
Submitted:
16 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
Senna rugosa is a species found in the Cerrado and used in folk medicine as a vermifuge and in the treatment of poisonous snakebites accidents. In this work, we identified the main secondary metabolites present in ethanolic extracts of the leaves (ELSR) and roots (ERSR) of S. rugosa and evaluated the potential cytoprotective effect against cellular macromolecular damage, as well as the cytotoxic properties of the extracts on the K562 and Jurkat leukemic cell lines. The identification of metabolites was done by liquid chromatography coupled with mass spectrometry. The antioxidant activities were investigated by direct ABTS•+ and DPPH• radical scavenging methods, protection against oxidative damage in proteins and DNA. The cytotoxic properties were investigated against healthy cells, isolated from human peripheral blood (PBMC) and leukemic cell lines. The leaf extract contains catechin, rutin, epigallocatechin derivatives, kaempferol glycosides, luteolin, and dimeric and trimeric procyanidins, while the root extract profile shows obtusichromoneside derivatives, 2-methoxystypandrone, stilbene derivatives, naphthopyranones, and flavanone derivatives. The extracts showed antioxidant activity, with IC50 of 4.86 ± 0.51 µg/mL and 8.33 ± 0.90 µg/mL in the ABTS assay for ELSR and ERSR, respectively. And in the DPPH• assay, the IC50 was 19.98 ± 1.96 µg/mL for ELSR and 13.37 ± 1.05 µg/mL for ERSR. The extracts protected macromolecules against oxidative damage at concentrations of 5 µg/mL. The cytotoxicity test against leukemic strains was observed after 24 and 48 hours of treatment. After 48 h, results against the K562 cell line demonstrate IC50 of 242.54 ± 2.38 µg/mL and 223.00 ± 2.34 µg/mL for ELSR and ERSR , respectively. While against the Jurkat cell line, these extracts showed IC50 of 171.45 ± 2.25 µg/mL and 189.30 ± 2.27 µg/mL, respectively Results on PBMC viability, demonstrate that the extracts showed selectivity for the leukemic cell lines. Together, our results reveal that the leaves and roots of S. rugosa have completely distinct and complex chemical compositions. This study expands the repertoire of identified chemical compounds and underscores their significant pharmacological potential in conditions related to oxidative stress and leukemia.
Keywords:
Natural Products
; Anticancer Properties
; Cancer
; medicinal plants
; Chemical profile
1. Introduction
Oxidative stress is characterized by an imbalance between reactive species and the endogenous antioxidant system [1,2]. The oxidative damage on macromolecules - proteins and nucleic acids, accentuates cellular aging and reduces the efficiency of DNA repair mechanisms, increasing the accumulation of mutations [3,4], associated with the development of pathologies such as cancer, among others [5]. The deleterious effects of oxidative stress can be controlled by antioxidants, conferring protection against reactive species of endogenous or exogenous origin [6,7]. The beneficial effects of antioxidants on physiology are widely known [8]. Furthermore, antioxidants can promote pro-oxidant effects in cancer cells by activating different cell death mechanisms [9,10].
Leukemia is a hematologic neoplasm affecting the hematopoietic system's cells [11]. Four main subtypes of leukemias are known and diagnosis varies according to the maturity and type of cell lineage [12]. In many cases of leukemias, translocations between chromosomes 9 and 22 are observed, leading to the fusion of the BCR and ABL1 genes. This fusion results in increased tyrosine-kinase activity [13] and the activation of several signaling pathways, such as Jak/Stat and Ras/Raf/Map-kinase, resulting in uncontrolled events during the cell cycle [14]. Lack of control over cell cycle progression can also occur due to silencing of suppressor (p53 and PTEN) and pro-apoptotic genes (BAX) [15].
Despite advances in the treatment of leukemia, about 20% of patients have a relapse rate of the disease or adverse reactions to treatment, highlighting the importance of developing new therapies [11,16]. Consequently, a high mortality rate is recorded. In 2020, more than 19 million cases occurred worldwide, with a mortality rate of 70% [17]. In this regard, the scientific community has intensified the search for new therapeutic agents with anticancer activity of natural origin. Recent studies have demonstrated the pharmacological potential of plants from the Brazilian Cerrado, a biome with the potential for discovering new phytochemicals [18].
Recent pharmacological studies have shown that the crude extracts or derivatives of the Senna genus possess, among others, antidiabetic, antioxidant, anti-inflammatory, antitumor, and anticancer activities [19,20,21]. Several studies on medicinal plants from Cerrado have presented crude plant extracts with anticancer properties [22,23,24,25]. The extracts of these species presented a varied composition of secondary metabolites, among them gallic acid, catechin, epigallocatechin, epicatechin, kaempferol heteroside, rutin, ellagic acid, anthraquinone, piceatanol, dimeric and trimeric proanthocyanidins. Several studies have demonstrated the effect of these compounds on their cytotoxic effects on cancer cells [26]. Senna rugosa (Fabaceae) is a plant species popularly known as amendoirana, alcacuz bravo, bico-de-corvo, boi gordo and paratudo, where its seeds are used to treat parasitic worms’ infestations [27,28] and its roots are used in the treatment of poisonous snake bites [29]. Recently, Cunha et al., [30] evaluated the chemical composition of the ethanolic extract of S. rugosa leaves and fractions obtained from the extract and identified the presence of mainly phenolic compounds, which include flavonols, anthraquinones and anthrones derivatives (particularly, rutin, emodin, aloe-emodin and barbaloin/isobarbaloin). In that study, antimicrobial, antifungal, and moderate antitumor activities against breast cancer cell lines were reported. There are reports that several species of the Senna genus possess anticancer activities against various tumor cell lines [21,31].
As the species has been used by the population in the treatment of pathologies, we proposed to determine the chemical constitution of the metabolites present in the leaves and roots of S. rugosa. In addition, we investigated the antioxidant properties and cytotoxic activities of the ethanolic extracts of S. rugosa leaves (ELSR) and roots (ERSR) against chronic (K562) and acute (Jurkat) human leukemia cell lines. We used human peripheral blood cells (PBMC) in this study to determine the selectivity of the extracts on leukemic cells compared to healthy human cells.
2. Results
This section may be divided by subheadings. It should provide a concise and precise description of the experimental results, their interpretation, as well as the experimental conclusions that can be drawn.
2.1. Chemical Composition of ELSR
The detailed phytochemical analysis of S. rugosa, conducted separately on its leaf extracts (Table 1 - ELSR) and root extracts (Table 1 - ERSR), revealed distinct chemical profiles between these two plant organs. In the leaf extracts, we identified a variety of flavan-3-ol dimers and trimers, epiafzelechin derivatives, and a range of flavonoids, including rutin, luteolin, and kaempferol 3-O-rutinoside, indicating a marked predominance of phenolic and flavonoids derivatives. In contrast, the root extracts presented a widely varied secondary metabolic, ranging from C-glycosylated chromones, notably of the obtusichromoneside class, to glycosylated naphthopyranones with skeletons compatible with rubrofusarin derivatives, stilbenoids, and possible glycosylated and sulfated flavonoids. The chromatographic profile is shown in Figure 1, while the retention times, molecular formula, and M+H are in Table 1.
2.2. Antioxidant Activity Assays
Table 2 presents the IC50 values for the ABTS•+ and DPPH• radicals, as well as the concentrations at which the ELSR and ERSR showed the maximum antioxidant activity. For the ABTS•+ radical, ELSR presented IC50 1.25 times lower when compared with the reference antioxidant BHT, presenting maximum activity at the same concentrations (50 µg/mL). ERSR presented IC50 1.37 times higher than BHT, however, its maximum activity was reached at half the concentration of BHT.
In the direct DPPH• radical scavenging assay, the IC50 of EFRS and ERSR were 3.5 and 5.3 times lower, respectively, than that calculated for BHT, demonstrating a higher antioxidant activity of the extracts compared to this reference compound. Both extracts showed maximum activity at concentrations 5 times lower than BHT.
2.3. Protection of ELSR and ERSR against Oxidative Damage in Macromolecules
ELSR and ERSR protected BSA protein against AAPH-induced oxidation at all concentrations evaluated (Figure 2). The images of the gels are representative and demonstrate the protection against oxidation after ELSR and ERSR treatments. Both extracts reduced protein oxidation, resulting from AAPH exposure when compared to the AAPH control, where BSA was incubated with AAPH alone.
ELSR and ERSR were able to protect DNA from H2O2 UV-induced fragmentation in a concentration-dependent manner (Figure 3-B). Figure 3A is a representative picture of the protection against fragmentation after ELSR and ERSR treatments. In the absence of extracts and at low concentrations (up to 25 µg/mL) fragmentation of plasmid DNA is observed. Reduction of fragmentation is visible from 50 µg/mL onwards. The reference antioxidants rutin and catechin caused a reduction in DNA fragmentation, however, rutin was more effective in the evaluated concentration.
2.4. Cytotoxicity against Leukemic Strains
PBMC cells and K562 and Jurkat leukemic cell lines were treated with different concentrations of the ELSR and ERSR for 24 and 48 h. Incubation of the ELSR and ERSR (Figure 4 A–D) with PBMC showed a minimal reduction in the cell viability of 17% on average, but only at the highest concentration of the extracts. None of the concentrations of the extracts caused a 50% reduction in the PBMC viability, suggesting that the extracts are selective to the leukemic cells. The result with the solvent control (0.1% ethanol) demonstrated that the cells' viability under these conditions was unaffected by the solvent. The results show that the extracts showed a cytotoxic effect against the leukemic cell lines in a concentration-dependent manner, with the K562 cell line being less sensitive than the Jurkat cell line, as observed through the IC50 values presented in Table 3.
3. Discussion
The phytochemical constituent profiles of ELSR and ERSR were investigated using HPLC techniques to identify relevant secondary metabolite groups and useful chemical markers. These multi-constituent profile markers are used for quality control of herbal preparations and subsequent validity checks.
An interesting observation in the present study is the high divergence observed in the phytochemical constituents identified between ELSR and ERSR. Besides, the identification for the first time of pharmacologically active metabolites from the roots of S. rugosa. This divergence in phytochemical profiles not only underscores the existence of a unique chemical diversity between the distinct parts of the plant but also reflects the intricate specialized metabolic networks characterizing the leaf and root tissues of S. rugosa.
The chemical profile of ELSR was characterized predominantly by the accumulation of flavan-3-ol derivatives, notably catechin (L-1). This compound, distinguished by its characteristic UV absorption at 280 nm and a molecular ion [M+H]+ at m/z 291.0870, was confirmed through comparison with an authentic standard. Proanthocyanidins, including derivatives like epiafzelechin-epiafzelechin (L-4, L-5, L-6) and trihydroxyflavan-epiafzelechin derivatives (L-8, L-9, L-10, L-12), highlight the plant ability to synthesize complex polymeric flavonoids. These compounds were annotated by their molecular masses and distinctive MS/MS fragmentation patterns.
The flavonols, such as Rutin (L-2) and Kaempferol 3-O-rutinoside (L-7), were identified by their UV spectra, MS/MS fragmentation patterns, and comparison with an authentic standard. Flavones like Luteolin (L-11) and its methoxylated derivative, 3-methoxyluteolin (L-13), underscore the plant's ability to synthesize a wide variety of flavonoid structures.
The stilbenoid, 3,3',5,5'-Tetrahydroxy-4-methoxystilbene (L-3), was detected at a low intensity and annotated by its UV absorption and molecular formula. This class has been reported multiple times within the genus [32].
Furthermore, a tetrahydroanthracene derivative (L-16), representing a less common class, was putatively characterized by its molecular weight and MS/MS fragments. It is possibly novel in the literature; due to the absence of standards, we were unable to determine its chemical structure. We recently identified a compound of this class in S. velutina [20].
The ERSR extract from the roots displayed a metabolite accumulation distinctly different from that observed in the leaves. Chromones were prominently featured, evidenced by the annotation of obtusichromoneside derivatives (R-2, R-3, R-7) and a chromone derivative (R-7), characterized by their UV spectra and mass fragmentation patterns. These compounds have previously been reported in other species of Senna and Cassia, a closely related taxon [33].
The analysis also highlighted a significant presence of naphthoquinone and naphthopyrones, with compounds such as 2-methoxystypandrone (R-4), a putative norrubrofusarin gentiobioside (R-8) (a possible novel compound), rubrofusarin gentiobioside (R-10), cassiaside B (R-12), and rubrofusarin-O-glucopyranoside (R-14) demonstrating the root's capability to synthesize complex naphtho-derived structures. These findings are particularly interesting due to the rare occurrence of such compounds in nature and their potential pharmacological applications.
The only compound observed in both the leaves and roots was R-6 (equivalent to L-3 in the leaf), but it demonstrated low chromatographic intensity, consistent with its observation in leaf extracts. Additionally, the detection of Flavanones and their derivatives, including hexahydroxy Flavanonol pentosyl-hexosyl derivatives (R-9, R-13), a putatively annotated hexahydroxy Flavanonol sulfate (R-11), and hexahydroxy-methoxy flavanonol sulfate (R-16), indicates a distinctive flavonoid accumulation pattern in the leaves and roots of S. rugosa. Some of these compounds, such as R-11 and R-15 do not match any structures described in the literature, suggesting the possibility that they are novel compounds. The structural diversity within this class highlights the intricate enzymatic processes involved in flavonoid modification.
Seven compounds of the roots remained unidentified (R-1, R-15, R-17, R-18, R-20, R-21, R-22), presenting opportunities for further research. These unknown entities, detected through their unique mass spectra and UV profiles, hint at the unexplored chemical space within S. rugosa, suggesting the presence of novel or rare metabolites awaiting discovery.
In summary, this detailed phytochemical investigation significantly enhances our comprehension of the chemical diversity present in S. rugosa, underscoring the plant's extensive biosynthetic capacities. It sets a foundation for future research aimed at elucidating the pharmacological properties and ecological significance of these compounds, while also indicating the potential for discovering novel metabolites within this species.
Interestingly, the antioxidant and anticancer activities of most of these compounds identified in ELSR, including their molecular mechanisms have already been proven in several studies. Notably, several in vitro, in vivo, and clinical studies have shown multiple anticancer actions of epigallocatechin, and a review of their potential therapeutic targets and their role in the therapy of various cancers have been the motive of several studies [34,35,36]. These include anti-proliferative, pro-apoptotic, anti-angiogenic, and anti-invasive activities. Another compound found in ELSR is rutin, which has shown ubiquitous pharmacological properties, including antioxidant, anti-inflammatory, antiangiogenic, pro-apoptotic, and antiproliferative activities, all of which may participate in the prevention and treatment of cancer [36].
In the report by Cunha et al., [37] several peaks were observed in the hydroethanolic extract, but only one compound was identified (rutin), making comparison with our results impossible. Thus, we report the presence of different phytochemicals not previously described for this species.
In the ERSR, several pharmacologically important compounds were also identified. Of the twenty-two peaks, 6 could not be identified based on the current technique employed. Among the identified compounds in ERSR were the chromones derivatives, particularly the obtusichromoneside derivatives. Several chromones derivatives have been credited with significant biological and pharmacological activities, including antioxidant, antiviral, anti-inflammatory, antitumor, antimicrobial, among others [38,39,40]. Duan, et al. [39] presented a comprehensive review of the antitumor activities of naturally occurring chromones. The antitumor mechanisms of these chromones include cytotoxicity (cell-cycle arrest, and cell death, RNA inhibition, and inhibition of signal transducers), antimetastatic, anti-angiogenesis, and immune regulation, among others. Moreover, some have been shown to possess cancer chemoprevention potential. Another equally important secondary metabolites found in ERSR are the naphthoquinones. These groups of naturally occurring bioactive compounds have been the subject of intense research on anticancer and antioxidant activities. [41,42]. The radical scavenging, antioxidant, and anticancer activities of these group of compounds are widely known. These compounds together may act in synergy to account for the antioxidant and prevent oxidative damage to the macromolecules employed in this study.
Genomic mutation of DNA is an essential component in the process of cancer development. Oxidative damage and the consequent modification of DNA bases can lead to point mutations, deletions, insertions, or chromosomal translocations that, in turn, can produce oncogene activation or tumor suppressor gene inactivation [43,44]. In this context, we evaluated the potentials for reactive species sequestration, antioxidant activity, DNA, and protein protective activity against oxidative damage induced by oxidizing agents.
DNA mutation is a critical step in carcinogenesis and elevated levels of oxidative DNA damage (8-hydroxyguanosine) have been noted in various tumors, which strongly implicates such damage in cancer etiology [45]. Furthermore, the increased production of reactive oxygen species (ROS) in various cancers has been shown to play several roles, such as in the activation of pro-tumourigenic signaling, increase in cell survival and proliferation, and drive DNA damage and genetic instability [46,47,48].
To evaluate the potential protective effect of ELSR and ERSR in mitigating oxidative damage to macromolecules, models of oxidative damage induced by hydroxide peroxide were used and ELSR and ERSR exhibited antioxidant activity, acting in the direct capture of reactive species. Furthermore, we demonstrated that the extracts were able to promote the protection of macromolecules against oxidative damage. These results indicate that the extracts can stabilize reactive species through electron or H+ donation, preventing nonspecific oxidoreduction reactions that propagate in the intracellular environment [6]. The antioxidant activity of ELSR and ERSR may be related to the presence of biologically active metabolites identified in both extracts. These metabolites are widely recognized for stabilizing reactive oxygen species through hydrogen donation and binding with metal ions [49,50,51].
In addition to the effect of direct radical scavenging, ELSR and ERSR were able to protect proteins and DNA from oxidative damage. Oxidation of macromolecules by reactive species is an important deleterious effect observed at the cellular level. Protein oxidation promotes the loss of function of receptors, enzymes, and other intracellular proteins, the formation of protein aggregates, and proteolysis.
The protective effect of S. rugosa extracts against oxidation of macromolecules suggests that intracellular oxidative damages can be reduced by ingesting the extracts and could present beneficial effects for human health. The intake of natural antioxidants is recognized for promoting a double beneficial effect on the organism: the protection of healthy cells against the deleterious action of reactive species and anticancer properties. Besides conferring protection to macromolecules, flavonoids, naphthoquinones, among others can regulate the expression of oncogenes and tumor suppressor genes, control cell proliferation, metastasis, and angiogenesis, induce cell cycle arrest, and promote apoptosis in cancer cells [36,41,42,51,52,53].
Thus, based on the results obtained, ELSR and ERSR may modulate the cellular environment promoting apoptosis activation in K562 and Jurkat cells. From this perspective, further studies are necessary to gain a better understanding of the mechanisms through which the extracts promote the death of leukemic cells.
The search for cytotoxic agents that exhibit selectivity in cancer treatment is increasing [54,55]. In this study, we found that ELSR and ERSR do not affect the viability of healthy cells using PBMC as a model, cells that could be affected during treatment with chemotherapeutic agents during the treatment of leukemias. Since IC50 was not obtained for this cell type, it is possible to infer that ELSR and ERSR exhibit at least 2-3-fold minimal selectivity for leukemic cells. This effect is desirable because the chemotherapeutic agents currently in use are not selective in their actions, causing side effects in the body, such as high toxicity in healthy cells [56]. Thus, we suggest that S. rugosa extracts possess potential therapeutic and chemopreventive effects.
Our results demonstrated that the extracts are more effective on the Jurkat cell line, a model of T-cell acute lymphoblastic leukemia. The differences found in our assays among the cell lines utilized might be attributable to mutations in each of the lines investigated. In acute lymphoblastic leukemias, such as Jurkat, silencing of tumor suppressor and pro-apoptotic genes such as p53, PTEN, and BAX is common [15,57]. BCR-ABL1 positive leukemias, as in the K562 cell model, exhibit increased tyrosine kinase activity [13].
4. Materials and Methods
4.1. Chemicals and Reagents
The reagents 2,2-diphenyl-1-picrylhydrazyl (DPPH), 2,2'-azinobis 3-ethylbenzthiazoline-6-sulfonic acid (ABTS), 2,2′-Azobis(2-methylpropionamidine) dihydrochloride (AAPH), ascorbic acid (AA), bovine serum albumin (BSA), hydroxytoluene butylate (BHT), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), ammonium persulfate and potassium persulfate were purchased from Sigma-Aldrich (São Paulo, Brazil). Electrophoresis reagents were purchased from Bio-Rad Laboratories (São Paulo, Brazil). RPMI-1640 culture medium (Rosewell Park Memorial Institute), antibiotics penicillin, streptomycin, and neomycin, and fetal bovine serum (FBS) were purchased from Gibco/Invitrogem (Minneapolis, USA). The other reagents used in this work were of analytical grade.
4.2. Collection and Preparation of Plant Material
The leaves and roots of S. rugosa were collected in the city of Dourados, MS, Brazil (22°05′45″ S 55°20′46″ W), and the permission to access genetic heritage (CTA) was registered in the Heritage SisGen under access code AA6FADF. An exsiccate specimen of the species was deposited in the Herbarium of the Universidade Federal da Grande Dourados with the registration number 4664. Leaves and roots of S. rugosa were washed with running water, dried in an oven with air circulation at 36 °C for 10 days, and ground in a knife mill. Leaf (ELSR) and root extracts of S. rugosa (ERSR) were prepared from 200 g of leaf or root powder macerated in 95% ethanol at a 1:7 (w/v) ratio at room temperature for 21 days.
Every seven days, the extract was filtered, the solvent was reserved, and a new aliquot of 95% ethanol was added to perform an exhaustive extraction of the compounds. The filtrates were pooled, concentrated in a rotary vacuum evaporator (Gehaka, São Paulo, SP, Brazil) at 37 ºC and lyophilized. The yields of the extracts were calculated. The yield of ELSR was 16.61% while that of ERSR was 1%.
Phytochemical Analyses
The ELSR and ERSR were analyzed in a liquid chromatograph (UFLC, Shimadzu) coupled to a diode array detector (DAD, Shimadzu) and to an electrospray ionization time-of-flight mass spectrometer. (ESI-QTOF-micrOTOF QII, Bruker Daltonics); positive ionization mode, with detection between 120-1200 Da. The sample was injected into a column C-18 (Kinetex, 2,6 μm, Phenomenex). The chromatographic and mass spectrometric parameters applied were the same used by Castro et al. [19]. We identified the metabolites in the ELSR and ERSR by analyzing their molecular mass, fragmentation patterns, and ultraviolet (UV) absorption spectra, comparing these characteristics with existing literature data. Whenever possible, we confirmed the identities of these compounds through comparative analysis with authentic standards.
4.3. Antioxidant Activity Assays
4.3.1. Direct ABTS•+ Radical Scavenging Assay
The antioxidant capacity of ELSR and ERSR was assessed using ABTS•+ assay [58]. A volume of 5 mL of ABTS solution (7 mM) and 88 μL of potassium persulfate solution (140 mM) were incubated for 12-16 h at room temperature in the dark. The ABTS•+ working solution was obtained by diluting the stock solution in distilled water and its absorbance was determined (0.70 ± 0.05 units) at 734 nm in a spectrophotometer. ELSR or ERSR were prepared at different concentrations (0.1-100 μg/mL), and 20 μL were mixed with 1,980 μL of the ABTS•+ working solution. The mixture was incubated for 6 min and absorbance was measured at 734 nm. Ascorbic acid (AA) and butylated hydroxytoluene (BHT) were used as reference antioxidants. A volume of 20 µL of distilled water was used for the determination of the absorbance of the negative control The percentage of ABTS•+ radical scavenging was calculated using the following equation 1:
Three independent experiments were performed in triplicate and the concentration of the extracts needed for the capture of 50% of the ABTS•+ radical (IC50) was determined, as well as the reference antioxidants.
4.3.2. Direct DPPH• Radical Scavenging Assay
The direct capture of DPPH• radical by ELSR and ERSR was evaluated according to Gupta and Gupta [59]. For this, 200 μL of the ELSRs and ERSRs at different concentrations (0.1-100 μg/mL) were diluted in 1,800 μL of 0.11 mM DPPH• solution prepared in 80% ethanol. The mixture was incubated for 30 min at room temperature in the dark. The absorbance at 517 nm was determined. Ascorbic acid and BHT were used as reference antioxidants. A 0.11 mM DPPH• solution plus 200 µL of 80% ethanol was used as a negative control. Three independent experiments were performed in triplicate and the percentage of direct DPPH• radical scavenging was calculated with the following equation 2:
The concentration of the extracts required for the capture of 50% of the DPPH• radical (IC50) was determined, as well as for the reference antioxidants.
The antioxidant activity assays were performed by diluting the extracts in a 10 mg/mL stock solution with 80% ethanol. From the stock solution, working solutions containing between 5 and 500 µg/mL were prepared in the solvents of each assay.
4.4. Protection against Oxidative Damage to the Macromolecules by ELSRs and ERSRs
4.4.1. AAPH-Induced Oxidation of Proteins
The protective effect of the extracts against AAPH-induced protein oxidation was evaluated according to the methodology of Mayo et al. [60], with minor modifications. A 3 µL aliquot of BSA (3 mg/mL, prepared in PBS) was pre-incubated with 3 µL of the ELSRs or ERSRs at different concentrations (5-500 µg/mL) for 30 min at 37 °C. Then, 3 µL of AAPH (120 mM, prepared in water) was added to the respective treatments, followed by further incubation for 120 min at 37 °C.
Then, 11 µL of sample buffer was added to each tube. The samples were incubated for 5 min at 95 °C and applied to a 12% SDS-PAGE polyacrylamide gel, separated at 200 V on the Mini-PROTEAN Tetra Cell system (Bio-Rad Laboratories). At the end of electrophoresis, the gels were stained, destained, and scanned in a Gel Doc EZ Imager (Bio-Rad Laboratories). The volume of the BSA bands was determined with the help of Image Lab software. Increased volume of the BSA bands was considered oxidative damage. BSA incubated with only AAPH was used as a positive control. The protection conferred by the different concentrations of the extracts was calculated by averaging four gels.
4.4.2. DNA Fragmentation induced by Hydrogen Peroxide (H2O2)
The protective effect of ELSR and ERSR against DNA fragmentation was evaluated using the method described by Kumar and Chattopadhyay [61] with modifications. A 4 µL aliquot of plasmid DNA (50 ng/µL) was incubated with 4 µL of ELSR or ERSR at different concentrations, diluted in PBS (5-500 µg/mL). Then, 4 µL of the oxidizing agent H2O2 at 30% was added to the respective treatments. The plasmid in the absence of UV light (CT), the plasmid in the presence of UV light (CTUV), the plasmid in the presence of the extracts plus UV light (ELSR or ERSR), and the control plasmid incubated only with hydrogen peroxide (H2O2) were used as negative control. The effect of other reference antioxidants (rutin and catechin - 150 µg/mL) on protecting DNA fragmentation was analyzed. The samples were incubated in a UVT-312 transilluminator at 302 nm at room temperature for 5 minutes. Then, the samples were applied to a 2% agarose gel containing ethidium bromide (10 mg/mL) and subjected to electrophoresis. Images of the gels were scanned using the Gel Doc™ EZ Imager photodocumentator (Bio-Rad Laboratories) and analyzed using Image Lab™ software. The results of 3 gels were expressed as a percentage of DNA fragmentation.
The oxidative damage assays were performed by diluting the extracts in a stock solution containing 0.01% of 80% ethanol supplemented with PBS. From the stock solution, working solutions containing between 5 and 500 µg/mL were prepared in the solvents of each assay.
4.5. Cell-Based Assays
4.5.1. Cell Culture
The K562 (Human Chronic Myelogenous Leukemia) and Jurkat (Human Acute Lymphoblastic Leukemia was grown in suspension in RPMI 1640 media supplemented with 10 nM 4-(2-hydroxyethyl) piperazine-1-ethanesulfonic acid (HEPES), 24 nM sodium bicarbonate, 10% fetal bovine serum (FBS) and with antibiotic Penicillin-streptomycin (10,000 U/ml), all from Gibco/Invitrogen, Minneapolis, MN, USA. All cells were kept in an incubator with a humidified atmosphere containing 5% CO2 at 37 °C.
4.5.2. Isolation of Mononuclear Cells from Human Peripheral Blood
The Research Ethics Committee of the Centro Universitário da Grande Dourados (UNIGRAN) approved Blood collection from a healthy nonsmoking donor, process number 123/12. Human peripheral blood mononuclear cells (PBMC) were obtained by centrifugation using Ficoll-Paque-1077 Premium reagent (1.073g/cm3). PBMC were suspended in RPMI-1640 medium supplemented with 20% SFB and 1% antibiotic and maintained at 37 °C in a humidified atmosphere containing 5% CO2.
4.5.3. Isolation of Mononuclear Cells from Human Peripheral Blood
The potential cytotoxicity of ELSR and ERSR was evaluated with the MTT reagent against leukemic cells (K562 and Jurkat) and PBMC. The K562, Jurkat (2x104 cells/well), and PBMC (12x104 cells/well) cells were plated in 96-well microplates and treated with different concentrations of the ELSR and ERSR (0-500 μg/mL solubilized in RPMI-1640 medium) for 24 and 48 h. After treatment, the microplates were centrifuged at 1,200 rpm for 5 min and the culture medium was removed. A volume of 100 µL of MTT (0.5 mg/mL, prepared in RPMI-1640 medium) was added to the wells and a 4 h incubation at 37 ºC was performed. The plates were centrifuged, had the medium removed, and received 100 µL of dimethyl sulfoxide (DMSO) to solubilize the formazan crystals. A control treatment was performed with 0.1% ethanol, the volume of solvent used for preparing the extracts. The absorbances were determined at 630 nm in a microplate reader (TP READER NM, Thermo Plate) and cell viability was calculated using the following equation 3:
The cell viability assay was performed by diluting the extracts in a stock solution containing 0.1% ethanol supplemented with culture medium.
4.6. Statistical Analyzes
Data were presented as mean ± standard error of the mean (SEM). Statistically significant differences between groups were analyzed using the Student t-test for comparison between two groups and analysis of variance (ANOVA), followed by Dunnett's test, for comparison of two or more groups, using Prism 7 Software (GraphPad Software, San Diego, CA, USA). Results were considered significant when p < 0.05.
5. Conclusions
Together, we demonstrated that ELSR and ERSR present flavonoid and naphthopyranones derivatives related to the observed antioxidant activities, capable of promoting direct radical scavenging and promoting the protection of macromolecules against oxidative damage. In addition, the ELSR and ERSR exhibited cytotoxicity to human leukemia cell lines without affecting the viability of healthy cells. Our results expand the chemical compounds identified in the leaves and roots of S. rugosa and their pharmacological potential in the management conditions related to oxidative stress and leukemia.
Author Contributions
Conceptualization, C.M.d.S., J.F.C., K.d.P.S. and E.L.d.S.; Methodology and validation, C.M.d.S., J.T.d.G.C., A.S.O, P.d.S.d.R., J.F.C. and C.F.R.d.O.; Formal analysis, C.M.d.S., J.T.d.G.C., K.d.P.S.and E.L.d.S.; Investigation, C.M.d.S., D.d.S.B., D.T.H.C., P.d.S.d.R. and A.S.O.; Resources, C.A.C., D.B.d.S., K.d.P.S. and E.L.d.S.; Data curation, C.M.d.S., C.A.C., D.B.d.S., K.d.P.S. and E.L.d.S.; Writing—original draft preparation, C.M.d.S. and E.L.d.S.; Writing—review and editing, D.d.S.B., D.T.H.C., A.S.O., S.O.B., C.A.C., D.B.d.S., K.d.P.S. and E.L.d.S.; Conceptualization and supervision, K.d.P.S. and E.L.d.S; Project administration, K.d.P.S. and E.L.d.S; Funding acquisition, C.A.C., D.B.d.S., K.d.P.S. and E.L.d.S. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by institutional funding from the Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul (FUNDECT, Brazil), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES, Brazil) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq, Brazil).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The experimental data used to support the results of this study are included in the article.
Acknowledgments
This work was funded by grants from the Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul (Foundation for Support to the Development of Education, Science and Technology of the State of Mato Grosso do Sul, FUNDECT); Coordination for the Improvement of Higher Education Personnel (CAPES); and the National Council for Scientific and Technological Development (CNPq); Instituto de Ciência e Tecnologia em Áreas Úmidas (INAU).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Nogueira, V.; Hay, N. Molecular Pathways: Reactive Oxygen Species Homeostasis in Cancer Cells and Implications for Cancer Therapy. Clin Cancer Res 2013, 19, 4309. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Alomar, S. Y.; Alwasel, S. H.; Nepovimova, E.; Kuca, K.; Valko, M. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Archives of Toxicology 2024, 98, 1323–1367. [Google Scholar] [CrossRef]
- Irwin, M. E.; Rivera-Del Valle, N.; Chandra, J. Redox Control of Leukemia: From Molecular Mechanisms to Therapeutic Opportunities. Antioxid Redox Signal 2013, 18, 1349. [Google Scholar] [CrossRef]
- Xie, W.; Ma, W.; Liu, P.; Zhou, F. Overview of thioredoxin system and targeted therapies for acute leukemia. Mitochondrion 2019, 47, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Badana, A. K.; Murali Mohan, G.; Shailender, G.; Malla, R. R. Reactive Oxygen Species: A Key Constituent in Cancer Survival. Biomark Insights 2018, 13. [Google Scholar] [CrossRef] [PubMed]
- Ighodaro, O. M.; Akinloye, O. A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alexandria Journal of Medicine 2018, 54, 287–293. [Google Scholar] [CrossRef]
- Pisoschi, A. M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur J Med Chem 2015, 97, 55–74. [Google Scholar] [CrossRef]
- Thyagarajan, A.; Sahu, R. P. Potential Contributions of Antioxidants to Cancer Therapy: Immunomodulation and Radiosensitization. Integr Cancer Ther 2018, 17, 210. [Google Scholar] [CrossRef]
- León-González, A. J.; Auger, C.; Schini-Kerth, V. B. Pro-oxidant activity of polyphenols and its implication on cancer chemoprevention and chemotherapy. Biochem Pharmacol 2015, 98, 371–380. [Google Scholar] [CrossRef]
- Muchtaridi, M.; Az-Zahra, F.; Wongso, H.; Setyawati, L. U.; Novitasari, D.; Ikram, E. H. K. Molecular Mechanism of Natural Food Antioxidants to Regulate ROS in Treating Cancer: A Review. Antioxidants 2024, 13. [Google Scholar] [CrossRef]
- Bhatnagar, N.; Qureshi, A.; Hall, G. Leukaemias: a review. Paediatr Child Health 2017, 27, 489–494. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, Y.; Ma, S. Racial Differences in Four Leukemia Subtypes: Comprehensive Descriptive Epidemiology. Sci Rep 2018, 8, 548. [Google Scholar] [CrossRef] [PubMed]
- Mwirigi, A.; Dillon, R.; Raj, K. Acute leukaemia. Medicine 2017, 45, 280–286. [Google Scholar] [CrossRef]
- O’Shea, J. J.; Schwartz, D. M.; Villarino, A. V.; Gadina, M.; McInnes, I. B.; Laurence, A. The JAK-STAT Pathway: Impact on Human Disease and Therapeutic Intervention. Annu Rev Med 2015, 66, 311. [Google Scholar] [CrossRef] [PubMed]
- Gioia, L.; Siddique, A.; Head, S. R.; Salomon, D. R.; Su, A. I. A genome-wide survey of mutations in the Jurkat cell line. BMC Genomics 2018, 19, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rose-Inman, H.; Kuehl, D. Acute Leukemia. Hematol Oncol Clin North Am 2017, 31, 1011–1028. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization Estimated number of new cases in 2020, worldwide, both sexes, all ages. Available online: https://gco.iarc.fr/today/online-analysis-table?v=2020&mode=cancer&mode_population=countries&population=900&populations=900&key=asr&sex=0&cancer=39&type=0&statistic=5&prevalence=0&population_group=0&ages_group&5B&5D=0&ages_group%5B%5D=17&group_cancer=1&in (accessed on 10 June 2022).
- De Giffoni De Carvalho, J. T.; Da Silva Baldivia, D.; Leite, D. F.; De Araújo, L. C. A.; De Toledo Espindola, P. P.; Antunes, K. A.; Rocha, P. S.; De Picoli Souza, K.; Dos Santos, E. L. Medicinal Plants from Brazilian Cerrado: Antioxidant and Anticancer Potential and Protection against Chemotherapy Toxicity. Oxid Med Cell Longev 2019, 2019. [Google Scholar] [CrossRef] [PubMed]
- Castro, D. T. H.; Campos, J. F.; Damião, M. J.; Torquato, H. F. V.; Paredes-Gamero, E. J.; Carollo, C. A.; Rodrigues, E. G.; de Picoli Souza, K.; dos Santos, E. L. Ethanolic Extract of Senna velutina Roots: Chemical Composition, In Vitro and In Vivo Antitumor Effects, and B16F10-Nex2 Melanoma Cell Death Mechanisms. Oxid Med Cell Longev 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Castro, D. T. H.; Leite, D. F.; da Silva Baldivia, D.; dos Santos, H. F.; Balogun, S. O.; da Silva, D. B.; Carollo, C. A.; de Picoli Souza, K.; dos Santos, E. L. Structural Characterization and Anticancer Activity of a New Anthraquinone from Senna velutina (Fabaceae). Pharmaceuticals 2023, 16, 951. [Google Scholar] [CrossRef]
- Oladeji, O. S.; Adelowo, F. E.; Oluyori, A. P. The genus Senna (Fabaceae): A review on its traditional uses, botany, phytochemistry, pharmacology and toxicology. South African Journal of Botany 2021, 138, 1–32. [Google Scholar] [CrossRef]
- Baldivia, D. da S.; Leite, D. F.; de Castro, D. T. H.; Campos, J. F.; Dos Santos, U. P.; Paredes-Gamero, E. J.; Carollo, C. A.; Silva, D. B.; Souza, K. de P.; Dos Santos, E. L. Evaluation of In Vitro Antioxidant and Anticancer Properties of the Aqueous Extract from the Stem Bark of Stryphnodendron adstringens. Int J Mol Sci 2018, 19. [Google Scholar]
- Campos, J. F.; De Castro, D. T. H.; Damiaõ, M. J.; Vieira Torquato, H. F.; Paredes-Gamero, E. J.; Carollo, C. A.; Estevinho, L. M.; De Picoli Souza, K.; Santos, E. L. Dos The Chemical Profile of Senna velutina Leaves and Their Antioxidant and Cytotoxic Effects. Oxid Med Cell Longev 2016, 2016. [Google Scholar] [CrossRef] [PubMed]
- Campos, J. F.; Espindola, P. P. de T.; Torquato, H. F. V.; Vital, W. D.; Justo, G. Z.; Silva, D. B.; Carollo, C. A.; de Picoli Souza, K.; Paredes-Gamero, E. J.; dos Santos, E. L. Leaf and Root Extracts from Campomanesia adamantium (Myrtaceae) Promote Apoptotic Death of Leukemic Cells via Activation of Intracellular Calcium and Caspase-3. Front Pharmacol 2017, 8. [Google Scholar]
- Casagrande, J. C.; Macorini, L. F. B.; Antunes, K. A.; Dos Santos, U. P.; Campos, J. F.; Dias-Júnior, N. M.; Sangalli, A.; Cardoso, C. A. L.; Do Carmo Vieira, M.; Rabelo, L. A.; et al. Antioxidant and cytotoxic activity of hydroethanolic extract from Jacaranda decurrens leaves. PLoS One 2014, 9. [Google Scholar] [CrossRef]
- Yang, C. S.; Wang, H. Cancer Preventive Activities of Tea Catechins. Molecules 2016, 21. [Google Scholar] [CrossRef] [PubMed]
- Farias, D. F.; Cavalheiro, M. G.; Viana, M. P.; Queiroz, V. A.; Rocha-Bezerra, L. C. B.; Vasconcelos, I. M.; Morais, S. M.; Carvalho, A. F. U. Water extracts of Brazilian leguminous seeds as rich sources of larvicidal compounds against Aedes aegypti L. An Acad Bras Cienc 2010, 82, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Ricardo, L. M.; De Paula-Souza, J.; Andrade, A.; Brandão, M. G. L. Plants from the Brazilian Traditional Medicine: species from the books of the Polish physician Piotr Czerniewicz (Pedro Luiz Napoleão Chernoviz, 1812–1881). Revista Brasileira de Farmacognosia 2017, 27, 388–400. [Google Scholar] [CrossRef]
- Rodrigues, V. E. G.; Carvalho, D. A. de Levantamento etnobotânico de plantas medicinais no domínio do Cerrado na região do Alto Rio Grande - Minas Gerais. Ciência e agrotecnologia 2001, 25, 102–123. [Google Scholar]
- Cunha, L. F.; Costa, C. M.; Barroso, P. R.; Kato, K. C.; Oliveira, F. De; Victor, C.; Filho, M.; Fernanda, C.; Grael, F.; Gregório, L. E.; et al. Pharmacognosy Phytochemical screening and biological assays of ethanolic leaf extract of Senna rugosa widely used in the popular medicine although not yet adequately investigated as to its phytoconstituents and Abstract Resumo Senna rugosa ( Fabaceae ) é. Rodriguésia 2020, 7, 2–15. [Google Scholar]
- Silva, J. G. A.; Silva, A. A.; Coutinho, I. D.; Pessoa, C. O.; Cavalheiro, A. J.; Silva, M. G. V. Chemical Profile and Cytotoxic Activity of Leaf Extracts from Senna spp. from Northeast of Brazil. J Braz Chem Soc 2016, 27, 1872–1880. [Google Scholar]
- Maia, I. R. D. O.; Trevisan, M. T. S.; Silva, M. G. D. V.; Breuer, A.; Owen, R. W. Characterization and quantitation of polyphenolic compounds in senna macranthera var pudibunda from the Northeast of Brazil. Nat Prod Commun 2019, 14. [Google Scholar] [CrossRef]
- Wang, S.; Feng, K.; Han, L.; Fang, X.; Zhang, Y.; Yu, H.; Pang, X. Glycosidic compounds from Cassia obtusifolia seeds and their inhibitory effects on OATs, OCTs and OATPs. Phytochem Lett 2019, 32, 105–109. [Google Scholar] [CrossRef]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular targets of epigallocatechin—gallate (EGCG): A special focus on signal transduction and cancer. Nutrients 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Almatrood, S. A.; Almatroudi, A.; Khan, A. A.; Alhumaydh, F. A.; Alsahl, M. A.; Rahmani, A. H. Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules 2020, 25. [Google Scholar] [CrossRef] [PubMed]
- Nouri, Z.; Fakhri, S.; Nouri, K.; Wallace, C. E.; Farzaei, M. H.; Bishayee, A. Targeting Multiple Signaling Pathways in Cancer: The Rutin Therapeutic Approach. Cancers (Basel) 2020, 12, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Cunha, L. F.; Costa, C. M.; Barroso, P. R.; Kato, K. C.; Oliveira, F. De; Victor, C.; Filho, M.; Fernanda, C.; Grael, F.; Gregório, L. E.; et al. Pharmacognosy Phytochemical screening and biological assays of ethanolic leaf extract of Senna rugosa widely used in the popular medicine although not yet adequately investigated as to its phytoconstituents and Abstract Resumo Senna rugosa ( Fabaceae ) é. Rodriguésia 2020, 7, 2–15. [Google Scholar]
- Yadav, P.; Parshad, B.; Manchanda, P.; Sharma, S. K. Chromones and their derivatives as radical scavengers: a remedy for cell impairment. Curr Top Med Chem 2014, 14, 2552–2575. [Google Scholar] [CrossRef] [PubMed]
- Duan, Y. di; Jiang, Y. yan; Guo, F. xia; Chen, L. xiao; Xu, L. lu; Zhang, W.; Liu, B. The antitumor activity of naturally occurring chromones: A review. Fitoterapia 2019, 135, 114–129. [Google Scholar] [CrossRef] [PubMed]
- Amen, Y.; Elsbaey, M.; Othman, A.; Sallam, M.; Shimizu, K. Naturally occurring chromone glycosides: Sources, bioactivities, and spectroscopic features. Molecules 2021, 26. [Google Scholar] [CrossRef]
- Qiu, H.; Wang, P.; Lin, H.; Tang, C.; Zhu, H.; Yang, Y. Naphthoquinones: A continuing source for discovery of therapeutic antineoplastic agents. Chem Biol Drug Des 2018, 91, 681–690. [Google Scholar] [CrossRef]
- Wellington, K. W. Understanding cancer and the anticancer activities of naphthoquinones – a review. RSC Adv 2015, 5, 20309–20338. [Google Scholar] [CrossRef]
- Klaunig, J. E. Oxidative Stress and Cancer. Curr Pharm Des 2019, 24, 4771–4778. [Google Scholar] [CrossRef] [PubMed]
- Toyokuni, S. Novel aspects of oxidative stress-associated carcinogenesis. Antioxid Redox Signal 2006, 8, 1373–1377. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Rhodes, C. J.; Moncol, J.; Izakovic, M.; Mazur, M. Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chem Biol Interact 2006, 160, 1–40. [Google Scholar] [CrossRef] [PubMed]
- Moloney, J. N.; Cotter, T. G. ROS signalling in the biology of cancer. Semin Cell Dev Biol 2018, 80, 50–64. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Abrahamse, H. Redox potential of antioxidants in cancer progression and prevention. Antioxidants 2020, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Gupta, S. C.; Tyagi, A. K. Reactive oxygen species (ROS) and cancer: Role of antioxidative nutraceuticals. Cancer Lett 2017, 387, 95–105. [Google Scholar] [CrossRef] [PubMed]
- Kaurinovic, B.; Vastag, D. Flavonoids and Phenolic Acids as Potential Natural Antioxidants. Antioxidants 2019. [Google Scholar]
- Nijveldt, R. J.; Van Nood, E.; Van Hoorn, D. E. C.; Boelens, P. G.; Van Norren, K.; Van Leeuwen, P. A. M. Flavonoids: a review of probable mechanisms of action and potential applications. Am J Clin Nutr 2001, 74, 418–425. [Google Scholar] [CrossRef]
- Panche, A. N.; Diwan, A. D.; Chandra, S. R. Flavonoids: an overview. J Nutr Sci 2016, 5. [Google Scholar] [CrossRef]
- Abotaleb, M.; Samuel, S. M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in Cancer and Apoptosis. Cancers (Basel) 2019, 11. [Google Scholar] [CrossRef]
- Farha, A. K.; Gan, R. Y.; Li, H. Bin; Wu, D. T.; Atanasov, A. G.; Gul, K.; Zhang, J. R.; Yang, Q. Q.; Corke, H. The anticancer potential of the dietary polyphenol rutin: Current status, challenges, and perspectives. Crit Rev Food Sci Nutr 2022, 62, 832–859. [Google Scholar] [CrossRef]
- Lucas, D. M.; Still, P. C.; Pérez, L. B.; Grever, M. R.; Kinghorn, A. D. Potential of Plant-Derived Natural Products in the Treatment of Leukemia and Lymphoma. Curr Drug Targets 2010, 11, 812. [Google Scholar] [CrossRef]
- Arévalo, C. M.; Cruz-Rodriguez, N.; Quijano, S.; Fiorentino, S. Plant-derived extracts and metabolic modulation in leukemia: a promising approach to overcome treatment resistance. Front Mol Biosci 2023, 10, 1229760. [Google Scholar] [CrossRef] [PubMed]
- Senapati, S.; Mahanta, A. K.; Kumar, S.; Maiti, P. Controlled drug delivery vehicles for cancer treatment and their performance. Signal Transduct Target Ther 2018, 3. [Google Scholar] [CrossRef] [PubMed]
- Mócsai, A.; Ruland, J.; Tybulewicz, V. L. J. The SYK tyrosine kinase: a crucial player in diverse biological functions. Nat Rev Immunol 2010, 10, 387. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic Biol Med 1999, 26, 1231–1237. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.; Gupta, R. K. Bioprotective properties of Dragon’s blood resin: in vitro evaluation of antioxidant activity and antimicrobial activity. BMC Complement Altern Med 2011, 11. [Google Scholar] [CrossRef]
- Mayo, J. C.; Tan, D. X.; Sainz, R. M.; Lopez-Burillo, S.; Reiter, R. J. Oxidative damage to catalase induced by peroxyl radicals: functional protection by melatonin and other antioxidants. Free Radic Res 2003, 37, 543–553. [Google Scholar] [CrossRef]
- Kumar, A.; Chattopadhyay, S. DNA damage protecting activity and antioxidant potential of pudina extract. Food Chem 2007, 100, 1377–1384. [Google Scholar] [CrossRef]
Figure 1.
Chemical profile of ethanolic extracts of the leaves (ELSR) and roots (ERSR) of S. rugosa represented by chromatogram in column C-18. * Column contaminants.
Figure 1.
Chemical profile of ethanolic extracts of the leaves (ELSR) and roots (ERSR) of S. rugosa represented by chromatogram in column C-18. * Column contaminants.
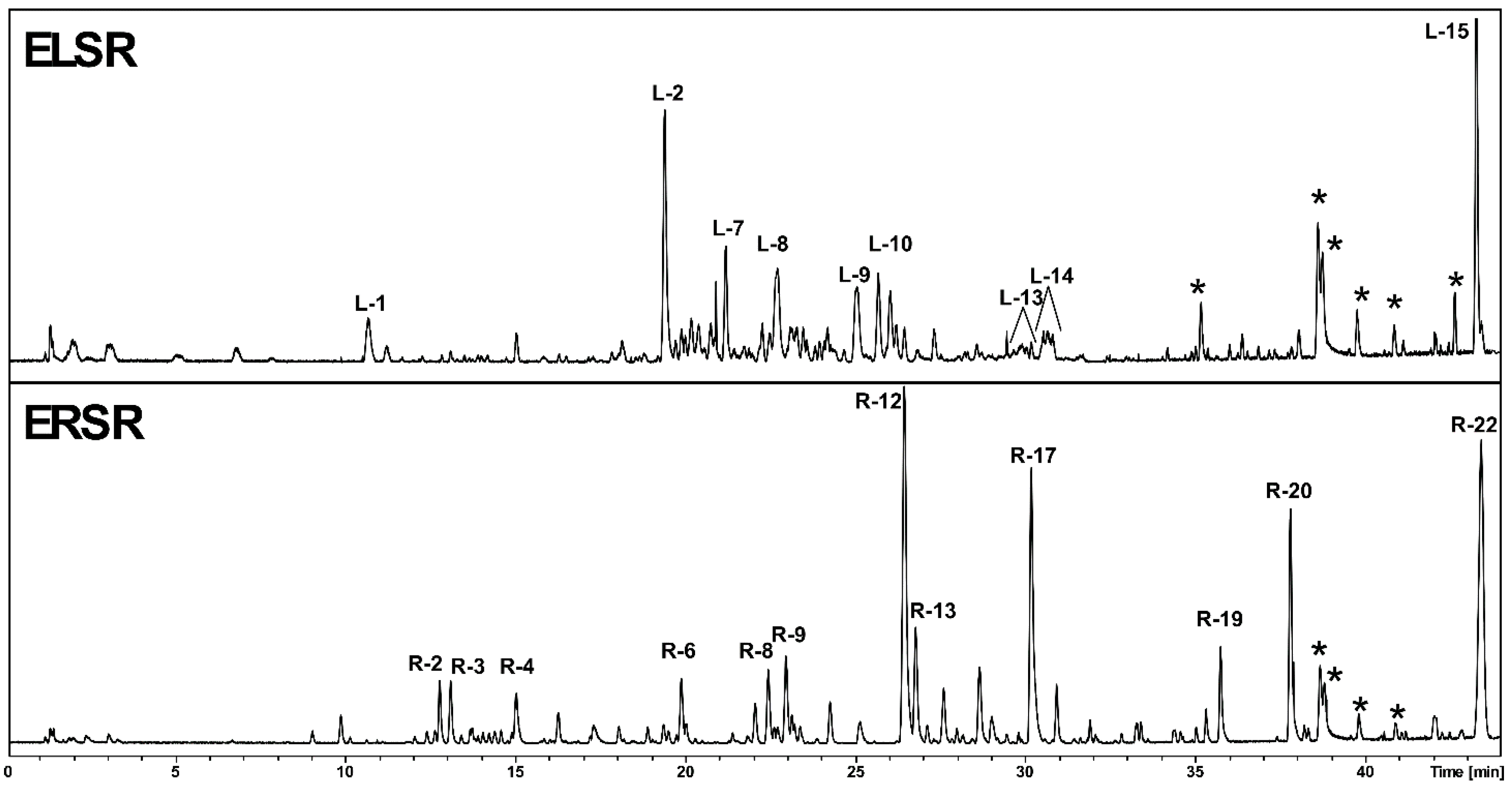
Figure 2.
Effect of the protection promoted by the extracts against AAPH-induced oxidation of BSA. BSA was incubated with different concentrations of ELSR (a) and ERSR (b). Data present results from four independent experiments, *** p < 0.0001 compared to control BSA incubated with AAPH alone (AAPH). CT: BSA control.
Figure 2.
Effect of the protection promoted by the extracts against AAPH-induced oxidation of BSA. BSA was incubated with different concentrations of ELSR (a) and ERSR (b). Data present results from four independent experiments, *** p < 0.0001 compared to control BSA incubated with AAPH alone (AAPH). CT: BSA control.
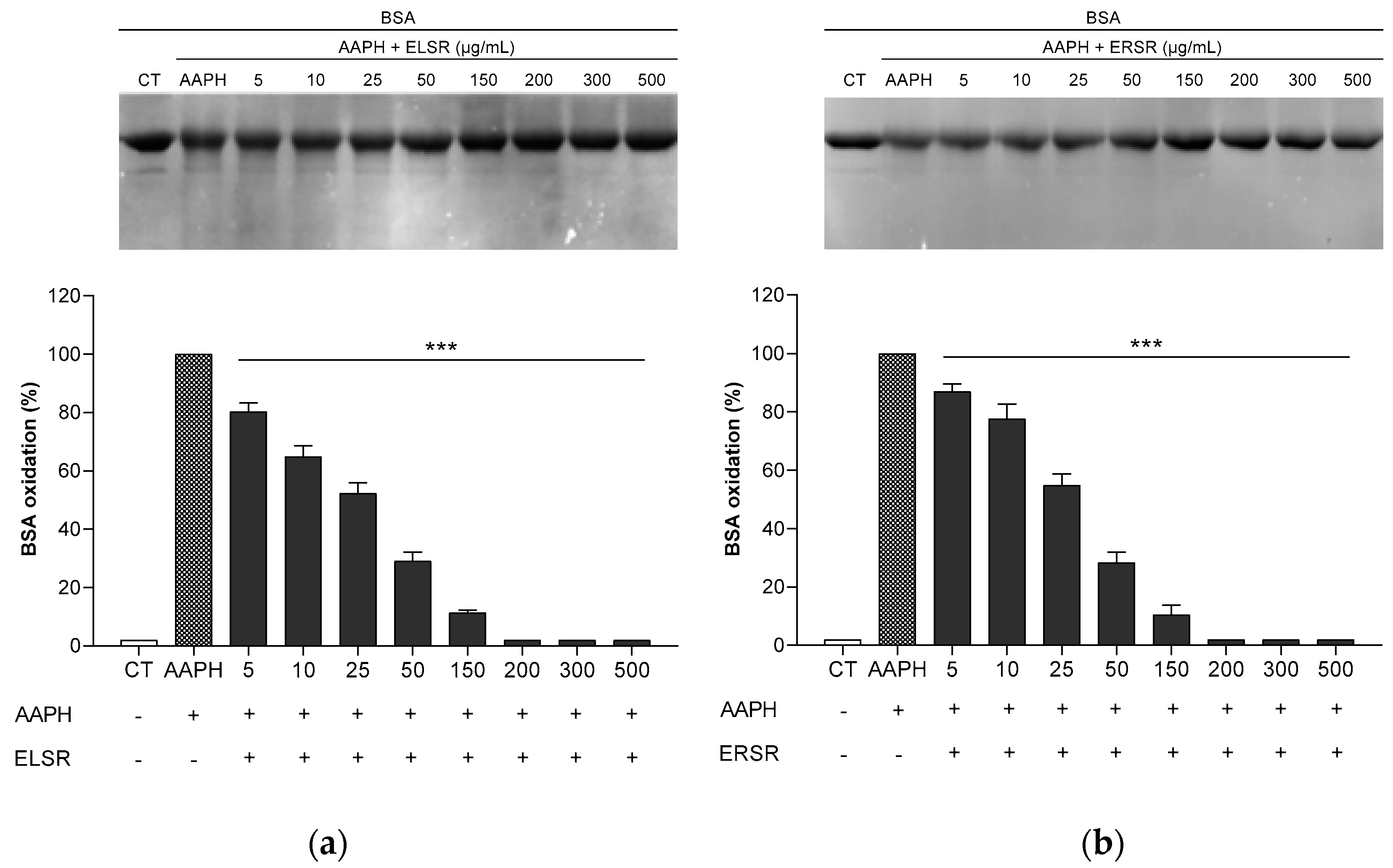
Figure 3.
Effect of ELSR and ERSR protection against DNA fragmentation induced by H2O2 and UV radiation. Representative image of plasmid DNA bands (a). Percent fragmentation of plasmid DNA incubated with different concentrations (5-500 µg/mL) of ELSR and ERSR (b). Results expressed a percentage of DNA fragmentation. PC: Plasmid control; PCUV: Plasmid + UV control; ELSR: Plasmid + ELSR 500 µg/mL; ERSR: Plasmid + ERSR 500 µg/mL; H2O2: plasmid + H2O2 30%; R: rutin; C: catechin. Data present results from three independent experiments,* p <0.05; ** p <0.01, and *** p <0.001 compared to control DNA incubated with H2O2 and subjected to UV.
Figure 3.
Effect of ELSR and ERSR protection against DNA fragmentation induced by H2O2 and UV radiation. Representative image of plasmid DNA bands (a). Percent fragmentation of plasmid DNA incubated with different concentrations (5-500 µg/mL) of ELSR and ERSR (b). Results expressed a percentage of DNA fragmentation. PC: Plasmid control; PCUV: Plasmid + UV control; ELSR: Plasmid + ELSR 500 µg/mL; ERSR: Plasmid + ERSR 500 µg/mL; H2O2: plasmid + H2O2 30%; R: rutin; C: catechin. Data present results from three independent experiments,* p <0.05; ** p <0.01, and *** p <0.001 compared to control DNA incubated with H2O2 and subjected to UV.
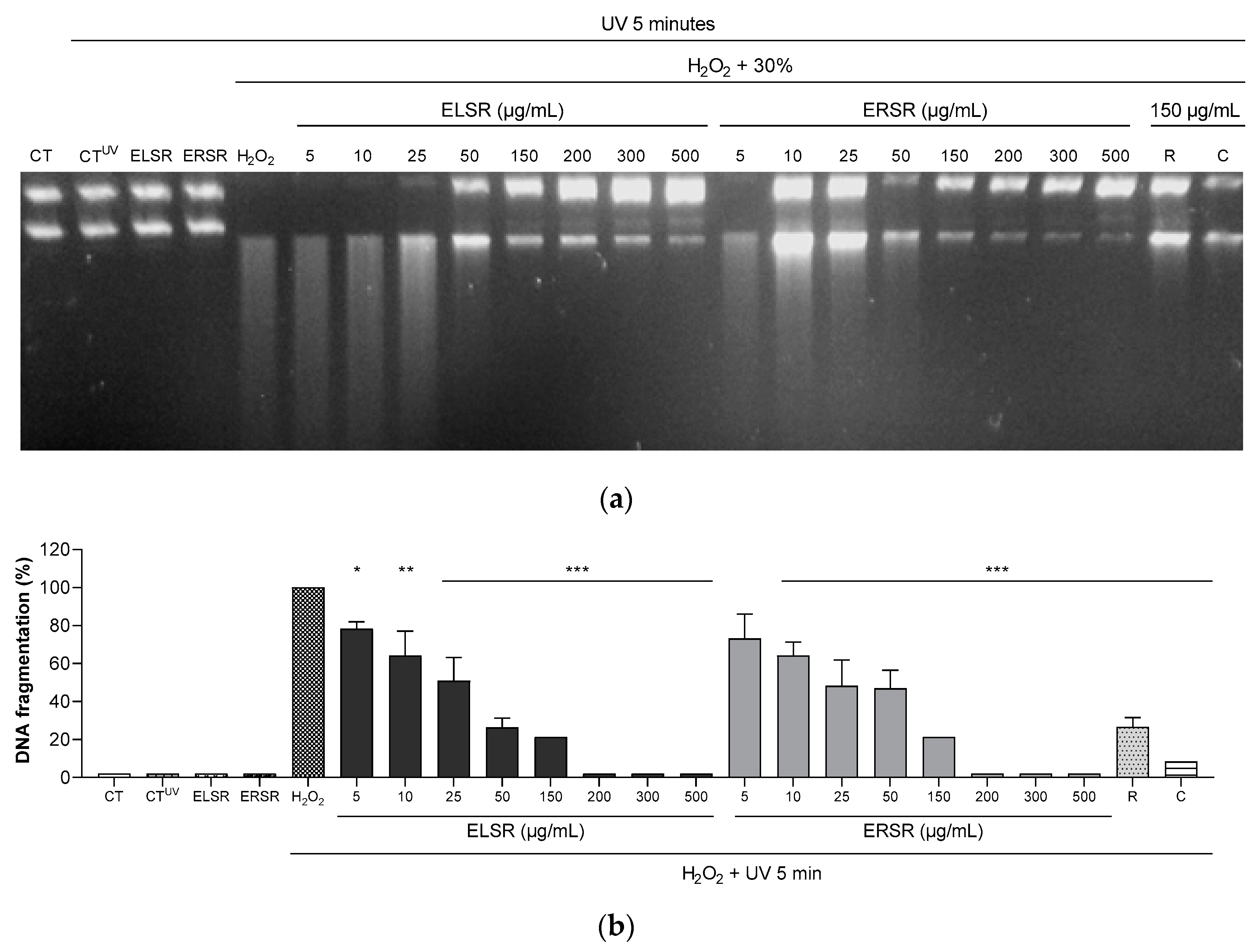
Figure 4.
Cell viability of PBMC, K562, and Jurkat cell lines after 24 (A, C) and 48 h (B, D) of treatment with different concentrations of the ELSR and ERSR (50 - 500 μg/mL). Data were expressed as the means ± SEM of three independent experiments. Results were considered significant when **** p< 0.0001 compared to control (CT) cells incubated with culture medium only.
Figure 4.
Cell viability of PBMC, K562, and Jurkat cell lines after 24 (A, C) and 48 h (B, D) of treatment with different concentrations of the ELSR and ERSR (50 - 500 μg/mL). Data were expressed as the means ± SEM of three independent experiments. Results were considered significant when **** p< 0.0001 compared to control (CT) cells incubated with culture medium only.
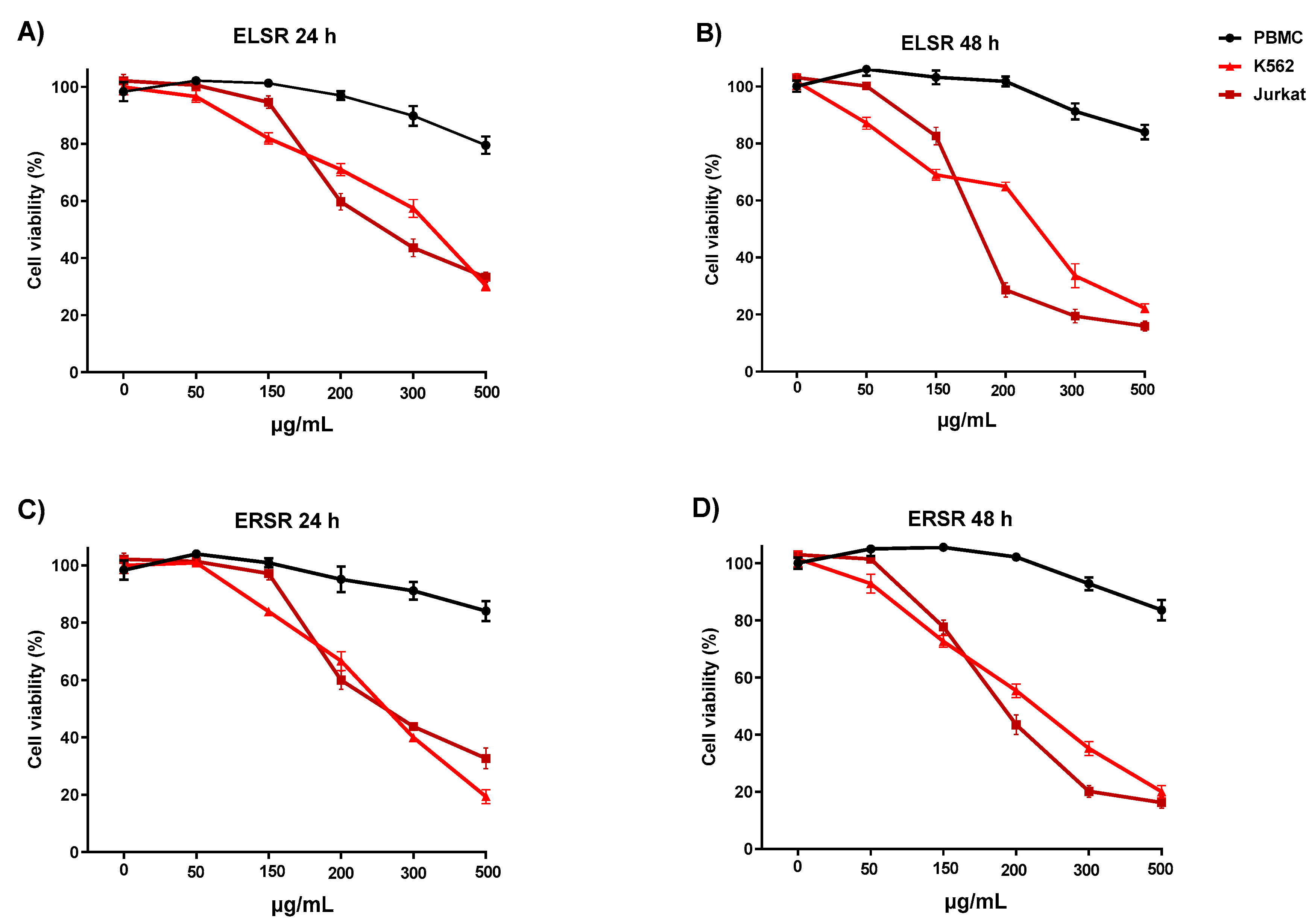

Table 1.
Chemical profile of ethanolic extracts of the leaves (ELSR) and roots (ERSR) of S. rugosa analyzed using UFLC-MS.
Table 1.
Chemical profile of ethanolic extracts of the leaves (ELSR) and roots (ERSR) of S. rugosa analyzed using UFLC-MS.
| ELSR | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Peak | Time(min) | UV | Formula | [M+H]+ | MS/MS | Classes | Compound | |||
| L-1 | 10.6 | 280 | C15H14O6 | 291.0870 | - | Flavan-3-ol | Catechin | |||
| L-2 | 19.4 | 256/353 | C27H30O16 | 611.1599 | 465 (C21H21O12), 449 (C21H21O11), 303 (C15H11O7) | Flavonol | Rutin | |||
| L-3 | 19.8 | 313 | C15H14O5 | 275.0914 | 225 (C14H9O3), 213 (C13H9O3), 197 (C13H9O2), 185 (C12H9O2), 157 (C11H9O) | Stilbene | 3,3',5,5'-Tetrahydroxy-4-methoxystilbene | |||
| L-4 | 20.1 | 280 | C30H26O10 | 547.1580 | 393 (C22H17O7), 285 (C16H13O5), 271 (C15H11O5), 259 (C14H11O5), 241 (C14H9O4), 147 (C9H7O2) | Proanthocyanidin | Epiafzelechin-epiafzelechin derivative | |||
| L-5 | 20.3 | 280 | C30H26O10 | 547.1582 | 393 (C22H17O7), 271 (C15H11O5), 241 (C14H9O4) | Proanthocyanidin | Epiafzelechin-epiafzelechin derivative | |||
| L-6 | 20.7 | 280 | C30H26O10 | 547.1589 | 409 (C22H17O8), 269 (C16H13O4), 243 (C14H11O4), 163 (C9H7O3) | Proanthocyanidin | Epiafzelechin-epiafzelechin derivative | |||
| L-7 | 21.1 | 265/347 | C27H30O15 | 595.1651 | 449 (C21H21O11), 287 (C15H11O6) | Flavonol | Kaempferol 3-O-rutinoside | |||
| L-8 | 22.6 | 280 | C30H26O9 | 531.1644 | 393 (C22H17O7), 269 (C16H13O4) | Proanthocyanidin | Trihydroxyflavan-epiafzelechin derivative | |||
| L-9 | 24.9 | 280 | C30H26O9 | 531.1648 | 393 (C22H17O7), 269 (C16H13O4) | Proanthocyanidin | Trihydroxyflavan-epiafzelechin derivative | |||
| L-10 | 25.6 | 280 | C30H26O9 | 531.1648 | 393 (C22H17O7), 269 (C16H13O4), 243 (C14H11O4), 207 (C11H11O4) | Proanthocyanidin | Trihydroxyflavan-epiafzelechin derivative | |||
| L-11 | 25.9 | 277/345 | C15H10O6 | 287.0546 | 241 (C14H9O4), 153 (C7H5O4) | Flavone | Luteolin | |||
| L-12 | 26.3 | 280 | C30H26O9 | 531.1651 | - | Proanthocyanidin | Trihydroxyflavan-epiafzelechin derivative | |||
| L-13 | 27.2 | 279/350 | C16H12O7 | 317.0662 | - | 3-metoxyluteolin | ||||
| L-14 | 29.2-30.1 | 280 | C45H38O13 | 787.2360 | - | Proanthocyanidin | Trimeric procyanidins | |||
| L-15 | 30.2-30.9 | 280 | C45H38O12 | 771.2412 | - | Proanthocyanidin | Trimeric procyanidins | |||
| L-16 | 43.2 | 406 | C34H40O9 | 593.2750 | 533 (C32H37O7), 461 (C29H33O5) | Tetrahydroanthracene | Dimeric tetrahydroanthracene derivative | |||
| ERSR | ||||||||||
| Peak | Time (min) | UV | Formula | [M+H]+ | MS/MS | Classes | Compound | |||
| R-1 | 9.8 | 253/298 | C24H32O15 | 561.1791 | 489 (C24H25O11), 423 (C20H23O10), 345 (C18H17O7), 291 (C15H15O6) | - | unknown | |||
| R-2 | 12.7 | 254/299 | C20H26O11 | 443.1542 | 323 (C16H19O7), 293 (C15H17O6), 235 (C12H11O5), 205 (C11H9O4) | Chromone | Obtusichromoneside derivative | |||
| R-3 | 13 | 252/295 | C20H26O11 | 443.1548 | 371 (C20H19O7), 259 (C14H11O5), 235 (C12H11O5), 205 (C11H9O4) | Chromone | Obtusichromoneside derivative | |||
| R-4 | 14.9 | 320 | C14H12O5 | 261.0757 | 197 (C13H9O2), 169 (C12H9O) | Naphthoquinone | 2-Methoxystypandrone | |||
| R-5 | 16.2 | 313 | C15H14O5 | 275.091 | 213 (C13H9O3), 197 (C13H9O2), 169 (C12H9O) | Stilbene | Tetrahydroxy-methoxy stilbene derivative | |||
| R-6 | 19.8 | 313 | C15H14O5 | 275.0914 | 225 (C14H9O3), 213 (C13H9O3), 197 (C13H9O2), 185 (C12H9O2), 157 (C11H9O) | Stilbene | 3,3',5,5'-Tetrahydroxy-4-methoxystilbene | |||
| R-7 | 22 | 280/334 | C16H14O6 | 303.0867 | 271 (C15H11O5), 243 (C14H11O4), 203 (C11H7O4) | Chromone | chromone deriative | |||
| R-8 | 22.3 | 278/326/400 | C25H28O14 | 553.1554 | 259 (C14H11O5) | Naphthopyrone | Putative norrubrofusarin gentiobioside | |||
| R-9 | 22.9 | 279/312/366 | C26H30O14 | 567.1699 | 273 (C15H13O5) | Flavanone | Putative hexahydroxy Flavanonol pentosyl-hexosyl | |||
| R-10 | 24.2 | 277/324/399 | C27H32O15 | 597.18 | 273 (C15H13O5) | Naphthopyrone | Rubrofusarin gentiobioside | |||
| R-11 | 25.1 | 276/310/369 | C15H12O8S | 353.0314 | 273 (C15H13O5), 230 (C13H10O4) | Flavanone | Putative hexahydroxy Flavanonol sulfate | |||
| R-12 | 26.4 | 277/324/395 | C26H30O14 | 567.1709 | 273 (C15H13O5) | Naphthopyrone | Cassiaside B | |||
| R-13 | 26.7 | 285/320/379 | C27H32O14 | 581.1864 | 449 (C22H25O10), 287 (C16H15O5) | Flavanone | Putative hexahydroxy-methoxy Flavanonol pentosyl-hexosyl | |||
| R-14 | 27.5 | 277/324/401 | C21H22O10 | 435.1286 | 273 (C15H13O5) | Naphthopyrone | Rubrofusarin-O-glucopyranoside | |||
| R-15 | 28.6 | 279/330/413 | C28H34O15 | 611.1963 | 449 (C22H25O10), 287 (C16H15O5) | - | unknown | |||
| R-16 | 28.9 | 285/321/377 | C16H14O8S | 367.0472 | 287 (C16H15O5) | Flavanone | Putative hexahydroxy-methoxy Flavanonol sulfate | |||
| R-17 | 30.1 | 258/278/331/411 | C27H32O14 | 581.1861 | 449 (C22H25O10), 419 (C21H23O9), 287 (C16H15O5) | - | unknown | |||
| R-18 | 30.8 | 279/330/411 | C22H24O10 | 449.1433 | 287 (C16H15O5) | - | unknown | |||
| R-19 | 35.7 | 277/325/402 | C15H12O5 | 273.0758 | 230 (C13H10O4) | Naphthopyrone | Rubrofusarin | |||
| R-20 | 37.7 | 281/337/425 | C16H15O5 | 287.0917 | 272 (C15H12O5), 254 (C15H10O4), 244 (C14H12O4), 226 (C15H10O3), 198 (C13H10O2) | - | unknown | |||
| R-21 | 43.4 | 410 | C36H44O9 | 621.308 | 561 (C34H41O7) | - | unknown | |||
| R-22 | 44.5 | 410 | C36H44O9 | 621.3077 | 561 (C34H41O7) | - | unknown | |||
Table 2.
IC50 values and maximum activity of the ELSR, ERSR, AA, and BHT.
| Samples | ABTS•+ | DPPH• | |||||
|---|---|---|---|---|---|---|---|
| IC50 (µg/mL) |
Maximum Activity (%) |
(µg/mL) | IC50 (µg/mL) |
Maximum Activity (%) |
(µg/mL) | ||
| AA | 1.32 + 0.04 | 99. 28 + 0.16 | 5 | 2.35 + 0.34 | 94. 88 +0.16 | 10 | |
| BHT | 6.08 + 0.77 | 98. 98 + 0.24 | 50 | 71.86 + 2.32 | 93. 16 +0.23 | 500 | |
| ELSR | 4.86 + 0.51 | 98. 15 + 0.53 | 50 | 19.98 + 1.96 | 95. 24 +0.20 | 100 | |
| ERSR | 8.33 + 0.90 | 98. 40 + 0.33 | 25 | 13.37 + 1.05 | 94. 98 +0.27 | 100 | |
Values are expressed as mean ± SEM (n=3). AA = ascorbic acid; BHT = butylated hydroxytoluene.
Table 3.
IC50 values were found for human leukemia cell lines K562 and Jurkat after treatment with ELSR and ERSR.
Table 3.
IC50 values were found for human leukemia cell lines K562 and Jurkat after treatment with ELSR and ERSR.
| Cell line | ELSR IC50 (μg/mL) |
ERSR IC50 (μg/mL) |
||
|---|---|---|---|---|
| 24 h | 48 h | 24 h | 48 h | |
| PBMC | ND | ND | ND | ND |
| K562 | 345.01 ± 2.53 | 242.54 ± 2,38 | 257.49 ± 2.41 | 223.00 ± 2.34 |
| Jurkat | 255.33 ± 2.40 | 171.45 ± 2,25 | 256.65 ± 2.40 | 189.30 ± 2.27 |
Values are expressed as mean ± SEM (n=3). ND = IC50 could not be determined.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



