
Submitted:
11 May 2024
Posted:
13 May 2024
You are already at the latest version
Preprints on COVID-19 and SARS-CoV-2
Abstract
Immunosuppressed individuals, such as people living with HIV (PLWH), remain vulnerable to severe COVID-19. We analyzed the persistence of specific SARS-CoV-2 humoral and cellular immune responses in a retrospective, cross-sectional study in PLWH on antiretroviral therapy. Among 104 participants, 70.2% had anti-S IgG antibodies and 55.8% had significant neutralizing activity against the Omicron variant in a surrogate virus neutralization test. Only 38.5% were vaccinated (8.76 ± 4.1 months prior), all displaying anti-S IgG, 75% with neutralizing antibodies and anti-S IgA. 29.8% had no SARS-CoV-2 serologic markers and displayed significantly lower CD4 counts and HIV viral load. Severe immunosuppression (present in 12.5% of participants) was linked to lower levels of detectable anti-S IgG (0.0003), anti-S IgA (p < 0.0001) and lack of neutralizing activity against Omicron variant (p < 0.0001). T-cell responses were present in 86.7% of tested participants, even in those lacking serological markers. In PLWH without severe immunosuppression, neutralizing antibodies and T-cell responses persisted for up to 9 months post-infection or vaccination. Advanced immunosuppression led to diminished humoral immunity, but retained cellular immune.
Keywords:
COVID-19
; SARS-CoV-2
; vaccination
; cellular
; humoral immune response
; immunosuppression
1. Introduction
Clinical trials and real-life studies have extensively evaluated the SARS-CoV-2 immune responses both after natural infection and after vaccination, demonstrating preserved protection against hospitalization and deaths, with diminishing efficacy against symptomatic infections caused by the continuously evolving, immune-evasive viral variants [1]. Nevertheless, there are several vulnerable populations who are still at risk for severe forms of COVID-19, including immunosuppressed persons. An analysis of the data gathered in the WHO Global Clinical Platform on COVID-19, revealed an independent correlation between HIV infection and severe outcomes of patients hospitalized with SARS-CoV-2 infection [2]. Consequently, the European AIDS Clinical Society strongly recommends vaccination of people living with HIV (PLWH), irrespective of CD4 count and HIV viral load. Variant adapted booster doses are also recommended with high priority for PLWH [3,4].
Passive immunization using anti-SARS-CoV-2 monoclonal antibodies, used as a possible pre-exposure prophylaxis in PLWH (especially in those unvaccinated, or with advanced immunosuppression), is no longer used due to the diminishing efficacy against the currently circulating SARS-CoV-2 variants [3].
In PLWH with controlled HIV infection, who are effectively treated, immunocompetent and with suppressed viral replication, SARS-CoV-2 infection has similar incidence rates and comparable evolution with the general population [5]. Nevertheless, PLWH with advanced HIV disease and severe immunodeficiency are prone to a more severe evolution of COVID-19, with higher risk of hospitalization and increased mortality [6,7]. After the emergence of the Omicron variant and its continuous antigenic drift, numerous breakthrough SARS-CoV-2 infections have been reported in all PLWH, including in vaccinated persons, with a higher frequency compared to the general population [8]. In addition, persistent SARS-CoV-2 infections, with continuous viral replication and high variability have been reported in immune-suppressed HIV infected persons, representing a potential source of new SARS-CoV-2 viral variants [1,9]. Repeated breakthrough infections could also potentially increase the HIV reservoirs in PLWH, even in those with undetectable HIV RNA in plasma on combination antiretroviral therapy (cART) [10].
Data on anti-SARS-CoV-2 immune responses efficiency in PLWH are controversial. While some studies indicate similar humoral and cellular immune responses to those developed in HIV-negative subjects [11,12,13,14,15], others suggest a decreased immune response efficiency in PLWH [16,17,18]. Severe immunosuppression decreases the ability to mount specific immune responses, both after infection and vaccination [5,19], and more severe breakthrough infections were diagnosed in PLWH even during the early COVID-19 waves [7,20]. As such, data regarding the SARS- CoV-2 immunity in HIV infected patients are still needed and are continuously gathered across diverse global regions with variable HIV demographics and healthcare standards [6].
The objective of the present study was to assess the seroprevalence of SARS-CoV-2 infection in non-hospitalized PLWH under antiretroviral therapy, and to analyze the persistence of specific SARS-CoV-2 humoral and cellular immune responses after vaccination and/or infection.
2. Materials and Methods
2.1. Patients and Samples
This retrospective, cross-sectional study included 104 HIV infected patients who were in active care in a regional HIV center, in Victor Babes Hospital for Tropical and Infectious Diseases, Bucharest, Romania, between April and August 2022. The study was conducted in accordance with the Declaration of Helsinki and approved by the Hospital’s Ethics Committee; all participants have signed an informed consent. 10 mL of venous blood on EDTAK3 anticoagulant were collected from all the patients. Serologic and virologic determinations were done in Stefan S. Nicolau Institute of Virology, Bucharest, Romania. Plasma was isolated by centrifugation and stored at - 20 °C for further analysis. Peripheral blood mononuclear cells (PBMCs) were isolated by density gradient centrifugation using Ficoll-Paque PREMIUM (GE Healthcare, Sweden), washed twice with phosphate saline buffer (PBS) and cryopreserved at – 80 °C, in AIM-V Medium (Thermo Fisher Scientific, USA) with 10 % DMSO, for further functional studies.
2.2. Detection of SARS-CoV-2 Humoral Immune Response
2.2.1. SARS-CoV-2 Anti-Spike (S) IgG and IgA, and Anti-Nucleocapsid (NCP) IgG Immunoassays
Anti-S IgG antibodies were tested using a quantitative immunoenzymatic assay (EUROIMMUN Medizinische Labordiagnostika AG, Lübeck, Germany), with results expressed in RU/mL, and transformed in BAU/mL using a conversion factor of 3.2 [21]. The results were considered negative if < 8 RU/mL (< 25.6 BAU/mL); borderline if between 8 - 11 RU/mL (25.6 – 35.2 BAU/mL); positive > 11 RU/mL (> 35.2 BAU/mL).
Anti-S IgA antibodies and anti-NCP IgG antibodies were tested with commercially semiquantitative enzyme-linked immunosorbent assays (EUROIMMUN Medizinische Labordiagnostika AG, Lübeck, Germany) according to the manufacturer protocols. Optical density (OD) was assessed at 450 nm and the results were expressed as reactivity (a ratio, calculated as the OD 450 of the patient’s sample over the OD 450 of a calibrator - an anti-S IgA positive sample or anti-NCP IgG positive sample provided by the manufacturer). The resulting ratio was considered negative if < 0.8; borderline if ≥ 0.8 to < 1.0, and positive if ≥ 1.1.
2.2.2. Surrogate SARS-CoV-2 Virus Neutralization Test
A SARS-CoV-2 surrogate virus neutralization test (GenScript cPass™ SARS-CoV-2 Neutralization Antibody Detection Kit, Genscript, US) was used to test the presence of neutralizing antibodies. This functional assay mimics the antibodies’ ability to block the interaction between the ACE2 receptor and the spike receptor-binding domain (RBD). The method was performed as previously validated [22]. Briefly, plasma samples were diluted 1:10, mixed with an equal volume of enzyme-conjugated RBD specific for the BA.4/BA.5 Omicron strain of SARS-CoV-2 (the predominant variant circulating worldwide in 2022), incubated at 37 °C for 30 min and transferred to a plate coated with recombinant human ACE2. After 15 min of incubation the plate was washed, and the enzyme substrate (100 μL tetramethylbenzidine) was added for 15 min at room temperature. Optical density (OD) was assessed at 450 nm and the samples’ neutralizing capacity was calculated using the following formula: % inhibition = [1 − (OD450 of the Sample / Mean OD450 of Negative Controls] × 100.
An inhibition value > 30% is considered positive for the SARS-CoV-2 neutralizing activity; values of 30 - 60% represents a low neutralizing activity, 60 - 90% - medium, and > 90% indicating a high SARS-CoV-2 neutralizing activity.
2.3. Detection of Cellular Immune Response
2.3.1. ELISpot Test
The IFN-γ and IL-2 secreting PBMCs were detected by an ELISpot assay (human IFN-gamma/IL-2 Dual-Color ELISpot Kit, R&D Systems, USA) according to the manufacturer protocol. Briefly, cryopreserved PBMCs were thawed and let to rest in AIM-V Medium (Thermo Fisher Scientific, USA) for two hours at 37 °C before plating. The cells were plated at 2×105 cells/well and were stimulated for 20 hours with 2 µg/mL of the recombinant BA.4/BA.5 S1 protein (R&D Systems, USA). Positive controls consisted of PBMCs stimulated with 2 µg/mL PepTivator® CMV pp65 (Miltenyi Biotec, USA) - a pool of 15-mer peptides with 11–amino acid overlap that covers the complete sequence of pp65 protein of cytomegalovirus (CMV), and negative control consisted of unstimulated PBMCs. Spots were counted using the AID ELISpot Reader System (Autoimmun Diagnostika GmbH, Germany). All assays were performed in duplicate. Results are reported as difference in spot forming units per 106 PBMCs between the spike peptide-stimulated PBMCs and negative controls with negative values set to zero (Δ SFU/106 PBMCs).
2.3.2. Flow Cytometry Analysis
The CD4/CD8 ratio was evaluated by flow-cytometry using tetraCHROME CD45-FITC/CD4-PE/CD8-ECD/CD3-PC5 antibody Cocktail (Beckman Coulter, USA) according to the manufacturer protocol. The samples were acquired on EPICS XL flow cytometer (Beckman Coulter, USA) and data were analyzed using Kaluza software (Beckman Coulter, USA).
2.4. Statistical Analysis
Data were analyzed using GraphPad Prism version 8 software. Multiple groups were compared using a two-way analysis of variance (ANOVA), followed by a Tukey multiple comparisons test. The non-parametric Spearman test was used for correlation analysis, with a significance threshold defined by p <0.05.
3. Results
3.1. General Characteristics of the Enrolled Subjects
The study included 104 PLWH (65.4% males, mean age 38.4 ± 0.98 years, mean duration of HIV infection 127.3 ± 10.55 months). HIV infection was acquired by sexual transmission (58.7% of cases, either hetero or homosexual), parenteral transmission (32.7% by injecting drug use, 4.8% parenterally infected during early childhood), and by vertical transmission (3.8% of cases). The mean CD4 count at the time of the study was 606.6 ± 32.62 cells/mm3, 27.9% of the patients had mild immunosuppression (CD4 200 - 500 cells/mm3) and 12.5% had severe immunosuppression (CD4 < 200 cells/mm3). The general characteristics of the study patients divided by the degree of immunosuppression are presented in Table 1.
All patients were treated with cART at the time of the study, with a mean length of antiretroviral therapy of 120.5 ± 9.9 months. All regimens included a backbone of 2 nucleoside analogues reverstranscriptase inhibitors (NRTI), combined with an integrase inhibitor (INSTI) in 74% of cases, a protease inhibitor (PI) in 18.3% of cases, and a non-nucleoside reverstranscriptase inhibitor (NNRTI) in 7.7% of cases. Only 60.6% of PLWH included in the study (63/104) had undetectable viral load, with significant differences according to the degree of immunosuppression, as shown in Table 1. Patients who were severely immunosuppressed have been diagnosed more recently, had lower CD4 nadir counts and higher HIV viral loads.
Out of 104 PLWH, only 40 (38.5%) were vaccinated against SARS-CoV-2 infection, either with an mRNA vaccine (Pfizer-BioNTech vaccine, BNT162b2 - 16 patients), or with a viral-vector based vaccine (Astra-Zeneca vaccine, ChAdOx1-S - 11 patients, or Janssen, Ad26.COV2.S - 13 patients). Just eight patients had received a booster dose; all boosters were monovalent mRNA vaccines, with the ancestral SARS-CoV-2 strain. The mean duration of time between the last vaccine dose and enrollment was 8.76 ± 4.1 months.
Only 11 subjects (10.6%) had been previously diagnosed with a SARS-CoV-2 infection, all with a mild clinical form. The mean time length between infection and enrollment was 9.36 ± 1.6 months.
3.2. SARS-CoV-2 Seroprevalence
Seventy-three patients (70.2%) had anti-Spike IgG antibodies (mean titer 477.3 ± 232 BAU/mL), 58 (55.8%) had also neutralizing antibody against the Omicron variant (an inhibition value > 30% in the surrogate SARS-CoV-2 virus neutralization test). Only 24 patients (23%) had anti-NCP antibodies (mean reactivity 2.3 ± 0.7) associated with anti-S IgG antibodies, indicating a prior SARS-CoV-2 infection.
Anti-S IgA antibodies were present in 62 patients (59.6%, with a mean reactivity 4.2 ± 1.1), always associated with anti-S IgG antibodies.
All vaccinated patients had anti-S IgG antibodies, 12 of them had also anti-NCP antibodies, signaling a prior SARS-CoV-2 infection and thus, hybrid immunity. Out of 40 vaccinated patients, 30 (75%) had detectable neutralizing antibodies against the Omicron variant (neutralizing activity in the surrogate SARS-CoV-2 virus neutralization test higher than 30%) and anti-S IgA antibodies.
The serologic profile of the previously SARS-CoV-2 infected patients was more heterogenous. Interestingly, 4 out of the 11 patients who declared past COVID-19, had no SARS-CoV-2 antibodies (negative for anti-S IgG and IgA, and for anti-NCP IgG), and 2 patients had only anti-S IgG antibodies (mean titer 636.2 ± 124 BAU/mL), with no detectable anti-NCP antibodies. Nevertheless, other 33 patients without a prior history of SARS-CoV-2 infection or vaccination, displayed anti-S IgG antibodies (with or without anti-NCP antibodies), indicating asymptomatic, undiagnosed infections.
The patients’ distribution according to their SARS-CoV-2 serologic status and vaccination/infection history is shown in Figure 1.
As seen in Figure 1, out of all enrolled patients, 31 have no SARS-CoV-2 serologic markers; they have significantly decreased CD4 counts (mean 443.8 ± 64.49 vs. 670.4 ± 35.3; p = 0.0015) and significantly lower percentages of undetectable HIV viral load (19.4% vs. 78.1%; p = 0.021) compared to those with detectable SARS-CoV-2 antibodies.
3.3. Antibody Response according to SARS-CoV-2 Vaccination/Infection Status
PLWH with hybrid immunity against SARS-CoV-2 (infection plus vaccination) have significantly higher levels of anti-S IgG antibodies (mean titer 653.3 BAU/mL vs. 412.4 BAU/mL in those infected only, and 464.3 BAU/mL in those vaccinated only, and higher neutralization capacity (82.96% vs. 60.97% in those infected only, and 57.06% in those vaccinated only), as observed in Figure 2. There is a significant correlation between the anti S IgG titers and the neutralizing activity against the Omicron variant. There are no significant differences in the antibodies titers according to the time since vaccination (vaccinated < 6 months vs. vaccinated 6 - 12 months ago vs. vaccinated > 12 months) (results not shown). The reactivity of anti-spike IgA antibodies is similar in all patients (vaccinated, infected or with hybrid immunity).
3.4. SARS-CoV-2 Humoral Immunity in Severe Immunosuppressed PLWH
As shown in Table 2, severe immunosuppression (CD4 count < 200 cells/mm3) is associated with significantly lower percentages of patients with detectable anti-S IgG (p = 0.0003), anti-S IgA antibodies (p < 0.0001), and with lack of neutralizing activity against Omicron variant (p < 0.0001). Patients with advanced immunosuppression have lower anti-S IgG antibodies mean titers (p = 0.005) and anti-S IgA antibodies reactivities (p = 0.002) compared to patients with mild or no immunosuppression; moreover, there is a significant correlation between the CD4/CD8 ratio and the titers of anti-S IgG and IgA antibodies (r = 0.353; p =0.0003 and r = 0.4208; p <0.0001, respectively, using non-parametric Spearman test).
3.5. SARS-CoV-2 Cellular Immunity
Cellular immunity was assessed in 15 PLWH similarly distributed in three groups: without immunosuppression - CD4 count > 500 cells/mm3, mild immunosuppression - CD4 count 200-500 cells/mm3, and severe immunosuppression - CD4 count < 200 cells/mm3. PBMCs from the patients with CD4 count lower than 200 cells/mm3 have inferior responses when stimulated with a recombinant Omicron BA.4/BA.5 S1 protein, as compared to patients with mild or absent immunosuppression (Figure 3A).
Detectable IFN-γ producing T cell responses were present in 13 out of 15 evaluated patients (86.7%), including two patients with CD4 < 200 cells/mm3 who did not display any serological markers of SARS-CoV-2 infection. There were no detectable IL-2 secreting T cells. Subjects without immunosuppression (> 500 CD4 cells/mm3) respond better to re-stimulation, although the cellular immune response displays individual heterogeneity. The non-parametric Spearman test did not detect any correlation between anti-SARS-CoV-2 specific cellular response and the CD4/CD8 ratio (r = 0.2151; p = 0.4384) (Figure 3B).
4. Discussion
We report that the majority of PLWH have preserved SARS-CoV-2 humoral and cellular immunity post-vaccination and post-infection, including anti-S IgA antibodies, neutralizing antibodies against Omicron variant, and IFN-γ secreting T cell responses. An important proportion of PLWH who have not been diagnosed with a prior SARS-CoV-2 infection and have not been SARS-CoV-2 vaccinated display serologic markers of SARS-CoV-2 infection, suggesting rather frequent asymptomatic infections, with preserved subsequent immunity.
Still, our results show important differences according to the degree of immunosuppression, with lower levels of seropositivity and lower antibodies titers in PLWH who are severely immunosuppressed (CD4 count < 200 cells/mm3), irrespective of their vaccination/infection status. A lower immunogenicity of SARS-CoV-2 vaccines has been demonstrated in numerous studies for PLWH who are severely immunocompromised, regardless of their HIV viral load [23,24,25,26]. On the contrary, PLWH on cART, with undetectable plasma HIV viral loads (less than 50 copies/mL), and CD4 counts of more than 350 cells/mm3, develop strong anti-SARS-CoV-2 humoral immune responses after vaccination, comparable in magnitude and persistence with those of people without HIV [12,23,24]. More recent studies have shown lower neutralization activity against all Omicron SARS-CoV-2 variants (SARS-CoV-2 B.1, B.1.617, BA.1, and BA.2) in PLWH with severe immunosuppression [27]. Nevertheless, the sub-variant specific neutralizing antibodies increase after a booster dose, reaching high levels in PLWH with high CD4 nadir and lower levels in PLWH with a low CD4 nadir; no substantial difference was detected in the rate of specific antibodies drop several months after variant specific booster vaccination in PLWH and HIV negative individuals [28,29].
A third of the PLWH enrolled in this study displayed no serologic markers for SARS-CoV-2, all had significantly decreased CD4 levels and tend to have detectable viral load under antiretroviral treatment. This might indicate either an (improbable) lack of exposure to SARS-CoV-2 infection, or previous asymptomatic infections, with a weaker immune response stimulation, followed by a complete waning of serologic markers. Data on decreased and volatile SARS-CoV-2 immune responses post-infection and post-vaccination have been also reported for patients with other causes of primary or secondary immunodeficiency (solid organ or stem cell transplantation recipients, patients on chimeric antigen receptor T cell therapy, or patients with chronic leukemias) [30,31,32]. The magnitude of the immune response in PLWH can be further influenced by the infecting viral variant, as some reports have demonstrated a higher neutralization activity against the Delta SARS-CoV-2 variant, but a very decreased one for the Beta variant [33]. We did not find any correlation between the presence and magnitude of the SARS-CoV-2 specific immune responses in PLHW and their age, gender or body mass index, although such associations were previously reported in the general population [34], probably due to the fact that most HIV infected persons included in this study are young, without associated comorbidities.
The majority of the HIV-positive individuals enrolled in the present study had detectable SARS-CoV-2 specific cellular immune responses following re-stimulation with a BA.4/BA.5 Spike recombinant protein. The cellular immune responses were higher in immunocompetent patients (CD4 > 500 cells/mm3). Nevertheless, SARS-CoV-2 specific cellular immune responses were also detected in two immunosuppressed PLWH, who had no serological markers of SARS-CoV-2 infection. This suggests a durable presence of cellular immunity irrespective of the individual's level of immunosuppression. Earlier studies have already demonstrated that PLWH, virally suppressed and on ART, present robust T cell response able to cross-recognize SARS-CoV-2 variants [12,17,35,36] even when the humoral immune response showed low or absent neutralization of SARS-CoV-2 [17].
Still, in PLWH with CD4 < 200 cells/mm3, the cellular immune response induced by mRNA vaccines was described as inferior to the one developed in PLWH without immunosuppression or in healthy controls [24]. Follow-up studies on the persistence of the cellular immune responses in PLWH, showed a decline 6 months after vaccination, but with no significant difference compared to healthy controls. In immunocompetent individuals, studies on the long-term durability of the cellular immune responses against SARS-CoV-2 suggest that memory T-cell responses initiated by the first viral infection retain significant cross-reactivity even after a span of 2 years [37,38].
In our study, PLWH with hybrid immunity (acquired after vaccination and infection) display the strongest immune responses against SARS-CoV-2, similar to the data available from immunocompetent persons [39,40]. This is in accordance with previous reports, showing a significant increase in antibody titers and neutralizing capacity after breakthrough SARS-CoV-2 infections in PLWH [41]. Noteworthy, in the present study, SARS-CoV-2 anti-S IgA antibodies, a surrogate marker for local immunity, were still present in most PLWH at 8 - 9 months after infection or vaccination; similar to the data reported in longitudinal follow-up studies in the general population, showing persistent adaptative systemic and local immune responses up to two years, with more durable and organized immune responses after recovery from COVID-19 [37]. Recent studies have also demonstrated that breakthrough infections with the currently circulating Omicron variants enhance both the breadth and persistence of all types of specific immune responses, including nasal spike-specific IgA levels [42]. Longitudinal studies have also demonstrated a rapid activation of SARS-CoV-2 specific cellular immune responses during breakthrough infections, explained both by the recall of CD4+ and CD8+ spike-specific memory cells, and de novo T cell responses [43]. Increased anti-SARS-CoV-2 IgA titers have been associated with better viral neutralization and protection against breakthrough infections [44,45]. Taken together, these results support prioritization of PLWH with severe immunosuppression for SARS-CoV-2 vaccination and variant adapted booster administration. A meta-analysis including 22 studies and 6522 PLWH reported that a second dose of SARS- CoV-2 vaccine is associated with an improved seroconversion, although with lower titres of antibodies in PLWH than in immunocompetent subjects [46]. Another meta-analysis based on 50 studies reported increases in the seroconversion rate achieved by PLWH after each new dose of vaccine, that remains however inferior to the one acquired in the general population [47]. In our study only 8 individuals received a vaccine booster dose, this low percentage reflects the general low level of booster uptake in Romania (< 10%) of the general population) [48]. Administration of additional vaccine doses (up to two variant adapted boosters) in PLWH, have been associated with an expansion of the specific humoral and cellular immune responses that are maintained up to 6 months [14]. Moreover, even in PLWH with CD4 < 200 cells/mm3, there is an increase of the humoral response after a third dose of SARS-CoV-2 vaccine, especially in the case of heterologous vaccination, while the cellular immune response appears to remain more stable [49]. Our results show that PLWH with preserved cellular immunity (> 500 CD4 cells/mm3) display improved cellular immune responses, with higher numbers of IFN-γ secreting T cells after stimulation with a SARS-CoV-2 Omicron S1 protein. The absence of IL-2 secreting cells might be explained by the mild or asymptomatic SARS-CoV-2 infections in our study participants. Previous reports have signaled that IL-2 secreting cells are less frequent in those with mild disease compared to those with severe or moderate forms of COVID-19 [50]. In addition, in vaccinated PLWH, SARS-CoV-2 specific T cells predominantly produce IFN-γ and TNF-α, while cells producing IL-2 are very rare [51].
The main limitation of this study is the relatively small number of participants who have been evaluated for the cellular immune response. Nevertheless, the data are still valuable, as they show persistence of the cellular immunity even after asymptomatic infections, and in persons without any serological markers of SARS-CoV-2 infection.
5. Conclusions
SARS-CoV-2 specific immune response in PLWH without immunosuppression is preserved, with neutralizing antibodies and T-cell responses persisting for up to 9 months after infection and/or vaccination. Similar to immunocompetent individuals, hybrid immunity (SARS-CoV-2 infection plus vaccination) is associated with stronger and more persistent immune responses. Longitudinal studies are needed to investigate the long-term durability of the SARS-CoV-2 immune response in the presence of multiple circulating viral variants.
Author Contributions
Conceptualization, S.R and C.S., methodology, S.R.; software, L.M.; validation, C.G., L.M. and A.P.; formal analysis, A.P.; investigation, C.P.; resources, C.P.; data curation, C.O.; writing—original draft preparation, C.S., writing—review and editing, C.S. and S.R.; visualization, L.M.; supervision, S.R.; project administration, C.P.; funding acquisition, S.R. All authors have read and agreed to the published version of the manuscript.
Funding
This work was financially supported by “Carol Davila” University of Medicine and Pharmacy Bucharest, Romania through Contract no. 33PFE/30.12.2021, funded by the Ministry of Research and Innovation within PNCDI III, Program 1—Development of the National RD system, Subprogram 1.2—Institutional Performance—RDI excellence funding projects and by a grant of the Ministry of Research, Innovation and Digitization, CNCS-UEFISCDI, project number PN-III-P1-1.1-PD-2021-0825, within PNCDI III. Publication of this paper was supported by the University of Medicine and Pharmacy Carol Davila, through the institutional program Publish not Perish.
Institutional Review Board Statement
The study was conducted in accordance with the Declaration of Helsinki and approved by the Ethics Committee of Dr Victor Babes Hospital of Infectious and Tropical Diseases, Bucharest, Romania (No. 5824/29.04.2022).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Data is unavailable due to privacy or ethical restrictions.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Dubey, A.; Choudhary, S.; Kumar, P.; Tomar, S. Emerging SARS-CoV-2 Variants: Genetic Variability and Clinical Implications. Current Microbiology 2021, 79, 20. [Google Scholar] [CrossRef]
- Bertagnolio, S.; Thwin, S.S.; Silva, R.; Nagarajan, S.; Jassat, W.; Fowler, R.; Haniffa, R.; Reveiz, L.; Ford, N.; Doherty, M.; Diaz, J. Clinical features of, and risk factors for, severe or fatal COVID-19 among people living with HIV admitted to hospital: Analysis of data from the WHO Global Clinical Platform of COVID-19. The Lancet HIV 2022, 9, e486–e495. [Google Scholar] [CrossRef]
- European AIDS Clinical Society EACS Guidelines Version 12.0, Oct 2023 PART IV 2023; p. 151.
- World Health Organization. SAGE updates COVID-19 vaccination guidance. Available online: https://www.who.int/groups/strategic-advisory-group-of-experts-on-immunization/covid-19-materials (accessed on 22 February 2024).
- Miller, K.W.; Gandhi, R.T. The severity of COVID-19 across the spectrum of HIV. Current opinion in HIV and AIDS 2023, 18, 119–125. [Google Scholar] [CrossRef]
- Höft, M.A.; Burgers, W.A.; Riou, C. The immune response to SARS-CoV-2 in people with HIV. Cellular & Molecular Immunology 2024, 21, 184–196. [Google Scholar] [CrossRef]
- Yang, X.; Sun, J.; Patel, R.C.; Zhang, J.; Guo, S.; Zheng, Q.; Olex, A.L.; Olatosi, B.; Weissman, S.B.; Islam, J.Y.; Chute, C.G.; Haendel, M.; Kirk, G.D.; Li, X. Associations between HIV infection and clinical spectrum of COVID-19: A population level analysis based on US National COVID Cohort Collaborative (N3C) data. The lancet. HIV 2021, 8, e690–e700. [Google Scholar] [CrossRef]
- Yang, X.; Zhang, J.; Liu, Z.; Chen, S.; Olatosi, B.; Poland, G.A.; Weissman, S.; Li, X. COVID-19 breakthrough infections among people living with and without HIV: A statewide cohort analysis. International Journal of Infectious Diseases 2024, 139, 21–27. [Google Scholar] [CrossRef]
- Cele, S.; Karim, F.; Lustig, G.; San, J.E.; Hermanus, T.; Tegally, H.; Snyman, J.; Moyo-Gwete, T.; Wilkinson, E.; Bernstein, M.; Khan, K.; Hwa, S.H.; Tilles, S.W.; Singh, L.; Giandhari, J.; Mthabela, N.; Mazibuko, M.; Ganga, Y.; Gosnell, B.I.; Karim, S.S.A.; Hanekom, W.; Van Voorhis, W.C.; Ndung'u, T.; Lessells, R.J.; Moore, P.L.; Moosa, M.S.; de Oliveira, T.; Sigal, A. SARS-CoV-2 prolonged infection during advanced HIV disease evolves extensive immune escape. Cell host & microbe 2022, 30, 154–162.e155. [Google Scholar] [CrossRef]
- Qu, M.-M.; Song, B.; Yang, B.-P.; Wang, Z.; Yu, M.; Zhang, Y.; Zhang, C.; Song, J.-W.; Fan, X.; Xu, R.; Zhang, J.-Y.; Zhou, C.-B.; Du, F.; Wang, F.-S.; Huang, H.-H.; Jiao, Y.-M. Effect of SARS-CoV-2 Breakthrough Infection on HIV Reservoirs and T-Cell Immune Recovery in 3-Dose Vaccinated People Living with HIV. Viruses 2023, 15, 2427. [Google Scholar] [CrossRef]
- Donadeu, L.; Tiraboschi, J.M.; Scévola, S.; Torija, A.; Meneghini, M.; Jouve, T.; Favà, A.; Calatayud, L.; Ardanuy, C.; Cidraque, I.; Preyer, R.; Strecker, K.; Lozano, J.J.; Podzamczer, D.; Crespo, E.; Bestard, O. Long-lasting adaptive immune memory specific to SARS-CoV-2 in convalescent coronavirus disease 2019 stable people with HIV. AIDS (London, England) 2022, 36, 1373–1382. [Google Scholar] [CrossRef]
- Frater, J.; Ewer, K.J.; Ogbe, A.; Pace, M.; Adele, S.; Adland, E.; Alagaratnam, J.; Aley, P.K.; Ali, M.; Ansari, M.A.; Bara, A.; Bittaye, M.; Broadhead, S.; Brown, A.; Brown, H.; Cappuccini, F.; Cooney, E.; Dejnirattisai, W.; Dold, C.; Fairhead, C.; Fok, H.; Folegatti, P.M.; Fowler, J.; Gibbs, C.; Goodman, A.L.; Jenkin, D.; Jones, M.; Makinson, R.; Marchevsky, N.G.; Mujadidi, Y.F.; Nguyen, H.; Parolini, L.; Petersen, C.; Plested, E.; Pollock, K.M.; Ramasamy, M.N.; Rhead, S.; Robinson, H.; Robinson, N.; Rongkard, P.; Ryan, F.; Serrano, S.; Tipoe, T.; Voysey, M.; Waters, A.; Zacharopoulou, P.; Barnes, E.; Dunachie, S.; Goulder, P.; Klenerman, P.; Screaton, G.R.; Winston, A.; Hill, A.V.S.; Gilbert, S.C.; Pollard, A.J.; Fidler, S.; Fox, J.; Lambe, T. Safety and immunogenicity of the ChAdOx1 nCoV-19 (AZD1222) vaccine against SARS-CoV-2 in HIV infection: A single-arm substudy of a phase 2/3 clinical trial. The lancet. HIV 2021, 8, e474–e485. [Google Scholar] [CrossRef]
- Oyaert, M.; De Scheerder, M.A.; Van Herrewege, S.; Laureys, G.; Van Assche, S.; Cambron, M.; Naesens, L.; Hoste, L.; Claes, K.; Haerynck, F.; Kerre, T.; Van Laecke, S.; Van Biesen, W.; Jacques, P.; Verhasselt, B.; Padalko, E. Evaluation of Humoral and Cellular Responses in SARS-CoV-2 mRNA Vaccinated Immunocompromised Patients. Front Immunol 2022, 13, 858399. [Google Scholar] [CrossRef]
- Oyaert, M.; De Scheerder, M.-A.; Van Herrewege, S.; Laureys, G.; Van Assche, S.; Cambron, M.; Naesens, L.; Hoste, L.; Claes, K.; Haerynck, F.; Kerre, T.; Van Laecke, S.; Jacques, P.; Padalko, E. Longevity of the humoral and cellular responses after SARS-CoV-2 booster vaccinations in immunocompromised patients. European Journal of Clinical Microbiology & Infectious Diseases 2024, 43, 177–185. [Google Scholar] [CrossRef]
- Woldemeskel, B.A.; Karaba, A.H.; Garliss, C.C.; Beck, E.J.; Aytenfisu, T.Y.; Johnston, T.S.; Laeyendecker, O.; Cox, A.L.; Blankson, J.N. Decay of coronavirus disease 2019 mRNA vaccine-induced immunity in people with HIV. AIDS (London, England) 2022, 36, 1315–1317. [Google Scholar] [CrossRef]
- Schmidt, K.G.; Harrer, E.G.; Tascilar, K.; Kübel, S.; El Kenz, B.; Hartmann, F.; Simon, D.; Schett, G.; Nganou-Makamdop, K.; Harrer, T. Characterization of Serum and Mucosal SARS-CoV-2-Antibodies in HIV-1-Infected Subjects after BNT162b2 mRNA Vaccination or SARS-CoV-2 Infection. Viruses 2022, 14, 651. [Google Scholar] [CrossRef] [PubMed]
- Touizer, E.; Alrubayyi, A.; Ford, R.; Hussain, N.; Gerber, P.P.; Shum, H.L.; Rees-Spear, C.; Muir, L.; Gea-Mallorquí, E.; Kopycinski, J.; Jankovic, D.; Jeffery-Smith, A.; Pinder, C.L.; Fox, T.A.; Williams, I.; Mullender, C.; Maan, I.; Waters, L.; Johnson, M.; Madge, S.; Youle, M.; Barber, T.J.; Burns, F.; Kinloch, S.; Rowland-Jones, S.; Gilson, R.; Matheson, N.J.; Morris, E.; Peppa, D.; McCoy, L.E. Attenuated humoral responses in HIV after SARS-CoV-2 vaccination linked to B cell defects and altered immune profiles. iScience 2023, 26, 105862. [Google Scholar] [CrossRef]
- Hensley, K.S.; Jongkees, M.J.; Geers, D.; GeurtsvanKessel, C.H.; Mueller, Y.M.; Dalm, V.; Papageorgiou, G.; Steggink, H.; Gorska, A.; Bogers, S.; den Hollander, J.G.; Bierman, W.F.W.; Gelinck, L.B.S.; Schippers, E.F.; Ammerlaan, H.S.M.; van der Valk, M.; van Vonderen, M.G.A.; Delsing, C.E.; Gisolf, E.H.; Bruns, A.H.W.; Lauw, F.N.; Berrevoets, M.A.H.; Sigaloff, K.C.E.; Soetekouw, R.; Branger, J.; de Mast, Q.; Lammers, A.J.J.; Lowe, S.H.; de Vries, R.D.; Katsikis, P.D.; Rijnders, B.J.A.; Brinkman, K.; Roukens, A.H.E.; Rokx, C. Correction: Immunogenicity and reactogenicity of SARS-CoV-2 vaccines in people living with HIV in the Netherlands: A nationwide prospective cohort study. PLoS medicine 2023, 20, e1004159. [Google Scholar] [CrossRef]
- Vergori, A.; Tavelli, A.; Matusali, G.; Azzini, A.M.; Augello, M.; Mazzotta, V.; Pellicanò, G.F.; Costantini, A.; Cascio, A.; De Vito, A.; Marconi, L.; Righi, E.; Sartor, A.; Pinnetti, C.; Maggi, F.; Bai, F.; Lanini, S.; Piconi, S.; Levy Hara, G.; Marchetti, G.; Giannella, M.; Tacconelli, E.; d'Arminio Monforte, A.; Antinori, A.; Cozzi-Lepri, A.; On Behalf Of The Vax-Icona-Orchestra, S. SARS-CoV-2 mRNA Vaccine Response in People Living with HIV According to CD4 Count and CD4/CD8 Ratio. Vaccines (Basel) 2023, 11. [Google Scholar] [CrossRef]
- Sun, J.; Zheng, Q.; Madhira, V.; Olex, A.L.; Anzalone, A.J.; Vinson, A.; Singh, J.A.; French, E.; Abraham, A.G.; Mathew, J.; Safdar, N.; Agarwal, G.; Fitzgerald, K.C.; Singh, N.; Topaloglu, U.; Chute, C.G.; Mannon, R.B.; Kirk, G.D.; Patel, R.C. Association Between Immune Dysfunction and COVID-19 Breakthrough Infection After SARS-CoV-2 Vaccination in the US. JAMA internal medicine 2022, 182, 153–162. [Google Scholar] [CrossRef]
- Infantino, M.; Pieri, M.; Nuccetelli, M.; Grossi, V.; Lari, B.; Tomassetti, F.; Calugi, G.; Pancani, S.; Benucci, M.; Casprini, P.; Manfredi, M.; Bernardini, S. The WHO International Standard for COVID-19 serological tests: Towards harmonization of anti-spike assays. International Immunopharmacology 2021, 100, 108095. [Google Scholar] [CrossRef]
- Meyer, B.; Reimerink, J.; Torriani, G.; Brouwer, F.; Godeke, G.J.; Yerly, S.; Hoogerwerf, M.; Vuilleumier, N.; Kaiser, L.; Eckerle, I.; Reusken, C. Validation and clinical evaluation of a SARS-CoV-2 surrogate virus neutralisation test (sVNT). Emerging microbes & infections 2020, 9, 2394–2403. [Google Scholar] [CrossRef]
- Nault, L.; Marchitto, L.; Goyette, G.; Tremblay-Sher, D.; Fortin, C.; Martel-Laferrière, V.; Trottier, B.; Richard, J.; Durand, M.; Kaufmann, D.; Finzi, A.; Tremblay, C. Covid-19 vaccine immunogenicity in people living with HIV-1. Vaccine 2022, 40, 3633–3637. [Google Scholar] [CrossRef]
- Antinori, A.; Cicalini, S.; Meschi, S.; Bordoni, V.; Lorenzini, P.; Vergori, A.; Lanini, S.; De Pascale, L.; Matusali, G.; Mariotti, D.; Cozzi Lepri, A.; Gallì, P.; Pinnetti, C.; Gagliardini, R.; Mazzotta, V.; Mastrorosa, I.; Grisetti, S.; Colavita, F.; Cimini, E.; Grilli, E.; Bellagamba, R.; Lapa, D.; Sacchi, A.; Marani, A.; Cerini, C.; Candela, C.; Fusto, M.; Puro, V.; Castilletti, C.; Agrati, C.; Girardi, E.; Vaia, F. Humoral and Cellular Immune Response Elicited by mRNA Vaccination Against Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) in People Living With Human Immunodeficiency Virus Receiving Antiretroviral Therapy Based on Current CD4 T-Lymphocyte Count. Clinical infectious diseases : An official publication of the Infectious Diseases Society of America 2022, 75, e552–e563. [Google Scholar] [CrossRef]
- Corma-Gómez, A.; Fernández-Fuertes, M.; García, E.; Fuentes-López, A.; Gómez-Ayerbe, C.; Rivero-Juárez, A.; Domínguez, C.; Santos, M.; Viñuela, L.; Palacios, R.; Real, L.M.; Rivero, A.; Macías, J.; Pineda, J.A.; García, F. Severe immunosuppression is related to poorer immunogenicity to SARS-CoV-2 vaccines among people living with HIV. Clinical microbiology and infection : The official publication of the European Society of Clinical Microbiology and Infectious Diseases 2022, 28, 1492–1498. [Google Scholar] [CrossRef]
- Benet, S.; Blanch-Lombarte, O.; Ainsua-Enrich, E.; Pedreño-Lopez, N.; Muñoz-Basagoiti, J.; Raïch-Regué, D.; Perez-Zsolt, D.; Peña, R.; Jiménez, E.; de la Concepción, M.L.R.; Ávila, C.; Cedeño, S.; Escribà, T.; Romero-Martín, L.; Alarcón-Soto, Y.; Rodriguez-Lozano, G.F.; Miranda, C.; González, S.; Bailón, L.; Blanco, J.; Massanella, M.; Brander, C.; Clotet, B.; Paredes, R.; Esteve, M.; Izquierdo-Useros, N.; Carrillo, J.; Prado, J.G.; Moltó, J.; Mothe, B. Limited Humoral and Specific T-Cell Responses After SARS-CoV-2 Vaccination in PWH With Poor Immune Reconstitution. The Journal of infectious diseases 2022, 226, 1913–1923. [Google Scholar] [CrossRef]
- Corma-Gómez, A.; Fernández-Fuertes, M.; Viñuela, L.; Domínguez, C.; Santos, M.; Fuentes-López, A.; Rojas, A.; Fernández-Pérez, N.; Martín-Carmona, J.; Serrano-Conde, E.; Real, L.M.; Mendoza, J.; Macías, J.; Pineda, J.A.; García, F. Reduced neutralizing antibody response to SARS-CoV-2 vaccine booster dose in people living with HIV with severe immunosuppression. J Med Virol 2023, 95, e28602. [Google Scholar] [CrossRef]
- Vergori, A.; Cozzi-Lepri, A.; Matusali, G.; Colavita, F.; Cicalini, S.; Gallì, P.; Garbuglia, A.R.; Fusto, M.; Puro, V.; Maggi, F.; Girardi, E.; Vaia, F.; Antinori, A. SARS-CoV-2 Omicron Variant Neutralization after Third Dose Vaccination in PLWH. Viruses 2022, 14. [Google Scholar] [CrossRef]
- Khan, K.; Karim, F.; Ganga, Y.; Bernstein, M.; Jule, Z.; Reedoy, K.; Cele, S.; Lustig, G.; Amoako, D.; Wolter, N.; Samsunder, N.; Sivro, A.; San, J.E.; Giandhari, J.; Tegally, H.; Pillay, S.; Naidoo, Y.; Mazibuko, M.; Miya, Y.; Ngcobo, N.; Manickchund, N.; Magula, N.; Karim, Q.A.; von Gottberg, A.; Abdool Karim, S.S.; Hanekom, W.; Gosnell, B.I.; Lessells, R.J.; de Oliveira, T.; Moosa, M.S.; Sigal, A. Omicron BA.4/BA.5 escape neutralizing immunity elicited by BA.1 infection. Nat Commun 2022, 13, 4686. [Google Scholar] [CrossRef]
- Shoham, S.; Batista, C.; Ben Amor, Y.; Ergonul, O.; Hassanain, M.; Hotez, P.; Kang, G.; Kim, J.H.; Lall, B.; Larson, H.J.; Naniche, D.; Sheahan, T.; Strub-Wourgaft, N.; Sow, S.O.; Wilder-Smith, A.; Yadav, P.; Bottazzi, M.E. Vaccines and therapeutics for immunocompromised patients with COVID-19. EClinicalMedicine 2023, 59, 101965. [Google Scholar] [CrossRef]
- Bergman, P.; Blennow, O.; Hansson, L.; Mielke, S.; Nowak, P.; Chen, P.; Söderdahl, G.; Österborg, A.; Smith, C.I.E.; Wullimann, D.; Vesterbacka, J.; Lindgren, G.; Blixt, L.; Friman, G.; Wahren-Borgström, E.; Nordlander, A.; Gomez, A.C.; Akber, M.; Valentini, D.; Norlin, A.C.; Thalme, A.; Bogdanovic, G.; Muschiol, S.; Nilsson, P.; Hober, S.; Loré, K.; Chen, M.S.; Buggert, M.; Ljunggren, H.G.; Ljungman, P.; Aleman, S. Safety and efficacy of the mRNA BNT162b2 vaccine against SARS-CoV-2 in five groups of immunocompromised patients and healthy controls in a prospective open-label clinical trial. EBioMedicine 2021, 74, 103705. [Google Scholar] [CrossRef]
- Rabinowich, L.; Grupper, A.; Baruch, R.; Ben-Yehoyada, M.; Halperin, T.; Turner, D.; Katchman, E.; Levi, S.; Houri, I.; Lubezky, N.; Shibolet, O.; Katchman, H. Low immunogenicity to SARS-CoV-2 vaccination among liver transplant recipients. Journal of hepatology 2021, 75, 435–438. [Google Scholar] [CrossRef]
- Pourcher, V.; Belin, L.; Soulie, C.; Rosenzwajg, M.; Marot, S.; Lacombe, K.; Valin, N.; Pialoux, G.; Calin, R.; Palacios, C.; Malet, I.; Zafilaza, K.; Tubiana, R.; Valantin, M.A.; Klatzmann, D.; Calvez, V.; Simon-Tillaux, N.; Marcelin, A.G. High seroconversion rate and SARS-CoV-2 Delta neutralization in people with HIV vaccinated with BNT162b2. AIDS (London, England) 2022, 36, 1545–1552. [Google Scholar] [CrossRef]
- Fernandes, M.d.C.R.; Vasconcelos, G.S.; de Melo, A.C.L.; Matsui, T.C.; Caetano, L.F.; de Carvalho Araújo, F.M.; Fonseca, M.H.G. Influence of age, gender, previous SARS-CoV-2 infection, and pre-existing diseases in antibody response after COVID-19 vaccination: A review. Molecular Immunology 2023, 156, 148–155. [Google Scholar] [CrossRef]
- Augello, M.; Bono, V.; Rovito, R.; Tincati, C.; d'Arminio Monforte, A.; Marchetti, G. Six-month immune responses to mRNA-1273 vaccine in combination antiretroviral therapy treated late presenter people with HIV according to previous SARS-CoV-2 infection. AIDS (London, England) 2023, 37, 1503–1517. [Google Scholar] [CrossRef]
- Fidler, S.; Fox, J.; Tipoe, T.; Longet, S.; Tipton, T.; Abeywickrema, M.; Adele, S.; Alagaratnam, J.; Ali, M.; Aley, P.K.; Aslam, S.; Balasubramanian, A.; Bara, A.; Bawa, T.; Brown, A.; Brown, H.; Cappuccini, F.; Davies, S.; Fowler, J.; Godfrey, L.; Goodman, A.L.; Hilario, K.; Hackstein, C.-P.; Mathew, M.; Mujadidi, Y.F.; Packham, A.; Petersen, C.; Plested, E.; Pollock, K.M.; Ramasamy, M.N.; Robinson, H.; Robinson, N.; Rongkard, P.; Sanders, H.; Serafimova, T.; Spence, N.; Waters, A.; Woods, D.; Zacharopoulou, P.; Barnes, E.; Dunachie, S.; Goulder, P.; Klenerman, P.; Winston, A.; Hill, A.V.S.; Gilbert, S.C.; Carroll, M.; Pollard, A.J.; Lambe, T.; Ogbe, A.; Frater, J. Booster Vaccination Against SARS-CoV-2 Induces Potent Immune Responses in People With Human Immunodeficiency Virus. Clinical Infectious Diseases 2022, 76, 201–209. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, Q.; Gu, X.; Ren, L.; Huang, T.; Li, Y.; Zhang, H.; Liu, Y.; Zhong, J.; Wang, X.; Chen, L.; Zhang, Y.; Li, D.; Fang, M.; Xu, L.; Li, H.; Wang, Z.; Li, H.; Bai, T.; Liu, W.; Peng, Y.; Dong, T.; Cao, B.; Wang, J. Durability and cross-reactive immune memory to SARS-CoV-2 in individuals 2 years after recovery from COVID-19: A longitudinal cohort study. The Lancet Microbe 2024, 5, e24–e33. [Google Scholar] [CrossRef]
- Ogbe, A.; Pace, M.; Bittaye, M.; Tipoe, T.; Adele, S.; Alagaratnam, J.; Aley, P.K.; Ansari, M.A.; Bara, A.; Broadhead, S.; Brown, A.; Brown, H.; Cappuccini, F.; Cinardo, P.; Dejnirattisai, W.; Ewer, K.J.; Fok, H.; Folegatti, P.M.; Fowler, J.; Godfrey, L.; Goodman, A.L.; Jackson, B.; Jenkin, D.; Jones, M.; Longet, S.; Makinson, R.A.; Marchevsky, N.G.; Mathew, M.; Mazzella, A.; Mujadidi, Y.F.; Parolini, L.; Petersen, C.; Plested, E.; Pollock, K.M.; Rajeswaran, T.; Ramasamy, M.N.; Rhead, S.; Robinson, H.; Robinson, N.; Sanders, H.; Serrano, S.; Tipton, T.; Waters, A.; Zacharopoulou, P.; Barnes, E.; Dunachie, S.; Goulder, P.; Klenerman, P.; Screaton, G.R.; Winston, A.; Hill, A.V.; Gilbert, S.C.; Carroll, M.; Pollard, A.J.; Fidler, S.; Fox, J.; Lambe, T.; Frater, J. Durability of ChAdOx1 nCoV-19 vaccination in people living with HIV. JCI insight 2022, 7. [Google Scholar] [CrossRef]
- Chivu-Economescu, M.; Bleotu, C.; Grancea, C.; Chiriac, D.; Botezatu, A.; Iancu, I.V.; Pitica, I.; Necula, L.G.; Neagu, A.; Matei, L.; Dragu, D.; Sultana, C.; Radu, E.L.; Nastasie, A.; Voicu, O.; Ataman, M.; Nedeianu, S.; Mambet, C.; Diaconu, C.C.; Ruta, S.M. Kinetics and persistence of cellular and humoral immune responses to SARS-CoV-2 vaccine in healthcare workers with or without prior COVID-19. Journal of cellular and molecular medicine 2022, 26, 1293–1305. [Google Scholar] [CrossRef]
- Ekström, N.; Leino, T.M.; Juutinen, A.; Lehtonen, T.; Haveri, A.; Liedes, O.; Vara, S.; Salo, H.; Palmu, A.A.; Nohynek, H.; Martelius, T.; Melin, M. Hybrid Immunity Improves the Immune Response after the Fourth COVID-19 Vaccine Dose in Individuals with Medical Conditions Predisposing to Severe COVID-19. Vaccines 2024, 12, 247. [Google Scholar] [CrossRef] [PubMed]
- Lapointe, H.R.; Mwimanzi, F.; Cheung, P.K.; Sang, Y.; Yaseen, F.; Speckmaier, S.; Barad, E.; Moran-Garcia, N.; Datwani, S.; Duncan, M.C.; Kalikawe, R.; Ennis, S.; Young, L.; Ganase, B.; Omondi, F.H.; Umviligihozo, G.; Dong, W.; Toy, J.; Sereda, P.; Burns, L.; Costiniuk, C.T.; Cooper, C.; Anis, A.H.; Leung, V.; Holmes, D.; DeMarco, M.L.; Simons, J.; Hedgcock, M.; Prystajecky, N.; Lowe, C.F.; Romney, M.G.; Barrios, R.; Guillemi, S.; Brumme, C.J.; Montaner, J.S.G.; Hull, M.; Harris, M.; Niikura, M.; Brockman, M.A.; Brumme, Z.L. Antibody response durability following three-dose coronavirus disease 2019 vaccination in people with HIV receiving suppressive antiretroviral therapy. AIDS (London, England) 2023, 37, 709–721. [Google Scholar] [CrossRef]
- Chen, Y.; Zhao, T.; Chen, L.; Jiang, G.; Geng, Y.; Li, W.; Yin, S.; Tong, X.; Tao, Y.; Ni, J.; Lu, Q.; Ning, M.; Wu, C. SARS-CoV-2 Omicron infection augments the magnitude and durability of systemic and mucosal immunity in triple-dose CoronaVac recipients. mBio 2024, 15, e0240723. [Google Scholar] [CrossRef]
- Koutsakos, M.; Reynaldi, A.; Lee, W.S.; Nguyen, J.; Amarasena, T.; Taiaroa, G.; Kinsella, P.; Liew, K.C.; Tran, T.; Kent, H.E.; Tan, H.X.; Rowntree, L.C.; Nguyen, T.H.O.; Thomas, P.G.; Kedzierska, K.; Petersen, J.; Rossjohn, J.; Williamson, D.A.; Khoury, D.; Davenport, M.P.; Kent, S.J.; Wheatley, A.K.; Juno, J.A. SARS-CoV-2 breakthrough infection induces rapid memory and de novo T cell responses. Immunity 2023, 56, 879–892. [Google Scholar] [CrossRef]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claër, L.; Quentric, P.; Fadlallah, J.; Devilliers, H.; Ghillani, P.; Gunn, C.; Hockett, R.; Mudumba, S.; Guihot, A.; Luyt, C.E.; Mayaux, J.; Beurton, A.; Fourati, S.; Bruel, T.; Schwartz, O.; Lacorte, J.M.; Yssel, H.; Parizot, C.; Dorgham, K.; Charneau, P.; Amoura, Z.; Gorochov, G. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Science translational medicine 2021, 13. [Google Scholar] [CrossRef]
- Denis, J.; Garnier, A.; Cheutin, L.; Ferrier, A.; Timera, H.; Jarjaval, F.; Hejl, C.; Billon-Denis, E.; , P.I.g.; Ricard, D.; Tournier, J.-N.; Trignol, A.; Mura, M. Long-term systemic and mucosal SARS-CoV-2 IgA response and its association with persistent smell and taste disorders. Frontiers in Immunology 2023, 14. [CrossRef]
- Yin, J.; Chen, Y.; Li, Y.; Wang, C.; Zhang, X. Immunogenicity and efficacy of COVID-19 vaccines in people living with HIV: A systematic review and meta-analysis. International Journal of Infectious Diseases 2022, 124, 212–223. [Google Scholar] [CrossRef]
- Zhou, Q.; Zeng, F.; Meng, Y.; Liu, Y.; Liu, H.; Deng, G. Serological response following COVID-19 vaccines in patients living with HIV: A dose-response meta-analysis. Scientific reports 2023, 13, 9893. [Google Scholar] [CrossRef]
- INSP. Raportare Vaccinari Impotriva COVID-19 Si RAPI Luna Noiembrie 06-30.11.2023 [in Romanian]. Available online: https://insp.gov.ro/wpfb-file/raportare-vaccinari-impotriva-covid-19-si-rapi_luna-noiembrie-06-30-11-2023-pdf/.
- Vergori, A.; Cozzi Lepri, A.; Cicalini, S.; Matusali, G.; Bordoni, V.; Lanini, S.; Meschi, S.; Iannazzo, R.; Mazzotta, V.; Colavita, F.; Mastrorosa, I.; Cimini, E.; Mariotti, D.; De Pascale, L.; Marani, A.; Gallì, P.; Garbuglia, A.; Castilletti, C.; Puro, V.; Agrati, C.; Girardi, E.; Vaia, F.; Antinori, A.; Amendola, A.; Baldini, F.; Bellagamba, R.; Bettini, A.; Bordi, L.; Camici, M.; Casetti, R.; Costantini, S.; Cristofanelli, F.; D’Alessio, C.; D’Aquila, V.; De Angelis, A.; De Zottis, F.; de Pascale, L.; Francalancia, M.; Fusto, M.; Gagliardini, R.; Gramigna, G.; Grassi, G.; Grilli, E.; Grisetti, S.; Iafrate, D.; Lapa, D.; Lorenzini, P.; Marani, A.; Masone, E.; Marongiu, S.; Mondi, A.; Notari, S.; Ottou, S.; Paulicelli, J.; Pellegrino, L.; Pinnetti, C.; Plazzi, M.M.; Possi, A.; Sacchi, A.; Tartaglia, E.; group, H.-V.s. Immunogenicity to COVID-19 mRNA vaccine third dose in people living with HIV. Nature Communications 2022, 13, 4922. [Google Scholar] [CrossRef]
- Lin, J.; Law, R.; Korosec, C.S.; Zhou, C.; Koh, W.H.; Ghaemi, M.S.; Samaan, P.; Ooi, H.K.; Matveev, V.; Yue, F.; Gingras, A.-C.; Estacio, A.; Buchholz, M.; Cheatley, P.L.; Mohammadi, A.; Kaul, R.; Pavinski, K.; Mubareka, S.; McGeer, A.J.; Leis, J.A.; Heffernan, J.M.; Ostrowski, M. Longitudinal Assessment of SARS-CoV-2-Specific T Cell Cytokine-Producing Responses for 1 Year Reveals Persistence of Multicytokine Proliferative Responses, with Greater Immunity Associated with Disease Severity. Journal of Virology 2022, 96, e00509–e00522. [Google Scholar] [CrossRef]
- Nkosi, T.; Chasara, C.; Papadopoulos, A.O.; Nguni, T.L.; Karim, F.; Moosa, M.-Y.S.; Gazy, I.; Jambo, K.; Team, C.K.; Hanekom, W.; Sigal, A.; Ndhlovu, Z.M. Unsuppressed HIV infection impairs T cell responses to SARS-CoV-2 infection and abrogates T cell cross-recognition. eLife 2022, 11, e78374. [Google Scholar] [CrossRef]
Figure 1.
Patients’ distribution by SARS-CoV-2 serologic status and vaccination/infection history. The classification was based on the presence or absence of SARS-CoV-2 serological markers, and self-declared vaccine history.
Figure 1.
Patients’ distribution by SARS-CoV-2 serologic status and vaccination/infection history. The classification was based on the presence or absence of SARS-CoV-2 serological markers, and self-declared vaccine history.
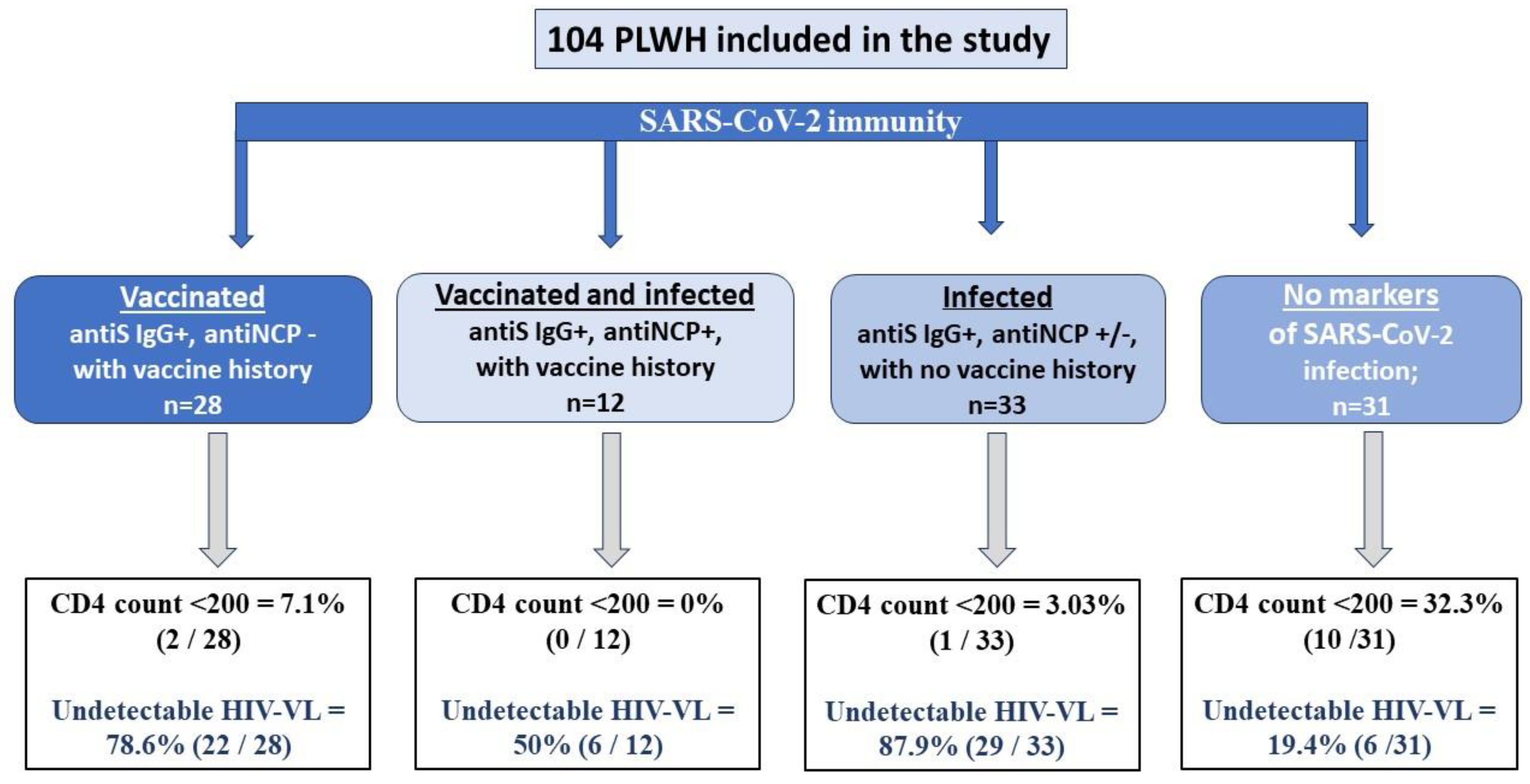
Figure 2.
Antibody responses according to SARS- CoV-2 infection/vaccination status. V = SARS-CoV-2 vaccinated; I = SARS-CoV-2 infected; V+I = SARS-CoV-2 vaccinated and infected (with hybrid immunity).
Figure 2.
Antibody responses according to SARS- CoV-2 infection/vaccination status. V = SARS-CoV-2 vaccinated; I = SARS-CoV-2 infected; V+I = SARS-CoV-2 vaccinated and infected (with hybrid immunity).
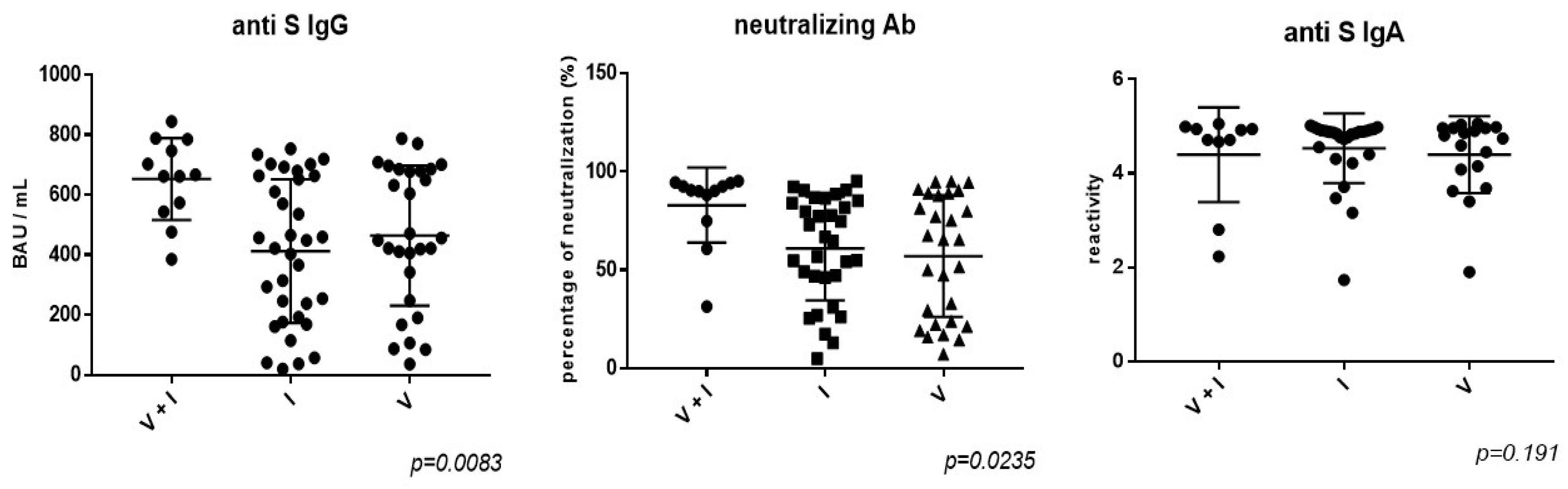
Figure 3.
SARS-CoV-2-specific T-cell responses in HIV-positive subjects. A. Number of IFN-γ SFU per 106 PBMCs by CD4 counts following stimulation with recombinant SARS-CoV-2 BA.4/BA.5 S1 protein. B. Correlation between CD4/CD8 ratio in HIV-infected individuals with their total SARS-CoV-2 responses. The non-parametric Spearman test was used for correlation analysis.
Figure 3.
SARS-CoV-2-specific T-cell responses in HIV-positive subjects. A. Number of IFN-γ SFU per 106 PBMCs by CD4 counts following stimulation with recombinant SARS-CoV-2 BA.4/BA.5 S1 protein. B. Correlation between CD4/CD8 ratio in HIV-infected individuals with their total SARS-CoV-2 responses. The non-parametric Spearman test was used for correlation analysis.
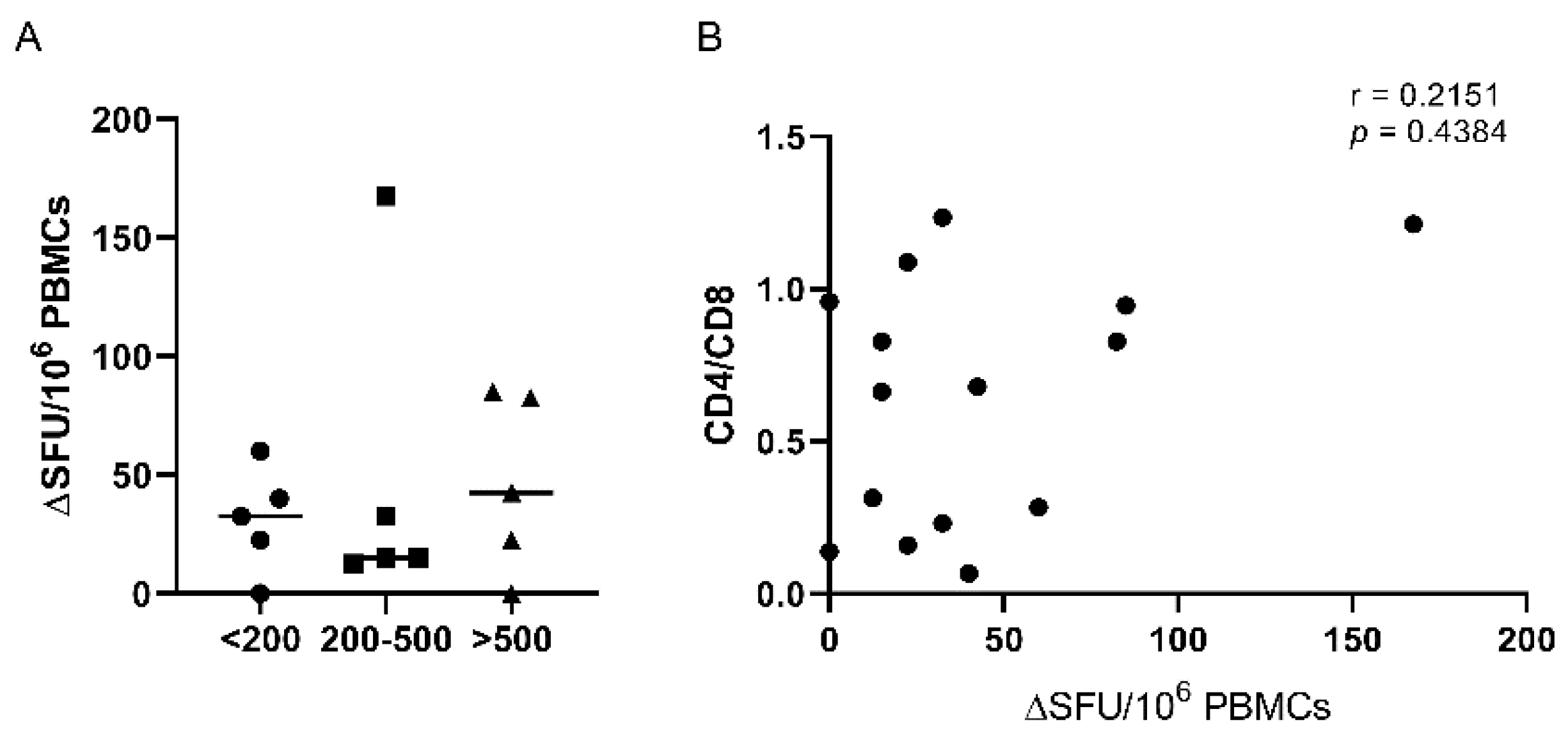

Table 1.
Characteristics of the study PLWH with or without immunosuppression.
| Patients’ immunological status | CD4 count> 500 n=62 |
CD4 count 200-500 n=29 |
CD4 count< 200 n=13 |
p |
|---|---|---|---|---|
| Male (%) | 41 (66.1%) | 20 (68.9%) | 9 (69.2%) | 0.9009 |
|
Age (years) mean ± SD |
38.3 ± 9.7 | 38.8 ± 12.3 | 38.3 ± 4.6 | 0.9736 |
| AIDS C3 (%) | 27.4 | 34.5 | 35 | 0.7107 |
|
HIV infection duration (months), median [IQR] |
132 [56-240] |
78.5 [23.5-144] |
72.5 [1-122] |
0.0376 |
| Age at HIV diagnosis (years), median [IQR] | 27 [14-35] |
26 [21-36] |
33.5 [24-37.5] |
0.5185 |
| cART treatment duration (month), mean ± SD | 108 [60-228] |
72 [36-204] |
60 [2-120] |
0.1343 |
| CD4 nadir (cells/mm3), mean ± SD | 328.9 ± 267.4 | 175.2 ± 120.5 | 86.8 ± 71.9 | 0.0003 |
|
CD4/CD8 ratio, mean ± SD |
1.16 ± 0.7 | 0.63 ± 0.3 | 0.13 ± 0.08 | <0.0001 |
|
HIV viral load (log10 HIV-RNA copies/mL), mean ± SD |
4.23 ± 3.9 | 5.1 ± 4.5 | 5.38 ± 4.6 | 0.005 |
|
Zenith HIV viral load (log10 copies/mL), mean ± SD |
5.65 ± 4.8 | 5.72 ± 4.9 | 6.1 ± 5.7 | 0.1023 |
|
Undetectable HIV RNA, n (%) |
45 (72.6%) | 16 (55.2%) | 2 (15.4%) | 0.0005 |
| Number of cART regimens, mean ± SD | 2.9 ± 2.3 | 2.96 ± 2.6 | 2.3 ± 1.6 | 0.6708 |
SD = Standard deviation; IQR = Interquartile range; cART = combination antiretroviral therapy.
Table 2.
Anti SARS-CoV-2 serological status according to the immunosuppression level.
| Patients’ immunological status | CD4 count >500 n=62 | CD4 count 200-500; n=29 |
CD4 count <200 n=13 |
p |
|---|---|---|---|---|
| Positive for anti-S IgG antibodies, n (%) | 49 (79.1%) | 21 (72.4%) | 3 (23.1%) | 0.0003 |
| Anti-S IgG titer (BAU/ml) mean ± SD | 377.6 ± 276.1 | 354.5 ± 301.1 | 106.8 ± 228.1 | 0.005 |
| Positive for anti-S IgA antibodies n (%) | 56 (90.3%) | 21 (72.4%) | 4 (30.8%) | <0.0001 |
| Anti-S IgA antibidies reactivity, mean ±SD | 3.6 ± 1.7 | 3.2 ± 1.9 | 1.4 ± 1.1 | 0.002 |
| Positive for neutralizing activity (>30%), n (%) | 51 (82.3%) | 23 (79.3%) | 2 (15.4%) | <0.0001 |
| SARS-CoV-2 neutralizing capacity, mean ± SD | 63.8 ± 27.6 | 64.2 ± 30.1 | 45.5 ± 29.7 | 0.041 |
SD = Standard deviation.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



