Submitted:
10 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
Background and Objectives: Mycobacterium bovis Bacillus Calmette–Guérin (BCG) vaccine administration has been suggested to prevent glucose metabolism abnormalities and fatty liver in genetically obese ob/ob mice; however, it is not clear whether the beneficial effects of BCG are also observed in the progression of glucose intolerance induced by a high-fat diet (HFD). Therefore, the effects of BCG vaccination on changes in glucose tolerance and insulin response were investigated in HFD-fed C57BL/6 mice. Materials and Methods: We used the BCG Tokyo 172 strain to determine its effects on abnormalities in glucose metabolism. For vaccination, five-week-old male mice were injected intraperitoneally with BCG and maintained on an HFD three weeks later. The mice were regularly subjected to intraperitoneal glucose tolerance and insulin tolerance tests (IGTT and ITT). These tests were also performed in mice transplanted with bone marrow cells from BCG-vaccinated donor mice. Results: Significant effects of BCG vaccination on blood glucose levels in IGTT and ITT were observed from week 12 of the experiment. BCG vaccination significantly improved changes in fasting glucose and insulin levels, insulin resistance indexes, and glucagon-to-insulin ratios by HFD at the end of the experiment. Significant inhibitory effects in IGTT and ITT on glucose intolerance were also observed by transplantation with bone marrow cells derived from BCG-vaccinated donor mice. Conclusions: BCG vaccination significantly delayed glucose intolerance progression, suggesting a beneficial effect of BCG on the pathogenesis of type 2 diabetes. It has also been demonstrated that the effects of BCG vaccination may be at least partially due to an immune memory (trained immunity) for hematopoietic stem and progenitor cells of the bone marrow.
Keywords:
BCG
; glucose intolerance
; insulin resistance
; trained immunity
; nonalcoholic fatty liver disease
1. Introduction
Obesity, especially visceral obesity, contributes to the pathogenesis of metabolic syndrome, a cluster of metabolic abnormalities including hyperlipidemia, hypertension, and insuline resistance (IR) [1]. It is well known that IR is the primary indicator of type 2 diabetes mellitus (T2DM). T2DM pahtogensis is also considered to be linked to the innate and adaptive immune systems, which are recognized as important etiological components in the development of IR [2,3,4]. Per the International Diabetes Federation, T2DM is the most common type of diabetes (accounting for approximately 90% of all cases), and in 2021, more than one in ten adults now have diabetes mellitus globally, and the number of people with this disease will continually increase in the future [5]. The increasing prevalence of this condition makes it a public health problem of paramout importance. T2DM imposes a significant personal and public health burden in terms of the number of people affected, complications, and expenses incurred by the national health and social care system [6]. Therefore, the discovery and development of new treatments that regulate glucose and metabolic homeostasis are urgently needed.
An imbalance in energy homeostasis is a hallmark of T2DM [7]. Altered immune surveillance and impaired host defenses have been observed in patients suffering from obesity and T2DM, which may predispose patients to infection caused by germs such a Mycobacterium tuberculosis (Mtb) [8,9]. Animal and human studies have also indicated an increased susceptibility to Mtb infection in type 1 diabetes mellitus (T1DM), which is commonly known as juvenile-onset diabetes and characterized by an absolute deficiency in insulin production by autoimmune destruction of islet β-cells [10,11]. To protect against Mtb infection and its progression to tuberculosis, an attenuated strain of M. bovis was used to develop a vaccine, the Bacillus Calmette–Guérin (BCG) vaccine over 100 years ago. The nonspecific effects of BCG were first used for bladder cancer treatment over 40 years ago [12]. Thereafter, the off-target effects of BCG have been shown to protect against infectious and noninfectious diseases, including T1DM [13,14,15,16,17,18]. It has been reported that repeated BCG vaccinations in long-term diabetics can restore blood sugars to near normal by resetting the immune system and by increasing glucose utilization through a metabolic shift to aerobic glycolysis, a high-glucose-utilization state. [18]. However, to the best of our knowledge, only a few studies have examined the effects of BCG vaccination on T2DM. It has also been reported that in leptin receptor-deficient db/db mice, multiple BCG injections significantly decreased blood glucose levels and increased glucose uptake in bone marrow cells [19]. Our previous study demonstrated that a single intravenous administration of BCG significantly decreased serum insulin levels and the insulin resistance index, homeostatic model assessment for insulin resistance (HOMA-IR) in leptin-deficient ob/ob mice [20]. However, to the best of our knowledge, no information is available on the effect of BCG vaccination on diet-induced glucose intolerance. In the current study, we investigated the effects of prior BCG vaccination on the progression of glucose intolerance in high-fat and chow diet-fed mice.
2. Materials and Methods
2.1. Animals, Diet, and Microorganisms
All animal experiments were approved by the Animal Care and Use Committee of the University of the Ryukyus (approval numbers: A2022003 and A2022007) and conducted per their guidelines. Male C57BL/6JmsSlc (CD45.2; referred to as B6-Ly5.2) mice were purchased from Japan SLC Inc. (Shizuoka, Japan). C57BL/6-Ly5.1 (CD45.1; referred to as B6-Ly5.1) mice were maintained in our animal laboratory. The mice were randomly housed in environmentally enriched cages (5 animals per cage) under a controlled environment (at 24°C ± 1°C in a 12 h day/night cycle with lights on from 07:00 to 19:00). After one week of adaptation, the mice were randomly divided into experimental groups for each experiment. All animals had free access to food and water during the experiment.
A commercial chow diet (12 kcal% fat, CE-2 diet) and a high-fat diet (HFD, 30 kcal% fat, Quick fat diet) were purchased from CLEA Japan, Inc. (Tokyo, Japan). The BCG Tokyo 172 strain was purchased from the Japan BCG Laboratory (Tokyo, Japan) and suspended at 5×108 colony-forming units (CFU)/mL in phosphate-buffered saline (PBS) before use.
2.2. BCG Vaccination in HFD-Fed Mice
For vaccination, male five-week-old-B6-Ly5.2 mice were intraperitoneally (i.p.) injected with BCG (5 × 107 CFU/100 µL) for the BCG group and with vehicle PBS for the chow and control groups. All mice were fed a chow diet for three weeks, after which they started to access a chow diet for the chow group and an HFD for the control and BCG groups during the experiment. After the thirty-week feeding period, the mice were sacrificed after 12 h of starvation by exsanguination from the heart under isoflurane anesthesia to minimize suffering.
2.3. Bone Marrow Transplantation from BCG Vaccinated Mice
Donor male B6-Ly5.1 mice were i.p. injected with vehicle PBS or BCG (5 × 107 CFU/100 µL) and then fed a chow diet for three weeks. Their bone marrow samples were obtained by flushing their femurs and tibias with Eagle’s minimum essential medium and resuspended in PBS for transplantation. Recipient 8-week-old male B6-Ly5.2 mice were treated with i.p. busulfan injection for five days (20 mg/kg body weight/day) and transplanted intravenously with 1 × 107 nucleated cells from the donor’s bone marrow 24 h after the last busulfan injection. In this experiment, mice transplanted with bone marrow cells derived from PBS-treated mice served as the PBS-BM group and those from BCG-vaccinated mice served as the BCG-BM group. After bone marrow transplantation, all mice were started to feed an HFD.
2.4. Intraperitoneal Glucose Tolerance Test and Insulin Tolerance Test
At 8, 12, and 24 weeks after the start of the experiment, intraperitoneal glucose tolerance test (IGTT) were performed to assess whether the mice exhibited alterations in peripheral glucose regulation. The mice fasted for 12 h and were injected i.p. with D-glucose (1 g/kg body weight). Their blood glucose levels were measured before and at 30, 60, 90, and 120-min post-glucose injection. For the insulin tolerance test (ITT), mice fasted for 4 h and were injectedwith human recombinant insulin (1 U/kg body weight; Wako Pure Chemical Industries, Ltd., Osaka, Japan) i.p., and blood glucose levels were measured before and at 30, 60, 90, and 120 min post-injection. For both tests, blood was obtained from the tail vein, and glucose levels were measured using a glucometer (Free Style Precision Neo, Abbott Laboratories, IL, US).
2.5. Measurement of Biochemical Parameters in Serum
Serum triglyceride (TG), total cholesterol and glucose levels, and the activities of hepatopathy indicators; the activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST), were measured using a commercial enzymatic kit (Wako Pure Chemical Industries, Ltd., Osaka, Japan). Serum insulin and glucagon levels were measured using enzyme-linked immunosorbent assay kits purchased from Morinaga Institute of Biological Science, Inc. (Kanagawa, Japan), and Wako Pure Chemical Industries, Ltd., respectively. The homeostatic indexes for the quantification of insulin resistance and beta cell function (HOMA-IR and HOMA-β) were calculated as previously described [21].
2.6. Measurement of Hepatic Lipid Levels
Hepatic lipids were extracted and purified using a previously reported method [22]. We determined hepatic TG levels using commercial enzymatic kits (Wako Pure Chemical Industries).
2.7. Histopathological Examination
The pancreas was excised and immediately fixed in 10% neutral formalin solution. Formalin-fixed samples were embedded in paraffin and cut into 4-µm-thick sections. Paraffinized tissue sections were stained with hematoxylin and eosin (H&E) per a standard protocol for microscopic evaluation. The sizes of islets were calculated from digital images using Image J software (NIH, MA, US)
2.8. Flow Cytometry
The monoclonal antibodies (mAb) used in this study included the APC/Cyanine7 -conjugated anti-mouse CD45.1 mAb (clone A20), PerCP-cyanine5.5-conjugated anti-mouse CD45.1 mAb (clone 104), and non-labeled anti-mouse CD16/CD32 mAb (clone 2.4G2) were bought from BioLegend Inc. (CA, US), Thermo Fisher Scientific Inc. (MA, US), and BD Biosciences (CA, US), respectively. Before staining with the labeled mAb, isolated splenocyte were preincubated with an anti-CD32/CD16 mAb (2.4G2, BD Biosciences) before staining to prevent nonspecific Fc receptor-mediated binding of mAbs. The stained cells were analyzed on a FACSCanto II flow cytometer and analyzed with the FACSDiva software program (BD Biosciences).
2.9. Statistical Analyses
All data are expressed as the mean ± SEM. To determine the significance of the differences among mean value in the three experimental groups, the differences among the mean values were inspected using Tukey–Kramer’s multiple comparison tests. The threshold for statistical significance was set at p < 0.05.
3. Results
3.1. Effect of BCG Vaccination on Glucose Intolerance in HFD-fed Mice.
To assess the effect of BCG vaccination on the progression of glucose dysmetabolism in HFD-fed mice, we performed IGTTs and ITTs to measure the ability of mice to retain circulatory glucose levels over time after administering glucose and insulin and calculated the area under the curve (AUC) from these results. Changes in the blood glucose level and AUC in the GTT at week 8 of the experimental period did not differ significantly among all experimental groups (Figure 1A). Although no significant differences were detected in fasting glucose levels, the blood glucose levels in the HFD-fed control group at all measurement time points after glucose administration and the AUC were significantly increased compared with those in the chow group at week 12. However, when comparing the mice in the HFD-fed groups, blood glucose levels at 90 and 120 min after glucose administration and the AUC in the BCG group were significantly lower than those in the control group. The IGTT performed at week 24 revealed that HFD feeding led to marked hyperglycemia; however, BCG vaccination significantly reduced these abnormal levels and suppressed the increase in the AUC. As shown in Figure 1B, blood glucose levels after 4 h of fasting in the chow and BCG groups were significantly lower than those in the control group, although there were no significant differences in the mean dietary intake (data not shown) among all experimental groups. Significant inhibitory effects of BCG vaccination on blood glucose levels after insulin administration were observed only at week 12 of the experimental period in this study.

Table 1.
Growth and serum parameters in high-fat diet-fed mice at the end of the experimental period.
Table 1.
Growth and serum parameters in high-fat diet-fed mice at the end of the experimental period.
| Parameters | Chow | Cntl | BCG |
|---|---|---|---|
| Growth | |||
| Final body weight (g) | 27.5 ± 0.6 a | 39.7 ± 1.8 b | 36.6 ± 1.5 b |
| Liver weight (g) | 0.95 ± 0.03 a | 1.70 ± 0.19 b | 1.30 ± 0.08 ab |
| Serum | |||
| Triglyceride (mg/dL) | 54.4 ± 4.9 a | 69.7 ± 3.3 b | 60.3 ± 6.1 b |
| Total cholesterol (mg/dL) | 160 ± 12 | 188 ± 8.6 | 161 ± 9.5 |
| AST (IU/L) | 29.3 ± 5.2 | 26.9 ± 5.6 | 21.8 ± 5.9 |
| ALT (IU/L) | 4.41 ± 1.24 | 4.02 ± 0.50 | 4.47 ± 0.44 |
| Hepatic | |||
| Triglyceride (mg/g liver) | 31.3 ± 0.6 a | 57.4 ± 6.6 b | 40.6 ± 5.6 b |
Chow; chow group, Cntl; control group, BCG; BCG group; AST; aspartate aminotransferase, ALT; alanine aminotransferase. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05). .
3.2. Effect of BCG Vaccination on Growth Parameters, and Blood Parameters, and Hepatic Lipid Contents
We assessed the effects of BCG on growth and serum parameters (Table 1). HFD consumption to significant increments in the final body weight, liver weight, serum TG level, and hepatic TG content. BCG vaccination tended to inhibit these increments in liver weight and hepatic TG content. The serum cholesterol level and hepatopathy indicators did not differ significantly among all experimental groups.
3.3. Effect of BCG Vaccination on Glucose Metabolism Parameters
Fasting serum glucose, insulin, and glucagon levels in the control group were significantly higher than those in the chow group (Figure 2A–C). A significant decrease in the glucagon-to-insulin ratio and significant increments in HOMA-IR and HOMA-β in the control group were observed compared with those in the chow group (Figure 2D–E). BCG vaccination significantly decreased the fasting serum glucose levels and insulin levels compared with the control group (Figure 2A, B). Although no significant effects of BCG vaccination on the glucagon levels were observed (Figure 2C), the HFD-induced decrease in the glucagon-to-insulin ratio was significantly alleviated by BCG vaccination (Figure 1D). HOMA-IR in the BCG group was significantly decreased compared with that in the control group; meanwhile, HOMA-β was did not differ significantly between the control and BCG groups (Figure 1E, F).
3.4. Effect of BCG Vaccination on Pancreatic Islet Size
As shown in Figure 3, the size of the pancreatic islet in the control group was larger than that in the chow group. These significant increments in islet size were inhibited by BCG-vaccinated mice.
3.5. Effect of Bone Marrow Transplantation from BCG-vaccinated Mice on Glucose Intolerance Progression
To assess whether immunomodulation induced by BCG vaccination affects the progression of glucose intolerance in HFD-fed mice, recipient B6-Ly5.2 mice were transplanted with bone marrow from donor B6-Ly5.1 mice, in which BCG vaccination performed three weeks earlier had caused immune changes. At week 8, the results of IGTT and ITT did not differ significantly between the PBS- and BCG-BM groups (Figure 4A, B). Blood glucose levels and AUCs were significantly decreased in the BCG-BM group compared with those in the PBS-BM group at week 12, although BCG vaccination did not affect the ITT results. Maximal blood glucose levels after 30 min of glucose administration were significantly decreased in the BCG-BM group compared with those in the PBS-BM group at week 24. Although blood glucose levels after insulin administration in the BCG-BM group tended to be lower than those in the PBS-BM group, the AUC of the BCG-BM group was significantly decreased compared with that of the PBS-BM group. More than 90% of the immune cells of recipient mice used in these studies were of donor origin (Figure 4C).
4. Discussion
BCG is a microorganism that was developed as a vaccine for tuberculosis 100 years ago, and its off-target effects have been found to range from cancer treatment to protection against infectious and noninfectious diseases [12,13,14,15,16,17,18]. Non-obese diabetic (NOD) mice are well-studied spontaneous models of autoimmune diabetes, and they mimic only some features of T1DM [17,23]. Three decades of research have indicated that BCG administration permanently cured diabetes when administered to NOD mice [23,24]. Many studies have shown that BCG can be used as a therapeutic promise for T1DM in humans [17,25,26,27]; meanwhile, others have suggested that BCG is not useful for T1DM [28,29,30]. T1DM onset is typically associated with the rapid loss of the pancreas function from the T cell autoimmune attack on the insulin-secreting cells of the islets of Langerhans. The impact of BCG on human blood sugars in T1DM appears to be driven by immune and immune-metabolic effects [31]. The transfer with immune cells from BCG-vaccinated NOD mice prevented the occurrence of overt diabetes in the recipients, while the transfer from untreated donors did not [23]. It has been demonstrated that BCG can reset the immune system on the cellular level by inducing suppressive regulatory T cells and killing the autoreactive cytotoxic T cells that attack insulin-secreting cells [32,33]. It is also known that aerobic glycolysis is suppressed in individuals with T1DM and oxidative phosphorylation, which is a metabolic pathway involving low glucose utilization, high ketone production, and high Krebs cycle utilization, is predominant instead. Recent studies have shown that BCG treatment switches systemic metabolism from overactivated oxidative phosphorylation to accelerated aerobic glycolysis, suggesting that they lead to the lowering of blood sugar levels [17,18]. This suggested that BCG induction of aerobic glycolysis has broader applicability to other forms of hyperglycemia including T2DM [17].
Our previous study suggested that intravenous BCG administration may be therapeutic in relation to the inhibition of fat accumulation and reduction of fasting insulin level and the insulin resistance index, HOMA-IR, in obese and T2DM model, leptin-deficient ob/ob mice [20]. Shpilsky et al. recently revealed a significant reduction in blood sugar levels and body weight gain by four BCG injections in rear footpads of db/db mice, a model of T2DM, although a single injection failed to have a clinical outcome [19]. Herein, we examined the beneficial effects of single and prior BCG vaccination on HFD-induced glucose intolerance in wild-type C57BL/6 mice. Although significant effects of HFD intake on the changes in blood sugar and AUC in both IGTT and ITT were observed from week 12 onward, blood glucose levels after 4 h of fasting were significantly lower in both chow and BCG groups than in the control group at week 8 (Figure 1). These data suggest that prior BCG vaccination not only lowers blood glucose levels, but also inhibits glucose dysmetabolism progression in HFD-fed mice.
At the end of the experimental period, fasting blood glucose, insulin levels, and HOMA-IR in BCG-vaccinated mice were significantly lower than in the control mice (Figure 2A, B, E). Fasting insulin levels and HOMA-IR are one approach to measuring IR [34]. T2DM is characterized by hyperglycemia, typically due to the interaction of IR and impaired beta cell function [35]. We observed that BCG vaccination does significantly not affect HOMA-β (Figure 2F). Pancreatic β-cell function is commonly estimated using HOMA-β, which in individuals with T2DM increases between years 4 and 3 before diagnosis and then decreases until diagnosis [36]. Glucose intolerance in HFD-fed mice with reduced insulin sensitivity is suggested to impair the β-cell functions in pancreatic islets, resulting in excessive β-cell proliferation and increased islet size [37,38]. It has also been reported that dietary intake of HFD to induce IR in rodents develops to increase the pancreatic islet size [39,40]. H&E staining of pancreatic islets were herein larger in the control group than in the chow group, and this increase was significantly diminished by BCG vaccination (Figure 3). These data suggest that prior BCG vaccination alleviates the development of β-cell dysfunction in HFD-fed mice and consequently improves insulin tolerance.
The glucagon-to-insulin ratio was ameliorated in the BCG group compared with that in the control group, although the serum glucagon levels were largely comparable between both experimental groups (Figure 2C, D). Recent studies have indicated that the lower the glucagon-to-insulin ratio, the greater the likelihood of suffering from nonalcoholic fatty liver disease (NAFLD), cardiovascular disease, and metabolic syndrome in patients with T2DM [41,42]. Intravenous BCG administration to ob/ob mice has been shown to improve the hepatic fat accumulation state and serum levels of high-molecular-weight adiponectin, which is considered a more sensitive marker of metabolic dysfunction [20,43]. In the present study, no significant differences were observed; however, inhibitory tendencies were observed in liver weight (p = 0.07) and hepatic TG content (p = 0.06) after BCG vaccination (Table 1). Therefore, these results suggest that pre-vaccination with BCG not only delays the progression of glucose dysmetabolism but also has beneficial effects on various diseases associated with lipid metabolism abnormalities, such as metabolic syndrome, in HFD-fed mice.
Although innate immune cells were usually considered to be able to respond de novo to stimuli but not to form immunological memories, which were previously considered only as a part of adaptive immunity, it has been found that even organisms without an adaptive immune response can protect themselves against reinfection with pathogens [44]. “Trained immunity,” a term first coined in 2011, refers to the immunological memory responses of innate immune cells in response to past pathogen infections [45]. This phenomenon was clarified in humans that BCG vaccination can lead to the epigenetic reprograming of monocytes, resulting in an enhanced proinflammatory responses to secondary invasion with unrelated pathogens [46]. It has also been reported that BCG vaccination induces persistent epigenetic, transcriptional, and functional changes in hematopoietic stem and progenitor cells in human bone marrow [47]. Several studies have suggested that BCG-induced trained immunity may be partly related to the heterologous beneficial off-target effects of BCG vaccination [48,49,50]. Therefore, we assessed whether BCG-induced trained immunity modulates the progression of glucose intolerance in HFD-fed mice. Bone marrow cells were harvested from donor B6-Ly5.1 mice vaccinated with BCG or administered with PBS three weeks previously and transplanted into busulfan-conditioned mice. The progression of HFD-induced glucose intolerance was inhibited in recipient B6-Ly5.2 with > 90% myeloid cells derived from BCG-vaccinated mice compared with those with myeloid cells not affected by BCG (Figure 4). These data suggest that the inhibitory effects of BCG vaccination on glucose intolerance in HFD-fed mice may be at least partially mediated by trained immunity. Faustman and colleagues reported that in vitro and in vivo BCG treatments can improve baseline glucose transport of monocytes, which is deficient in T1DM individual [51], and also reported that glucose uptake of peripheral monocytes isolated from T2DM subject and that of bone marrow cells of T2DM models, db/db mice were augmented by in vitro BCG treatment [19]. Therefore, it is suggested that such changes in glucose metabolism in bone marrow cells and their derived myeloid cells are relevant to the results of this study. The main limitation of our study is that it did not identify the molecular mechanisms and key functional cells for the inhibitory effects of BCG vaccination on glucose metabolism abnormalities, and these remain to be addressed in future studies.
5. Conclusions
Our study aimed to investigate, for the first time, the effects of prior BCG vaccination on glucose intolerance progression induced by HFD intake in normal mice. BCG vaccination significantly delayed the progression of glucose intolerance and tended to inhibit hepatic lipid accumulation and reductions in the glucagon-to-insulin ratio, suggesting a beneficial effect of BCG on the development of T2DM and NAFLD, which are frequently associated with features of the metabolic syndrome. Furthermore, it has been shown that the effects of BCG vaccination may be at least partially due to an immune memory, and trained immunity, for hematopoietic stem and progenitor cells of bone marrow. In this study, we used the BCG Tokyo 172 strain to determine its effects on abnormalities in glucose metabolism. However, it has been suggested that the importance of the BCG dose, BCG strain, and timing of BCG administration for the efficacy in human T1DM [17]. Further studies are required to understand the differences in dose administration and timing, and strains of the BCG vaccine, together with the identification of molecular mechanisms and key functional cells of the beneficial effects of BCG vaccination.
Author Contributions
H.A. and M.I. made a substantial contribution to assembling, analyzing, and interpreting the data, and writing the manuscript. M.I. contributed to planning the experiments and discussing the results. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by JSPS KAKENHI Grant Number 22K11878.
Institutional Review Board Statement
All animal experiments were approved by the Animal Care and Use Committee of the University of the Ryukyus (approval numbers: A2022003 and A2022007) and conducted in accordance with their guidelines.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in this article.
Acknowledgments
We would like to thank Japan BCG Laboratory and Goro Matsuzaki for preparing the BCG suspension.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Formiguera, X.; Canton, A. Obesity: epidemiology and clinical aspects. Best Pract Res Clin Gastroenterol 2004, 18, 1125-1146. [CrossRef]
- Patel, P.S.; Buras, E.D.; Balasubramanyam, A. The Role of the Immune System in Obesity and Insulin Resistance. Journal of Obesity 2013, 2013, 1-9. [CrossRef]
- Pedicino, D.; Francesca, A.; Alessandro, V.; Trotta, F.; Liuzzo, G. Type 2 Diabetes, Immunity and Cardiovascular Risk: A Complex Relationship. InTech: 2012.
- Zhou, T.; Hu, Z.; Yang, S.; Sun, L.; Yu, Z.; Wang, G. Role of Adaptive and Innate Immunity in Type 2 Diabetes Mellitus. Journal of Diabetes Research 2018, 2018, 1-9. [CrossRef]
- Sun, H.; Saeedi, P.; Karuranga, S.; Pinkepank, M.; Ogurtsova, K.; Duncan, B.B.; Stein, C.; Basit, A.; Chan, J.C.N.; Mbanya, J.C.; et al. IDF Diabetes Atlas: Global, regional and country-level diabetes prevalence estimates for 2021 and projections for 2045. Diabetes Research and Clinical Practice 2022, 183, 109119. [CrossRef]
- Sinclair, A.; Saeedi, P.; Kaundal, A.; Karuranga, S.; Malanda, B.; Williams, R. Diabetes and global ageing among 65-99-year-old adults: Findings from the International Diabetes Federation Diabetes Atlas, 9(th) edition. Diabetes Res Clin Pract 2020, 162, 108078. [CrossRef]
- Gao, A.W.; Cantó, C.; Houtkooper, R.H. Mitochondrial response to nutrient availability and its role in metabolic disease. EMBO Molecular Medicine 2014, 6, 580-589. [CrossRef]
- Lachmandas, E.; Van Den Heuvel, C.N.A.M.; Damen, M.S.M.A.; Cleophas, M.C.P.; Netea, M.G.; Van Crevel, R. Diabetes Mellitus and Increased Tuberculosis Susceptibility: The Role of Short-Chain Fatty Acids. Journal of Diabetes Research 2016, 2016, 1-15. [CrossRef]
- Radhakrishnan, R.K.; Thandi, R.S.; Tripathi, D.; Paidipally, P.; McAllister, M.K.; Mulik, S.; Samten, B.; Vankayalapati, R. BCG vaccination reduces the mortality of Mycobacterium tuberculosis–infected type 2 diabetes mellitus mice. JCI Insight 2020, 5. [CrossRef]
- Jiang, Y.; Zhang, W.; Wei, M.; Yin, D.; Tang, Y.; Jia, W.; Wang, C.; Guo, J.; Li, A.; Gong, Y. Associations between type 1 diabetes and pulmonary tuberculosis: a bidirectional mendelian randomization study. Diabetology & Metabolic Syndrome 2024, 16. [CrossRef]
- Sugawara, I.; Mizuno, S. Higher Susceptibility of Type 1 Diabetic Rats to Mycobacterium tuberculosis Infection. The Tohoku Journal of Experimental Medicine 2008, 216, 363-370. [CrossRef]
- Jamshidi, P.; Danaei, B.; Mohammadzadeh, B.; Arbabi, M.; Nayebzade, A.; Sechi, L.A.; Nasiri, M.J. BCG Vaccination and the Risk of Type 1 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Pathogens 2023, 12, 581. [CrossRef]
- Arts, R.J.W.; Moorlag, S.; Novakovic, B.; Li, Y.; Wang, S.Y.; Oosting, M.; Kumar, V.; Xavier, R.J.; Wijmenga, C.; Joosten, L.A.B.; et al. BCG Vaccination Protects against Experimental Viral Infection in Humans through the Induction of Cytokines Associated with Trained Immunity. Cell Host Microbe 2018, 23, 89-100 e105. [CrossRef]
- Blok, B.A.; Arts, R.J.W.; Van Crevel, R.; Benn, C.S.; Netea, M.G. Trained innate immunity as underlying mechanism for the long-term, nonspecific effects of vaccines. Journal of Leukocyte Biology 2015, 98, 347-356. [CrossRef]
- Ijaz, M.U.; Vaziri, F.; Wan, Y.-J.Y. Effects of Bacillus Calmette-Gu erin on immunometabolism, microbiome and liver diseases. Liver Research 2023, 116-123. [CrossRef]
- Angelidou, A.; Pittet, L.F.; Faustman, D.; Curtis, N.; Levy, O. BCG vaccine’s off-target effects on allergic, inflammatory, and autoimmune diseases: Worth another shot? Journal of Allergy and Clinical Immunology 2022, 149, 51-54. [CrossRef]
- Kühtreiber, W.M.; Tran, L.; Kim, T.; Dybala, M.; Nguyen, B.; Plager, S.; Huang, D.; Janes, S.; Defusco, A.; Baum, D.; et al. Long-term reduction in hyperglycemia in advanced type 1 diabetes: the value of induced aerobic glycolysis with BCG vaccinations. npj Vaccines 2018, 3. [CrossRef]
- Kühtreiber, W.M.; Faustman, D.L. BCG Therapy for Type 1 Diabetes: Restoration of Balanced Immunity and Metabolism. Trends in Endocrinology & Metabolism 2019, 30, 80-92. [CrossRef]
- Shpilsky, G.F.; Takahashi, H.; Aristarkhova, A.; Weil, M.; Ng, N.; Nelson, K.J.; Lee, A.; Zheng, H.; Kühtreiber, W.M.; Faustman, D.L. Bacillus Calmette-Guerin ‘s beneficial impact on glucose metabolism: Evidence for broad based applications. iScience 2021, 24, 103150. [CrossRef]
- Inafuku, M.; Matsuzaki, G.; Oku, H. Intravenous Mycobacterium Bovis Bacillus Calmette-Guérin Ameliorates Nonalcoholic Fatty Liver Disease in Obese, Diabetic ob/ob Mice. PLOS ONE 2015, 10, e0128676. [CrossRef]
- Akbarian, F.; Rahmani, M.; Tavalaee, M.; Abedpoor, N.; Taki, M.; Ghaedi, K.; Nasr-Esfahani, M.H. Effect of Different High-Fat and Advanced Glycation End-Products Diets in Obesity and Diabetes-Prone C57BL/6 Mice on Sperm Function. Int J Fertil Steril 2021, 15, 226-233. [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. The Journal of biological chemistry 1957, 226, 497-509.
- Harada, M.; Kishimoto, Y.; Makino, S. Prevention of overt diabetes and insulitis in NOD mice by a single BCG vaccination. Diabetes Res Clin Pract 1990, 8, 85-89. [CrossRef]
- Shehadeh, N.; Etzioni, A.; Cahana, A.; Teninboum, G.; Gorodetsky, B.; Barzilai, D.; Karnieli, E. Repeated BCG vaccination is more effective than a single dose in preventing diabetes in non-obese diabetic (NOD) mice. Isr J Med Sci 1997, 33, 711-715.
- Doupis, J.; Kolokathis, K.; Markopoulou, E.; Efthymiou, V.; Festas, G.; Papandreopoulou, V.; Kallinikou, C.; Antikidou, D.; Gemistou, G.; Angelopoulos, T. The Role of Pediatric BCG Vaccine in Type 1 Diabetes Onset. Diabetes Therapy 2021, 12, 2971-2976. [CrossRef]
- Faustman, D.; Faustman, D.; ScienceDirect. The value of BCG and TNF in autoimmunity, Second edition. ed.; Academic Press: London, United Kingdom ; San Diego, CA, United States, 2018.
- Lazebnik, T.; Bunimovich-Mendrazitsky, S.; Kiselyov, A. Clinically Relevant Mathematical Model for the BCG-based Treatment Of Type 1 Diabetes. 2021. [CrossRef]
- Chang, Y.-C.; Lin, C.-J.; Hsiao, Y.-H.; Chang, Y.-H.; Liu, S.-J.; Hsu, H.-Y. Therapeutic Effects of BCG Vaccination on Type 1 Diabetes Mellitus: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Journal of Diabetes Research 2020, 2020, 1-8. [CrossRef]
- Moghtaderi, M.; Zarei, P.; Shakerian, B.; Babaei, M.; Mostafavi, A.; Modaressi, M. The Non-Significant Benefit of BCG Vaccination for the Treatment of Iranian Patients with Type 1 Diabetes up to 48 Weeks: A Controversial Result. Medical Journal of The Islamic Republic of Iran 2021. [CrossRef]
- Allen, H.F.; Klingensmith, G.J.; Jensen, P.; Simoes, E.; Hayward, A.; Chase, H.P. Effect of Bacillus Calmette-Guerin vaccination on new-onset type 1 diabetes. A randomized clinical study. Diabetes Care 1999, 22, 1703-1707. [CrossRef]
- Faustman, D.L. Benefits of BCG-induced metabolic switch from oxidative phosphorylation to aerobic glycolysis in autoimmune and nervous system diseases. Journal of Internal Medicine 2020, 288, 641-650. [CrossRef]
- Faustman, D.L.; Wang, L.; Okubo, Y.; Burger, D.; Ban, L.; Man, G.; Zheng, H.; Schoenfeld, D.; Pompei, R.; Avruch, J.; et al. Proof-of-Concept, Randomized, Controlled Clinical Trial of Bacillus-Calmette-Guerin for Treatment of Long-Term Type 1 Diabetes. PLoS ONE 2012, 7, e41756. [CrossRef]
- Keefe, R.C.; Takahashi, H.; Tran, L.; Nelson, K.; Ng, N.; Kühtreiber, W.M.; Faustman, D.L. BCG therapy is associated with long-term, durable induction of Treg signature genes by epigenetic modulation. Scientific Reports 2021, 11. [CrossRef]
- Singh, B.; Saxena, A. Surrogate markers of insulin resistance: A review. World J Diabetes 2010, 1, 36-47. [CrossRef]
- Esser, N.; Utzschneider, K.M.; Kahn, S.E. Early beta cell dysfunction vs insulin hypersecretion as the primary event in the pathogenesis of dysglycaemia. Diabetologia 2020, 63, 2007-2021. [CrossRef]
- Tabak, A.G.; Jokela, M.; Akbaraly, T.N.; Brunner, E.J.; Kivimaki, M.; Witte, D.R. Trajectories of glycaemia, insulin sensitivity, and insulin secretion before diagnosis of type 2 diabetes: an analysis from the Whitehall II study. Lancet 2009, 373, 2215-2221. [CrossRef]
- Matveyenko, A.V.; Gurlo, T.; Daval, M.; Butler, A.E.; Butler, P.C. Successful Versus Failed Adaptation to High-Fat Diet–Induced Insulin Resistance. Diabetes 2009, 58, 906-916. [CrossRef]
- Butler, A.E.; Janson, J.; Bonner-Weir, S.; Ritzel, R.; Rizza, R.A.; Butler, P.C. Beta-cell deficit and increased beta-cell apoptosis in humans with type 2 diabetes. Diabetes 2003, 52, 102-110. [CrossRef]
- Yang, H.-W.; Son, M.; Choi, J.; Oh, S.; Jeon, Y.-J.; Byun, K.; Ryu, B.M. Ishige okamurae reduces blood glucose levels in high-fat diet mice and improves glucose metabolism in the skeletal muscle and pancreas. Fisheries and Aquatic Sciences 2020, 23. [CrossRef]
- Hull, R.L.; Kodama, K.; Utzschneider, K.M.; Carr, D.B.; Prigeon, R.L.; Kahn, S.E. Dietary-fat-induced obesity in mice results in beta cell hyperplasia but not increased insulin release: evidence for specificity of impaired beta cell adaptation. Diabetologia 2005, 48, 1350-1358. [CrossRef]
- Moh Moh, M.A.; Jung, C.H.; Lee, B.; Choi, D.; Kim, B.Y.; Kim, C.H.; Kang, S.K.; Mok, J.O. Association of glucagon-to-insulin ratio and nonalcoholic fatty liver disease in patients with type 2 diabetes mellitus. Diab Vasc Dis Res 2019, 16, 186-195. [CrossRef]
- Bang, J.; Lee, S.A.; Koh, G.; Yoo, S. Association of Glucagon to Insulin Ratio and Metabolic Syndrome in Patients with Type 2 Diabetes. Journal of Clinical Medicine 2023, 12, 5806. [CrossRef]
- Lee, E.E.; Sears, D.D.; Liu, J.; Jin, H.; Tu, X.M.; Eyler, L.T.; Jeste, D.V. A novel biomarker of cardiometabolic pathology in schizophrenia? J Psychiatr Res 2019, 117, 31-37. [CrossRef]
- Kurtz, J.; Franz, K. Innate defence: evidence for memory in invertebrate immunity. Nature 2003, 425, 37-38. [CrossRef]
- Netea, M.G.; Joosten, L.A.; Latz, E.; Mills, K.H.; Natoli, G.; Stunnenberg, H.G.; O’Neill, L.A.; Xavier, R.J. Trained immunity: A program of innate immune memory in health and disease. Science 2016, 352, aaf1098. [CrossRef]
- Kleinnijenhuis, J.; Quintin, J.; Preijers, F.; Joosten, L.A.B.; Ifrim, D.C.; Saeed, S.; Jacobs, C.; Van Loenhout, J.; De Jong, D.; Stunnenberg, H.G.; et al. Bacille Calmette-Guérin induces NOD2-dependent nonspecific protection from reinfection via epigenetic reprogramming of monocytes. Proceedings of the National Academy of Sciences 2012, 109, 17537-17542. [CrossRef]
- Cirovic, B.; de Bree, L.C.J.; Groh, L.; Blok, B.A.; Chan, J.; van der Velden, W.; Bremmers, M.E.J.; van Crevel, R.; Handler, K.; Picelli, S.; et al. BCG Vaccination in Humans Elicits Trained Immunity via the Hematopoietic Progenitor Compartment. Cell Host Microbe 2020, 28, 322-334 e325. [CrossRef]
- Wu, Y.; Zhang, X.; Zhou, L.; Lu, J.; Zhu, F.; Li, J. Research progress in the off-target effects of Bacille Calmette-Guerin vaccine. Chin Med J (Engl) 2023. [CrossRef]
- van Puffelen, J.H.; Keating, S.T.; Oosterwijk, E.; van der Heijden, A.G.; Netea, M.G.; Joosten, L.A.B.; Vermeulen, S.H. Trained immunity as a molecular mechanism for BCG immunotherapy in bladder cancer. Nat Rev Urol 2020, 17, 513-525. [CrossRef]
- Atallah, A.; Grossman, A.; Nauman, R.W.; Paré, J.F.; Khan, A.; Siemens, D.R.; Cotechini, T.; Graham, C.H. Systemic versus localized <scp>Bacillus Calmette Guérin</scp> immunotherapy of bladder cancer promotes an anti-tumoral microenvironment: Novel role of trained immunity. International Journal of Cancer 2024. [CrossRef]
- Kühtreiber, W.M.; Takahashi, H.; Keefe, R.C.; Song, Y.; Tran, L.; Luck, T.G.; Shpilsky, G.; Moore, L.; Sinton, S.M.; Graham, J.C.; et al. BCG Vaccinations Upregulate Myc, a Central Switch for Improved Glucose Metabolism in Diabetes. iScience 2020, 23, 101085. [CrossRef]
Figure 1.
Effect of BCG vaccination on the progression of glucose intolerance in high-fat diet-fed mice. (A) Effect of BCG vaccination on the intraperitoneal glucose tolerance test (IGTT). (B) Effect of BCG vaccination on the insulin tolerance test (ITT). Chow; chow group, Cntl; control group, BCG; BCG group. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05). .
Figure 1.
Effect of BCG vaccination on the progression of glucose intolerance in high-fat diet-fed mice. (A) Effect of BCG vaccination on the intraperitoneal glucose tolerance test (IGTT). (B) Effect of BCG vaccination on the insulin tolerance test (ITT). Chow; chow group, Cntl; control group, BCG; BCG group. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05). .
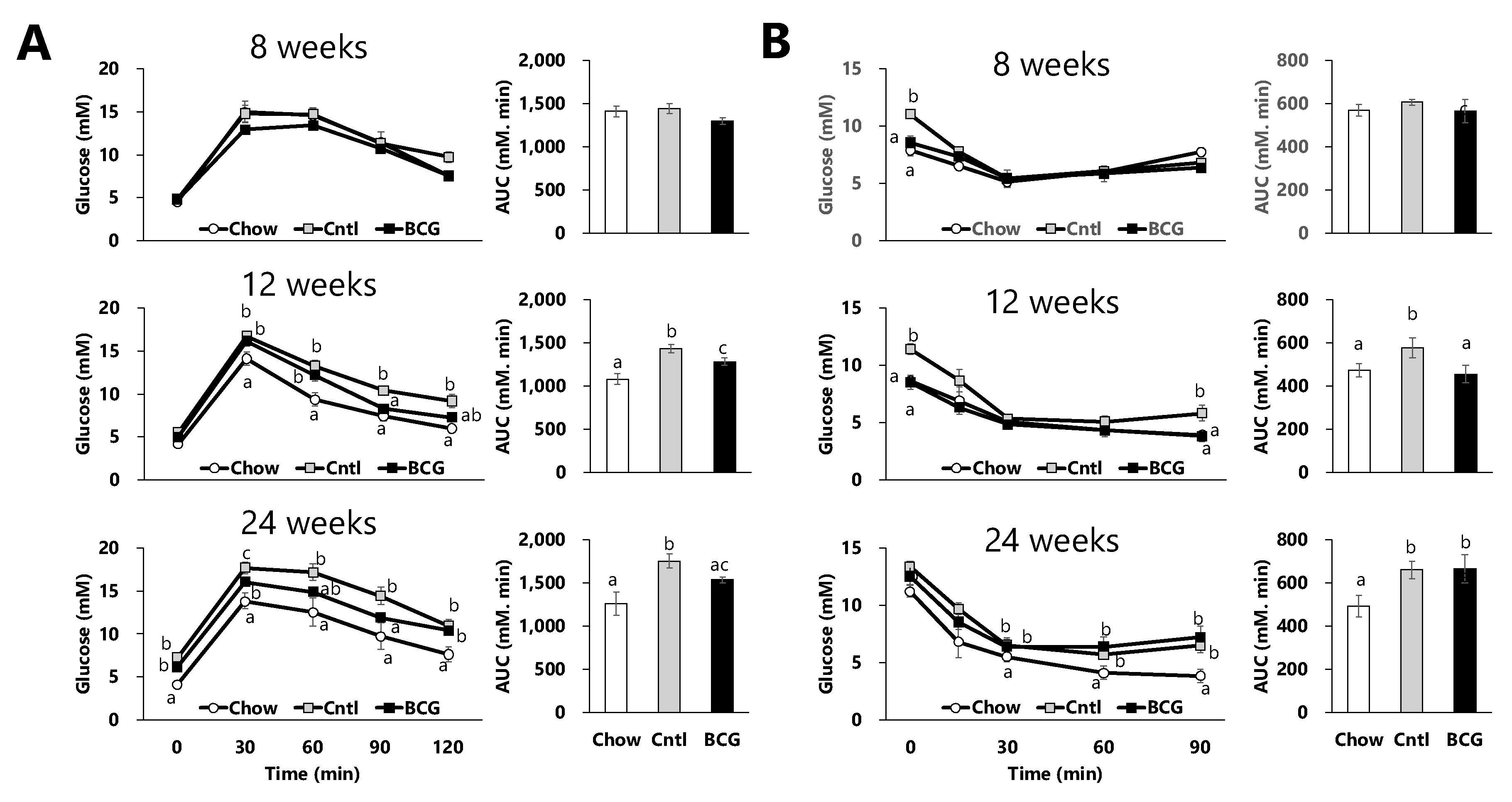
Figure 2.
Effect of BCG vaccination on serum parameters related to glucose metabolism in high fat diet-fed mice at the end of the experiment. (A) Serum glucose level. (B) Serum insulin level. (C) Serum glucagon level. (D) the glucagon-to-insulin ratio. (E) Homeostatic model assessment for insulin resistance (HOMA-IR). (F) Homeostatic model assessment for beta cell function (HOMA-β). Chow; chow group, Cntl; control group, BCG; BCG group. Data are presented as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05). .
Figure 2.
Effect of BCG vaccination on serum parameters related to glucose metabolism in high fat diet-fed mice at the end of the experiment. (A) Serum glucose level. (B) Serum insulin level. (C) Serum glucagon level. (D) the glucagon-to-insulin ratio. (E) Homeostatic model assessment for insulin resistance (HOMA-IR). (F) Homeostatic model assessment for beta cell function (HOMA-β). Chow; chow group, Cntl; control group, BCG; BCG group. Data are presented as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05). .
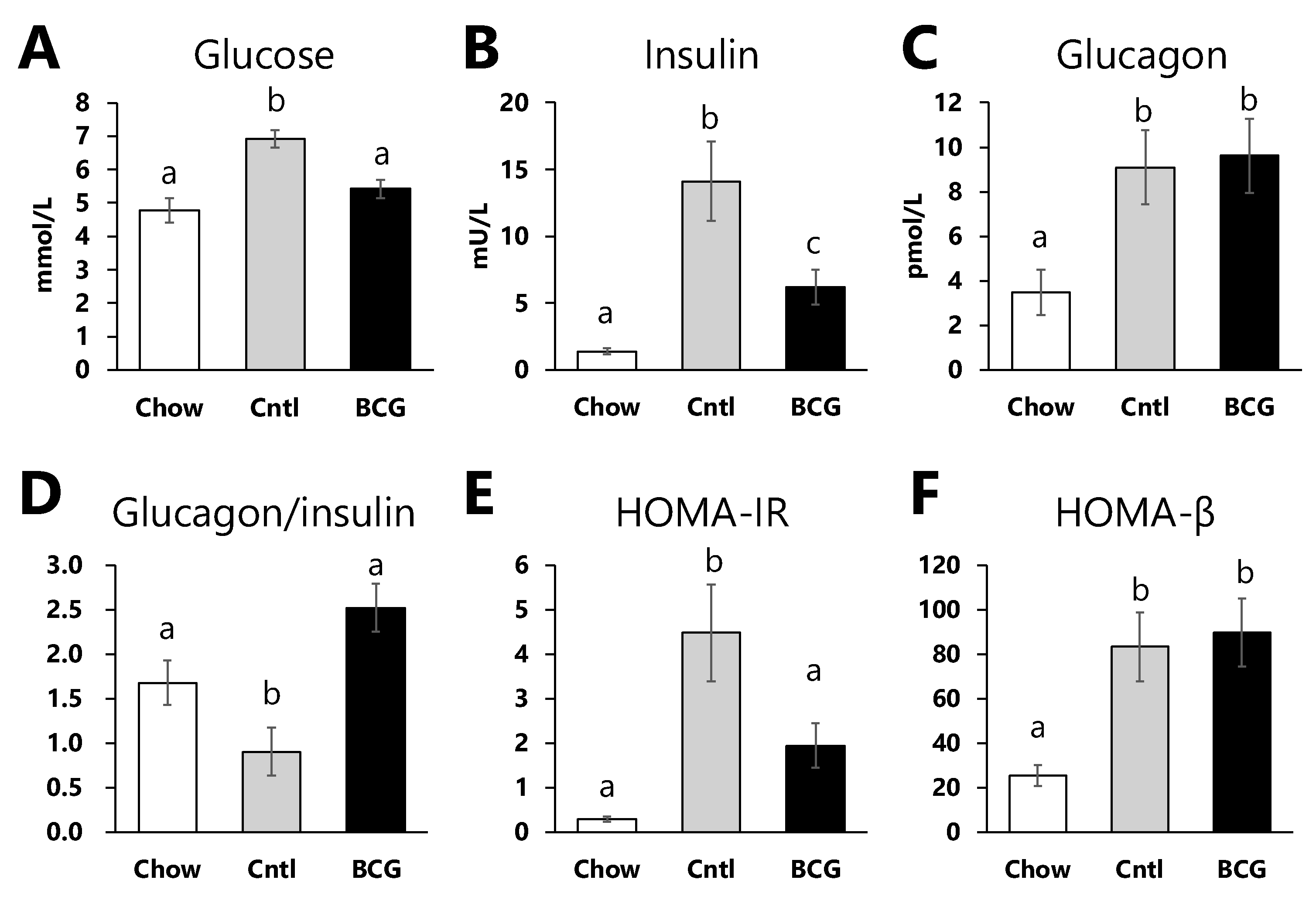
Figure 3.
Effect of BCG vaccination on pancreatic islet size in high-fat diet-fed mice at the end of the experiment. (A) Representative H&E histology of pancreatic samples from mice in each experimental group (scale bar = 100 μm). (B) The size of pancreatic islets. Chow; chow group, Cntl; control group, BCG; BCG group. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05).
Figure 3.
Effect of BCG vaccination on pancreatic islet size in high-fat diet-fed mice at the end of the experiment. (A) Representative H&E histology of pancreatic samples from mice in each experimental group (scale bar = 100 μm). (B) The size of pancreatic islets. Chow; chow group, Cntl; control group, BCG; BCG group. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05).
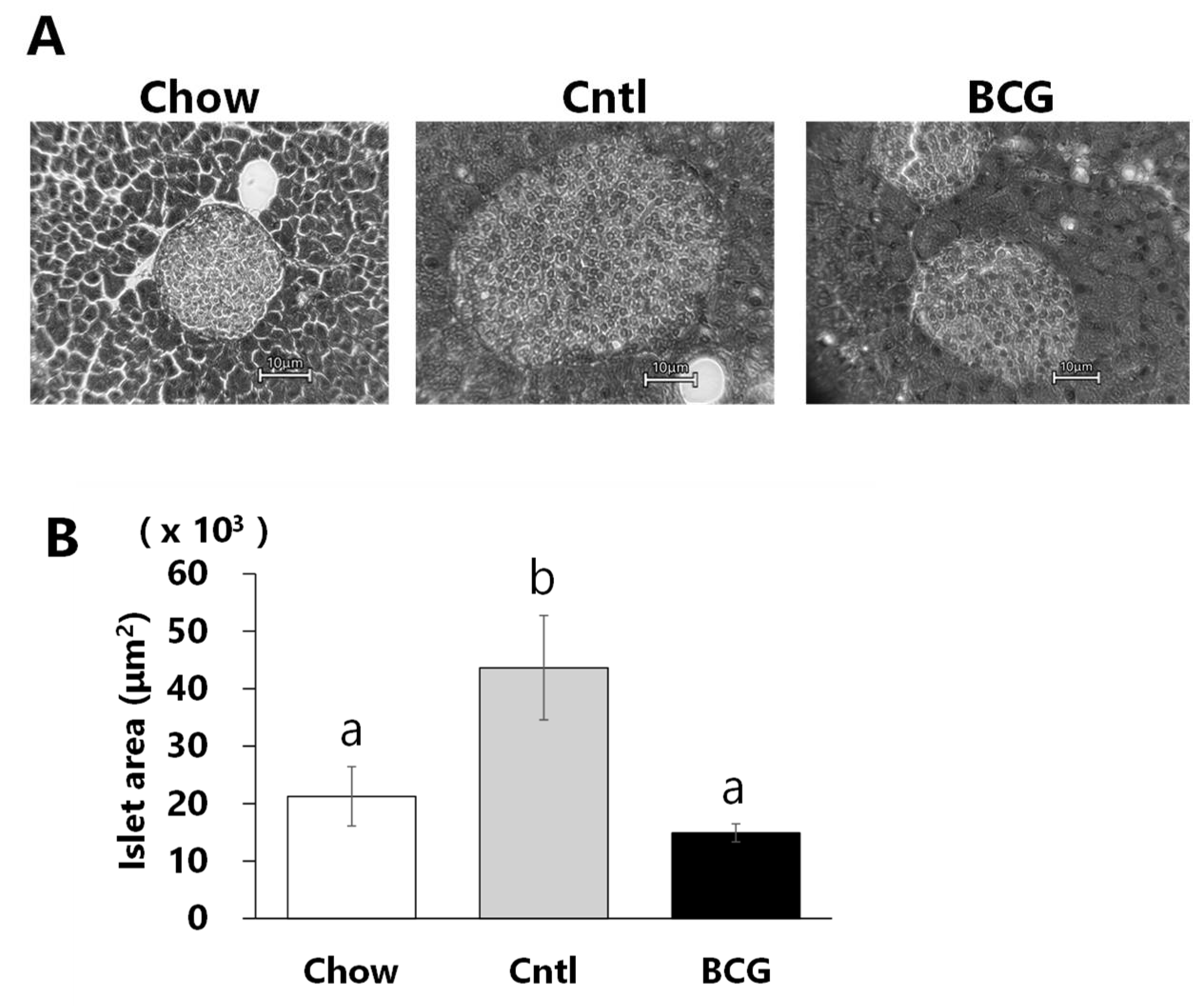
Figure 4.
Effect of bone marrow transplantation from BCG-vaccinated mice on the progression of glucose intolerance in high-fat diet-fed mice. (A) Effect of BCG vaccination on intraperitoneal glucose tolerance test (IGTT). (B) Effect of BCG vaccination on insulin tolerance test (ITT). (C) Representative flow cytometry plot of CD45.1 (Ly5.1) and CD45.2 (Ly5.2). PBS; PBS-BM group that transplanted with bone marrow from PBS-treated mice, BCG; BCG-BM group that transplanted with bone marrow from BCG-vaccinated mice. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05).
Figure 4.
Effect of bone marrow transplantation from BCG-vaccinated mice on the progression of glucose intolerance in high-fat diet-fed mice. (A) Effect of BCG vaccination on intraperitoneal glucose tolerance test (IGTT). (B) Effect of BCG vaccination on insulin tolerance test (ITT). (C) Representative flow cytometry plot of CD45.1 (Ly5.1) and CD45.2 (Ly5.2). PBS; PBS-BM group that transplanted with bone marrow from PBS-treated mice, BCG; BCG-BM group that transplanted with bone marrow from BCG-vaccinated mice. Data are shown as the mean ± SEM. Different letters indicate significant differences among the experimental groups using Tukey–Kramer’s multiple comparison test (p < 0.05).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



