Submitted:
10 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
Cotton fiber, a crucial and sustainable resource for global textile production, undergoes a complex five-stage developmental process, encompassing initiation, elongation, transition, secondary cell wall biosynthesis, and maturation. These elongated single-cell fibers originate from the outer ovule epidermis. The development of cotton fibers involves intricate changes in gene expression and physiological processes, resulting in a nearly pure cellulose product vital for the global cotton industry. Decoding genes associated with fiber development enhances our understanding of cotton fiber mechanisms and facilitates the cultivation of varieties with enhanced quality. In recent decades, advanced omics approaches, including genomics, transcriptomics, and proteomics, have played a pivotal role in identifying genes and gene products linked to cotton fiber development, including the MYB transcription factor family which coordinates cotton fiber development. Molecular studies have revealed transcription factors like MYB, WRKY, Homeodomain Leucine Zipper (HD-ZIP) and basic Helix-Loop-Helix (bHLH) influencing fiber initiation and elongation. The intricate interplay of phytohormones like auxin, gibberellic acid (GA), brassinosteroid (BR), jasmonic acid (JA), ethylene, abscisic acid (ABA), and cytokinin is explored, providing a comprehensive perspective on shaping cotton fibers. Numerous candidate genes and cellular processes affecting various aspects of fiber development hold promise for genetic engineering or marker-assisted breeding to improve fiber quality. This review presents a comprehensive overview of key achievements in cotton molecular biology, with a specific emphasis on recent advancements in understanding transcription factors and phytohormones involved in cotton fiber initiation and elongation.
Keywords:
cotton fiber
; initiation
; elongation
; transcription factors
; phytohormones
1. Introduction
Cotton (Gossypium spp.) is a globally cultivated and economically significant fiber crop, playing a pivotal role in the textile industry and making substantial contributions to the global economy. The quality of cotton fibers directly influences the excellence of cotton-based textiles. Derived from single-celled epidermal seed trichomes produced by four domesticated species within the Gossypium genus, cotton stands as the most widely utilized plant textile globally. The genus, encompassing over 50 species spread across the tropics and subtropics, exhibits rich species diversity and morphological variation [1]. This diversity, especially in the context of the domestication experiment involving four species and two ploidy states within Gossypium, serves as a natural and powerful system for studying genetic, genomic, and genotype-to-phenotype transitions. Notably, Gossypium hirsutum cotton exhibits exceptional adaptability to various environmental conditions and is renowned for its capacity to yield substantial quantities of high-quality fiber, constituting approximately 95% of all planted cotton [2], while Gossypium barbadense produces luxury textiles with fine, lengthy, and robust fibers [3]. Additionally, Gossypium hirsutum requires fewer resources and displays enhanced resistance to pests and diseases compared to Gossypium barbadense. Despite domesticated diploids producing inferior fibers, they persist in local cultivation due to their adaptation to specific regional conditions [3].
Each cotton fiber is a single, elongated cotton seed coat epidermal cell. The differentiation and development of cotton fiber cells are highly complex, divided into five overlapping stages: fiber initiation [days post-anthesis (DPA), -1~3 DPA], fiber cell elongation (1~16 DPA), transition (16~20 DPA), secondary cell wall (SCW) synthesis (20~40 DPA), and fiber cell maturation (40~50 DAP) [4,5,6] (Figure 1). Transition is a distinct developmental stage that occurs between primary and secondary wall synthesis. Over several days, a stage-specific transcriptome underpins the distinctive cellular and biochemical status of the fiber cell. The galacturonosyltransferases (GAUTs) gene family, which is a critical participant in the pectin synthesis pathway, plays an important role in elongation, transition, and cell wall synthesis [7]. The often mentioned ‘maturation’ phase of fiber development is not well characterized. After SCW cellulose synthesis stops, the fiber continues to dehydrate within the closed boll and the lateral packing of cellulose increases (Figure 1A). Morphological and embryological studies, coupled with advanced microscopy techniques, have significantly contributed to our understanding of cotton fiber development. In Gossypium hirsutum, long lint fibers primarily initiate before or on the day of anthesis, while ovule epidermal cells starting at or after 3 DPA produce shorter fibers known as linters or fuzz (Figure 1B). Fiber mutants with inhibited fiber initial growth have demonstrated that the first detectable sign of fiber initiation occurs on the day of anthesis (0 DPA), with approximately 25% of epidermal cells contributing to fiber initials [8]. Vibrational sum frequency generation, attenuated total refection infrared (ATR-IR), Fourier transform Raman (FT-Raman) spectroscopy and X-ray diffraction (XRD) were all used to study the cellulose component in mature and air-dry fibers from two species, Gossypium hirsutum and Gossypium barbadense [9].
Understanding the intricate processes of initiation and elongation in cotton fibers involves the coordination of multiple genes and pathways. The integration of next generation sequencing technology, including mRNA sequencing and genome sequencing with innovative phenotype evaluation methods, has emerged as a transformative tool in comprehending the biological mechanisms governing cotton fiber development. For example, the 102 TBL (Trichome Birefringence Like) gene family members were evaluated for their involvement in fiber development from Gossypium hirsutum with two GhTBL genes (GhTBL7 and GhTBL58) showing differential expression at 10 DPA fiber. Silencing these genes resulted in a significant reduction in the fiber length at 10 DPA, suggesting a potential role for these genes in fiber elongation [10]. Recently, a total of 143 GhHMAs (heavy metal-binding domain) were detected by genome-wide identification in Gossypium hirsutum. A gene expression profile provided essential clues for the function of GhHMA genes in cotton fiber development and response to various abiotic stresses. GhHMA26 was predominantly expressed in 10 DPA fiber cells and the relative expression was higher than any other GhHMAs which indicate that GhHMA26 may positively regulate fiber elongation [11]. Genome-wide analysis of the serine carboxypeptidase-like protein family reveals Ga09G1039 is involved in fiber elongation in cotton with overexpression of Ga09G1039 significantly increasing the length of stem trichomes [12]. RNA sequencing (RNA-seq) data and N6-methyladenosine sequencing (m6A-seq) data showed that methylation of m6A affected the mRNA stability of these fiber elongation-related genes including the transcription factor GhMYB44. Overexpression of GhMYB44 reduced fiber elongation, whereas the silencing of GhMYB44 produced longer fibers [13]. Comparative phosphor-proteomic analysis between two cotton varieties, J7-1 and J14-1, revealed that phosphorylation of sucrose synthase GhSUS2 by Ca2+ dependent protein kinases GhCPK84/93 affects cotton fiber development. Moreover, ABA could promote the transcription and translation of GhCPK84 and GhCPK93, thereby enhancing the phosphorylation of GhSUS2 to impede fiber elongation [14]. A genome-wide association study (GWAS) identified a ~6.2 kb insertion, larINDELFZ, positioned at the end of chromosome 8 in fuzzless Gossypium arboreum. Comprised of a ~5.0 kb repetitive sequence and a ~1.2 kb fragment translocated from chromosome 12, this remote insertion was predicted to function as an enhancer located ~18 kb upstream of the dominant-repressor GaFZ (Ga08G0121). This finding unveiled a novel regulator of fiber/trichome development, shedding light on the significance of noncoding sequences in cotton. The large-fragment insertion activates the GaFZ gene and is associated with fuzz and trichome reduction in Gossypium arboreum [15]. Genome-wide exploration identified a total of 125, 73 and 71 full-length Catharanthus roseus receptor-like kinase 1-like (CrRLK1L) family genes in Gossypium hirsutum, Gossypium arboreum and Gossypium raimondii, respectively, with some of GhCrRLK1Ls preferentially expressed in fibers at the different stages. GhCrRLK1L104 was highly expressed in fibers at 30 DPA and overexpression of GhCrRLK1L104 in Arabidopsis increased the trichomes length which indicate its function in cell elongation [16].
Recently, fiber osmoregulation has emerged as a crucial player in regulating cotton fiber initiation and elongation. A two-year field experiment was conducted to determine whether potassium ameliorates Gossypium hirsutum fiber length by regulating osmotic and K+/Na+ homeostasis under salt stress [17]. Another study showed that low soil available phosphorus (AP) contents (P0: 3 ±0.5; P1: 6 ± 0.5 mg kg-1) inhibited the fiber cell elongation leading to reduce the maximum velocity of fiber elongation (VLmax) and fiber length, mainly due to lower malate content and V-H+-PPase activities [18]. A cell wall–localized β-1,3-glucanase, GhGLU18 was found to promote fiber elongation and cell wall thickening by degrading callose and enhancing polysaccharide metabolism [19]. Calcium is also involved in fiber development. In vitro ovule culture demonstrated Ca2+ rescued the shorter-fiber phenotype of GhIQD10 overexpression lines. GhIQD10 was expressed mainly in the transition period of cotton fiber development. GhIQD10 interacted with GhCaM7 and the interaction was inhibited by Ca2+ [20].
Phytohormones, including gibberellin acid (GA), auxin, cytokinin, brassinosteroid (BR), abscisic acid (ABA), ethylene, jasmonic acid (JA), cytokinin, salicylic acid, and strigolactone (SL), are small endogenous signaling molecules in plants [21]. Many of these hormones directly participate in fiber initiation and elongation. For example, strigolactones are a class of carotenoid-derived plant hormones that modulate cotton fiber elongation and secondary cell wall thickening. The endogenous SLs were significantly higher in fibers 20 DPA. Exogenous SLs significantly increased fiber length and cell wall thickness [22]. Fiber-specific expression of GhOR1Del, a positive regulator of carotenoid accumulation, was found to upregulate the carotenoid level in cotton fiber simultaneously increasing the contents of carotenoids, ABA, and ethylene in elongating fibers [23].
This manuscript provides a comprehensive review of the essential roles played by various transcription factors, such as MYB, WRKY, HD-ZIP and bHLH transcription factors, and phytohormones including auxin, GA, BR, JA, ethylene, ABA and cytokinin during fiber initiation and elongation. The review offers an in-depth analysis of their contributions to cotton fiber development, emphasizing their interplay and regulatory mechanisms. Understanding these factors holds immense potential for advancing our knowledge and optimizing cotton cultivation for improved fiber quality and yield.
2. Fiberless Mutants Identification
Cotton fibers originate from single cells within embryo epidermal cells, with only 25~30% of them ultimately developing into fibers [5]. The initiation and final development of fibers play a crucial role in determining cotton yield. Stewart (1975) observed a fiber density of approximately 3,300 fibers per mm2, with the ratio of fiber initials to total epidermal cells being 1:3.7 at anthesis [24]. Two types of fibers exist: lint fibers and fuzz fibers, with lint fibers possessing higher economic value. Interestingly, there is no observable phenotype difference when both types of fibers initiate on the epidermal surface [25]. Lint fibers initiate on or a day before the day of anthesis (-1 ~ 0 DPA) and elongate to 2~3.5 cm, while fuzz fibers initiate 3~5 DPA and only reach around 5~10 mm in length [24].
Several fiberless or fuzzless mutants, including four dominant (Li1, Li2, N1, and Fbl) and three recessive (n2, sma-4 (ha), and sma-4 (fz)) mutants, have been studied to identify genes and understand their interactions in the molecular mechanism of fiber initiation and elongation [26]. By crossing fuzzless and/or lintless mutants, the N1, n2, Li3, and Li4 loci were identified to control the presence or absence of lint or fuzz. For instance, N1N1 confers the presence of fuzz, n2n2 inhibits fuzz initiation and development, and duplicate gene pairs Li3Li3 and Li4Li4 determine the presence of lint. Homozygosity for li3li3 and li4li4 may also inhibit fuzz development [27]. Comparative scanning electron microscopy studies of fiber development in a normal TM-1 genotype and the near-isogenic Li1 mutant at 1 DAP and 3 DAP revealed minimal differences during early stages, suggesting that Li1 gene expression occurs later, probably during the elongation phase [28]. Cross-pollination of N1, n2, and n3 in upland cotton lines produced fiberless seeds, such as MD17 and SL1-7-1 [29]. The mutant exhibited lower short fiber content and better yarn quality than the wild type cultivar [30]. The Li2 short fiber mutation is located within a terminal deletion of chromosome 18 in cotton [31]. siRNA-induced silencing of a family of RanBP1s inhibits the elongation of cotton fiber cells in the Li2 mutant [32].
XZ142FLM, a natural fiberless mutant with well-studied genes responsible for the fiberless phenotype, stands in contrast to GhVIN1-RNAi (GhVIN1i), one of the few fiberless cotton lines associated with sugar metabolism and signaling generated through reverse genetics. Comparative transcriptome analysis between the natural fiberless mutant XZ142FLM and the transgenic fiberless line GhVIN1i, obtained by RNAi silencing of GhVIN1, identified common differentially expressed genes (DEGs) in ovules during fiber initiation. The respective DEGs were enriched in several identical pathways related to fiber initiation, revealing shared molecular regulatory networks controlling fiber initiation [33]. Several genes related to fiber initiation or elongation, such as GhMML3, GhVIN1, GhMYB25, GhHD-1, and GhHOX3, exhibited similar expression patterns in mutant XZ142FLM and the RNAi mutant GhVIN1i during fiber early development. This suggests the operation of similar mechanisms for fiber initiation in these two fiberless lines. The study sheds light on the regulatory networks mediated by GhMML3 and GhVIN1 in controlling fiber initiation in cotton [33].
In a study investigating natural antisense transcripts and siRNA control over fiber development, researchers discovered that small RNA derived from the GhMML3_A12 locus can induce self-cleavage of GhMML3_A12 mRNA, leading to the production of naked seeds and subsequent inhibition of lint fiber in N1 plants [34]. Employing a map-based cloning strategy for the first time in tetraploid cotton, they successfully cloned the naked seed mutant gene (N1) encoding a MYBMIXTA-like transcription factor 3 (MML3)/GhMYB25-like on chromosome A12, known as GhMML3_A12, associated with fuzz fiber development [34]. Phenotypic and genotypic analysis of MYB25-like alleles in cottons exhibiting various fiber phenotypes and their crossed progeny revealed that both MYB25-like_At and MYB25-like_Dt are linked to lint development. Fuzz development, on the other hand, is primarily determined by the expression level of MYB25-like_Dt at approximately 3 DPA, making MYB25-like_Dt a strong candidate for N2. Recently, cotton microtubule-associated protein GhMAP20L5 was reported to mediate fiber elongation through the interaction with the tubulin GhTUB13. In the RNA silencing plants, GhMAP20L5 expression was repressed at least 28.1% in comparison with its null plants at different development stages (0 DPA, 6 DPA, 12 DPA, and 18 DPA) which resulted in reducing fiber elongation rate, fiber length and lint percentage [35].
3. Transcriptional Regulation
3.1. MYB Transcription Factors
MYB family genes play pivotal roles across diverse biological processes, including cell cycle control, hormone signaling, secondary metabolism, meristem formation, cellular morphogenesis, and responses to abiotic stress, among others. In the context of cotton, MYB has been established as a key regulator of fiber development. A study by Suo et al. (2003) identified 55 MYBs in developing cotton ovules both before and after fiber initiation (-3 DPA to 3 DPA) [36]. GhMYB25, identified through transcriptome comparisons between wild-type and fiberless cotton mutants is implicated in this developmental pathway [25,37]. GhMYB25 is expressed in fiber initials during the initiation and elongation stages. Both GhMML3 (GhMYB25-like) and GhMYB25 were down-regulated in the outer integument of 0 DPA ovules in fuzzless or fiberless mutants, indicating their close relation to fiber initiation [38] (Figure 2). Mutants with RNAi-induced silencing of GhMYB25 exhibited shifts in the timing of rapid fiber elongation, resulting in the formation of shorter fibers. Additionally, noticeable reductions in trichomes on various plant parts and decreased seed production were observed. GhMYB25 was also identified in diploid species Gossypium arboreum (A2 genome), Gossypium raimondii (D5 genome), and the allotetraploid Gossypium hirsutum cv. Coker 315 (AD genome). Its suppression resulted in fewer fiber initials with delayed expansion, while over-expression led to more fiber initials without altering final fiber length. In the regulatory cascade controlling fiber initiation, GhMYB25-like acts upstream of GhMYB25 [39]. GhMYB25-like expression was reduced in fl mutant ovules, resulting in fiberless seeds, but over-expression did not increase fiber initials. RNAi of GhMYB25-like resulted in fiberless cotton seeds without affecting trichome development elsewhere [39].
The R2R3 MYB family transcription factor, GhMYB109, shares structural similarities with Arabidopsis AtGL1, a known regulator of leaf trichome development [40]. Further analysis of GhMYB109 expression in cotton fiber initial cells and elongating fibers suggested its direct involvement in fiber initiation and elongation [36] (Figure 2). Transgenic reporter gene analysis confirmed the fiber-specific expression of a 2-kb GhMYB109 promoter, while antisense-mediated suppression of GhMYB109 led to a substantial reduction in fiber length [40]. GhMYB109 operates downstream of initiation-related GhMYB25-like and GhMYB25 [39]. Antisense suppression of GhMYB109 leads to a reduction in fiber length. Furthermore, the diminished expression of GhMYB109 is associated with the suppression of genes involved in fiber elongation, such as GhACO1 and GhACO2, which contribute to ethylene (ET) biosynthesis, and GhTUB1 and GhACT1, encoding cytoskeletal proteins. Consequently, it is likely that GhMYB109 acts upstream of phytohormonal and cytoskeletal changes during fiber elongation [40]. An R3-MYB gene, GhCPC, was identified through cDNA microarray analysis. The overexpression of GhCPC not only delayed fiber initiation but also resulted in a significant reduction in fiber length. Experiments suggest that GhCPC may negatively regulate cotton fiber initiation and early elongation through a potential CPC-MYC1-TTG1/4 complex [41].
Guan and colleagues (2011) proposed a specific pairing of a transcription factor and its downstream target based on the co-expression of GhMYB2 and GhRDL1 in Arabidopsis. This co-expression induced ectopic seed and silique trichomes. GhMYB2 functioned similarly to Arabidopsis GL1 which targets the GhRDL1 promoter and positively regulates fiber development [42] (Figure 2). A variant within the MYB5_A12 homolog has been classified within the GL1 functional group. During the fiber initiation stage (-3 to 0 DPA) and the early elongation stage (5 to 10 DPA) in fibers, transcript levels of MYB5_A12 were higher in G. barbadense compared to Gossypium hirsutum. Both GhMYB5_A12 and GbMYB5_A12 were identified as contributors to the fiber development network, interacting with EGL3 and HOX3 [43]. GhMYB30 has been established as a key regulator in fiber development through multiple pathways. It was identified as a crucial regulator of GhMUR3, responsible for encoding a xyloglucan galactosyltransferase essential for xyloglucan synthesis during fiber elongation, GhMYB30 was also found to directly bind to the GhMUR3 promoter, activating GhMUR3 expression (Figure 2). Additionally, GhMYB46 was confirmed as a target gene of GhMYB30 through EMSA, with GhMYB46 exhibiting a significant increase in GhMYB30-silenced lines, indicating the inhibitory role of GhMYB30 on GhMYB46 expression during fiber development [44] (Figure 2).
Sucrose serves as a direct carbon source during fiber elongation. In a study investigating the role of GhMYB212 in regulating sucrose transportation into expanding fibers, GhMYB212 RNAi plants (GhMYB212i) exhibited reduced accumulation of sucrose and glucose in developing fibers, resulting in shorter fibers and a lower lint index. RNA-seq and protein−DNA binding assays indicated that GhMYB212 was closely linked to sucrose and starch transportation and metabolism pathways, directly controlling the expression of the sucrose transporter gene GhSWEET12 [45]. Interestingly, exogenous sucrose supplementation in ovule cultures did not rescue the shorter fiber phenotype of GhMYB212i. Current investigations support the notion that GhMYB212 functions as the primary regulator of fiber elongation by controlling the expression of GhSWEET12 [46] (Figure 2). m6A, the most abundant methylation modification in eukaryotic mRNA, showed elevated expression levels in the short fiber mutants Li2. The methylation of m6A affected the mRNA stability of numerous genes related to fiber elongation, including the transcription factor GhMYB44. Overexpression of GhMYB44 led to reduced fiber elongation, while the silencing of GhMYB44 resulted in the production of longer fibers [13] (Figure 2).
3.2. MYBMIXTA-like (MML) Transcription Factors
Transcription factors, particularly MYBMIXTA-like (MML), GhMML3, GhMML4, and GhMML7, play central roles in regulating fiber initiation [47] (Figure 2). Evidence suggests that MIXTA-like MYB transcription factors, such as GhMML4 and GhMML7, act as master regulators of cotton fiber initiation.
GhMML3, identified as a crucial regulator of fiber initiation, was found to be downregulated in the fiberless mutant XZ142FLM, which specifically governs fiber initiation [39] (Figure 2). Positioned in tandem with GhMML3 is GhMML4_D12, another MIXTA gene responsible for fuzz fiber development. These closely related MIXTA genes play a pivotal role in directing the production of fiber initiation in two distinct cell forms: lint and fuzz fibers. It is suggested that they might influence the same metabolic pathway in different cell types. Recent map-based cloning analyses have confirmed the significant role of GhMML3 in fiber initiation in various fiber mutants, including N1NSM, n2NSM (fuzzless but linted), and fiberless XZ142FLM. Specifically, Wan et al. (2016) localized the mutated dominant N1 gene of the fuzzless mutant N1NSM to chromosome A12, the same locus as GhMML3_At. Virus-induced gene silencing of GhMML3_At resulted in little to no fuzz fibers in cotton, indicating that GhMML3_At primarily regulates fuzz fiber initiation [34].
Through map-based cloning, the fuzzless mutant N1 was mapped to the locus linked to the MYBMIXTA-like TF 3 (MML3)/GhMYB25-like gene (located on chromosome A12) with fuzz development [34]. Further studies revealed that this locus contains two MML genes, MML3 and MML4, arranged tandemly to control fuzz initiation [34] (Figure 2). The lint fiber development gene (Li3) encodes GhMML4_D12 on chromosome D12. Decreasing the expression of GhMML4_D12 in n2NSM plants resulted in a significant reduction in epidermal cell prominence and lint fiber production [48]. Zhu et al. (2018), analyzing interspecific populations, suggested that GhMML3_Dt is the best candidate allele for the mutated recessive n2 gene, mainly regulating fuzz fiber development, while both GhMML3_At and GhMML3_Dt are associated with lint fiber development [49].
3.3. WRKY Transcription Factors
As one of the most extensive transcription factor families in plants, WRKY transcription factors play essential roles in cotton fiber development by regulating genes associated with fiber growth and quality. Studies have underscored the significance of WRKY genes across the initiation, elongation, and maturation phases, showcasing their regulatory functions. In an extensive comparative study conducted by Ding et al. (2015), WRKY transcription factor genes linked to fiber development were identified in the diploid cotton species Gossypium raimondii and Gossypium arboreum. A total of 112 WRKY genes were pinpointed in Gossypium raimondii, while 109 WRKY genes were identified in Gossypium arboreum. Transcriptome analysis revealed the involvement of many WRKY genes in specific fiber development processes and displaying distinct expression patterns between the two species [50]. One such gene, GhWRKY53, was identified as being fiber initiation-related (Figure 2). The heterologous expression of GhWRKY53 in Arabidopsis significantly increased trichome density. A yeast two-hybrid experiment identified twelve proteins interacting with GhWRKY53 from the cotton fiber cDNA library. These findings establish a foundation for further exploration of GhWRKY53’s role in cotton fiber development and offers new targets for studying putative group III WRKY genes in Gossypium hirsutum [51]. In another study by Wang et al. (2021), the investigation focused on GhWRKY16, a fiber preferential WRKY transcription factor in cotton. Experiments demonstrated that GhWRKY16 positively regulated both fiber initiation and elongation (Figure 2). Silencing GhWRKY16 in transgenic cotton lines resulted in a significant reduction in fiber protrusions on the ovule and shorter fibers compared to the wild-type. During the early stages of fiber development, GhWRKY16 was found to directly bind to the promoters of genes such as GhHOX3, GhMYB109, GhCesA6D-D11, and GhMYB25. The phosphorylation of GhWRKY16 by GhMPK3-1 was identified as essential for the transcriptional activation of downstream genes during the cotton fiber development process [52].
3.4. HD-ZIP Transcription Factors
HD-Zip proteins exhibit diverse and sometimes overlapping functions, spanning stress responses to morphogenesis and development. A decline in GhHD1 transcripts was associated with delayed fiber initiation and impaired trichome formation, while its overexpression led to enhanced fiber initiation with no discernible effect on leaf trichomes [39]. Through a genome-wide analysis, thirteen HD-ZIP IV genes were identified in Gossypium arboreum and 26 in Gossypium hirsutum [53]. Among these, three genes encoding transcription factors, GhHOX1, GhHOX2, and GhHOX3, were isolated from both cotton species [54]. GaHOX1 serves as a homolog of Arabidopsis GLABRA2 (GL2), a gene essential for cell expansion, branching, and cell wall maturation in leaf trichomes [54]. As a member of the class IV homeodomain-leucine zipper (HD-ZIP) family of transcription factors, GaHOX1 exhibits robust expression in various Gossypium arboreum tissues, with peak expression in 0 DPA ovule epidermal cells and 1 DPA fibers (Figure 2). In Gossypium hirsutum, its highest expression occurs during early fiber elongation. GaHOX1 demonstrated a potential role in fiber initiation by complementing the trichome-less phenotype of the Arabidopsis gl2-2 mutant [54].
Experimental findings from transgenic cotton plants revealed that GhHOX3 may serve as a central regulator of fiber elongation. Silencing this gene resulted in an up to 80% reduction in fiber length, yielding fuzz-like short fibers on the seeds. Conversely, overexpression of GhHOX3 led to an extension of fiber length [55]. GhHOX3 controls cotton fiber elongation in a hormone GA dependent manner (Figure 2). In the presence of GA, the HOX3 protein interacts with the HD-ZIP protein (GhHD1) and the GA repressor DELLA protein (GhSLR1). The GhHOX3-GhHD1 complex exhibits higher transcriptional activity, while GhSLR1 competitively binds to GhHOX3, impeding the transcriptional activation necessary for transmitting the GA signal for fiber cell elongation [53,56] (Figure 2).
Additionally, GhHOX4 has been identified as playing a crucial role in fiber elongation. Overexpressing GhHOX4 in cotton results in longer fibers, whereas transgenic cotton with silenced GhHOX4 displays a “shorter fiber” phenotype compared to the wild type. GhHOX4 directly activates two target genes, GhEXLB1D and GhXTH2D, promoting fiber elongation. Conversely, phosphatidic acid (PA), associated with cell signaling and metabolism, interacts with GhHOX4, hindering fiber elongation (Figure 2). These findings suggest that GhHOX4 positively regulates fiber elongation, while PA may play a role in the transition from fiber elongation to secondary cell wall formation by negatively modulating GhHOX4 in cotton [57].
3.5. bHLH Transcription Factors
Basic helix–loop–helix (bHLH) transcription factors play pivotal roles in regulating plant cell cycle and elongation processes. GhDEL65, a bHLH protein derived from Gossypium hirsutum, is a functional homolog of Arabidopsis GLABRA3 (GL3) and enhancer of GLABRA3 (EGL3), contributing to the regulation of fiber development. GhDEL65 transcripts were detected in 0 ∼ 1 DPA ovules, with notable abundance in 3 DPA fibers, indicating a potential role in early fiber development [58] (Figure 2). Ectopic expression of GhDEL65 in the Arabidopsis gl3egl3 double mutant partially rescued trichome development, confirming GhDEL65’s involvement in fiber development. Interaction studies revealed GhDEL65’s association with cotton R2R3 MYB transcription factors GhMYB2 and GhMYB3, as well as the WD40 protein GhTTG3, suggesting the existence of a MYB-bHLH-WD40 protein complex in cotton fiber cells [58] (Figure 2). Both allotetraploid cottons, Gossypium hirsutum and Gossypium barbadense, contain two subgenomes, At and Dt. In tetraploid cotton, the bHLH transcription factor GhPRE1A, expressed specifically in fiber cells from its At subgenome, is presumed to play a role in regulating fiber elongation. Simultaneously, its Dt subgenome homolog remains inactive due to a TATA-box fragment deletion in its promoter region [59]. A recently identified gene, GhbHLH282, not only plays a role in regulating fiber development but is also involved in brassinosteroid signaling [60].
Two additional bHLH transcription factors, fiber-related protein 2 (GhFP2) and ACTIVATOR FOR CELL ELONGATION 1 (GhACE1), have been reported to interact with each other, positively influencing fiber elongation [61] (Figure 2). Overexpression of GhFP2 in cotton hindered fiber elongation, resulting in shortened fiber length. Conversely, the suppression of GhFP2 expression in cotton promoted fiber development, leading to longer fibers compared to the wild type. GhACE1 promotes fiber elongation by activating the expression of GhPIP2;7 and GhEXP8, but its transcriptional activation on downstream genes may be impeded by GhFP2 [61].
3.6. Other Transcription Factors
Additionally, several other transcription factor genes, such as TCP, WD40, KNOX, CCCH Zinc Finger and C2H2 genes play crucial roles in controlling the fiber development in cotton. In a recent discovery, miR319-regulated CIN-type TCP genes, such as GhTCP4, were identified for their role in modulating elongation of cotton fiber cells [62] (Figure 2). GhTCP4 interacts antagonistically with GhHOX3, influencing the growth of cotton fiber elongation (Figure 2). During the initial stages of cotton fiber development, miR319 is abundant, maintaining its target TCPs at low levels, while GhHOX3 actively promotes the elongation of fiber cells. The overexpression of a miR319-resistant form results in reduced fiber cell elongation and shorter fibers [62]. A MYB–basic helix–loop–helix (bHLH)–WD40 gene, GhWDR, can rescue Arabidopsis trichome mutant ttg1 and interact with GhMML4 (GhMML4_D12) to regulate spinnable lint production [63]. The Cotton class II KNOX, GhKNL1, acts as a transcriptional repressor in fiber development. Silencing GhKNL1 in transgenic cotton led to longer fibers with thicker secondary cell walls, whereas dominant repression transgenic lines of GhKNL1 exhibited the opposite fiber phenotype compared to controls (Figure 2). Moreover, GhKNL1 was found to suppress the expression of GhEXPA2D/4A-1/4D-1/13A by binding to their promoters, thereby regulating cotton fiber elongation [64] (Figure 2). In another notable finding, a novel tandem CCCH Zinc Finger (TZF), GhTZF2, was discovered to regulate cotton fiber development. GhTZF2 exhibited high expression in ovule cells during the very early stages of fiber development, and knockout lines of GhTZF2 produced significantly shorter fibers with thinner cell walls, underscoring its crucial role in the modulation of cotton fiber development [33]. Recently, Liu and colleagues found that Zinc Finger Protein8 (GhZFP8) encoding a C2H2 transcription factor were abundant at 3, 6 and 30 DAP during the fiber elongation. The interference of GhZFP8 inhibited the boll expansion and fibers elongation [65] (Figure 2).
3.7. Omic Tools for Studying the Fiber Initiation and Elongation
Through two-dimensional gel electrophoresis, a total of 235 proteins displayed varying abundance between 5 to 25 DPA, forming four distinct abundance patterns throughout fiber development [66]. Contributing to the regulation of the dynamic fiber proteome, particularly in protein degradation, a cotton RING-type ubiquitin ligase (E3), GhRING1, was identified. The transcriptional activity of GhRING1 increased from 5 to 15 DPA, followed by a decrease from 15 to 23 DPA. Additionally, recombinant GhRING1 exhibited ubiquitin ligase activity in vitro [5,6,67]. To investigate the molecular mechanisms underlying cotton fiber elongation, a combination of microarray technology and quantitative real-time PCR (qRT-PCR) was employed to analyze DEGs in the Li1 mutant compared to the wild-type. A total of 1915 DEGs were identified, with 984 up-regulated genes and 931 down-regulated genes reaching their peak expression at 5 DPA. Among these, numerous transcription factors, including zinc finger, MYB, and basic-leucine zipper, as well as hormones (ACO and ABP) and other proteins, were identified. This comprehensive analysis contributes to a better understanding of the molecular basis of early fiber elongation [68].
By a genome-wide comparative transcriptome analysis using the Affymetrix cotton GeneChip in isogenic fuzzy-lintless (Fl) and normal fuzzy linted (FL) lines of Gossypium arboreum at 0 and 10 DPA, a multitude of transcription factors involved in fiber initiation and elongation were identified. Specifically, at 0 dpa, transcription factors such as AP2-EREBP, C2H2, C3H, HB, and WRKY were found to be upregulated. Conversely, at 10 dpa in the mutant Fl line, transcription factors including AP2-EREBP, AUX/IAA, bHLH, C2H2, C3H, HB, MYB, NAC, Orphans, PLATZ, and WRKY were downregulated (Hande et al. 2017). Through the re-mapping of over 380 cotton RNAseq datasets using consistent mapping strategies that encompass approximately 400-fold coverage of the genome, a total of 47 transcription factor binding sites were identified, with Dof, GATA, and ZF-HD exhibiting prominent binding frequencies across all developmental stages. Notably, the binding frequency of Homeodomain, MYB/Myb related, NAC, and WRKY TFs was relatively higher during the initiation and development phases of fiber growth [69].
Employing single-cell RNA sequencing (scRNA-seq), single-cell assay for transposase-accessible chromatin with high-throughput sequencing (scATAC-seq), and laser capture microdissection (LCM) coupled with RNA-seq (LCM-seq), Wang et al. effectively characterized the transcriptome of fiber cells and established the developmental trajectory of the fiber cell lineage in cotton ovules during the primary developmental stage. Notably, the findings indicate that the small peptide GhRALF1 and the transcription factor GhTCP14 rhythmically control fiber cell growth, potentially through the modulation of hormone signaling, extracellular pH, and the metabolism of mitochondria and protein translation [70].
By utilizing DNase I hypersensitive sites sequencing (DNase-Seq) and RNA-seq data, a high-confidence regulatory network was constructed for cotton ovules at 0 and 3 DPA, as well as fibers at 8, 12, 15, and 18 DPA. Distinct chromatin accessibilities were observed in the ovules (0 and 3 DPA) compared to the fiber elongation stages (8, 12, 15, and 18 DPA). The researchers constructed a robust regulatory network of transcription factors associated with ovule and fiber development, leveraging chromatin accessibility and gene co-expression networks. Within this network, the identification of a novel transcription factor, WRKY46, emerged, suggesting its potential role in shaping fiber development by regulating lignin content [71] (Figure 2). The analysis of cis-regulatory modules also revealed the impact of hormones on fiber development, emphasizing the regulatory divergence of transcription factor motifs. This study unveiled dynamic chromatin accessibility during ovule and fiber developmental stages, highlighting the involvement of numerous transcription factors in fiber development [71].
4. Phytohormonal Regulation
Phytohormones play pivotal roles as small endogenous signaling molecules in plants, and their influence on cotton fiber development has been a subject of considerable research. Among these hormones, auxin, gibberellin (GA), jasmonic acid (JA), ethylene, cytokinin, abscisic acid (ABA), and brassinosteroid (BR) have been identified as key regulators in various stages of fiber development. Auxin, ethylene, GA, JA, and BR have been identified as positive regulators, playing crucial roles in fiber initiation and elongation processes. Conversely, CK and ABA exert a negative influence on fiber growth. In in vitro culturing of cotton ovules, the addition of exogenous Auxin and GA induces fiber initiation, emphasizing its positive regulatory role. On the contrary, abscisic acid inhibits fiber growth in this system. Notably, both auxin and GA have the capacity to overcome the inhibitory effects on total fiber production induced by ABA [72]. This intricate interplay between various phytohormones highlights the complexity of regulatory networks governing cotton fiber development. Understanding these hormone-mediated pathways is crucial for devising strategies to optimize fiber yield and quality in cotton cultivation.
4.1. Auxin
During the development of cotton ovules, the concentration of indole-3-acetic acid (IAA) exhibits distinct temporal dynamics. Prior to anthesis, there is a notable surge in IAA levels, peaking at 2 DPA, followed by a gradual decrease to approximately one-fifth of the initial concentration. Subsequently, a rapid increase is observed between 7~9 DPA [73]. Maintaining a consistent supply of IAA within this critical time frame has been shown to yield significant effects on fiber development. Chen and Guan reported that such supplementation resulted in an increase in the number of fiber cell initials by 14–19% while concurrently reducing the number of fuzz fibers [74]. To investigate the potential causal link between auxin accumulation around anthesis and lint-fiber development, Zhang et al. employed the FBP7 promoter, an epidermal-specific promoter active in the cotton ovule epidermis from -2 to 10 DPA. The transgenic plants carrying FBP7::iaaL demonstrated a significant reduction in the number of fiber initials in 0 DPA ovules, providing experimental evidence that verifies the crucial role of IAA in initiating fiber cells [75]. The utilization of the synthetic response element DR5, consisting of a tandem direct repeat of 11 bp containing the auxin responsive element TGTCTC, has provided insights into the dynamics of IAA accumulation in cotton ovules during fiber cell differentiation and initiation (from -2 to 2 DPA). The expression of DR5::GUS indicates that IAA accumulation in fiber initials commences before flower opening [76]. These findings underscore the intricate regulatory network involving the Aux/IAA gene family, Auxin response factors (ARFs), and IAA accumulation in cotton fiber development. In a study by Gokani and Thaker, an analysis of fibers was conducted on three cotton cultivars: Gossypium hirsutum hybrid-4 (H-4), hybrid-8 (H-8), and Gossypium arboreum G.Cot-15. This analysis revealed the involvement of auxin in fiber elongation, both in vitro and in vivo [77].
ARFs represent essential components in the auxin signaling pathway, orchestrating the expression of early auxin-responsive genes through binding to auxin response elements (AuxRE). In Gossypium hirsutum, the expression pattern of ARFs showed significant upregulation in the GhARF2 and GhARF18 subfamilies during the fiber initiation stage (Figure 3). Subsequent yeast-one-hybrid studies confirmed the involvement of six downstream transcription factors in this regulatory network [78]. GhARF2b exhibits specific expression in developing ovules and fibers. When GhARF2b is overexpressed using a fiber-specific promoter, it hinders fiber cell elongation while simultaneously promoting initiation. Conversely, the downregulation of GhARF2b through RNAi leads to a reduction in the number of fibers but an increase in their length [79].
The Aux/IAA gene family plays a crucial role in interacting with auxin response factors to coordinate the regulation of auxin response genes throughout plant development. Han et al. conducted a study identifying nine GhAux genes, ranging from GhAux1 to GhAux9. Notably, GhAux4, GhAux5, GhAux6, and GhAux7 exhibit preferential expression in ovules on the day of anthesis, while GhAux1, GhAux2, and GhAux3 are abundantly expressed in vegetative organs. Additionally, GhAux8 and GhIAA16 display preferential expression during fiber developmental stages, with GhAux8 prominent in early fiber elongation stages and GhIAA16 during fiber initiation and secondary cell wall thickening stages [80] (Figure 3). Expression profiling further strengthens the pivotal role of auxin in the elongation of fiber cells. Specifically, the expression of the auxin binding protein GhABP significantly increases by approximately 59-fold from 0 to 10 DPA. Additional analyses reveal that GhABP expression is exclusive to elongated fibroblasts, with no presence in undifferentiated epidermal cells [81] (Figure 3).
Auxin is known to activate Rac-like GTPases and TCP transcription factors which, in turn, trigger the expression of auxin-responsive genes in plants [82,83]. Specifically, in the context of cotton fiber development, the Rac genes GhRacA and GhRacB exhibit elevated accumulation during the initiation and elongation stages of seed fibers (Figure 2 and Figure 3). This observation suggests a significant involvement of GhRacA and GhRacB in the early phases of fiber development, potentially acting as key mediators in the auxin-stimulated processes [82,84]. The introduction of exogenous indole-3-acetic acid (IAA) specifically triggers increased expression of GhTCP14, prominently present during the fiber cell initiation and elongation stages [83] (Figure 2 and Figure 3). GhTCP14, in turn, directly binds to the promoters of genes encoding crucial components such as the auxin uptake carrier (AUX1), the auxin response protein (IAA3), and the auxin efflux carrier (PIN2). This direct interaction suggests that GhTCP14 likely plays a role in IAA-mediated differentiation and elongation of cotton fiber cells, providing a key connection between TCP transcription factors and the intricate processes involved in cotton fiber development [83].
Polar auxin transport, primarily facilitated by PIN-FORMED (PIN) proteins, plays a crucial role in auxin-triggered organogenesis across various plant species. In cotton, the mRNA of GhPIN3a was identified in the outer integument of ovules, encompassing fiber cells. Suppression of GhPIN1a was found to hinder both fiber initiation and elongation, emphasizing the significance of GhPIN-mediated auxin transport in the accumulation of fiber-specific auxin in cotton [85] (Figure 3). Recent investigations have highlighted the cell-specific degradation of GhPIN3a, a process guiding the establishment of the auxin gradient in cotton ovule epidermal cells. This degradation is linked to the preferential expression of GhROP6 GTPase in fiber cells. GhROP6, in turn, reduces the abundance of GhPIN3a at the plasma membrane and facilitates intracellular proteolysis of GhPIN3a. Overexpression and activation of GhROP6 lead to enhanced cell elongation, resulting in a significant improvement in cotton fiber length [86] (Figure 3).
4.2. Gibberellic Acid (GA)
GA serves as a pivotal plant hormone regulating diverse developmental processes, including root and stem elongation, seed germination, trichome development, flowering, and fruit ripening. Like auxin, GA exhibits a positive influence on cotton fiber initiation and elongation. The application of exogenous GAs has been shown to promote both fiber initiation and elongation in cultured ovules. In contrast, the introduction of the GA biosynthesis inhibitor paclobutrazol leads to a reduction in the number and length of fibers compared to controls without hormone treatment [87,88,89,90]. The endogenous level of GA3, a bioactive gibberellic acid form, is notably higher in long-staple cotton varieties compared to medium- and short-fiber varieties [91].
Transcriptomic analyses have unveiled the up-regulation of genes involved in GA metabolism and signaling, such as those encoding GA 20-oxidase (GA20ox) during fiber initiation and elongation (Figure 3). Notably, the overexpression of GhGA20ox1 in cotton results in a significant increase in endogenous bioactive GA content in 0 DPA ovules and 10 DPA fibers. This enhancement correlates with an increased number of fiber initials and elongating fibers at 0 to 3 DPA [92]. In transgenic fiber cells overexpressing GA 20-oxidase, the sucrose synthase gene GhSusA1 exhibits significantly higher transcript levels and sucrose synthase activity compared to wild-type fiber cells [93]. Additionally, the exogenous application of bioactive GA enhances GhSusA1 transcription in both fiber cells and hypocotyls [93]. These findings indicate that GA regulates the expression of sucrose synthase genes, thereby promoting cotton fiber cell development.
DELLA proteins, act as negative regulators within the GA signaling pathway and engage in interactions with transcription factors or key regulatory proteins. These interactions serve to impede their binding to target genes or dampen their transcriptional activation activities [94]. In cotton, the DELLA protein GhGAI1 is transcribed at higher levels in −1 ~ 3 DPA ovules of the fuzzless-lintless mutant XZ142FLM compared to its wild-type counterpart, suggesting a negative regulatory role during cotton fiber initiation [95]. In the presence of GA, it binds the receptor of GID1, initiating the ubiquitin-mediated degradation of DELLA proteins. This process liberates essential transcription factors, promoting the expression of GA-responsive genes [94,96]. Cotton DELLA proteins, GhGID1 and GhSLR1, exhibit specific expression in fiber cells, and their interaction is triggered in response to GA. The ectopic expression of GhSLR1 in Arabidopsis induces a dwarf plant phenotype and amplifies the transcription of GA-responsive genes [91,97]. The transcription factor GhHOX3 emerged as a central regulator in the GA signaling pathway [55]. In conditions of low GA levels, the DELLA protein GhSLR1 specifically interacts with GhHOX3, impeding GhHOX3 from regulating its target genes. However, under high GA levels, GhSLR1 protein degradation releases GhHOX3, enabling its interaction with GhHD1. This interaction promotes the expression of two cell wall genes, GhRDL1 and GhEXPA1, thereby contributing to cotton fiber elongation [55]. It was discovered recently that GA stimulated fiber elongation through the degradation of GhSLR1 resulting in the free of the two transcription factors GhBLH1and GhZFP8 [98]. GhBLH1 binds to GhKCS12 promoter and activates its expression to enhance VLCFAs biosynthesis. GhZFP8 activate the GhSDCP1 which will upregulate the expression of GhPIF3 gene associated with fiber elongation [98].
4.3. Brassinosteroids (BR)
BR categorized as steroid hormones in plants, assume crucial roles in cotton fiber development. The application of a low concentration of brassinolide, a bioactive BR derived from plants, notably enhances fiber cell elongation in cotton. Conversely, the use of brassinazole (BRZ), a BR biosynthesis inhibitor, significantly impedes fiber cell development in vitro. In this context, brassinolide (BL) has been observed to stimulate the expression of genes associated with elongation, such as EXP, XTH, AGP, and GhTUB1, while brassinazole (BRZ) has the opposite effect, inhibiting their expression [99] (Figure 3). Furthermore, the application of finasteride, a steroid 5α-reductase inhibitor, significantly hinders fiber elongation. Notably, this inhibitory effect can be reversed by the application of BR, highlighting the regulatory role of BR in cotton fiber development [100]. Applying brassinazole externally to cotton floral buds results in significant abnormalities in fiber cell differentiation. Steroid reduction, a crucial step in brassinosteroid (BR) biosynthesis catalyzed by steroid 5α-reductase (DET), is recognized as a major rate-limiting process. The expression levels of GhSMT1 and GhDET2 mRNA in cotton ovules increase from 0 DPA to 10 DPA and subsequently decrease at 20 DPA ovules of the fiberless mutant compared to the wild-type [101]. GhDET2, a cotton steroid 5α-reductase, exhibits heightened expression levels during both fiber cell initiation and elongation phases (Figure 3). Silencing of cotton GhDET2 inhibits fiber initiation, while the upregulation of GhDET2 expression driven by the seed coat-specific promoter pFBP7 enhances fiber numbers [100]. Additional genes related to BR biosynthesis, such as GhPAG1, plays a role in cotton fiber development by modulating the concentration of endogenous BRs [102]. The pag1 mutant characterized by dwarfism and diminished fiber length attributed to a pronounced inhibition of cell elongation and expansion. Application of BL effectively restored its growth and promoted fiber elongation [102].
Overexpression of Gh14-3-3 L in cotton promotes fiber elongation, resulting in enhanced mature fiber length, while silencing Gh14-3-3 L significantly hampers the initiation and elongation of fiber cells [103] (Figure 3). This reduction in fiber length can be partially restored by the exogenous application of brassinosteroid (BR). Subsequent investigations revealed that external BR application induces the expression of brassinazole-resistant 1 (GhBZR1), and the phosphorylation of BZR1 by GhBIN2 kinase facilitates its binding to an acidic regulatory protein Gh14-3-3. Moreover, Gh14-3-3 L was found to interact with GhBZR1, and the 14-3-3-regulated GhBZR1 directly binds to the promoters of GhXTH1 and GhEXP, thereby regulating gene expression during the fiber cell elongation stage [103] (Figure 3).
Brassinosteroids (BRs) also play a role in regulating fiber growth through the involvement of the transcription factors. The cotton bHLH protein GhFP1 has been identified as a positive regulator of fiber elongation (Figure 3). Transgenic cotton overexpressing GhFP1 displayed significantly longer fiber length compared to the wild type, while the suppression of GhFP1 expression hindered fiber elongation. Notably, the expression of brassinosteroid (BR)-related genes was markedly upregulated in fibers of GhFP1-overexpressing cotton but downregulated in GhFP1-silenced fibers. The BR content in the transgenic fibers exhibited significant alterations compared to that in the wild type [104]. Brassinosteroids (BRs) play a crucial role in plant growth and development through the action of BRI1-EMS-SUPPRESSOR1 (BES1)/BRASSINAZOLE-RESISTANT1 (BZR1) transcription factors (Figure 3). In-depth insights into the regulation of cotton fiber development by GhBES1.4 were obtained through combined analysis of DAP-seq and RNA-seq data from GhBES1.4 overexpression and RNAi lines. GhBES1.4 overexpression positively influenced fiber elongation, while GhBES1.4 silencing resulted in reduced fiber length. The integrated approach of GWAS, RNA-seq, and DAP-seq identified seven genes directly regulated by GhBES1.4, including Cytochrome P450 84A1 (GhCYP84A1) and 3-hydroxy-3-methylglutaryl-coenzyme A reductase 1 (GhHMG1), both promoting cotton fiber elongation [105]. Furthermore, BR controls cotton fiber elongation by modulating very-long-chain fatty acid (VLCFA) biosynthesis. GhBES1.4 regulates endogenous VLCFA contents and directly binds to BR RESPONSE ELEMENTS (BRREs) in the GhKCS10_At promoter region (Figure 3). This interaction regulates GhKCS10_At expression, leading to increased endogenous VLCFA contents. GhKCS10_At overexpression promotes cotton fiber elongation, while GhKCS10_At silencing inhibits fiber growth, emphasizing GhKCS10_At’s positive regulatory role in fiber elongation [106].
4.4. Jasmonic Acid (JA)
The influence of JA on fiber initiation follows a dose-dependent pattern. Cotton ovules cultivated in a medium containing 0.001 μM JA exhibited increased fiber cell initiation, while a higher concentration of 2.5 μM inhibited fiber initiation. This suggests that maintaining an optimal concentration of JA is crucial for effective fiber initiation [107] (Figure 3).
In exploring the molecular aspects, four members of the allene-oxide cyclase (AOC) family, integral to JA biosynthesis, were concurrently upregulated during fiber initiation, particularly at -1 DAP in linted-fuzzed TM-1 compared to other tissues and organs (Figure 3). Real-time quantitative PCR analysis of different fiber mutants indicated that the expression levels of four JA biosynthesis enzymes, the AOC genes (GhAOC1~GhAOC4), were higher in −1 DPA ovules of fiberless mutants compared to linted–fuzzless and linted–fuzzed lines. These genes exhibited increased expression under JA treatment, with predominant expression in −3~1 DPA ovules, particularly at −1 DPA, signifying their crucial regulatory role during fiber initiation [108]. This aligns with the outcomes observed in vitro ovule cultures with different concentrations of JA.
The JASMONATE ZIM-DOMAIN (JAZ) protein stands out as a pivotal inhibitory factor in the jasmonic acid (JA) signaling pathway. Specifically, GhJAZ2 exhibits heightened expression during the fiber initiation stage, and the upregulation of GhJAZ2 expression in cotton has been found to impede fiber initials [109] (Figure 3). This suppression is achieved through the interaction of GhJAZ2 with GhMYB25-like, leading to the inhibition of GhMYB25-like activity. Furthermore, GhJAZ2 interacts with various proteins, including GhGL1, GhMYC2, and GhWD40 (Figure 3) which constitute the core components of the WD-repeat/bHLH/MYB transcriptional complex, known to be integral to fiber development. This interaction adds a layer of complexity to understanding the regulatory mechanisms underlying cotton fiber development [109].
4.5. Ethylene
Ethylene biosynthesis emerges as a pivotal pathway up-regulated during cotton fiber cell elongation, as revealed by physiological and gene expression analyses [101]. Gene expression profiling experiments highlighted the heightened expression of 1-Aminocyclopropane-1-Carboxylic Acid Oxidase (ACO1-ACO3) genes, crucial for ethylene production, particularly during the 10-15 DPA phase of fiber elongation. Experiments involving the exogenous application of ethylene reported increased fiber cell expansion, while the ethylene inhibitor 2-aminoethoxyvinyl glycine (AVG) hindered fiber growth [101] (Figure 3). These findings underscore the significant role of ethylene in supporting cotton fiber growth and elongation. Moreover, ethylene is proposed to enhance cell elongation by upregulating the expression of tubulin, sucrose synthase, and expansin genes [101].
Ethylene appears to play a dual role in fostering fiber elongation by facilitating the generation of hydrogen peroxide (H2O2), identified as a reactive oxygen species (ROS) with a substantial impact on in vitro fiber cell elongation [110] (Figure 3). Ascorbate peroxidase (APX), an enzyme responsible for scavenging ROS, participates in regulating intracellular ROS levels [111]. In the context of wild-type cotton, GhAPX1 exhibits heightened expression in 5 DPA fiber cells compared to ovules in the fl mutant [112]. The application of exogenous H2O2 significantly triggers the transcription of GhAPX1 and augments APX activity [110]. Furthermore, ethylene, when externally applied, stimulates the production of H2O2 during fiber elongation, suggesting a downstream connection between H2O2-induced cotton fiber elongation and the ethylene signaling pathway. It becomes apparent that ROS regulate the accumulation of Ca2+, thereby fostering fiber elongation, primarily by promoting ethylene production [113]. The overexpression of calmodulin GhCaM7 has been identified as a promoter of fiber elongation, whereas GhCaM7 RNAi plants exhibit delayed fiber initiation and inhibited fiber elongation [114] (Figure 3). These findings collectively underscore the intricate interplay of ethylene, ROS, and calmodulin in regulating essential processes during cotton fiber development.
Saturated very long-chain fatty acids (VLCFAs) play an important role in orchestrating the regulation of cotton fiber growth by promoting ethylene production [115]. Ethylene can counteract the inhibition of fiber cell elongation induced by 2-chloro-N-[ethoxymethyl]-N-[2-ethyl-6-methyl-phenyl]-acetamide (ACE), an inhibitor of VLCFA biosynthesis. Remarkably, VLCFA is unable to overcome the inhibition caused by AVG. In vitro application of C24:0 fatty acids has been shown to induce a substantial increase in ACO transcripts, resulting in a significant surge in ethylene production. The overexpression of KCS6, a pivotal gene in VLCFA biosynthesis within Upland cotton, yields a remarkable increase in the final length of the fiber (approximately 6.0–12%), suggesting that VLCFAs may operate upstream in the ethylene pathway [113]. These findings highlight the intricate interconnection between VLCFAs, ethylene production, and the regulation of essential processes governing cotton fiber development.
4.6. Abscisic Acid (ABA)
Past investigations have demonstrated that the application of ABA in vitro not only hinders the initiation of cotton fiber cells but also impedes the elongation of cotton fibers [116] (Figure 3). The application of exogenous abscisic acid (ABA) in vitro was found to inhibit cotton fiber development, while treatment with an ABA inhibitor (ABAI) promoted fiber development [117]. In a recent investigation, the physiological changes and proteomic profiles of Gossypium hirsutum ovules were examined after 20 days of ABA or ABAI treatment. The results revealed significant alterations compared to the control with fiber length notably reduced under ABA treatment and increased under ABAI treatment [117]. This inhibitory effect is linked to an elevation in ABA levels. Analyzing the endogenous ABA content in different fiber cells revealed a gradual increase during the initiation and elongation stages of fiber cells (0–10 DPA), followed by a decline in the period of rapid elongation (10–20 DPA). Subsequently, ABA levels returned to their original low state during the maturation stage (30–50 DPA) [118]. Research findings indicate an elevated level of ABA in mature cotton fruits compared to their younger, healthy counterparts [119]. In line with these discoveries, the concentration of ABA in fruits exhibits a low point at the time of anthesis, decreasing over the subsequent two days. It then undergoes a substantial increase, reaching up to 15-fold between 2 DPA and 10 DPA, before decreasing again and becoming undetectable up to 30 DPA. From 30 to 50 DPA, there is a renewed surge in ABA levels [5]. These findings collectively underscore the intricate regulatory role of ABA at various stages of cotton fiber development. Throughout development, the ABA content in short-staple fibers consistently surpasses that in long-staple fibers [120]. Furthermore, the levels of endogenous ABA in cotton ovules exhibit a positive correlation with short fiber production [121]. The short fiber cotton mutant Li1 exhibits a significantly higher accumulation of ABA in 0 DPA ovules compared to the wild type [122]. At the early fiber initiation stage in the Xu142 fl mutant, a notable increase in ABA levels was observed [123]. This aligns with the previously mentioned correlation between endogenous ABA levels in cotton ovules and short fiber production. Additionally, the short fiber mutant Li2 exhibited a significantly higher deposition of ABA in 0 DPA ovules compared to the wild type [122]. These findings collectively lead to the conclusion that there exists an inverse relationship between ABA content and fiber length. Dasani and Thaker conducted a comprehensive analysis on fibers from three cotton cultivars, Gossypium hirsutum hybrid-4 (H-4), hybrid-8 (H8), and Gossypium arboreum G.Cot-15, to elucidate the role of ABA in fiber elongation under both in vitro and in vivo conditions [124].
Recently, within transgenic cotton featuring the fiber-specific promoter proSCFP driving the expression of GhOR1Del, a positive regulator of carotenoid accumulation, notable alterations in hormonal content were observed during fiber elongation. Specifically, there was an increase in both abscisic acid (ABA) and ethylene levels in elongating fibers. The downstream regulator of ABA, GhbZIP27a, demonstrated a capacity to stimulate the expression of the ethylene synthase gene GhACO3 by binding to its promoter. This interaction strongly suggests that ABA plays a role in promoting fiber elongation by augmenting the production of ethylene [23].
A comprehensive proteomic analysis identified a total of 7321 proteins, including 365 and 69 differentially abundant proteins in the ABA versus control and ABAI versus control comparisons, respectively. Notably, proteins associated with phenylpropanoid biosynthesis were upregulated after ABA treatment, suggesting a crucial role of this pathway in the response to ABA. Additionally, three auxin-related proteins were upregulated, indicating the potential involvement of auxin in the regulation of fiber development under ABAI treatment. Furthermore, the enrichment of indole alkaloid biosynthesis in the ABAI group hinted at a possible promotion of fiber elongation [117]. These findings highlight the highly interconnected network of ABA, its inhibitor, and the diverse molecular pathways influencing cotton fiber development at both physiological and proteomic levels.
4.7. Cytokinin
Cytokinin plays a pivotal role in regulating various aspects of plant development, including cell division, tissue and organ senescence, and apical dominance. While cytokinin is initially present at a relatively low concentration in unfertilized ovules, its levels progressively rise after anthesis. Notably, the introduction of exogenous cytokinins into ovule culture medium has been found to significantly enhance ovule growth but concurrently inhibits the elongation of fiber cells [116] (Figure 3). Interestingly, the accumulation of endogenous cytokinin is predominantly observed in wild-type ovules, with limited presence in fiber cells [125]. To modulate cytokinin levels, one approach involved the overexpression of isopentenyl transferase (IPT), a rate-limiting enzyme crucial in cytokinin biosynthesis [126]. This experiment provided insights into the intricate balance required for cytokinin regulation and its differential impact on ovule and fiber cell development in plants. Cytokinin oxidase/dehydrogenase (CKX), a pivotal enzyme responsible for the cleavage of the unsaturated side chain of cytokinin N6, is a significant negative regulator in cytokinin metabolism. Suppressing CKX expression was shown to elevate endogenous cytokinin levels in plants [127]. Utilizing RNAi technology to silence GhCKX transcripts in transgenic cotton plants resulted in a notable increase in seed number and a slight enhancement in fiber yield [128]. The enhanced expression of the GhIPT gene in cotton, whether controlled by the CaMV35S promoter or a seed-specific promoter, leads to a substantial increase in cytokinin accumulation. Surprisingly, this heightened cytokinin level does not exhibit any discernible impact on fiber yield or quality [49]. In summary, cytokinin exhibits a crucial role in ovule development, yet paradoxically, it appears to play a negative role in the growth of fiber cells in cotton. These findings highlight the intricate and context-dependent influence of cytokinin in multiple aspects of cotton plant physiology.
5. Conclusions
Gaining insight into the underlying genetic and molecular mechanisms of fiber initiation and elongation in cotton is a crucial step to further success in breeding higher quality cotton fiber and elevating the overall fiber yield. An in-depth exploration of the intricate mechanisms dictating fiber development enables researchers and breeders to develop improved cultivars with traits modified at the genomic level. The significance of fiber length and strength cannot be overstated, as they serve as the paramount parameters shaping the quality of cotton and thereby exerting a direct effect on the performance and quality of textiles derived from these fibers. The onset of fiber initiation is intricately linked to lint formation, directly impacting the overall cotton yield. Among the various molecular mechanisms governing fiber development, transcriptional regulation and phytohormonal regulation emerge as the two key players guiding this intricate process.
It is imperative to delve into the molecular networks orchestrating fiber initiation, elongation, and the subsequent maturation of secondary walls in cotton fibers. Transcriptional regulation to control gene expression and phytohormonal regulation, of plant hormones, constitute the backbone of these interrelated processes. Unraveling the genetic factors and signaling pathways associated with these mechanisms is indispensable for deciphering the genetic architecture that underlies optimal fiber length and strength. By understanding the interplay between these molecular elements, researchers can develop more targeted breeding strategies, leveraging this knowledge to produce cotton cultivars with superior fiber attributes. The identification of key genes and pathways through advanced molecular techniques facilitates the selection of cotton varieties that exhibit the desired fiber characteristics.
Author Contributions
Conceptualization, F. B. writing—original draft preparation, J. S. writing-review and editing. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by U. S. Department of Agriculture, Agricultural Research Service project 6066-21000-053-000D.
Acknowledgments
We would like to thank Dr. Candace Haigler from North Carolina State University and Dr. David Fang from USDA-ARS-SRRC, New Orleans for providing cotton fiber development images. We thank Dr. L. Curtis Hannah from University of Florida, Dr. Chunda Feng and Dr. Abdu Abdelraheem from USDA-ARS, Stoneville, MS for critical review of the manuscript and their comments. We thank Dr. Jeff Gustin from USDA-ARS, Urbana, IL and Dr. Candace Haigler for their expert discussions for parts of the manuscript. The findings and conclusions in this publication are those of the authors and should not be construed to represent any official USDA or U. S. Government determination or policy.
Conflicts of Interest
The authors declare no conflict of interest.
EEO/Non-Discrimination
USDA is an equal opportunity provider and employer.
References
- Wendel, J.F.; Grover, C.E. Taxonomy and evolution of the cotton genus, Gossypium. Cotton 2015, 57, 25–44. [Google Scholar]
- Chen, Z.J.; et al. Toward sequencing cotton (Gossypium) genomes. Plant Physiol 2007, 145, 1303–1310. [Google Scholar] [CrossRef] [PubMed]
- Constable, G.; et al. Cotton breeding for fiber quality improvement. Industrial crops: Breeding for bioenergy and bioproducts 2015, 191–232. [Google Scholar]
- Basra, A.S.; C.P. Malik, Development of the Cotton Fiber, in International Review of Cytology, G.H. Bourne, J.F. Danielli, and K.W. Jeon, Editors. 1984, Academic Press. p. 65-113.
- Haigler, C.H.; et al. Cotton fiber: a powerful single-cell model for cell wall and cellulose research. Frontiers in Plant Science 2012, 3. [Google Scholar] [CrossRef]
- Stiff, M.R.; Haigler, C.H. Recent advances in cotton fiber development. Flowering and fruiting in cotton. Tennessee: The Cotton Foundation 2012, 163–192. [Google Scholar]
- Fan, S.; et al. Evolution of pectin synthesis relevant galacturonosyltransferase gene family and its expression during cotton fiber development. Journal of Cotton Research 2021, 4, NA. [Google Scholar] [CrossRef]
- Taliercio, E.W.; Boykin, D. Analysis of gene expression in cotton fiber initials. BMC Plant Biol 2007, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; et al. Comprehensive analysis of cellulose content, crystallinity, and lateral packing in Gossypium hirsutum and Gossypium barbadense cotton fibers using sum frequency generation, infrared and Raman spectroscopy, and X-ray diffraction. Cellulose 2015, 22. [Google Scholar] [CrossRef]
- Kabir, N.; et al. Functional characterization of TBL genes revealed the role of GhTBL7 and GhTBL58 in cotton fiber elongation. Int J Biol Macromol 2023, 241, 124571. [Google Scholar] [CrossRef]
- Wu, C.; et al. Genome-wide analysis elucidates the roles of GhHMA genes in different abiotic stresses and fiber development in upland cotton. Plant Physiology and Biochemistry 2023, 194, 281–301. [Google Scholar] [CrossRef]
- Fu, G.; et al. Genome-wide analysis of the serine carboxypeptidase-like protein family reveals Ga09G1039 is involved in fiber elongation in cotton. Plant Physiology and Biochemistry 2023, 107759. [Google Scholar] [CrossRef] [PubMed]
- Xing, K.; et al. N(6) -Methyladenosine mRNA modification regulates transcripts stability associated with cotton fiber elongation. Plant J 2023, 115, 967–985. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; et al. Comparative phosphoproteomic analysis reveals that phosphorylation of sucrose synthase GhSUS2 by Ca2+-dependent protein kinases GhCPK84/93 affects cotton fiber development. Journal of Experimental Botany 2023, 74, 1836–1852. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; et al. Large-fragment insertion activates gene GaFZ (Ga08G0121) and is associated with the fuzz and trichome reduction in cotton (Gossypium arboreum). Plant Biotechnology Journal 2021, 19, 1110–1124. [Google Scholar] [CrossRef] [PubMed]
- Zuo, D.; et al. Genome-wide identification of cotton CrRLK1L family genes and potential function of GhCrRLK1L104 in cell elongation. Current Plant Biology 2024, 37, 100325. [Google Scholar] [CrossRef]
- Yu, K.; et al. Potassium ameliorates cotton (Gossypium hirsutum L.) fiber length by regulating osmotic and K+/Na+ homeostasis under salt stress. Physiologia Plantarum 2023, 175, e13842. [Google Scholar] [CrossRef] [PubMed]
- Sun, M., C. Zheng, and W. Feng, Low soil available phosphorus level reduces cotton fiber length via osmoregulation. Frontiers in Plant Science 2023, 14, 1254103. [CrossRef]
- Fang, S.; et al. A cell wall-localized beta-1,3-glucanase promotes fiber cell elongation and secondary cell wall deposition. Plant Physiol 2023, 194, 106–123. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; et al. GhIQD10 interacts with GhCaM7 to control cotton fiber elongation via calcium signaling. The Crop Journal 2023, 11, 447–456. [Google Scholar] [CrossRef]
- Davière, J.M.; P. Achard, A Pivotal Role of DELLAs in Regulating Multiple Hormone Signals. Molecular Plant 2016, 9, 10–20. [CrossRef]
- Wen, Y.-z.; et al. Strigolactones modulate cotton fiber elongation and secondary cell wall thickening. Journal of Integrative Agriculture. 2023. [Google Scholar]
- Zeng, J.; et al. Fiber-specific increase of carotenoid content promotes cotton fiber elongation by increasing abscisic acid and ethylene biosynthesis. The Crop Journal. 2023. [Google Scholar]
- Stewart, J.M. Fiber initiation on the cotton ovule (Gossypium hirsutum). American Journal of Botany 1975, 62, 723–730. [Google Scholar] [CrossRef]
- Wu, Y.; et al. Expression profiling identifies genes expressed early during lint fibre initiation in cotton. Plant Cell Physiol 2006, 47, 107–127. [Google Scholar] [CrossRef] [PubMed]
- Kohel, R.J., E.V. Narbuth, and C.R. Benedict, Fiber Development of Ligon Lintless-2 Mutant of Cotton. Crop science 1992, 32, 733–735. [CrossRef]
- Du, X.M.; et al. Genetic analysis of presence and absence of lint and fuzz in cotton. Plant Breeding 2001, 120, 519–522. [Google Scholar] [CrossRef]
- Karaca, M.; et al. Simple sequence repeat (SSR) markers linked to the Ligon lintless (Li 1) mutant in cotton. Journal of Heredity 2002, 93, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Turley, R.B.; R.H. Kloth, The inheritance model for the fiberless trait in upland cotton (Gossypium hirsutum L.) line SL1-7-1: variation on a theme. Euphytica 2008, 164, 123–132. [CrossRef]
- Bechere, E., D.L. Auld, and E. Hequet, Development of ‘naked-tufted’ seed coat mutants for potential use in cotton production. Euphytica 2009, 167, 333–339. [CrossRef]
- Patel, J.D.; et al. The Ligon lintless-2 short fiber mutation is located within a terminal deletion of chromosome 18 in cotton. Plant Physiology 2020, 183, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Naoumkina, M.; et al. A deletion/duplication in the Ligon lintless-2 locus induces siRNAs that inhibit cotton fiber cell elongation. Plant Physiology 2022, 190, 1792–1805. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; et al. A Novel Tandem Zinc Finger Protein in Gossypium hirsutum, GhTZF2, Interacts with GhMORF8 to Regulate Cotton Fiber Cell Development. Agronomy 2023, 13, 519. [Google Scholar] [CrossRef]
- Wan, Q.; et al. Small interfering RNAs from bidirectional transcripts of GhMML3_A12 regulate cotton fiber development. New Phytol 2016, 210, 1298–1310. [Google Scholar] [CrossRef] [PubMed]
- Song, Q.; et al. Cotton microtubule-associated protein GhMAP20L5 mediates fiber elongation through the interaction with the tubulin GhTUB13. Plant Science 2023, 327, 111545. [Google Scholar] [CrossRef] [PubMed]
- Suo, J.; et al. Identification of GhMYB109 encoding a R2R3 MYB transcription factor that expressed specifically in fiber initials and elongating fibers of cotton (Gossypium hirsutum L.). Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression 2003, 1630, 25–34. [Google Scholar] [CrossRef]
- Lee, J.J.; et al. Developmental and gene expression analyses of a cotton naked seed mutant. Planta 2006, 223, 418–432. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; et al. Laser capture microdissection and cDNA microarrays used to generate gene expression profiles of the rapidly expanding fibre initial cells on the surface of cotton ovules. Planta 2007, 226, 1475–1490. [Google Scholar] [CrossRef] [PubMed]
- Walford, S.A.; et al. GhMYB25-like: a key factor in early cotton fibre development. Plant J 2011, 65, 785–797. [Google Scholar] [CrossRef] [PubMed]
- Pu, L.; et al. The R2R3 MYB transcription factor GhMYB109 is required for cotton fiber development. Genetics 2008, 180, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Liu, B., Y. Zhu, and T. Zhang, The R3-MYB gene GhCPC negatively regulates cotton fiber elongation. PLoS One 2015, 10, e0116272.
- Guan, X.; et al. Activation of Arabidopsis seed hair development by cotton fiber-related genes. PLoS One 2011, 6, e21301. [Google Scholar] [CrossRef]
- Wang, N.; et al. Genetic variation in MYB5_A12 is associated with fibre initiation and elongation in tetraploid cotton. Plant Biotechnology Journal 2021, 19, 1892. [Google Scholar] [CrossRef]
- Wu, A.; et al. GhMYB30-GhMUR3 affects fiber elongation and secondary wall thickening in cotton. Plant J 2024, 117, 694–712. [Google Scholar] [CrossRef] [PubMed]
- Ruan, J.; et al. Jasmonic Acid Signaling Pathway in Plants. Int J Mol Sci 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; et al. Cotton fiber elongation requires the transcription factor Gh MYB 212 to regulate sucrose transportation into expanding fibers. New Phytologist 2019, 222, 864–881. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; et al. Recent Advances and Future Perspectives in Cotton Research. Annu Rev Plant Biol 2021, 72, 437–462. [Google Scholar] [CrossRef]
- Wu, H.; et al. Genetics and evolution of MIXTA genes regulating cotton lint fiber development. New Phytol 2018, 217, 883–895. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.H.; et al. Genetic dissection of the fuzzless seed trait in. Journal of Experimental Botany 2018, 69, 997–1009. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; et al. Genome-wide investigation and transcriptome analysis of the WRKY gene family in Gossypium. Mol Genet Genomics 2015, 290, 151–171. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; et al. Identification of the Group III WRKY Subfamily and the Functional Analysis of GhWRKY53 in Gossypium hirsutum L. Plants 2021, 10, 1235. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.-N.; et al. Phosphorylation of WRKY16 by MPK3-1 is essential for its transcriptional activity during fiber initiation and elongation in cotton (Gossypium hirsutum). The Plant Cell 2021, 33, 2736–2752. [Google Scholar] [CrossRef]
- Chen, E.; et al. Genome-wide analysis of the HD-ZIP IV transcription factor family in Gossypium arboreum and GaHDG11 involved in osmotic tolerance in transgenic Arabidopsis. Mol Genet Genomics 2017, 292, 593–609. [Google Scholar] [CrossRef]
- Guan, X.Y.; et al. The HD-Zip IV gene GaHOX1 from cotton is a functional homologue of the Arabidopsis GLABRA2. Physiologia plantarum 2008, 134, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.M.; et al. Control of cotton fibre elongation by a homeodomain transcription factor GhHOX3. Nat Commun 2014, 5, 5519. [Google Scholar] [CrossRef] [PubMed]
- Walford, S.A.; et al. Epidermal cell differentiation in cotton mediated by the homeodomain leucine zipper gene, GhHD-1. Plant J 2012, 71, 464–478. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; et al. Phosphatidic acid interacts with an HD-ZIP transcription factor GhHOX4 to influence its function in fiber elongation of cotton (Gossypium hirsutum). Plant J 2024. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, X.X.; et al. Functional characterization of a basic helix-loop-helix (bHLH) transcription factor GhDEL65 from cotton (Gossypium hirsutum). Physiol Plant 2016, 158, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; et al. Core cis-element variation confers subgenome-biased expression of a transcription factor that functions in cotton fiber elongation. New Phytologist 2018, 218, 1061–1075. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; et al. Characterization of bHLH/HLH genes that are involved in brassinosteroid (BR) signaling in fiber development of cotton (Gossypium hirsutum). BMC plant biology 2018, 18, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lu, R.; et al. The bHLH/HLH transcription factors GhFP2 and GhACE1 antagonistically regulate fiber elongation in cotton. Plant Physiology 2022, 189, 628–643. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.F.; et al. The miR319-Targeted GhTCP4 Promotes the Transition from Cell Elongation to Wall Thickening in Cotton Fiber. Mol Plant 2020, 13, 1063–1077. [Google Scholar] [CrossRef]
- Tian, Y.; et al. The transcription factor MML4_D12 regulates fiber development through interplay with the WD40-repeat protein WDR in cotton. Journal of Experimental Botany 2020, 71, 3499–3511. [Google Scholar] [CrossRef]
- Wang, Y.; et al. GhKNL1 controls fiber elongation and secondary cell wall synthesis by repressing its downstream genes in cotton (Gossypium hirsutum). J Integr Plant Biol 2022, 64, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; et al. Zinc Finger Protein8 (GhZFP8) Regulates the Initiation of Trichomes in Arabidopsis and the Development of Fiber in Cotton. Plants 2024, 13, 492. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.-W.; et al. Comparative proteomic analysis provides new insights into the fiber elongating process in cotton. Journal of Proteome Research 2008, 7, 4623–4637. [Google Scholar] [CrossRef]
- Ho, M.H.; et al. Characterization and promoter analysis of a cotton RING-type ubiquitin ligase (E3) gene. Mol Biotechnol 2010, 46, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Salih, H.; et al. Characterization of the early fiber development gene, Ligon-lintless 1 (Li1), using microarray. Plant Gene 2016, 6, 59–66. [Google Scholar] [CrossRef]
- Prasad, P.; et al. Transcriptional Landscape of Cotton Fiber Development and Its Alliance With Fiber-Associated Traits. Front Plant Sci 2022, 13, 811655. [Google Scholar] [CrossRef]
- Wang, D.; et al. Cell-specific clock-controlled gene expression program regulates rhythmic fiber cell growth in cotton. Genome Biol 2023, 24, 49. [Google Scholar] [CrossRef]
- Bao, Y.; et al. Genome-wide chromatin accessibility landscape and dynamics of transcription factor networks during ovule and fiber development in cotton. BMC Biol 2023, 21, 165. [Google Scholar] [CrossRef] [PubMed]
- Beasley, C.A.; I.P. Ting, Effects of plant growth substances on in vitro fiber development from unfertilized cotton ovules. American Journal of Botany 1974, 61, 188–194. [CrossRef]
- Guinn, G.; D.L. Brummett, Changes in abscisic acid and indoleacetic acid before and after anthesis relative to changes in abscission rates of cotton fruiting forms. Plant physiology 1988, 87, 629–631. [CrossRef]
- Chen, Z.J.; X. Guan, Auxin boost for cotton. Nat Biotechnol 2011, 29, 407–409. [CrossRef] [PubMed]
- Zhang, M.; et al. Spatiotemporal manipulation of auxin biosynthesis in cotton ovule epidermal cells enhances fiber yield and quality. Nat Biotechnol 2011, 29, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; et al. Auxin Regulates Cotton Fiber Initiation via GhPIN-Mediated Auxin Transport. Plant Cell Physiol 2017, 58, 385–397. [Google Scholar] [CrossRef] [PubMed]
- Gokani, S.J.; V.S. Thaker, Role of gibberellic acid in cotton fibre development. The Journal of Agricultural Science 2002, 138, 255–260. [CrossRef]
- Xiao, G.; et al. Genome-wide identification of the GhARF gene family reveals that GhARF2 and GhARF18 are involved in cotton fibre cell initiation. Journal of experimental botany 2018, 69, 4323–4337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; et al. Characterization of cotton ARF factors and the role of GhARF2b in fiber development. BMC Genomics 2021, 22, 202. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; et al. Cloning and expression analysis of novel Aux/IAA family genes in Gossypium hirsutum. Gene 2012, 503, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-G.; et al. ABP1 is required for organized cell elongation and division in Arabidopsis embryogenesis. Genes & development 2001, 15, 902–911. [Google Scholar]
- Tao, L.Z., A.Y. Cheung, and H.M. Wu, Plant Rac-like GTPases are activated by auxin and mediate auxin-responsive gene expression. Plant Cell 2002, 14, 2745–2760. [CrossRef]
- Wang, M.Y.; et al. The cotton transcription factor TCP14 functions in auxin-mediated epidermal cell differentiation and elongation. Plant Physiol 2013, 162, 1669–1680. [Google Scholar] [CrossRef] [PubMed]
- Li, I.X.; et al. [Cloning and expression analysis of two Rac genes from cotton (Gossypium hirsutum L)]. Yi Chuan Xue Bao 2005, 32, 72–78. [Google Scholar]
- Zhang, M.; et al. PIN-formed protein, a door to reveal the mechanism for auxin-triggered initiation of cotton fiber. Plant Signal Behav 2017, 12, e1319031. [Google Scholar] [CrossRef] [PubMed]
- Xi, J.; et al. GhROP6 GTPase modulates auxin accumulation in cotton fibers by regulating cell-specific GhPIN3a localization. J Exp Bot 2023, 74, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; B.A. Triplett, Cotton fiber growth in planta and in vitro. Models for plant cell elongation and cell wall biogenesis. Plant Physiol 2001, 127, 1361–1366. [CrossRef]
- Gialvalis, S.; R.W. Seagull, Plant hormones alter fiber initiation in unfertilized, cultured ovules of Gossypium hirsutum. J. Cotton Sci 2001, 5, 252–258.
- Seagull, R.W.; S. Giavalis, Pre-and post-anthesis application of exogenous hormones alters fiber production in Gossypium hirsutum L. cultivar Maxxa GTO. 2004.
- Liao, W.-b.; et al. Isolation and characterization of a GAI/RGA-like gene from Gossypium hirsutum. Plant growth regulation 2009, 58, 35–45. [Google Scholar] [CrossRef]
- Aleman, L.; et al. Functional analysis of cotton orthologs of GA signal transduction factors GID1 and SLR1. Plant Mol Biol 2008, 68, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.H.; et al. Gibberellin 20-oxidase promotes initiation and elongation of cotton fibers by regulating gibberellin synthesis. J Plant Physiol 2010, 167, 829–837. [Google Scholar] [CrossRef]
- Bai, W.Q.; et al. Gibberellin overproduction promotes sucrose synthase expression and secondary cell wall deposition in cotton fibers. PLoS One 2014, 9, e96537. [Google Scholar] [CrossRef]
- Daviere, J.M.; P. Achard, Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [CrossRef]
- Hu, M.-y.; et al. Brassinosteroids and auxin down-regulate DELLA genes in fiber initiation and elongation of cotton. Agricultural Sciences in China 2011, 10, 1168–1176. [Google Scholar] [CrossRef]
- Sun, T.P. Gibberellin metabolism, perception and signaling pathways in Arabidopsis. Arabidopsis Book 2008, 6, e0103. [Google Scholar] [CrossRef]
- Jing, D.; et al. Cloning and Expression Profile of Gibberellin Insensitive Dwarf GID1 Homologous Genes from Cotton. Acta Agronomica Sinica 2009, 35, 1822–1830. [Google Scholar]
- He, P.; et al. Gibberellin acid promotes single-celled fiber elongation through the activation of two signaling cascades in cotton. Developmental Cell, 2024.
- Sun, Y.; et al. Brassinosteroid regulates fiber development on cultured cotton ovules. Plant Cell Physiol 2005, 46, 1384–1391. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; et al. GhDET2, a steroid 5alpha-reductase, plays an important role in cotton fiber cell initiation and elongation. Plant J 2007, 51, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.H.; et al. Transcriptome profiling, molecular biological, and physiological studies reveal a major role for ethylene in cotton fiber cell elongation. Plant Cell 2006, 18, 651–664. [Google Scholar] [CrossRef]
- Yang, Z.; et al. PAG1, a cotton brassinosteroid catabolism gene, modulates fiber elongation. New Phytol 2014, 203, 437–448. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; et al. Cotton (Gossypium hirsutum) 14-3-3 proteins participate in regulation of fibre initiation and elongation by modulating brassinosteroid signalling. Plant Biotechnol J 2015, 13, 269–280. [Google Scholar] [CrossRef]
- Liu, Z.H.; et al. A basic helix-loop-helix protein (GhFP1) promotes fibre elongation of cotton (Gossypium hirsutum) by modulating brassinosteroid biosynthesis and signalling. New Phytol 2020, 225, 2439–2452. [Google Scholar] [CrossRef]
- Liu, L.; et al. A brassinosteroid transcriptional regulatory network participates in regulating fiber elongation in cotton. Plant Physiol 2023, 191, 1985–2000. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; et al. Brassinosteroids regulate cotton fiber elongation by modulating very-long-chain fatty acid biosynthesis. Plant Cell 2023, 35, 2114–2131. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; et al. Exogenous jasmonic acid inhibits cotton fiber elongation. Journal of Plant Growth Regulation 2012, 31, 599–605. [Google Scholar] [CrossRef]
- Wang, L.; et al. Comparative Transcriptomics Reveals Jasmonic Acid-Associated Metabolism Related to Cotton Fiber Initiation. PLoS One 2015, 10, e0129854. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; et al. GhJAZ2 negatively regulates cotton fiber initiation by interacting with the R2R3-MYB transcription factor GhMYB25-like. Plant J 2016, 88, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.-M., C.-Y. Hu, and Y.-X. Zhu, The ascorbate peroxidase regulated by H2O2 and ethylene is involved in cotton fiber cell elongation by modulating ROS homeostasis. Plant signaling & behavior 2008, 3, 194–196.
- Jiang, L.; et al. A role for APX1 gene in lead tolerance in Arabidopsis thaliana. Plant Sci 2017, 256, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; et al. A cotton ascorbate peroxidase is involved in hydrogen peroxide homeostasis during fibre cell development. New Phytologist 2007, 175, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.-M.; Y.-X. Zhu, How cotton fibers elongate: a tale of linear cell-growth mode. Current opinion in plant biology 2011, 14, 106–111. [CrossRef] [PubMed]
- Tang, W.; et al. The calcium sensor GhCaM7 promotes cotton fiber elongation by modulating reactive oxygen species (ROS) production. New Phytol 2014, 202, 509–520. [Google Scholar] [CrossRef]
- Qin, Y.M.; et al. Saturated very-long-chain fatty acids promote cotton fiber and Arabidopsis cell elongation by activating ethylene biosynthesis. Plant Cell 2007, 19, 3692–3704. [Google Scholar] [CrossRef] [PubMed]
- Beasley, C.A.; et al. A quantitative procedure for estimating cotton fiber growth. Stain Technol 1974, 49, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; et al. Comparative proteomic analysis identified proteins and the phenylpropanoid biosynthesis pathway involved in the response to ABA treatment in cotton fiber development. Sci Rep 2023, 13, 1488. [Google Scholar] [CrossRef] [PubMed]
- Davis, L.A.; F.T. Addicott, Abscisic Acid: correlations with abscission and with development in the cotton fruit. Plant Physiol 1972, 49, 644–648. [CrossRef] [PubMed]
- Gokani, S.J., R. Kumar, and V.S. Thaker, Potential role of abscisic acid in cotton fiber and ovule development. Journal of plant growth regulation 1998, 17, 1–5. [CrossRef]
- Nayyar, H.; et al. Hormonal regulation of cotton fibre elongation in Gossypium arboreum L. in vitro and in vivo. Biochemie und Physiologie der Pflanzen 1989, 185, 415–421. [Google Scholar] [CrossRef]
- Ma, Q., P. Hedden, and Q. Zhang, Heterosis in rice seedlings: its relationship to gibberellin content and expression of gibberellin metabolism and signaling genes. Plant Physiol 2011, 156, 1905–1920. [CrossRef] [PubMed]
- Gilbert, M.K.; et al. A transcript profiling approach reveals an abscisic acid-specific glycosyltransferase (UGT73C14) induced in developing fiber of Ligon lintless-2 mutant of cotton (Gossypium hirsutum L). PLoS One 2013, 8, e75268. [Google Scholar] [CrossRef]
- Zhang, H.; et al. Effect of phytohormones on fiber initiation of cotton ovule. Acta physiologiae plantarum 2009, 31, 979–986. [Google Scholar] [CrossRef]
- Dasani, S.H.; V.S. Thaker, Role of abscisic acid in cotton fiber development. Russian Journal of Plant Physiology 2006, 53, 62–67. [CrossRef]
- Chen, J.G.; et al. Levels of cytokinins in the ovules of cotton mutants with altered fiber development. Journal of Plant Growth Regulation 1997, 16, 181–185. [Google Scholar] [CrossRef]
- Spallek, T.; et al. Interspecies hormonal control of host root morphology by parasitic plants. Proc Natl Acad Sci U S A 2017, 114, 5283–5288. [Google Scholar] [CrossRef] [PubMed]
- Niemann, M.C.E.; et al. The cytokinin oxidase/dehydrogenase CKX1 is a membrane-bound protein requiring homooligomerization in the endoplasmic reticulum for its cellular activity. Plant Physiology 2018, 176, 2024–2039. [Google Scholar] [CrossRef] [PubMed]
- Jones, R.J.; B.M.N. Schreiber, Role and function of cytokinin oxidase in plants. Plant Growth Regulation 1997, 23, 123–134. [CrossRef]
Figure 1.
Cotton fiber development. (A) Progression of cotton flower and boll development. Fiber developments are classified into five stages including initiation, elongation, transition, secondary cell wall (SCW) biosynthesis and maturation. DPA, days post-anthesis. Adapted with permission from Fang et al. 2018, Crop Science 58: 2214-2228, (B) Images of fibers at progressive developmental stages: SEM of fiber initials (bar = 10 µm) and fibers during early elongation (bar=100 µm); TEM of the first stage of fiber thickening during the transition stage (three adjacent fibers are shown; bar=300 nm) and near the end of secondary cell wall synthesis (bar=1 µm); and light micrograph of cross-sectioned mature, dried, fibers. Images excerpted with permission from Figure 1 in Stiff and Haigler, 2012, https://www.cotton.org/foundation/upload/F-F-Chapter-10.pdf.
Figure 1.
Cotton fiber development. (A) Progression of cotton flower and boll development. Fiber developments are classified into five stages including initiation, elongation, transition, secondary cell wall (SCW) biosynthesis and maturation. DPA, days post-anthesis. Adapted with permission from Fang et al. 2018, Crop Science 58: 2214-2228, (B) Images of fibers at progressive developmental stages: SEM of fiber initials (bar = 10 µm) and fibers during early elongation (bar=100 µm); TEM of the first stage of fiber thickening during the transition stage (three adjacent fibers are shown; bar=300 nm) and near the end of secondary cell wall synthesis (bar=1 µm); and light micrograph of cross-sectioned mature, dried, fibers. Images excerpted with permission from Figure 1 in Stiff and Haigler, 2012, https://www.cotton.org/foundation/upload/F-F-Chapter-10.pdf.

Figure 2.
Key transcription factors that regulate fiber initiation (A) and elongation (B). The light blue color and arrows indicate the positive regulation, and the light grey color and bars represent the negative regulation.
Figure 2.
Key transcription factors that regulate fiber initiation (A) and elongation (B). The light blue color and arrows indicate the positive regulation, and the light grey color and bars represent the negative regulation.
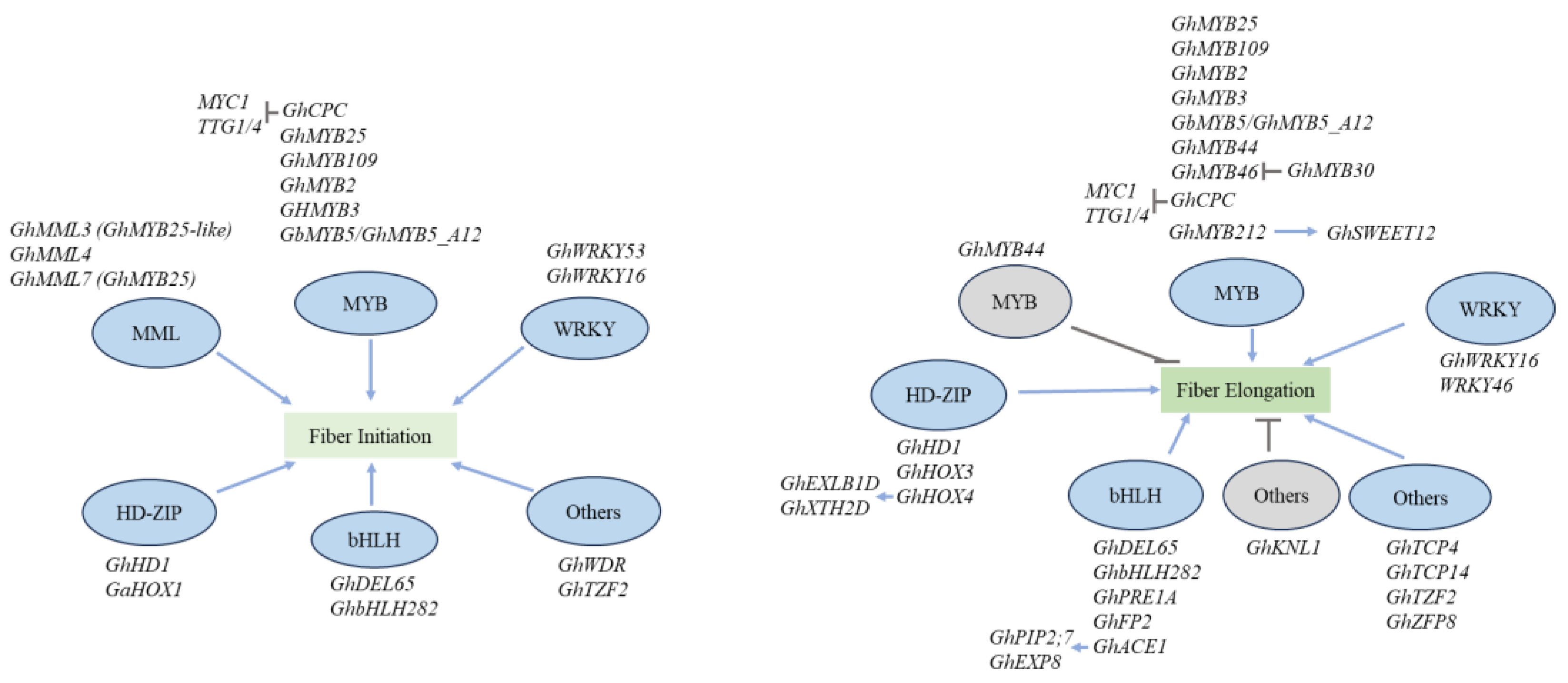
Figure 3.
Phytohormones regulation of fiber initiation (A) and elongation (B). The light blue color and arrows indicate the positive regulation, and the light grey color and bars represent the negative regulation. GA, gibberellic acid; BR, brassinosteroids, JA, jasmonic acid; ABA, abscisic acid. ABA and cytokinin both have negative regulation on fiber initiation and elongation. Cytokinin promotes the growth of ovule.
Figure 3.
Phytohormones regulation of fiber initiation (A) and elongation (B). The light blue color and arrows indicate the positive regulation, and the light grey color and bars represent the negative regulation. GA, gibberellic acid; BR, brassinosteroids, JA, jasmonic acid; ABA, abscisic acid. ABA and cytokinin both have negative regulation on fiber initiation and elongation. Cytokinin promotes the growth of ovule.
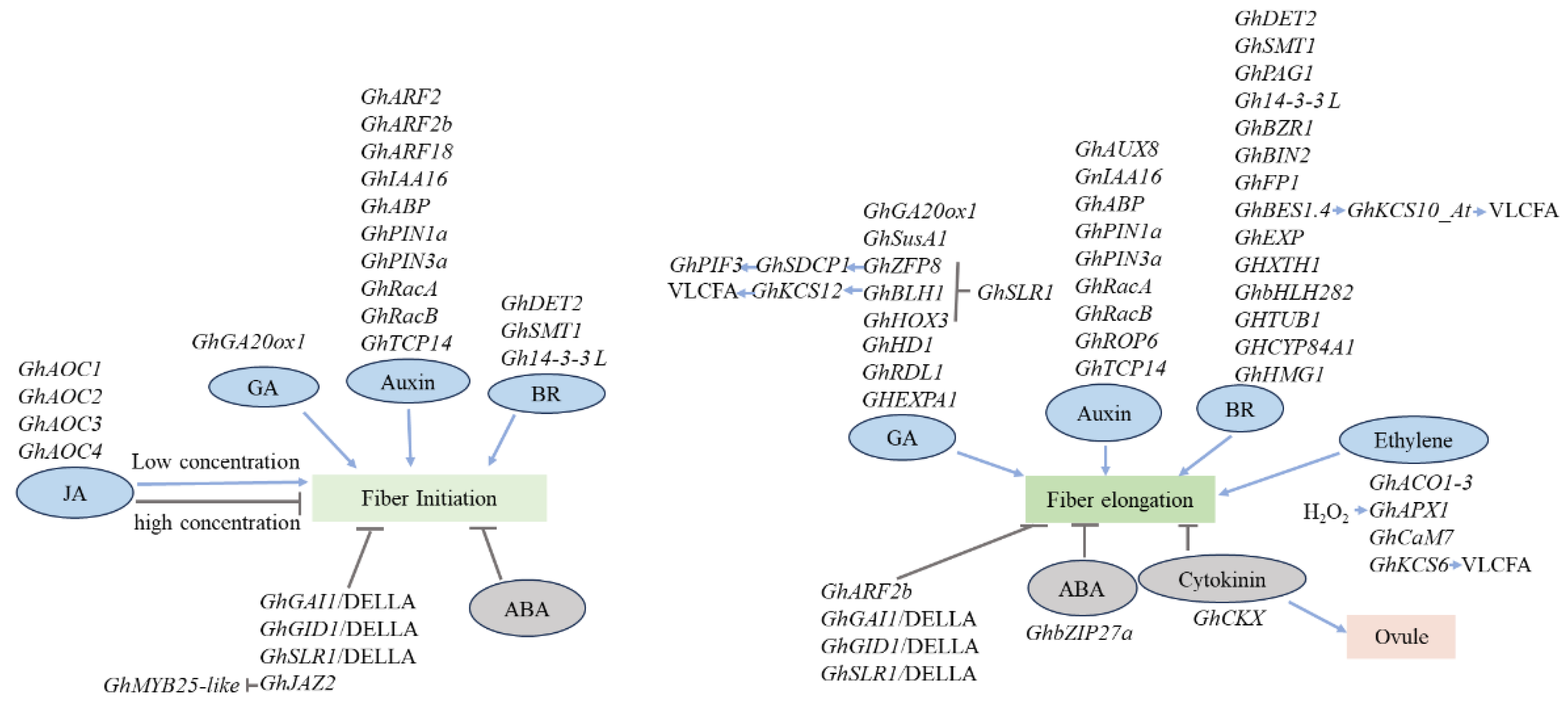
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



