
Submitted:
17 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
Portal myofibroblasts (PMF) are known to be critical in bile duct injury, but their role in liver fibrogenesis remains underexplored. Thy-1, an adhesion molecule integral to fibroblasts, relevant in fibrogenesis and detectable in serum as soluble Thy-1 (sThy-1), is expressed by PMF but not hepatic stellate cells in vitro. This study aimed to elucidate the correlation between Thy-1 expression in liver histology and sThy-1 levels in serum with the degree of liver fibrosis in patients with primary biliary cholangitis (PBC) and metabolic dysfunction associated steatotic liver disease (MASLD).
Liver histology samples were stained for Thy-1 and sThy-1 was measured using an enzyme-linked immunosorbent assay (ELISA).
In patients with PBC and MASLD, portal Thy-1 expression significantly correlated with increased fibrosis. The sinusoidal Thy-1 expression also significantly correlated with increased fibrosis in patients with PBC (p
Keywords:
liver fibrosis
; primary biliary cholangitis
; metabolic dysfunction-associated liver disease
; Thy-1
; soluble Thy-1
1. Introduction
Liver fibrosis is the common endpoint of various chronic liver injuries and critically determines the prognosis of chronic liver diseases. Despite its significance, the mechanisms driving liver fibrogenesis remain incompletely understood, as evidenced by the current lack of effective anti-fibrotic treatments. Myofibroblasts are central to the fibrotic process in all fibrotic diseases, functioning as matrix-producing cells with contractile properties and expressing proteins common to smooth muscle cells, such as alpha-smooth muscle actin (SMA) [1,2]. Hepatic stellate cells (HSC) and portal myofibroblasts (PMF) are the two primary myofibroblast sources in the liver [3,4,5]. While the role of HSC in fibrogenesis has been studied more extensively, the importance of PMF has only more recently come to attention [6]. PMF have been proposed to deposit matrix specifically in biliary fibrosis and to have a role in place of, or in addition to, hepatic stellate cell-derived myofibroblasts [5,7]. Therefore, PMF and HSC are supposed to responding uniquely to specific stimuli and performing distinct functions. However, the role of PMFs remains poorly defined because of a lack of markers clearly distinguishing this cell population from HSCs at the stage of myofibroblasts [1,6,8,9].
Thymocyte differentiation antigen 1 (Thy-1, or CD90), known for its presence on various cell types and its diverse roles in cellular biology, emerges as a pivotal molecule in fibrogenesis. Its presence or absence has significant effects on cellular biology, including the control of many core functions of fibroblasts relevant to fibrogenesis, including the deposition of extracellular matrix, proliferation, apoptosis, cytokine and growth factor expression and responsiveness, cell adhesion, migration, and myofibroblast differentiation [10]. However, the effects vary between cell types, tissues and organs, and between similar tissues in different species [11], with pro-fibrogenic functions of Thy1+ fibroblasts observed in some studies [12] and anti-fibrogenic effects in others [13,14].
Within the context of liver fibrogenesis, Thy-1 has been identified as a marker for liver PMF, particularly in the periportal area, distinguishing them from HSC [1,15,16,17], and has been identified as a gene that is overexpressed in PMFs [15], pointing to its relevance in liver fibrogenesis. On the other hand, a more recent study investigated the role of mesothelin and Thy-1 in the activation of fibroblasts across multiple organs and demonstrated that mesothelin -/- mice were protected from cholestatic fibrosis but Thy-1 -/- mice were more susceptible to fibrosis, suggesting that the mesothelin-Thy-1 signaling complex is critical for tissue fibroblasts [18]. Whether these findings translate to in vivo circumstances in humans is yet to be seen.
The shedding of Thy-1 from fibroblasts surfaces, correlating with a shift towards a profibrogenic myofibroblast phenotype, has been a focus of recent studies [19,20]. An enzyme-linked-immunosorbent-assay (ELISA) for detecting soluble Thy-1 (sThy-1) has revealed elevated levels in systemic fibrotic diseases, correlating with disease severity [21]. However, the relevance of sThy-1 in liver diseases remains unexplored.
Primary biliary cholangitis (PBC) and metabolic dysfunction-associated steatotic liver disease (MASLD) offer unique insights into liver fibrosis. PBC, a rare chronic inflammation of small biliary ducts, shows periportal fibrosis that expands to sinusoids with disease progression [22].
MASLD, a complex metabolic disorder characterized by excessive hepatocyte fat accumulation leading to steatohepatitis with a hepatocellular pattern of liver injury, shows perisinusoidal fibrosis that expands to portal fields with disease progression [23]. It has become a leading cause of cirrhosis in the western world. In the context of PBC and MASLD, two diseases with distinct fibrosis patterns, this study aims to elucidate Thy-1’s presence in liver histology and serum across different fibrosis stages, seeking new insights into mechanisms of liver fibrogenesis.
2. Patients and Methods
2.1. Patients and Samples
This retrospective monocentric study was conducted at the University Hospital of Bern, a tertiary care center in Switzerland. We included adult patients who received medical care between 2011 and 2022 with a diagnosis of PBC. Eligibility criteria included written informed consent and no concurrent chronic liver diseases. For the control group, we selected 50 patients with MASLD, ensuring a similar fibrosis grading distribution to the PBC patients. Patient samples were sourced from the Hepatology Biobank at the University Hospital Bern, Switzerland. All patients underwent liver biopsy concurrent with or within two years of their blood sample collection. Diagnosis criteria for PBC followed the guidelines of the European Association for the Study of the Liver [24]. Similarly, MASLD diagnosis conformed to the guidelines of the European Association for the Study of the Liver [23]. Definition of portal hypertension was either hepatic venous pressure gradient ≥ 5 mmHg, dilatation of the portal venous system on sonography, splenomegaly, platelets <150 G/L or oesophageal varices. Control samples from healthy individuals matched the age and sex distribution of the PBC patients and were obtained from the blood donation center.
2.2. Laboratory Methods
For laboratory analysis, sThy-1 levels were measured using an enzyme-linked immunosorbent assay (ELISA) as described previously [25]. Liver samples underwent staining using a Thy-1 antibody (CD90 [5E10] from BD Bioscience #550402) on a BOND Research RX staining platform from Leica Biosystems. The antibody dilution was 1:100, with a 30-minute incubation at room temperature and a 30-minute heat-induced epitope retrieval in Tris EDTA buffer at 95 °C. Additionally, an anti-alpha-smooth muscle actin antibody (SMA) was used as a marker for myofibroblasts. The SMA antibody [1A4] from Sigma #A2547 was used on a BOND Max III in KPATH Diagnostics, with a dilution of 1:8000 and a 15-minute room temperature incubation without heat-induced epitope retrieval. For fibrosis assessment, Masson’s trichome staining was employed. All tissue samples were evaluated by an experienced liver pathologist using a semi-quantitative approach based on four staining intensity categories: negative, weak, moderate, and strong (Supplementary Figure 1). The degree of fibrosis in PBC was categorized according to METAVIR. The degree of fibrosis in MASLD was categorized using the Brunt and EPOS classification systems [26,27]. Patients were categorized into three disease groups based on fibrosis stages: Mild fibrosis (METAVIR F0/1 and Brunt 1/EPOS 1), moderate fibrosis (METAVIR F2 and Brunt 2/EPOS 2 and Brunt 3 if EPOS 3) and advanced fibrosis (METAVIR F3/4 and Brunt 3 if EPOS 4, Brunt 4/EPOS 5-6).
2.3. Statistical Analysis
Categorical data are presented as numbers and percentages, while continuous data are expressed as median and interquartile range (IQR). We used the Chi-square test or Fisher’s exact test for comparing categorical data, depending on the data distribution. For quantitative data, the Wilcoxon rank sum test or the Kruskal-Wallis test was applied. We adjusted the alpha level for paired comparisons using the Bonferroni correction. Associations were assessed with ordinal logistic regression, reported as odds ratio (OR) and 95% confidence intervals (CI). Brant’s test verified the proportionality of odds in our model. A logarithmic transformation of sThy-1 measurements was applied to approximate a normal distribution, minimize the impact of outliers and enhance interpretability. A p-value < 0.05 (two-tailed) was considered statistically significant. All analyses were performed using R software, Version 4.1.2.
Clinical data were missing at random, as most, but not all parameters were available for all patients at the time point of sampling. Missing data is indicated separately in the results section.
2.4. Ethical Approval
The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Cantonal Ethics Commission of Bern, Switzerland (approval number 2021-02237). All participants provided informed consent.
3. Results
3.1. Patient Characteristics
3.1.1. Study Inclusion
Among the 2809 patients in the Hepatology Biobank, 69 with PBC met the inclusion criteria and 49 MASLD patients were selected as controls to match the fibrosis distribution of the PBC group. The median time between liver biopsy and blood sample collection was 0 days (0;0) for PBC and MASLD patients.
3.1.2. Baseline Characteristics
Table 1 presents the baseline characteristics of the study population at the time of sample collection. As healthy controls were matched to PBC patients, these two groups were predominantly female, differing significantly from the MASLD group. PBC patients exhibited more pronounced cholestasis and higher immunoglobulin M levels, both of which are characteristic features of this disease. MASLD patients had a higher baseline body mass index as well as ferritin levels, indicative of liver inflammation. M30-Apoptosens, a marker of hepatocyte apoptosis, was similarly elevated in both groups. INR was normal in both groups, however, significantly lower in the PBC group in comparison to the MASLD group. There were no significant differences in bilirubin, liver stiffness measurements, histological fibrosis classification, or the presence of portal hypertension between the two groups (Table 1).
3.2. Liver Histology
We were not able to retrieve and reevaluate liver biopsies of two patients with PBC and advanced fibrosis, three patients with MASLD and mild fibrosis and one patient with MASLD and advanced fibrosis, hence, from these patients we were only able to analyze the blood samples.
3.2.1. Thy-1 Expression in PBC and MASLD Patients
Figure 1 illustrates a significant correlation between liver fibrosis and Thy-1 expression intensity for PBC (Figure 1A) and MASLD (Figure 1B) patients. In PBC, a higher likelihood of advanced fibrosis is associated with increased Thy-1 expression (OR 10.37, 95% CI 4.30-30.36, p<0.001), while in MASLD, this correlation is significant but with a lower effect size (OR 3.8, 95% CI 1.63-10.02, p=0.004). Adjusting for sex and age at baseline did not alter these findings (Table 2).
3.2.2. Localization of Thy-1 According to Fibrosis Stages
Figure 2 shows the portal and sinusoidal localization of the Thy-1 expression in relation to fibrosis grading. In PBC, Thy-1 expression was present in the portal field across all fibrosis stages. In those with mild fibrosis, 85.7% (18 patients) exhibited portal Thy-1 expression, increasing to 95.5% (21 patients) in the moderate fibrosis group, and reaching 100% (24 patients) in the advanced fibrosis group (Figure 2A). As the Thy-1 expression was already high in the mild fibrosis group, the correlation with the increase in fibrosis grading was not significant (OR 8.36, 95% CI 1.01-171.78, p=0.068). Notably, the four patients lacking portal Thy-1 expression only demonstrated weak overall Thy-1 intensity. Conversely, sinusoidal Thy-1 expression in PBC was observed in 19% (four patients) with mild fibrosis, 31.8% (seven patients) with moderate fibrosis, and 70.83% (17 patients) with advanced fibrosis (Figure 2C). This association was statistically significant (OR 6.05, 95% CI 2.3-17.11, p<0.001), indicating an increase in sinusoidal Thy-1 expression with advancing fibrosis.
In MASLD, the detection of Thy-1 in the portal area was present in 44.4% (four patients) in patients with mild fibrosis, increasing to 86.4% (19 patients) in moderate fibrosis, and 100% (14 patients) in advanced fibrosis, as shown in Figure 2B. It is important to note that the eight patients who did not exhibit portal Thy-1 expression demonstrated only weak Thy-1 intensity. The association between portal Thy-1 expression and fibrosis grading was significant (OR 15.78, 95% CI 3.10-103.73, p=0.002). Sinusoidal Thy-1 expression, was present in 11.1% (one patient) with mild fibrosis, 18.2% (four patients) with moderate fibrosis, and 42.8% (six patients) with advanced fibrosis (Figure 2D). This relationship between fibrosis grading and sinusoidal Thy-1 expression in MASLD was not statistically significant (OR 3.73, 95% CI 0.98-15.55, p=0.059). The associations did not change relevantly if adjusted for sex and age (Table 2).
One patient did not have a portal SMA positivity. All other patients had SMA expression portal and sinusoidal. Therefore, the Thy-1 expression portal and sinusoidal is also reflecting the co-localization of Thy-1 with SMA.
3.3. Soluble Thy-1
A logarithmic transformation of sThy-1 measurements was applied to approximate a normal distribution, minimize the impact of outliers and enhance interpretability. The original measurements are available in supplementary Table 1.
3.3.1. Soluble Thy-1 in Serum in Patients with PBC and MASLD According to Fibrosis Groups
As illustrated in Figure 3, soluble Thy-1 levels in serum significantly differ across fibrosis stages within PBC and MASLD groups. Advanced fibrosis in PBC is linked to the highest sThy-1 levels (p=0.043). No significant differences were observed between the other fibrosis groups, as detailed in Figure 3 and Supplementary Table 2. However, the relationship between sThy-1 levels and fibrosis severity was statistically significant (OR 3.762, 95% CI 1.49-10.93, p=0.008), a finding that remained consistent after adjusting for sex and age (OR 3.613, 95% CI 1.403-10.696, p=0.012).
For MASLD patients, significant variations in median sThy-1 levels across fibrosis stages were observed, with the most pronounced levels in the advanced fibrosis group (8.455 pg/ml [8.255; 8.618]), as illustrated in Figure 3 and Supplementary Table 2. This association between sThy-1 levels and fibrosis grading was notably strong (OR 10.661, 95% CI 2.46-64.75, p=0.005), and remained significant after adjustments for sex and age (OR 9.277, 95% CI 1.86-64.75, p=0.013).
No significant variance in sThy-1 levels was found between PBC and MASLD patients across comparable fibrosis stages (data not shown).
3.3.2. Soluble Thy-1 in Serum in Patients with PBC and MASLD According to Thy-1 Intensity on Liver Histology
In PBC, while median sThy-1 levels did not show significant differences among the various Thy-1 staining intensities on liver histology, as depicted in Figure 4 and detailed in Supplementary Table 2, the overall trend across these groups revealed a significant association between sThy-1 levels and Thy-1 staining intensity (OR 2.833, 95% CI 1.11-7.77, p=0.034), which remained significant after adjusting for sex and age (OR 2.799, 95% CI 1.08-7.79, p=0.040).
Conversely, in MASLD patients, no significant difference in median sThy-1 levels was observed across Thy-1 intensity groups on liver histology (Figure 4 and Supplementary Table 2), and similarly, the overall association between sThy-1 levels and Thy-1 staining intensity did not reach statistical significance (OR 2.688, 95% CI 0.71-12.47, p=0.167), a finding that persisted upon adjustment for sex and age (OR 2.202, 95% CI 0.47-12.57, p=0.338).
4. Discussion
Our study reveals a significant expansion of Thy-1 positive cells in liver fibrogenesis, notably in biliary liver injuries, underscoring the nuanced roles of these cells across different liver diseases.
Patients with PBC more often were females, showed a more pronounced cholestasis as well as elevated immunoglobulin M levels than patients with MASLD, all of which are typical demographic and laboratory features of PBC [22]. As we matched the healthy controls regarding age and sex to the PBC controls, there is a significantly higher proportion of females in the healthy controls in comparison to the MASLD group, with no difference, however, regarding age at baseline. As expected, patients with MASLD had a significantly higher body mass index at baseline as well as higher ferritin levels, which is an indicator of liver inflammation in MASLD [28]. Importantly, there was no significant difference between the level of apoptosis, fibrosis and portal hypertension between PBC and MASLD patients. These findings indicate that we had two typical disease groups with similar fibrosis stages, which is central to the interpretation of the results.
Despite PBC being a disease with a biliary type of injury and MASLD a disease with a hepatocellular type of injury, the correlation between an increase in fibrosis grading and an increase in the intensity of Thy-1 expression on liver histology was significant in both diseases. Our findings corroborate those of Dezsὅ et al. (34), who identified Thy-1 decorated myofibroblasts in models of cholangiofibrosis and hepatotoxic cirrhosis, and extended these observations by demonstrating the significant involvement of Thy-1-positive cells even in the early stages of liver fibrogenesis [29]. Complementing Lemoinne et al. [1], who emphasized the role of PMFs in fibrogenesis without using Thy-1 as a marker, our study not only confirms the increase of PMFs as fibrosis progresses but also establishes Thy-1 expression as a distinct marker for these cells in liver tissue samples from PBC and MASLD patients.
In patients with PBC the Thy-1 expression was portal in almost all patients, regardless of fibrosis grading. In patients with MASLD, however, the portal expression of Thy-1 significantly increased with fibrosis progression. This might indicate that portal Thy-1 positive cells are activated earlier in the context of a biliary type of injury, even in situations, where fibrosis is not yet pronounced.
The sinusoidal presence of Thy-1 positive cells was clearly associated with the severity of fibrosis in patients with PBC, indicating, that Thy-1 positive cells are expanding in the liver during fibrogenesis. This is in line with the data from rat models by Dudas et al., which suggest, that Thy-1 may induce myofibroblast migration along the liver sinusoids [15]. In MASLD patients, although the sinusoidal presence of Thy-1 expressing cells increased with fibrosis progression, this was observed in a smaller proportion of patients and did not reach statistical significance. Dudas et al. had similar findings in a CCL4-induced liver injury model in rats, where Thy-1 reaction was seen primarily in the periportal area and not significantly sinusoidal [15]. These findings are furthermore in line with the observation from Iwaisako et al., who found that activated PMFs were shown to comprise 70% of myofibroblasts at the onset of the bile duct ligation mouse model of cholestatic fibrosis, while HSCs were increasingly activated with fibrosis progression [9]. Our findings might therefore indicate that also in humans the Thy-1 positive fibroblasts are more specifically activated and involved in the process of fibrogenesis in patients with a biliary type of liver injury. Given that Thy-1 expression correlates with fibrosis severity, targeting the Thy-1 signaling pathways or modulating Thy-1 expression could offer new strategies for fibrosis treatment. However, the exact regulatory capacities of Thy-1 positive cells, whether fibrotic or anti-fibrotic [11], remain to be fully understood. Our findings underscore the urgency for further research to explore these pathways, potentially leading to targeted therapies that can mitigate fibrosis in liver diseases.
SMA was present portal and sinusoidal in all patients and all fibrosis stages, with the exception of one sinusoidal area. Thy-1 however, was present portal and sinusoidal depending on fibrosis grading and disease subgroup. These results are in line with previous findings by Dezsὅ [29] and Dudas and colleagues [15], who described SMA staining exceeding Thy-1 staining. In the work from Lemoinne et al. described above [1] the COL15A1 positive cells, identified as PMFs, also expressed SMA in cirrhotic livers, but not in normal livers, with SMA staining largely exceeding that of COL15A1, which is in line with our findings, with the limitation, that we used another marker for PMFs. This only partial co-localization with SMA indicates that Thy-1 positive cells are only a subpopulation of myofibroblasts in the liver, which is consistent with previous findings, that HSC are the major contributors to the entire population of hepatic myofibroblasts [6].
Soluble Thy-1 measurements significantly differed between early and advanced fibrosis in both diseases, with, interestingly, a higher significance level in patients with MASLD. The mechanism of shedding of Thy-1 is not yet understood well [11] and it can be hypothesized, that it is influenced by different types of inflammatory activities in the liver and therefore might be more pronounced in MASLD. Of note, despite the observed association between sThy-1 levels and fibrosis grading, the variability of sThy-1 levels suggests limitations in its utility as a non-invasive surrogate marker for liver fibrosis. Several factors could contribute to this variability, including the heterogeneous nature of liver diseases and the complex biology of Thy-1, which may have differential expression patterns and shedding mechanisms under varying pathological conditions [11]. Further research is necessary to elucidate these mechanisms and evaluate the potential of sThy-1 in a broader spectrum of liver diseases.
In the correlation between the intensity of Thy-1 seen in liver histology and the sThy-1 levels, we could not find significant differences between the staining intensities and the sThy-1 levels in MASLD and PBC patients with, however a significant association in the regression in patients with PBC.
Our data indicates that Thy-1 positive cells are expanding in the liver during fibrogenesis, more significantly in patients with a biliary type of liver injury, but also in patients with a hepatocellular type of liver injury. Furthermore, we have shown that the release of sThy-1 from Thy-1 positive cells is present but our data suggests that it is not relevant enough for it to be a non-invasive surrogate marker for liver fibrosis.
Thus, our research not only aligns with existing findings regarding the role in fibrogenesis of Thy-1-positive cells but also advances our understanding by delineating their activity across different liver disease contexts and its correlation to sThy-1.
Strengths and Limitations of the Study
Our study enhances the understanding of Thy-1’s role in liver fibrosis through comprehensive analysis of Thy-1 expression in liver histologies and sThy-1 levels, providing novel insights across various stages of liver fibrosis. To our knowledge, this represents the largest study of patients with PBC and MASLD analyzed for Thy-1 expression in the liver and the first study where histological Thy-1 expression and soluble Thy-1 measurements were conducted simultaneously in humans.
The significance of our study is further underscored by the rarity of PBC as a liver disease and the challenges inherent in measuring sThy-1 in experimental models. Enzymatic digestion during cell isolation can lead to the detachment of the Thy-1 molecule, potentially reassociating with other cells and yielding misleading results in cell suspension [29].
A limitation of our study is the absence of further characterization of Thy-1 positive cells with additional markers to deepen our understanding of these cells’ roles in fibrogenesis. Additionally, the observed variability in sThy-1 measurements underscores the need for further validation of sThy-1 as a biomarker. However, these limitations pave the way for future research directions, including the detailed study of Thy-1 signaling pathways and the exploration of sThy-1 in a wider array of liver conditions. Embracing these opportunities can significantly advance our understanding and treatment of liver fibrosis.
5. Conclusions
Our findings underscore the pivotal role of Thy-1 positive cells in liver fibrogenesis, with a marked presence in biliary types of liver injury. Despite the observed shedding of sThy-1, its variability suggests limitations for its use as a non-invasive marker for fibrosis. These insights not only contribute to our understanding of liver fibrosis mechanisms but also highlight the critical need for further research to explore the therapeutic potential targeting Thy-1. Identifying modulatory pathways that could be specifically targeted offers a promising avenue for developing novel treatments for liver fibrosis.
Abbreviations
CI, 95% confidence interval; ELISA, enzyme-linked immunosorbent assay; HC, healthy controls; IgM, immunoglobulin M; MASLD, metabolic dysfunction-associated steatotic liver disease; ns, not significant; OR, odds ratio; PBC, primary biliary cholangitis; SMA, smooth muscle actin antibody; sThy-1, soluble Thy1
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org., Supplementary Table 1. Soluble Thy-1 measurements in relation to fibrosis groups and Thy-1 intensities in patients with PBC and MASLD. HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis. Supplementary Table 2 . Significance levels comparing median soluble Thy-1 levels in the different fibrosis groups and according to Thy-1 intensities in patients with PBC and MASLD. Calculations were done with the Wilcoxon rank sum test. The alpha level for the paired comparisons was adjusted applying the Bonferroni correction. * indicates a significant p-value (<0.05). Supplementary Figure 1. Thy-1 staining on liver tissue. The analysis was semi-quantitative with four categories of staining intensity: negative, weak, moderate and strong.
Authors’ Contributions
KM: conception and design of the study, data management, acquisition of data, analysis and interpretation of data, drafting the article, final approval of the submitted version. SA: laboratory analysis, revising article critically for important intellectual content, final approval of the submitted version. BR: acquisition of data. KF: conception, revising article critically for important intellectual content, final approval of the submitted version. MM: conception and design of the study, histological analysis. SN: conception and design of the study, interpretation of data, revising article critically for important intellectual content, final approval of the submitted version.
Financial disclosure
KM has financial support from the Swiss Liver Foundation. The funding source had no influence in the writing of the report.
Acknowledgments
KM thanks the Swiss Liver Foundation for supporting her work. We thank Mattia Branca from the Clinical Trials Unit, University of Bern, Switzerland for the statistical support.
Declaration of competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Lemoinne S, Thabut D, Housset C. Portal myofibroblasts connect angiogenesis and fibrosis in liver. Cell Tissue Res. 2016;365(3):583-9. [CrossRef]
- Lemoinne S, Cadoret A, El Mourabit H, Thabut D, Housset C. Origins and functions of liver myofibroblasts. Biochim Biophys Acta. 2013;1832(7):948-54. [CrossRef]
- Friedman SL. Hepatic stellate cells: protean, multifunctional, and enigmatic cells of the liver. Physiol Rev. 2008;88(1):125-72. [CrossRef]
- Ramadori G, Saile B. Mesenchymal cells in the liver--one cell type or two? Liver. 2002;22(4):283-94.
- Kinnman N, Francoz C, Barbu V, Wendum D, Rey C, Hultcrantz R, et al. The myofibroblastic conversion of peribiliary fibrogenic cells distinct from hepatic stellate cells is stimulated by platelet-derived growth factor during liver fibrogenesis. Lab Invest. 2003;83(2):163-73. [CrossRef]
- Wells RG. The portal fibroblast: not just a poor man’s stellate cell. Gastroenterology. 2014;147(1):41-7. [CrossRef]
- Dranoff JA, Wells RG. Portal fibroblasts: Underappreciated mediators of biliary fibrosis. Hepatology. 2010;51(4):1438-44. [CrossRef]
- Hinz B, Phan SH, Thannickal VJ, Prunotto M, Desmouliere A, Varga J, et al. Recent developments in myofibroblast biology: paradigms for connective tissue remodeling. Am J Pathol. 2012;180(4):1340-55. [CrossRef]
- Iwaisako K, Jiang C, Zhang M, Cong M, Moore-Morris TJ, Park TJ, et al. Origin of myofibroblasts in the fibrotic liver in mice. Proc Natl Acad Sci U S A. 2014;111(32):E3297-305. [CrossRef]
- Hu P, Leyton L, Hagood JS, Barker TH. Thy-1-Integrin Interactions in cis and Trans Mediate Distinctive Signaling. Front Cell Dev Biol. 2022;10:928510. [CrossRef]
- Bradley JE, Ramirez G, Hagood JS. Roles and regulation of Thy-1, a context-dependent modulator of cell phenotype. Biofactors. 2009;35(3):258-65.
- Koumas L, Smith TJ, Feldon S, Blumberg N, Phipps RP. Thy-1 expression in human fibroblast subsets defines myofibroblastic or lipofibroblastic phenotypes. Am J Pathol. 2003;163(4):1291-300. [CrossRef]
- Sanders YY, Kumbla P, Hagood JS. Enhanced myofibroblastic differentiation and survival in Thy-1(-) lung fibroblasts. Am J Respir Cell Mol Biol. 2007;36(2):226-35. [CrossRef]
- Li Y, Song D, Mao L, Abraham DM, Bursac N. Lack of Thy1 defines a pathogenic fraction of cardiac fibroblasts in heart failure. Biomaterials. 2020;236:119824. [CrossRef]
- Dudas J, Mansuroglu T, Batusic D, Ramadori G. Thy-1 is expressed in myofibroblasts but not found in hepatic stellate cells following liver injury. Histochem Cell Biol. 2009;131(1):115-27. [CrossRef]
- Dudas J, Mansuroglu T, Batusic D, Saile B, Ramadori G. Thy-1 is an in vivo and in vitro marker of liver myofibroblasts. Cell Tissue Res. 2007;329(3):503-14. [CrossRef]
- Katsumata LW, Miyajima A, Itoh T. Portal fibroblasts marked by the surface antigen Thy1 contribute to fibrosis in mouse models of cholestatic liver injury. Hepatol Commun. 2017;1(3):198-214. [CrossRef]
- Nishio T, Koyama Y, Liu X, Rosenthal SB, Yamamoto G, Fuji H, et al. Immunotherapy-based targeting of MSLN(+) activated portal fibroblasts is a strategy for treatment of cholestatic liver fibrosis. Proc Natl Acad Sci U S A. 2021;118(29). [CrossRef]
- Shahin AA, Anwar S, Elawar AH, Sharaf AE, Hamid MA, Eleinin AA, et al. Circulating soluble adhesion molecules in patients with systemic sclerosis: correlation between circulating soluble vascular cell adhesion molecule-1 (sVCAM-1) and impaired left ventricular diastolic function. Rheumatol Int. 2000;20(1):21-4. [CrossRef]
- Hagood JS, Prabhakaran P, Kumbla P, Salazar L, MacEwen MW, Barker TH, et al. Loss of fibroblast Thy-1 expression correlates with lung fibrogenesis. Am J Pathol. 2005;167(2):365-79. [CrossRef]
- Kollert F, Christoph S, Probst C, Budweiser S, Bannert B, Binder M, et al. Soluble CD90 as a potential marker of pulmonary involvement in systemic sclerosis. Arthritis Care Res (Hoboken). 2013;65(2):281-7. [CrossRef]
- European Association for the Study of the Liver. Electronic address eee, European Association for the Study of the L. EASL Clinical Practice Guidelines: The diagnosis and management of patients with primary biliary cholangitis. J Hepatol. 2017;67(1):145-72.
- European Association for the Study of the L, European Association for the Study of D, European Association for the Study of O. EASL-EASD-EASO Clinical Practice Guidelines for the Management of Non-Alcoholic Fatty Liver Disease. Obes Facts. 2016;9(2):65-90.
- European Association for the Study of the Liver. Electronic address eee, European Association for the Study of the L. EASL 2017 Clinical Practice Guidelines on the management of hepatitis B virus infection. J Hepatol. 2017;67(2):370-98.
- Saalbach A, Wetzig T, Haustein UF, Anderegg U. Detection of human soluble Thy-1 in serum by ELISA. Fibroblasts and activated endothelial cells are a possible source of soluble Thy-1 in serum. Cell Tissue Res. 1999;298(2):307-15.
- Pierre Bedossa JA, et al. The EPoS staging system is a reproducible 7-tier fibrosis score for NAFLD adapted both to glass slides and digitized images (e-slides). J Hepatol. 2018;68.
- Kleiner DE, Brunt EM, Van Natta M, Behling C, Contos MJ, Cummings OW, et al. Design and validation of a histological scoring system for nonalcoholic fatty liver disease. Hepatology. 2005;41(6):1313-21. [CrossRef]
- Kowdley KV, Belt P, Wilson LA, Yeh MM, Neuschwander-Tetri BA, Chalasani N, et al. Serum ferritin is an independent predictor of histologic severity and advanced fibrosis in patients with nonalcoholic fatty liver disease. Hepatology. 2012;55(1):77-85. [CrossRef]
- Dezso K, Jelnes P, Laszlo V, Baghy K, Bodor C, Paku S, et al. Thy-1 is expressed in hepatic myofibroblasts and not oval cells in stem cell-mediated liver regeneration. Am J Pathol. 2007;171(5):1529-37. [CrossRef]
Figure 1.
Intensity of Thy-1 expression on liver histology in patients with PBC and MASLD. Significant correlation between the Thy-1 intensity and the fibrosis grading in A) patients with PBC (OR 10.37, 95% CI 4.30-30.36, p <0.0001) and B) patients with MASLD (OR 3.8, 95% CI 1.63-10.02, p=0.004). MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis.
Figure 1.
Intensity of Thy-1 expression on liver histology in patients with PBC and MASLD. Significant correlation between the Thy-1 intensity and the fibrosis grading in A) patients with PBC (OR 10.37, 95% CI 4.30-30.36, p <0.0001) and B) patients with MASLD (OR 3.8, 95% CI 1.63-10.02, p=0.004). MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis.
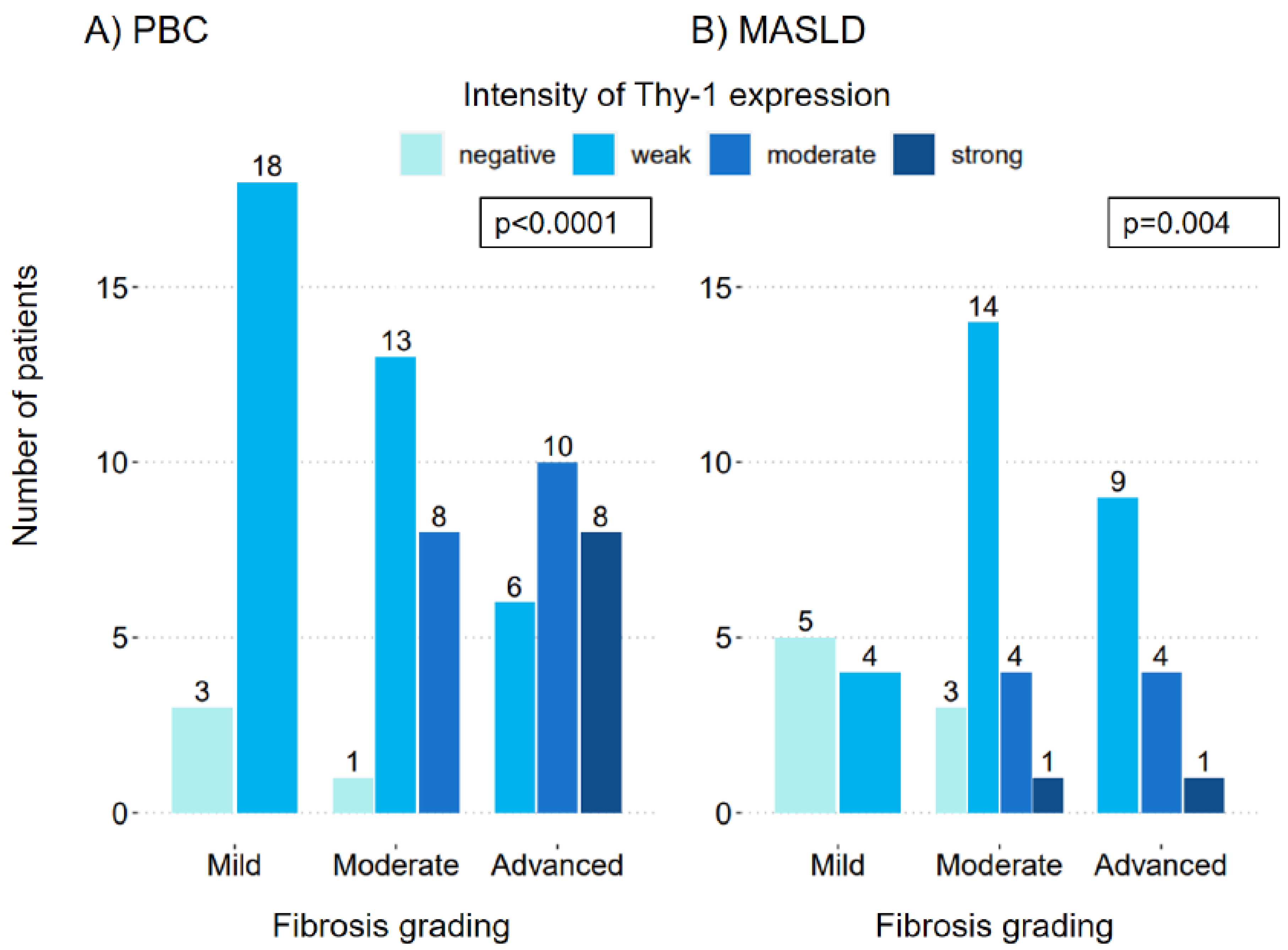
Figure 2.
Localization of Thy-1 expression on liver histology. Portal Thy-1 expression was present in most patients with PBC (OR 8.36, CI 1.01-171.78, p=0.068) (A) and MASLD (OR 15.78, CI 3.10-103.73, p=0.002) (B) even in patients with mild fibrosis. Sinusoidal expression of Thy-1 was significantly more present in patients with advanced fibrosis in patients with PBC (OR 6.05, CI 2.30-17.11, p<0.001) (C), but not significantly correlated in patients with MASLD (OR 3.73, CI 0.98-15.55, p=0.059) (D). MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis; SMA, smooth muscle actin antibody.
Figure 2.
Localization of Thy-1 expression on liver histology. Portal Thy-1 expression was present in most patients with PBC (OR 8.36, CI 1.01-171.78, p=0.068) (A) and MASLD (OR 15.78, CI 3.10-103.73, p=0.002) (B) even in patients with mild fibrosis. Sinusoidal expression of Thy-1 was significantly more present in patients with advanced fibrosis in patients with PBC (OR 6.05, CI 2.30-17.11, p<0.001) (C), but not significantly correlated in patients with MASLD (OR 3.73, CI 0.98-15.55, p=0.059) (D). MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis; SMA, smooth muscle actin antibody.
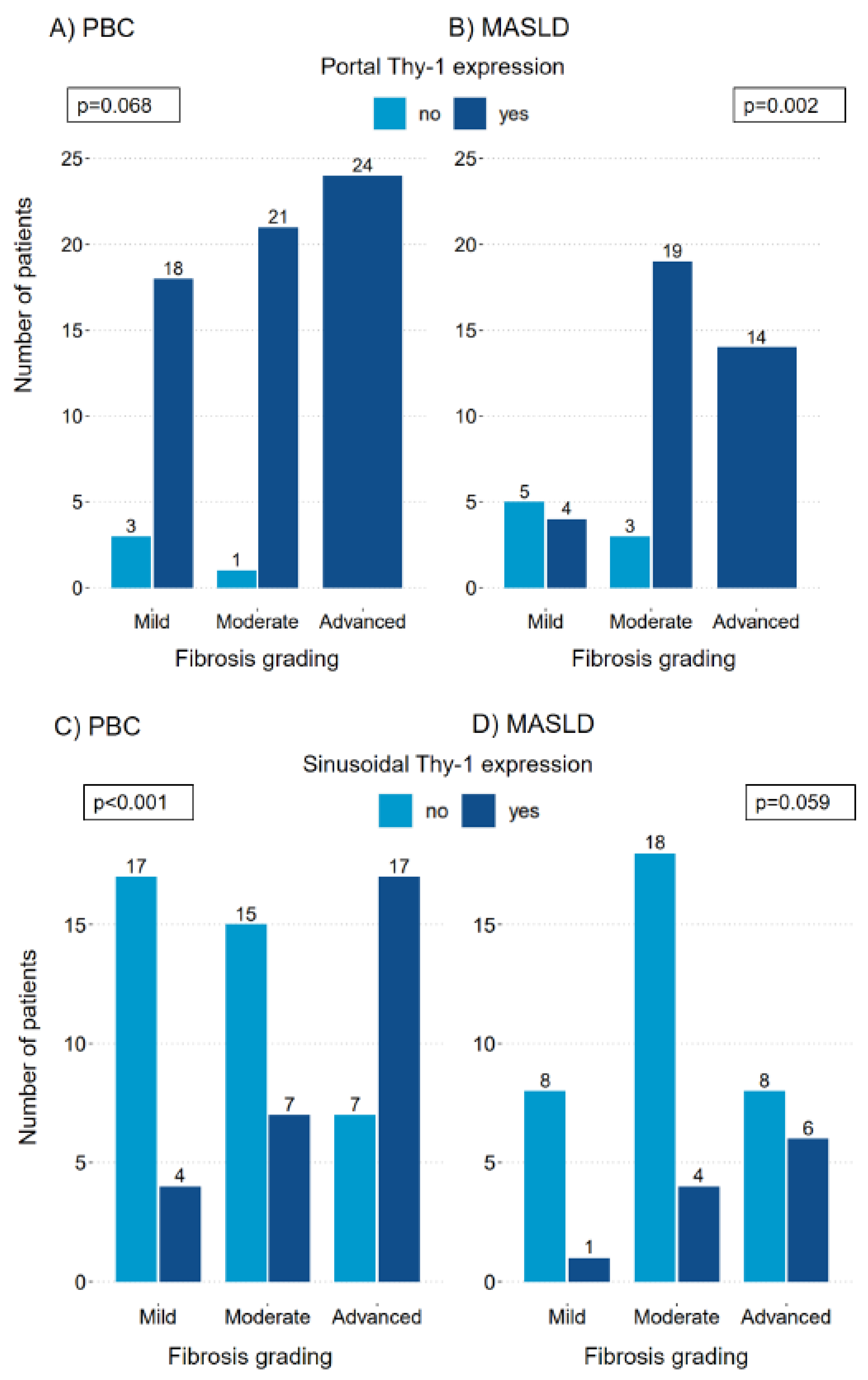
Figure 3.
Transformed soluble Thy-1 measurements in relation to fibrosis groups in patients with PBC and MASLD. The median (IQR25-IQR75) sThy-1 measurements according to fibrosis groups in patients with PBC (A) and MASLD (B). Significant differences between groups are indicated. ns indicates a value ≥0.05, * indicates a p-value of <0.05, ** indicates a p-value <0.01, *** indicates a p-value <0.001. HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; ns, not significant; PBC, primary biliary cholangitis.
Figure 3.
Transformed soluble Thy-1 measurements in relation to fibrosis groups in patients with PBC and MASLD. The median (IQR25-IQR75) sThy-1 measurements according to fibrosis groups in patients with PBC (A) and MASLD (B). Significant differences between groups are indicated. ns indicates a value ≥0.05, * indicates a p-value of <0.05, ** indicates a p-value <0.01, *** indicates a p-value <0.001. HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; ns, not significant; PBC, primary biliary cholangitis.
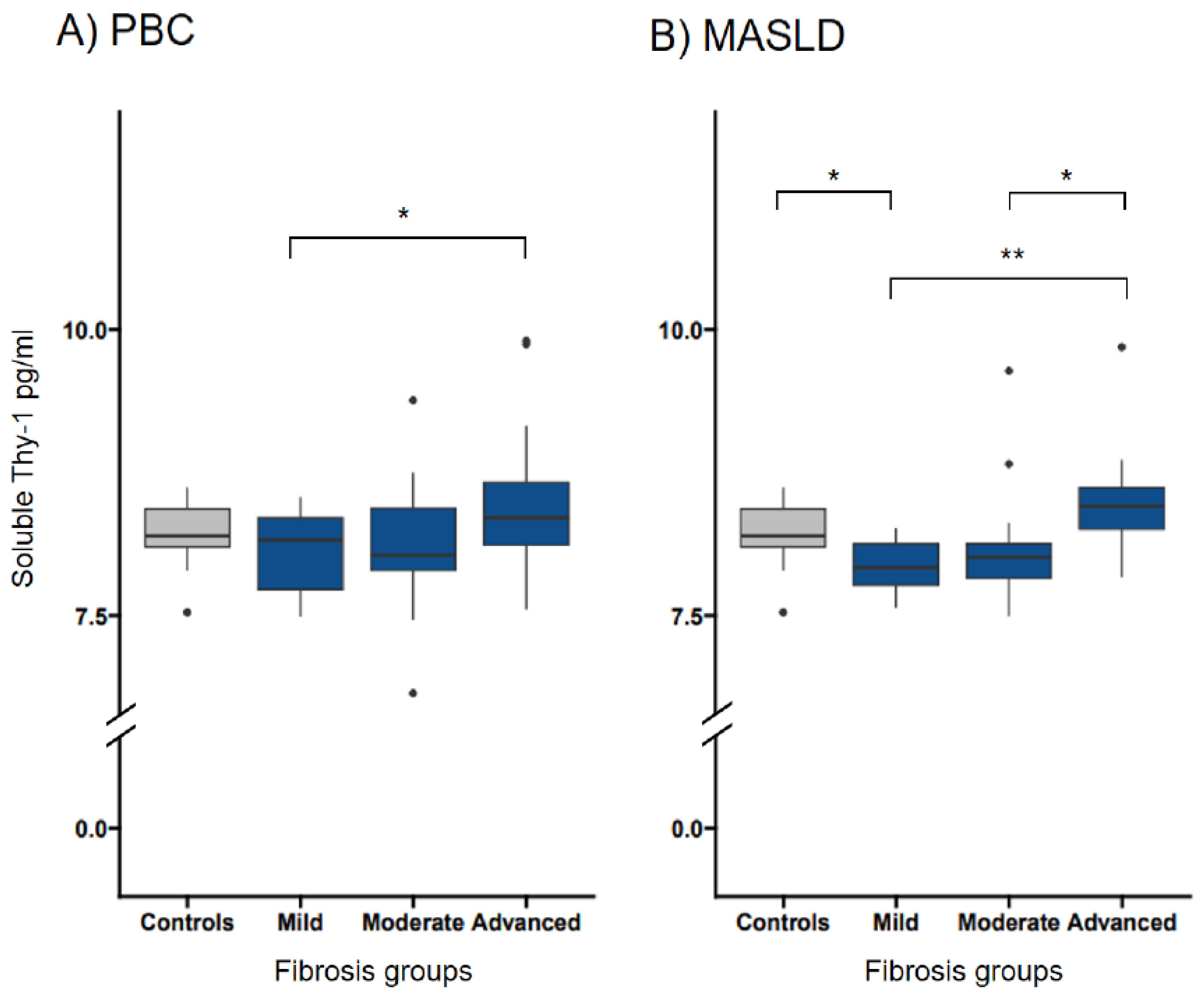
Figure 4.
Transformed soluble Thy-1 measurements in relation to Thy-1 intensities in patients with PBC and MASLD. The median (IQR25-IQR75) sThy-1 measurements according to Thy-1 intensities in patients with PBC (A) and MASLD (B). Significant differences between groups are indicated. ns indicates a value ≥0.05,* indicates a p-value of <0.05, ** indicates a p-value <0.01, *** indicates a p-value <0.001. HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; ns, not significant; PBC, primary biliary cholangitis.
Figure 4.
Transformed soluble Thy-1 measurements in relation to Thy-1 intensities in patients with PBC and MASLD. The median (IQR25-IQR75) sThy-1 measurements according to Thy-1 intensities in patients with PBC (A) and MASLD (B). Significant differences between groups are indicated. ns indicates a value ≥0.05,* indicates a p-value of <0.05, ** indicates a p-value <0.01, *** indicates a p-value <0.001. HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; ns, not significant; PBC, primary biliary cholangitis.

Table 1.
Baseline characteristics from patients with primary biliary cholangitis, metabolic dysfunction-associated steatotic liver disease and healthy controls at the time point of sample acquisition.
Table 1.
Baseline characteristics from patients with primary biliary cholangitis, metabolic dysfunction-associated steatotic liver disease and healthy controls at the time point of sample acquisition.
| PBC n=69 |
MASLD n=49 |
Healthy controls n=20 |
Missing values PBC/MASLD/HC |
p-value | |
|---|---|---|---|---|---|
| Demographics | |||||
| Female sex, n (%) | 57 (82.6) | 19 (38.8) | 18 (90) | 0(0)/0(0)/0(0) | <0.001* |
| Age at baseline, years | 59 (46-66) | 55.0 (44.0-60.0) | 53.5 (49.5-56.3) | 0(0)/0(0)/0(0) | 0.091 |
| Body mass index, kg/m2 | 26.3 (23-30) | 30.7 (27.8-35.0) | 0(0)/0(0)/20(100) | <0.001* | |
| Liver assessment | |||||
| ALT, x ULN | 1.7 (1-2.5) | 1.3 (0.9-2.0) | 0(0)/0(0)/20(100) | 0.203 | |
| Alkaline phosphatase, x ULN | 1.7 (1.1-2.8) | 0.7 (0.5-1.0) | 0(0)/0(0)/20(100) | <0.001* | |
| gGT, x ULN | 5.5 (3.2-9.5) | 1.5 (0.9-3.5) | 0(0)/0(0)/20(100) | <0.001* | |
| Bilirubin total, μmol/l (<17 μmol/l) | 11 (8-16) | 11 (7.75-14) | 0(0)/1(2)/20(100) | 0.541 | |
| INR (<1.2) | 1 (1-1.04) | 1.03 (1-1.1) | 0(0)/0(0)/20(100) | 0.008* | |
| Immunoglobulin M g/l (0.4-2.3 g/l) | 3.18 (2-4.87) | 1.42 (0.76-1.75) | 2(2.9)/23(47)/20(100) | <0.001* | |
| Ferritin µg/l (10-120 µg/l) | 109 (65.5-202) | 286 (130-547) | 26(37.7)/4(8.2)/20(100) | 0.002* | |
| M30-Apoptosens, U/L (<200) | 287 (183-423) | 280.5 (172-567.8) | 48(69.6)/17(34.7)/20(100) | 0.848 | |
| Fibroscan at baseline, kPa | 7.95 (4.95-14.8) | 8.8 (6.4-15.38) | 3(4.4)/5(10.2)/20(100) | 0.273 | |
| Signs of portal hypertension, n (%) | 15 (21.7) | 11 (22.4) | 0(0)/0(0)/20(100) | 1.000 | |
| Histological fibrosis grading | |||||
| Mild fibrosis, n (%) | 21 (30.4) | 12 (24.5) | 0(0)/0(0)/20(100) | 0.616 | |
| Moderate fibrosis, n (%) | 22 (31.9) | 22 (44.9) | 0(0)/0(0)/20(100) | 0.212 | |
| Advanced fibrosis, n (%) | 26 (37.6) | 15 (30.6) | 0(0)/0(0)/20(100) | 0.550 |
Definition of portal hypertension: hepatic venous pressure gradient ≥ 5 mmHg, dilatation of the portal venous system on sonography, splenomegaly, platelets <150 G/L or oesophageal varices. Mild fibrosis=METAVIR F0/1 or Brunt 1/EPOS 1; moderate fibrosis=METAVIR F2 or Brunt 2/EPOS 2 and Brunt 3 if EPOS 3; advanced fibrosis=METAVIR F3/4 or Brunt 3 if EPOS 4, Brunt 4/EPOS 5-6. Categorical variables presented as n (%). Continuous data presented as median and interquartile range (IQR25-IQR75). * indicates a significant p-value (<0.05). HC, healthy controls; MASLD, metabolic dysfunction-associated steatotic liver disease; PBC, primary biliary cholangitis.
Table 2.
Assessment of the correlation between fibrosis grading and Thy-1 intensity using ordinal logistic regression.
Table 2.
Assessment of the correlation between fibrosis grading and Thy-1 intensity using ordinal logistic regression.
| Odds ratio | p-value | CI | |
|---|---|---|---|
| Primary biliary cholangitis | |||
| Association between fibrosis grading and the following parameters | |||
| Thy-1 expression overall | 10.37 | <0.001* | 4.30-30.36 |
| Thy-1 expression overall adjusted | 10.78 | <0.001* | 4.42-32.04 |
| Thy-1 expression portal | 8.36 | 0.068 | 1.01-171.78 |
| Thy-1 expression portal adjusted | 8.42 | 0.071 | 1.07-175.40 |
| Thy-1 expression sinusoidal | 6.05 | <0.001* | 2.30-17.11 |
| Thy-1 expression sinusoidal adjusted | 6.48 | <0.001* | 2.42-18.67 |
| Metabolic dysfunction-associated steatotic liver disease | |||
| Association between fibrosis grading and the following parameters | |||
| Thy-1 expression overall | 3.8 | 0.004* | 1.63-10.02 |
| Thy-1 expression overall adjusted | 3.46 | 0.008* | 1.45-9.32 |
| Thy-1 expression portal | 15.78 | 0.002* | 3.10-103.73 |
| Thy-1 expression portal adjusted | 16.84 | 0.002* | 3.12-118.61 |
| Thy-1 expression sinusoidal | 3.73 | 0.059 | 0.98-15.55 |
| Thy-1 expression sinusoidal adjusted | 3.06 | 0.148 | 0.69-14.86 |
There was no change in significance after adjusting for sex and age at baseline. * indicates a significant p-value (<0.05). CI, 95% confidence interval; SMA, smooth muscle actin antibody.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



