 inhibitors [212,213]. Our preliminary data encourage the further development of more potent NLRP3 inhibitors based on this chalcone scaffold, which could lead to the development of novel treatments for epilepsy and other inflammatory diseases.
8. Conclusions and Perspectives
Scientific evidence supports the role of NLRP3 in epileptic seizures and the use of NLRP3 inhibitors seems promising in understanding the biological mechanisms behind epilepsy, leading to the development of biomarkers for early detection and more targeted, effective treatments to reduce seizures in high-risk individuals with epilepsy. Clinical trials are underway to investigate the efficacy and safety of these inhibitors in humans, and if these trials are successful, NLRP3 inhibitors may eventually become standard medical care for patients with epilepsy.
However, there is still much to be investigated before NLRP3 inhibitors are officially approved for the treatment of epilepsy.
Author Contributions
Conceptualization, V.S. I.H. and M.S.; validation, I.H. P.T., R.S. M.S. and VS; investigation, V.S., I.H. and P.T.; data curation, I.H., P.T. and D.M.B., JZ; writing—original draft preparation, I.H. and D.M.B.; writing (editing and reviewing)—I.H., P.T., D.M.B., J.Z., R.S., M.S. V.S..; visualization, I.H., P.T. and V.S.; supervision, I.H. and V.S.; project administration, R.M. and M.S.; funding acquisition, R.M., M.S. and V.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the Research Department of the Kansas City VA Medical Center (KCVAMC) and Midwest Veterans Biomedical Research Foundation (MVBRF) at KCVAMC, Kansas City, MO, 64128.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We would like to thank the research and development office staff for their valuable and helpful comments on this manuscript. Graphical abstracts and figures were created on Scientific Image and Illustration Software|BioRender. The authors apologize to investigators whose contributions could not be appropriately cited because of space limitations.
Conflicts of Interest
The authors declare no conflicts of interest.
Abbreviations
| ASM | Antiseizure medication |
| SRS | Spontaneous recurrent seizure |
| BBB | Blood-brain barrier |
| TBI | Traumatic brain injury |
| DAMP | Damage-associated molecular pattern |
| PAMP | Pathogen-associated molecular pattern |
| ATP | Adenosine triphosphate |
| HMGB1 | High mobility group box1 |
| ROS | Reactive oxygen species |
| CARD | Caspase-recruitment domain |
| ASC | Apoptosis-associated speck-like protein containing a CARD. |
| PRR | Pattern-recognition receptor |
| NBD | Nucleotide binding domain |
| LRR-CR | Leucine-rich-repeat-containing receptor. |
| AIM2 | Absent-in-melanoma 2 |
| PYD | Pyrin domain |
| NOD | Nucleotide-binding oligomerization domain |
| NLR | Nod-like receptor |
| NLRP | Nod-like receptor protein |
| NLRP3 | NLR family pyrin domain containing 3. |
| NLRC4 | NLR family CARD domain-containing protein 4 |
| CNS | Central nervous system |
| NACHT | NAIP (neuronal apoptosis inhibitor protein), C2TA (MHC class 2 transcription activator), HET-E (incompatibility locus protein from Podospora anserina) and TP1 (telomerase-associated protein). |
| HD | Helical domain |
| WHD | Winged helix domain |
| NIMA | Never in mitosis A |
| NEK7 | NIMA related kinase 7 |
| NF-κB | Nuclear factor kappa-light-chain-enhancer of activated B cells |
| EM | Electron microscopy |
| IL | Interleukin |
| LPS | Lipopolysaccharide |
| miRNA | MicroRNA |
| P2RX7 | P2X purinoceptor 7 |
| ERS | Endoplasmic reticulum stress |
| GSDMD | Gasdermin D |
| TLR | Toll-like receptors |
| TRIF | TIR-domain-containing adapter-inducing interferon-β |
| RIPK1 | Receptor-interacting serine/threonine-protein kinase 1 |
| FADD | Fas-associated protein with death domain |
| KA | Kainic acid |
| TLE | Temporal lobe epilepsy |
| SE | Status epilepticus |
| PTZ | Pentylenetetrazol |
| GABA | γ-aminobutyric acid |
| CAPS | Cryopyrin-associated periodic syndromes. |
| MCC950 | 1-(1,2,3,5,6,7-Hexahydro-s-indacen-4-yl)-3-[4-(2-hydroxypropan-2-yl)furan-2-yl]sulfonylurea |
| BMDM | Bone marrow-derived macrophage |
| HMDM | Human monocyte derived macrophage |
| CY-09 | 4-[[4-Oxo-2-thioxo-3-[[3-(trifluoromethyl) phenyl] methyl] -5-thiazolidinylidene] methyl] benzoic acid |
| FDA | Food and drug administration |
| BHB | Beta-hydroxybutyrate |
| STAT3 | Signal transducer and activator of transcription 3 |
References
- Loscher, W.; Klitgaard, H.; Twyman, R.E.; Schmidt, D. New avenues for anti-epileptic drug discovery and development. Nat Rev Drug Discov 2013, 12, 757-776. [CrossRef]
- Lukawski, K.; Gryta, P.; Luszczki, J.; Czuczwar, S.J. Exploring the latest avenues for antiepileptic drug discovery and development. Expert Opin Drug Discov 2016, 11, 369-382. [CrossRef]
- Hong, Y.; Wei, C.; Fu, M.; Li, X.; Zhang, H.; Yao, B. MCC950 alleviates seizure severity and angiogenesis by inhibiting NLRP3/ IL-1beta signaling pathway-mediated pyroptosis in mouse model of epilepsy. Int Immunopharmacol 2024, 126, 111236. [CrossRef]
- Mesraoua, B.; Abou-Khalil, B.; Hosni Khodair, R.; Melikyan, G.; Al Hail, H.; Asadi-Pooya, A.A. Seizure clusters. J Drug Assess 2021, 10, 86-90. [CrossRef]
- Guiard, B.P.; Di Giovanni, G. Central serotonin-2A (5-HT2A) receptor dysfunction in depression and epilepsy: the missing link? Front Pharmacol 2015, 6, 46. [CrossRef]
- Henning, O.; Medalen, T.E.M.; Nakken, K.O.; Lossius, M.I. How often do doctors discuss drug withdrawal with their seizure-free patients with epilepsy? Epilepsy Behav 2020, 108, 107095. [CrossRef]
- Loscher, W.; Klein, P. The Pharmacology and Clinical Efficacy of Antiseizure Medications: From Bromide Salts to Cenobamate and Beyond. CNS Drugs 2021, 35, 935-963. [CrossRef]
- Lerche, H. Drug-resistant epilepsy - time to target mechanisms. Nat Rev Neurol 2020, 16, 595-596. [CrossRef]
- Alyu, F.; Dikmen, M. Inflammatory aspects of epileptogenesis: contribution of molecular inflammatory mechanisms. Acta Neuropsychiatr 2017, 29, 1-16. [CrossRef]
- Goldberg, E.M.; Coulter, D.A. Mechanisms of epileptogenesis: a convergence on neural circuit dysfunction. Nat Rev Neurosci 2013, 14, 337-349. [CrossRef]
- Sung, P.S.; Lin, P.Y.; Liu, C.H.; Su, H.C.; Tsai, K.J. Neuroinflammation and Neurogenesis in Alzheimer's Disease and Potential Therapeutic Approaches. Int J Mol Sci 2020, 21, 701. [CrossRef]
- Fuster-Matanzo, A.; Llorens-Martin, M.; Hernandez, F.; Avila, J. Role of neuroinflammation in adult neurogenesis and Alzheimer disease: therapeutic approaches. Mediators Inflamm 2013, 2013, 260925. [CrossRef]
- Yong, H.Y.F.; Rawji, K.S.; Ghorbani, S.; Xue, M.; Yong, V.W. The benefits of neuroinflammation for the repair of the injured central nervous system. Cell Mol Immunol 2019, 16, 540-546. [CrossRef]
- Zhou, Q.; Lin, L.; Li, H.; Wang, H.; Jiang, S.; Huang, P.; Lin, Q.; Chen, X.; Deng, Y. Melatonin Reduces Neuroinflammation and Improves Axonal Hypomyelination by Modulating M1/M2 Microglia Polarization via JAK2-STAT3-Telomerase Pathway in Postnatal Rats Exposed to Lipopolysaccharide. Mol Neurobiol 2021, 58, 6552-6576. [CrossRef]
- Bollaerts, I.; Van Houcke, J.; Andries, L.; De Groef, L.; Moons, L. Neuroinflammation as Fuel for Axonal Regeneration in the Injured Vertebrate Central Nervous System. Mediators Inflamm 2017, 2017, 9478542. [CrossRef]
- Shi, Y.; Wei, B.; Li, L.; Wang, B.; Sun, M. Th17 cells and inflammation in neurological disorders: Possible mechanisms of action. Front Immunol 2022, 13, 932152. [CrossRef]
- Soltani Khaboushan, A.; Yazdanpanah, N.; Rezaei, N. Neuroinflammation and Proinflammatory Cytokines in Epileptogenesis. Mol Neurobiol 2022, 59, 1724-1743. [CrossRef]
- Vezzani, A.; Balosso, S.; Ravizza, T. Neuroinflammatory pathways as treatment targets and biomarkers in epilepsy. Nat Rev Neurol 2019, 15, 459-472. [CrossRef]
- Rana, A.; Musto, A.E. The role of inflammation in the development of epilepsy. J Neuroinflammation 2018, 15, 144. [CrossRef]
- Mahfoz, A.M.; Shahzad, N. Neuroinflammation impact in epileptogenesis and new treatment strategy. Behav Pharmacol 2019, 30, 661-675. [CrossRef]
- Banjara, M.; Ghosh, C. Sterile Neuroinflammation and Strategies for Therapeutic Intervention. Int J Inflam 2017, 2017, 8385961. [CrossRef]
- Relja, B.; Land, W.G. Damage-associated molecular patterns in trauma. Eur J Trauma Emerg Surg 2020, 46, 751-775. [CrossRef]
- Gong, T.; Liu, L.; Jiang, W.; Zhou, R. DAMP-sensing receptors in sterile inflammation and inflammatory diseases. Nat Rev Immunol 2020, 20, 95-112. [CrossRef]
- Fleshner, M.; Frank, M.; Maier, S.F. Danger Signals and Inflammasomes: Stress-Evoked Sterile Inflammation in Mood Disorders. Neuropsychopharmacology 2017, 42, 36-45. [CrossRef]
- Martinon, F.; Burns, K.; Tschopp, J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Mol Cell 2002, 10, 417-426. [CrossRef]
- von Moltke, J.; Ayres, J.S.; Kofoed, E.M.; Chavarria-Smith, J.; Vance, R.E. Recognition of bacteria by inflammasomes. Annu Rev Immunol 2013, 31, 73-106. [CrossRef]
- Hoffmann, S.; Beyer, C. A Fatal Alliance between Microglia, Inflammasomes, and Central Pain. Int J Mol Sci 2020, 21, 3764. [CrossRef]
- Trendelenburg, G. Acute neurodegeneration and the inflammasome: central processor for danger signals and the inflammatory response? J Cereb Blood Flow Metab 2008, 28, 867-881. [CrossRef]
- Mortezaee, K.; Khanlarkhani, N.; Beyer, C.; Zendedel, A. Inflammasome: Its role in traumatic brain and spinal cord injury. J Cell Physiol 2018, 233, 5160-5169. [CrossRef]
- Slowik, A.; Lammerding, L.; Hoffmann, S.; Beyer, C. Brain inflammasomes in stroke and depressive disorders: Regulation by oestrogen. J Neuroendocrinol 2018, 30, e12482. [CrossRef]
- Walsh, J.G.; Muruve, D.A.; Power, C. Inflammasomes in the CNS. Nat Rev Neurosci 2014, 15, 84-97. [CrossRef]
- Yap, J.K.Y.; Pickard, B.S.; Chan, E.W.L.; Gan, S.Y. The Role of Neuronal NLRP1 Inflammasome in Alzheimer's Disease: Bringing Neurons into the Neuroinflammation Game. Mol Neurobiol 2019, 56, 7741-7753. [CrossRef]
- Tan, C.C.; Zhang, J.G.; Tan, M.S.; Chen, H.; Meng, D.W.; Jiang, T.; Meng, X.F.; Li, Y.; Sun, Z.; Li, M.M.; et al. NLRP1 inflammasome is activated in patients with medial temporal lobe epilepsy and contributes to neuronal pyroptosis in amygdala kindling-induced rat model. J Neuroinflammation 2015, 12, 18. [CrossRef]
- de Rivero Vaccari, J.P.; Lotocki, G.; Alonso, O.F.; Bramlett, H.M.; Dietrich, W.D.; Keane, R.W. Therapeutic neutralization of the NLRP1 inflammasome reduces the innate immune response and improves histopathology after traumatic brain injury. J Cereb Blood Flow Metab 2009, 29, 1251-1261. [CrossRef]
- de Rivero Vaccari, J.P.; Dietrich, W.D.; Keane, R.W. Activation and regulation of cellular inflammasomes: gaps in our knowledge for central nervous system injury. J Cereb Blood Flow Metab 2014, 34, 369-375. [CrossRef]
- Minkiewicz, J.; de Rivero Vaccari, J.P.; Keane, R.W. Human astrocytes express a novel NLRP2 inflammasome. Glia 2013, 61, 1113-1121. [CrossRef]
- Ducza, L.; Szucs, P.; Hegedus, K.; Bakk, E.; Gajtko, A.; Weber, I.; Hollo, K. NLRP2 Is Overexpressed in Spinal Astrocytes at the Peak of Mechanical Pain Sensitivity during Complete Freund Adjuvant-Induced Persistent Pain. Int J Mol Sci 2021, 22, 1408. [CrossRef]
- Zhang, Z.; Guo, P.; Huang, S.; Jia, Z.; Chen, T.; Liu, X.; Feng, H.; Chen, Y. Inhibiting Microglia-Derived NLRP3 Alleviates Subependymal Edema and Cognitive Dysfunction in Posthemorrhagic Hydrocephalus after Intracerebral Hemorrhage via AMPK/Beclin-1 Pathway. Oxid Med Cell Longev 2022, 2022, 4177317. [CrossRef]
- Shi, F.; Yang, Y.; Kouadir, M.; Fu, Y.; Yang, L.; Zhou, X.; Yin, X.; Zhao, D. Inhibition of phagocytosis and lysosomal acidification suppresses neurotoxic prion peptide-induced NALP3 inflammasome activation in BV2 microglia. J Neuroimmunol 2013, 260, 121-125. [CrossRef]
- Hanslik, K.L.; Ulland, T.K. The Role of Microglia and the Nlrp3 Inflammasome in Alzheimer's Disease. Front Neurol 2020, 11, 570711. [CrossRef]
- Gustin, A.; Kirchmeyer, M.; Koncina, E.; Felten, P.; Losciuto, S.; Heurtaux, T.; Tardivel, A.; Heuschling, P.; Dostert, C. NLRP3 Inflammasome Is Expressed and Functional in Mouse Brain Microglia but Not in Astrocytes. PLoS One 2015, 10, e0130624. [CrossRef]
- Lim, J.; Kim, M.J.; Park, Y.; Ahn, J.W.; Hwang, S.J.; Moon, J.S.; Cho, K.G.; Kwack, K. Upregulation of the NLRC4 inflammasome contributes to poor prognosis in glioma patients. Sci Rep 2019, 9, 7895. [CrossRef]
- Freeman, L.; Guo, H.; David, C.N.; Brickey, W.J.; Jha, S.; Ting, J.P. NLR members NLRC4 and NLRP3 mediate sterile inflammasome activation in microglia and astrocytes. J Exp Med 2017, 214, 1351-1370. [CrossRef]
- Wu, P.J.; Liu, H.Y.; Huang, T.N.; Hsueh, Y.P. AIM 2 inflammasomes regulate neuronal morphology and influence anxiety and memory in mice. Sci Rep 2016, 6, 32405. [CrossRef]
- Adamczak, S.E.; de Rivero Vaccari, J.P.; Dale, G.; Brand, F.J., 3rd; Nonner, D.; Bullock, M.R.; Dahl, G.P.; Dietrich, W.D.; Keane, R.W. Pyroptotic neuronal cell death mediated by the AIM2 inflammasome. J Cereb Blood Flow Metab 2014, 34, 621-629. [CrossRef]
- Sharif, H.; Wang, L.; Wang, W.L.; Magupalli, V.G.; Andreeva, L.; Qiao, Q.; Hauenstein, A.V.; Wu, Z.; Nunez, G.; Mao, Y.; et al. Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome. Nature 2019, 570, 338-343. [CrossRef]
- He, Y.; Zeng, M.Y.; Yang, D.; Motro, B.; Nunez, G. NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 2016, 530, 354-357. [CrossRef]
- Shi, H.; Wang, Y.; Li, X.; Zhan, X.; Tang, M.; Fina, M.; Su, L.; Pratt, D.; Bu, C.H.; Hildebrand, S.; et al. NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat Immunol 2016, 17, 250-258. [CrossRef]
- Liu, G.; Chen, X.; Wang, Q.; Yuan, L. NEK7: a potential therapy target for NLRP3-related diseases. Biosci Trends 2020, 14, 74-82. [CrossRef]
- Xu, J.; Lu, L.; Li, L. NEK7: a novel promising therapy target for NLRP3-related inflammatory diseases. Acta Biochim Biophys Sin (Shanghai) 2016, 48, 966-968. [CrossRef]
- El-Sharkawy, L.Y.; Brough, D.; Freeman, S. Inhibiting the NLRP3 Inflammasome. Molecules 2020, 25. [CrossRef]
- Mangan, M.S.J.; Olhava, E.J.; Roush, W.R.; Seidel, H.M.; Glick, G.D.; Latz, E. Targeting the NLRP3 inflammasome in inflammatory diseases. Nat Rev Drug Discov 2018, 17, 588-606. [CrossRef]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kappaB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J Immunol 2009, 183, 787-791. [CrossRef]
- Wei, M.; Wang, L.; Wu, T.; Xi, J.; Han, Y.; Yang, X.; Zhang, D.; Fang, Q.; Tang, B. NLRP3 Activation Was Regulated by DNA Methylation Modification during Mycobacterium tuberculosis Infection. Biomed Res Int 2016, 2016, 4323281. [CrossRef]
- Haneklaus, M.; O'Neil, J.D.; Clark, A.R.; Masters, S.L.; O'Neill, L.A.J. The RNA-binding protein Tristetraprolin (TTP) is a critical negative regulator of the NLRP3 inflammasome. J Biol Chem 2017, 292, 6869-6881. [CrossRef]
- Bauernfeind, F.; Rieger, A.; Schildberg, F.A.; Knolle, P.A.; Schmid-Burgk, J.L.; Hornung, V. NLRP3 inflammasome activity is negatively controlled by miR-223. J Immunol 2012, 189, 4175-4181. [CrossRef]
- Seok, J.K.; Kang, H.C.; Cho, Y.Y.; Lee, H.S.; Lee, J.Y. Regulation of the NLRP3 Inflammasome by Post-Translational Modifications and Small Molecules. Front Immunol 2020, 11, 618231. [CrossRef]
- Schroder, K.; Zhou, R.; Tschopp, J. The NLRP3 inflammasome: a sensor for metabolic danger? Science 2010, 327, 296-300. [CrossRef]
- de Vasconcelos, N.M.; Lamkanfi, M. Recent Insights on Inflammasomes, Gasdermin Pores, and Pyroptosis. Cold Spring Harb Perspect Biol 2020, 12, a036392. [CrossRef]
- Fernandes-Alnemri, T.; Wu, J.; Yu, J.W.; Datta, P.; Miller, B.; Jankowski, W.; Rosenberg, S.; Zhang, J.; Alnemri, E.S. The pyroptosome: a supramolecular assembly of ASC dimers mediating inflammatory cell death via caspase-1 activation. Cell Death Differ 2007, 14, 1590-1604. [CrossRef]
- Kayagaki, N.; Stowe, I.B.; Lee, B.L.; O'Rourke, K.; Anderson, K.; Warming, S.; Cuellar, T.; Haley, B.; Roose-Girma, M.; Phung, Q.T.; et al. Caspase-11 cleaves gasdermin D for non-canonical inflammasome signalling. Nature 2015, 526, 666-671. [CrossRef]
- Yi, Y.S. Caspase-11 Non-Canonical Inflammasome: Emerging Activator and Regulator of Infection-Mediated Inflammatory Responses. Int J Mol Sci 2020, 21, 2736. [CrossRef]
- Vigano, E.; Diamond, C.E.; Spreafico, R.; Balachander, A.; Sobota, R.M.; Mortellaro, A. Human caspase-4 and caspase-5 regulate the one-step non-canonical inflammasome activation in monocytes. Nat Commun 2015, 6, 8761. [CrossRef]
- Santos, J.C.; Dick, M.S.; Lagrange, B.; Degrandi, D.; Pfeffer, K.; Yamamoto, M.; Meunier, E.; Pelczar, P.; Henry, T.; Broz, P. LPS targets host guanylate-binding proteins to the bacterial outer membrane for non-canonical inflammasome activation. EMBO J 2018, 37, e98089. [CrossRef]
- Gaidt, M.M.; Ebert, T.S.; Chauhan, D.; Schmidt, T.; Schmid-Burgk, J.L.; Rapino, F.; Robertson, A.A.; Cooper, M.A.; Graf, T.; Hornung, V. Human Monocytes Engage an Alternative Inflammasome Pathway. Immunity 2016, 44, 833-846. [CrossRef]
- Gurung, P.; Kanneganti, T.D. Novel roles for caspase-8 in IL-1beta and inflammasome regulation. Am J Pathol 2015, 185, 17-25. [CrossRef]
- Yang, Y.; Wang, H.; Kouadir, M.; Song, H.; Shi, F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis 2019, 10, 128. [CrossRef]
- He, Q.; Jiang, L.; Man, S.; Wu, L.; Hu, Y.; Chen, W. Curcumin Reduces Neuronal Loss and Inhibits the NLRP3 Inflammasome Activation in an Epileptic Rat Model. Curr Neurovasc Res 2018, 15, 186-192. [CrossRef]
- Lin, W.S.; Hsu, T.R. Hypothesis: Febrile infection-related epilepsy syndrome is a microglial NLRP3 inflammasome/IL-1 axis-driven autoinflammatory syndrome. Clin Transl Immunology 2021, 10, e1299. [CrossRef]
- Meng, X.F.; Tan, L.; Tan, M.S.; Jiang, T.; Tan, C.C.; Li, M.M.; Wang, H.F.; Yu, J.T. Inhibition of the NLRP3 inflammasome provides neuroprotection in rats following amygdala kindling-induced status epilepticus. J Neuroinflammation 2014, 11, 212. [CrossRef]
- Xiang, T.; Luo, X.; Ye, L.; Huang, H.; Wu, Y. Klotho alleviates NLRP3 inflammasome-mediated neuroinflammation in a temporal lobe epilepsy rat model by activating the Nrf2 signaling pathway. Epilepsy Behav 2022, 128, 108509. [CrossRef]
- Shen, K.; Mao, Q.; Yin, X.; Zhang, C.; Jin, Y.; Deng, A.; Gu, Z.; Chen, B. NLRP3 Inflammasome Activation Leads to Epileptic Neuronal Apoptosis. Curr Neurovasc Res 2018, 15, 276-281. [CrossRef]
- Yue, J.; Wei, Y.J.; Yang, X.L.; Liu, S.Y.; Yang, H.; Zhang, C.Q. NLRP3 inflammasome and endoplasmic reticulum stress in the epileptogenic zone in temporal lobe epilepsy: molecular insights into their interdependence. Neuropathol Appl Neurobiol 2020, 46, 770-785. [CrossRef]
- Wang, L.; Ding, J.; Zhu, C.; Guo, B.; Yang, W.; He, W.; Li, X.; Wang, Y.; Li, W.; Wang, F.; et al. Semaglutide attenuates seizure severity and ameliorates cognitive dysfunction by blocking the NLR family pyrin domain containing 3 inflammasome in pentylenetetrazolekindled mice. Int J Mol Med 2021, 48, 219. [CrossRef]
- Wang, Z.; Zhou, L.; An, D.; Xu, W.; Wu, C.; Sha, S.; Li, Y.; Zhu, Y.; Chen, A.; Du, Y.; et al. TRPV4-induced inflammatory response is involved in neuronal death in pilocarpine model of temporal lobe epilepsy in mice. Cell Death Dis 2019, 10, 386. [CrossRef]
- Magalhaes, D.M.; Pereira, N.; Rombo, D.M.; Beltrao-Cavacas, C.; Sebastiao, A.M.; Valente, C.A. Ex vivo model of epilepsy in organotypic slices-a new tool for drug screening. J Neuroinflammation 2018, 15, 203. [CrossRef]
- Jiang, Q.; Tang, G.; Zhong, X.M.; Ding, D.R.; Wang, H.; Li, J.N. Role of Stat3 in NLRP3/caspase-1-mediated hippocampal neuronal pyroptosis in epileptic mice. Synapse 2021, 75, e22221. [CrossRef]
- Rong, S.; Wan, D.; Fan, Y.; Liu, S.; Sun, K.; Huo, J.; Zhang, P.; Li, X.; Xie, X.; Wang, F.; et al. Amentoflavone Affects Epileptogenesis and Exerts Neuroprotective Effects by Inhibiting NLRP3 Inflammasome. Front Pharmacol 2019, 10, 856. [CrossRef]
- Zhu, X.; Dong, J.; Xia, Z.; Zhang, A.; Chao, J.; Yao, H. Repeated restraint stress increases seizure susceptibility by activation of hippocampal endoplasmic reticulum stress. Neurochem Int 2017, 110, 25-37. [CrossRef]
- Yue, J.; He, J.; Wei, Y.; Shen, K.; Wu, K.; Yang, X.; Liu, S.; Zhang, C.; Yang, H. Decreased expression of Rev-Erbalpha in the epileptic foci of temporal lobe epilepsy and activation of Rev-Erbalpha have anti-inflammatory and neuroprotective effects in the pilocarpine model. J Neuroinflammation 2020, 17, 43. [CrossRef]
- Wu C, Z.G., Chen L, Kim S, Yu J, Hu G, Chen J, Huang Y, Zheng G, Huang S. . The Role of NLRP3 and IL-1β in Refractory Epilepsy Brain Injury. Front Neurol. 2020, 10, 1418. [CrossRef]
- Cristina de Brito Toscano, E.; Leandro Marciano Vieira, E.; Boni Rocha Dias, B.; Vidigal Caliari, M.; Paula Goncalves, A.; Varela Giannetti, A.; Mauricio Siqueira, J.; Kimie Suemoto, C.; Elaine Paraizo Leite, R.; Nitrini, R.; et al. NLRP3 and NLRP1 inflammasomes are up-regulated in patients with mesial temporal lobe epilepsy and may contribute to overexpression of caspase-1 and IL-beta in sclerotic hippocampi. Brain Res 2021, 1752, 147230. [CrossRef]
- Wu, J., Chen, L., Wang, S., Li, Y., Liu,L., Chen, G., Wang, S. and Zhou, S. Negative Regulation of Autophagy in Activating Nucleotide-Binding Oligomerization Domain-Like Receptor Family Pyrin Domain-Containing 3 Inflammasomes in the Hippocampus of an Epilepsy Rat Model. Nanosci. Nanotechnol. Lett. 2019, 11, 947-959. [CrossRef]
- Liu, Z.; Xian, H.; Ye, X.; Chen, J.; Ma, Y.; Huang, W. Increased levels of NLRP3 in children with febrile seizures. Brain Dev 2020, 42, 336-341. [CrossRef]
- Zhang, H.; Yu, S.; Xia, L.; Peng, X.; Wang, S.; Yao, B. NLRP3 Inflammasome Activation Enhances ADK Expression to Accelerate Epilepsy in Mice. Neurochem Res 2022, 47, 713-722. [CrossRef]
- Pohlentz, M.S.; Muller, P.; Cases-Cunillera, S.; Opitz, T.; Surges, R.; Hamed, M.; Vatter, H.; Schoch, S.; Becker, A.J.; Pitsch, J. Characterisation of NLRP3 pathway-related neuroinflammation in temporal lobe epilepsy. PLoS One 2022, 17, e0271995. [CrossRef]
- Dixit, A.B.; Banerjee, J.; Srivastava, A.; Tripathi, M.; Sarkar, C.; Kakkar, A.; Jain, M.; Chandra, P.S. RNA-seq analysis of hippocampal tissues reveals novel candidate genes for drug refractory epilepsy in patients with MTLE-HS. Genomics 2016, 107, 178-188. [CrossRef]
- Gao, B.; Wu, Y.; Yang, Y.J.; Li, W.Z.; Dong, K.; Zhou, J.; Yin, Y.Y.; Huang, D.K.; Wu, W.N. Sinomenine exerts anticonvulsant profile and neuroprotective activity in pentylenetetrazole kindled rats: involvement of inhibition of NLRP1 inflammasome. J Neuroinflammation 2018, 15, 152. [CrossRef]
- Samadianzakaria, A.; Abdolmaleki, Z.; Faedmaleki, F. The effect of valproic acid and furosemide on the regulation of the inflammasome complex (NLRP1 and NLRP3 mRNA) in the brain of epileptic animal model. Brain Res Bull 2022, 191, 20-29. [CrossRef]
- Cosford, N.D.; Meinke, P.T.; Stauderman, K.A.; Hess, S.D. Recent advances in the modulation of voltage-gated ion channels for the treatment of epilepsy. Curr Drug Targets CNS Neurol Disord 2002, 1, 81-104. [CrossRef]
- Sills, G.J.; Rogawski, M.A. Mechanisms of action of currently used antiseizure drugs. Neuropharmacology 2020, 168, 107966. [CrossRef]
- Treiman, D.M. GABAergic mechanisms in epilepsy. Epilepsia 2001, 42 Suppl 3, 8-12. [CrossRef]
- Romoli, M.; Mazzocchetti, P.; D'Alonzo, R.; Siliquini, S.; Rinaldi, V.E.; Verrotti, A.; Calabresi, P.; Costa, C. Valproic Acid and Epilepsy: From Molecular Mechanisms to Clinical Evidences. Curr Neuropharmacol 2019, 17, 926-946. [CrossRef]
- Johannessen, C.U.; Johannessen, S.I. Valproate: past, present, and future. CNS Drug Rev 2003, 9, 199-216. [CrossRef]
- Glauser, T.A.; Cnaan, A.; Shinnar, S.; Hirtz, D.G.; Dlugos, D.; Masur, D.; Clark, P.O.; Adamson, P.C.; Childhood Absence Epilepsy Study, T. Ethosuximide, valproic acid, and lamotrigine in childhood absence epilepsy: initial monotherapy outcomes at 12 months. Epilepsia 2013, 54, 141-155. [CrossRef]
- Brigo, F.; Igwe, S.C.; Bragazzi, N.L.; Lattanzi, S. Clonazepam monotherapy for treating people with newly diagnosed epilepsy. Cochrane Database Syst Rev 2022, 2, CD013028. [CrossRef]
- Jenner, P.; Pratt, J.A.; Marsden, C.D. Mechanism of action of clonazepam in myoclonus in relation to effects on GABA and 5-HT. Adv Neurol 1986, 43, 629-643.
- Nardou, R.; Yamamoto, S.; Chazal, G.; Bhar, A.; Ferrand, N.; Dulac, O.; Ben-Ari, Y.; Khalilov, I. Neuronal chloride accumulation and excitatory GABA underlie aggravation of neonatal epileptiform activities by phenobarbital. Brain 2011, 134, 987-1002. [CrossRef]
- Zhang, L.L.; Zeng, L.N.; Li, Y.P. Side effects of phenobarbital in epilepsy: a systematic review. Epileptic Disord 2011, 13, 349-365. [CrossRef]
- Contreras-Garcia, I.J.; Cardenas-Rodriguez, N.; Romo-Mancillas, A.; Bandala, C.; Zamudio, S.R.; Gomez-Manzo, S.; Hernandez-Ochoa, B.; Mendoza-Torreblanca, J.G.; Pichardo-Macias, L.A. Levetiracetam Mechanisms of Action: From Molecules to Systems. Pharmaceuticals (Basel) 2022, 15, 475. [CrossRef]
- Itoh, K.; Ishihara, Y.; Komori, R.; Nochi, H.; Taniguchi, R.; Chiba, Y.; Ueno, M.; Takata-Tsuji, F.; Dohgu, S.; Kataoka, Y. Levetiracetam treatment influences blood-brain barrier failure associated with angiogenesis and inflammatory responses in the acute phase of epileptogenesis in post-status epilepticus mice. Brain Res 2016, 1652, 1-13. [CrossRef]
- Ebrahimi, H.A.; Ebrahimi, F. The effect of lamotrigine on epilepsy. Iran J Neurol 2012, 11, 162-163.
- Goldsmith, D.R.; Wagstaff, A.J.; Ibbotson, T.; Perry, C.M. Lamotrigine: a review of its use in bipolar disorder. Drugs 2003, 63, 2029-2050. [CrossRef]
- Marson, A.; Burnside, G.; Appleton, R.; Smith, D.; Leach, J.P.; Sills, G.; Tudur-Smith, C.; Plumpton, C.; Hughes, D.A.; Williamson, P.; et al. The SANAD II study of the effectiveness and cost-effectiveness of levetiracetam, zonisamide, or lamotrigine for newly diagnosed focal epilepsy: an open-label, non-inferiority, multicentre, phase 4, randomised controlled trial. Lancet 2021, 397, 1363-1374. [CrossRef]
- Guerrini, R.; Parmeggiani, L. Topiramate and its clinical applications in epilepsy. Expert Opin Pharmacother 2006, 7, 811-823. [CrossRef]
- Davies, J.A. Mechanisms of action of antiepileptic drugs. Seizure 1995, 4, 267-271. [CrossRef]
- Huang, C.R.; Chuang, H.Y.; Chen, N.C.; Chen, S.F.; Hsu, C.Y.; Chuang, Y.C. Zonisamide Therapy Reduces Metabolic Consequences and Diminishes Nonalcoholic Fatty Liver Disease in Patients with Epilepsy. J Clin Med 2021, 10. [CrossRef]
- Bang, L.M.; Goa, K.L. Spotlight on oxcarbazepine in epilepsy. CNS Drugs 2004, 18, 57-61. [CrossRef]
- Chang, X.C.; Yuan, H.; Wang, Y.; Xu, H.Q.; Hong, W.K.; Zheng, R.Y. Eslicarbazepine acetate add-on for drug-resistant partial epilepsy. Cochrane Database Syst Rev 2017, 10, CD008907. [CrossRef]
- Landmark, C.J.; Rektorli, L.; Burns, M.L.; Revdal, E.; Johannessen, S.I.; Brodtkorb, E. Pharmacokinetic data on brivaracetam, lacosamide and perampanel during pregnancy and lactation. Epileptic Disord 2021, 23, 426-431. [CrossRef]
- D'Andrea Meira, I.; Romao, T.T.; Pires do Prado, H.J.; Kruger, L.T.; Pires, M.E.P.; da Conceicao, P.O. Ketogenic Diet and Epilepsy: What We Know So Far. Front Neurosci 2019, 13, 5. [CrossRef]
- Gonzalez, H.F.J.; Yengo-Kahn, A.; Englot, D.J. Vagus Nerve Stimulation for the Treatment of Epilepsy. Neurosurg Clin N Am 2019, 30, 219-230. [CrossRef]
- Ohemeng, K.K.; Parham, K. Vagal Nerve Stimulation: Indications, Implantation, and Outcomes. Otolaryngol Clin North Am 2020, 53, 127-143. [CrossRef]
- Wallace, S.J. Newer antiepileptic drugs: advantages and disadvantages. Brain Dev 2001, 23, 277-283. [CrossRef]
- Farhat, G.; Yamout, B.; Mikati, M.A.; Demirjian, S.; Sawaya, R.; El-Hajj Fuleihan, G. Effect of antiepileptic drugs on bone density in ambulatory patients. Neurology 2002, 58, 1348-1353. [CrossRef]
- Paulson, O.B.; Gyory, A.; Hertz, M.M. Blood-brain barrier transfer and cerebral uptake of antiepileptic drugs. Clin Pharmacol Ther 1982, 32, 466-477. [CrossRef]
- Lin, T.Y.; Hung, C.Y.; Chiu, K.M.; Lee, M.Y.; Lu, C.W.; Wang, S.J. Neferine, an Alkaloid from Lotus Seed Embryos, Exerts Antiseizure and Neuroprotective Effects in a Kainic Acid-Induced Seizure Model in Rats. Int J Mol Sci 2022, 23. [CrossRef]
- Han, X.; Sun, S.; Sun, Y.; Song, Q.; Zhu, J.; Song, N.; Chen, M.; Sun, T.; Xia, M.; Ding, J.; et al. Small molecule-driven NLRP3 inflammation inhibition via interplay between ubiquitination and autophagy: implications for Parkinson disease. Autophagy 2019, 15, 1860-1881. [CrossRef]
- Sharma, B.; Satija, G.; Madan, A.; Garg, M.; Alam, M.M.; Shaquiquzzaman, M.; Khanna, S.; Tiwari, P.; Parvez, S.; Iqubal, A.; et al. Role of NLRP3 Inflammasome and Its Inhibitors as Emerging Therapeutic Drug Candidate for Alzheimer's Disease: a Review of Mechanism of Activation, Regulation, and Inhibition. Inflammation 2022, 46, 1-32. [CrossRef]
- Tezcan, G.; Garanina, E.E.; Alsaadi, M.; Gilazieva, Z.E.; Martinova, E.V.; Markelova, M.I.; Arkhipova, S.S.; Hamza, S.; McIntyre, A.; Rizvanov, A.A.; et al. Therapeutic Potential of Pharmacological Targeting NLRP3 Inflammasome Complex in Cancer. Front Immunol 2020, 11, 607881. [CrossRef]
- Wu, A.G.; Zhou, X.G.; Qiao, G.; Yu, L.; Tang, Y.; Yan, L.; Qiu, W.Q.; Pan, R.; Yu, C.L.; Law, B.Y.; et al. Targeting microglial autophagic degradation in NLRP3 inflammasome-mediated neurodegenerative diseases. Ageing Res Rev 2021, 65, 101202. [CrossRef]
- Chen, Q.L.; Yin, H.R.; He, Q.Y.; Wang, Y. Targeting the NLRP3 inflammasome as new therapeutic avenue for inflammatory bowel disease. Biomed Pharmacother 2021, 138, 111442. [CrossRef]
- Coll, R.C.; Schroder, K.; Pelegrin, P. NLRP3 and pyroptosis blockers for treating inflammatory diseases. Trends Pharmacol Sci 2022, 43, 653-668. [CrossRef]
- Schwaid, A.G.; Spencer, K.B. Strategies for Targeting the NLRP3 Inflammasome in the Clinical and Preclinical Space. J Med Chem 2021, 64, 101-122. [CrossRef]
- Coll, R.C.; Robertson, A.A.; Chae, J.J.; Higgins, S.C.; Munoz-Planillo, R.; Inserra, M.C.; Vetter, I.; Dungan, L.S.; Monks, B.G.; Stutz, A.; et al. A small-molecule inhibitor of the NLRP3 inflammasome for the treatment of inflammatory diseases. Nat Med 2015, 21, 248-255. [CrossRef]
- Coll, R.C.; Hill, J.R.; Day, C.J.; Zamoshnikova, A.; Boucher, D.; Massey, N.L.; Chitty, J.L.; Fraser, J.A.; Jennings, M.P.; Robertson, A.A.B.; et al. MCC950 directly targets the NLRP3 ATP-hydrolysis motif for inflammasome inhibition. Nat Chem Biol 2019, 15, 556-559. [CrossRef]
- Tapia-Abellan, A.; Angosto-Bazarra, D.; Martinez-Banaclocha, H.; de Torre-Minguela, C.; Ceron-Carrasco, J.P.; Perez-Sanchez, H.; Arostegui, J.I.; Pelegrin, P. MCC950 closes the active conformation of NLRP3 to an inactive state. Nat Chem Biol 2019, 15, 560-564. [CrossRef]
- Umiker, B.; Lee, H.H.; Cope, J.; Ajami, N.J.; Laine, J.P.; Fregeau, C.; Ferguson, H.; Alves, S.E.; Sciammetta, N.; Kleinschek, M.; et al. The NLRP3 inflammasome mediates DSS-induced intestinal inflammation in Nod2 knockout mice. Innate Immun 2019, 25, 132-143. [CrossRef]
- Saber, S.; El-Kader, E.M.A. Novel complementary coloprotective effects of metformin and MCC950 by modulating HSP90/NLRP3 interaction and inducing autophagy in rats. Inflammopharmacology 2021, 29, 237-251. [CrossRef]
- Dempsey, C.; Rubio Araiz, A.; Bryson, K.J.; Finucane, O.; Larkin, C.; Mills, E.L.; Robertson, A.A.B.; Cooper, M.A.; O'Neill, L.A.J.; Lynch, M.A. Inhibiting the NLRP3 inflammasome with MCC950 promotes non-phlogistic clearance of amyloid-beta and cognitive function in APP/PS1 mice. Brain Behav Immun 2017, 61, 306-316. [CrossRef]
- Li, J.; Zhuang, L.; Luo, X.; Liang, J.; Sun, E.; He, Y. Protection of MCC950 against Alzheimer's disease via inhibiting neuronal pyroptosis in SAMP8 mice. Exp Brain Res 2020, 238, 2603-2614. [CrossRef]
- Guo, C.; Fu, R.; Wang, S.; Huang, Y.; Li, X.; Zhou, M.; Zhao, J.; Yang, N. NLRP3 inflammasome activation contributes to the pathogenesis of rheumatoid arthritis. Clin Exp Immunol 2018, 194, 231-243. [CrossRef]
- Li, R.; Zhang, J.; Wang, Q.; Cheng, M.; Lin, B. TPM1 mediates inflammation downstream of TREM2 via the PKA/CREB signaling pathway. J Neuroinflammation 2022, 19, 257. [CrossRef]
- Gao, R.; Shi, H.; Chang, S.; Gao, Y.; Li, X.; Lv, C.; Yang, H.; Xiang, H.; Yang, J.; Xu, L.; et al. The selective NLRP3-inflammasome inhibitor MCC950 reduces myocardial fibrosis and improves cardiac remodeling in a mouse model of myocardial infarction. Int Immunopharmacol 2019, 74, 105575. [CrossRef]
- Xu, L.; Zhang, C.; He, D.; Jiang, N.; Bai, Y.; Xin, Y. Rapamycin and MCC950 modified gut microbiota in experimental autoimmune encephalomyelitis mouse by brain gut axis. Life Sci 2020, 253, 117747. [CrossRef]
- Zhang, X.; Xu, A.; Lv, J.; Zhang, Q.; Ran, Y.; Wei, C.; Wu, J. Development of small molecule inhibitors targeting NLRP3 inflammasome pathway for inflammatory diseases. Eur J Med Chem 2020, 185, 111822. [CrossRef]
- Ismael, S.; Nasoohi, S.; Ishrat, T. MCC950, the Selective Inhibitor of Nucleotide Oligomerization Domain-Like Receptor Protein-3 Inflammasome, Protects Mice against Traumatic Brain Injury. J Neurotrauma 2018, 35, 1294-1303. [CrossRef]
- Engel, T.; Sanz-Rodgriguez, A.; Jimenez-Mateos, E.M.; Concannon, C.G.; Jimenez-Pacheco, A.; Moran, C.; Mesuret, G.; Petit, E.; Delanty, N.; Farrell, M.A.; et al. CHOP regulates the p53-MDM2 axis and is required for neuronal survival after seizures. Brain 2013, 136, 577-592. [CrossRef]
- Abais, J.M.; Xia, M.; Zhang, Y.; Boini, K.M.; Li, P.L. Redox regulation of NLRP3 inflammasomes: ROS as trigger or effector? Antioxid Redox Signal 2015, 22, 1111-1129. [CrossRef]
- Paul, R.; Choudhury, A.; Chandra Boruah, D.; Devi, R.; Bhattacharya, P.; Choudhury, M.D.; Borah, A. Hypercholesterolemia causes psychomotor abnormalities in mice and alterations in cortico-striatal biogenic amine neurotransmitters: Relevance to Parkinson's disease. Neurochem Int 2017, 108, 15-26. [CrossRef]
- Zhu, X.; Dong, J.; Han, B.; Huang, R.; Zhang, A.; Xia, Z.; Chang, H.; Chao, J.; Yao, H. Neuronal Nitric Oxide Synthase Contributes to PTZ Kindling Epilepsy-Induced Hippocampal Endoplasmic Reticulum Stress and Oxidative Damage. Front Cell Neurosci 2017, 11, 377. [CrossRef]
- Jiang, H.; He, H.; Chen, Y.; Huang, W.; Cheng, J.; Ye, J.; Wang, A.; Tao, J.; Wang, C.; Liu, Q.; et al. Identification of a selective and direct NLRP3 inhibitor to treat inflammatory disorders. J Exp Med 2017, 214, 3219-3238. [CrossRef]
- Qiao, J.; Wu, X.; Luo, Q.; Wei, G.; Xu, M.; Wu, Y.; Liu, Y.; Li, X.; Zi, J.; Ju, W.; et al. NLRP3 regulates platelet integrin alphaIIbbeta3 outside-in signaling, hemostasis and arterial thrombosis. Haematologica 2018, 103, 1568-1576. [CrossRef]
- Shen, K.; Jiang, W.; Zhang, C.; Cai, L.; Wang, Q.; Yu, H.; Tang, Z.; Gu, Z.; Chen, B. Molecular Mechanism of a Specific NLRP3 Inhibitor to Alleviate Seizure Severity Induced by Pentylenetetrazole. Curr Mol Pharmacol 2021, 14, 579-586. [CrossRef]
- Zhu, Q.; Zheng, F.; You, W.; Kang, X.; Chen, C.; Pan, Z.; Zhou, J.; Hu, W. Expression of Histone H1 in Rats with Traumatic Brain Injury and the Effect of the NLRP3 Inflammasome Pathway. World Neurosurg 2023, 171, e286-e290. [CrossRef]
- Wang, Y.; Liu, Y.J.; Zhang, M.M.; Zhou, H.; Gao, Y.H.; Cheng, W.J.; Ye, Z.W.; Yuan, Z.Y.; Xu, G.H.; Li, C.F.; et al. CY-09 Alleviates the Depression-like Behaviors via Inhibiting NLRP3 Inflammasome-Mediated Neuroinflammation in Lipopolysaccharide-Induced Mice. ACS Chem Neurosci 2022, 13, 3291-3302. [CrossRef]
- Riddle, M.C. Editorial: sulfonylureas differ in effects on ischemic preconditioning--is it time to retire glyburide? J Clin Endocrinol Metab 2003, 88, 528-530. [CrossRef]
- Perregaux, D.G.; McNiff, P.; Laliberte, R.; Hawryluk, N.; Peurano, H.; Stam, E.; Eggler, J.; Griffiths, R.; Dombroski, M.A.; Gabel, C.A. Identification and characterization of a novel class of interleukin-1 post-translational processing inhibitors. J Pharmacol Exp Ther 2001, 299, 187-197.
- Coelho, F.R.; Cavriani, G.; Soares, A.L.; Teixeira, S.A.; Almeida, P.C.; Sudo-Hayashi, L.S.; Muscara, M.N.; Oliveira-Filho, R.M.; Vargaftig, B.B.; Tavares-de-Lima, W. Lymphatic-borne IL-1beta and the inducible isoform of nitric oxide synthase trigger the bronchial hyporesponsiveness after intestinal ischema/reperfusion in rats. Shock 2007, 28, 694-699. [CrossRef]
- Lamkanfi, M.; Mueller, J.L.; Vitari, A.C.; Misaghi, S.; Fedorova, A.; Deshayes, K.; Lee, W.P.; Hoffman, H.M.; Dixit, V.M. Glyburide inhibits the Cryopyrin/Nalp3 inflammasome. J Cell Biol 2009, 187, 61-70. [CrossRef]
- Masters, S.L.; Dunne, A.; Subramanian, S.L.; Hull, R.L.; Tannahill, G.M.; Sharp, F.A.; Becker, C.; Franchi, L.; Yoshihara, E.; Chen, Z.; et al. Activation of the NLRP3 inflammasome by islet amyloid polypeptide provides a mechanism for enhanced IL-1beta in type 2 diabetes. Nat Immunol 2010, 11, 897-904. [CrossRef]
- Lebreton, F.; Berishvili, E.; Parnaud, G.; Rouget, C.; Bosco, D.; Berney, T.; Lavallard, V. NLRP3 inflammasome is expressed and regulated in human islets. Cell Death Dis 2018, 9, 726. [CrossRef]
- Liu, H.; Gu, C.; Liu, M.; Liu, G.; Wang, Y. NEK7 mediated assembly and activation of NLRP3 inflammasome downstream of potassium efflux in ventilator-induced lung injury. Biochem Pharmacol 2020, 177, 113998. [CrossRef]
- Liu, K.; Zhu, J.; Chang, Y.; Lin, Z.; Shi, Z.; Li, X.; Chen, X.; Lin, C.; Pan, S.; Huang, K. Attenuation of cerebral edema facilitates recovery of glymphatic system function after status epilepticus. JCI Insight 2021, 6. [CrossRef]
- Shafaroodi, H.; Barati, S.; Ghasemi, M.; Almasirad, A.; Moezi, L. A role for ATP-sensitive potassium channels in the anticonvulsant effects of triamterene in mice. Epilepsy Res 2016, 121, 8-13. [CrossRef]
- Goldberg, E.L.; Asher, J.L.; Molony, R.D.; Shaw, A.C.; Zeiss, C.J.; Wang, C.; Morozova-Roche, L.A.; Herzog, R.I.; Iwasaki, A.; Dixit, V.D. beta-Hydroxybutyrate Deactivates Neutrophil NLRP3 Inflammasome to Relieve Gout Flares. Cell Rep 2017, 18, 2077-2087. [CrossRef]
- Jiang, Z.; Yin, X.; Wang, M.; Wang, Y.; Li, F.; Gao, Y.; Han, G.; Gao, Z.; Wang, Z. beta-Hydroxybutyrate alleviates pyroptosis in MPP(+)/MPTP-induced Parkinson's disease models via inhibiting STAT3/NLRP3/GSDMD pathway. Int Immunopharmacol 2022, 113, 109451. [CrossRef]
- Lutas, A.; Yellen, G. The ketogenic diet: metabolic influences on brain excitability and epilepsy. Trends Neurosci 2013, 36, 32-40. [CrossRef]
- Miyauchi, T.; Uchida, Y.; Kadono, K.; Hirao, H.; Kawasoe, J.; Watanabe, T.; Ueda, S.; Okajima, H.; Terajima, H.; Uemoto, S. Up-regulation of FOXO1 and reduced inflammation by beta-hydroxybutyric acid are essential diet restriction benefits against liver injury. Proc Natl Acad Sci U S A 2019, 116, 13533-13542. [CrossRef]
- Shippy, D.C.; Wilhelm, C.; Viharkumar, P.A.; Raife, T.J.; Ulland, T.K. beta-Hydroxybutyrate inhibits inflammasome activation to attenuate Alzheimer's disease pathology. J Neuroinflammation 2020, 17, 280. [CrossRef]
- Youm, Y.H.; Nguyen, K.Y.; Grant, R.W.; Goldberg, E.L.; Bodogai, M.; Kim, D.; D'Agostino, D.; Planavsky, N.; Lupfer, C.; Kanneganti, T.D.; et al. The ketone metabolite beta-hydroxybutyrate blocks NLRP3 inflammasome-mediated inflammatory disease. Nat Med 2015, 21, 263-269. [CrossRef]
- Kim, D.Y.; Simeone, K.A.; Simeone, T.A.; Pandya, J.D.; Wilke, J.C.; Ahn, Y.; Geddes, J.W.; Sullivan, P.G.; Rho, J.M. Ketone bodies mediate antiseizure effects through mitochondrial permeability transition. Ann Neurol 2015, 78, 77-87. [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 173, 1728-1741 e1713. [CrossRef]
- Oronsky, B.; Takahashi, L.; Gordon, R.; Cabrales, P.; Caroen, S.; Reid, T. RRx-001: a chimeric triple action NLRP3 inhibitor, Nrf2 inducer, and nitric oxide superagonist. Front Oncol 2023, 13, 1204143. [CrossRef]
- Chen, Y.; He, H.; Lin, B.; Chen, Y.; Deng, X.; Jiang, W.; Zhou, R. RRx-001 ameliorates inflammatory diseases by acting as a potent covalent NLRP3 inhibitor. Cell Mol Immunol 2021, 18, 1425-1436. [CrossRef]
- Reid T, O.B., Caroen S, Cabrales P. . The direct NLRP3 inhibitor and Phase 3 small molecule anticancer agent, RRx-001, protects aged triple transgenic Alzheimer’s disease model mice from CNS degeneration and cognitive decline. Alzheimer's Dement 2022, 18,e061516. [CrossRef]
- Sun, Y.; Ma, J.; Li, D.; Li, P.; Zhou, X.; Li, Y.; He, Z.; Qin, L.; Liang, L.; Luo, X. Interleukin-10 inhibits interleukin-1beta production and inflammasome activation of microglia in epileptic seizures. J Neuroinflammation 2019, 16, 66. [CrossRef]
- Li, Q.; Feng, H.; Wang, H.; Wang, Y.; Mou, W.; Xu, G.; Zhang, P.; Li, R.; Shi, W.; Wang, Z.; et al. Licochalcone B specifically inhibits the NLRP3 inflammasome by disrupting NEK7-NLRP3 interaction. EMBO Rep 2022, 23, e53499. [CrossRef]
- Honda, H.; Nagai, Y.; Matsunaga, T.; Okamoto, N.; Watanabe, Y.; Tsuneyama, K.; Hayashi, H.; Fujii, I.; Ikutani, M.; Hirai, Y.; et al. Isoliquiritigenin is a potent inhibitor of NLRP3 inflammasome activation and diet-induced adipose tissue inflammation. J Leukoc Biol 2014, 96, 1087-1100. [CrossRef]
- Zeng, J.; Chen, Y.; Ding, R.; Feng, L.; Fu, Z.; Yang, S.; Deng, X.; Xie, Z.; Zheng, S. Isoliquiritigenin alleviates early brain injury after experimental intracerebral hemorrhage via suppressing ROS- and/or NF-kappaB-mediated NLRP3 inflammasome activation by promoting Nrf2 antioxidant pathway. J Neuroinflammation 2017, 14, 119. [CrossRef]
- Wang, K.; Lv, Q.; Miao, Y.M.; Qiao, S.M.; Dai, Y.; Wei, Z.F. Cardamonin, a natural flavone, alleviates inflammatory bowel disease by the inhibition of NLRP3 inflammasome activation via an AhR/Nrf2/NQO1 pathway. Biochem Pharmacol 2018, 155, 494-509. [CrossRef]
- Mohseni-Moghaddam, P.; Sadr, S.S.; Roghani, M.; Arabzadeh, S.; Khamse, S.; Zamani, E.; Hosseini, M.; Moradi, F. Huperzine A ameliorates cognitive dysfunction and neuroinflammation in kainic acid-induced epileptic rats by antioxidant activity and NLRP3/caspase-1 pathway inhibition. Clin Exp Pharmacol Physiol 2019, 46, 360-372. [CrossRef]
- Liu, R.; Wu, S.; Guo, C.; Hu, Z.; Peng, J.; Guo, K.; Zhang, X.; Li, J. Ibuprofen Exerts Antiepileptic and Neuroprotective Effects in the Rat Model of Pentylenetetrazol-Induced Epilepsy via the COX-2/NLRP3/IL-18 Pathway. Neurochem Res 2020, 45, 2516-2526. [CrossRef]
- Aghaie, F.; Moradifar, F.; Hosseini, A. Rapamycin attenuates depression and anxiety-like behaviors through modulation of the NLRP3 pathway in pentylenetetrazole-kindled male Wistar rats. Fundam Clin Pharmacol 2021, 35, 1045-1054. [CrossRef]
- Xia, S.; Yang, P.; Li, F.; Yu, Q.; Kuang, W.; Zhu, Y.; Lu, J.; Wu, H.; Li, L.; Huang, H. Chaihu-Longgu-Muli Decoction exerts an antiepileptic effect in rats by improving pyroptosis in hippocampal neurons. J Ethnopharmacol 2021, 270, 113794. [CrossRef]
- He, H.; Jiang, H.; Chen, Y.; Ye, J.; Wang, A.; Wang, C.; Liu, Q.; Liang, G.; Deng, X.; Jiang, W.; et al. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat Commun 2018, 9, 2550. [CrossRef]
- Juliana, C.; Fernandes-Alnemri, T.; Wu, J.; Datta, P.; Solorzano, L.; Yu, J.W.; Meng, R.; Quong, A.A.; Latz, E.; Scott, C.P.; et al. Anti-inflammatory compounds parthenolide and Bay 11-7082 are direct inhibitors of the inflammasome. J Biol Chem 2010, 285, 9792-9802. [CrossRef]
- Wu, L.; Chen, M.; Li, M.; Wang, Y.; Li, Y.; Zheng, L.; Ke, Z.; Liu, K.; Qiao, Y.; Shi, X. Oridonin alleviates kanamycin-related hearing loss by inhibiting NLRP3/caspase-1/gasdermin D-induced inflammasome activation and hair cell pyroptosis. Mol Immunol 2022, 149, 66-76. [CrossRef]
- Agarwal, N.B.; Jain, S.; Nagpal, D.; Agarwal, N.K.; Mediratta, P.K.; Sharma, K.K. Liposomal formulation of curcumin attenuates seizures in different experimental models of epilepsy in mice. Fundam Clin Pharmacol 2013, 27, 169-172. [CrossRef]
- Bharal, N.; Sahaya, K.; Jain, S.; Mediratta, P.K.; Sharma, K.K. Curcumin has anticonvulsant activity on increasing current electroshock seizures in mice. Phytother Res 2008, 22, 1660-1664. [CrossRef]
- Choudhary, K.M.; Mishra, A.; Poroikov, V.V.; Goel, R.K. Ameliorative effect of Curcumin on seizure severity, depression like behavior, learning and memory deficit in post-pentylenetetrazole-kindled mice. Eur J Pharmacol 2013, 704, 33-40. [CrossRef]
- Gupta, Y.K.; Briyal, S.; Sharma, M. Protective effect of curcumin against kainic acid induced seizures and oxidative stress in rats. Indian J Physiol Pharmacol 2009, 53, 39-46.
- Hashemian, M.; Anissian, D.; Ghasemi-Kasman, M.; Akbari, A.; Khalili-Fomeshi, M.; Ghasemi, S.; Ahmadi, F.; Moghadamnia, A.A.; Ebrahimpour, A. Curcumin-loaded chitosan-alginate-STPP nanoparticles ameliorate memory deficits and reduce glial activation in pentylenetetrazol-induced kindling model of epilepsy. Prog Neuropsychopharmacol Biol Psychiatry 2017, 79, 462-471. [CrossRef]
- Yin, H.; Guo, Q.; Li, X.; Tang, T.; Li, C.; Wang, H.; Sun, Y.; Feng, Q.; Ma, C.; Gao, C.; et al. Curcumin Suppresses IL-1beta Secretion and Prevents Inflammation through Inhibition of the NLRP3 Inflammasome. J Immunol 2018, 200, 2835-2846. [CrossRef]
- Yang, Z.; Zhong, L.; Xian, R.; Yuan, B. MicroRNA-223 regulates inflammation and brain injury via feedback to NLRP3 inflammasome after intracerebral hemorrhage. Mol Immunol 2015, 65, 267-276. [CrossRef]
- Haneklaus, M.; Gerlic, M.; Kurowska-Stolarska, M.; Rainey, A.A.; Pich, D.; McInnes, I.B.; Hammerschmidt, W.; O'Neill, L.A.; Masters, S.L. Cutting edge: miR-223 and EBV miR-BART15 regulate the NLRP3 inflammasome and IL-1beta production. J Immunol 2012, 189, 3795-3799. [CrossRef]
- Qingfeng Meng, X.H., Hui Sun, Hao Wang, Zhenfeng Luan, Shoudong Wang. miR-223 regulates myocardial ischemia-reperfusion damage via targeting NLRP3 in vitro and in vivo. Int J Clin Exp Med 2018, 11, 2004-2013.
- Wang, R.; Li, Q.; He, Y.; Yang, Y.; Ma, Q.; Li, C. miR-29c-3p inhibits microglial NLRP3 inflammasome activation by targeting NFAT5 in Parkinson's disease. Genes Cells 2020, 25, 364-374. [CrossRef]
- Chen, D.; Dixon, B.J.; Doycheva, D.M.; Li, B.; Zhang, Y.; Hu, Q.; He, Y.; Guo, Z.; Nowrangi, D.; Flores, J.; et al. IRE1alpha inhibition decreased TXNIP/NLRP3 inflammasome activation through miR-17-5p after neonatal hypoxic-ischemic brain injury in rats. J Neuroinflammation 2018, 15, 32. [CrossRef]
- Gong, L.; Han, Y.; Chen, R.; Yang, P.; Zhang, C. LncRNA ZNF883-Mediated NLRP3 Inflammasome Activation and Epilepsy Development Involve USP47 Upregulation. Mol Neurobiol 2022, 59, 5207-5221. [CrossRef]
- Zhang, A.; Lu, Y.; Yuan, L.; Zhang, P.; Zou, D.; Wei, F.; Chen, X. miR-29a-5p Alleviates Traumatic Brain Injury- (TBI-) Induced Permeability Disruption via Regulating NLRP3 Pathway. Dis Markers 2021, 2021, 9556513. [CrossRef]
- Zamani, P.; Oskuee, R.K.; Atkin, S.L.; Navashenaq, J.G.; Sahebkar, A. MicroRNAs as important regulators of the NLRP3 inflammasome. Prog Biophys Mol Biol 2020, 150, 50-61. [CrossRef]
- Kennedy, C.R.; Goya Grocin, A.; Kovacic, T.; Singh, R.; Ward, J.A.; Shenoy, A.R.; Tate, E.W. A Probe for NLRP3 Inflammasome Inhibitor MCC950 Identifies Carbonic Anhydrase 2 as a Novel Target. ACS Chem Biol 2021, 16, 982-990. [CrossRef]
- Kadioglu, O.; Saeed, M.; Kuete, V.; Greten, H.J.; Efferth, T. Oridonin Targets Multiple Drug-Resistant Tumor Cells as Determined by in Silico and in Vitro Analyses. Front Pharmacol 2018, 9, 355. [CrossRef]
- Agarwal, S.; Pethani, J.P.; Shah, H.A.; Vyas, V.; Sasane, S.; Bhavsar, H.; Bandyopadhyay, D.; Giri, P.; Viswanathan, K.; Jain, M.R.; et al. Identification of a novel orally bioavailable NLRP3 inflammasome inhibitor. Bioorg Med Chem Lett 2020, 30, 127571. [CrossRef]
- Agarwal, S.; Sasane, S.; Shah, H.A.; Pethani, J.P.; Deshmukh, P.; Vyas, V.; Iyer, P.; Bhavsar, H.; Viswanathan, K.; Bandyopadhyay, D.; et al. Discovery of N-Cyano-sulfoximineurea Derivatives as Potent and Orally Bioavailable NLRP3 Inflammasome Inhibitors. ACS Med Chem Lett 2020, 11, 414-418. [CrossRef]
- Smolak, P.; Nguyen, M.; Diamond, C.; Wescott, H.; Doedens, J.R.; Schooley, K.; Snouwaert, J.N.; Bock, M.G.; Harrison, D.; Watt, A.P.; et al. Target Cell Activation of a Structurally Novel NOD-Like Receptor Pyrin Domain-Containing Protein 3 Inhibitor NT-0796 Enhances Potency. J Pharmacol Exp Ther 2024, 388, 798-812. [CrossRef]
- Yetisgin, A.A.; Cetinel, S.; Zuvin, M.; Kosar, A.; Kutlu, O. Therapeutic Nanoparticles and Their Targeted Delivery Applications. Molecules 2020, 25, 2193. [CrossRef]
- Mancuso, R.; Citterio, L.A.; Agostini, S.; Marventano, I.; La Rosa, F.; Re, F.; Seneci, P.; Saresella, M.; Clerici, M. Glibenclamide-Loaded Nanoparticles Reduce NLRP3 Inflammasome Activation and Modulate miR-223-3p/miR-7-1-5p Expression in THP-1 Cells. Pharmaceuticals (Basel) 2023, 16, 1590. [CrossRef]
- Nandi, D.; Debnath, M.; Forster, J., 3rd; Pandey, A.; Bharadwaj, H.; Patel, R.; Kulkarni, A. Nanoparticle-mediated co-delivery of inflammasome inhibitors provides protection against sepsis. Nanoscale 2024, 16, 4678-4690. [CrossRef]
- Tang, J.; Li, T.; Xiong, X.; Yang, Q.; Su, Z.; Zheng, M.; Chen, Q. Colchicine delivered by a novel nanoparticle platform alleviates atherosclerosis by targeted inhibition of NF-kappaB/NLRP3 pathways in inflammatory endothelial cells. J Nanobiotechnology 2023, 21, 460. [CrossRef]
- Chen, X.; Zhou, Y.; Yu, J. Exosome-like Nanoparticles from Ginger Rhizomes Inhibited NLRP3 Inflammasome Activation. Mol Pharm 2019, 16, 2690-2699. [CrossRef]
- Liu, B.; Li, X.; Yu, H.; Shi, X.; Zhou, Y.; Alvarez, S.; Naldrett, M.J.; Kachman, S.D.; Ro, S.H.; Sun, X.; et al. Therapeutic potential of garlic chive-derived vesicle-like nanoparticles in NLRP3 inflammasome-mediated inflammatory diseases. Theranostics 2021, 11, 9311-9330. [CrossRef]
- Muhammad, W.; Zhu, J.; Zhai, Z.; Xie, J.; Zhou, J.; Feng, X.; Feng, B.; Pan, Q.; Li, S.; Venkatesan, R.; et al. ROS-responsive polymer nanoparticles with enhanced loading of dexamethasone effectively modulate the lung injury microenvironment. Acta Biomater 2022, 148, 258-270. [CrossRef]
- Liu, X.; Lu, B.; Fu, J.; Zhu, X.; Song, E.; Song, Y. Amorphous silica nanoparticles induce inflammation via activation of NLRP3 inflammasome and HMGB1/TLR4/MYD88/NF-kb signaling pathway in HUVEC cells. J Hazard Mater 2021, 404, 124050. [CrossRef]
- Sharma, B.; McLeland, C.B.; Potter, T.M.; Stern, S.T.; Adiseshaiah, P.P. Assessing NLRP3 Inflammasome Activation by Nanoparticles. Methods Mol Biol 2018, 1682, 135-147. [CrossRef]
- Shirasuna, K.; Karasawa, T.; Takahashi, M. Exogenous nanoparticles and endogenous crystalline molecules as danger signals for the NLRP3 inflammasomes. J Cell Physiol 2019, 234, 5436-5450. [CrossRef]
- Wang, X.; Lin, J.; Wang, Z.; Li, Z.; Wang, M. Possible therapeutic targets for NLRP3 inflammasome-induced breast cancer. Discov Oncol 2023, 14, 93. [CrossRef]
- Debnath, M.; Forster, J., 3rd; Ramesh, A.; Kulkarni, A. Protein Corona Formation on Lipid Nanoparticles Negatively Affects the NLRP3 Inflammasome Activation. Bioconjug Chem 2023, 34, 1766-1779. [CrossRef]
- Rajendran, G.; Bhanu, D.; Aruchamy, B.; Ramani, P.; Pandurangan, N.; Bobba, K.N.; Oh, E.J.; Chung, H.Y.; Gangadaran, P.; Ahn, B.C. Chalcone: A Promising Bioactive Scaffold in Medicinal Chemistry. Pharmaceuticals (Basel) 2022, 15, 1250. [CrossRef]
- Thapa, P.; Upadhyay, S.P.; Suo, W.Z.; Singh, V.; Gurung, P.; Lee, E.S.; Sharma, R.; Sharma, M. Chalcone and its analogs: Therapeutic and diagnostic applications in Alzheimer's disease. Bioorg Chem 2021, 108, 104681. [CrossRef]
- Thapa, P.; Upadhyay, S.P.; Singh, V.; Boinpelly, V.C.; Zhou, J.; Johnson, D.K.; Gurung, P.; Lee, E.S.; Sharma, R.; Sharma, M. Chalcone: A potential scaffold for NLRP3 inflammasome inhibitors. Eur J Med Chem Rep 2023, 7. [CrossRef]
- Zhang, C.; Yue, H.; Sun, P.; Hua, L.; Liang, S.; Ou, Y.; Wu, D.; Wu, X.; Chen, H.; Hao, Y.; et al. Discovery of chalcone analogues as novel NLRP3 inflammasome inhibitors with potent anti-inflammation activities. Eur J Med Chem 2021, 219, 113417. [CrossRef]
- Jiang, W.; Li, M.; He, F.; Zhou, S.; Zhu, L. Targeting the NLRP3 inflammasome to attenuate spinal cord injury in mice. J Neuroinflammation 2017, 14, 207. [CrossRef]
- Zhao, X.J.; Zhu, H.Y.; Wang, X.L.; Lu, X.W.; Pan, C.L.; Xu, L.; Liu, X.; Xu, N.; Zhang, Z.Y. Oridonin Ameliorates Traumatic Brain Injury-Induced Neurological Damage by Improving Mitochondrial Function and Antioxidant Capacity and Suppressing Neuroinflammation through the Nrf2 Pathway. J Neurotrauma 2022, 39, 530-543. [CrossRef]
Figure 2.
Chemical structures of clinically approved antiseizure medications discussed in this review.
Figure 2.
Chemical structures of clinically approved antiseizure medications discussed in this review.
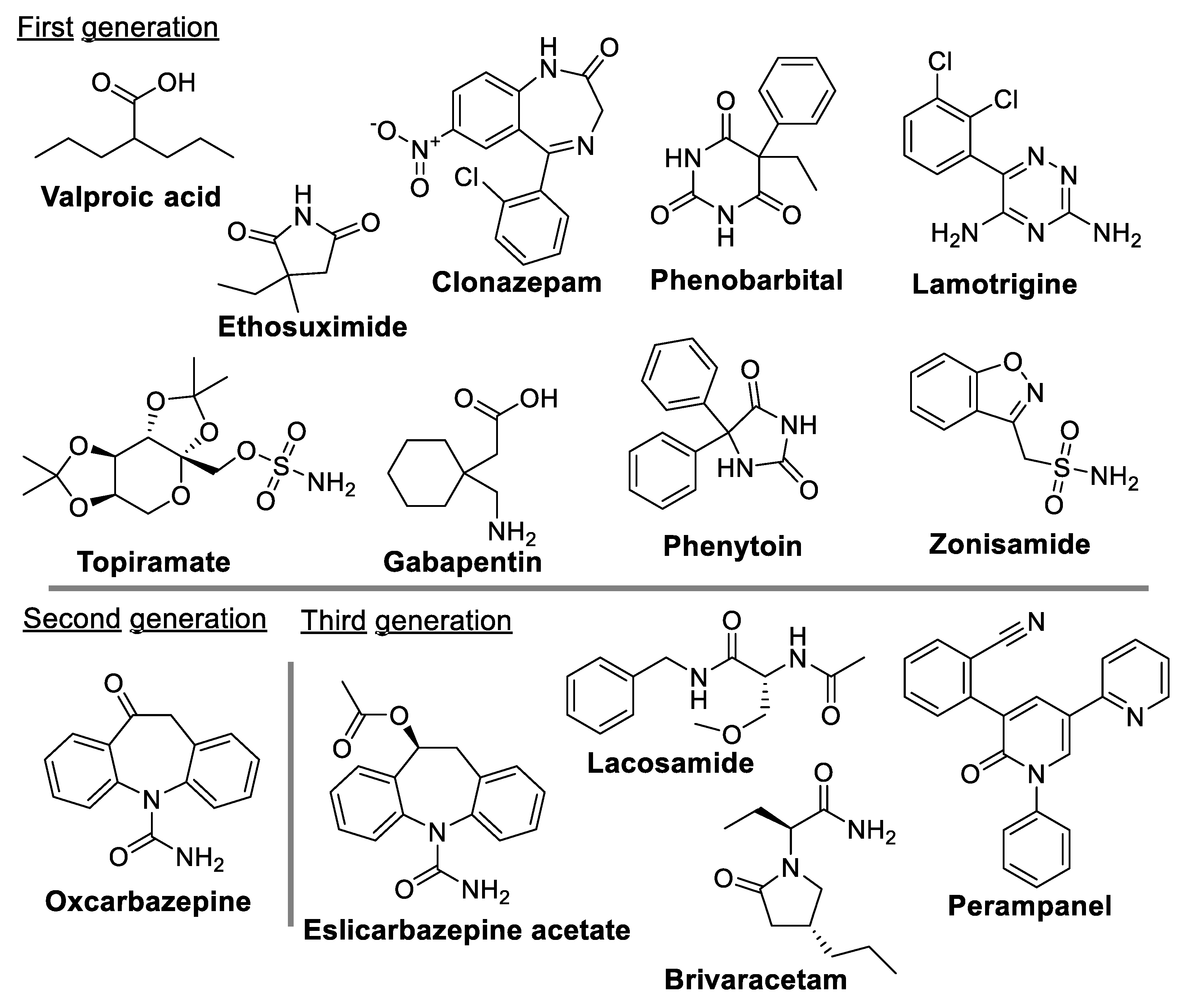
Figure 3.
Chemical structure of different NLRP3 inflammasome inhibitors discussed in this review.
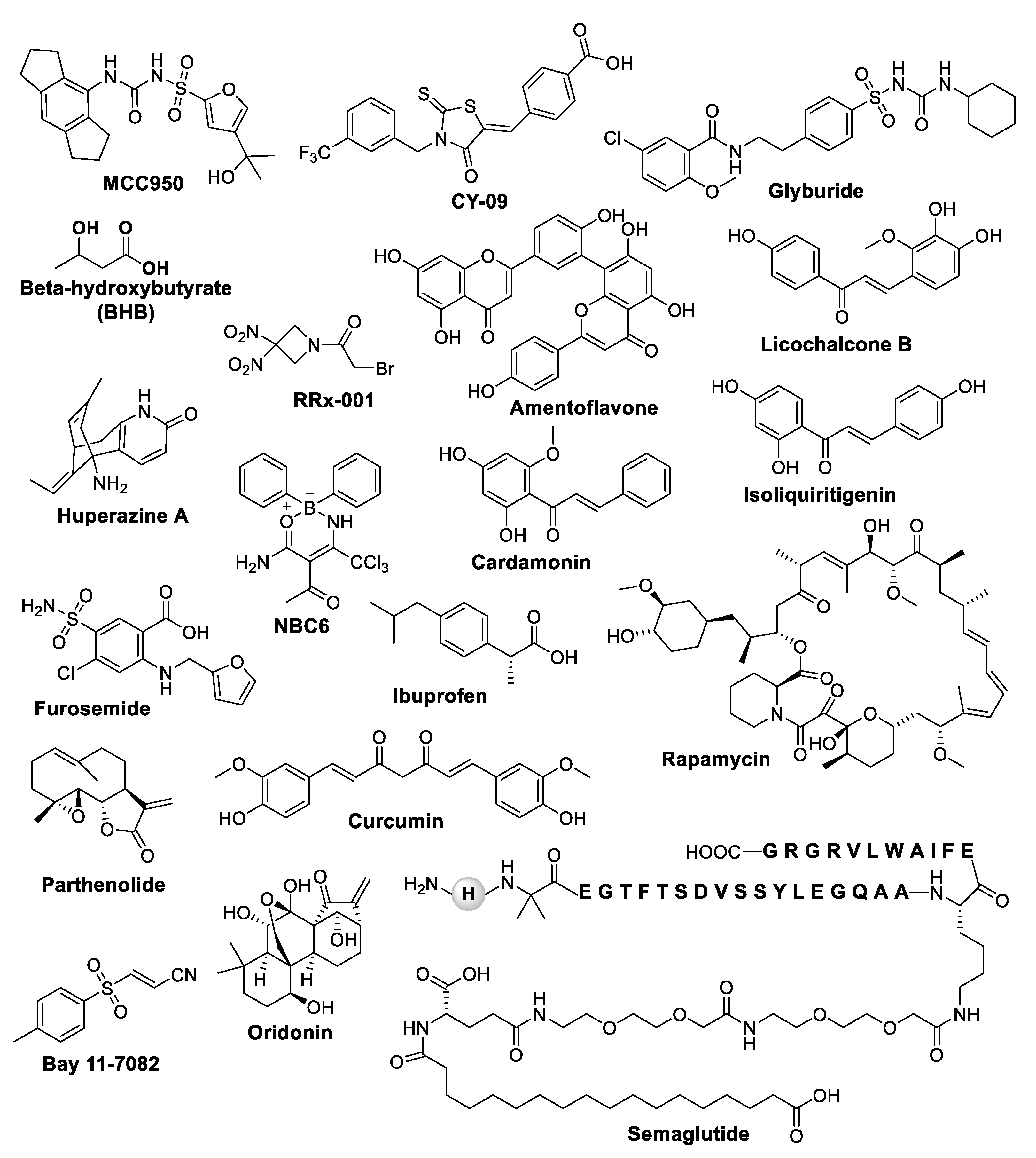
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



