
Submitted:
10 May 2024
Posted:
13 May 2024
You are already at the latest version
Abstract
In a constantly changing environment brought about by the climate crisis and escalated anthropogenic perturbations driven by the growing population, harmful algal bloom dynamics and their impacts are expected to shift, necessitating adaptive management strategies and comprehensive research efforts. Similar to primary productivity, HABs have been thought to be driven primarily by major nutrients such as N, P, and Si. However, recent investigations on the role and importance of micronutrients as limiting factors in aquatic environments have been highlighted. This paper provides a review of metal and phytoplankton interactions, with a specific emphasis on pertinent information on the influence of trace nutrients on growth, toxin production, and other underlying mechanisms related to the dynamics of HABs. Low to near-depleted levels of essential nutrients including Fe, Cu, Zn, Se, Mn, Co, and Mo, negatively impact cell growth and proliferation of various marine and freshwater HAB species. However, evidence shows that at elevated levels, these trace elements along with other non-essential ones, could still cause toxic effects to certain HAB species manifested by decreased photosynthetic activities, oxidative stress, ultrastructure damage, and cyst formation. Interestingly, while elevated levels of these metals mostly result in increased toxin production, Co (i.e., yessotoxins, gymnodimine, and palytoxins) and Mn (i.e., isodomoic acid, okadaic and diol esters) enrichments revealed otherwise. In addition to toxin production, releasing dissolved organic matter (DOM) including dissolved organic carbon (DOC) and humic substances was observed as an adaptation strategy, since these organic compounds have been proven to chelate metals in the water column, thereby reducing metal-induced toxicity. Whilst current research center on free metal toxicity of specific essential elements such as in Cu and Zn, a comprehensive account of how trace metals contribute to the growth, toxin production, and other metabolic processes under conditions reflective of in-situ scenarios of HAB-prone areas, would yield new perspectives on the roles of trace metals in HABs. With the growing demands of the global population for food security and sustainability, substantial pressure is exerted on the agriculture and aquaculture sector, highlighting the need for effective communication of information regarding the interactions of macro- and micronutrients with HABs to improve existing policies and practices.
Keywords:
Metal bioavailability
; toxin production
; nutrient limitation
; harmful algal bloom dynamics
; metal contamination
; metal-phytoplankton feedback interaction
1. Introduction
Harmful algal blooms (HABs) have been an ever-evolving concern globally, and are characterized by their unpredictable occurrences, diverse impacts, and complex underlying mechanisms [1,2,3,4]. As drastic changes in climate patterns coupled with increasing anthropogenic disturbances continue, the frequency, intensity, duration, and distribution of HAB events are expected to shift leading to deleterious consequences in aquatic ecosystems[5,6,7,8]. These outcomes include oxygen depletion, mechanical damages (e.g., disruption of fish gills), and production of toxic substances affecting not only their immediate consumers but also those in higher trophic levels and even inflicting fatalities on humans[4,9]. Persistent HAB events can shut down aquaculture facilities as well as economically important fishing zones over a long period, resulting in massive economic losses[10,11], highlighting the need for further development of robust detection tools.
While progress has facilitated thorough research on mitigating the impacts of HABs, obstacles to the advancement of early warning systems persist. For instance, most detection models to date, primarily utilize oceanographic data that are combined solely with macronutrient loading data [12,13,14]. Recent investigations on the role and importance of micronutrients as limiting factors in aquatic environments have been highlighted [1,15,16,17]. Hence, the lack of inclusion of these parameters could lead to potential gaps in the overall species-specific prediction of HAB events. Advancements in monitoring technologies and analytical capability constantly reveal new insights into the drivers and dynamics of HABs. The dynamic nature of HABs underscores the need for adaptive management strategies and multi-pronged research efforts to mitigate their impacts on the environment, public health, and economies worldwide.
In this review, we will expand the current knowledge base for the dynamics of HABs, focusing on the roles of trace nutrients on growth, toxin production, and other underlying mechanisms related to bloom dynamics.
2. Frequency and Diversification of HABs
While extensive discussions about the worldwide spread of HABs only began recently, historical records dating back centuries ago document HAB occurrences and HAB-associated poisoning [18,19,20]. Through the years, some factions in the scientific community have been skeptical about the persistence of HABs as a global phenomenon as supported by the argument that increase in reported incidence is due to heightened awareness and monitoring efforts (see Figure 1) [1,3,4,21,22]. Interestingly, recent statistical analysis on global HAB events has found that once distribution data were normalized against variations in regional monitoring activities conducted, no uniform global trend exists, suggesting that observable trends should be considered on a regional scale as well as species level [3]. Despite debates surrounding HAB frequencies, a pattern of diversification in HAB occurrences over time has been noted across various regions [23,24,25,26]. This phenomenon impacts new sites and is attributed to climate change and intensified anthropogenic disturbances, which in turn contribute to the persistence of HABs in coastal and freshwater ecosystems (Figure 1).
2.1. Major Drivers: Changing Climate and the Global Population
Occurrences of algal blooms and the natural succession in phytoplankton community structure are shaped by nutrient loading from seasonal rains, flooding, upwelling [9,27], and atmospheric deposition from major deserts [28]. In oligotrophic oceans where nitrogen is limited, the bloom and bust of nitrogen-fixing organisms initiate succession of organisms up to higher trophic levels. The imbalance in some key physicochemical parameters necessary for growth initiates harmful blooms, which are associated with climate crisis coupled with increased anthropogenic perturbations that exacerbate the impact of algal blooms across regions [5,7,8,29].
Drastic changes in global climate patterns affecting temperatures [30,31], sea levels [11], rainfall and precipitation [32,33], atmospheric CO2 concentrations [34], acidification [35,36], and water stratification [37,38] influence species-specific algal bloom dynamics. This outcome is best illustrated by algal blooms following volcanic eruptions or inundation brought by typhoons.
Furthermore, apart from the direct impacts of urbanization and expanding human settlements, the increasing global population has led to massively growing demands for food production, placing substantial pressure primarily on the agriculture and aquaculture sectors. Under varied socio-economic and climate change scenarios, the projected global food demand is expected to increase from 35% to 56% between 2010 and 2050 [39]. Currently, such demands are met by expansion and increased productivity of both sectors.
According to FAO in 2003, fertilizer consumption will increase by 1% per annum from 138 million tonnes in 1997/99 to 188 million tonnes in 2030 [40]. As of 2022, however, the demand for fertilizer has already exceeded the 2030 prediction and was reported at 200 million tonnes [41]. The heavy use and reliance on fertilizer has an unwanted outcome because poorly managed agricultural nutrient enrichments lead to higher levels of nutrient run-off, including key elements like nitrogen [42,43,44,45] and phosphorus [46,47,48,49], entering bodies of water.
Similar trends in market demands for commercial feeds for livestock and aquaculture facilities were observed [50]. Interestingly, these demands are met by around 30% of cereal crop production utilizing about 40% of the world’s arable land [51,52], and around 23% of capture fishery [53]. This complex competition between feed and food production is deemed as one of the major bottlenecks in achieving sustainable food production systems [50]. Moreover, excess macronutrients, trace elements, and other chemicals from the unregulated use of feed products contribute further to eutrophication, impacting various aquatic ecosystems [54,55].
2.2. Roles of Macronutrients in Bloom Dynamics
Eutrophication has been identified as one of the major culprits of algal bloom events since macronutrients from agricultural runoffs and aquaculture wastes including nitrogen, phosphorus, and to some extent, silica, play critical roles in species-specific bloom dynamics of phytoplankton species. Variations in concentration and ratio of these major nutrients lead to diverse plankton community compositions, potentially giving rise to either HAB or non-HAB bloom formations.
Nitrogen (N) plays a fundamental metabolic role in phytoplankton physiology, serving as a building block for various cellular components such as nucleic acids, proteins, and chlorophyll, which are essential for algal growth and biomass production [56,57]. Various nitrogen enrichment experiments recorded high cell densities and faster growth rates of different microalgal species [56,58,59]. In addition, phytoplankton community structure shifts were reported as a result of increased nutrient loading, primarily N [60,61,62,63]. In oligotrophic areas such as the tropical Pacific [64,65] and North Atlantic [66], phytoplankton biomass has increased after the addition of N during in situ enrichment studies. Although several studies have shown that fast-growing diatoms and cyanobacteria dominate marine [61,62] and estuarine ecosystems [67] with high N concentrations, other phytoplankton taxa were also found to dominate in certain areas. Aquatic ecosystems with high N:P ratios were found to have dinoflagellates as primary producers [68,69] whilst other cultivation experiments reported that N:P ratios have no significant effect on cell abundance, reiterating that it is the N sources alone that significantly affects growth [59,70]. Apart from the fundamental role of N in cell division and growth as well as in macromolecule synthesis, N is also essential for dinoflagellate toxin metabolism, which has been of interest over the years due to deleterious impacts to the environment as well as in public health [71,72,73,74,75,76].
Phosphorus (P) has several key roles in cellular metabolism particularly in processes related to energy transfer, nucleic acid synthesis, and cell membrane structure [77]. It serves as a key constituent of adenosine triphosphate (ATP), which fuels cellular metabolism by providing energy for biochemical reactions within cells [78]. P is also essential for the synthesis of nucleic acids (DNA and RNA), which are crucial for cell growth, division, and protein synthesis [79]. Furthermore, P is a structural component of cell membranes, phospholipids, and various cellular molecules that contribute to the integrity and fluidity of cell membranes. Ding et al., [80] demonstrated the ability of P limitation to change nutrient limiting conditions for HAB formation. These metabolic functions underscore its significance as a limiting nutrient in marine ecosystems with implications for phytoplankton productivity and ecosystem dynamics [66,81,82].
Silica (Si) has a more species-specific role in phytoplankton particularly in diatoms. It serves as a structural component of their frustules, which are external siliceous cell walls providing mechanical support and protection from predators. These siliceous structures also regulate the uptake and transport of essential nutrients, such as nitrogen, phosphorus, and carbon through specialized silica transporters [83,84]. Silica metabolism is closely linked to photosynthesis and carbon fixation processes in phytoplankton, ultimately impacting primary productivity and the biogeochemical cycling of nutrients in aquatic ecosystems [85,86,87].
These variations in species-specific responses to the major nutrient enrichments can generally be explained by differences in the species physiology [61], theory of resource competition [88], and the widely observed Redfield ratio, stating that elemental composition of phytoplankton biomass across environments follows a ratio of 106:16:1 for C:N:P [89,90,91]. Under sufficient N and P supply in marine environments, maximum phytoplankton growth rate can be achieved with the Redfield ratio [92]. Though, it has been reported that nutrient requirements by phytoplankton result in deviations from the Redfield ratio [60,91]. As such, the silicate requirement by diatoms merits the inclusion of Si as part of this ratio [93].
Backed by global documentation, primary productivity was always thought to be driven by the availability of macronutrients primarily N, P, and Si. However, results from research spanning the past decades revealed that trace nutrient availability also impacts productivity and phytoplankton community structure in marine and freshwater environments across the globe [94,95,96,97,98]. Advancements in monitoring technology and analytical tools have permitted exploring the influence of trace elements on algal productivity and various cellular and biochemical processes. Gathered data from such investigations hold the potential to enhance our understanding of HABs dynamics.
2.3. Current Efforts for Algal Bloom Detection
In the pursuit of mitigating the detrimental impacts of HABs, there has been a substantial focus on development and enhancement of HAB prediction, detection, and other early-warning systems. Current methods involve the use of satellite imagery, bathymetry data, in-situ physicochemical data, and even machine learning for algorithms used in modeling [13,99,100,101]. In marine environments, physicochemical parameters such as DO, temperature, salinity, Chl A, and pH are the few primary considerations for in-situ prediction of HABs. These factors are most often selected due to their high correlation with HAB occurrences, the availability of data during all periods of fish kill or toxic bloom, and the capacity to obtain the real-time parameters [102]. Recently, the inclusion of macronutrients such as N, P, and Si in the regular monitoring of HAB-prone areas as additional prediction parameters has been highlighted [103]. Incorporating nutrient loads of particulate phosphorus (PP) to other physicochemical parameters improved prediction models in HAB formation in Lake Erie [104]. Furthermore, nitrogen (N) loads were also found to be a valuable parameter in the prediction of HAB formation in the same lake [105]. More complex models utilize different sources and types of nutrients as part of the predictive drivers for the modeling of cyanobacterial HABs [106]. The contribution of nutrient loading to eutrophication in coastal systems has been demonstrated and has warranted its inclusion as a predictive driver in several HAB modeling papers [42,107]. The predictive capabilities of models are reliant on existing data, which includes results from bottle-incubation and mesocosm experiments. However, continuous, and long-term in situ observations are still necessary in fine-tuning these models, especially in a changing climate [14]. Increasing evidence of the roles of metals in the initiation and development of HABs underscores the importance of its inclusion in long-term monitoring [12,108]. Furthermore, studies detailing the effects of metals on growth, toxin production, and allelopathic effects may improve prediction models. This is especially true for those involving complex biological systems that not only consider the growth of a species capable of forming harmful blooms but also trophic succession and the upstream risks that come with HAB events.
3. Trace Metals
3.1. Overview of Trace Metal Distribution and Bioavailability
Concentrations of many trace metals typically increase with depth of the water column and proximity to terrestrial ecosystems [109,110,111] due to natural and anthropogenic inputs such as continental dust [44,112,113,114,115], shelf sediments [116,117,118], hydrothermal circulation at ocean ridges [119,120,121], glaciers [122,123,124], atmospheric ash [125,126,127], coastal sediments and riverine outputs [128,129,130]. Interestingly, recent studies have also explored the potential contribution of fecal materials of marine mammals, seabirds, and other migratory animals in trace metal deposition in the oceans [131,132,133].
Most metals including iron (Fe), zinc (Zn), copper (Cu), nickel (Ni), selenium (Se), and cadmium (Cd) have a typical nutrient-type distribution similar to macronutrients [111,134,135]. These metals generally increase in concentrations with depth and proximity to landmasses [136]. Co, similar to Fe, has a nutrient-like profile. However, its concentrations in deep waters do not necessarily increase due to organic complexation [137,138,139].
Other metals such as manganese (Mn), chromium (Cr), molybdenum (Mo), and vanadium (V) have different behaviors in seawater and are largely conservative [140,141,142,143]. Mn has an established maximum layer in surface waters and in the oxygen minimum zone, while the lowest concentrations are in deep waters [140]. Cr is found to be slightly depleted in subsurface waters. However, it was found that decoupling of Cr happens in deeper waters as observed in subtropical North Atlantic [144]. Mo and V are under-explored elements due to their largely conservative nature. Mo concentration, in the form of molybdate (MoO42-), is influenced by salinity rather than depth and shows little to no differences in its vertical distribution [145,146]. However, recent studies have reported the non-conservative nature of Mo, especially in its horizontal distribution [146]. In some cases, dissolved V concentrations are presented as non-conservative oceanic profiles, but variability is generally observed across ocean basins. The variations are attributed to factors such as riverine inputs, reducing shelf environments, and chemically driven particle interactions, all of which highlight the call to further investigate the V cycle [147].
In most freshwater lakes, the seasonal release of macronutrients and Fe species from sediments occurs due to thermal stratification, upwelling, and mixing [148,149,150]. However, anthropogenic sources such as wastewater discharge, mining, and changing land-use practices caused by urbanization have impacted metal deposition in such habitats [151,152]. Wastewater discharge from human settlements contributes to the accumulation of Cd, Ni, and Zn, with 60% of the total claimed to be recycled by phytoplankton [152,153]. In these ecosystems, the spatiotemporal distribution of trace nutrients exhibits significant variability owing to variations in bathymetry, mixing patterns, temperature regimes, and other physicochemical parameters [154,155,156]. Ultimately, this variability is influenced by fluctuating levels of disturbances from nearby anthropogenic settlements e.g., water runoffs, aerosols, etc. [153,157].
Trace metal elements exist in several chemical species influencing their chemistry, biological availability, and hence, status as a limiting nutrient [15]. Most metals occur as cations in complexes with inorganic or organic ligands. Essential trace elements such as Fe, Cu, Mn, and Co, exist in more than one oxidation state resulting in varied reaction energy requirements and reaction rates affecting their bioavailability [111,158]. For example, the most biologically available form of Fe is ferrous iron (Fe2+), which is highly soluble in anoxic waters. However, it can be rapidly oxidized to weakly soluble ferric iron (Fe3+) in neutral to slightly alkaline oxygenated waters [159,160,161]. Other metals including Zn, Ni, and Cd exist as soluble divalent cations complexed with inorganics such as chlorides, hydroxides, carbonates, and other organic chelators (binding agents). The chemical speciation of these trace metals in the water column affects their biological availability for phytoplankton uptake and assimilation [162].
3.2. Overview of Roles of Trace Metals in Photosynthetic Microorganisms
Bioavailable trace elements are essential for phytoplankton growth and metabolism resulting in varied productivities across ocean basins, coastal regions, and other aquatic ecosystems. A growing number of literature (Table 1) demonstrates the roles of essential trace metals–Fe, Mn, Ni, Co, Cu, and Zn, as well as species-specific essential metals including Cd, Mo, and V, based on their function as active centers or structural support in enzymes involved in various biochemical pathways [96,98,163,164,165,166,167,168,169,170].
Fe is widely accepted as a key micronutrient that limits phytoplankton productivity in vast oceanic regions [171] and this can be attributed to the high Fe requirements of various phytoplankton species [95,96]. Fe serves as a cofactor for a wide range of essential enzymes required in different metabolic processes. It is integral to the structure and function of pigments, electron transport systems, and other components of the photosynthetic machinery. Additionally, Fe-containing enzymes are crucial for nutrient acquisition and assimilation, respiration, and RNA and DNA synthesis. All these processes comprise a pipeline for the conversion of nutrients into biomass and energy. Hence, influencing ecosystem-level interactions and various biogeochemical cycles.
Moreover, Fe along with other trace elements (Table 1) contribute to various antioxidative defense mechanisms and other metabolic processes to ensure phytoplankton survival and growth, thus shaping ecosystem dynamics [96,97,167,172]. Although a wide range of metabolic roles in phytoplankton has been well documented during the past decades, the foundation of phytoplankton metal physiology mostly come from studies on diatoms [97,168,173,174,175,176,177,178,179,180,181] and recently, cyanobacteria including Prochlorococcus [182,183,184], Synechococcus [185], Crocosphaera [186,187], Cyanothece [188], Trichodesmium [186,189,190,191,192], and several others.
In recent years, investigations on the effects of trace metal concentrations on photosynthetic microorganisms have expanded to other taxa–picoeukaryotes, dinoflagellates, green algae, prymnesiophytes, and other nano-picophytoplankton, due to their significant roles in biogeochemical cycles. Studies have shown that metal requirements vary among different phytoplankton functional types, suggesting that the current understanding of metal functions in diatoms and diazotrophs might not apply to others. For example, in similarly designed studies conducted on diatoms and a picoeukaryote, Pelagomonas calceolata, the Fe quotas exhibited a 100-fold difference with P. calceolata showing minimal physiological plasticity [193]. Similar observations were reported with another picoeukaryote, Ostreococcus lucimarinus, wherein iron limitation resulted in 68% reduction in iron quota, which is just a fraction of plasticity reported in diatoms [184]. In a study investigating the metal composition of 15 major marine phytoplankton groups, systematic differences were observed in diatoms, chlorophytes, and coccolithophores [95]. Differences were also observed between strains isolated from coastal and oceanic regions [194]. Moreover, several members of Symbiodiniaceae, a family of marine dinoflagellates that are notable for their symbiotic relationships with reef-building corals [195], have shown different metal requirements when subjected to various metal conditions, indicating the complex roles of trace nutrients in their growth [57,189,190,191] and other metabolic processes [196].
The variations in trace metal quotas and metal requirements of these taxa reflect the complexity of phytoplankton metal physiology, highlighting the need for further investigations of trace elements and their roles in phytoplankton growth, metabolic processes, bloom dynamics, and other ecosystem-level interactions.
4. Trace Metals and Harmful Algal Blooms
The diverse roles of metals in the physiology of phytoplankton indicate numerous possibilities on how it would affect HAB events and their causative species. Discerning the effects of metals on HABs is complicated as they can affect either the individual cells or the HAB events. Aside from their usual effect on growth, metals may also be vital in other processes that support the proliferation of HABs such as toxin production, dissolution of and protection against other metals, and synergistic effects with other compounds produced [175,197,198,199,200]. Several pieces of evidence reveal that the relationship between metals and HABs is not unidirectional, since some species can produce compounds that affect the bioavailability of metals in the environment [175,201]. Nonetheless, exposure to high concentrations can still lead to toxicity [202,203]. Describing the effects of metal on these organisms entails a meticulous degree of planning and experimentation to elucidate underlying mechanisms.
A growing number of bottle incubation experiments, mesocosm studies, and field investigations have been reported to visualize the role of trace metals on certain marine and freshwater HAB species (see Table 2 and Table 3). Bottle incubation experiments entail culturing HAB-causing species, aiming to uncover how certain species respond physiologically to various growth factors like metals, macronutrients, micronutrients, or their combinations. Meanwhile, mesocosm experiments, conducted on a larger scale, strive to replicate real-world conditions more closely while maintaining adequate control to distinguish responses among treatments. While these approaches enable researchers to delve into fundamental biological inquiries, these may only infer the complex interconnectivity of processes involved in actual HAB occurrence and persistence. Thus, regardless of logistical challenges, emphasis is placed on in situ investigations such as metal concentration monitoring before, during, and after the onslaught of HABs. Complementing bottle incubations with mesocosm and field investigations on various physicochemical parameters can provide more extensive elucidation of bloom dynamics in a feedback loop.
4.1. Iron (Fe)
This section will highlight conducted research focused on how metal availability influences HABs and HAB-causing organisms (Table 2 and Table 3). Discussions will mainly focus on Fe, Co, Zn, Se, and other metals, both essential and non-essential.
Among metals, Fe is the most heavily studied in terms of its effects on the physiology of HABs. Growth studies indicate that Fe is an essential metal for both marine and freshwater HAB causative species [16,204,205,206]. Phytoplankton are heavily reliant on Fe for its role on the major photosynthetic complexes and as cofactors for many physiologically important enzymes. Bottle experiments show that decreasing Fe concentrations cause a decrease in physiological activities related to growth and proliferation. Specifically, Fe depletion or low Fe conditions have been observed to decrease cell density and growth rate on freshwater Microcystis aeruginosa [207,208,209,210,211] and marine HAB species, Alexandrium minutum [197,209], Alexandrium catenella [212], Protoceratium reticulatum [213], Pseudo-nitzschia australis [214], Chattonella antiqua [215], Heterosigma akashiwo [215], Heterocapsa circularisquama [215], Scrippsiella trochoidea [206], Gymnodinium sanguineum [216,217]. Interestingly, synergistic effects of Fe and humic compounds in improving growth rates have been established for M. aeruginosa [208]. Nevertheless, elevated Fe concentrations can still result in toxicity for certain HAB species [209,212]. The role of Fe in enzymes essential for photosynthetic activities and various metabolic processes including nutrient uptake and assimilation, underscore its significance in the universal requirements of phytoplankton for Fe [94,171]. Hence, mitigation strategies targeting HABs should consider the significance of Fe in phytoplankton physiology.
Moreover, increasing correlations between Fe levels and toxins produced by specific HAB species were reported. The freshwater M. aeruginosa, a known cyanoHAB species, is capable of producing microcystin, a hepatotoxic and carcinogenic compound [218,219]. Incubation experiments revealed contrasting trends for production of microcystin. Lukač & Aegerter [207] have shown increased toxin production at lower or reduced Fe concentrations while other investigations on the same species presented otherwise. Facey et al. [211] reported reduced microcystin-LR in Fe-deficient media after 20 days. Moreover, Alexova et al. [220] have shown decreased toxin levels in increasing Fe concentrations. This evidence highlights the complex yet potential role of Fe on microcystin production which stipulates further studies to elucidate this relationship. Nonetheless, the possible role and importance of Fe in the alleviation of damage caused by M. aeruginosa is highlighted.
Other forms of toxic substances produced by HAB species shed some light on the potential metabolic role of Fe on toxin production. Alterations in toxin production associated with Paralytic Shellfish Poisoning (PSP) were also observed in different Alexandrium species. Decrease in Fe leads to increased PSP toxin content, PSP toxicity per cell, and changes in PSP toxin profile of Alexandrium tamarense [209]. Interestingly, a gradient of Fe concentrations tested on A. catenella and A. minutum revealed highest toxin levels were observed not in the lowest Fe treatment tested but at the most optimal [197,212]. Comparison between the three studies show that the highest PSP toxin concentration per cell was observed at 1 nmol L-1 Fe for A. tamarense while 1000 nmol L-1 Fe for A. catenella [209,212]. Although the PSP toxin profile was elucidated for both strains, changes to the PSP toxin profile were observed only on A. minutum with an increase in GTX 1+4 and decrease in both STX and GTX 2+3 [197]. A. catenella was only able to produce GTX 1-4, with the ratios of these toxins being similar throughout Fe treatments [212]. These findings show the different requirements and sensitivities of varying Alexandrium species towards Fe. Elevated PSP toxin generation in low Fe conditions may serve as a stress response, with heightened toxin production serving as a method for alternative Fe storage through metal-ligand interactions, where the toxin itself acts as a ligand. However, the observed decrease in toxin production at high Fe levels can be attributed to toxicity effects exerted by Fe and the subsequent decrease in related physiological activities. Further studies are needed to pinpoint the exact mechanism on how Fe affects PSP toxin production.
Though we see that Fe affects the toxin production of some HABs species, this is not necessarily true for all. P. reticulatum, a known yessotoxin (YTX) producer, requires Fe for its production. However, cells subjected to increasing Fe levels have shown neither increase nor decrease in YTX production [213]. Congruent with other toxic HABs species, an increase in the production of domoic acid was observed for both Pseudo-nitzschia multiseries and P. australis under Fe-limited treatments [214]. In contrast, an argument on the importance of Fe in the production of domoic acid (DA) by Pseudo-nitzschia was raised due to positive correlations between increasing Fe concentrations and domoic acid production [221]. This is further evidenced by the Fe-deplete treatments that reveal comparatively lower DA. Although the two studies give contrasting results, this may be due to interspecific variability between different strains of the species. Consequently, it was shown by Sobrinho et al. [222] that both excessive Fe amendments and Fe-deficiency would lead to comparatively higher DA concentrations between treatments. These findings suggest that increases in DA production is mediated by either Fe-deficiency or Fe-mediated toxicity. Upregulated DA production under scarcity conditions may be a means of self-protection for Pseudo-nitzschia as observed in other nutrient-deplete experiments [223,224,225]. On the other hand, the surge of DA production under high Fe stress can also be seen as a survival response as evidenced by the chelating capabilities of DA towards several different metal [175,198,214].
Investigations of the relationship between toxin production and Fe concentrations reveal complexity and specificity of responses not only among species but specific strains within these species. Furthermore, repercussions of toxin production and concentration cascade towards higher trophic organisms. Understanding the influences of Fe in toxin production would help deepen understanding of HABs and how they affect other organisms in the field.
Comprehension of Fe concentrations in the field and its relationship to blooms is necessary in understanding the dynamics and triggers of HABs. Concentration of Fe in the environment has been established as one of the driving factors in progress and proliferation of HABs. Linkages between atmospheric transport of Fe and algal blooms have already been substantiated [226]. As an essential nutrient needed by planktonic blooms, drawdown of Fe to produce biomass has been observed in several studies as a negative correlation between biomass and dissolved Fe concentrations [198,215,227]. The lowest dissolved Fe concentrations were observed during peak biomass of a cyanobacterial HAB event [228]. Algal bloom sites showed reduced Fe concentrations, which includes the exchangeable/acid-soluble fraction, reducible fraction, and residual fraction as compared to sites without algal blooms [229]. However, no differences in patterns of the three Fe fractions were observed during algal bloom formation. These field studies demonstrate the necessity of Fe in the progress of these blooms.
Aside from the drawdown of Fe by phytoplankton as a requirement for their growth, massive algal blooms also alter the physical parameters of the environment in which they exist in. Thamdrup et al. [230] has outlined the effects of algal blooms to Fe concentrations as follows. Heterotrophic organisms devour the organic matter provided by the bloom which leads to anoxic conditions at the topmost layer in shallow coastal sediments. The lack of oxygen causes bacterial organisms to use Mn-oxide and Fe-oxide as oxidizing agents resulting in dissolution of Fe and Mn. Interactions of Fe and H2S during algal blooms also lead to control of available Fe during algal blooms [231]. Subsequent anaerobic experiments derived from algal bloom-simulated conditions prove the dissolution of metals from sediments, most notably Fe [80,232,233]. The indirect effects of HABs to Fe introduce a layer of complexity in the dynamics and interactions of the two. Further studies are needed to substantiate other possible indirect effects that may exist.
Utilization of toxins and other compounds produced by HABs are not only limited to their allelopathic effects against other phytoplankton but may also provide other environmental advantages for the organism. Domoic acid produced by several species of Pseudo-nitzschia is capable of chelating Fe and other metals [175]. This chelating mechanism of DA helps in the provision of bioavailable Fe and in the decrease of Cu concentrations to pre-toxic levels [175,214]. Though the mechanism of Fe chelation by domoic acid may reduce bioavailable Fe, several strains of Pseudo-nitzschia have been shown to perform high affinity Fe acquisition with the aid of Cu and domoic acid [198]. Microcystins, brevetoxins, okadaic acid, and pahayokoides were found to form complexes with Fe, suggesting the ability of HABS to control bioavailability of metals within their environment [201]. Aside from its toxicity, Microcystis aggregates in surface water which exacerbates damage it causes during blooms. This scum formation is assisted by microcystin production [234,235,236]. The consequence of Fe concentrations towards microcystin production described earlier spill over to scum formation during blooms, effectively heightening the threat posed by M. aeruginosa.
The dependence of HABs on Fe concentrations provides a possible means for the suppression of cyanobacterial blooms. The greater Fe requirement of cyanobacteria as compared to other algae proves perilous under low concentrations of Fe [162,204,237,238]. Additionally, the role of phosphorus itself and in conjunction with Fe in the trigger of cyanobacterial HABs has also been well established [228,239,240]. Several contrasting studies have described the use of Fe and phosphorus in the control of HABs through various mechanisms. Orihel et al. [241] showed through mesocosm experiments that additional Fe concentrations in eutrophic lakes lead to the reduced pore water P concentrations. Decreased P concentrations were then shown to be directly correlated to decreased phytoplankton and periphyton growth. In contrast, direct addition of Fe chelators to a eutrophic lake has been proposed to control cyanobacterial growth. This exploits the direct relationship between Fe and cyanobacterial growth [242]. Further evidence from Leung et al. [228] demonstrates the drawdown of dissolved Fe during peak growth times of cyanobacterial HABs indicating the necessity of Fe in the growth of cyanobacterial blooms. However, in the same paper, the role of increasing phosphorus concentrations was also attributed to the triggering of the bloom event. These studies suggest the complexity of bacterial blooms in the field and the control of nutrients that limit them.
The relationship between Fe and HABs have been described in terms of the metal effect on growth, senescence, and toxin production. The compounds produced by HABs, and algal bloom events affect the bioavailability of Fe which shows their two-way relationship. The universal need for Fe by phytoplankton reveals the possible use of the metal in the mitigation of HABs. Differences in the reaction of varying species towards Fe merit continuous research on the topic and sustained data gathering would fine tune our understanding of HAB dynamics.
4.2. Copper (Cu)
Copper has long been established as an essential nutrient for phytoplankton growth. The role of Cu as a metal center for plastocyanin and for other oxidative stress enzymes highlights the importance of the metal in the physiology of HABs. Several studies have shown correlations between growth of marine HABs species like Pseudo-nitzschia australis [243], Pseudo-nitzschia delicatissima [244], Alexandrium minutum [197], and Aureococcus anophagefferens [245] with adequate Cu concentrations. Furthermore, Cu has been observed to be a limiting agent for HAB occurrences in a hypereutrophic lake [246]. Under Cu starvation, multiple strategies are deployed for compensation and conservation of physiological functions. For some HABs, most of these strategies aim to preserve survival of the organism. The stabilization and maintenance of photosynthesis at the PSII step [247] and preference for pigment and lipid reconstruction [248] were observed for P. delicatissima. This alludes to be a compensation for the role of Cu in plastocyanin in the electron transfer chain of photosynthesis. Transcriptome modifications involving lipid pathways were observed for Chlamydomonas reinhardtii under Cu deficient stresses [249]. Cu as a nutrient is required by HABs for growth at the cellular level and must adjust when facing Cu starvation. However, the high reactivity of Cu dictates a need for optimal concentrations of this nutrient for it to be most effective. Toxicity occurs when concentrations exceed the threshold an organism can be exposed to.
The sensitivity of phytoplankton to high Cu concentrations is evidenced by decreasing growth population to increasing Cu levels. Marine and estuarine populations of Gymnodinium breve [250], A. minutum [197,251], Pseudo-nitzschia sp. [175,198], Pseudo-nitzchia multiseries [252,253], P. delicatissima [244,248], P. australis [214], Amphidinium carterae [254], Prorocentrum micans [254], Alexandrium catenella [255,256], Ostreopsis ovata [256], Cochlodinium polykrikoides [257,258,259], and Karenia brevis [201] have exhibited toxicity to high Cu concentrations. Furthermore, some freshwater HAB species such as A. anophagefferens [245], Anabaenopsis [260], Closterium ehrenbergii [261], Lyngbya wollei [262], and freshwater Microcystis aeruginosa [65,203,207,210,211,260,263,264,265] also show decreased growth in high Cu concentrations. Aside from its senescent capabilities, induction of temporary cyst formation was observed in both O. ovata and A. catenella upon exposure to high Cu concentrations [255,256].
Responses to varying Cu concentrations were different for toxin producing HAB species. Increasing Cu concentrations induced lower production of YTX by Protoceratium reticulatum along with decreased growth [243]. Instances wherein Cu concentrations did not significantly affect toxin production was observed in microcystin production by M. aeruginosa [207], gymnodimine production by Karenia selliformis [243], and PSP toxin production in A. catenella [256]. However, PSP toxin production in a different species, A. minutum, was found to be maximized at optimal Cu concentrations and both low- and high-concentration Cu treatments induced lower toxin production [197]. Increasing Cu concentration caused higher okadaic acid production in Prorocentrum lima [243] and palytoxin and related compounds in both Ostreopsis siamensis [243] and O. ovata [256]. This increase can also be seen in the production of domoic acid by Pseudo-nitzschia. This toxin is documented to form complexes with metal ions and is considered as a means of surviving Cu toxicity by several Pseudo-nitzschia species [175,198,252]. Furthermore, compounding requirements of silicate and Cu also lead to an increase in domoic acid production in Pseudo-nitzschia sp. [253]. Aside from domoic acid, other HAB-produced toxins such as microcystin, pahayokoides, brevetoxins, and okadaic acid were able to form complexes with Cu and other metal ions [201,266,267,268]. This further indicates the capability of numerous HAB species to control and mitigate metal toxicity in the environment. Aside from their toxins, dissolved organic matter (DOM) and dissolved organic carbon (DOC) produced by phytoplankton may also function as metal chelators in the environment through their organic ligands [15,137,269]. DOC production by M. aeruginosa [202] and A. minutum [255] was enhanced under moderate Cu exposure. Although the DOC may play a role in decreasing Cu toxicity, it is highlighted in both studies that the highest concentrations of Cu used, exhibited total inhibition for both the organisms. This suggests that production of DOC cannot entirely negate effects of Cu toxicity. HAB species may utilize this characteristic of the toxins they produce to chelate metals in their environment, alleviating effects of toxic metals like Cu [214].
Consequences of Cu and its toxicity towards phytoplankton at high concentrations has paved the way for its possible use as an algaecide in the control of HABs. M. aeruginosa was determined to be sensitive to high concentrations of Cu [270]. Exposure to high doses of Cu was seen with cell lysis related release of K+ ions, increased reactive oxygen species production, increased superoxide dismutase and catalase activity, and upregulation of H2O2 treatment genes that are highly associated with oxidative stress [263,265]. Consequently, several studies have already outlined the use of copper sulfate as a means to control HAB blooms of M. aeruginosa [202,203,260,271,272]. This mitigative measure was also applied to other HAB-causing organisms. Reduced photosynthetic ability and increases in reactive oxygen species were perceived in both C. ehrenbergii [261] and C. polykrikoides [258]. Downregulation of photosystem genes and upregulation of mitochondrial genes and other genes that are involved in translation, spliceosome, and/or signal transduction were also further identified in C. polykrikoides [259]. Blooms of Anabaenopsis were also deemed susceptible to Cu-algicide treatments [203]. In the mat forming Lyngbya wollei, decreased filament viability was discovered upon exposure to high Cu concentrations [262] countering the mechanism in which L. wollei usually causes harm to the environment.
Description of the roles of Cu as both an essential and extremely toxic metal help us understand its unique dynamics with HABs. Studies regarding its use as an algicide is plentiful and provides us useful information on ways to mitigate these deleterious events. Although several papers have already described the effects of Cu starvation, its role as an essential nutrient should be further explored.
4.3. Zinc (Zn)
Zinc has been widely regarded as an important micronutrient due to its function in metalloenzymes for photosynthetic (carbonic anhydrase), translational (RNA polymerase, tRNA synthetase, reverse transcriptase) and antioxidative activities (superoxide dismutase, alcohol dehydrogenase) (Table 1). Zn is observed to be a necessary component in the growth of both freshwater HABs species such as Aureococcus anophagefferens [245], Microcystis aeruginosa [207], and marine HABs like Pseudo-nitzschia [243], Karenia selliformis [243], and Protoceratium reticulatum [243].
Although many physiological functions provided by Zn promote growth and relief from oxidative stress, high concentrations of Zn may still cause toxicity to organisms. In M. aeruginosa, high concentrations of Zn inhibited its growth [273]. This is further substantiated by an increase in dissolved organic carbon production [65], increase in toxin production [207], and upregulation of genes responsible for the production of microcystin [274]. Dissolved organic carbon and microcystins are capable of metal chelation - this adaptation of the organism is exhibited to decrease toxicity possibly exerted by Zn [214,255]. Synergistic effects of Zn and Cu [275] and Zn and Cd [276] toward decrease in cell division and photosynthetic activity were also described. Adaptations for oxidative stress response such as enhanced esterase and superoxide dismutase activity and increase in malondialdehyde content were noted as a response to Zn toxicity [276].
Marine HAB species also exhibited Zn stress-specific responses as an adaptation to toxicity. Increase in the domoic acid isomer-C production was observed in Pseudo-nitzschia under optimal Zn conditions [243]. Alternatively, elevated Zn levels decreased domoic acid production, which is contrasting to responses by Pseudo-nitzschia under the stress toxicity by other metal ions [248,252,253]. Decrease in toxin production was also observed for P. reticulatum [243].
Interestingly, Alexandrium minutum was observed to increase release in fluorescent dissolved organic matter in response to moderate Zn concentrations [277]. Fluorescent dissolved organic matter has also been documented to be capable of metal chelation, which was seen as a response to metal toxicity. Ultimately, Zn levels above threshold still lead to toxicity as evidenced by the formation of temporary cysts. Inspection of protein components under Zn toxicity in Alexandrium catenella reveals that Zn toxicity targets photosynthetic (Rubisco, peridinin chlorophyll-A, and ferredoxin NADP reductase), cellular signaling (calmodulin), and antioxidative processes (superoxide dismutases) [278]. Zn concentrations in the environment are also greatly affected during algal bloom development, duration, and decay. It has been established that drawdown of Zn by phytoplankton causes reduced concentrations during the onset of an algal bloom [153,232,279]. PCA component analysis suggests careful reconsideration of the relationship between Zn and algal bloom formation [280]. Inconsistent concentrations of Zn during onset are proof of other underlying mechanisms that govern over environmental Zn concentrations. A possible substitution of Co instead of Zn in carbonic anhydrase [96] may diminish the need for Zn during algal blooms. Subsequently, the decay of blooms reintroduces Zn into the system via reductive dissolution of Mn oxides in sediments [232,280]. This is supported by positive correlation of Zn concentrations and bloom-related DOC levels [281].
Much like Cu, several studies have already detailed the distinctive capabilities of Zn as both an essential nutrient and a toxic metal. Optimal concentrations reveal the benefits Zn poses towards HABs and their toxin production. However, utilization and concentrations of the metal during blooms remain poorly understood. Subsequent studies should be established to further divulge the relationship of Zn and HABs and their interactions.
4.4. Selenium (Se)
The biochemistry of Se has piqued interest due to intrinsic requirements by diverse organisms in different trophic levels. Phytoplankton necessitates an optimum Se concentration to support growth and this requirement is attributed to the production of Se-containing proteins [282]. There is limited work done on the investigations of the interdependence of Se availability and phytoplankton growth. The limited reports, however, still clearly present effects by Se on algal growth and dynamics of phytoplankton blooms [283,284,285,286]. Two pioneering works done on Gymnodinium nagasakiense, a dinoflagellate species persistent in Japanese waters, reflected good correlation between Se concentrations backed by enhanced cell growth when bay waters were amended with Se [283,284]. Subsequent work on other organisms as detailed in a review by Araie & Shiraiwa [282] (and references therein), show the relevance of Se but highlighted the need for further studies to better understand the mechanisms and bring to light its biological relevance.
The complexity of Se biochemistry in microalgae is depicted in the study on the intraspecific variabilities of Gymnodinium catenatum [285]. Se exists in nature with different isotopes and speciation ranging from the inorganic (selenite and selenate) to organic forms including selenomethionine, selenocysteine, and other selenoproteins. Phytoplankton preferentially takes up selenite, Se(IV), over selenate, Se(VI), using an ATP-dependent high affinity transporter that functions as an active transport system [282,286,287,288,289]. In the study by Doblin et al. [285], five strains from multiple origins were subjected to Se enrichment experiments and the physiological responses of Se-treated strains were compared with non-Se amended treatments. The results show variability in the growth performances of strains collected from different years and localities. These findings underscore the discrepancies in Se requirements, which is tempting to use to infer on the possible requirements for other trace metals as well. But what this clearly shows is that the Se requirement of different bloom-forming organisms needs to be further investigated.
Se is introduced to the environment through natural and anthropogenic sources with remarkably low concentrations. In open waters, selenite concentrations often range between 0.1 to 0.2 nM while selenate concentrations may be from 0.1 to 1.0 nM [290]. The narrow concentration ranges at which Se occur in aquatic environments may have dictated the window of essentiality of the metal for phytoplankton use because photosynthetic organisms need an optimum selenite supply at about 1 nM, beyond which Se begins to exert toxic effects on the organism [288]. Se is notable among trace metals because of this narrow usability for biological functions. The natural distribution of Se and its biogeochemical cycling in open waters is heavily influenced by phytoplankton because of preferential uptake of selenite, reduction and incorporation into proteins inside the cells of primary producers, and the complex regeneration from organic to inorganic Se compounds during algal decomposition [290]. In the many steps of Se transformations performed by microalgae, the role of excreted compounds comprising 80% of dissolved Se in the ocean is poorly characterized [291,292,293].
In terms of Se requirement by HAB-forming organisms, an excellent study organism is Aureococcus anophagefferens because it has the distinction of having the largest and most diverse known selenoproteome [289]. In ecophysiological studies conducted on the harmful pelagophyte that included cell cultures, genomic analyses, and ecosystem studies, it was demonstrated that dissolved Se were elevated before and after A. anophagefferens blooms but were significantly reduced during mid-bloom at about 0.05 nM. Validation of this observation was done using enrichment experiments by spiking seawater during the different stages of the bloom of the pelagophyte. The results show positive impacts of Se enrichment only in seawater during mid-bloom indicating Se limitation for the organisms at this stage. The results exemplify the role of Se in brown blooms and puts premium in the conduct of complementary experiments to provide a holistic understanding of interactions of environmental parameters with ecological responses of critically important organisms.
4.5. Other Essentials (Mn, Co, Ni, Cd, Mo, V)
The crucial roles of other essential metals on algal physiology are recognized but the current knowledge on their influence on HABs are quite scarce. In this section, we delve into the uses of other essential metals by HAB-causing species.
4.5.1. Manganese (Mn)
The major use for Mn is primarily in photosynthesis, respiration, and antioxidative defense mechanisms with MnSOD, a major reactive oxygen species detoxifying enzyme. With these functions, Mn affects growth rates, biomass production, and ultimately bloom dynamics of HAB species [294]. Mn enrichment experiments conducted by Rhodes et al. [243] reported increases in biomass of marine HAB species namely Ostreopsis siamensis, Karenia selliformis, and Prorocentrum lima. Interestingly, despite the limited known metabolic functions of Mn (Table 1), a significant decrease in toxin production–isodomoic acid and okadaic and diol esters of marine Pseudo-nitzchia australis and P. lima, respectively, has been reported [243]. Similar trends were also observed in freshwater Microcystis aeruginosa, where Mn depletion results in limited growth [211]. However, Lukač & Aegerter [207] reported that Mn does not significantly affect toxin yields, indicating a need for additional research in this area.
4.5.2. Cobalt (Co)
Similarly, studies regarding the role of Co in HABs are relatively limited and it was previously reported that the need for Co-containing cofactors is low in phytoplankton [193]. Early investigations conducted are focused solely on the effects of Co on growth of a bloom forming Prymnesiophycean flagellate, Chrysochromulina polylepis [295,296]. The underlying biochemical basis remained largely unknown [162,193,297] until succeeding studies have shed light on the metabolic role of Co and its potential influence on HAB formation. Co serves as an integral component of cobalamin or vitamin B12, an essential micronutrient for enzymes involved in N2 fixation [298]. It also assists methionine synthase in DNA synthesis and methylmalonyl CoA, which is required for inorganic C assimilation [299]. Extensive research on various marine microalgae have revealed that many HAB species including Aureococcus anophagefferens, Prorocentrum minimum, Karenia mikimotoi, Chatonella marina, Fibrocasa japonica, among others, exhibit auxotrophy for vitamin B, particularly B12, resulting in elevated vitamin B quotas [300]. These findings demonstrate the potential ecological significance of B-vitamins, including B1 and B7, in the regulation of HABs. Moreover, while Co is required in trace amounts, its availability can influence growth and proliferation of certain algal species [182]. Co enrichment experiments have shown increased cell densities in marine HAB species, Pseudo-nitzchia australis, Karenia selliformis, Ostreopsis siamensis, and Prorocentrum reticulatum [243] with a general trend of decreasing toxin production e.g., yessotoxins, gymnodimine, and palytoxins. However, Mitrovic et al. [213] observed no significant differences in growth and yessotoxin production of P. reticulatum, further suggesting that Co has species-specific influences on algal growth.
4.5.3. Nickel (Ni)
Ni has been considered as a common contaminant for aquatic ecosystems [301] and in fact, some algal bloom events were linked to Ni concentrations [153,302]. The known function of Ni is for nitrogen assimilation in the form of urease [303] and oxidative defense through the action of Ni-SOD [304,305,306]. Laboratory investigations revealed that Ni additions result in elevated growth of Aureococcus anophagefferens [245] and it is consistent with the genomic study on the same alga [305]. About 82% of cellular Ni in A. anophagefferens was sequestered through surface adsorption resulting in internalization of Ni, directly affecting metabolism and cell division, and thus potentially contributing to the persistence of brown tide development [245]. Other research on the influence of Ni on HABs demonstrates varying effects of Ni availability on the species under investigation. Ni addition starting from 3 μg L-1 Ni affected the growth of freshwater Mirocystis aeruginosa, with complete inhibition observed at 24 μg L-1 Ni [307]. Moreover, Ni exposure led to an increase in macromolecule production, activities of antioxidant biomarkers, e.g., catalase (CAT), glutathione peroxidase (GPx), superoxide dismutase (SOD), and toxic microcystin, which is in contrast with a previous report [207]. Nonetheless, these studies provide evidence for the link between Ni supply and algal bloom development.
4.5.4. Cadmium (Cd)
Cd is one of few metals that are widely regarded as non-essential and toxic owing to health effects associated with long-term exposure to it. Earlier studies on diatoms [165,168,193] and other phytoplankton species [308,309] have otherwise shown that Cd may also promote algal growth via metal substitutions when bioavailability of essential Zn is limited or depleted. A couple of reported bloom events were associated with Cd contamination [153,264,310]. Dinoflagellate HAB species including Chatonella marina, Chatonella ovata, Gymnodinium catenatum, and Gymnodinium sanguineum were found to be the culprits of massive mollusks and fish kill events in Kun Kaak Bay, Mexico, that coincided with high sediment Cd concentrations reaching up to 4 μg g-1 [310] from potential metal leaching [295,311]. Another HAB-forming dinoflagellate, Lingulodinium polyedrum, was found to be the dominant species of the HAB events in Todos Santos Bay, Mexico coinciding with high average Cd concentration (0.37 ± 0.41 to 1.02 ± 0.99 nmol kg-1 [312]. Tolerance to Cd levels has been reported in A. anophagefferens up to pCd values of 10.33 (defined as pCd = - log [Cd2+]) but further increases in Cd results in growth inhibition relative to the control: 21% (p = 0.001) and 67% (p < 0.001) at pCd 9.33 and 8.33, respectively (Wang et al., 2012). Similar sensitivity to Cd was also observed in another marine dinoflagellate, Alexandrium catenella, in which drastic growth inhibition was observed starting from pCd 7.88 (30%) up to pCd 6.36 (100%), where growth is completely inhibited [255]. Cyst formation was observed in pCd value of 7.88 and A. catenella released extracellular dissolved organic matter (DOM) as a defense mechanism to Cd toxicity at pCd 6.36. Other phytoplankton species have been reported to employ similar strategies via release of chelating molecules to protect cells from metals [313,314,315]. Interestingly, in freshwater M. aeruginosa, Cd exhibited no apparent negative effects on growth [253] as well as on microcystin production [207]. Moreover, increased floating rates and buoyancy of M. aeruginosa at high Cd concentrations provide insights on its potential contribution to facilitate development of cyanobacterial blooms, highlighting the need for further research on the role of Cd in algal blooms [264].
4.5.5. Molybdenum (Mo) and Vanadium (V)
In phytoplankton metal physiology, both Mo and V are understudied. Nonetheless, growing concerns regarding contamination from Mo and V due to extensive industrial activities have sparked renewed interest in studying them in aquatic ecosystems [316,317,318]. Previous research has shown that Mo serves as a constituent of nitrate reductase and nitrogenase enzymes, participating in nitrate assimilation and N2 fixation [319]. Likewise, V has been identified as essential nutrient influencing photosynthesis [320,321,322], cell division [323], nitrogenase and nitrate reductase activities [324] as well as bromoperoxidase enzymes containing V at the active site [163,164]. The existing knowledge base about Mo and V mostly come from research on freshwater green algae, blue-green algae [325,326], and some diatoms [322] because the major Mo species, MoO42−, is known to be generally less available in seawater than in freshwater [327].
Limited information is available concerning the involvement of Mo and V in HABs, although efforts to expand our current understanding have been made. Barros et al. [328] have explored the potential influence of molybdate (MoO42 −) to sulfate (SO42-) ratio on redox metabolism and viability of marine dinoflagellate Lingulodium polyedrum. Research revealed augmented xanthine oxidase (XO) and nitrate reductase (NR) activities, lipid peroxidation, protein carbonylation, as well as photosynthetic protein synthesis as a response to elevated [Mo]:[sulfate] ratios. These observations suggest that increasing [Mo]:[sulfate] ratios impose gradual oxidative stress conditions to L. polyedrum cultures. Moreover, Facey et al. [211] explored the role of Mo on growth and toxin production of freshwater M. aeruginosa, however, the reported results show no notable influence on growth and microcystin production. Meanwhile, studies on V in aquatic environments are mostly focused on redox geochemistry and mobility providing limited evidence on linkage between V and HABs [329]. A better understanding of redox metabolism in dinoflagellates as well as physiological functions of Mo and V in both marine and freshwater HABs could better inform on the roles of these metals in mitigating the impacts of HABs.
4.6. Non-Essentials (Pb, Hg, As, Sn, Ti, Zr)
Since HABs do not utilize non-essential metals, the major focus of encountering them is addressing the toxicity associated with these metals. The tolerance, survival, and adaptive mechanism of phytoplankton and HAB species are the aspects commonly investigated in the relationship of HABs and non-essential metals. Although the available data is considerably limited, this section will discuss HABs and their varying interactions with non-essential metals.
4.6.1. Lead (Pb)
Anthropogenic activities have been the primary reason for increased Pb contamination in both freshwater and oceanic ecosystems [330,331,332]. Pb is a nonessential element and toxicity has been documented for this metal. Photosynthetic systems, antioxidative activities, and changes in extracellular morphology are some of the primary responses of phytoplankton with regards to Pb stress [333]. The marine HAB species Alexandrium catenella was observed to have reduced protein expression, especially those for photosynthetic functions (RUBISCO and Ferredoxin-NADP+ reductase) and oxidative stress management (superoxide dismutase) when exposed to Pb [278]. In contrast, ATP synthase was upregulated indicating that this may serve as a survival mechanism to cope with metal stress induced by Pb. For Cochlodinum polykrikoides, tolerance was observed in lower Pb concentrations, but total inhibition was demonstrated in higher concentrations [257]. In the freshwater HAB species Microcystis aeruginosa, high Pb concentrations with accompanying high pH conditions result in decreased cell division and growth inhibition [264]. Additionally, chlorophyll content and cell fluorescence in M. aeruginosa were also reported to decrease under elevated Pb concentrations [334]. However, the same study has shown tolerance to high Pb levels and a corresponding accumulation of intracellular Pb.
Sun et al., [233] described the mobilization of Pb from sediments during an algal bloom in the following steps: (1) algal blooms create anaerobic environments from its increased biomass, (2) the resulting reductive conditions causes the solubilization of Fe/Mn oxides, which induces a chemically mediated release of Pb, (3) bloom-decay derived dissolved organic matter complexes Pb effectively decreasing dissolved Pb concentrations, (4) the lower Pb levels allow for algal growth and subsequent bloom, and eventually (5) the collapse of the algal bloom creates anoxic conditions and the cycle repeats again. The effects of Pb towards the growth of HAB species and the development of blooms underscore its importance in understanding HAB dynamics. Underlying mechanisms governing the cycling of Pb in the context of algal blooms need to be further investigated. The adaptability to elevated Pb levels poses concerns of trophic transfer to higher organisms owing to phytoplankton serving as the base of the food chain and with the known biomagnification potential of Pb.
4.6.2. Mercury (Hg)
Mercury is a common pollutant and is most toxic in its organic form. In phytoplankton, Hg will bind to cytosolic ligands and distribute into the organelles [335] wherein it may affect photosynthesis [336,337,338] and increase reactive oxygen species inside the cell [339,340]. As a consequence of toxicity, phytoplankton has devised mechanisms to counter this toxicity, which includes the conversion of ionic mercury to elemental mercury [341]. Even with this mechanism, several other factors may also affect the Hg composition during blooms. Unfortunately, increases in monomethyl mercury (MMHg) percentages were observed during an algal bloom in a high-altitude lake [342]. This increase in methylmercury may lead to cascading effects up the trophic ladder since this form can cross lipid bilayers and the blood-brain barrier. Transfer and bioaccumulation of methylmercury to organisms threaten organisms at the top of the food chain. Pickhardt et al. [343] demonstrated that increasing algal density reduces the amount of methylmercury found in phytoplankton by 2-3 folds. MMHg levels may have been diluted due to the increasing number of organisms that would uptake the metal. This denotes lesser Hg accumulation in fish in algal bloom environments as compared to algal bloom-free environments. The interaction between mercury and algal blooms remains relatively understudied, future studies should fortify our understanding of the cascading effects of the two in food webs.
4.6.3. Arsenic (As)
Arsenic is a highly toxic carcinogenic metal found ubiquitously in rocks and sediments exposed to bodies of water [344]. It is known to be toxic to humans and other organisms and has no utilization in physiological functions [345,346]. Mechanisms of toxicity for As revolves around arsenate (V) and phosphorus being very biochemically similar [347,348,349]. Arsenate (V) interferes with energy metabolism with its substitution in the adenotriphosphate cycle. This was clearly exhibited by the HAB species Chlamydomonas reinhardtii, wherein As toxicity was observed to be characteristically similar to phosphate deficiency [345]. Mechanistic adaption involving As employed by phytoplankton centers around the conversion of As(V) to As(III), and monomethyl and dimethyl arsenic species [350,351,352]. As such, studies regarding As and algal blooms revolve mainly on As species transformation and correlation studies with phosphorus concentrations.
During bloom development, arsenate was rapidly reduced to arsenite and other methylated species [351]. The rate of reduction depended on pre-existing As concentrations, dominant phytoplankton, season, and the degree of decline in phosphorus concentrations. Contrary to the onset of algal blooms, the decay stage of the event aggravates the release of As and other labile metals from sediments. Strong anoxic conditions [353], reduction of Fe and Mn oxides [240], and microbial action [354] were observed to cause an increase in As concentrations during algal bloom decay. Released Fe-sulfides from the decaying bloom facilitated As reduction, which was then mobilized to the aqueous phase [240,353]. Aside from the conversion of As(V) to As(III), a strain of M. aeruginosa isolated from a lake with naturally high As concentration was found to be resistant to As concentrations of 10-5 mol/L As(III) and 10-3 mol/L As(V) [355]. This observation was accompanied by increased total microcystin production under low As concentrations. The role of algal blooms in the cycling and transformation of As to its various forms remains relatively understudied. Furthermore, synergistic effects of toxic algal blooms and As poisoning may be on the horizon as adaptations to As toxicity may occur.
4.6.4. Tin (Sn), Titanium (Ti), and Zirconium (Zr)
Interests in the effects of other non-essential metals have recently been put in the limelight. The group IV metals tin, titanium, and zirconium are relatively understudied in terms of biological relevance, but these metals have interesting properties. The possibility of group IV metals to incur changes in phytoplankton has been exhibited in the case of germanium (Ge), which is able to modify the cell shape of silica-rich diatoms [356]. This modification may have been achieved by Ge by mimicking the chemical properties of Si and replacing it in cellular structures in the phytoplankton. This ability to stimulate structural changes was also caused by Sn, Ti, and Zr in the freshwater diatom, Synedra acus [357]. Aside from morphological changes these metals can induce, their inhibitory properties towards phytoplankton are also demonstrated. Zr salts were able to reduce phytoplankton populations through inactivation of phosphorous [358,359]. Chloride salts of Sn (II) and Sn(IV) exhibited toxicity towards Synechocystis [360]. Furthermore, Titanium dioxide (TiO2) nanoparticles have shown remarkable toxicity to natural phytoplankton assemblages [361]. Reductions on the maximal photosystem II quantum yields and induction of oxidative stress were observed upon introduction to TiO2 [362]. Nano-TiO2 particles were also described to induce changes in phytoplanktonic percent composition denoting species-specific responses towards the nanoparticle [363]. Though studies focusing on these metals and algal blooms are lacking, increasing evidence of interactive effects merits further studies to be conducted on these metals.
5. Gaps and Recommendations
The current knowledge base on phytoplankton physiology primarily comes from research conducted on larger diatoms and cyanobacterial diazotrophs. Over the past decades, investigations have expanded to other phytoplankton functional types including dinoflagellates, picoeukaryotes, green algae, non-diazotrophic cyanobacteria, and others, shedding some light on the roles of trace metals on dynamics of HABs. However, variations in trace metal requirements, modes of nutrient uptake and assimilation, redox metabolisms, and toxin production calls for further investigations on the role of trace metals as well as vitamins on phytoplankton growth, metabolic processes, bloom dynamics and other ecosystem-level interactions. Studies describing the effects of trace elements towards HABs and their toxin production leave much to be desired. A general focus of current studies hinges on the toxicity of certain essential nutrients such as Cu and Zn when supply is in excess. Detailing the role of metals towards growth and toxin production under deficient conditions demonstrative of actual scenarios in coastal environments prone to HAB occurrences would provide new insights into the importance of metals in these processes.
The initiation and dynamics of phytoplankton blooms will be further complicated in the scenario of the future ocean with acidification and global warming looming to cause complex and interdependent responses in phytoplankton and their growth factors. Trace metal cycling and availability will be influenced since solubility and reactivity of metals are controlled by pH and temperature changes. Pinpointing specific responses of phytoplankton groups to these environmental changes will require carefully-designed experiments that may use complementary and orthogonal studies for holistic and cohesive understanding that will permit inference of projected ecological changes of algal blooms. Nowadays, omics strategies are more readily accessible and field- or laboratory-derived physiological and physicochemical information backed with genomics or transcriptomics data will be more valuable and provide more definitive insight into underlying processes that we wish to bring to light. To address the paucity in data about the role of metals in phytoplankton growth, structure, and community succession, more research is needed especially focusing on algal species of concern. In the context of HABs, further trace metal research should consider environmentally-relevant amendments when conducting metal enrichment work and conduct of field-based studies should be guided by timing and frequency to capture information that is illustrative of ambient conditions. It is expected that expanding agriculture and aquaculture activities will continue to exert pressure and shape ecological niches in coastal environments. It is then imperative that studies incorporate these considerations when putting forward plans for monitoring bloom occurrences so that sampling designs will factor in spatial, temporal, and seasonal variabilities for more accurate data, especially when these will be inputted into predictive models. More importantly, data gathered on the interactive effects of trace metals and other nutrients on harmful algal blooms should be communicated to stakeholders so that translation into policies and everyday practices will be implemented.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Acknowledgments
We express our gratitude to the University of the Philippines System, the Marine Science Institute, Department of Science and Technology (DOST), Philippine Council for Agriculture Aquatic and Natural Resource Research and Development (DOST-PCAARD), and the RVA-Marine Microalgal Culture Collection (RVA-MMCC) for their support to our research group.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Anderson, D. HABs in a Changing World: A Perspective on Harmful Algal Blooms, Their Impacts, and Research and Management in a Dynamic Era of Climactic and Environmental Change. Harmful Algae 2012 Proc. 15th Int. Conf. Harmful Algae Oct. 29 - Novemb. 2 2012 CECO Chang. Gyeongnam Korea Int. Conf. Harmful Algae 15th 2012 Chang. Gyeongnam Kore 2014, 2012, 3–17. [Google Scholar]
- Anderson, C.R.; Moore, S.K.; Tomlinson, M.C.; Silke, J.; Cusack, C.K. Living with Harmful Algal Blooms in a Changing World. In Coastal and Marine Hazards, Risks, and Disasters; Elsevier, 2015; pp. 495–561 ISBN 978-0-12-396483-0.
- Hallegraeff, G.M.; Anderson, D.M.; Belin, C.; Bottein, M.-Y.D.; Bresnan, E.; Chinain, M.; Enevoldsen, H.; Iwataki, M.; Karlson, B.; McKenzie, C.H.; et al. Perceived Global Increase in Algal Blooms Is Attributable to Intensified Monitoring and Emerging Bloom Impacts. Commun. Earth Environ. 2021, 2, 117. [Google Scholar] [CrossRef]
- Hallegraeff, G.M.; Anderson, D.M. ; Cembella Manual on Harmful Marine Microalgae; 2nd ed.; UNESCO Publishing: Landais, France, 2004; ISBN 92-3-103948-2. [Google Scholar]
- Anderson, D.M.; Burkholder, J.M.; Cochlan, W.P.; Glibert, P.M.; Gobler, C.J.; Heil, C.A.; Kudela, R.M.; Parsons, M.L.; Rensel, J.E.J.; Townsend, D.W.; et al. Harmful Algal Blooms and Eutrophication: Examining Linkages from Selected Coastal Regions of the United States. Harmful Algae 2008, 8, 39–53. [Google Scholar] [CrossRef] [PubMed]
- Azanza, R.V.; Fukuyo, Y.; Yap, L.G.; Takayama, H. Prorocentrum Minimum Bloom and Its Possible Link to a Massive Fish Kill in Bolinao, Pangasinan, Northern Philippines. Harmful Algae 2005, 4, 519–524. [Google Scholar] [CrossRef]
- Paerl, H.W. Assessing and Managing Nutrient-Enhanced Eutrophication in Estuarine and Coastal Waters: Interactive Effects of Human and Climatic Perturbations. Ecol. Eng. 2006, 26, 40–54. [Google Scholar] [CrossRef]
- Paerl, H.W.; Havens, K.E.; Xu, H.; Zhu, G.; McCarthy, M.J.; Newell, S.E.; Scott, J.T.; Hall, N.S.; Otten, T.G.; Qin, B. Mitigating Eutrophication and Toxic Cyanobacterial Blooms in Large Lakes: The Evolution of a Dual Nutrient (N and P) Reduction Paradigm. Hydrobiologia 2020, 847, 4359–4375. [Google Scholar] [CrossRef]
- Sellner, K.G.; Doucette, G.J.; Kirkpatrick, G.J. Harmful Algal Blooms: Causes, Impacts and Detection. J. Ind. Microbiol. Biotechnol. 2003, 30, 383–406. [Google Scholar] [CrossRef]
- Brown, A.R.; Lilley, M.; Shutler, J.; Lowe, C.; Artioli, Y.; Torres, R.; Berdalet, E.; Tyler, C.R. Assessing Risks and Mitigating Impacts of Harmful Algal Blooms on Mariculture and Marine Fisheries. Rev. Aquac. 2020, 12, 1663–1688. [Google Scholar] [CrossRef]
- Heil, C.A.; Muni-Morgan, A.L. Florida’s Harmful Algal Bloom (HAB) Problem: Escalating Risks to Human, Environmental and Economic Health With Climate Change. Front. Ecol. Evol. 2021, 9, 646080. [Google Scholar] [CrossRef]
- Shirokova, L.S.; Kunhel, L.; Rols, J.-L.; Pokrovsky, O.S. Experimental Modeling of Cyanobacterial Bloom in a Thermokarst Lake: Fate of Organic Carbon, Trace Metal, and Carbon Sequestration Potential. Aquat. Geochem. 2015, 21, 487–511. [Google Scholar] [CrossRef]
- Kim, J.S.; Seo, I.W.; Baek, D. Modeling Spatial Variability of Harmful Algal Bloom in Regulated Rivers Using a Depth-Averaged 2D Numerical Model. J. Hydro-Environ. Res. 2018, 20, 63–76. [Google Scholar] [CrossRef]
- Ralston, D.K.; Moore, S.K. Modeling Harmful Algal Blooms in a Changing Climate. Harmful Algae 2020, 91, 101729. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Processes Regulating Cellular Metal Accumulation and Physiological Effects: Phytoplankton as Model Systems. Sci. Total Environ. 1998, 219, 165–181. [Google Scholar] [CrossRef]
- Gerringa, L.J.A.; De Baar, H.J.W.; Timmermans, K.R. A Comparison of Iron Limitation of Phytoplankton in Natural Oceanic Waters and Laboratory Media Conditioned with EDTA. Mar. Chem. 2000, 68, 335–346. [Google Scholar] [CrossRef]
- Firme, G.F.; Rue, E.L.; Weeks, D.A.; Bruland, K.W.; Hutchins, D.A. Spatial and Temporal Variability in Phytoplankton Iron Limitation along the California Coast and Consequences for Si, N, and C Biogeochemistry. Glob. Biogeochem. Cycles 2003, 17, 2001GB001824. [Google Scholar] [CrossRef]
- de Quiros, P.F. The Voyages of Pedro Fernadez de Quiros 1595 to 1606 (No. 14); Hakluyt Society: London, 1904. [Google Scholar]
- Koenigswald, W.V.; Braun, A.; Pfeiffer, T. Cyanobacteria and Seasonal Death: A New Taphonomic Model for the Eocene Messel Lake. Paläontol. Z. 2004, 78, 417–424. [Google Scholar] [CrossRef]
- Bargu, S.; Silver, M.W.; Ohman, M.D.; Benitez-Nelson, C.R.; Garrison, D.L. Mystery behind Hitchcock’s Birds. Nat. Geosci. 2012, 5, 2–3. [Google Scholar] [CrossRef]
- Smayda, T.J.; Villareal, T.A. The 1985 ‘Brown-Tide’ and the Open Phytoplankton Niche in Narragansett Bay During Summer. In Novel Phytoplankton Blooms; Cosper, E.M., Bricelj, V.M., Carpenter, E.J., Eds.; Springer Berlin Heidelberg: Berlin, Heidelberg, 1989; pp. 159–187. ISBN 978-3-642-75282-7. [Google Scholar]
- Hallegraeff, G.M. A Review of Harmful Algal Blooms and Their Apparent Global Increase. Phycologia 1993, 32, 79–99. [Google Scholar] [CrossRef]
- Anderson, D.M.; Fensin, E.; Gobler, C.J.; Hoeglund, A.E.; Hubbard, K.A.; Kulis, D.M.; Landsberg, J.H.; Lefebvre, K.A.; Provoost, P.; Richlen, M.L.; et al. Marine Harmful Algal Blooms (HABs) in the United States: History, Current Status and Future Trends. Harmful Algae 2021, 102, 101975. [Google Scholar] [CrossRef]
- Karlson, B.; Andersen, P.; Arneborg, L.; Cembella, A.; Eikrem, W.; John, U.; West, J.J.; Klemm, K.; Kobos, J.; Lehtinen, S.; et al. Harmful Algal Blooms and Their Effects in Coastal Seas of Northern Europe. Harmful Algae 2021, 102, 101989. [Google Scholar] [CrossRef]
- Yñiguez, A.T.; Lim, P.T.; Leaw, C.P.; Jipanin, S.J.; Iwataki, M.; Benico, G.; Azanza, R.V. Over 30 Years of HABs in the Philippines and Malaysia: What Have We Learned? Harmful Algae 2021, 102, 101776. [Google Scholar] [CrossRef] [PubMed]
- Li, X.-Y.; Yu, R.-C.; Richardson, A.J.; Sun, C.; Eriksen, R.; Kong, F.-Z.; Zhou, Z.-X.; Geng, H.-X.; Zhang, Q.-C.; Zhou, M.-J. Marked Shifts of Harmful Algal Blooms in the Bohai Sea Linked with Combined Impacts of Environmental Changes. Harmful Algae 2023, 121, 102370. [Google Scholar] [CrossRef]
- Paerl, H.W.; Justić, D. Primary Producers. In Treatise on Estuarine and Coastal Science; Elsevier, 2011; pp. 23–42 ISBN 978-0-08-087885-0.
- Herut, B.; Krom, M.D.; Pan, G.; Mortimer, R. Atmospheric Input of Nitrogen and Phosphorus to the Southeast Mediterranean: Sources, Fluxes, and Possible Impact. Limnol. Oceanogr. 1999, 44, 1683–1692. [Google Scholar] [CrossRef]
- Hamilton, D.P.; Salmaso, N.; Paerl, H.W. Mitigating Harmful Cyanobacterial Blooms: Strategies for Control of Nitrogen and Phosphorus Loads. Aquat. Ecol. 2016, 50, 351–366. [Google Scholar] [CrossRef]
- Paerl, H.W.; Huisman, J. Blooms Like It Hot. Science 2008, 320, 57–58. [Google Scholar] [CrossRef] [PubMed]
- Gobler, C.J.; Doherty, O.M.; Hattenrath-Lehmann, T.K.; Griffith, A.W.; Kang, Y.; Litaker, R.W. Ocean Warming since 1982 Has Expanded the Niche of Toxic Algal Blooms in the North Atlantic and North Pacific Oceans. Proc. Natl. Acad. Sci. 2017, 114, 4975–4980. [Google Scholar] [CrossRef] [PubMed]
- León-Muñoz, J.; Urbina, M.A.; Garreaud, R.; Iriarte, J.L. Hydroclimatic Conditions Trigger Record Harmful Algal Bloom in Western Patagonia (Summer 2016). Sci. Rep. 2018, 8, 1330. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.C.; Michalak, A.M. Exploring Temperature and Precipitation Impacts on Harmful Algal Blooms across Continental U.S. Lakes. Limnol. Oceanogr. 2020, 65, 992–1009. [Google Scholar] [CrossRef]
- Beardall, J.; Stojkovic, S.; Larsen, S. Living in a High CO 2 World: Impacts of Global Climate Change on Marine Phytoplankton. Plant Ecol. Divers. 2009, 2, 191–205. [Google Scholar] [CrossRef]
- Dutkiewicz, S.; Morris, J.J.; Follows, M.J.; Scott, J.; Levitan, O.; Dyhrman, S.T.; Berman-Frank, I. Impact of Ocean Acidification on the Structure of Future Phytoplankton Communities. Nat. Clim. Change 2015, 5, 1002–1006. [Google Scholar] [CrossRef]
- Flynn, K.J.; Clark, D.R.; Mitra, A.; Fabian, H.; Hansen, P.J.; Glibert, P.M.; Wheeler, G.L.; Stoecker, D.K.; Blackford, J.C.; Brownlee, C. Ocean Acidification with (de)Eutrophication Will Alter Future Phytoplankton Growth and Succession. Proc. R. Soc. B Biol. Sci. 2015, 282, 20142604. [Google Scholar] [CrossRef] [PubMed]
- Timmerman, A.H.V.; McManus, M.A.; Cheriton, O.M.; Cowen, R.K.; Greer, A.T.; Kudela, R.M.; Ruttenberg, K.; Sevadjian, J. Hidden Thin Layers of Toxic Diatoms in a Coastal Bay. Deep Sea Res. Part II Top. Stud. Oceanogr. 2014, 101, 129–140. [Google Scholar] [CrossRef]
- Raine, R.; Berdalet, E.; Yamazaki, H.; Jenkinson, I.; Reguera, B. Key Questions and Recent Research Advances on Harmful Algal Blooms in Stratified Systems. In Global Ecology and Oceanography of Harmful Algal Blooms; Glibert, P.M., Berdalet, E., Burford, M.A., Pitcher, G.C., Zhou, M., Eds.; Ecological Studies; Springer International Publishing: Cham, 2018; Vol. 232, pp. 165–186. ISBN 978-3-319-70068-7. [Google Scholar]
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A Meta-Analysis of Projected Global Food Demand and Population at Risk of Hunger for the Period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Bruinsma, J. World Agriculture: Towards 2015/2030 an FAO Perspective; Earthscan publications: London, 2003; ISBN 978-92-5-104835-1. [Google Scholar]
- FAO World Fertilizer Trends and Outlook to 2022; FAO: Rome, 2022.
- Glibert, P.M.; Harrison, J.; Heil, C.; Seitzinger, S. Escalating Worldwide Use of Urea – A Global Change Contributing to Coastal Eutrophication. Biogeochemistry 2006, 77, 441–463. [Google Scholar] [CrossRef]
- Yue, F.-J.; Li, S.-L.; Liu, C.-Q.; Zhao, Z.-Q.; Ding, H. Tracing Nitrate Sources with Dual Isotopes and Long Term Monitoring of Nitrogen Species in the Yellow River, China. Sci. Rep. 2017, 7, 8537. [Google Scholar] [CrossRef] [PubMed]
- Tamborski, J.; Brown, C.; Bokuniewicz, H.; Cochran, J.K.; Rasbury, E.T. Investigating Boron Isotopes for Identifying Nitrogen Sources Supplied by Submarine Groundwater Discharge to Coastal Waters. Front. Environ. Sci. 2020, 8, 126. [Google Scholar] [CrossRef]
- Sarma, V.V.S.S.; Krishna, M.S.; Srinivas, T.N.R. Sources of Organic Matter and Tracing of Nutrient Pollution in the Coastal Bay of Bengal. Mar. Pollut. Bull. 2020, 159, 111477. [Google Scholar] [CrossRef] [PubMed]
- Csathó, P.; Sisák, I.; Radimszky, L.; Lushaj, S.; Spiegel, H.; Nikolova, M.T.; Nikolov, N.; ČErmák, P.; Klir, J.; Astover, A.; et al. Agriculture as a Source of Phosphorus Causing Eutrophication in Central and Eastern Europe. Soil Use Manag. 2007, 23, 36–56. [Google Scholar] [CrossRef]
- Ishida, T.; Uehara, Y.; Iwata, T.; Cid-Andres, A.P.; Asano, S.; Ikeya, T.; Osaka, K.; Ide, J.; Privaldos, O.L.A.; Jesus, I.B.B.D.; et al. Identification of Phosphorus Sources in a Watershed Using a Phosphate Oxygen Isoscape Approach. Environ. Sci. Technol. 2019, 53, 4707–4716. [Google Scholar] [CrossRef]
- Oelsner, G.P.; Stets, E.G. Recent Trends in Nutrient and Sediment Loading to Coastal Areas of the Conterminous U.S.: Insights and Global Context. Sci. Total Environ. 2019, 654, 1225–1240. [Google Scholar] [CrossRef]
- Li, Q.; Yuan, H.; Li, H.; Main, C.; Anton, J.; Jaisi, D.P. Tracing the Sources of Phosphorus along the Salinity Gradient in a Coastal Estuary Using Multi-Isotope Proxies. Sci. Total Environ. 2021, 792, 148353. [Google Scholar] [CrossRef] [PubMed]
- Makkar, H.P.S. Review: Feed Demand Landscape and Implications of Food-Not Feed Strategy for Food Security and Climate Change. Animal 2018, 12, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Mottet, A.; De Haan, C.; Falcucci, A.; Tempio, G.; Opio, C.; Gerber, P. Livestock: On Our Plates or Eating at Our Table? A New Analysis of the Feed/Food Debate. Glob. Food Secur. 2017, 14, 1–8. [Google Scholar] [CrossRef]
- FAO Food and Agriculture Data Statistics (FAOSTAT); 2022;
- FAO The State of World Fisheries and Aquaculture: Sustainability in Action; 2020;
- Alonso-Rodrı́guez, R.; Páez-Osuna, F. Nutrients, Phytoplankton and Harmful Algal Blooms in Shrimp Ponds: A Review with Special Reference to the Situation in the Gulf of California. Aquaculture 2003, 219, 317–336. [Google Scholar] [CrossRef]
- San Diego-McGlone, M.L.; Azanza, R.V.; Villanoy, C.L.; Jacinto, G.S. Eutrophic Waters, Algal Bloom and Fish Kill in Fish Farming Areas in Bolinao, Pangasinan, Philippines. Mar. Pollut. Bull. 2008, 57, 295–301. [Google Scholar] [CrossRef] [PubMed]
- Yodsuwan, N.; Sawayama, S.; Sirisansaneeyakul, S. Effect of Nitrogen Concentration on Growth, Lipid Production and Fatty Acid Profiles of the Marine Diatom Phaeodactylum Tricornutum. Agric. Nat. Resour. 2017, 51, 190–197. [Google Scholar] [CrossRef]
- Reich, H.G.; Rodriguez, I.B.; LaJeunesse, T.C.; Ho, T.-Y. Endosymbiotic Dinoflagellates Pump Iron: Differences in Iron and Other Trace Metal Needs among the Symbiodiniaceae. Coral Reefs 2020, 39, 915–927. [Google Scholar] [CrossRef]
- Baek, S.H.; Shimode, S.; Han, M.-S.; Kikuchi, T. Growth of Dinoflagellates, Ceratium Furca and Ceratium Fusus in Sagami Bay, Japan: The Role of Nutrients. Harmful Algae 2008, 7, 729–739. [Google Scholar] [CrossRef]
- Longo, S.; Sibat, M.; Darius, H.T.; Hess, P.; Chinain, M. Effects of pH and Nutrients (Nitrogen) on Growth and Toxin Profile of the Ciguatera-Causing Dinoflagellate Gambierdiscus Polynesiensis (Dinophyceae). Toxins 2020, 12, 767. [Google Scholar] [CrossRef]
- Anderson, D.M.; Glibert, P.M.; Burkholder, J.M. Harmful Algal Blooms and Eutrophication: Nutrient Sources, Composition, and Consequences. Estuaries 2002, 25, 704–726. [Google Scholar] [CrossRef]
- Lagus, A. Species-Specific Differences in Phytoplankton Responses to N and P Enrichments and the N:P Ratio in the Archipelago Sea, Northern Baltic Sea. J. Plankton Res. 2004, 26, 779–798. [Google Scholar] [CrossRef]
- Li, Q.; Legendre, L.; Jiao, N. Phytoplankton Responses to Nitrogen and Iron Limitation in the Tropical and Subtropical Pacific Ocean. J. Plankton Res. 2015, 37, 306–319. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T. Throwing Fuel on the Fire: Synergistic Effects of Excessive Nitrogen Inputs and Global Warming on Harmful Algal Blooms. Environ. Sci. Technol. 2010, 44, 7756–7758. [Google Scholar] [CrossRef] [PubMed]
- Moutin, T.; Karl, D.M.; Duhamel, S.; Rimmelin, P.; Raimbault, P.; Van Mooy, B.A.S.; Claustre, H. Phosphate Availability and the Ultimate Control of New Nitrogen Input by Nitrogen Fixation in the Tropical Pacific Ocean. Biogeosciences 2008, 5, 95–109. [Google Scholar] [CrossRef]
- Wu, H.; Lin, L.; Shen, G.; Li, M. Heavy-Metal Pollution Alters Dissolved Organic Matter Released by Bloom-Forming Microcystis Aeruginosa. RSC Adv. 2017, 7, 18421–18427. [Google Scholar] [CrossRef]
- Moore, C.M.; Mills, M.M.; Langlois, R.; Milne, A.; Achterberg, E.P.; La Roche, J.; Geider, R.J. Relative Influence of Nitrogen and Phosphorous Availability on Phytoplankton Physiology and Productivity in the Oligotrophic Sub-tropical North Atlantic Ocean. Limnol. Oceanogr. 2008, 53, 291–305. [Google Scholar] [CrossRef]
- Piehler, M.F.; Twomey, L.J.; Hall, N.S.; Paerl, H.W. Impacts of Inorganic Nutrient Enrichment on Phytoplankton Community Structure and Function in Pamlico Sound, NC, USA. Estuar. Coast. Shelf Sci. 2004, 61, 197–209. [Google Scholar] [CrossRef]
- Burson, A.; Stomp, M.; Akil, L.; Brussaard, C.P.D.; Huisman, J. Unbalanced Reduction of Nutrient Loads Has Created an Offshore Gradient from Phosphorus to Nitrogen Limitation in the N Orth S Ea. Limnol. Oceanogr. 2016, 61, 869–888. [Google Scholar] [CrossRef]
- Song, Y.; Guo, Y.; Liu, H.; Zhang, G.; Zhang, X.; Thangaraj, S.; Sun, J. Water Quality Shifts the Dominant Phytoplankton Group from Diatoms to Dinoflagellates in the Coastal Ecosystem of the Bohai Bay. Mar. Pollut. Bull. 2022, 183, 114078. [Google Scholar] [CrossRef]
- Hernández-Sandoval, F.E.; Bustillos-Guzmán, J.J.; Band-Schmidt, C.J.; Núñez-Vázquez, E.J.; López-Cortés, D.J.; Fernández-Herrera, L.J.; Poot-Delgado, C.A.; Moreno-Legorreta, M. Effect of Different N:P Ratios on the Growth, Toxicity, and Toxin Profile of Gymnodinium Catenatum (Dinophyceae) Strains from the Gulf of California. Toxins 2022, 14, 501. [Google Scholar] [CrossRef]
- Leong, S.C.Y.; Murata, A.; Nagashima, Y.; Taguchi, S. Variability in Toxicity of the Dinoflagellate Alexandrium Tamarense in Response to Different Nitrogen Sources and Concentrations. Toxicon 2004, 43, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Hattenrath, T.K.; Anderson, D.M.; Gobler, C.J. The Influence of Anthropogenic Nitrogen Loading and Meteorological Conditions on the Dynamics and Toxicity of Alexandrium Fundyense Blooms in a New York (USA) Estuary. Harmful Algae 2010, 9, 402–412. [Google Scholar] [CrossRef]
- Hattenrath-Lehmann, T.K.; Marcoval, M.A.; Mittlesdorf, H.; Goleski, J.A.; Wang, Z.; Haynes, B.; Morton, S.L.; Gobler, C.J. Nitrogenous Nutrients Promote the Growth and Toxicity of Dinophysis Acuminata during Estuarine Bloom Events. PLOS ONE 2015, 10, e0124148. [Google Scholar] [CrossRef] [PubMed]
- 7Zhuang, Y.; Zhang, H.; Hannick, L.; Lin, S. Metatranscriptome Profiling Reveals Versatile N-Nutrient Utilization, CO2 Limitation, Oxidative Stress, and Active Toxin Production in an Alexandrium Fundyense Bloom. Harmful Algae 2015, 42, 60–70. [Google Scholar] [CrossRef]
- Barnard, M.A.; Chaffin, J.D.; Plaas, H.E.; Boyer, G.L.; Wei, B.; Wilhelm, S.W.; Rossignol, K.L.; Braddy, J.S.; Bullerjahn, G.S.; Bridgeman, T.B.; et al. Roles of Nutrient Limitation on Western Lake Erie CyanoHAB Toxin Production. Toxins 2021, 13, 47. [Google Scholar] [CrossRef] [PubMed]
- Wagner, N.D.; Quach, E.; Buscho, S.; Ricciardelli, A.; Kannan, A.; Naung, S.W.; Phillip, G.; Sheppard, B.; Ferguson, L.; Allen, A.; et al. Nitrogen Form, Concentration, and Micronutrient Availability Affect Microcystin Production in Cyanobacterial Blooms. Harmful Algae 2021, 103, 102002. [Google Scholar] [CrossRef] [PubMed]
- Cooper, G.M.; Adams, K.W. The Cell: A Molecular Approach; Ninth edition.; Oxford University Press: New York, NY, 2023; ISBN 978-0-19-758374-6. [Google Scholar]
- Falkowski, P.G.; Raven, J.A. Aquatic Photosynthesis; 2nd ed.; Princeton University Press: Princeton, 2007; ISBN 978-0-691-11550-4.
- Feng, T.-Y.; Yang, Z.-K.; Zheng, J.-W.; Xie, Y.; Li, D.-W.; Murugan, S.B.; Yang, W.-D.; Liu, J.-S.; Li, H.-Y. Examination of Metabolic Responses to Phosphorus Limitation via Proteomic Analyses in the Marine Diatom Phaeodactylum Tricornutum. Sci. Rep. 2015, 5, 10373. [Google Scholar] [CrossRef]
- Ding, S.; Chen, M.; Gong, M.; Fan, X.; Qin, B.; Xu, H.; Gao, S.; Jin, Z.; Tsang, D.C.W.; Zhang, C. Internal Phosphorus Loading from Sediments Causes Seasonal Nitrogen Limitation for Harmful Algal Blooms. Sci. Total Environ. 2018, 625, 872–884. [Google Scholar] [CrossRef]
- Lin, S.; Litaker, R.W.; Sunda, W.G. Phosphorus Physiological Ecology and Molecular Mechanisms in Marine Phytoplankton. J. Phycol. 2016, 52, 10–36. [Google Scholar] [CrossRef]
- Paerl, H.W.; Scott, J.T.; McCarthy, M.J.; Newell, S.E.; Gardner, W.S.; Havens, K.E.; Hoffman, D.K.; Wilhelm, S.W.; Wurtsbaugh, W.A. It Takes Two to Tango: When and Where Dual Nutrient (N & P) Reductions Are Needed to Protect Lakes and Downstream Ecosystems. Environ. Sci. Technol. 2016, 50, 10805–10813. [Google Scholar] [CrossRef]
- Simpson, T.L.; Volcani, B.E. Silicon and Siliceous Structures in Biological Systems; Springer New York: New York, NY, 1981; ISBN 978-1-4612-5944-2. [Google Scholar]
- Martin-Jézéquel, V.; Hildebrand, M.; Brzezinski, M.A. SILICON METABOLISM IN DIATOMS: IMPLICATIONS FOR GROWTH. J. Phycol. 2000, 36, 821–840. [Google Scholar] [CrossRef]
- Yool, A.; Tyrrell, T. Role of Diatoms in Regulating the Ocean’s Silicon Cycle. Glob. Biogeochem. Cycles 2003, 17, 2002GB002018. [Google Scholar] [CrossRef]
- Ragueneau, O.; Schultes, S.; Bidle, K.; Claquin, P.; Moriceau, B. Si and C Interactions in the World Ocean: Importance of Ecological Processes and Implications for the Role of Diatoms in the Biological Pump. Glob. Biogeochem. Cycles 2006, 20, 2006GB002688. [Google Scholar] [CrossRef]
- Tréguer, P.J.; De La Rocha, C.L. The World Ocean Silica Cycle. Annu. Rev. Mar. Sci. 2013, 5, 477–501. [Google Scholar] [CrossRef] [PubMed]
- Litchman, E. Resource Competition and the Ecological Success of Phytoplankton. In Evolution of Primary Producers in the Sea; Elsevier, 2007; pp. 351–375 ISBN 978-0-12-370518-1.
- Redfield, A. On the Proportions of Organic Derivatives in Sea Water and Their Relation to Their Planktonic Composition. In James Johnstone Memorial Volume; Woods Hole Oceanographic Institution: Liverpool, 1934; pp. 176–192. [Google Scholar]
- Takahashi, T.; Broecker, W.S.; Langer, S. Redfield Ratio Based on Chemical Data from Isopycnal Surfaces. J. Geophys. Res. Oceans 1985, 90, 6907–6924. [Google Scholar] [CrossRef]
- Geider, R.J.; Roche, J.L. Redfield Revisited: Variability of C[Ratio ]N[Ratio ]P in Marine Microalgae and Its Biochemical Basis. Eur. J. Phycol. 2002, 37, 1–17. [Google Scholar] [CrossRef]
- Goldman, J. On Phytoplankton Growth Rates and Particulate C: N: P Ratios at Low Light1. Limnol. Oceanogr. 1986. [Google Scholar] [CrossRef]
- Price, N.M. The Elemental Stoichiometry and Composition of an Iron-Limited Diatom. Limnol. Oceanogr. 2005, 50, 1159–1171. [Google Scholar] [CrossRef]
- Morel, F.; Rueter, J.; Price, N. Iron Nutrition of Phytoplankton and Its Possible Importance in the Ecology of Ocean Regions with High Nutrient and Low Biomass. Oceanography 1991, 4, 56–61. [Google Scholar] [CrossRef]
- Ho, T.; Quigg, A.; Finkel, Z.V.; Milligan, A.J.; Wyman, K.; Falkowski, P.G.; Morel, F.M.M. THE ELEMENTAL COMPOSITION OF SOME MARINE PHYTOPLANKTON 1. J. Phycol. 2003, 39, 1145–1159. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Milligan, A.J.; Saito, M.A. 6.05 Marine Bioinorganic Chemistry: The Role of Trace Metals in the Oceanic Cycles of Major Nutrients. 2003.
- Sunda, W.G. Feedback Interactions between Trace Metal Nutrients and Phytoplankton in the Ocean. Front. Microbiol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Twining, B.S.; Baines, S.B. The Trace Metal Composition of Marine Phytoplankton. Annu. Rev. Mar. Sci. 2013, 5, 191–215. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Penta, B.; Dieterle, D.A.; Bissett, W.P. Predictive Ecological Modeling of Harmful Algal Blooms. Hum. Ecol. Risk Assess. Int. J. 2001, 7, 1369–1383. [Google Scholar] [CrossRef]
- Villanoy, C.L.; Azanza, R.V.; Altemerano, A.; Casil, A.L. Attempts to Model the Bloom Dynamics of Pyrodinium, a Tropical Toxic Dinoflagellate. Harmful Algae 2006, 5, 156–183. [Google Scholar] [CrossRef]
- Hill, P.R.; Kumar, A.; Temimi, M.; Bull, D.R. HABNet: Machine Learning, Remote Sensing-Based Detection of Harmful Algal Blooms. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 3229–3239. [Google Scholar] [CrossRef]
- Yñiguez, A.T.; Ottong, Z.J. Predicting Fish Kills and Toxic Blooms in an Intensive Mariculture Site in the Philippines Using a Machine Learning Model. Sci. Total Environ. 2020, 707, 136–173. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.; Bao, M.; Lou, X.; Zhou, Z.; Yin, K. Monitoring, Modeling and Projection of Harmful Algal Blooms in China. Harmful Algae 2022, 111, 102164. [Google Scholar] [CrossRef] [PubMed]
- Bertani, I.; Obenour, D.R.; Steger, C.E.; Stow, C.A.; Gronewold, A.D.; Scavia, D. Probabilistically Assessing the Role of Nutrient Loading in Harmful Algal Bloom Formation in Western Lake Erie. J. Gt. Lakes Res. 2016, 42, 1184–1192. [Google Scholar] [CrossRef]
- Ai, H.; Zhang, K.; Sun, J.; Zhang, H. Short-Term Lake Erie Algal Bloom Prediction by Classification and Regression Models. Water Res. 2023, 232, 119710. [Google Scholar] [CrossRef]
- Iiames, J.S.; Salls, W.B.; Mehaffey, M.H.; Nash, M.S.; Christensen, J.R.; Schaeffer, B.A. Modeling Anthropogenic and Environmental Influences on Freshwater Harmful Algal Bloom Development Detected by MERIS Over the Central United States. Water Resour. Res. 2021, 57, e2020WR028946. [Google Scholar] [CrossRef]
- Wang, H.; Bouwman, A.F.; Van Gils, J.; Vilmin, L.; Beusen, A.H.W.; Wang, J.; Liu, X.; Yu, Z.; Ran, X. Hindcasting Harmful Algal Bloom Risk Due to Land-Based Nutrient Pollution in the Eastern Chinese Coastal Seas. Water Res. 2023, 231, 119669. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Chen, J.; Sun, X.; Li, D.; Liu, C.; Weng, H. Algae Explosive Growth Mechanism Enabling Weather-like Forecast of Harmful Algal Blooms. Sci. Rep. 2018, 8, 9923. [Google Scholar] [CrossRef] [PubMed]
- Bruland, K.W.; Franks, R.P. Mn, Ni, Cu, Zn and Cd in the Western North Atlantic. In Trace Metals in Sea Water; Wong, C.S., Boyle, E., Bruland, K.W., Burton, J.D., Goldberg, E.D., Eds.; Springer US: Boston, MA, 1983; pp. 395–414. ISBN 978-1-4757-6866-4. [Google Scholar]
- Whitfield, M. Interactions between Phytoplankton and Trace Metals in the Ocean. In Advances in Marine Biology; Elsevier, 2001; Vol. 41, pp. 1–128 ISBN 978-0-12-026141-3.
- John, S.G.; Sunda, W.G. Trace Metal Nutrients. In Encyclopedia of Ocean Sciences; Elsevier, 2019; pp. 208–217 ISBN 978-0-12-813082-7.
- Blain, S.; Quéguiner, B.; Armand, L.; Belviso, S.; Bombled, B.; Bopp, L.; Bowie, A.; Brunet, C.; Brussaard, C.; Carlotti, F.; et al. Effect of Natural Iron Fertilization on Carbon Sequestration in the Southern Ocean. Nature 2007, 446, 1070–1074. [Google Scholar] [CrossRef] [PubMed]
- Pollard, R.; Sanders, R.; Lucas, M.; Statham, P. The Crozet Natural Iron Bloom and Export Experiment (CROZEX). Deep Sea Res. Part II Top. Stud. Oceanogr. 2007, 54, 1905–1914. [Google Scholar] [CrossRef]
- Jacques, D.; Šimůnek, J.; Mallants, D.; Van Genuchten, M.Th. Modelling Coupled Water Flow, Solute Transport and Geochemical Reactions Affecting Heavy Metal Migration in a Podzol Soil. Geoderma 2008, 145, 449–461. [Google Scholar] [CrossRef]
- Moore, J.K.; Braucher, O. Sedimentary and Mineral Dust Sources of Dissolved Iron to the World Ocean. Biogeosciences 2008, 5, 631–656. [Google Scholar] [CrossRef]
- Elrod, V.A.; Berelson, W.M.; Coale, K.H.; Johnson, K.S. The Flux of Iron from Continental Shelf Sediments: A Missing Source for Global Budgets. Geophys. Res. Lett. 2004, 31, 2004GL020216. [Google Scholar] [CrossRef]
- Chase, Z.; Johnson, K.S.; Elrod, V.A.; Plant, J.N.; Fitzwater, S.E.; Pickell, L.; Sakamoto, C.M. Manganese and Iron Distributions off Central California Influenced by Upwelling and Shelf Width. Mar. Chem. 2005, 95, 235–254. [Google Scholar] [CrossRef]
- Lohan, M.C.; Bruland, K.W. Elevated Fe(II) and Dissolved Fe in Hypoxic Shelf Waters off Oregon and Washington: An Enhanced Source of Iron to Coastal Upwelling Regimes. Environ. Sci. Technol. 2008, 42, 6462–6468. [Google Scholar] [CrossRef]
- Von Damm, K.L.; Edmond, J.M.; Grant, B.; Measures, C.I.; Walden, B.; Weiss, R.F. Chemistry of Submarine Hydrothermal Solutions at 21 °N, East Pacific Rise. Geochim. Cosmochim. Acta 1985, 49, 2197–2220. [Google Scholar] [CrossRef]
- Fisher, A.T.; Davis, E.E.; Hutnak, M.; Spiess, V.; Zühlsdorff, L.; Cherkaoui, A.; Christiansen, L.; Edwards, K.; Macdonald, R.; Villinger, H.; et al. Hydrothermal Recharge and Discharge across 50 Km Guided by Seamounts on a Young Ridge Flank. Nature 2003, 421, 618–621. [Google Scholar] [CrossRef] [PubMed]
- Middag, R.; De Baar, H.J.W.; Laan, P.; Huhn, O. The Effects of Continental Margins and Water Mass Circulation on the Distribution of Dissolved Aluminum and Manganese in Drake Passage. J. Geophys. Res. Oceans 2012, 117, 2011JC007434. [Google Scholar] [CrossRef]
- Shaw, T.J.; Smith, K.L.; Hexel, C.R.; Dudgeon, R.; Sherman, A.D.; Vernet, M.; Kaufmann, R.S. 234Th-Based Carbon Export around Free-Drifting Icebergs in the Southern Ocean. Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 1384–1391. [Google Scholar] [CrossRef]
- Li, X.; He, X.; Kang, S.; Sillanpää, M.; Ding, Y.; Han, T.; Wu, Q.; Yu, Z.; Qin, D. Diurnal Dynamics of Minor and Trace Elements in Stream Water Draining Dongkemadi Glacier on the Tibetan Plateau and Its Environmental Implications. J. Hydrol. 2016, 541, 1104–1118. [Google Scholar] [CrossRef]
- Krause, J.; Hopwood, M.J.; Höfer, J.; Krisch, S.; Achterberg, E.P.; Alarcón, E.; Carroll, D.; González, H.E.; Juul-Pedersen, T.; Liu, T.; et al. Trace Element (Fe, Co, Ni and Cu) Dynamics Across the Salinity Gradient in Arctic and Antarctic Glacier Fjords. Front. Earth Sci. 2021, 9, 725279. [Google Scholar] [CrossRef]
- Mahowald, N.; Kohfeld, K.; Hansson, M.; Balkanski, Y.; Harrison, S.P.; Prentice, I.C.; Schulz, M.; Rodhe, H. Dust Sources and Deposition during the Last Glacial Maximum and Current Climate: A Comparison of Model Results with Paleodata from Ice Cores and Marine Sediments. J. Geophys. Res. Atmospheres 1999, 104, 15895–15916. [Google Scholar] [CrossRef]
- 1Chase, Z.; Paytan, A.; Beck, A.; Biller, D.; Bruland, K.; Measures, C.; Sañudo-Wilhelmy, S. Evaluating the Impact of Atmospheric Deposition on Dissolved Trace-Metals in the Gulf of Aqaba, Red Sea. Mar. Chem. 2011, 126, 256–268. [Google Scholar] [CrossRef]
- Liao, W.; Ho, T. Particulate Trace Metal Composition and Sources in the Kuroshio Adjacent to the East China Sea: The Importance of Aerosol Deposition. J. Geophys. Res. Oceans 2018, 123, 6207–6223. [Google Scholar] [CrossRef]
- Millward, G.E.; Morris, A.W.; Tappin, A.D. Trace Metals at Two Sites in the Southern North Sea: Results from a Sediment Resuspension Study. Cont. Shelf Res. 1998, 18, 1381–1400. [Google Scholar] [CrossRef]
- Berger, C.J.M.; Lippiatt, S.M.; Lawrence, M.G.; Bruland, K.W. Application of a Chemical Leach Technique for Estimating Labile Particulate Aluminum, Iron, and Manganese in the Columbia River Plume and Coastal Waters off Oregon and Washington. J. Geophys. Res. Oceans 2008, 113, 2007JC004703. [Google Scholar] [CrossRef]
- Li, T.; Sun, G.; Yang, C.; Liang, K.; Ma, S.; Huang, L.; Luo, W. Source Apportionment and Source-to-Sink Transport of Major and Trace Elements in Coastal Sediments: Combining Positive Matrix Factorization and Sediment Trend Analysis. Sci. Total Environ. 2019, 651, 344–356. [Google Scholar] [CrossRef]
- Sparaventi, E.; Rodríguez-Romero, A.; Barbosa, A.; Ramajo, L.; Tovar-Sánchez, A. Trace Elements in Antarctic Penguins and the Potential Role of Guano as Source of Recycled Metals in the Southern Ocean. Chemosphere 2021, 285, 131423. [Google Scholar] [CrossRef] [PubMed]
- Alba-González, P.; Álvarez-Salgado, X.A.; Cobelo-García, A.; Kaal, J.; Teira, E. Faeces of Marine Birds and Mammals as Substrates for Microbial Plankton Communities. Mar. Environ. Res. 2022, 174, 105560. [Google Scholar] [CrossRef] [PubMed]
- De La Peña-Lastra, S.; Pérez-Alberti, A.; Ferreira, T.O.; Huerta-Díaz, M.Á.; Otero, X.L. Global Deposition of Potentially Toxic Metals via Faecal Material in Seabird Colonies. Sci. Rep. 2022, 12, 22392. [Google Scholar] [CrossRef] [PubMed]
- Sugimura, Y.; Suzuki, Y.; Miyake, Y. The Content of Selenium and Its Chemical Form in Sea Water. J. Oceanogr. 1976, 32, 235–241. [Google Scholar] [CrossRef]
- Duan, L.; Song, J.; Li, X.; Yuan, H.; Xu, S. Distribution of Selenium and Its Relationship to the Eco-Environment in Bohai Bay Seawater. Mar. Chem. 2010, 121, 87–99. [Google Scholar] [CrossRef]
- Nozaki, Y. Rare Earth Elements and Their Isotopes in the Ocean. In Encyclopedia of Ocean Sciences; Steele, J.H., Ed.; Academic Press: Oxford, 2001; pp. 2354–2366. ISBN 978-0-12-227430-5. [Google Scholar]
- Moffett, J.W.; Ho, J. Oxidation of Cobalt and Manganese in Seawater via a Common Microbially Catalyzed Pathway. Geochim. Cosmochim. Acta 1996, 60, 3415–3424. [Google Scholar] [CrossRef]
- Saito, M.A.; Moffett, J.W. Complexation of Cobalt by Natural Organic Ligands in the Sargasso Sea as Determined by a New High-Sensitivity Electrochemical Cobalt Speciation Method Suitable for Open Ocean Work. Mar. Chem. 2001, 75, 49–68. [Google Scholar] [CrossRef]
- Noble, A.E.; Ohnemus, D.C.; Hawco, N.J.; Lam, P.J.; Saito, M.A. Coastal Sources, Sinks and Strong Organic Complexation of Dissolved Cobalt within the US North Atlantic GEOTRACES Transect GA03. Biogeosciences 2017, 14, 2715–2739. [Google Scholar] [CrossRef]
- Landing, W.M.; Bruland, K.W. The Contrasting Biogeochemistry of Iron and Manganese in the Pacific Ocean. Geochim. Cosmochim. Acta 1987, 51, 29–43. [Google Scholar] [CrossRef]
- Ho, P.; Lee, J.-M.; Heller, M.I.; Lam, P.J.; Shiller, A.M. The Distribution of Dissolved and Particulate Mo and V along the U.S. GEOTRACES East Pacific Zonal Transect (GP16): The Roles of Oxides and Biogenic Particles in Their Distributions in the Oxygen Deficient Zone and the Hydrothermal Plume. Mar. Chem. 2018, 201, 242–255. [Google Scholar] [CrossRef]
- Rickli, J.; Janssen, D.J.; Hassler, C.; Ellwood, M.J.; Jaccard, S.L. Chromium Biogeochemistry and Stable Isotope Distribution in the Southern Ocean. Geochim. Cosmochim. Acta 2019, 262, 188–206. [Google Scholar] [CrossRef]
- Chmiel, R.; Lanning, N.; Laubach, A.; Lee, J.-M.; Fitzsimmons, J.; Hatta, M.; Jenkins, W.; Lam, P.; McIlvin, M.; Tagliabue, A.; et al. Major Processes of the Dissolved Cobalt Cycle in the North and Equatorial Pacific Ocean. Biogeosciences 2022, 19, 2365–2395. [Google Scholar] [CrossRef]
- Wang, W.; Goring-Harford, H.; Kunde, K.; Woodward, E.M.S.; Lohan, M.C.; Connelly, D.P.; James, R.H. Biogeochemical Cycling of Chromium and Chromium Isotopes in the Sub-Tropical North Atlantic Ocean. Front. Mar. Sci. 2023, 10, 1165304. [Google Scholar] [CrossRef]
- Collier, R.W. Molybdenum in the Northeast Pacific Ocean1. Limnol. Oceanogr. 1985, 30, 1351–1354. [Google Scholar] [CrossRef]
- Fan, J.; Duan, L.; Yin, M.; Yuan, H.; Li, X. Nonconservative Behavior of Dissolved Molybdenum and Its Potential Role in Nitrogen Cycling in the Bohai and Yellow Seas. Front. Mar. Sci. 2022, 9, 1094846. [Google Scholar] [CrossRef]
- Whitmore, L.M.; Morton, P.L.; Twining, B.S.; Shiller, A.M. Vanadium Cycling in the Western Arctic Ocean Is Influenced by Shelf-Basin Connectivity. Mar. Chem. 2019, 216, 103701. [Google Scholar] [CrossRef]
- Bormans, M. Spatial and Temporal Variability in Cyanobacterial Populations Controlled by Physical Processes. J. Plankton Res. 2004, 27, 61–70. [Google Scholar] [CrossRef]
- Molot, L.A.; Watson, S.B.; Creed, I.F.; Trick, C.G.; McCabe, S.K.; Verschoor, M.J.; Sorichetti, R.J.; Powe, C.; Venkiteswaran, J.J.; Schiff, S.L. A Novel Model for Cyanobacteria Bloom Formation: The Critical Role of Anoxia and Ferrous Iron. Freshw. Biol. 2014, 59, 1323–1340. [Google Scholar] [CrossRef]
- Müller, S.; Mitrovic, S.M.; Baldwin, D.S. Oxygen and Dissolved Organic Carbon Control Release of N, P and Fe from the Sediments of a Shallow, Polymictic Lake. J. Soils Sediments 2016, 16, 1109–1120. [Google Scholar] [CrossRef]
- Paerl, H.W.; Hall, N.S.; Calandrino, E.S. Controlling Harmful Cyanobacterial Blooms in a World Experiencing Anthropogenic and Climatic-Induced Change. Sci. Total Environ. 2011, 409, 1739–1745. [Google Scholar] [CrossRef] [PubMed]
- Facey, J.A.; Apte, S.C.; Mitrovic, S.M. A Review of the Effect of Trace Metals on Freshwater Cyanobacterial Growth and Toxin Production. Toxins 2019, 11, 643. [Google Scholar] [CrossRef] [PubMed]
- Luoma, S.N.; Van Geen, A.; Lee, B.-G.; Cloern, J.E. Metal Uptake by Phytoplankton during a Bloom in South San Francisco Bay: Implications for Metal Cycling in Estuaries. Limnol. Oceanogr. 1998, 43, 1007–1016. [Google Scholar] [CrossRef]
- Blais, JulesM.; Kalff, J. Atmospheric Loading of Zn, Cu, Ni, Cr, and Pb to Lake Sediments: The Role of Catchment, Lake Morphometry, and Physico-Chemical Properties of the Elements. Biogeochemistry 1993, 23. [Google Scholar] [CrossRef]
- Giles, C.D.; Isles, P.D.F.; Manley, T.; Xu, Y.; Druschel, G.K.; Schroth, A.W. The Mobility of Phosphorus, Iron, and Manganese through the Sediment–Water Continuum of a Shallow Eutrophic Freshwater Lake under Stratified and Mixed Water-Column Conditions. Biogeochemistry 2016, 127, 15–34. [Google Scholar] [CrossRef]
- Saleem, M.; Iqbal, J.; Shah, M.H. Seasonal Variations, Risk Assessment and Multivariate Analysis of Trace Metals in the Freshwater Reservoirs of Pakistan. Chemosphere 2019, 216, 715–724. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Wang, Q.; Tian, X.; Zhu, X.; Dong, X.; Wu, Z.; Yuan, Y. Spatiotemporal Variation and Ecological Risk Assessment of Sediment Heavy Metals in Two Hydrologically Connected Lakes. Front. Ecol. Evol. 2022, 10, 1005194. [Google Scholar] [CrossRef]
- Sunda, W.G. Trace Metal/Phytoplankton Interactions in the Sea. In Chemistry of Aquatic Systems: Local and Global Perspectives; Bidoglio, G., Stumm, W., Eds.; Springer Netherlands: Dordrecht, 1994; ISBN 978-90-481-4410-5. [Google Scholar]
- Anderson, M.A.; Morel, F.M.M. The Influence of Aqueous Iron Chemistry on the Uptake of Iron by the Coastal Diatom Thalassiosira Weissflogii1. Limnol. Oceanogr. 1982, 27, 789–813. [Google Scholar] [CrossRef]
- Sunda, W.G. Trace Element Nutrients. In Encyclopedia of Ocean Sciences; Elsevier, 2001; pp. 75–86 ISBN 978-0-12-374473-9.
- Maldonado, M.T.; Strzepek, R.F.; Sander, S.; Boyd, P.W. Acquisition of Iron Bound to Strong Organic Complexes, with Different Fe Binding Groups and Photochemical Reactivities, by Plankton Communities in Fe-Limited Subantarctic Waters. Glob. Biogeochem. Cycles 2005, 19. [Google Scholar] [CrossRef]
- Sunda, W.G. Trace Metals and Harmful Algal Blooms. In Ecology of Harmful Algae; Granéli, E., Turner, J.T., Eds.; Ecological Studies; Springer Berlin Heidelberg, 2006; Vol. 189, pp. 203–214 ISBN 978-3-540-32209-2.
- Wever, R.; De Boer, E.; Plat, H.; E. Krenn, B. Vanadium — an Element Involved in the Biosynthesis of Halogenated Compounds and Nitrogen Fixation. FEBS Lett. 1987, 216, 1–3. [Google Scholar] [CrossRef]
- Kentemich, T.; Danneberg, G.; Hundeshagen, B.; Bothe, H. Evidence for the Occurrence of the Alternative, Vanadium-Containing Nitrogenase in the Cyanobacterium Anabaena Variabilis. FEMS Microbiol. Lett. 1988, 51, 19–24. [Google Scholar] [CrossRef]
- Price, N.M.; Morel, F.M.M. Cadmium and Cobalt Substitution for Zinc in a Marine Diatom. Nature 1990, 344, 658–660. [Google Scholar] [CrossRef]
- Raven, J.A. Predictions of Mn and Fe Use Efficiencies of Phototrophic Growth as a Function of Light Availability for Growth and of C Assimilation Pathway. New Phytol. 1990, 116, 1–18. [Google Scholar] [CrossRef]
- Raven, J.A.; Evans, M.C.W.; Korb, R.E. The Role of Trace Metals in Photosynthetic Electron Transport in O2-Evolving Organisms. Photosynth. Res. 1999, 60, 111–150. [Google Scholar] [CrossRef]
- Lane, T.W.; Morel, F.M.M. A Biological Function for Cadmium in Marine Diatoms. Proc. Natl. Acad. Sci. 2000, 97, 4627–4631. [Google Scholar] [CrossRef] [PubMed]
- Wolfe-Simon, F.; Grzebyk, D.; Schofield, O.; Falkowski, P.G. THE ROLE AND EVOLUTION OF SUPEROXIDE DISMUTASES IN ALGAE 1. J. Phycol. 2005, 41, 453–465. [Google Scholar] [CrossRef]
- Ji, Y.; Sherrell, R.M. Differential Effects of Phosphorus Limitation on Cellular Metals in Chlorella and Microcystis. Limnol. Oceanogr. 2008, 53, 1790–1804. [Google Scholar] [CrossRef]
- Rue, E.L.; Bruland, K.W. Complexation of Iron(II1) by Natural Organic Ligands in the Central North Pacific as Determined by a New Competitive Ligand Equilibration/Adsorptive Cathodic Stripping Voltammetric Method. Mar. Chem. 1995, 50, 117–138. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Lam, P.J.; Saito, M.A. Trace Metal Substitution in Marine Phytoplankton. Annu. Rev. Earth Planet. Sci. 2020, 48, 491–517. [Google Scholar] [CrossRef]
- Rachlin, J.W.; Jensen, T.E.; Warkentine, B. The Growth Response of the Diatom Navicula Incerta to Selected Concentrations of the Metals: Cadmium, Copper, Lead and Zinc. Bull. Torrey Bot. Club 1983, 110, 217. [Google Scholar] [CrossRef]
- Morel, F.M.M.; Reinfelder, J.R.; Roberts, S.B.; Chamberlain, C.P.; Lee, J.G.; Yee, D. Zinc and Carbon Co-Limitation of Marine Phytoplankton. Nature 1994, 369, 740–742. [Google Scholar] [CrossRef]
- Rue, E.; Bruland, K. Domoic Acid Binds Iron and Copper: A Possible Role for the Toxin Produced by the Marine Diatom Pseudo-Nitzschia. Mar. Chem. 2001, 76, 127–134. [Google Scholar] [CrossRef]
- Maldonado, M.T.; Price, N.M. REDUCTION AND TRANSPORT OF ORGANICALLY BOUND IRON BY THALASSIOSIRA OCEANICA (BACILLARIOPHYCEAE). J. Phycol. 2001, 37, 298–310. [Google Scholar] [CrossRef]
- Leblanc, K.; Hare, C.E.; Boyd, P.W.; Bruland, K.W.; Sohst, B.; Pickmere, S.; Lohan, M.C.; Buck, K.; Ellwood, M.; Hutchins, D.A. Fe and Zn Effects on the Si Cycle and Diatom Community Structure in Two Contrasting High and Low-Silicate HNLC Areas. Deep Sea Res. Part Oceanogr. Res. Pap. 2005, 52, 1842–1864. [Google Scholar] [CrossRef]
- Iwade, S.; Kuma, K.; Isoda, Y.; Yoshida, M.; Kudo, I.; Nishioka, J.; Suzuki, K. Effect of High Iron Concentrations on Iron Uptake and Growth of a Coastal Diatom Chaetoceros Sociale. Aquat. Microb. Ecol. 2006, 43, 177–191. [Google Scholar] [CrossRef]
- Guo, J.; Annett, A.L.; Taylor, R.L.; Lapi, S.; Ruth, T.J.; Maldonado, M.T. COPPER-UPTAKE KINETICS OF COASTAL AND OCEANIC DIATOMS 1. J. Phycol. 2010, 46, 1218–1228. [Google Scholar] [CrossRef]
- Varela, D.E.; Willers, V.; Crawford, D.W. EFFECT OF ZINC AVAILABILITY ON GROWTH, MORPHOLOGY, AND NUTRIENT INCORPORATION IN A COASTAL AND AN OCEANIC DIATOM1: ZINC EFFECTS ON MARINE DIATOMS. J. Phycol. 2011, 47, 302–312. [Google Scholar] [CrossRef] [PubMed]
- Twining, B.S.; Baines, S.B.; Vogt, S.; Nelson, D.M. Role of Diatoms in Nickel Biogeochemistry in the Ocean. Glob. Biogeochem. Cycles 2012, 26, 2011GB004233. [Google Scholar] [CrossRef]
- Saito, M.A.; Moffett, J.W.; Chisholm, S.W.; Waterbury, J.B. Cobalt Limitation and Uptake in Prochlorococcus. Limnol. Oceanogr. 2002, 47, 1629–1636. [Google Scholar] [CrossRef]
- Hawco, N.J.; McIlvin, M.M.; Bundy, R.M.; Tagliabue, A.; Goepfert, T.J.; Moran, D.M.; Valentin-Alvarado, L.; DiTullio, G.R.; Saito, M.A. Minimal Cobalt Metabolism in the Marine Cyanobacterium Prochlorococcus. Proc. Natl. Acad. Sci. 2020, 117, 15740–15747. [Google Scholar] [CrossRef]
- Sofen, L.E.; Antipova, O.A.; Ellwood, M.J.; Gilbert, N.E.; LeCleir, G.R.; Lohan, M.C.; Mahaffey, C.; Mann, E.L.; Ohnemus, D.C.; Wilhelm, S.W.; et al. Trace Metal Contents of Autotrophic Flagellates from Contrasting Open-ocean Ecosystems. Limnol. Oceanogr. Lett. 2022, 7, 354–362. [Google Scholar] [CrossRef]
- Cox, Alysia D.; Saito, Mak A., M.A. Proteomic Responses of Oceanic Synechococcus WH8102 to Phosphate and Zinc Scarcity and Cadmium Additions. Front. Microbiol. 2013. [Google Scholar] [CrossRef] [PubMed]
- Tuit, C.; Waterbury, J.; Ravizza, G. Diel Variation of Molybdenum and Iron in Marine Diazotrophic Cyanobacteria. Limnol. Oceanogr. 2004, 49, 978–990. [Google Scholar] [CrossRef]
- Yang, N.; Merkel, C.A.; Lin, Y.-A.; Levine, N.M.; Hawco, N.J.; Jiang, H.-B.; Qu, P.-P.; DeMers, M.A.; Webb, E.A.; Fu, F.-X.; et al. Warming Iron-Limited Oceans Enhance Nitrogen Fixation and Drive Biogeographic Specialization of the Globally Important Cyanobacterium Crocosphaera. Front. Mar. Sci. 2021, 8, 628363. [Google Scholar] [CrossRef]
- Tuo, S.; Rodriguez, I.B.; Ho, T. H 2 Accumulation and N 2 Fixation Variation by Ni Limitation in Cyanothece. Limnol. Oceanogr. 2020, 65, 377–386. [Google Scholar] [CrossRef]
- Rodriguez, I.B.; Lin, S.; Ho, J.; Ho, T.-Y. Effects of Trace Metal Concentrations on the Growth of the Coral Endosymbiont Symbiodinium Kawagutii. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef]
- Rodriguez, I.B.; Ho, T.-Y. Interactive Effects of Spectral Quality and Trace Metal Availability on the Growth of Trichodesmium and Symbiodinium. PLOS ONE 2017, 12, e0188777. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, I.B.; Ho, T.-Y. Trace Metal Requirements and Interactions in Symbiodinium Kawagutii. Front. Microbiol. 2018, 9, 142. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Rodriguez, I.B.; Chen, Y.L.; Zehr, J.P.; Chen, Y.; Hsu, S.D.; Yang, S.; Ho, T. Nickel Superoxide Dismutase Protects Nitrogen Fixation in Trichodesmium. Limnol. Oceanogr. Lett. 2022, 7, 363–371. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Iron Uptake and Growth Limitation in Oceanic and Coastal Phytoplankton. Mar. Chem. 1995, 50, 189–206. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Interrelated Influence of Iron, Light and Cell Size on Marine Phytoplankton Growth. Nature 1997, 390, 389–392. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580.e6. [Google Scholar] [CrossRef]
- Li, T.; Lin, X.; Yu, L.; Lin, S.; Rodriguez, I.B.; Ho, T.-Y. RNA-Seq Profiling of Fugacium Kawagutii Reveals Strong Responses in Metabolic Processes and Symbiosis Potential to Deficiencies of Iron and Other Trace Metals. Sci. Total Environ. 2020, 705, 135767. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Lu, Y.H. Influence of Environmental and Nutritional Factors on Growth, Toxicity, and Toxin Pro®le of Dino¯agellate Alexandrium Minutum. Toxicon 2000, 38, 1491–1503. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G.; Cochlan, W.P.; Hughes, M.P.; Trainer, V.L. Domoic Acid: The Synergy of Iron, Copper, and the Toxicity of Diatoms. Limnol. Oceanogr. 2005, 50, 1908–1917. [Google Scholar] [CrossRef]
- Hochmuth, J.D.; Asselman, J.; De Schamphelaere, K.A.C. Are Interactive Effects of Harmful Algal Blooms and Copper Pollution a Concern for Water Quality Management? Water Res. 2014, 60, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, X.; Wu, G.; Qin, B.; Tefsen, B.; Wells, M. Toxins from Harmful Algal Blooms: How Copper and Iron Render Chalkophore a Predictor of Microcystin Production. Water Res. 2023, 244, 120490. [Google Scholar] [CrossRef] [PubMed]
- Li, S. Interactions of Toxic Metals with Algal Toxins Derived from Harmful Algal Blooms. Master of Science Chemistry, Florida International University, 2011.
- Wu, H.; Wei, G.; Tan, X.; Li, L.; Li, M. Species-Dependent Variation in Sensitivity of Microcystis Species to Copper Sulfate: Implication in Algal Toxicity of Copper and Controls of Blooms. Sci. Rep. 2017, 7, 40393. [Google Scholar] [CrossRef]
- Shen, X.; Zhang, H.; He, X.; Shi, H.; Stephan, C.; Jiang, H.; Wan, C.; Eichholz, T. Evaluating the Treatment Effectiveness of Copper-Based Algaecides on Toxic Algae Microcystis Aeruginosa Using Single Cell-Inductively Coupled Plasma-Mass Spectrometry. Anal. Bioanal. Chem. 2019, 411, 5531–5543. [Google Scholar] [CrossRef]
- Tang, Y.Z.; Koch, F.; Gobler, C.J. Most Harmful Algal Bloom Species Are Vitamin B 1 and B 12 Auxotrophs. Proc. Natl. Acad. Sci. 2010, 107, 20756–20761. [Google Scholar] [CrossRef]
- Herzi, F.; Jean, N.; Sakka Hlaili, A.; Mounier, S. Three-dimensional (3-D) Fluorescence Spectroscopy Analysis of the Fluorescent Dissolved Organic Matter Released by the Marine Toxic Dinoflagellate A Lexandrium Catenella Exposed to Metal Stress by Zinc or Lead. J. Phycol. 2014, 50, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Couet, D.; Pringault, O.; Bancon-Montigny, C.; Briant, N.; Elbaz Poulichet, F.; Delpoux, S.; Kefi-Daly Yahia, O.; Hela, B.; Charaf, M.; Hervé, F.; et al. Effects of Copper and Butyltin Compounds on the Growth, Photosynthetic Activity and Toxin Production of Two HAB Dinoflagellates: The Planktonic Alexandrium Catenella and the Benthic Ostreopsis Cf. Ovata. Aquat. Toxicol. 2018, 196, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Herzi, F.; Jean, N.; Zhao, H.; Mounier, S.; Mabrouk, H.H.; Hlaili, A.S. Copper and Cadmium Effects on Growth and Extracellular Exudation of the Marine Toxic Dinoflagellate Alexandrium Catenella: 3D-Fluorescence Spectroscopy Approach. Chemosphere 2013, 93, 1230–1239. [Google Scholar] [CrossRef] [PubMed]
- Jean, N.; Dumont, E.; Herzi, F.; Balliau, T.; Laabir, M.; Masseret, E.; Mounier, S. Modifications of the Soluble Proteome of a Mediterranean Strain of the Invasive Neurotoxic Dinoflagellate Alexandrium Catenella under Metal Stress Conditions. Aquat. Toxicol. Amst. Neth. 2017, 188, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Yarimizu, K.; Mardones, J.I.; Paredes-Mella, J.; Norambuena-Subiabre, L.; Carrano, C.J.; Maruyama, F. The Effect of Iron on Chilean Alexandrium Catenella Growth and Paralytic Shellfish Toxin Production as Related to Algal Blooms. BioMetals 2022, 35, 39–51. [Google Scholar] [CrossRef]
- Doblin, M.; Blackburn, S.; Hallegraeff, G. Comparative Study of Selenium Requirements of Three Phytoplankton Species: Gymnodinium Catenatum, Alexandrium Minutum (Dinophyta) and Chaetoceros Cf. Tenuissimus (Bacillariophyta). J. Plankton Res. 1999, 21, 1153–1169. [Google Scholar] [CrossRef]
- Long, M.; Holland, A.; Planquette, H.; González Santana, D.; Whitby, H.; Soudant, P.; Sarthou, G.; Hégaret, H.; Jolley, D.F. Effects of Copper on the Dinoflagellate Alexandrium Minutum and Its Allelochemical Potency. Aquat. Toxicol. 2019, 210, 251–261. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Chen, F.; Li, H.; Xiang, W.; Li, Y.; Jiang, Y. Effect of Iron on Growth, Biochemical Composition and Paralytic Shellfish Poisoning Toxins Production of Alexandrium Tamarense. Harmful Algae 2010, 9, 98–104. [Google Scholar] [CrossRef]
- Lage, O.M.; Parente, A.M.; Soares, H.M.V.M.; Vasconcelos, M.T.S.D.; Salema, R. Some Effects of Copper on the Dinoflagellates Amphidinium Carterae and Prorocentrum Micans in Batch Culture. Eur. J. Phycol. 1994, 29, 253–260. [Google Scholar] [CrossRef]
- Wang, B.; Axe, L.; Michalopoulou, Z.-H.; Wei, L. Effects of Cd, Cu, Ni, and Zn on Brown Tide Alga Aureococcus Anophagefferens Growth and Metal Accumulation. Environ. Sci. Technol. 2012, 46, 517–524. [Google Scholar] [CrossRef]
- Gobler, C.J.; Lobanov, A.V.; Tang, Y.-Z.; Turanov, A.A.; Zhang, Y.; Doblin, M.; Taylor, G.T.; Sañudo-Wilhelmy, S.A.; Grigoriev, I.V.; Gladyshev, V.N. The Central Role of Selenium in the Biochemistry and Ecology of the Harmful Pelagophyte, Aureococcus Anophagefferens. ISME J. 2013, 7, 1333–1343. [Google Scholar] [CrossRef]
- Gobler, C.J.; Donat, J.R.; Consolvo, J.A.; Sañudo-Wilhelmy, S.A. Physicochemical Speciation of Iron during Coastal Algal Blooms. Mar. Chem. 2002, 77, 71–89. [Google Scholar] [CrossRef]
- Anderson, D.M.; Lively, J.S.; Vaccaro, R.F. Copper Complexation during Spring Phytoplankton Blooms in Coastal Waters. J. Mar. Res. 1984, 42, 677–695. [Google Scholar] [CrossRef]
- Naito, K.; Matsui, M.; Imai, I. Effects of Organic Iron Complexes on the Growth of Red Tide Causative Phytoplankton. In Proceedings of the Oceans ’04 MTS/IEEE Techno-Ocean ’04 (IEEE Cat. No.04CH37600); Vol. 3; IEEE: Kobe, Japan, 2004; pp. 1774–1780. [Google Scholar]
- García-Hernández, J.; García-Rico, L.; Jara-Marini, M.E.; Barraza-Guardado, R.; Hudson Weaver, A. Concentrations of Heavy Metals in Sediment and Organisms during a Harmful Algal Bloom (HAB) at Kun Kaak Bay, Sonora, Mexico. Mar. Pollut. Bull. 2005, 50, 733–739. [Google Scholar] [CrossRef]
- Sanders, J.G.; Riedel, G.F. Trace Element Transformation during the Development of an Estuarine Algal Bloom. Estuaries 1993, 16, 521. [Google Scholar] [CrossRef]
- Ebenezer, V.; Lim, W.A.; Ki, J.-S. Effects of the Algicides CuSO4 and NaOCl on Various Physiological Parameters in the Harmful Dinoflagellate Cochlodinium Polykrikoides. J. Appl. Phycol. 2014, 26, 2357–2365. [Google Scholar] [CrossRef]
- Guo, R.; Wang, H.; Suh, Y.S.; Ki, J.-S. Transcriptomic Profiles Reveal the Genome-Wide Responses of the Harmful Dinoflagellate Cochlodinium Polykrikoides When Exposed to the Algicide Copper Sulfate. BMC Genomics 2016, 17, 29. [Google Scholar] [CrossRef]
- Ebenezer, V.; Ki, J.-S. Evaluation of the Sub-Lethal Toxicity of Cu, Pb, Bisphenol A and Polychlorinated Biphenyl to the Marine Dinoflagellate Cochlodinium Polykrikoides. ALGAE 2012, 27, 63–70. [Google Scholar] [CrossRef]
- Martin, D.F.; Olander, W.K. Effects of Copper, Titanium and Zirconium on the Growth Rates of the Red Tide Organism, Gymnodinium Breve. Environ. Lett. 1971, 2, 135–142. [Google Scholar] [CrossRef]
- Doblin, M.A. Intraspecific Variation in the Selenium Requirement of Different Geographic Strains of the Toxic Dinoflagellate Gymnodinium Catenatum. J. Plankton Res. 2000, 22, 421–432. [Google Scholar] [CrossRef]
- Han, K.H.; Kim, H.J.; Li, Z.; Youn, J.Y.; Kwak, K.Y.; Seo, M.H.; Hwang, J.; Lee, S.D.; Yun, S.M.; Oh, S.J.; et al. Effects of Different Nutrient and Trace Metal Concentrations on Growth of the Toxic Dinoflagellate Gymnodinium Catenatum Isolated from Korean Coastal Waters. Sustainability 2020, 12, 4992. [Google Scholar] [CrossRef]
- Ishimaru, T.; Takeuchi, T.; Fukuyo, Y.; Kodama, M. The Selenium Requirement of Gymnodinium Nagasakiense. In Okaichi, T.Anderson, D.M. and Nemoto, T. In Red tides: Biology, Environmental Science and Toxicology; Elsevier: New York, NY, 1989; pp. 357–360. [Google Scholar]
- Koike, Y.; Nakaguchi, Y.; Hiraki, K.; Takeuchi, T.; Kokubo, T.; Ishimaru, T. Species and Concentrations of Selenium and Nutrients in Tanabe Bay during Red Tide Due toGymnodinium Nagasakiense. J. Oceanogr. 1993, 49, 641–656. [Google Scholar] [CrossRef]
- Doucette, G.; Harrison, P. Some Effects of Iron and Nitrogen Stress on the Red Tide Dinoflagellate Gymnodinium Sanguineum. Mar. Ecol. Prog. Ser. 1990, 62, 293–306. [Google Scholar] [CrossRef]
- Doucette, G.J.; Harrison, P.J. Aspects of Iron and Nitrogen Nutrition in the Red Tide Dinoflagellate Gymnodiniumsanguineum. Mar. Biol. 1991, 110, 165–173. [Google Scholar] [CrossRef]
- Rhodes, L.; Selwood, A.; McNabb, P.; Briggs, L.; Adamson, J.; Van Ginkel, R.; Laczka, O. Trace Metal Effects on the Production of Biotoxins by Microalgae. Afr. J. Mar. Sci. 2006, 28, 393–397. [Google Scholar] [CrossRef]
- Gutierrez-Mejia, E.; Lares, M.L.; Huerta-Diaz, M.A.; Delgadillo-Hinojosa, F. Cadmium and Phosphate Variability during Algal Blooms of the Dinoflagellate Lingulodinium Polyedrum in Todos Santos Bay, Baja California, Mexico. Sci. Total Environ. 2016, 541, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Barros, M.P.; Hollnagel, H.C.; Glavina, A.B.; Soares, C.O.; Ganini, D.; Dagenais-Bellefeuille, S.; Morse, D.; Colepicolo, P. Molybdate:Sulfate Ratio Affects Redox Metabolism and Viability of the Dinoflagellate Lingulodinium Polyedrum. Aquat. Toxicol. 2013, 142–143, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, S.M.; Fernández Amandi, M.; McKenzie, L.; Furey, A.; James, K.J. Effects of Selenium, Iron and Cobalt Addition to Growth and Yessotoxin Production of the Toxic Marine Dinoflagellate Protoceratium Reticulatum in Culture. J. Exp. Mar. Biol. Ecol. 2004, 313, 337–351. [Google Scholar] [CrossRef]
- Tian, R.; Lin, Q.; Li, D.; Zhang, W.; Zhao, X. Atmospheric Transport of Nutrients during a Harmful Algal Bloom Event. Reg. Stud. Mar. Sci. 2020, 34, 101007. [Google Scholar] [CrossRef]
- Maldonado, M.T.; Hughes, M.P.; Rue, E.L.; Wells, M.L. The Effect of Fe and Cu on Growth and Domoic Acid Production by Pseudo-Nitzschiamultiseries and Pseudo-nitzschia Australis. Limnol. Oceanogr. 2002, 47, 515–526. [Google Scholar] [CrossRef]
- Lelong, A.; Bucciarelli, E.; Hégaret, H.; Soudant, P. Iron and Copper Limitations Differently Affect Growth Rates and Photosynthetic and Physiological Parameters of the Marine Diatom Pseudo-nitzschia Delicatissima. Limnol. Oceanogr. 2013, 58, 613–623. [Google Scholar] [CrossRef]
- Long, M.; Lelong, A.; Bucciarelli, E.; Le Grand, F.; Hégaret, H.; Soudant, P. Physiological Adaptation of the Diatom Pseudo-Nitzschia Delicatissima under Copper Starvation. Mar. Environ. Res. 2023, 188, 105995. [Google Scholar] [CrossRef] [PubMed]
- Sobrinho, B.F.; De Camargo, L.M.; Sandrini-Neto, L.; Kleemann, C.R.; Machado, E.D.C.; Mafra, L.L. Growth, Toxin Production and Allelopathic Effects of Pseudo-Nitzschia Multiseries under Iron-Enriched Conditions. Mar. Drugs 2017, 15, 331. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.S.; Wikfors, G.H. Control of Domoic Acid Toxin Expression in Pseudo-Nitzschia Multiseries by Copper and Silica: Relevance to Mussel Aquaculture in New England (USA). Mar. Environ. Res. 2013, 83, 23–28. [Google Scholar] [CrossRef]
- Ladizinsky, N.L.; Smith J., G. Accumulation of Domoic Acid by the Coastal Diatom Pseudo-Nitzschia Multiseries: A Possible Copper Complexation Strategy. J. Phycol. 2000, 36, 41–41. [Google Scholar] [CrossRef]
- Zhuo-Ping, C.; Wei-Wei, H.; Min, A.; Shun-Shan, D. Coupled Effects of Irradiance and Iron on the Growth of a Harmful Algal Bloom-Causing Microalga Scrippsiella Trochoidea. Acta Ecol. Sin. 2009, 29, 297–301. [Google Scholar] [CrossRef]
- Ho, T.-Y. Nickel Limitation of Nitrogen Fixation in Trichodesmium. Limnol. Oceanogr. 2013, 58, 112–120. [Google Scholar] [CrossRef]
- Greenfield, D.I.; Duquette, A.; Goodson, A.; Keppler, C.J.; Williams, S.H.; Brock, L.M.; Stackley, K.D.; White, D.; Wilde, S.B. The Effects of Three Chemical Algaecides on Cell Numbers and Toxin Content of the Cyanobacteria Microcystis Aeruginosa and Anabaenopsis Sp. Environ. Manage. 2014, 54, 1110–1120. [Google Scholar] [CrossRef]
- Molot, L.A.; Li, G.; Findlay, D.L.; Watson, S.B. Iron-mediated Suppression of Bloom-forming Cyanobacteria by Oxine in a Eutrophic Lake. Freshw. Biol. 2010, 55, 1102–1117. [Google Scholar] [CrossRef]
- Cusick, K.D.; Wetzel, R.K.; Minkin, S.C.; Dodani, S.C.; Wilhelm, S.W.; Sayler, G.S. Paralytic Shellfish Toxins Inhibit Copper Uptake in Chlamydomonas Reinhardtii. Environ. Toxicol. Chem. 2013, 32, 1388–1395. [Google Scholar] [CrossRef]
- Wang, H.; Sathasivam, R.; Ki, J.-S. Physiological Effects of Copper on the Freshwater Alga Closterium Ehrenbergii Meneghini (Conjugatophyceae) and Its Potential Use in Toxicity Assessments. ALGAE 2017, 32, 131–137. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Deng, J.; Cheng, Y.; Chen, Z.; Qin, B.; Tefsen, B.; Wells, M. A New Perspective of Copper-Iron Effects on Bloom-Forming Algae in a Highly Impacted Environment. Water Res. 2021, 195, 116889. [Google Scholar] [CrossRef]
- Bishop, W.M.; Lynch, C.L.; Willis, B.E.; Cope, W.G. Copper-Based Aquatic Algaecide Adsorption and Accumulation Kinetics: Influence of Exposure Concentration and Duration for Controlling the Cyanobacterium Lyngbya Wollei. Bull. Environ. Contam. Toxicol. 2017, 99, 365–371. [Google Scholar] [CrossRef]
- Jin, Z.; Ding, S.; Sun, Q.; Gao, S.; Fu, Z.; Gong, M.; Lin, J.; Wang, D.; Wang, Y. High Resolution Spatiotemporal Sampling as a Tool for Comprehensive Assessment of Zinc Mobility and Pollution in Sediments of a Eutrophic Lake. J. Hazard. Mater. 2019, 364, 182–191. [Google Scholar] [CrossRef]
- Lukač, M.; Aegerter, R. Influence of Trace Metals on Growth and Toxin Production of Microcystis Aeruginosa. Toxicon 1993, 31, 293–305. [Google Scholar] [CrossRef]
- Facey, J.A.; Violi, J.P.; King, J.J.; Sarowar, C.; Apte, S.C.; Mitrovic, S.M. The Influence of Micronutrient Trace Metals on Microcystis Aeruginosa Growth and Toxin Production. Toxins 2022, 14, 812. [Google Scholar] [CrossRef]
- Gu, P.; Li, Q.; Zhang, W.; Zheng, Z.; Luo, X. Effects of Different Metal Ions (Ca, Cu, Pb, Cd) on Formation of Cyanobacterial Blooms. Ecotoxicol. Environ. Saf. 2020, 189, 109976. [Google Scholar] [CrossRef]
- Zeng, L.; Yan, C.; Guo, J.; Zhen, Z.; Zhao, Y.; Wang, D. Influence of Algal Blooms Decay on Arsenic Dynamics at the Sediment-Water Interface of a Shallow Lake. Chemosphere 2019, 219, 1014–1023. [Google Scholar] [CrossRef]
- Yu, S.; Xu, C.; Tang, T.; Zhang, Y.; Effiong, K.; Hu, J.; Bi, Y.; Xiao, X. Down-Regulation of Iron/Zinc Ion Transport and Toxin Synthesis in Microcystis Aeruginosa Exposed to 5,4′-Dihydroxyflavone. J. Hazard. Mater. 2023, 460, 132396. [Google Scholar] [CrossRef] [PubMed]
- Gouvêa, S.P.; Boyer, G.L.; Twiss, M.R. Influence of Ultraviolet Radiation, Copper, and Zinc on Microcystin Content in Microcystis Aeruginosa (Cyanobacteria). Harmful Algae 2008, 7, 194–205. [Google Scholar] [CrossRef]
- Deng, J.; Fu, D.; Hu, W.; Lu, X.; Wu, Y.; Bryan, H. Physiological Responses and Accumulation Ability of Microcystis Aeruginosa to Zinc and Cadmium: Implications for Bioremediation of Heavy Metal Pollution. Bioresour. Technol. 2020, 303, 122963. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Ruiz, E.B.; Martínez-Jerónimo, F. How Do Toxic Metals Affect Harmful Cyanobacteria? An Integrative Study with a Toxigenic Strain of Microcystis Aeruginosa Exposed to Nickel Stress. Ecotoxicol. Environ. Saf. 2016, 133, 36–46. [Google Scholar] [CrossRef]
- Alexova, R.; Fujii, M.; Birch, D.; Cheng, J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Iron Uptake and Toxin Synthesis in the Bloom-Forming Microcystis Aeruginosa under Iron Limitation. Environ. Microbiol. 2011, 13, 1064–1077. [Google Scholar] [CrossRef] [PubMed]
- Alexova, R.; Dang, T.C.; Fujii, M.; Raftery, M.J.; Waite, T.D.; Ferrari, B.C.; Neilan, B.A. Specific Global Responses to N and Fe Nutrition in Toxic and Non-Toxic Microcystis Aeruginosa. Environ. Microbiol. 2016, 18, 401–413. [Google Scholar] [CrossRef]
- Kosakowska, A.; Nędzi, M.; Pempkowiak, J. Responses of the Toxic Cyanobacterium Microcystis Aeruginosa to Iron and Humic Substances. Plant Physiol. Biochem. 2007, 45, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Bishop, W.M.; Willis, B.E.; Richardson, R.J. Sensitivity of Microcystis Aeruginosa Strains to Copper and Influence of Phosphorus. J Aquat Plant Manage 2019. [Google Scholar]
- Kansole, M.; Lin, T.-F. Impacts of Hydrogen Peroxide and Copper Sulfate on the Control of Microcystis Aeruginosa and MC-LR and the Inhibition of MC-LR Degrading Bacterium Bacillus Sp. Water 2017, 9, 255. [Google Scholar] [CrossRef]
- Liu, H.; Chen, S.; Zhang, H.; Wang, N.; Ma, B.; Liu, X.; Niu, L.; Yang, F.; Xu, Y.; Zhang, X. Effects of Copper Sulfate Algaecide on the Cell Growth, Physiological Characteristics, the Metabolic Activity of Microcystis Aeruginosa and Raw Water Application. J. Hazard. Mater. 2023, 445, 130604. [Google Scholar] [CrossRef]
- Qian, H.; Yu, S.; Sun, Z.; Xie, X.; Liu, W.; Fu, Z. Effects of Copper Sulfate, Hydrogen Peroxide and N-Phenyl-2-Naphthylamine on Oxidative Stress and the Expression of Genes Involved Photosynthesis and Microcystin Disposition in Microcystis Aeruginosa. Aquat. Toxicol. 2010, 99, 405–412. [Google Scholar] [CrossRef]
- Burger, H.; Dickson, S.; Awad, J.; Marzouk, J.; Van Leeuwen, J. Investigation of Cyanobacteria Blooms in Paper Mill Wastewaters and Assessment of Zinc as a Control Agent. Int. J. Environ. Sci. Technol. 2022, 19, 1105–1120. [Google Scholar] [CrossRef]
- Perez, J.L.; Chu, T. Effect of Zinc on Microcystis Aeruginosa UTEX LB 2385 and Its Toxin Production. Toxins 2020, 12, 92. [Google Scholar] [CrossRef] [PubMed]
- ]Gong, Y.; Ao, H.; Liu, B.; Wen, S.; Wang, Z.; Hu, D.; Zhang, X.; Song, L.; Liu, J. Effects of Inorganic Arsenic on Growth and Microcystin Production of a Microcystis Strain Isolated from an Algal Bloom in Dianchi Lake, China. Chin. Sci. Bull. 2011, 56, 2337–2342. [Google Scholar] [CrossRef]
- Morton, S.D.; Lee, T.H. Algal Blooms. Possible Effects of Iron. Environ. Sci. Technol. 1974, 8, 673–674. [Google Scholar] [CrossRef]
- Wells, M.L.; Trick, C.G. Controlling Iron Availability to Phytoplankton in Iron-Replete Coastal Waters. Mar. Chem. 2004, 86, 1–13. [Google Scholar] [CrossRef]
- Dawson, R.M. The Toxicology of Microcystins. Toxicon 1998, 36, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Vasconcelos, V. Molecular Mechanisms of Microcystin Toxicity in Animal Cells. Int. J. Mol. Sci. 2010, 11, 268–287. [Google Scholar] [CrossRef]
- Bates, S.S.; Léger, C.; Satchwell, M.; Boyer, G.L. THE EFFECTS OF IRON ON DOMOIC ACID PRODUCTION BY PSEUDO-NITZSCHIA MULTISERIES. In Proceedings of the Proceeding of the 9th International Conference on Harmful Algal Blooms; Intergov. Oceanogr. Comm., Paris: Tasmania, 2001; pp. 320–323. [Google Scholar]
- Pan, Y.; Subba Rao, D.V.; Mann, K.H. Changes in Domoic Acid Production and Cellular Chemical Composition of the Toxigenic Diatom Pseudo-Nitzschia Multiseries Under Phosphate Limitation1. J. Phycol. 1996, 32, 371–381. [Google Scholar] [CrossRef]
- Fehling, J.; Davidson, K.; Bolch, C.J.; Bates, S.S. Growth and Domoic Acid Production by Pseudo-Nitzschia Seriata (Bacillariophyceae) Under Phosphate and Silicate Limitation1. J. Phycol. 2004, 40, 674–683. [Google Scholar] [CrossRef]
- Sun, J.; Hutchins, D.A.; Feng, Y.; Seubert, E.L.; Caron, D.A.; Fu, F.-X. Effects of Changing pCO2 and Phosphate Availability on Domoic Acid Production and Physiology of the Marine Harmful Bloom Diatom Pseudo-Nitzschia Multiseries. Limnol. Oceanogr. 2011, 56, 829–840. [Google Scholar] [CrossRef]
- Kai, N.; Naito, K.; Mito, S.; Miyahara, K.; Sakamoto, S. Distribution and Behavior of Harmful Algae and Trace Metals in Harima-Nada, Japan.; Japanese Society of Fisheries Science, 2017; Vol. 4006.
- Leung, T.; Wilkinson, G.M.; Swanner, E.D. Iron Availability Allows Sustained Cyanobacterial Blooms: A Dual-Lake Case Study. Inland Waters 2021, 11, 417–429. [Google Scholar] [CrossRef]
- Wang, S.; Diao, X.; He, L. Effects of Algal Bloom Formation, Outbreak, and Extinction on Heavy Metal Fractionation in the Surficial Sediments of Chaohu Lake. Environ. Sci. Pollut. Res. 2015, 22, 14269–14279. [Google Scholar] [CrossRef]
- Thamdrup, B.; Fossing, H.; Jørgensen, B.B. Manganese, Iron and Sulfur Cycling in a Coastal Marine Sediment, Aarhus Bay, Denmark. Geochim. Cosmochim. Acta 1994, 58, 5115–5129. [Google Scholar] [CrossRef]
- Schoemann, V.; De Baar, H.J.W.; De Jong, J.T.M.; Lancelot, C. Effects of Phytoplankton Blooms on the Cycling of Manganese and Iron in Coastal Waters. Limnol. Oceanogr. 1998, 43, 1427–1441. [Google Scholar] [CrossRef]
- Sun, Q.; Lin, J.; Ding, S.; Gao, S.; Gao, M.; Wang, Y.; Zhang, C. A Comprehensive Understanding of Enhanced Pb Mobilization in Sediments Caused by Algal Blooms. Sci. Total Environ. 2019, 691, 969–980. [Google Scholar] [CrossRef]
- Sedmak, B.; Kosi, G. The Role of Microcystins in Heavy Cyanobacterial Bloom Formation. J. Plankton Res. 1998, 20, 691–708. [Google Scholar] [CrossRef]
- Kemp, A.; John, J. Microcystins Associated with Microcystis Dominated Blooms in the Southwest Wetlands, Western Australia. Environ. Toxicol. 2006, 21, 125–130. [Google Scholar] [CrossRef]
- Steiner, K.; Wood, S.; Puddick, J.; Hawes, I.; Dietrich, D.; Hamilton, D. A Comparison of Bacterial Community Structure, Activity and Microcystins Associated with Formation and Breakdown of a Cyanobacterial Scum. Aquat. Microb. Ecol. 2017, 80, 243–256. [Google Scholar] [CrossRef]
- Brand, L.E. Minimum Iron Requirements of Marine Phytoplankton and the Implications for the Biogeochemical Control of New Production. Limnol. Oceanogr. 1991, 36, 1756–1771. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Quigg, A.; Finkel, Z.V.; Irwin, A.J.; Haramaty, L. Nitrogen-Fixation Strategies and Fe Requirements in Cyanobacteria. Limnol. Oceanogr. 2007, 52, 2260–2269. [Google Scholar] [CrossRef]
- Chen, M.; Ding, S.; Chen, X.; Sun, Q.; Fan, X.; Lin, J.; Ren, M.; Yang, L.; Zhang, C. Mechanisms Driving Phosphorus Release during Algal Blooms Based on Hourly Changes in Iron and Phosphorus Concentrations in Sediments. Water Res. 2018, 133, 153–164. [Google Scholar] [CrossRef]
- Yao, Y.; Han, X.; Chen, Y.; Li, D. The Variations of Labile Arsenic Diffusion Driven by Algal Bloom Decomposition in Eutrophic Lake Ecosystems. Sci. Total Environ. 2022, 842, 156703. [Google Scholar] [CrossRef]
- Orihel, D.M.; Schindler, D.W.; Ballard, N.C.; Wilson, L.R.; Vinebrooke, R.D. Experimental Iron Amendment Suppresses Toxic Cyanobacteria in a Hypereutrophic Lake. Ecol. Appl. 2016, 26, 1517–1534. [Google Scholar] [CrossRef]
- Zhang, X.; Li, B.; Xu, H.; Wells, M.; Tefsen, B.; Qin, B. Effect of Micronutrients on Algae in Different Regions of Taihu, a Large, Spatially Diverse, Hypereutrophic Lake. Water Res. 2019, 151, 500–514. [Google Scholar] [CrossRef]
- Lelong, A.; Jolley, D.F.; Soudant, P.; Hégaret, H. Impact of Copper Exposure on Pseudo-Nitzschia Spp. Physiology and Domoic Acid Production. Aquat. Toxicol. 2012, 118–119, 37–47. [Google Scholar] [CrossRef]
- Castruita, M.; Casero, D.; Karpowicz, S.J.; Kropat, J.; Vieler, A.; Hsieh, S.I.; Yan, W.; Cokus, S.; Loo, J.A.; Benning, C.; et al. Systems Biology Approach in Chlamydomonas Reveals Connections between Copper Nutrition and Multiple Metabolic Steps. Plant Cell 2011, 23, 1273–1292. [Google Scholar] [CrossRef]
- Humble, A.V.; Gadd, G.M.; Codd, G.A. Binding of Copper and Zinc to Three Cyanobacterial Microcystins Quantified by Differential Pulse Polarography. Water Res. 1997, 31, 1679–1686. [Google Scholar] [CrossRef]
- Yan, F.; Ozsoz, M.; Sadik, O.A. Electrochemical and Conformational Studies of Microcystin–LR. Anal. Chim. Acta 2000, 409, 247–255. [Google Scholar] [CrossRef]
- Saito, K.; Sei, Y.; Miki, S.; Yamaguchi, K. Detection of Microcystin–Metal Complexes by Using Cryospray Ionization-Fourier Transform Ion Cyclotron Resonance Mass Spectrometry. Toxicon 2008, 51, 1496–1498. [Google Scholar] [CrossRef]
- Le Faucheur, S.; Schildknecht, F.; Behra, R.; Sigg, L. Thiols in Scenedesmus Vacuolatus upon Exposure to Metals and Metalloids. Aquat. Toxicol. 2006, 80, 355–361. [Google Scholar] [CrossRef]
- Yang, F.; Hu, Y.; Qiu, G.; Li, Q.; Wang, G. Complexation of Copper Algaecide and Algal Organic Matter in Algae-Laden Water: Insights into Complex Metal–Organic Interactions. Environ. Pollut. 2023, 333, 122032. [Google Scholar] [CrossRef]
- Zwolsman, J.J.G.; van Eck, G.T.M. Geochemistry of Major Elements and Trace Metals in Suspended Matter of the Scheldt Estuary, Southwest Netherlands. Mar. Chem. 1999, 66, 91–111. [Google Scholar] [CrossRef]
- Luengen, A.C.; Raimondi, P.T.; Flegal, A.R. Contrasting Biogeochemistry of Six Trace Metals during the Rise and Decay of a Spring Phytoplankton Bloom in San Francisco Bay. Limnol. Oceanogr. 2007, 52, 1112–1130. [Google Scholar] [CrossRef]
- Kuwabara, J.S.; Chang, C.C.Y.; Cloern, J.E.; Fries, T.L.; Davis, J.A.; Luoma, S.N. Trace Metal Associations in the Water Column of South San Francisco Bay, California. Estuar. Coast. Shelf Sci. 1989, 28, 307–325. [Google Scholar] [CrossRef]
- Araie, H.; Shiraiwa, Y. Selenium Utilization Strategy by Microalgae. Molecules 2009, 14, 4880–4891. [Google Scholar] [CrossRef]
- Boyer, G.L.; Brand, L.E. Trace Elements and Harmful Algal Blooms. In Physiological Ecology of Harmful Algal Blooms; Springer Verlag: Heidelberg, 1989; Vol. 41, pp. 498–508. [Google Scholar]
- Chang, Y.; Wu, Y.; Zhang, J.; Wang, X.; Jiang, S.; Cao, W.; Wang, X.; Qu, J.; Zhang, Z.; Jin, J.; et al. Effects of Algal Blooms on Selenium Species Dynamics: A Case Study in the Changjiang Estuary, China. Sci. Total Environ. 2021, 768, 144235. [Google Scholar] [CrossRef]
- Conde, J.E.; Sanz Alaejos, M. Selenium Concentrations in Natural and Environmental Waters. Chem. Rev. 1997, 97, 1979–2004. [Google Scholar] [CrossRef]
- Danbara, A.; Shiraiwa, Y. The Requirement of Selenium for the Growth of Marine Coccolithophorids, Emiliania Huxleyi, Gephyrocapsa Oceanica and Helladosphaera Sp. (Prymnesiophyceae). Plant Cell Physiol. 1999, 40, 762–766. [Google Scholar] [CrossRef]
- Cutter, G.A.; Bruland, K.W. The Marine Biogeochemistry of Selenium: A Re-evaluation1. Limnol. Oceanogr. 1984, 29, 1179–1192. [Google Scholar] [CrossRef]
- Yang, Y.; Hu, M. Uptake and Transformation of Selenium by Marine Phytoplankton. J. Oceanogr. Taiwan Strait 1996, 15, 225–231. [Google Scholar]
- Ivanenko, N.V. The Role of Microorganisms in Transformation of Selenium in Marine Waters. Russ. J. Mar. Biol. 2018, 44, 87–93. [Google Scholar] [CrossRef]
- Chang, Y.; Müller, M.; Wu, Y.; Jiang, S.; Cao, W.W.; Qu, J.G.; Ren, J.L.; Wang, X.N.; Rao, E.M.; Wang, X.L.; et al. Distribution and Behaviour of Dissolved Selenium in Tropical Peatland-Draining Rivers and Estuaries of Malaysia. Biogeosciences 2020, 17, 1133–1145. [Google Scholar] [CrossRef]
- Peers, G.; Price, N.M. A Role for Manganese in Superoxide Dismutases and Growth of Iron-deficient Diatoms. Limnol. Oceanogr. 2004, 49, 1774–1783. [Google Scholar] [CrossRef]
- Graneli, E.; Haraldsson, C. Can Increased Leaching of Trace Metals from Acidified Areas Influence Phytoplankton Growth in Coastal Waters? 1994.
- Granéli, E.; Risinger, L. Effects of Cobalt and Vitamin B12 on the Growth of Chrysochromulina Polylepis (Prymnesiophyceae). Mar. Ecol. Prog. Ser. 1994, 113, 177–183. [Google Scholar] [CrossRef]
- Saito, M.A.; Rocap, G.; Moffett, J.W. Production of Cobalt Binding Ligands in a Synechococcus Feature at the Costa Rica Upwelling Dome. Limnol. Oceanogr. 2005, 50, 279–290. [Google Scholar] [CrossRef]
- Sañudo-Wilhelmy, S.A.; Gómez-Consarnau, L.; Suffridge, C.; Webb, E.A. The Role of B Vitamins in Marine Biogeochemistry. Annu. Rev. Mar. Sci. 2014, 6, 339–367. [Google Scholar] [CrossRef]
- Roth, J.; Lawrence, J.; Bobik, T. COBALAMIN (COENZYME B 12 ): Synthesis and Biological Significance. Annu. Rev. Microbiol. 1996, 50, 137–181. [Google Scholar] [CrossRef]
- Förstner, U.; Wittmann, G.T.; Prosi, F.; Lierde, J.H. van Metal Pollution in the Aquatic Environment; Springer study edition; 2., rev. ed., 2. print.; Springer: Berlin Heidelberg New York Tokyo, 1983; ISBN 978-3-540-12856-4.
- Tovar-Sanchez, A.; Sañudo-Wilhelmy, S.A. Influence of the Amazon River on Dissolved and Intra-Cellular Metal Concentrations in <I>Trichodesmium</I> Colonies along the Western Boundary of the Sub-Tropical North Atlantic Ocean. Biogeosciences 2011, 8, 217–225. [Google Scholar] [CrossRef]
- Price, N.M.; Morel, F.M.M. Colimitation of Phytoplankton Growth by Nickel and Nitrogen. Limnol. Oceanogr. 1991, 36, 1071–1077. [Google Scholar] [CrossRef]
- Dupont, C.L.; Buck, K.N.; Palenik, B.; Barbeau, K. Nickel Utilization in Phytoplankton Assemblages from Contrasting Oceanic Regimes. Deep Sea Res. Part Oceanogr. Res. Pap. 2010, 57, 553–566. [Google Scholar] [CrossRef]
- Gobler, C.J.; Berry, D.L.; Dyhrman, S.T.; Wilhelm, S.W.; Salamov, A.; Lobanov, A.V.; Zhang, Y.; Collier, J.L.; Wurch, L.L.; Kustka, A.B.; et al. Niche of Harmful Alga Aureococcus Anophagefferens Revealed through Ecogenomics. Proc. Natl. Acad. Sci. 2011, 108, 4352–4357. [Google Scholar] [CrossRef]
- Sunda, W.G.; Huntsman, S.A. Effect of Zn, Mn, and Fe on Cd Accumulation in Phytoplankton: Implications for Oceanic Cd Cycling. Limnol. Oceanogr. 2000, 45, 1501–1516. [Google Scholar] [CrossRef]
- Xu, Y.; Morel, F.M.M. Cadmium in Marine Phytoplankton. In Cadmium: From Toxicity to Essentiality; Sigel, A., Sigel, H., Sigel, R.K., Eds.; Metal Ions in Life Sciences; Springer Netherlands: Dordrecht, 2013; Vol. 11, pp. 509–528. ISBN 978-94-007-5178-1. [Google Scholar]
- Granéli, E.; Persson, H.; Edler, L. Connection between Trace Metals, Chelators and Red Tide Blooms in the Laholm Bay, SE Kattegat—an Experimental Approach. Mar. Environ. Res. 1986, 18, 61–78. [Google Scholar] [CrossRef]
- Kaplan, D. Absorption and Adsorption of Heavy Metals by Microalgae. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; Wiley, 2013; pp. 602–611 ISBN 978-0-470-67389-8.
- McIntyre, A.M.; Guéguen, C. Binding Interactions of Algal-Derived Dissolved Organic Matter with Metal Ions. Chemosphere 2013, 90, 620–626. [Google Scholar] [CrossRef]
- Mühlenbruch, M.; Grossart, H.; Eigemann, F.; Voss, M. Mini-review: Phytoplankton-derived Polysaccharides in the Marine Environment and Their Interactions with Heterotrophic Bacteria. Environ. Microbiol. 2018, 20, 2671–2685. [Google Scholar] [CrossRef] [PubMed]
- Tulcan, R.X.S.; Ouyang, W.; Lin, C.; He, M.; Wang, B. Vanadium Pollution and Health Risks in Marine Ecosystems: Anthropogenic Sources over Natural Contributions. Water Res. 2021, 207, 117838. [Google Scholar] [CrossRef] [PubMed]
- Yeh, G.; Lin, C.; Nguyen, D.-H.; Hoang, H.-G.; Shern, J.-C.; Hsiao, P.-J. A Five-Year Investigation of Water Quality and Heavy Metal Mass Flux of an Industrially Affected River. Environ. Sci. Pollut. Res. 2022, 29, 12465–12472. [Google Scholar] [CrossRef] [PubMed]
- Tulcan, R.X.S.; Ouyang, W.; Guo, Z.; Lin, C.; Gu, X.; Wang, A.; Wang, B. Watershed Seasonality Regulating Vanadium Concentrations and Ecological Risks in the Coastal Aquatic Habitats of the Northwest Pacific. Environ. Pollut. 2023, 322, 121145. [Google Scholar] [CrossRef] [PubMed]
- Raven, J.A. The Iron and Molybdenum Use Efficiencies of Plant Growth with Different Energy, Carbon and Nitrogen Sources. New Phytol. 1988, 109, 279–287. [Google Scholar] [CrossRef]
- Wilhelm, C.; Wild, A. The Variability of the Photosynthetic Unit in Chlorella II. The Effect of Light Intensity and Cell Development on Photosynthesis, P-700 and Cytochrome f in Homocontinuous and Synchronous Cultures of Chlorella. J. Plant Physiol. 1984, 115, 125–135. [Google Scholar] [CrossRef]
- Gilmour, D.J.; Kaaden, R.; Gimmler, H. Vanadate Inhibition of ATPases of Dunaliella Parva in Vitro and in Vivo. J. Plant Physiol. 1985, 118, 111–126. [Google Scholar] [CrossRef]
- Nalewajko, C.; Lee, K.; Jack, T.R. Effects of Vanadium on Freshwater Phytoplankton Photosynthesis. Water. Air. Soil Pollut. 1995, 81, 93–105. [Google Scholar] [CrossRef]
- Meisch, H.-U.; Benzschawel, H. The Role of Vanadium in Green Plants: III. Influence on Cell Division of Chlorella. Arch. Microbiol. 1978, 116, 91–95. [Google Scholar] [CrossRef]
- Vaishampayan, A. VANADIUM AS A TRACE ELEMENT IN THE BLUE-GREEN ALGA, NOSTOC MUSCORUM : INFLUENCE ON NITROGENASE AND NITRATE REDUCTASE. New Phytol. 1983, 95, 55–60. [Google Scholar] [CrossRef]
- Arnon, D.I.; Wessel, G. Vanadium as an Essential Element for Green Plants. Nature 1953, 172, 1039–1040. [Google Scholar] [CrossRef]
- Patrick, R. Effects of Trace Minerals in the Aquatic Ecosystem. Am. Sci. 1978, 66, 185–191. [Google Scholar]
- Howarth, R.W.; Cole, J.J. Molybdenum Availability, Nitrogen Limitation, and Phytoplankton Growth in Natural Waters. Science 1985, 229, 653–655. [Google Scholar] [CrossRef]
- Wang, Y.; Zhu, D.; Li, C.; You, X.; Zhou, L.; Zhang, L.; Xiao, J.; Chen, M.; Ding, S.; Hang, X. Cyanobacterial Blooms Increase the Release of Vanadium through Iron Reduction and Dissolved Organic Matter Complexation in the Sediment of Eutrophic Lakes. Water Res. 2023, 243, 120377. [Google Scholar] [CrossRef]
- Schaule, B.K.; Patterson, C.C. Lead Concentrations in the Northeast Pacific: Evidence for Global Anthropogenic Perturbations. Earth Planet. Sci. Lett. 1981, 54, 97–116. [Google Scholar] [CrossRef]
- Reuer, M.K.; Weiss, D.J. Anthropogenic Lead Dynamics in the Terrestrial and Marine Environment. Philos. Trans. R. Soc. Lond. Ser. Math. Phys. Eng. Sci. 2002, 360, 2889–2904. [Google Scholar] [CrossRef]
- Lin, Y.-C.; Chang-Chien, G.-P.; Chiang, P.-C.; Chen, W.-H.; Lin, Y.-C. Multivariate Analysis of Heavy Metal Contaminations in Seawater and Sediments from a Heavily Industrialized Harbor in Southern Taiwan. Mar. Pollut. Bull. 2013, 76, 266–275. [Google Scholar] [CrossRef]
- Mu, W.; Chen, Y.; Liu, Y.; Pan, X.; Fan, Y. Toxicological Effects of Cadmium and Lead on Two Freshwater Diatoms. Environ. Toxicol. Pharmacol. 2018, 59, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Rzymski, P.; Poniedzialek, B.; Niedzielski, P.; Tabaczewski, P.; Wiktorowicz, K. Cadmium and Lead Toxicity and Bioaccumulation in Microcystis Aeruginosa. Front. Environ. Sci. Eng. 2014, 8, 427–432. [Google Scholar] [CrossRef]
- Deb, S.C.; Fukushima, T. Metals in Aquatic Ecosystems: Mechanisms of Uptake, Accumulation and release-Ecotoxicological Perspectives. Int. J. Environ. Stud. 1999, 56, 385–417. [Google Scholar] [CrossRef]
- Kamp-Nielsen, L. The Effect of Deleterious Concentrations of Mercury on the Photosynthesis and Growth of Chlorella Pyrenoidosa. Physiol. Plant. 1971, 24, 556–561. [Google Scholar] [CrossRef]
- Le Faucheur, S.; Campbell, P.G.C.; Fortin, C.; Slaveykova, V.I. Interactions between Mercury and Phytoplankton: Speciation, Bioavailability, and Internal Handling. Environ. Toxicol. Chem. 2014, 33, 1211–1224. [Google Scholar] [CrossRef]
- Protopopov, F.F.; Todorenko, D.A.; Nikolaev, I.N.; Alekseev, A.A.; Bratkovskaya, L.B.; Matorin, D.N. The Fluorescence of Phytoplankton Chlorophyll from the Moskva River in the Presence of Mercury Ions. Biophysics 2021, 66, 779–785. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, W.-X. Thiol Compounds Induction Kinetics in Marine Phytoplankton during and after Mercury Exposure. J. Hazard. Mater. 2012, 217–218, 271–278. [Google Scholar] [CrossRef]
- Satoh, M.; Matsumoto, Y. Mercury-Induced Oxidative Stress in Marine Phytoplankton Tetreselmis Tetrahele (Prasinophyceae). Nat. Sci. Res. 2008, 22, 57–63. [Google Scholar]
- Mason, R.P.; Morel, F.M.M.; Hemond, H.F. The Role of Microorganisms in Elemental Mercury Formation in Natural Waters. Water. Air. Soil Pollut. 1995, 80, 775–787. [Google Scholar] [CrossRef]
- Achá, D.; Guédron, S.; Amouroux, D.; Point, D.; Lazzaro, X.; Fernandez, P.E.; Sarret, G. Algal Bloom Exacerbates Hydrogen Sulfide and Methylmercury Contamination in the Emblematic High-Altitude Lake Titicaca. Geosciences 2018, 8, 438. [Google Scholar] [CrossRef]
- Pickhardt, P.C.; Folt, C.L.; Chen, C.Y.; Klaue, B.; Blum, J.D. Algal Blooms Reduce the Uptake of Toxic Methylmercury in Freshwater Food Webs. Proc. Natl. Acad. Sci. 2002, 99, 4419–4423. [Google Scholar] [CrossRef]
- Fendorf, S.; Michael, H.A.; van Geen, A. Spatial and Temporal Variations of Groundwater Arsenic in South and Southeast Asia. Science 2010, 328, 1123–1127. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Healey, F.P. Effects of Arsenate on Growth and Phosphorus Metabolism of Phytoplankton1,2. J. Phycol. 1978, 14, 337–341. [Google Scholar] [CrossRef]
- Sabath, E. 18 - Arsenic, Kidney, and Urinary Bladder Disorders. In Handbook of Arsenic Toxicology (Second Edition); Flora, S.J.S., Ed.; Academic Press: Oxford, 2023; pp. 485–500. ISBN 978-0-323-89847-8. [Google Scholar]
- Donohue, J. Some Comparisons among Ring Compounds of Phosphorus and Arsenic. Acta Crystallogr. 1962, 15, 708–713. [Google Scholar] [CrossRef]
- Nyulászi, L.; Veszprémi, T. Photoelectron Spectroscopic Study of the Aromaticity of Phosphorus and Arsenic Compounds. J. Mol. Struct. 1995, 347, 57–71. [Google Scholar] [CrossRef]
- Nava, P.; Ahlrichs, R. Theoretical Investigation of Clusters of Phosphorus and Arsenic: Fascination and Temptation of High Symmetries. Chem. – Eur. J. 2008, 14, 4039–4045. [Google Scholar] [CrossRef] [PubMed]
- Andreae, M. Arsenic Speciation in Seawater and Interstitial Waters: The Influence of Biological-chemical Interactions on the Chemistry of a Trace Element. Limnol. Oceanogr. 1979, 24, 440–452. [Google Scholar] [CrossRef]
- Hellweger, F.L.; Farley, K.J.; Lall, U.; Di Toro, D.M. Greedy Algae Reduce Arsenate. Limnol. Oceanogr. 2003, 48, 2275–2288. [Google Scholar] [CrossRef]
- Duan, L.; Song, J.; Zhang, Y.; Yuan, H.; Li, X.; Sun, L. Role of Marine Algal Blooms in the Release of Arsenic at the Sediment-Seawater Interface: Evidence from Microcosm Experiments. Water Res. 2023, 244, 120508. [Google Scholar] [CrossRef]
- Safonova, T.A.; Annenkov, V.V.; Chebykin, E.P.; Danilovtseva, E.N.; Likhoshway, Y.V.; Grachev, M.A. Aberration of Morphogenesis of Siliceous Frustule Elements of the Diatom Synedra Acus in the Presence of Germanic Acid. Biochem. Biokhimiia 2007, 72, 1261–1269. [Google Scholar] [CrossRef]
- Basharina, T.N.; Danilovtseva, E.N.; Zelinskiy, S.N.; Klimenkov, I.V.; Likhoshway, Y.V.; Annenkov, V.V. The Effect of Titanium, Zirconium and Tin on the Growth of Diatom Synedra Acus and Morphology of Its Silica Valves. Silicon 2012, 4, 239–249. [Google Scholar] [CrossRef]
- Sanville, W.D.; Powers, C.F.; Schuytema, G.S.; Stay, F.S.; Lauer, W.L. Phosphorus Inactivation by Zirconium in a Eutrophic Pond. J. Water Pollut. Control Fed. 1982, 54, 434–443. [Google Scholar]
- Couture, P.; Blaise, C.; Cluis, D.; Bastien, C. Zirconium Toxicity Assessment Using Bacteria, Algae and Fish Assays. Water. Air. Soil Pollut. 1989, 47, 87–100. [Google Scholar] [CrossRef]
- Pawlik-Skowrońska, B.; Kaczorowska, R.; Skowroński, T. The Impact of Inorganic Tin on the Planktonic Cyanobacterium Synechocystis Aquatilis: The Effect of pH and Humic Acid. Environ. Pollut. 1997, 97, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Yallop, M.; Wang, Y.; Masuda, S.; Daniels, J.; Ockenden, A.; Masani, H.; Scott, T.B.; Xie, F.; Ryan, M.; Jones, C.; et al. Quantifying Impacts of Titanium Dioxide Nanoparticles on Natural Assemblages of Riverine Phytobenthos and Phytoplankton in an Outdoor Setting. Sci. Total Environ. 2022, 831, 154616. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Juneau, P.; Lian, Y.; Zhang, W.; Wang, S.; Wang, C.; Shu, L.; Yan, Q.; He, Z.; Xu, K. Effects of Titanium Dioxide Nanoparticles on Photosynthetic and Antioxidative Processes of Scenedesmus Obliquus. Plants 2020, 9, 1748. [Google Scholar] [CrossRef]
- Kulacki, K.J.; Cardinale, B.J. Effects of Nano-Titanium Dioxide on Freshwater Algal Population Dynamics. PLOS ONE 2012, 7, e47130. [Google Scholar] [CrossRef]
Figure 1.
Global distribution of HAB events from 1980 to 2024. (Left) Map showing location of reported HAB events generated from IOC-Unesco Harmful Algae Information System. (Right) Number of reported HAB events and syndrome caused. Presented data include the following reported events-water discoloration, seafood toxins, coastal foam/mucilage, high phytoplankton concentration, and mass mortalities. Retrieved April 29, 2024, from https://data.hais.ioc-unesco.org/.
Figure 1.
Global distribution of HAB events from 1980 to 2024. (Left) Map showing location of reported HAB events generated from IOC-Unesco Harmful Algae Information System. (Right) Number of reported HAB events and syndrome caused. Presented data include the following reported events-water discoloration, seafood toxins, coastal foam/mucilage, high phytoplankton concentration, and mass mortalities. Retrieved April 29, 2024, from https://data.hais.ioc-unesco.org/.
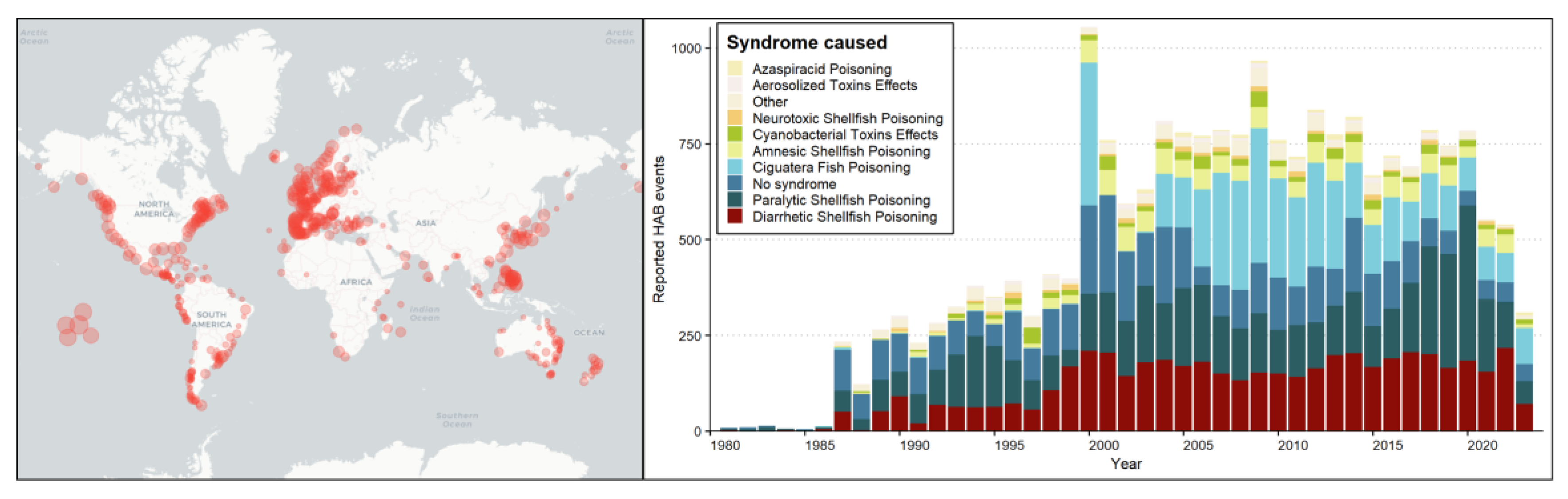

Table 1.
List of metals and associated metalloenzymes in photosynthetic microorganisms.
| Metal | Metalloenzyme | Metal | Metalloenzyme | |
|---|---|---|---|---|
|
Fe |
Fe-S proteins (e.g., aconitase) |
Mn |
Oxygen evolving enzymes | |
| Ferredoxin | Superoxide dismutase | |||
| Superoxide dismutase | Arginase | |||
| Nitrate reductase | Phosphotransferases | |||
| Nitrite reductase |
Cu |
Plastocyanin | ||
| Chelatase | Ferrous oxidase | |||
| Peroxidase | Amine oxidase | |||
|
Zn |
Carbonic anhydrase | Cytochrome oxidase | ||
| Alkaline phosphatase | Ascorbate oxidase | |||
| RNA polymerase | Superoxide dismutase | |||
| tRNA synthase |
Co |
Carbonic anhydrase | ||
| Reverse transcriptase | Cobalamin | |||
| Carboxypeptidase | Alkaline phosphatase | |||
| Superoxide dismutase | Se | Gluthathione peroxidase | ||
| Alcohol dehydrogenase | V | Bromoperoxidase | ||
| Aminopeptidase | Nitrogenase | |||
| Ni | Urease | Cd | Carbonic anhydrase | |
| Superoxide dismutase | Alkaline phosphatase |
Table 2.
List of research focusing on trace metals in specific HAB species in marine and brackish ecosystems.
Table 2.
List of research focusing on trace metals in specific HAB species in marine and brackish ecosystems.
| Species | Metals | Reference |
|---|---|---|
| Akashiwo sanguinea | Co (Vitamin B12) | [204] |
| Alexandrium | Zn | [205] |
| Alexandrium catenella | Cu, Sn (as butyltin) | [206] |
| Cu, Cd | [207] | |
| Zn, Pb | [208] | |
| Fe | [209] | |
| Co (Vitamin B12) | [204] | |
| Alexandrium minutum | Fe, Cu | [197] |
| Se | [210] | |
| Cu | [211] | |
| Co (Vitamin B12) | [204] | |
| Alexandrium tamarense | Fe | [212] |
| Amphidinium carterae | Cu | [213] |
| Aurecoccus anophagefferens | Zn, Cu, Ni, Cd | [214] |
| Se | [215] | |
| Co (Vitamin B12) | [204] | |
| Fe | [216] | |
| Aureoumbra lagunensis | Co (Vitamin B12) | [204] |
| Chaetoceros compressus | Cu | [217] |
| Chatonella antiqua | Fe | [218] |
| Chatonella marina | Zn, Cu, Cd, Pb, Hg | [219] |
| Co (Vitamin B12) | [204] | |
| Chatonella ovata | Zn, Cu, Cd, Pb, Hg | [219] |
| Choomonas | Cu, As | [220] |
| Cochlodinium polykrikodes | Cu | [221,222] |
| Cu and Pb | [223] | |
| Co (Vitamin B12) | [204] | |
| Dinophysis acuminata | Cu | [217] |
| Fibrocasa japonica | Co (Vitamin B12) | [204] |
| Gymnodinium aureolum | Co (Vitamin B12) | [204] |
| Gymnodinium breve | Cu, Ti, Zr | [224] |
| Gymnodium catenatum | Se | [210,225] |
| Fe, Cu, Se | [226] | |
| Zn, Cu, Cd, Pb, Hg | [219] | |
| Gymnodinium instriatum | Co (Vitamin B12) | [204] |
| Gymnodinium nagasakiense | Se | [227,228] |
| Gymnodinium sanguineum | Fe | [229,230] |
| Zn, Cu, Cd, Pb, Hg | [219] | |
| Heterosigma akashiwo | Fe | [218] |
| Heterocapsa artica | Co (Vitamin B12) | [204] |
| Heterocapsa circularisquama | Fe | [218] |
| Heterocapsa triquetra | Cu | [217] |
| Co (Vitamin B12) | [204] | |
| Karenia brevis | Fe, Cu, Co, Cd, Hg | [201] |
| Karenia mikimotoi | Co (Vitamin B) | [204] |
| Karenia selliformis | Mn, Zn, Cu, Co, Se | [231] |
| Karlodinium veneficum | Co (Vitamin B12) | [204] |
| Katodinium rotundatum | Cu, As | [220] |
| Lingulodium polyedrum | Fe | [209] |
| Cd | [232] | |
| Mo | [233] | |
| Ostreopsis cf. ovata | Cu, Sn (as butyltin) | [206] |
| Ostreopsis siamensis | Mn, Zn, Cu, Co, Se | [231] |
| Phaeocystis globosa | Co (Vitamin B12) | [204] |
| Polykrikos hartmannii | Co (Vitamin B12) | [204] |
| Protoceratium reticulatum | Mn, Zn, Cu, Co, Se | [231] |
| Fe, Co, Se | [234] | |
| Prorocentrum dentatum | Fe | [235] |
| Prorocentrum donghaiense | Co (Vitamin B12) | [204] |
| Prorocentrum lima | Mn, Zn, Cu, Co, Se | [231] |
| Prorocentrum micans | Cu | [213] |
| Prorocentrum minimum | Co (Vitamin B12) | [204] |
| Pseudo-nitzschia sp. | Fe, Cu | [175,198] |
| Pseudo-nitzschia australis | Mn, Zn, Cu, Co, Se | [231] |
| Fe, Cu | [236] | |
| Pseudo-nitzschia delicatissima | Fe, Cu | [237] |
| Cu | [238] | |
| Pseudo-nitzschia multiseries | Fe | [239] |
| Cu | [240,241] | |
| Co (Vitamin B12) | [204] | |
| Pseudo-nitzschia pungens | Co (Vitamin B12) | [204] |
| Rhizosolenia setigera | Cu | [217] |
| Scrippsiella trochoidea | Fe | [242] |
| Co (Vitamin B12) | [204] | |
| Skeletonema costatum | Fe | [235] |
| Takayama acrotrocha | Co (Vitamin B12) | [204] |
| Thalassiosira pseudonana | Cu, As | [220] |
| Trichodesmium | Ni | [190,243] |
Table 3.
List of research focusing on trace metals in specific HAB species in freshwater ecosystems.
Table 3.
List of research focusing on trace metals in specific HAB species in freshwater ecosystems.
| Species | Metals | Reference |
|---|---|---|
| Anabaena lemmermannii | Cu | [199] |
| Anabaenopsis sp. | Cu | [244] |
| Aphanizomenon schindlerii | Fe | [245] |
| Aphanizonmenon sp. | Cu | [199] |
| Chlamydomonas reinhardtii | Cu | [199,246] |
| Cylindrospermopsis raciboskii | Cu | [199] |
| Closterium ehrenbergii | Cu | [247] |
| Desmodesmus | Cu, Fe | [248] |
| Lyngbya wollei | Cu | [249] |
| Microcystis sp. | Cu, Fe | [248] |
| Zn | [250] | |
| Microcystis aeruginosa | Fe, Zn, Al, Cd, Cr, Cu, Mn, Ni, and Sn | [251] |
| Co, Cu, Fe, Mn, Mo | [252] | |
| Ca, Cu, Pb, Cd | [253] | |
| Fe, Mn, As | [254] | |
| Fe, Zn | [255] | |
| Fe, Cu | [200] | |
| Cu, Zn | [202,256] | |
| Zn, Cd | [257] | |
| Ni | [258] | |
| Fe | [259,260,261] | |
| Cu | [199,202,203,244,262,263,264,265] | |
| Zn | [266,267] | |
| As | [268] | |
| Oscillatoria sp | Cu | [199] |
| Pseudokirchneriella subcapitata | Cu | [199] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



