Submitted:
11 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
Kombucha is a two-stage fermented sweetened-tea beverage that uses yeast and lactic acid bacteria (LAB) to convert sugars into ethanol and lactate, and acetic acid bacteria (AAB) to oxidize ethanol to acetate. Its popularity as a beverage grew from claims of health-benefits derived from this vibrant microbial bioconversion. While recent studies have shed light on the diversity of cultures during a Kombucha fermentation, there is limited information on the diversity, and especially viability, of cultures in retail beverages that advertise the presence of Kombucha and probiotic cultures. In this study, 12 Kombucha beverages produced by different manufacturers throughout the US were purchased and microbially characterized. Eight of the beverages contained viable Kombucha cultures, while 3 of the remaining 4 had viable Bacillus cultures as added probiotics. Amplicon profiling revealed all contained Kombucha yeast and bacteria cells. The dominant yeast detected were Lachancea cidri (10/12), Brettanomyces (9/12), Malassezia (6/12) and Saccharomyces (5/12). Dominant LAB included Liquorilactobacillus and Oenococcus oeni, and AAB were Komagataeibacter, Gluconobacter and Acetobacter. One beverage had a significant amount of Zymomonas mobilis, an ethanol producing bacterium from Agave cactus. While Kombucha beverages differ in the types and viability of cultures, all except one beverage contained detectable viable cells.
Keywords:
Fermentation
; Probiotic
; Cultures
; Amplicon profiling
; Viable plate count
1. Introduction
Kombucha is a fermented tea beverage composed of a SCOBY (symbiotic relationship of bacteria and yeast), water, tea, and sugar. It is reported to be originally developed during the Chinese Tsin Dynasty (249 – 206 BC), where it was thought to be associated with health and longevity [1]. While the precise country of origin is unknown, its current name comes from the Japanese word ‘Kombu’ that referred to Luminaria japonica which was reported to be an edible fungus, but is actually a marine alga [2]. It was originally referred to as ‘tea fungus’, due to the surface cellulose based pellicle that forms during production resembling a fungal mat [3]. This traditional fermented beverage has also been referred to as Teekvass, Cajnij, Hongo, Teeschwamm, and Wunderpilz in different countries worldwide [4]. The historical association of its nutritional benefits precipitated its subsequent introduction into Japan, Korea and Eastern Europe, where it gained many legendary health attributes over the centuries [5]. While reports on the global history of Kombucha are not consistent throughout, a common health benefit theme prevails. More recently, it has become popular in many Western countries and has consistently gained popularity in the US market over the last two plus decades. Its market share in the US reached 1 billion dollars in 2021 and is projected to continue growing by 16.2% annually for the next decade [6].
The potential health benefits that give Kombucha its popularity come from the presence of viable cultures of fermentative bacteria and yeast as well as bioactive components such as phenolic compounds and organic acids formed during the fermentation [7]. Like other traditional fermented foods, such as yogurt, kefir and kimchi, it is viewed as a probiotic food due to the presence of viable cultures [8]. However, Kombucha has not been researched as much as other fermented foods as to if and how the live and active cultures in Kombucha play a role in health benefits. However, the presence of lactic acid bacteria (LAB) has been reported in Kombucha, substantiating its probiotic potential [9,10]. Studies examining the diversity of microbes during Kombucha production have occurred, but few studies have looked at the viability and diversity of cultures in retail Kombucha drinks that are consumed by the public. Recently, six different retail Kombuchas in New Zealand were analyzed and found only three contained viable cultures [11]. Studies have reported on inconsistencies in product labels of retail Kombucha drinks regarding the term ‘probiotic’ and actual viable cultures present, indicating a growing need to further standardize labeling to reflect the probiotic potential of available Kombucha products more accurately [12].
Early microbial studies with Kombucha found that a defined culture of two yeasts and one acetic acid bacterium (AAB) was minimally required for a successful Kombucha fermentation [13]. Typically, there are a lot more fermentative bacteria and yeast within Kombucha drinks, which likely provide additional potential health benefits and flavors. These include genera of AAB (Komagataeibacter, Gluconobacter and Acetobacter) [14], LAB (Lactobacillus, Oenococcus, and Lactococcus) [10], and yeasts (Saccharomyces, Zygosaccharomyces, Torulaspora and Brettanomyces) [9]. Defined probiotic cultures are sometimes added in commercial Kombuchas to potentially enhance the nutraceutical aspect of the beverage [12].
The health benefits attributed to Kombucha are impressive, ranging from prevention of different diseases to enhancing mental attributes [15]. However human clinical trials are currently very limited and are needed to substantiate the potential health claims from cell culture and animal studies [15,16]. Many studies suggest Kombucha may offer many holistic benefits regarding colon cancer, diabetes, and inflammatory improvements. For example, an invitro study by Rasouli et al., (2021) using a cancer cell line HCT-116 compared the use of Kombucha to the anticancer drug doxorubicin for the treatment of colon cancer [17]. They found that Kombucha was able to increase the expression of a variety of genes to overall help slow down and stop the growth of cancer cells more effectively than doxorubicin. Another invitro study by Xu et al., (2022) used a mouse model to explore the mechanism of how Kombucha could be used to protect against diabetes [18]. They showed a decrease in insulin resistance and improved beta cell function, thus improving diabetes symptoms in mice. Perhaps the most surprising but strongest potential health benefits of Kombucha is its strong anti-inflammatory effects including from lipopolysaccharide (LPS) challenges. Using a mouse model Wang et al., (2021) showed a strong correlation between Kombucha feeding and reduced tumor necrosis factor alpha (TNF-α) levels, which is a potent inflammatory cytokine specifically induced by LPS [19]. The study also found a reduction in inflammatory interleukins (IL-1ß and IL-6) and a significant reduction in LPS induced sepsis in this mouse study. Given that Kombucha fermentation involves high levels of gram negative bacteria, which are a source of dietary LPS, this anti-inflammatory effect is very promising. Recent studies examining the ability of dietary Kombucha to modulate the gut microbiota provide a likely mechanism for the observed anti-inflammatory effects that contribute to the potential health benefits [20].
Studies in recent years looking at the diversity of microorganisms during Kombucha fermentations have primarily utilized non-culturing molecular approaches, using either metagenomic or meta-amplicon sequencing. Arikan et al., (2020) used 16S rRNA amplicon sequencing to reveal Komagataeibacter as the most prominent bacterial genus from two homemade Kombuchas produced in Turkey [21]. This AAB genus was first described in 2012 and is most closely related to the genus Gluconacetobacter [22]. Another study utilized these approaches to examine the microbial diversity in nine retail Kombucha samples purchased in Los Angeles, CA, USA and found Komagataeibacter, Gluconacetobacter and Acetobacter were the dominant AAB present and Brettanomyces the dominant yeast [23]. While this latter study did not examine culture viability, it was assumed throughout that the cultures detected were viable. Given inactivated microbial cells can contain DNA for significant, but unknown lengths of time it is important to examine this further. Therefore, the objective of this current study was to combine culturing and non-culturing molecular techniques to examine both the viability and diversity in different Kombucha beverages available in retail stores in the US.
2. Materials and Methods
2.1. Selection of Kombucha Beverages
Twelve different Kombucha beverages from different manufacturers were purchased in various grocery stores within the Twin Cities (Minneapolis and St. Paul, MN). Table 1 details the 12 Kombuchas and their listed relevant ingredients and processes. The ‘best before’ dates on each beverage were all more than five months from the date of purchase. These represent all the major brands available in this market as well as the brands with the largest market share in the USA [23]. All samples were maintained at 4°C and used for culturing studies within two weeks of purchase.
2.2. Viable Culture Analysis
The following selective and non-selective media were used for quantitative culture analysis of the kombucha samples. BHI (Brain Heart Infusion) agar media was used as a general non-selective medium. MRS (de Man–Rogosa–Sharpe) medium was for lactic acid bacteria (LAB) and Bacillus probiotics. Acetic acid bacteria (AAB) selective medium (ABS) (Kim et al., 2019) was used for growth of AAB. Potato Dextrose Agar (PDA) supplemented with 25 mg/ml chloramphenicol was used for selective growth of yeast (PDA-Y). MacConkey agar (MAC) plates were used to test for the presence of any fecal coliform bacteria. Serial 1:10 dilutions of each Kombucha drink were generated using sterile peptone water up to 10−4 and spread plated onto each agar medium. All plates were incubated at 30◦C aerobically for both bacteria and yeast, and MAC plates were also incubated at 37◦C. After 48 hours, colony forming units (CFU) were calculated and representatives for different types of microbes were inoculated into BHI broth media for further analysis.
2.3. Analytical Techniques
Gram stains were conducted using freshly grown cultures for each of the different microorganisms representing different colony types from different agar media and viewed under a light microscope at 1000X magnification. All pH measurements of fresh Kombucha beverages were obtained with newly calibrated pH probes using a Mettler Toledo SevenEasy instrument (Columbus, OH).
2.4. Molecular Identification of Select Kombucha Colony Isolates
Colonies were inoculated into BHI broths and when fully grown, 1.5 ml of culture was pelleted using an Eppendorf centrifuge and the supernatant was completely removed. Pellets were then resuspended in 500 µl molecular grade water and 200 µl glass beads (< 106 µm diameter; Sigma, St Louis, MO) and cells were disrupted in a Minibeadbeater-8 (Biospec Products, Bartlesville, OK) for 20 seconds at maximum speed. Tubes were immediately placed on ice for debris to settle and 100 µl of the DNA enriched supernatant was transferred to a fresh tube and diluted 1:10 to provide the template for PCR.
The 16S rRNA gene was amplified using primers: 27F, 5′-AGAGTTTGATCMTGGCTCAG-3′ and 1525R, 5′-AAGGAGGTGATCCAGCC-3′ with Taq DNA polymerase in a reaction volume of 50µl using a Robocycler (Stratagene, La Jolla, CA) with the following conditions: 1 cycle of 92◦C for 2 min and 35 cycles of 92◦C X 1 min; 50◦C X 30 seconds and 72◦C X 1 min. One percent agarose gels in 1X TAE buffer (Bio-Rad, Hercules, CA) were used for electrophoresis of PCR samples. Bands were cut out and the DNA was gel extracted using a kit from New England BioLabs (Ipswich, MA) according to manufacturer’s instructions. Purified DNAs were quantified using a Nanodrop spectrophotometer (Thermo Scientific, Waltham, MA) and Sanger sequencing of amplicons was conducted at ACGT DNA Sequencing Services (Wheeling, IL).
2.5. Amplicon Profiling of Kombucha Beverages
Fresh beverages were initially mixed by shaking to ensure microbial cells were properly suspended. Five ml of samples were pelleted by centrifugation to pellet any microbial cells. Pellets were resuspended in 1 ml molecular grade water and pelleted by centrifugation to ensure removal of all liquid. Pellets were then resuspended in warm MBL buffer of the DNeasy PowerFood Microbial DNA isolation Kit (Qiagen, Redwood City, CA). The manufacturer’s instructions were followed for total DNA isolation except the disruption of cells using the PowerBead tubes was conducted with a Minibeadbeater-8 (Biospec Products, Bartlesville, OK) for 20 seconds at maximum speed. The V3-V4 region of the bacterial 16S rRNA gene and the ITS1 (region between the fungal 18S and 5.8S rRNA genes) were used for profiling the total bacterial and yeast diversity in each Kombucha. All amplifications and paired end sequencing reactions were conducted at the University of Minnesota Genomics Center using techniques previously described [24].
Four FASTQ raw sequence files were generated for each sample corresponding to the paired end reads for each amplicon. Trimming and merging of paired sequences was conducted using Geneious Prime® 2024.0.3 Software [25]. Sequences were further quality controlled using the BBDuk plugin from the Joint Genome Institute (Department of Energy) in this software using a minimum sequence quality score of 32. Following assembly of identical sequences into taxonomical groups species identifications were assigned to each group using BLAST database searches from the NCBI (National Center for Biotechnology Information). Relative abundances were assigned based on the number of sequence reads corresponding to each taxonomic grouping.
3. Results
3.1. Viable Culture Analysis of Kom 1 – Kom 12
Fermented foods have a long tradition of containing potentially healthy bioactive compounds produced from the fermentation as well as live microbial fermentative cultures. While fermented dairy beverages have a long history of containing live and active fermentative cultures, there is considerably less known about the viability of cultures available in commercially available Kombucha beverages. To investigate this, 12 Kombucha beverages were examined for relevant Kombucha cultures including yeast, AAB, LAB, and added probiotics (where relevant). Table 2 details the viable plate counts of the 12 Kombuchas on various media as well as the pH measurements of each. Kom 11 was the only product that contained no detectable viable cells on any plate. All other products contained viable cells at levels of 103 to 107 cfu/ml. However, 3 of these Kombucha’s (Kom 1, Kom 3 and Kom 6) did not have any viable Kombucha fermentative cultures (AAB, LAB or yeast) but did contain added Bacillus cultures as probiotics. Two of the products (Kom 2 and Kom 7) contained colonies on MacConkey agar plates, which were included as they are routinely utilized to test for fecal coliform bacteria in the food industry.
3.2. Microscopic Analysis of Different Cultures Isolated from the Selective Media
Selected colonies from different Kombuchas and different agar media were cultured in BHI broths for gram staining and microscopic analysis. This revealed that at least some yeast could grow on all agar media, including MacConkey agar. The two Kombucha samples that had growth on this medium (Kom 2 and Kom 7) both exhibited similar colony morphologies, suggesting a single type of yeast species. Microscopic analysis of these colonies also revealed similar morphologies substantiating this result (Figure 1A). In addition, MRS medium, which is selective for LAB also showed yeast growth as well as viable gram positive coccoid cells in chains, indicative of LAB (Figure 1B). This distinctive budding yeast morphology was also seen from different Kombucha’s as well as from colonies on the AAB selective medium, ACS. A short gram negative rod which came from a colony on the ACS plate from Kom 4 was used for PCR amplification of its 16S rRNA gene and sequence analysis of this gene indicated it was an AAB, Acetobacter tropicalis, which is consistent with its observed morphology. While most yeast morphologies observed microscopically were single cells, some elongated/filamentous cellular structures were also seen (Figure 1C).
3.3. Bacterial and Fungal Diversity Obtained from Direct Amplicon Profiling of Each Kombucha
Kombucha production involves both anaerobic and aerobic fermentation activities, whereby sugars are first fermented to ethanol, primarily by yeast, and to organic acids by LAB. Subsequently the AAB aerobically converts ethanol to acetic acid and therefore the functional Kombucha cultures are composed of both fungal (yeast) and bacterial components (LAB and AAB). While 4 of the 12 Kombucha’s did not have any detectable viable bacteria or yeast fermentative cultures, the amplicon profiling did reveal Kombucha cultures for all 12 of the samples, indicating that intact non-viable cells were present. The relative abundance of different amplicon sequences gave a good picture of the dominant types involved and some samples were much more diversely represented than others. It should be noted that diversity in a specific food fermentation is significantly lower than microbial diversity in a dynamic natural ecosystem, such as soil or feces and often may be dominated by one or more genera. Figure 2 compares the dominant fungal genera observed in the 12 Kombucha samples and illustrates this differential, where many samples are primarily dominated by one or two genera while others are less so. The Brettanomyces yeast genus was the most dominant one in 5 samples (Kom8 – Kom 12) and was also a significant member of 4 of the other 7 samples. Two species were represented, B. bruxellensis the most common, and B. anomalus (Table 3). The other most prominently represented dominant yeast genus was Lachancea, which was found in 10 of the 12 samples and represented the most dominant yeast genus in Kom 4 and Kom 7 (Figure 2). It was just represented by one species, L. cidri indicating the prevalence of this yeast species in Kombucha brewing in the USA currently. The prominent ethanol producing yeast genus, Saccharomyces was present in 5 samples, with S. cerevisiae the most common species and was one of the two most dominant yeast species in Kom 9 and Kom 12. The overall relative abundance in the 12 Kombuchas, using a minimum abundance of 1% per sample, revealed 24 different yeast/fungal species, representing 18 genera (Table 3).
Five of the 12 Kombucha beverages listed added ‘Bacillus’ probiotics as an ingredient. Kom 1, Kom 5 and Kom 6 listed B. coagulans, while Kom 2 and Kom 3 listed B. subtilis. In a recent reclassification of some Bacillus species, and species of other genera, B. coagulans was included in the new genus Heyndrickxia and thus is represented in the BLAST database under this name (Narsing Rao et al., 2023). The 16S rRNA amplicon profiling was consistent with this as the added culture was dominant in all cases (Figure 3). In Kom 1, Kom 3 and Kom 6, it was so numerically dominant that the AAB and LAB were essentially not represented in Table 4 for these samples as it was limited to 1% relative abundance. However, the expected repertoire of AAB and LAB were present at < 1% relative abundance in these samples (data not shown).
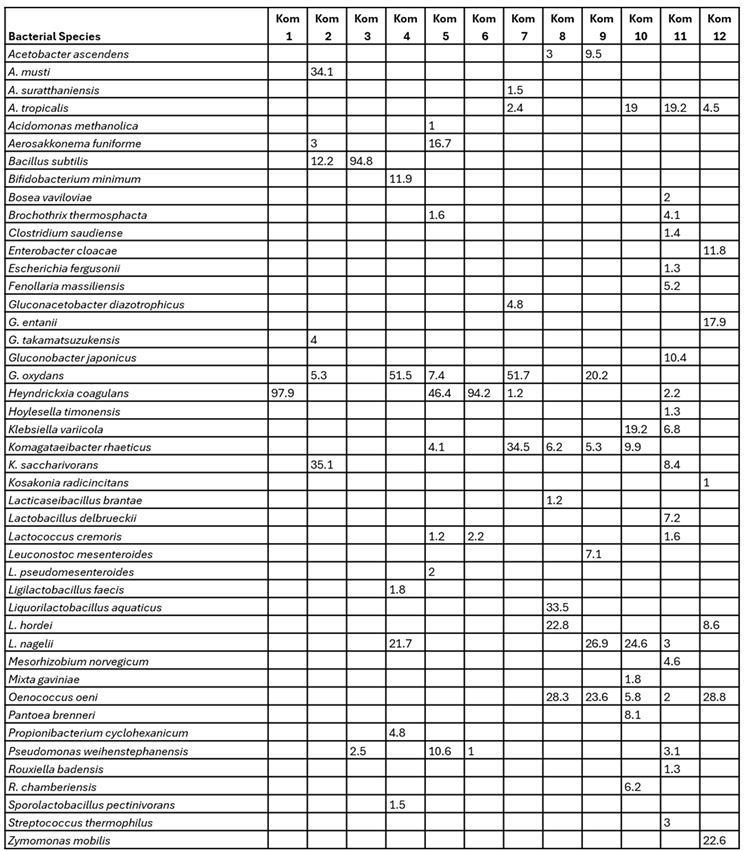
The major AAB genera represented were Acetobacter, Gluconobacter, Gluconacetobacter and Komagataeibacter which is consistent with other studies on Kombucha. The most dominant AAB genera in each Kombucha sample were Gluconobacter (Kom 1, Kom 4, Kom 5, Kom 6, Kom 7 and Kom 9), Komagataeibacter (Kom 2, Kom 3 and Kom 8), Acetobacter (Kom 10 and Kom 11) and Gluconacetobacter (Kom 12). Of the detected LAB genera, Liqorilactobacillus and Oenococcus were the two genera to comprise significant relative abundance in many of the Kombucha’s (Kom 4, Kom 8, Kom 9, Kom 10 and Kom 12) (Figure 3). Lactobacillus, Leuconostoc and Lactococcus) were represented at lower abundances in a few others (Kom 5, Kom 6 and Kom 11). The overall diversity of bacteria in the 12 Kombucha’s was greater than for fungi with 45 species representing 34 genera (Table 4).

Table 4.
Relative abundance of bacterial species identified (minimum 1%) from amplicon profiling for each Kombucha sample.
Table 4.
Relative abundance of bacterial species identified (minimum 1%) from amplicon profiling for each Kombucha sample.
4. Discussion
Kombucha has become an increasingly popular beverage at retail outlets in the US primarily due to its perceived health benefits from the bioactive components produced from the yeast and bacterial cultures during its fermentation process. This process is a two stage fermentation process whereby yeast and LAB first ferment simple sugars into ethanol and lactic acid and subsequently AAB aerobically oxidizes ethanol into acetic acid. The final beverage should be acidic and protected from foodborne bacterial pathogens. Two previous studies on retail available Kombucha reported pH ranges of beverages in New Zealand as 3.2 – 3.9 [11] and in Los Angeles, CA as 3.0 – 3.2 [23]. In this current study, the pH ranges of the 12 Kombucha beverages analyzed were 3.0 – 3.8, illustrating the protective acidic profile of these beverages against food pathogens. The appearance of colonies on MacConkey agar plates for Kom 2 and Kom 7 was initially surprising as this medium and variants thereof are widely used in food microbiology laboratories for the detection of fecal coliform Enterobacteriaceae [26,27]. However, some lactose fermenting strains of yeast with tolerance to bile salts can also form colonies on this medium [28].
The presence of viable cultures being present in Kombucha beverages depends on their acid tolerance over time. While fermentative yeast exhibit a higher tolerance than fermentative bacteria in general, many AAB and LAB can withstand acid environments for different time periods, with some AAB having evolved higher tolerances to acetic acid [29]. Yeast and AAB are frequently isolated from Kombucha, including commercial beverages, demonstrating their survival abilities under these conditions [11]. This is consistent with findings in this study based on microscopic analysis of colonies obtained on the different media. The molecular identification of one of these isolates from Kom 4 that exhibited a gram negative rod phenotype, indicated it was a strain of Acetobacter tropicalis. While this species was not detected in the major AAB of Kom 4 using the direct amplicon profiling (Table 4), it was present at lower relative abundances (0.2% of total reads) demonstrating the differential tolerance of individual AAB to acetic acid. LAB are generally more sensitive to prolonged exposure to pH < 4.0 and LAB fermented vegetable foods that are allowed to ferment until completion, such as sauerkraut, rapidly lose viability of LAB. However, some LAB can remain viable in the pH range 3 – 4 for extended periods. While most studies that examined viable cultures in Kombucha focused on AAB and yeast, some LAB genera such as Pediococcus have been isolated from Kombucha [30]. In this current study, a colony from an MRS plate of Kom 12 exhibited gram positive cocci cells arranged in chains, which is indicative of some LAB genera (Figure 1B). However, as Oenococcus oeni was the most dominant bacterial culture identified from the amplicon profiling of Kom 12 (Figure 3), it is most likely this organism. This is best known as an acid tolerant LAB organism important for the malolactic conversion of malic acid to lactic acid during the aging phase of wine production [31]. It has also been frequently isolated from Kombucha [32,33].
The dominant fungal genus in the most Kombucha beverages identified in this study was Brettanomyces and was detected in 9 of the 12 samples above the 1% relative abundance level (Table 3). This is consistent with microbial studies on Kombucha since first reported in a study by Mayser et al., (1995) where it was identified in 56% of samples tested [2]. Since then, it is one of the more frequent genera reported from Kombucha studies throughout the world. Another dominant yeast identified among the Kombucha beverages in this study was Lachancea cidri, which was detected in 10 of the 12 samples at significant levels (Figure 2). This species has not been reported previously as part of a Kombucha microbial consortium and is more frequently found in cider fermentations [9,34]. However, another species of Lachancea, L. fermentati has been found in a few Kombucha studies [35,36,37]. The fungus Malassezia with yeast-like cells was also a notable component in 6 of the 12 Kombucha beverages and represented > 35% of the relative abundance of Kom 1, Kom 3 and Kom 6. This fungus has also been reported as a component of Kombucha microbial studies previously [38,39]. The natural habitat for Malassezia fungi is the skin of humans and animals, where it is frequently associated with various dermatological disorders [40]. Its presence in some Kombucha beverages at such high levels is surprising. Torulaspora microellipsoides, an ethanol producing yeast of the Saccharomycetaceae family, was the major fungal representative of only one sample Kom 2 (66.5% abundance level). While not frequently reported as a dominant yeast component for Kombucha, it was detected at low levels in Kom 3 as well as in previous Kombucha studies [9]. Given it is an environmental yeast often associated with viticulture its dominance in a Kombucha fermentation is not surprising and illustrates the diversity of potential functional yeast varieties that can form the basis of a Kombucha culture.
Yeast are the primary ethanol producers in all Kombucha fermentations and were detected in all Kombucha beverages in this study. However, Kom 12 was unique in that it also had a significant proportion of its bacterial population (22.6% relative abundance) composed of the ethanol producing gram negative bacterium, Zymomonas mobilis [41]. This bacterium is found on certain plants, such as the Agave catus in Mexico and is best known for its involvement in the primary ethanol fermentation of Agave during Tequila production [42]. Its presence in the SCOBY used for the Kom 12 fermentation may reflect insights into its origin. Liquorilactobacillus was the most numerically dominant bacterial genus detected in Kom8, Kom 9 and Kom 10 (Figure 3). This LAB has previously been seen to predominate in a Kombucha microbial consortia during its evolution over 3 years and may reflect beverages produced with a mature SCOBY inoculum rather than a freshly generated one [43]. This is also reflective of its relatively high tolerance to acid compared it other LAB and is also of interest for its potential probiotic characteristics [44].
The term ‘probiotic’ was used on the beverage container by 10 of the 12 Kombuchas (Table 1). This term implies the ingestion of live microorganisms to confer a health benefit on the host and was first used in this context by Fuller (1989) [45]. While the use of the term by different authors has had numerous variations, all comply with ingesting a viable organism that can positively modulate the intestinal microflora of the host to confer a health benefit [46]. While four of the Kombucha’s did not have viable Kombucha fermentative cultures, likely due to pasteurization or other processing steps to terminate the fermentation, three of them did contain viable Bacillus cultures as probiotics (Figure 4). As these are endospore formers, they would survive these processing steps. It is ironic that the one beverage that did not contain any detectable viable organisms (Kom 11) used the brand name ‘Live Probiotic Kombucha’.
Author Contributions
ENO and DJO designed the study, carried out the experiments and data analysis, and wrote the manuscript. DJO supervised all aspects of the work and finalized all revisions.
Funding
UMN UROP provided student support funding to initiate this research. This research received no external sponsored funding.
Data Availability Statement
All data presented in the study are available in the main body of the manuscript or can be requested from the corresponding author.
Acknowledgments
The staff of the University of Minnesota Genomics Center are appreciated for their technical assistance with amplicon sequencing. We are grateful for the Undergraduate Research Opportunities Program (UROP) at the UMN for approving a student grant to ENO to initiate this research.
Conflicts of Interest
The authors declare no conflict of interes.
References
- de Oliveira:, P.V.; da Silva Júnior, A.H.; de Oliveira, C.R.S.; Assumpção, C.F.; Ogeda, C.H. Kombucha Benefits, Risks and Regulatory Frameworks: A Review. Food Chemistry Advances 2023, 2, 100288. [Google Scholar] [CrossRef]
- Mayser, P.; Fromme, S.; Leitzmann, G.; Gründer, K. The Yeast Spectrum of the ‘Tea Fungus Kombucha’: Das Hefespektrum Des ‘Teepilzes Kombucha. ’ Mycoses 1995, 38, 289–295. [Google Scholar] [CrossRef] [PubMed]
- Blanc, P.J. Characterization of the Tea Fungus Metabolites. Biotechnol Lett 1996, 18, 139–142. [Google Scholar] [CrossRef]
- Cambell-Platt, G. G. Campbell-Platt: Fermented Foods of the World. A Dictionary and Guide. 291 Seiten. Butterworth, London, Boston, Durban u. a. 1987. Preis: 35,— £ (Hardcover). Food/Nahrung 1989, 33, 304–304. [Google Scholar] [CrossRef]
- Jayabalan, R.; Malbaša, R.V.; Lončar, E.S.; Vitas, J.S.; Sathishkumar, M. A Review on Kombucha Tea—Microbiology, Composition, Fermentation, Beneficial Effects, Toxicity, and Tea Fungus. Comprehensive Reviews in Food Science and Food Safety 2014, 13, 538–550. [Google Scholar] [CrossRef]
- Kombucha Market Size, Share & Trends Report, 2022-2030 Available online: https://www.grandviewresearch.com/industry-analysis/kombucha-market (accessed on 18 April 2024).
- Chong, A.Q.; Lau, S.W.; Chin, N.L.; Talib, R.A.; Basha, R.K. Fermented Beverage Benefits: A Comprehensive Review and Comparison of Kombucha and Kefir Microbiome. Microorganisms 2023, 11, 1344. [Google Scholar] [CrossRef]
- Fabricio, M.F.; Vargas, B.K.; Tischer, B.; Wagner, R.; Ribeiro, S.R.; Cordeiro, N.; Flôres, S.H.; Záchia Ayub, M.A. Revamping Kombucha Production: Achieving Consistency and Probiotic Potential through a Tailor-Made Microbial Consortium. International Journal of Gastronomy and Food Science 2023, 34, 100844. [Google Scholar] [CrossRef]
- Coton, M.; Pawtowski, A.; Taminiau, B.; Burgaud, G.; Deniel, F.; Coulloumme-Labarthe, L.; Fall, A.; Daube, G.; Coton, E. Unraveling Microbial Ecology of Industrial-Scale Kombucha Fermentations by Metabarcoding and Culture-Based Methods. FEMS Microbiology Ecology 2017, 93, fix048. [Google Scholar] [CrossRef] [PubMed]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-Based Analysis of the Bacterial and Fungal Compositions of Multiple Kombucha (Tea Fungus) Samples. Food Microbiology 2014, 38, 171–178. [Google Scholar] [CrossRef]
- Wang, B.; Rutherfurd-Markwick, K.; Zhang, X.-X.; Mutukumira, A.N. Isolation and Characterisation of Dominant Acetic Acid Bacteria and Yeast Isolated from Kombucha Samples at Point of Sale in New Zealand. Current Research in Food Science 2022, 5, 835–844. [Google Scholar] [CrossRef]
- Harrison, K.; Navarro, R.; Jensen, K.; Cayler, W.; Nielsen, T.; Curtin, C. Live, Probiotic, or Neither? Microbial Composition of Retail-Available Kombucha and “Hard” Kombucha in the Pacific Northwest of the United States. Beverages 2023, 9, 59. [Google Scholar] [CrossRef]
- Hesseltine, C.W. A Millennium of Fungi, Food, and Fermentation. Mycologia 1965, 57, 149–197. [Google Scholar] [CrossRef]
- De Roos, J.; De Vuyst, L. Acetic Acid Bacteria in Fermented Foods and Beverages. Curr Opin Biotechnol 2018, 49, 115–119. [Google Scholar] [CrossRef]
- Kapp, J.M.; Sumner, W. Kombucha: A Systematic Review of the Empirical Evidence of Human Health Benefit. Annals of Epidemiology 2019, 30, 66–70. [Google Scholar] [CrossRef] [PubMed]
- Morales, D. Biological Activities of Kombucha Beverages: The Need of Clinical Evidence. Trends in Food Science & Technology 2020, 105, 323–333. [Google Scholar] [CrossRef]
- Rasouli, L.; Aryaeian, N.; Gorjian, M.; Nourbakhsh, M.; Amiri, F. Evaluation of Cytotoxicity and Anticancer Activity of Kombucha and Doxorubicin Combination Therapy on Colorectal Cancer Cell Line HCT-116. J Educ Health Promot 2021, 10, 376. [Google Scholar] [CrossRef]
- Xu, S.; Wang, Y.; Wang, J.; Geng, W. Kombucha Reduces Hyperglycemia in Type 2 Diabetes of Mice by Regulating Gut Microbiota and Its Metabolites. Foods 2022, 11, 754. [Google Scholar] [CrossRef]
- Wang, P.; Feng, Z.; Sang, X.; Chen, W.; Zhang, X.; Xiao, J.; Chen, Y.; Chen, Q.; Yang, M.; Su, J. Kombucha Ameliorates LPS-Induced Sepsis in a Mouse Model. Food Funct. 2021, 12, 10263–10280. [Google Scholar] [CrossRef]
- Costa, M.A. de C.; Vilela, D.L. de S.; Fraiz, G.M.; Lopes, I.L.; Coelho, A.I.M.; Castro, L.C.V.; Martin, J.G.P. Effect of Kombucha Intake on the Gut Microbiota and Obesity-Related Comorbidities: A Systematic Review. Crit Rev Food Sci Nutr 2023, 63, 3851–3866. [Google Scholar] [CrossRef]
- Arıkan, M.; Mitchell, A.L.; Finn, R.D.; Gürel, F. Microbial Composition of Kombucha Determined Using Amplicon Sequencing and Shotgun Metagenomics. J Food Sci 2020, 85, 455–464. [Google Scholar] [CrossRef]
- Yamada, Y.; Yukphan, P.; Vu, H.T.L.; Muramatsu, Y.; Ochaikul, D.; Nakagawa, Y. Subdivision of the Genus Gluconacetobacter Yamada, Hoshino and Ishikawa 1998: The Proposal of Komagatabacter Gen. Nov., for Strains Accommodated to the Gluconacetobacter Xylinus Group in the α-Proteobacteria. Ann Microbiol 2012, 62, 849–859. [Google Scholar] [CrossRef]
- Yang, J.; Lagishetty, V.; Kurnia, P.; Henning, S.M.; Ahdoot, A.I.; Jacobs, J.P. Microbial and Chemical Profiles of Commercial Kombucha Products. Nutrients 2022, 14, 670. [Google Scholar] [CrossRef] [PubMed]
- Gohl, D.M.; Vangay, P.; Garbe, J.; MacLean, A.; Hauge, A.; Becker, A.; Gould, T.J.; Clayton, J.B.; Johnson, T.J.; Hunter, R.; et al. Systematic Improvement of Amplicon Marker Gene Methods for Increased Accuracy in Microbiome Studies. Nat Biotechnol 2016, 34, 942–949. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- de Boer, E. Update on Media for Isolation of Enterobacteriaceae from Foods. Int J Food Microbiol 1998, 45, 43–53. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-S.; Kim, Y.-J.; Chon, J.-W.; Kim, D.-H.; Kim, K.-Y.; Seo, K.-H. Citrobacter Braakii: A Major Cause of False-Positive Results on MacConkey and Levine’s Eosin Methylene Blue Selective Agars Used for the Isolation of Escherichia Coli from Fresh Vegetable Samples. Journal of Food Safety 2016, 36, 33–37. [Google Scholar] [CrossRef]
- Hong, S.-M.; Kwon, H.-J.; Park, S.-J.; Seong, W.-J.; Kim, I.; Kim, J.-H. Genomic and Probiotic Characterization of SJP-SNU Strain of Pichia Kudriavzevii. AMB Express 2018, 8, 80. [Google Scholar] [CrossRef] [PubMed]
- Trček, J.; Mira, N.P.; Jarboe, L.R. Adaptation and Tolerance of Bacteria against Acetic Acid. Appl Microbiol Biotechnol 2015, 99, 6215–6229. [Google Scholar] [CrossRef] [PubMed]
- Diguță, C.F.; Nițoi, G.D.; Matei, F.; Luță, G.; Cornea, C.P. The Biotechnological Potential of Pediococcus Spp. Isolated from Kombucha Microbial Consortium. Foods 2020, 9, 1780. [Google Scholar] [CrossRef]
- Guzzo, J.; Coucheney, F.; Pierre, F.; Fortier, L.-C.; Delams, F.; Divies, C.; Tourdot-Maréchal, R. Acidophilic Behaviour of the Malolactic Bacterium Oenococcus Oeni. Sciences Des Aliments - SCI ALIMENT 2002, 22, 107–111. [Google Scholar] [CrossRef]
- Balmaseda, A.; Lorentzen, M.; Dutilh, L.; Bauduin, R.; Guichard, H.; Ollivier, S.; Miot-Sertier, C.; Lucas, P.M. Alcoholic Fermentation Drives the Selection of Oenococcus Oeni Strains in Wine but Not in Cider. International Journal of Food Microbiology 2023, 400, 110276. [Google Scholar] [CrossRef] [PubMed]
- Lorentzen, M.P.; Campbell-Sills, H.; Jorgensen, T.S.; Nielsen, T.K.; Coton, M.; Coton, E.; Hansen, L.; Lucas, P.M. Expanding the Biodiversity of Oenococcus Oeni through Comparative Genomics of Apple Cider and Kombucha Strains. BMC Genomics 2019, 20, 330. [Google Scholar] [CrossRef] [PubMed]
- Villarreal, P.; O’Donnell, S.; Agier, N.; Muñoz-Guzman, F.; Benavides-Parra, J.; Urbina, K.; Peña, T.A.; Solomon, M.; Nespolo, R.F.; Fischer, G.; et al. Domestication Signatures in the Non-Conventional Yeast Lachancea Cidri. mSystems 9, e01058-23. [CrossRef]
- Bellut, K.; Krogerus, K.; Arendt, E.K. Lachancea Fermentati Strains Isolated From Kombucha: Fundamental Insights, and Practical Application in Low Alcohol Beer Brewing. Front Microbiol 2020, 11, 764. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha Tea Fermentation: Microbial and Biochemical Dynamics. International Journal of Food Microbiology 2016, 220, 63–72. [Google Scholar] [CrossRef]
- Harrison, K.; Curtin, C. Microbial Composition of SCOBY Starter Cultures Used by Commercial Kombucha Brewers in North America. Microorganisms 2021, 9, 1060. [Google Scholar] [CrossRef]
- Barbosa, C.D.; Trovatti Uetanabaro, A.P.; Rodrigues Santos, W.C.; Caetano, R.G.; Albano, H.; Kato, R.; Cosenza, G.P.; Azeredo, A.; Góes-Neto, A.; Rosa, C.A.; et al. Microbial–Physicochemical Integrated Analysis of Kombucha Fermentation. LWT 2021, 148, 111788. [Google Scholar] [CrossRef]
- Kaashyap, M.; Cohen, M.; Mantri, N. Microbial Diversity and Characteristics of Kombucha as Revealed by Metagenomic and Physicochemical Analysis. Nutrients 2021, 13, 4446. [Google Scholar] [CrossRef]
- Prohic, A.; Jovovic Sadikovic, T.; Krupalija-Fazlic, M.; Kuskunovic-Vlahovljak, S. Malassezia Species in Healthy Skin and in Dermatological Conditions. Int J Dermatol 2016, 55, 494–504. [Google Scholar] [CrossRef]
- Panesar, P.S.; Marwaha, S.S.; Kennedy, J.F. Zymomonas Mobilis: An Alternative Ethanol Producer. Journal of Chemical Technology & Biotechnology 2006, 81, 623–635. [Google Scholar] [CrossRef]
- Escalante-Minakata, P.; Blaschek, H.P.; Barba de la Rosa, A.P.; Santos, L.; De León-Rodríguez, A. Identification of Yeast and Bacteria Involved in the Mezcal Fermentation of Agave Salmiana. Lett Appl Microbiol 2008, 46, 626–630. [Google Scholar] [CrossRef]
- Mas, P.; Tran, T.; Verdier, F.; Martin, A.; Alexandre, H.; Grandvalet, C.; Tourdot-Maréchal, R. Evolution in Composition of Kombucha Consortia over Three Consecutive Years in Production Context. Foods 2022, 11, 614. [Google Scholar] [CrossRef] [PubMed]
- Ruíz-Ramírez, Y.; Guadarrama-Mendoza, P.C.; Escalante, A.; Giles-Gómez, M.; Valadez-Blanco, R. Probiotic Activity Traits in Vitro and Production of Antimicrobial Peptides by Lactobacillaceae Isolates from Pulque Using Lactobacillus Acidophilus NCFM as Control. Braz J Microbiol 2022, 53, 921–933. [Google Scholar] [CrossRef] [PubMed]
- Fuller, R. Probiotics in Man and Animals. J Appl Bacteriol 1989, 66, 365–378. [Google Scholar]
- O’Sullivan, D.J. Genomics Can Advance the Potential for Probiotic Cultures to Improve Liver and Overall Health. Curr Pharm Des 2008, 14, 1376–1381. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
A: Isolated colonies from a MAC plate of a 100 dilution of Kom 7 with arrow pointing to microscopic view of the indicated colony. B: Isolated colonies from an MRS plate of a 10−3 dilution of Kom 12 with arrows pointing to microscopic views of the indicated colonies. C: Microscopic views of colonies depicting different yeast cellular morphologies from various Kombucha’s.
Figure 1.
A: Isolated colonies from a MAC plate of a 100 dilution of Kom 7 with arrow pointing to microscopic view of the indicated colony. B: Isolated colonies from an MRS plate of a 10−3 dilution of Kom 12 with arrows pointing to microscopic views of the indicated colonies. C: Microscopic views of colonies depicting different yeast cellular morphologies from various Kombucha’s.
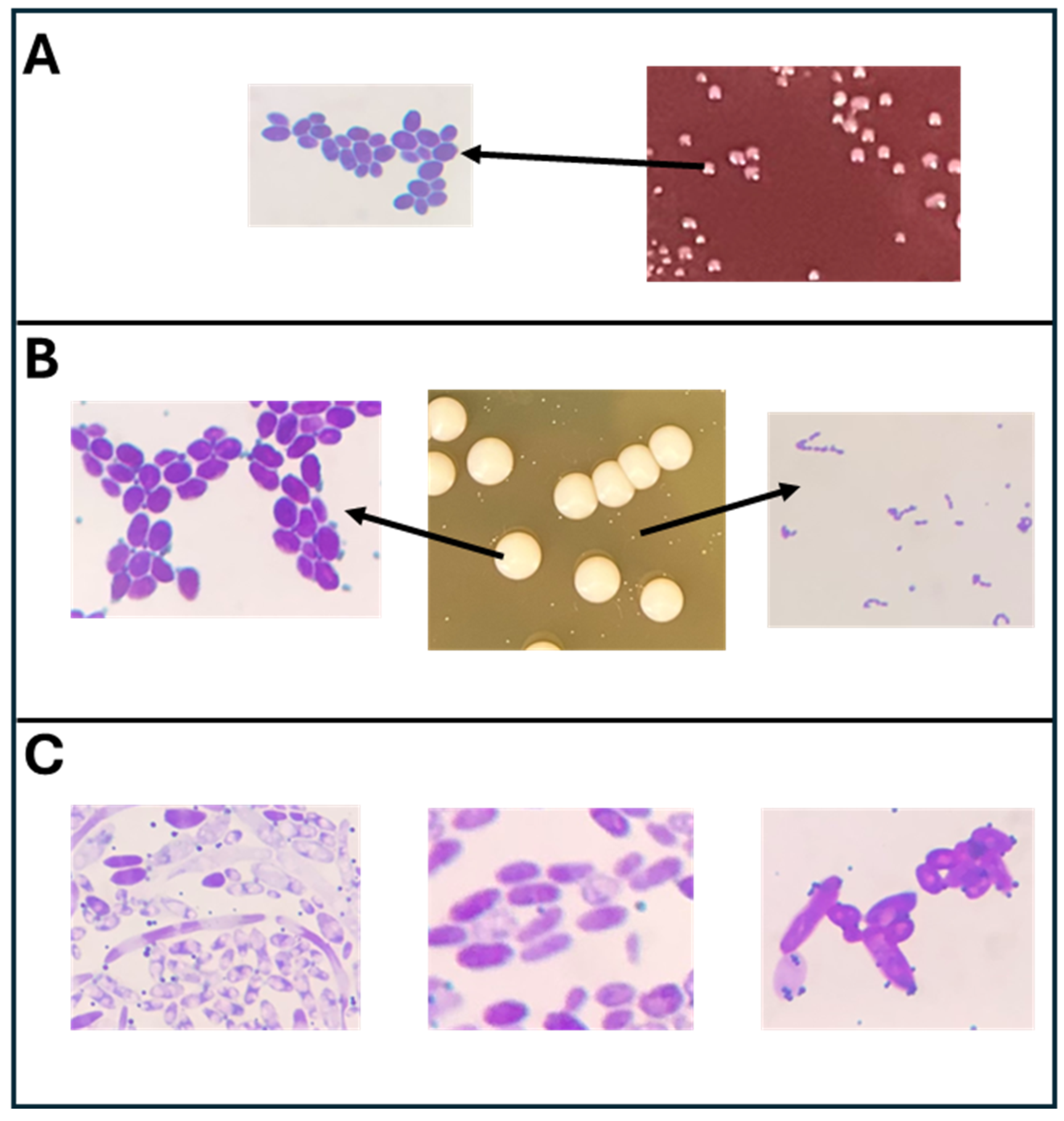
Figure 2.
Graphical representation of the numerically dominant genera of yeast cells identified in the 12 Kombucha beverages from ITS amplicon profiling.
Figure 2.
Graphical representation of the numerically dominant genera of yeast cells identified in the 12 Kombucha beverages from ITS amplicon profiling.
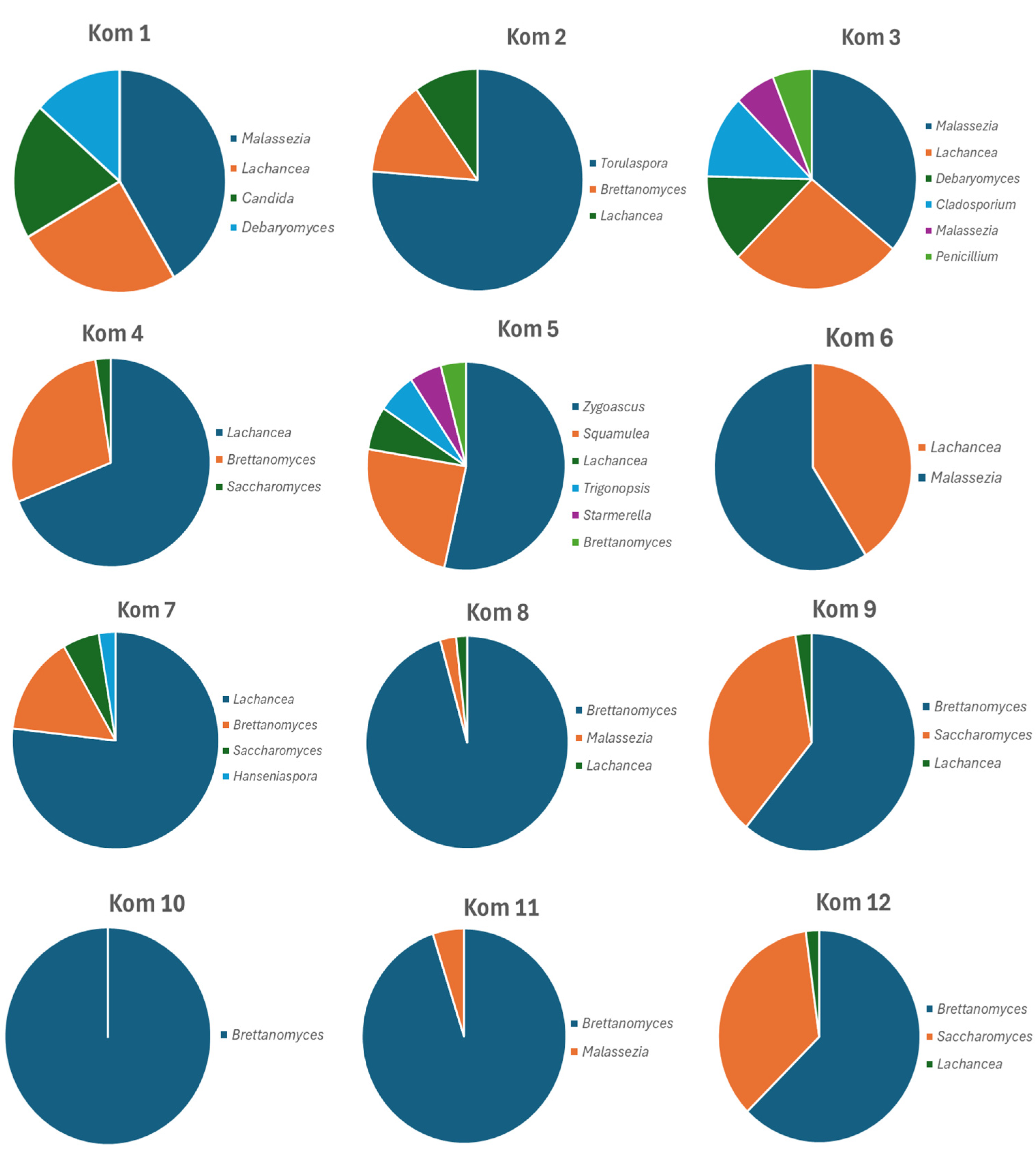
Figure 3.
Graphical representation of the numerically dominant genera of bacterial cells identified in the 12 Kombucha beverages from 16S rRNA gene amplicon profiling.
Figure 3.
Graphical representation of the numerically dominant genera of bacterial cells identified in the 12 Kombucha beverages from 16S rRNA gene amplicon profiling.
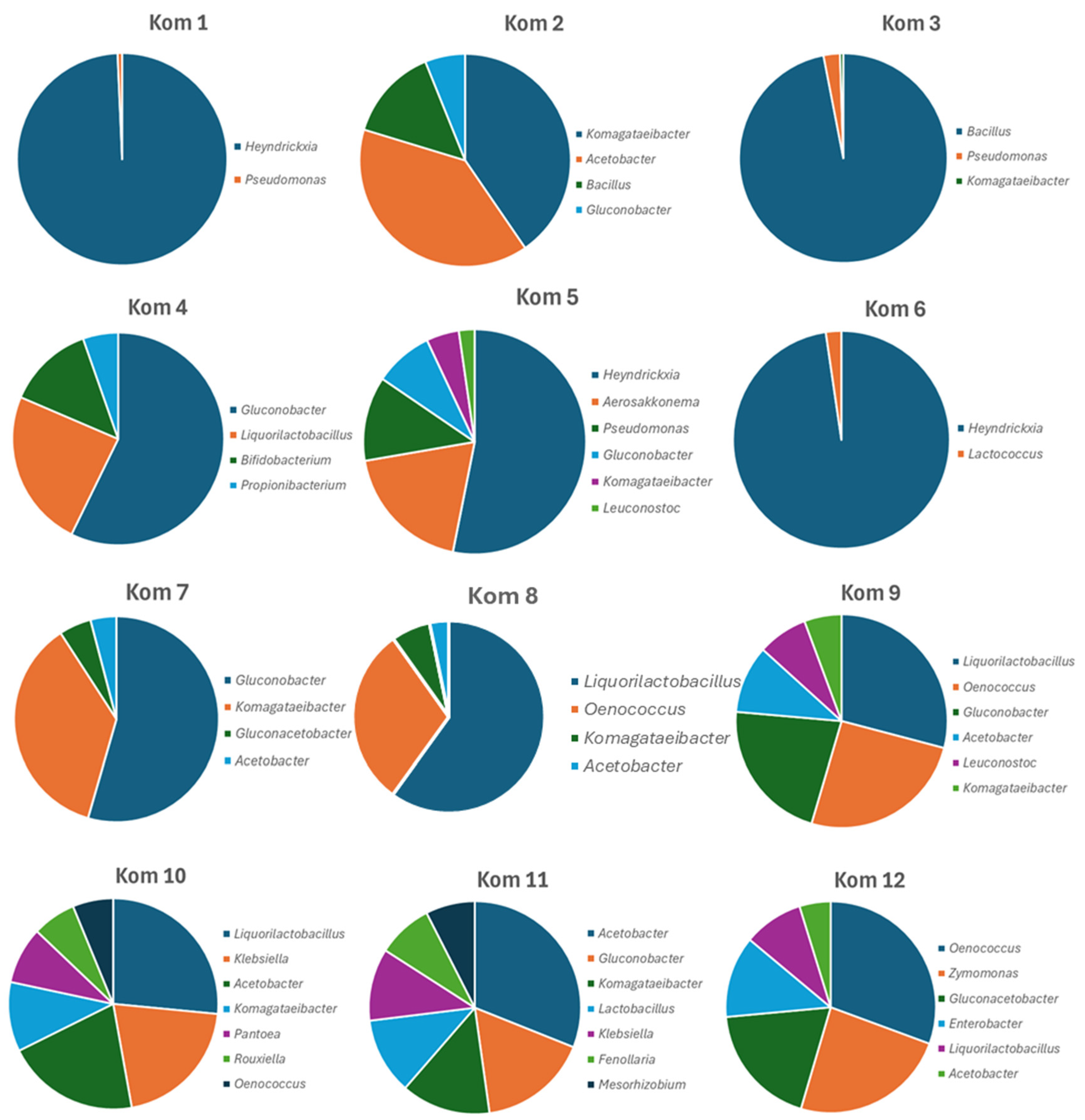
Figure 4.
Viable culture profile of the 12 Kombucha beverages analyzed in this study. Green, represents viable Kombucha cultures (Yeast, AAB and LAB). Red, represents viable added B. coagulans or B. subtilis. Gray, indicates no viable cells detected.
Figure 4.
Viable culture profile of the 12 Kombucha beverages analyzed in this study. Green, represents viable Kombucha cultures (Yeast, AAB and LAB). Red, represents viable added B. coagulans or B. subtilis. Gray, indicates no viable cells detected.

Table 1.
Commercial Kombucha beverages used in this study.
| Kombucha beverage | State | Relevant ingredients listed regarding microbial fermentation | Relevant label claims |
|---|---|---|---|
| Kom 1b | CA | Kombucha cultures, Bacillus coagulans MTCC5856, cane sugar, black tea, green tea, lime juice and grape juice, | “Bubbly probiotic tea” |
| Kom 2b | WA | Kombucha culture, Bacillus subtilis, cane sugar, tea blend, ginger juice, whole lemon puree | “2 billion CFU Probiotic Cultures” |
| Kom 3b | OR | Kombucha cultures, Bacillus subtilis, green tea, black tea, cane sugar, apple, white grape and passion fruit juice, mango puree | “Probiotic Kombucha” “2 billion Probiotic Cultures” |
| Kom 4b | OR | Kombucha culture, currant, Oolong tea, Elderberry, Hibiscus, Blueberry, Goji Berry, Strawberry, Raspberry, Cane Sugar | “Fizzy Probiotic Tea” |
| Kom 5b | CA | Kombucha culture, black tea, green tea, kiwi and ginger juice, Bacillus coagulans GBI-306086 | “9 billion Living Probiotics” |
| Kom 6b | CA | Kombucha culture, Bacillus coagulans MTCC 5856, black and green tea, cane sugar, stevia leaf extract, green coffee bean extract | “Live Probiotics” |
| Kom 7b | IL | Black tea, cane sugar, cherry and lemon juice concentrate | “Live Probiotics & Enzymes” “Heirloom cultures” |
| Kom 8c | MN | Black and green tea, cane sugar, apple juice | “Live Probiotics” “Active Kombucha Culture” |
| Kom 9c | WI | Kombucha culture, green tea, hibiscus flowers, cane sugar | “Forage Kombucha is Alive” “Kombucha Culture” |
| Kom 10b | MN | Peach oolong tea, mango turmeric tea, cane sugar | “Non-pasteurized” “Fresh Brewed” |
| Kom 11b | TX | Kombucha culture, sugar, stevia extract | “Live Sparkling Probiotic Kombucha" |
| Kom 12b | WI | Kombucha culture, black tea, safflower, cane sugar | “Contains millions of living probiotic cultures” “Live Kombucha Culture” |
b, bottled beverage; c, canned beverage.
Table 2.
pH readings and viable microbial plate counts obtained on the different agar media for the 12 Kombucha beverages.
Table 2.
pH readings and viable microbial plate counts obtained on the different agar media for the 12 Kombucha beverages.
| Kombucha Sample | pH Reading | Total CFU BHI (cfu/ml) |
Total CFU MRS (cfu/ml) | Total CFU ABS (cfu/ml) |
Total CFU PDA-Y (cfu/ml) |
Total CFU MAC (cfu/ml) |
|---|---|---|---|---|---|---|
| Kom 1 | 3.2 | 3.2 × 105 | 3.5 × 105 | nc1 | nc | nc |
| Kom 2 | 3.5 | 3.4 × 106 | 3.1 × 106 | 1.7 × 105 | 1.7 × 105 | 1.0 × 104 |
| Kom 3 | 3.7 | 5.0 × 105 | 2.5 × 106 | nc | nc | nc |
| Kom 4 | 3.3 | 3.0 × 103 | 5.5 × 103 | 1.2 × 104 | 1.2 × 104 | nc |
| Kom 5 | 3.1 | 1.1 × 104 | 3.3 × 103 | 7.0 × 103 | 1.1 × 104 | nc |
| Kom 6 | 3.2 | 1.0 × 103 | 1.0 × 103 | nc | nc | nc |
| Kom 7 | 3.3 | 5.5 × 105 | 9.7 × 105 | 8.5 × 105 | 4.7 × 105 | 3.0 × 103 |
| Kom 8 | 3.6 | 2.0 × 105 | 6.3 × 105 | 5.2 × 106 | 1.5 × 106 | nc |
| Kom 9 | 3.0 | 2.4 × 106 | 1.5 × 106 | 1.6 × 106 | 2.0 × 106 | nc |
| Kom 10 | 3.4 | 1.1 × 106 | 2.2 × 106 | 3.8 × 106 | 6.0 × 106 | nc |
| Kom 11 | 3.5 | nc | nc | nc | nc | nc |
| Kom 12 | 3.8 | 2.3 × 106 | 1.9 × 106 | 3.1 × 106 | 2.1 × 106 | nc |
1, no colonies.
Table 3.
Relative abundance of fungal species identified (minimum 1%) from amplicon profiling for each Kombucha sample.
Table 3.
Relative abundance of fungal species identified (minimum 1%) from amplicon profiling for each Kombucha sample.
| Fungal Species | Kom 1 | Kom 2 | Kom 3 | Kom 4 | Kom 5 | Kom 6 | Kom 7 | Kom 8 | Kom 9 | Kom 10 | Kom 11 | Kom 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Allodekkera sacchari | 7.1 | |||||||||||
| Brettanomyces anomalus | 14.7 | 4.1 | 11.8 | 3.5 | 35.2 | |||||||
| B. bruxellensis | 12.2 | 26.5 | 3.8 | 88.7 | 48.9 | 95.1 | 94.5 | 26.6 | ||||
| Candida mesenterica | 5 | 1.6 | ||||||||||
| C. parapsilosis | 14 | |||||||||||
| Cladosporium angustiherbarum | 3 | 11.5 | ||||||||||
| Cyberlindnera jadinii | 3.5 | |||||||||||
| Debaryomyces psychrosporus | 13 | 12.2 | ||||||||||
| Gibellulopsis serrae | 1.2 | |||||||||||
| Hanseniaspora valbyensis | 2.6 | |||||||||||
| Lachancea cidri | 24.1 | 8.5 | 25 | 67.8 | 6.2 | 38.5 | 75.5 | 1.7 | 2.5 | 2.1 | ||
| Malassezia arunalokei | 14.5 | 1.6 | 38.5 | |||||||||
| M. globosa | 5.9 | 14.4 | 2.5 | |||||||||
| M. restricta | 25.2 | 33.8 | 2.4 | 2.4 | 2 | |||||||
| M. slooffiae | 3 | |||||||||||
| Penicillium hordei | 5.7 | |||||||||||
| Pichia cecembensis | 1.8 | |||||||||||
| Saccharomyces bayanus | 2.5 | |||||||||||
| S. cerevisiae | 2.1 | 5.6 | 36.5 | 34.8 | ||||||||
| Squamulea flakusii | 22.2 | |||||||||||
| Starmerella davenportii | 4.8 | |||||||||||
| Torulaspora microellipsoides | 66.5 | 2 | ||||||||||
| Trigonopsis variabilis | 5.8 | |||||||||||
| Zygoascus hellenicus | 49.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



