
Submitted:
17 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
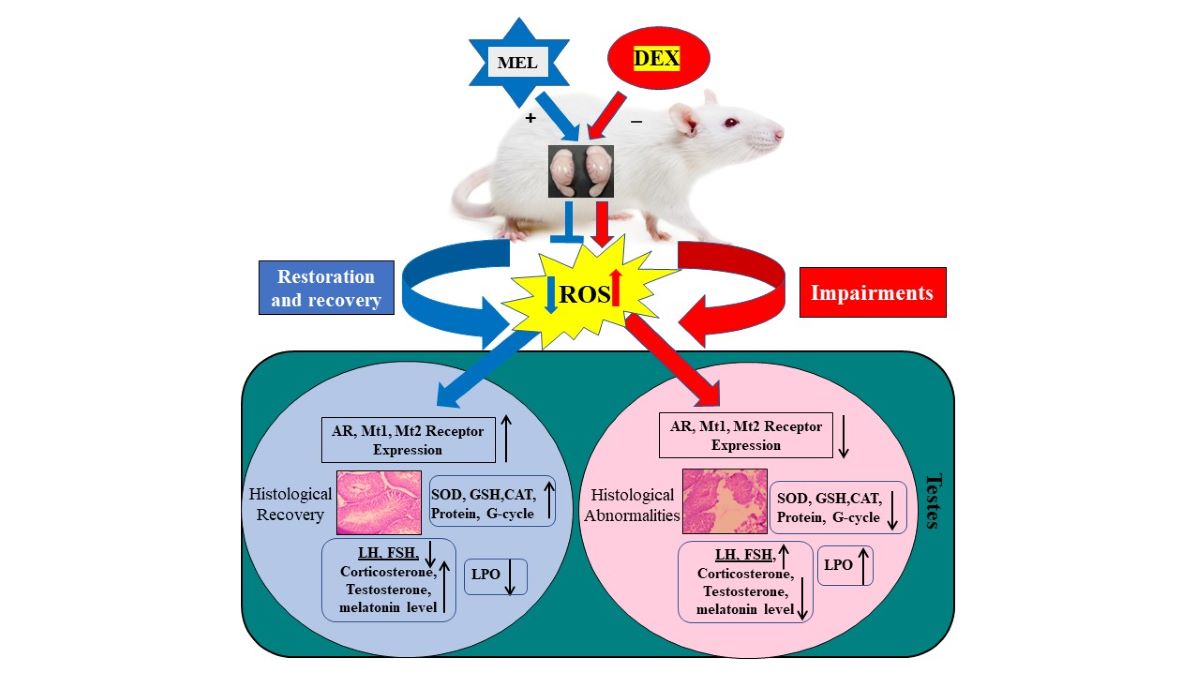
The present study aims to elucidate the effect of melatonin (MEL) on dexamethasone (DEX)-induced reproductive impairments in rats. Rats were procured and acclimatized with adequate food and water ad libitum. Twenty-four rats were divided into four groups (six in each) of Control, DEX (70µg/100g bw), DEX plus MEL, and MEL (200µg/100g bw) for 21 days. After completion, rats were sacrificed, and testes were dissected, weighed, and processed for biochemical, and receptor expression. Serum was collected for hormonal assay. DEX-treated rats showed a significant increase in lipid peroxidation (TBARS), a decrease in weight of the body and testes, and the level of antioxidative enzymes (superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPX), glutathione oxidase (GSH), glutathione reductase (GR), glucose 6-phosphate dehydrogenase (G6PDH), glutathione S-transferase (GST)), total cellular protein content, serum testosterone, corticosterone, LH, FSH, melatonin levels, AR, MT1, and MT2 receptor expression. Marked cellular alterations were noted in the testis histology of the DEX group. Melatonin treated in DEX rats showed recovery and restoration in gravimetric, serum hormonal, biochemical parameters, receptor expression, and cellularity of testes. Therefore, findings suggest that melatonin improved defence against DEX-induced testicular damages suggesting its therapeutic potential for the management of male reproductive health.
Keywords:
melatonin receptor
; androgen receptor
; oxidative stress
; male infertility
; antioxidant enzyme
1. Introduction
The hypothalamus-pituitary-pineal-gonadal (HPPG) axis is recognized for its important role in regulating reproductive functions. This interconnected communication called reciprocal cross-talk helps to keep both the reproductive and endocrine systems within precise boundaries and could contribute to coordinating the one mega-event reproduction. Melatonin (N-acetyl 5-methoxy tryptamine), the pineal hormone known as the ‘chemical expression of darkness’ is an indole amine synthesized within the pinealocytes and derived from the amino acid tryptophan plays a significant role in male reproduction. Melatonin is classified as a mitochondria-targeted antioxidant, functioning as a “firewall” against free radicals [1]. It has a crucial role in governing physiological rhythms and as well as being connected to the management of various disorders, including depression, insomnia, and Alzheimer’s disease [2]. It performs a clock and calendar function within the body [2]. Melatonin exerts its cellular effect by interacting with free radicals via scavenging [3], and serving as an important co-factor for quinone reductase-2 (QR-2) [4]. It functions as a regulator of the reproductive system by influencing various processes such as spermatogenesis, testosterone production, and sexual behavior. Free radicals are perilous substances generated by various natural bodily processes such as breathing, digestion, metabolizing drugs and alcohol, and converting lipids into energy. Typically, our bodies have a built-in defense system of antioxidants to neutralize these free radicals. If this system cannot cope properly, free radicals can initiate a damaging chain of events within the body when this defense system malfunctions. According to findings it was stated that, this sequence may cause damage to DNA, disruption of important cellular processes, breakdown of cell membranes, inhibition of vital enzymes, and interference with the creation of energy [5].
Reproduction is critically important in male rats, and the testes play key roles in this process. The testes are the integral components of the male rat's reproductive system, ensuring the continuation of the species through successful mating and fertilization. Of all infertility cases, “male factor” infertility accounts for approximately half of couple infertility (40–50% of instances), and inadequate sperm parameters can be seen in up to 2% of all males [6]. Although the etiology of infertility in up to 60% of cases remains unclear and is referred to as idiopathic [7]. Numerous causative elements have been found in infertile men. Lifestyle and human exposure to free radicals, environmental pollutants, and drug treatment could adversely affect the male reproductive system [8,9]. Glucocorticoids (GCs) are the major steroid hormones secreted by the adrenal gland. Prolonged treatment with GCs was found to cause side effects that can cause endocrine abnormalities via interfering with the anterior hypothalamic–pituitary–gonadal (HPG) axis [10,11]. Dexamethasone (DEX) is a synthetic glucocorticoid widely used in therapy due to its immunosuppressive and anti-inflammatory effects [12]. These hormones have potent anti-inflammatory and immune-system inhibitory effects and because of this characteristic, they are the most prescribed medication globally [13] and are increasingly prescribed [14]. Research has demonstrated that dexamethasone can impact several bodily systems, including the reproductive system [15]. The Leydig cells in the testes produce steroids, which supply the plasma level of testosterone. Receptors for glucocorticoids are made by Leydig cells thus, testicular tissue is the primary site of glucocorticoid action [16]. The drug passes through the cell membrane to bind to its receptors in the cytoplasm, and the drug-receptor complex subsequently reaches the cell nucleus. Functional AR was needed for Sertoli cells to maintain normal spermatogenesis and testosterone production and ensure normal male fertility, by supplying Sertoli cell-specific AR to mice lacking the AR [17]. Therefore, the expression of AR was regarded as an important symbol of androgen in cell regulation. A few research have till investigated for the effect of DEX on AR expression in the testes in our knowledge. The study aimed to investigate the neurohormone melatonin effect on reproductive impairments induced by dexamethasone in rats. For the determination of the effect of melatonin on oxidative damage and antioxidant enzymes, biochemical parameters were performed in the testes. Serum testosterone, corticosterone, LH, FSH, and melatonin were analyzed by ELISA Kit to determine the effect of melatonin on DEX-induced oxidative damage. AR, MT1, and MT2 expression levels were analyzed in the reproductive tissue testes using q RT-PCR. Melatonin in mammals mediated it actions through the membrane receptors Mt1 and Mt2. Despite the fact that melatonin has also been shown to influence testicular activity in all vertebrates, the idea that melatonin acts locally in male gonads has very little evidence. Glucocorticoids are well known for their immunosuppressive effects, but some studies indicate that they have differential effects on reproductive functions. However, how glucocorticoid influences the expression of melatonin and androgen receptors in DEX-induced animals is not clear. Hence, in our present studies, we evaluated the effect of melatonin on synthetic glucocorticoids and androgen receptor expression in testes. From our research, some evidence has been implicated suggesting that melatonin and its receptors (Mt1 & Mt2) expression, laterally interaction with androgen receptor (AR) expression in the testes, and perform a significant role in mediating reproductive functions.
2. Material and Methods
All procedures were carried out following the standard national and international guidelines and regulations and were permitted by the Institutional Animal Ethics Committee (IAEC) of SLT Institute of Pharmaceutical Sciences, Guru Ghasidas Vishwavidyalaya, Bilaspur (Registration Number: 994/ GO/Re/S/06/CPCSEA) C.G., India.
2.1. Animal Procurement and Maintenance
Male Wistar albino rats have been imported from the Defence Research and Development Establishment (DRDE), Gwalior, India. Research was performed on rats aged 3-4 months and weighing 200 ± 10 g. Rats were kept in plastic cages under ideal conditions of humidity (55–60%) and temperature (25±2°C) and were exposed to the natural day-night cycle (~12 h/12 h) and kept under standard laboratory conditions. After an acclimatization period of fourteen days, animals were divided into four groups: Control (CON), Dexamethasone (DEX), Dexamethasone along with Melatonin (MEL), and Melatonin (MEL) alone.
2.2. Experimental Design

Table 1.
caption.
| Sl. No. | Groups | Treatment | No. of animals | Duration |
|---|---|---|---|---|
| 1. | Control (CON) | Normal saline | 06 | 21 days |
| 2. | Dexamethasone (DEX) | 70 µg/100 g body weights dexamethasone | 06 | |
| 3. | Dexamethasone (DEX) + Melatonin (MEL) | 70 µg/100 g body weights DEX and 200µg/100g body weight MEL | 06 | |
| 4. | Melatonin (MEL) | 200µg/100g body weight MEL | 06 |
2.3. Drugs and Treatment
DEX and MEL were first dissolved in a few drops of absolute ethanol individually and then the working concentration was prepared as desired from the stock by diluting 0.9% saline and injected i.p. (reference). The volume of DEX and MEL injected daily was 0.5 mL at 10 to 11 AM and 5 to 6 PM respectively for twenty-one days. The control group animals were treated with 0.9% normal saline.
2.4. Tissue Collection and Processing
After completion of the experiment, deep ether anesthesia was used to euthanize the rats from each experimental group. From the heart the blood was collected through the syringe and serum was collected by centrifugation and stored at – 20 ℃ for ELISA (Enzyme-Linked Immunosorbent Assay). The bilateral testes were carefully excised under sterile conditions, and one was put in RNA Later by immediately removing the adhering fat from each group and was stored at 4℃ overnight and then transferred to -20℃. After being dried using blotting paper, the testes were weighed. Chilled normal saline was used to clean the organs and photographed for morphological analysis, and the testes were preserved in Bouin's solution for histological studies. The organ testes were subjected to biochemical assay, including lipid peroxidation (LPO), total protein content, catalase (CAT), superoxide dismutase (SOD), reduced glutathione (GSH), glutathione peroxidase (GPX), glutathione reductase (GR), Glucose-6-phosphate dehydrogenase (G6PDH), and glutathione-S-transferase (GST).
2.5. Parameters
2.5.1. q RT-PCR
Total RNA Extraction and reverse transcription
Sample Lyses and Phase Separation
To examine the expression of Mt1, Mt2, and AR genes in the testes were harvested and kept with RNA Later at 40C overnight. Total RNA isolation was achieved by homogenization, and after phase separation, the upper layer containing RNA was collected. Precipitation and elution were performed to obtain pure RNA (SPINeasyTMRNA Kit). The frozen tissue samples were homogenized using liquid nitrogen and transferred to a clean, nuclease-free 1.5 ml microcentrifuge tube. 1 ml Lysis Buffer R was added and mixed by vortexing then centrifuged at 14,000 × g for 5 min. Transfer lysate supernatant to a clean, nuclease-free 2 mL microcentrifuge tube. An equal volume of absolute ethanol was added (e.g., 750 μL ethanol to 750 μL Lysate supernatant) and mixed well by pipetting up and down. the mixture (750 μL) was transferred to column R with a collection tube. Centrifuged at 14,000 × g for 1 min. The above two steps were repeated to load the remaining mixture. A Wash Buffer of 500μl was added to the column. Centrifuged at 14,000 × g for 1 min. For DNase I digestion 1.5 ml of clean microcentrifuge tube was taken, and 5 μL of DNase I solution was added to 75 μL DNase I Buffer per preparation and mixed well then 80 μL was added to the center of the column membrane. At room temperature, it was incubated for 15 minutes. A Wash Buffer of 500μl was added to the column. Centrifuged at 14,000 × g for 1 min. Again, centrifuged at 14,000 × g for an additional 1 min to dry the column. The collection tube was removed and placed the column onto a clean 1.5 ml microcentrifuge tube. 100μl of Nuclease-free water was added to the center of the membrane. At room temperature, it was incubated for 1 min. Centrifuge at 8,000 × g for 1-2 min to elute RNA. Eluted RNA was collected in the microcentrifuge tube and pure RNA was obtained. The RNA concentration was assessed with a nanodrop at the A280/A260 wavelength. The purity of the RNA was confirmed by observing a ratio falling within the range of 1 to 2.
For the synthesis of full-length first-strand cDNA, cDNA synthesis was done by using a NeoScript 1st strand cDNA synthesis Kit. So, the first step was RNA denaturation in which we made a total of 8 μL by adding Oligo-(dT), VN (50uM)- 1 μL, total RNA (10pg-5ug)- 3 μL and RNase free water- 4 μL. This was incubated at 65°C for 5min and then chilled on ice immediately for 2min. In step two RNase-free PCR tube was taken and the following components were mixed in a PCR tube by gentle pipetting. Mixture from step one- 8 μL, 2X RT mix 10 μL, and Neo Script 1st strand Enzyme mix 2 μL. In step 3 place in L18 Gradient Thermocycler PCR (TaKaRa PCR; Thermal Cycler DiceTM Touch) for the 1st strand cDNA synthesis and run for 25°C, 5min; 55°C 45min; 85°C, 5min. qPCR was done in the L18 Gradient Thermocycler (TaKaRa PCR; Thermal Cycler DiceTM Touch) system using the master mix. Each reaction included 20 ng cDNA, 10pocomole of forward and reverse primers, and 2X Fast SYBR Green PCR Master Mix (TB Green Premix Ex Taq II, # RR820A). 10μl reaction volumes were used according as follows: Fast SYBR master mix (5 μL), nuclease-free water (3 μL), RT product (20 ng/μL-1 μL), F+R primer (10 pM-1) making a total volume of (10 μL).
A Tata MD CHECK Express real-time PCR machine was used for the running of the programme i.e. (95°C → 30sec), and 40 cycles (95°C → 5sec, 58°C → 30sec) followed by Melting curve 600C to 990C. For data analysis, MyGo pro-PCR software 3.6 was used for experimental setup and data analysis. Based on the threshold cycle of all the concerned genes including the reference gene the fold change in gene expression was calculated. The 2^-ΔΔCT method was applied to measure the fold change in all genes.
For the expression of MT1, MT2, and AR, q RT-PCR was done
Table 2.
Sequence of AR, MT1, MT2, and GAPDH Primers used in q RT-PCR.
| Sl. No. | Primer | Forward & Reverse | 5ꞌ – 3ꞌ sequence |
|---|---|---|---|
| 1. | AR | Forward | AGGAACTTGATCGCATCATTGC |
| Reverse | CTGCCATCATTTCAGGAA | ||
| 2. | MT1 | Forward | CAGTACGACCCCCGGATCTA |
| Reverse | GGCAATCGTGTACGCCG | ||
| 3. | MT2 | Forward | ATGTTCGCAGTGTTTGTGGTTT |
| Reverse | ACTGCAAGGCCAATACAGTTGA | ||
| 4. | GAPDH | Forward | CCTCAAGATTGTCAGCAATG |
| Reverse | CAGTCTTCTGAGTGGCAGTG |
AR, Androgen receptor, MT, Melatonin receptor, GAPDH, Glyceraldehyde 3-phosphate dehydrogenase.
2.5.2. Hormonal Assay
The blood sample was drawn from the heart of anesthetized subjects using a 2mL syringe, followed by centrifugation for 15 min. at 3000 rpm. The resulting serum was carefully pipetted out from the tube. Levels of serum testosterone, corticosterone, LH, FSH, and melatonin in all experimental groups were calculated using the instructions provided in the respective commercial kits: CSB – 55-TESMS-E01, 55-CORMS-E01, 11-LUTHU-E01, 11-FSHHU-E01, 12-MNTL-96-U respectively.
2.5.3. Histological Preparation
The tissue samples testis was rinsed with saline, and fixed in Bouin's solution. To get rid of extra picric acid, they were put in a small container with running water for 24 hours after being left for 2 to 8 hours. Subsequently, the tissue was stored in 70% alcohol until needed. The testicle was halved vertically, embedded in paraffin wax, and cut into sections of 5 µm thickness. The wax was removed from these paraffin pieces using xylene before being rehydrated in ethanol. After that, they were mounted in DPX resin after being stained with hematoxylin and eosin (H&E). The sections were studied and photographed under a fluorescence microscope (Axioscope-5, Carl-Zeiss, Germany).
2.5.4. Single-Cell Gel Electrophoresis (Comet Assay)
The comet assay was done following the method of [18] with some modifications [19]. The testes of both control and experimental group rats were separated and immediately stored at −80°C for comet assays. After that, these samples were homogenized in a pH 7.5 cold buffer that contained 24 mM disodium ethylenediaminetetraacetic acid (Na2EDTA) at pH 13-, and 75-mM sodium chloride (NaCl) to make a 10% tissue solution. The testis samples were treated and stored on ice using a homogenizer. Six microliters of the homogenate were put on 0.5% low melting agarose, which was then layered on frosted slides between layers of 0.6% normal melting agarose and another layer of 0.5% low melting agarose. Throughout the gel layer polymerization process, the slides were maintained on ice. A lysis solution was submerged in the slides comprising 1% sodium sarcosine, 2.5 M NaCl, 100 mM Na2EDTA, 10 mM Tris-HCl, 1% Triton X-100, and 10% DMSO at 4°C after the 0.6% agarose layer hardened. To aid in DNA unwinding, the slides were placed in an electrophoresis buffer (0.3 M NaOH, 1 mM Na2EDTA, pH 13) for ten minutes after an hour had passed. After that, 10 minutes of electrophoresis and a supply of 24 volts were carried out. After that, the slides were stained with 20 mg/mL ethidium bromide after being neutralized with Tris-HCl solution at pH 7.5. For analysis, every slide was examined using a fluorescence microscope (Axioscope-5, Carl-Zeiss, Germany).
2.5.5. Biochemical Parameters
Lipid Peroxidation Assay
The lipid peroxidation assay was used to determine MDA levels [20]. The primary technique makes use of spectrophotometric analysis of the color produced by the interaction of thiobarbituric acid (TBA) with malondialdehyde (MDA), an outcome of lipid breakdown. To do this, 100 mg of tissues were homogenized in 1 mL of a pH 7.4 20 mM Tris-HCl buffer. Three milliliters of 1% phosphoric acid and one milliliter of 0.6% TBA were added to an equivalent volume of homogenate and BHT (250 µL). After 45 minutes of heating, the sample combination underwent 10 minutes of centrifugation at 1500 rpm. It was extruding the supernatant that allowed for the measurement of optical density at 535.
Analysis of Protein
The protein content of the testes tissue homogenates was determined using the [21] technique. Solution A: Mixture of Na2CO3 and Cu2SO4 (dissolved in distilled water respectively). Solution B: Sodium Potassium Tartrates (dissolved in distilled water). Solution C: Mixture of Solution A and Solution B (Freshly prepared).
Superoxide Dismutase (SOD) Assay
The antioxidant enzyme superoxide dismutase (SOD) is in charge of balancing the superoxide anion that the body produces. Following the methodology of Ohkhawa [22], the overall SOD activity was determined. The principle of the method is based on the Xanthine oxidase system’s inhibition of Nitroblue Tetrazolium (NBT) reduction as a source of superoxide. The tissue was homogenated with potassium buffer which has a pH of 7.4. The sample spun at 3500 rpm for 15 minutes. The supernatant was removed, and 1.2 mL of sodium pyrophosphate, 0.1 mL PMS, and 0.3 mL NBT were added to the sample solution. 0.2 mL of NADH and 1 mL of glacial acetic acid were also mixed in. At 560 nm, the absorption was then determined.
Estimation of Catalase (CAT) Activity
Beers and Sizer's method was used to quantify CAT activity [23]. The principle is based on determining the rate constant for hydrogen peroxide breakdown. By measuring the absorbance change per minute, the enzyme's rate constant has been found. The tissue was homogenated using phosphate buffer and followed by a 15-minute centrifugation at 3000 rpm. The supernatant was extracted. The mixture was mixed in the dark with 3 mL H2O2 and phosphate buffer. A 240 nm measurement was made for the absorbance.
Reduced Glutathione (GSH) Assay
Glutathione is a vital antioxidant that donates one of its electrons to reduce hydroperoxides. Glutathione activity was assessed using the Sedlak and Lindsay [24]. To start the enzymatic reaction of reduced glutathione, H2O2 was introduced. A mixture of 0.1 gm of tissue and 1 mL of sucrose was mixed, and then 1.5 mL of a 0.2 molar Tris-Buffer was incorporated into the sample. To this mixture, 0.1 mL of DTNB was added, and the volume was adjusted to 10 mL by the addition of methanol. The solution was left to incubate for 30 minutes and was subsequently centrifuged for 15 minutes at 3000 rpm. After centrifugation, 200 µL of the supernatant was extracted and at 412 nm, the absorbance was determined.
Glutathione Reductase (GR) Assay
Glutathione reductase activity was measured following Goldberg and Spooner's [25] protocol. The glutathione reductase enzyme catalyzes the following reaction: although the GR process is reversible, the strongly preferred result is the production of GSH. The measurement of catalytic activity involves following the oxidation of NADPH, which causes a drop in absorbance at 340 nm. A reaction mixture (1.5 mL of phosphate buffer, 0.3 mM), including 0.5 mL of EDTA, 0.2 mL of GSSG, and 0.1 mL of NADPH was added one after the other in a stepwise manner to 0.2 mL of tissue homogenate in a test tube. At 340 nm, OD was taken. Results were given in units/min/mg of protein.
Glutathione Peroxidase (GPX) Assay
The GPX was performed according to Paglia and Valentine [26]. H2O2 was exposed to a predefined quantity of enzyme preparation in the presence of GSH for a specified period. 0.4 mL of 0.4 M phosphate buffer and 0.2 mL of tissue homogenate were combined in a test tube, and tissue homogenate (0.2 mL) was mixed with 0.1 mL 2.5 mM H2O2, 0.2 mL GSH, and 0.4 mL 0.4 M phosphate buffer. After that, the homogenate received this mixture. Ten minutes were spent heating the reaction mixture to 37°C. Following the addition of 0.5 mL of 10% TCA to halt the reaction, the tubes were centrifuged at 2000 rpm. After centrifugation, the pellet was taken out of the mixture and the supernatant was transferred to another test tube. Following that, 1 mL of 0.04% DTNB and 0.3 mL of 0.3 mM disodium hydrogen phosphate were combined with this supernatant. At 420 nm, the absorption was taken. The glutathione oxidized nanomoles per milligram of protein were used to express the GPX activity (µmol/unit/mg protein). Protein results were given as units/min/mg.
Glutathione-6- Peroxidase (G6PDH) Assay
From the testes, the glucose 6-phosphate dehydrogenase was assessed using a modified Ellis and Kirkman [27] technique. The cells were homogenized and broken down to isolate this enzyme. This enzyme turns Glucose-6-phosphate into 6-phosphogluconate Along the way, the coenzyme NADP is converted into NADPH, which increases the rate of absorption at 340 nm. The reaction mixture contained 1.0 ml of tris buffer, 0.1 ml of magnesium chloride, 0.1 mL of NADP+, 0.5 ml of phenazine methosulfate, and 0.4 ml of a dye solution. The tissue was homogenized, and 3.3 mL of the supernatant was obtained and added. After adding 0.5 mL of glucose-6-phosphate, the reaction mixture was left to oxidize for 10 minutes at room temperature. After that, absorbance values at 640 nm were determined using a UV spectrometer at minute intervals throughout 3.5 minutes.
Glutathione- S-Transferase (GST) Assay
The procedure described by Habig [28] was used to measure the activity of glutathione-S-transferase (GST). The catalytic reaction of GST, GSH, and the GST substrate CDNB (1-chloro-2,4-dinitrobenzene) interact throughout this process. For CDNB and glutathione to interact, active GST must exist under certain conditions. Through the observation of variations in absorbance at 340 nm, the glutathione-S-transferase activity was identified. Phosphate buffer, 0.1 mL of GSH, 0.1 mL of CDNB, and 0.1 mL of each make up the entire reaction mixture. 100 µl of the sample was added to begin the reaction. Using distilled water as the blank for at least three minutes, the rate of absorbance at 340 nm was measured using a spectrophotometer (Perkin Elmer) every fifteen seconds. The tissue sample was utilized to look at nonspecific substrate binding whereas the test combination, which contained all the reagents, served as a reference. Unit/Min/Mg Protein was used to express the results.
2.6. Analysis of Data
The resulting data were presented as Mean ± SD. The SPSS computer program was used for statistical analysis. A one-way ANOVA test was applied to the data to detect significant differences. Secondly, Tukey’s post-hoc test was used to compare the groups. Differences were considered significant at P<0.05, P <0.01 and P<0.001.
3. Results
3.1. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Body and Organ Weight
The gravimetric analysis showed that dexamethasone-induced a significant decrease in body weight and organ (testes) weight as compared to control rats. Treatment of the dexamethasone with exogenous melatonin ameliorates body weight and testes weight concerning the testes weight of the dexamethasone (DEX) group rats (Figure 1; Figure 2)
3.2. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Morphological Changes
The melatonin-treated group showed no change in morphology as compared with the control group, whereas DEX treated group showed shrinkage as compared to the control group. Melatonin with DEX showed recovery in morphology when compared to the DEX-treated group. (Figure 3)
3.3. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Receptor Expression
3.3.1. Effect of Dexamethasone (DEX) and Melatonin (MEL) on AR Receptor Expression
In the current study, AR protein was expressed in the testes of each rat in the experimental group, and administering DEX decreased the expression of AR. However, AR expression in DEX with MEL there was a was significant increased. These findings suggest that DEX when combined with MEL. Has a notable impact on AR expression levels. (Figure 4)
3.3.2. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Mt1 Receptor Expression
In the testes of the DEX-treated group, the activity of Mt1 decreased. Melatonin given to the DEX-treated group rat significantly recovered the gene activity in both tissues. melatonin didn’t affect Mt1 gene activity in the control group. The MEL group showed the same gene expression as that of the CON one in the testes. (Figure 5)
3.3.3. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Mt2 Receptor Expression
The DEX-treated group’s testes and pancreas showed a reduction in Mt2 expression. Rats in the DEX-treated group showed a considerable recovery in gene expression in both organ after receiving melatonin. The control group’s Mt2 gene expression was unaffected by melatonin. The MEL group had the same gene expression in the testes and pancreas compared to the CON group. (Figure 6)
3.4. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Hormonal Assay
3.4.1. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Circulatory Level of LH
The serum concentration of luteinizing hormone (LH) exhibited elevation in the group subjected to dexamethasone (DEX) treatment, in contrast to the control group. This hormonal alteration was mitigated upon supplementation with exogenous melatonin in the DEX-treated group. Notably, the administration of melatonin alone in the experimental group did not yield a discernible difference in serum LH levels when compared to those of the control group (Figure 7).
3.4.2. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Circulatory Level of FSH
The serum concentration in follicular stimulating hormone (FSH) exhibits evolution in the group subjected to dexamethasone (DEX) treatment, in contrast to the control group. The hormonal alteration was mitigated upon the supplementation was exogenous melatonin in the DEX-treated group. Notably, melatonin treatment alone in the experimental group didn’t significantly affect in serum follicular stimulating hormone (FSH) levels compared to the control group. (Figure 8)
3.4.3. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Serum Level of Testosterone
The study revealed a decline in serum testosterone levels in the dexamethasone (DEX) group compared to the control group. However, providing exogenous melatonin treatment to the DEX group restored the circulating testosterone levels of those of the control groups. Animals treated only with melatonin showed a considerably smaller difference from the control group (Figure 9).
3.4.4. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Circulatory Level of Corticosterone
In the DEX-treated group, the circulatory corticosterone level decreased significantly compared to the that of control group, while exogenous administration of melatonin showed a significant decrease in the serum corticosterone level (Figure 10).
3.4.5. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Circulatory Level of Melatonin
Compared to the control group the DEX-treated group showed a decrease in the serum melatonin level. Conversely, the group treated with DEX along with exogenous melatonin exhibited a trend toward melatonin restoration. The same hormonal levels were shown in the melatonin-treated rats as the control one (Figure 11).
3.5. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Histological Changes
Histological examination of the testes in the control group revealed normal structure of the seminiferous tubules and seminiferous epithelium, with active spermatogenesis. Similarly, treatment with melatonin alone did not cause any notable changes in testicular histology, as indicated by the maintained normal architecture of the seminiferous epithelium. Different degree of germ cell degeneration, including degenerating cells, disruption of the germinal epithelium, and its loosing and detachment, were seen following DEX treatment. Nevertheless, these degenerative alterations in the seminiferous tubules were reduced when melatonin was given in addition to DEX (Figure 12)
3.6. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Comet Assay
Damage to heritable DNA was measured through comet assay. There was intact DNA in the CON group, but the DEX-treated group showed damaged DNA presented by tail formation and the CON group’s comet count was lower than that of the DEX-treated group. Melatonin treated to DEX group rats showed less DNA damage compared to the dexamethasone-treated groups. (Figure 13)
3.7. Markers of Oxidative Stress and Tissue Biochemical Status
3.7.1. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Lipid Peroxidation (TBARS level)
The end product of the lipid peroxidation is reactive aldehydes, malondialdehyde (MDA) formed by oxidative stress. MDA is a major bioactive marker of lipid peroxidation. The TBARS reacts with the MDA which forms a fluorescent product formed per mg of testes. The concentration of MDA is found to be higher in the testes tissue that was treated with dexamethasone than in the controlled group. The testes treated with exogenous melatonin show significantly lower levels of concentration of MDA when compared to the dexamethasone-treated rats. (Figure 14)
3.7.2. Effect of Dexamethasone (DEX) and Melatonin (MEL) on Protein Content
There was a significant decrease in the total protein content level in the dexamethasone-treated group of rats while comparing to the control one. Treatment of rats with exogenous melatonin ameliorates the level of protein content from the dexamethasone (DEX) group rats. (Figure 15)
3.7.3. Effect of Dexamethasone (DEX) and Melatonin (MEL) on the Antioxidant Status
SOD Enzyme Activity
In the dexamethasone-treated rats, it shows a significant decline in the SOD enzyme activity in comparison to the control rats. While in the case of exogenous melatonin-treated rats the SOD enzyme activity is comparatively higher than the testes of the dexamethasone-treated group rats. (Figure 16)
Catalase Enzyme Activity
Catalase is an antioxidant enzyme that stimulates the breakdown of H2O2 formed in the tissue. Treatment of dexamethasone decreases the catalytic activity in the testes in comparison to the control group rats' testes. Exogenous testes elevate the catalytic activity of the SOD enzyme when compared to the dexamethasone-treated rats. (Figure 17)
Reduced Glutathione Level (GSH)
The dexamethasone-treated rats showed a significant decrease in the level of glutathione in comparison to the control rats. In the case of exogenous melatonin-treated rats, the glutathione level is comparatively elevated than the testes of the dexamethasone-treated group rats. (Figure 18)
Glutathione Reductase (GR) Level
Rats administered with dexamethasone exhibited a notable reduction in GR activity in the testes. Conversely, there was a significant restoration in the GR activity to levels comparable to those of the CON group. (Figure 19)
Glutathione Peroxidase (GPX) Level
Rats treated with dexamethasone displayed a significant reduction in testicular GPX activity. The use of melatonin in the DEX-treated group, however, significantly boosted the rate of GPX activity in the DEX group (Figure 20).
Glutathione Peroxidase (G6PDH) Level
G6PDH activity was determined to be significantly lower in the reproductive tissue testes. However, treatment with melatonin successfully reinstated G6PDH activity to levels similar to those seen in the control group, as depicted in (Figure 21)
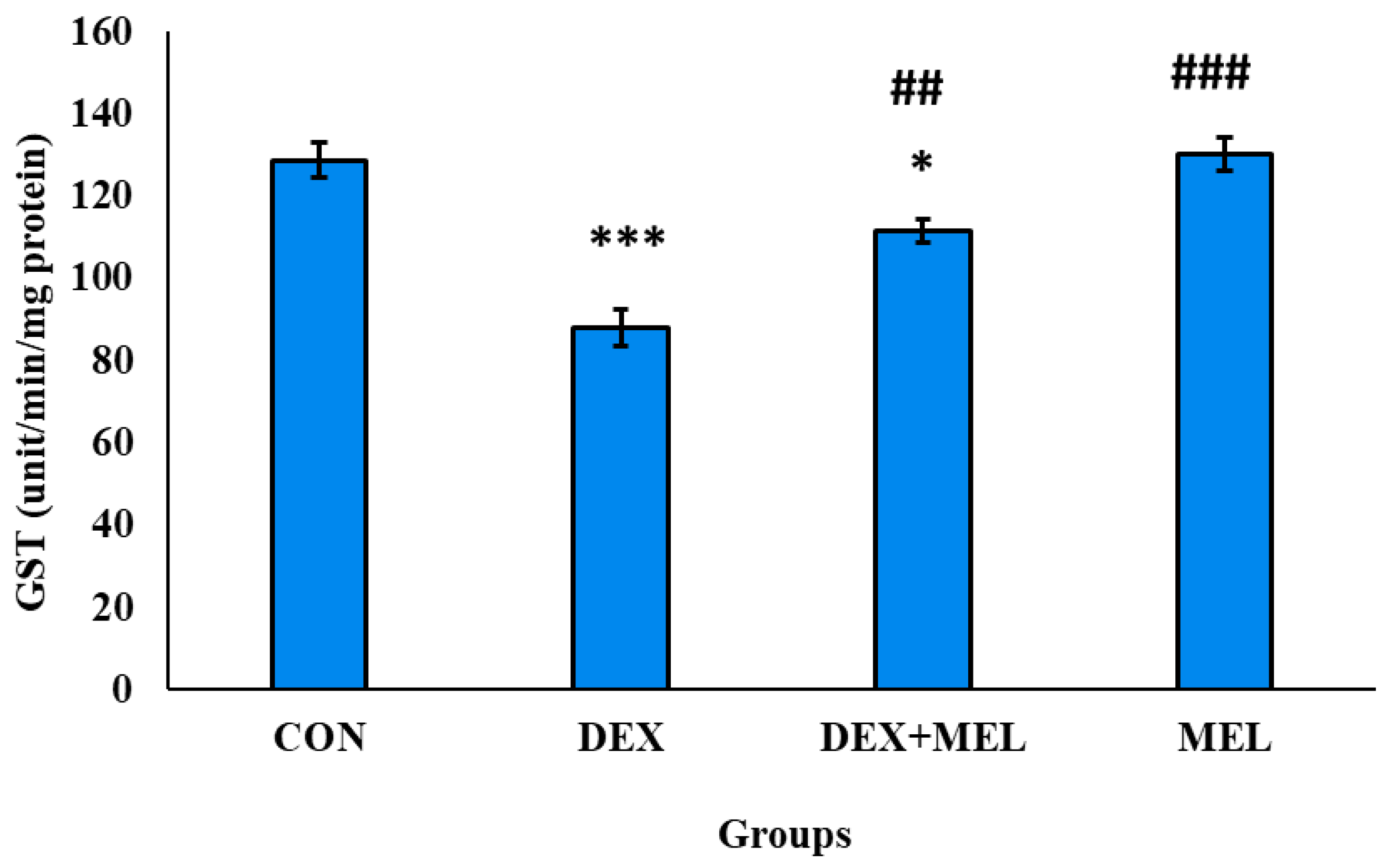
Glutathione S Transferase (GST) Level
Testicular GST activity was significantly reduced in rat testes treated with dexamethasone. However, the rats administered with MEL treated with DEX significantly restored the GST activity. (Figure 22)
4. Discussion
In earlier studies, it was found that dexamethasone administration decreased the rat's body weight [29,30]. This weight reduction can be partly attributed to decreased food intake, as demonstrated in earlier studies [31,32], Additionally, the weight loss observed in these animals is the cause of increased muscle loss and the breakdown of proteins and fats [33]. Following the administration of DEX, sperm production may be interrupted, resulting in a reduction in the size of seminiferous tubules and a notable fall in testicular weight [34,35]. Administering melatonin restored both body weight gain and the weight of testes in rats intoxicated with DEX, bringing them close to normal levels. Therefore, melatonin exhibited a notable effect in mitigating the body weight loss seen in the DEX-treated rat groups. It's plausible that decreased proteolysis occurred in rats when given both dexamethasone and melatonin [36], and the notable improvement in body weight, and testes weight could be credited to melatonin's antioxidant properties, which help prevent cellular damage caused by free radical generation in the reproductive tissue testes. In the DEX-treated group, there was shrinkage in the reproductive tissue [37]. In the control group, there was no such shrinkage found, this is because free radicals attack the cell membrane of these organs which disrupts the structure and finally leads to shrinkage in the testes in the DEX-treated animal compared to the normal saline-treated group. However, in MEL treated group due to a depletion in lipid peroxidation and free radical attack, there was no shrinkage was seen in the testes as the usual structure was regained after the melatonin treatment. In this study, DEX led to significant changes in the histological structure of the testes and a significant decrease in testosterone levels and these findings were consistent with other reports for animals treated with DEX [38,39]. The degradation of Leydig cells and severe oxidative stress could be the cause of these disruptions [40,41,42]. The biochemical changes brought on by DEX-induced toxicity on the testis function indices were verified by the histological changes in the testicular cells in our study [34]. Research demonstrates that DEX suppresses the expression of the gene that codes for the steroidogenic acute regulatory protein (StAR), which is involved in the transportation of cholesterol [43] inside Leydig cells, therefore decreasing the synthesis of testosterone.
Prior studies have shown that DEX might alter a body's healthy equilibrium and disrupt the endocrine system [44]. The current study's outcome indicated that rats exposed to DEX had higher concentrations of FSH and LH. Furthermore, the levels of LH and FSH regulated Sertoli and Leydig cells [44]. As testosterone levels dropped in the DEX group, the synthesis of testosterone was inhibited, which in turn reduced Leydig cells' response to LH. Conversely, the increase in serum FSH levels suggested that spermatogenesis had been destroyed in the experimental group and that this was due to either germ cell loss or injury to the Sertoli cells [45]. A decrease in testosterone levels might be brought on by a rise in cortisol levels, which may inhibit Leydig cells by binding to GC receptors on the cell surface [46]. The findings of this investigation align with other studies that have demonstrated that DEX is a potent modulator of apoptosis and that DEX significantly elevates gonadotropin-releasing hormone, FSH, and LH levels [35,38].
Dexamethasone increases the production of free radicals like ROS, upsetting the balance between oxidants and antioxidants within cells, and leading to oxidative stress [47] (Sadeghzadeh et al., 2020). This stress causes damage to the plasma membrane through lipid peroxidation and interferes with germ-cell communication [48]. A key biomarker indicating lipid peroxidation is TBARS, which was observed to increase in the DEX experimental group. Studies reported that injecting dexamethasone triggered lipid peroxidation and reduced antioxidant activity. Also, a similar study found that dexamethasone caused a higher ROS level and lower mRNA level of either catalase or SOD. Dexamethasone treatment increases MDA levels and decreases SOD levels in testicular [41]. In the current investigation, the total antioxidation ability of the DEX group was significantly lower than that of the CON group. Glutathione (GSH) processes H2O2 and organic peroxides, thereby acting as an important antioxidant. The DEX group in our study showed decreased GSH, which is involved in glyoxalase reactions, ribonucleotide reductions to deoxyribonucleotides, protein regulation, and gene expression through the thiol-disulfide exchange [49].
The present investigation’s DEX administration produced a remarkable drop in testicular GSH content, CAT, and SOD activity when compared to the control group [34,50]. The decreased GSH content in DEX control rats could be due to the decline in the activity of G-6-PD which is directly involved in the metabolism of GSH [51] and the accumulation of ROS. G-6-PDH has a direct link to the synthesis of glutathione its decreased activity causes increased oxidative stress in the tissues, which ultimately results in cell death [34,52]. The other antioxidant enzymes; GR, GPX, and GST antioxidant enzymes, were also shown to be drastically reduced as a result of the high generations of the free radical load. The malondialdehyde levels also reached the control level in the dexamethasone alongside the melatonin-treated group. In other words, melatonin was able to compensate for the undesired effects of dexamethasone. Melatonin has been highly gained a lot of recognition recently due to its antioxidant properties [53], and considering its small size and lipoprotein structure, it may easily cross the biological membranes to reach every area of the cell, including cytosol, mitochondria, nucleus, and DNA to prevent oxidative damage [53]. Lipid peroxidation and all antioxidant enzymes appeared normal in the DEX + MEL group, suggesting that MEL has a significant antioxidant effect in testes as melatonin increases the activity of antioxidant enzymes such as superoxide dismutase, glutathione peroxidase, and glutathione reductase, and as a result, increases the antioxidant potential of the cell [53,54] and reduces DNA fragmentation, lipid peroxidation of the plasma membrane and also the ROS levels. The androgen receptor (AR) which binds to androgen, is essential to androgen action because it may bind DNA and promote the transcription of androgen-dependent genes [55]. Prostate cancer and male infertility have been connected to abnormalities in the AR signaling system. Male genital abnormalities or infertility were linked to mutations or deletions of the AR genes [56]. According to studies conducted in vitro, FSH significantly influences the expression of AR mRNA and the production of proteins in cultivated Sertoli cells [57]. We inferred that the abnormal increase in AR expression caused by DEX resulted from two factors. Firstly, DEX-induced damage to Leydig and Sertoli cells led to decreased testosterone synthesis and concentration, thereby impairing the regulatory role of testosterone on AR expression. Consequently, DEX exposure disrupted the HPG axis, reducing AR expression and weakening the interaction between testosterone and AR. This notably suppressed testicular function in the Wistar albino rats, affecting testicular growth and maintenance [58]. Research has discovered that melatonin can regulate the mRNA expression of the androgen receptor, indicating a possible involvement in the control of steroid receptors. Research provided additional evidence for this, demonstrating the age-dependent effects of darkness exposure or melatonin administration on prostatic androgen receptors. These results suggest that melatonin might play a role in regulating androgen receptor expression in male rats. The majority of melatonin’s effect in mammals is mediated by Mt1 and Mt2 receptors, and melatonin can modify these receptors in response to physiological demands [59]. It is well established that melatonin membrane receptors on the testes control reproductive activity. The expression of Mt1 and Mt2 receptors has been reported in rat testis and Sertoli cells It has been documented that rat testis and Sertoli cells express Mt1 and Mt2 receptors [60,61]. Sertoli cells play a crucial part in spermatogenesis efficiency and fertility [62] (McLachlan et al.,1996). Melatonin increases the responsiveness of the Sertoli cell to FSH during gonadal development, which may help to prevent testicular damage [63]. Studies on neurodegenerative illness have revealed that Mt1 expression and melatonin plasma levels are altered [64]. In our results, the group treated with DEX had a noteworthy reduction in the expression of both Mt1 and Mt2 receptors.
As DEX causes oxidative stress which plays a crucial role in testicular toxicity and germ cell apoptosis which can be blocked by the administration of neurohormone melatonin. Melatonin can protect testes and Leydig cells against oxidative stress, partly through Mt1/Mt2 activation. The recovery of melatonin receptors (Mt1, and Mt2) in the testes lies behind the reason that melatonin also acts as an antioxidative reagent. The oxidative stress generated by DEX exposure was restored after the administration of exogenous melatonin. So, the testicular cells containing Mt1, and Mt2 started expressing again. It was suggested that prolonged melatonin treatment upregulates the Mt1 and Mt2 receptor expression in lymphoid tissues of tropical rodents. Long-term melatonin administration appears to upregulate Mt1 and Mt2 receptor expression in tropical rodent lymphoid tissues [65]. Our findings corroborate other research that found that melatonin medication shows the course of illness by increasing apoptosis in response to melatonin receptor loss.
5. Conclusions
The findings indicated that rats administered with dexamethasone suffered adverse effects on their reproductive health, such as increased lipid peroxidation, diminished organ weight, lowered hormone levels, and alterations in cellular structures. Conversely, rats treated with melatonin in DEX treated group exhibited enhancements across all assessed factors, indicating a potential therapeutic role for melatonin in addressing male reproductive health concerns induced by glucocorticoids. In summary melatonin takes the edge off dexamethasone-induced gonadal functions and improve receptor expression androgen (AR) and melatonin (Mt1/Mt2) via modulating antioxidative enzymes and its protective role.
Conflicts of Interest and Disclosure Statement
All authors declare that they do not have any conflict of interest that could inappropriately influence this manuscript.
Author Contributions
SR conceived the ideas and designed the study. SP performed biochemical and molecular evaluation and preparation of histological slides. SP was also involved in data analysis, interpretation, and drafting of the manuscript. The authors have read and approved the final version of the manuscript. No writing assistance was utilized in the production of this manuscript.
Funding
The author is grateful to the University Grants Commission (Selection and Award Bureau), New Delhi, for providing the Rajiv Gandhi National Fellowship for Other Backward Classes (RGNFOBC) during this study.
Acknowledgments
All authors thank the Department of Zoology, Guru Ghasidas Vishwavidyalaya, Bilaspur, Chhattisgarh, India, for providing support and space and authors special thanks to the Hereditary Biosciences, Bhubaneswar, Odisha for providing lab facilities to perform q RT PCR experiment.
References
- Galano A: Reiter, RJ. Melatonin and its metabolites vs oxidative stress: From individual actions to collective protection. Journal of Pineal Research. 2018 Aug;65(1):e12514.
- Singh, M. and Jadhav, H.R., 2014. Melatonin: functions and ligands. Drug discovery today, 19(9), pp.1410-1418.
- Tan DX, Manchester LC, Reiter RJ et al. Melatonin directly scavenges hydrogen peroxide: a potentially new metabolic pathway of melatonin biotransformation. Free Radic Biol Med 2000; 29:1177–1185.
- Tan DX, Manchester LC, Terron MP et al. Melatonin as a naturally occurring co-substrate of quinone reductase-2, the putative melatonin membrane receptor: hypothesis and significance. J Pineal Res 2007; 43:317–320.
- Kurutas EB, Senoglu M, Yuksel KZ, Unsal V, Altun I. Oxidative/nitrosative stress in patients with modic changes: preliminary controlled study. Spine. 2015 Jul 15;40(14):1101-7.
- Saleh, S.R.; Manaa, A.; Sheta, E.; Ghareeb, D.A.; Abd-Elmonem, N.M. The Synergetic. Effect of Egyptian Portulaca oleracea L. (Purslane) and Cichorium intybus L. (Chicory) Extracts against Glucocorticoid-Induced Testicular Toxicity in Rats through Attenuation of Oxidative Reactions and Autophagy. Antioxidants 2022, 11, 1272. [Google Scholar] [CrossRef] [PubMed]
- Ventimiglia, E.; Pozzi, E.; Capogrosso, P.; Boeri, L.; Alfano, M.; Cazzaniga, W.; Matloob, R.; Abbate, C.; Viganò, P.; Montorsi, F.; et al. Extensive Assessment of Underlying Etiological Factors in Primary Infertile Men Reduces the Proportion of Men With Idiopathic Infertility. Front. Endocrinol. 2021, 12, 801125 [PubMed]. [Google Scholar] [CrossRef]
- Sharma, A. Male infertility; evidences, risk factors, causes, diagnosis and management in human. Ann. Clin. Lab. Res. 2017, 5, 188. [Google Scholar] [CrossRef]
- Lettieri, G.; D’Agostino, G.; Mele, E.; Cardito, C.; Esposito, R.; Cimmino, A.; Giarra, A.; Trifuoggi, M.; Raimondo, S.; Notari, T.; et al. Discovery of the Involvement in DNA Oxidative Damage of Human Sperm Nuclear Basic Proteins of Healthy Young Men Living in Polluted Areas. Int. J. Mol. Sci. 2020, 21, 4198. [Google Scholar] [CrossRef] [PubMed]
- Orazizadeh, M.; Khorsandi, L.; Hashemitabar, M. Toxic effects of dexamethasone on mouse testicular germ cells. Andrologia 2010, 42, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Hanafy AM, Khalil HA. Influence of Chronic Dexamethasone Administration on Reproductive Parameters and Semen Traits in Male of Japanese Quail. Asian J Poultry Sci. 2015;9(4):223-232. [CrossRef]
- Sadi-Guettaf, H.; Bekkouche, F.H. Dexamethasone: Impact on testicular activity. Int. J. Med. Health Sci. 2014, 8, 759–762. [Google Scholar]
- Buckingham, JC. Glucocorticoids: exemplars of multi-tasking. British journal of pharmacology. 2006; 147(S1):S258-S68.
- Hashemitabar M, Orazizadeh M, Khorsandi L. Effect of Dexamethasone on Fas Ligand Expression in Mouse Testicular Germ Cells. ZUMS Journal. 2008;16(62):17-26.
- Danek, J. , 2008. Effect of dexamethasone on the changes of semen quality induced by endotoxin in stallion. Bulletin of the Veterinary Institute in Puawy, 52, 581–589.
- Gametchu B, Watson CS. Correlation of membrane glucocorticoid receptor levels with glucocorticoid-induced apoptotic competence using mutant leukemic and lymphoma cells lines. Journal of cellular biochemistry. 2002;87(2):133-46.
- Chang CS, Chen YT, Yeh SD, Xu QQ, Wang RS, Guillou F (2004) Infertility with defective spermatogenesis and hypotestosteronemia in male mice lacking the androgen receptor in Sertoli cells. Proc Natl Acad Sci 101:6876–6881.
- Tice RR, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae Y, Rojas E, Ryu JC, Sasaki YF. Single cell gel/comet assay: guidelines for in vitro and in vivo genetic toxicology testing. Environ Mol Mutagen 2000; 35: 206-221.
- Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res 1988; 175(1):184-191.
- H. Ohkawa, N. H. Ohkawa, N. Ohishi, K. Yagi, Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 95 (1979) 351-358.
- Lowry, N.J. Rosebrough, AL. Farr, J. Rose, Randall, Protein measurement with the Folin phenol reagent, J. Biol. Chem. 193 (1951) 265-275.
- P. Kakkar, B. P. Kakkar, B. Das, P.N.Viswanathan, A modified spectrophotometric assay of superoxide dismutase. Indian J. Biochem. Biophys. 21 (1983) 192-205.
- R.F. Beers, I.W. R.F. Beers, I.W. Sizer, A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase, J, Biol. Chem. 195 (1952) 133-140.
- Sedlak J, Lindsay RH. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Anal Biochem. 1968; 25:192-205. [CrossRef]
- Goldberg, D. M. (1984). Glutathione reductase. Methods of enzymatic analysis, 3, 258-265.
- Paglia, D. E., & Valentine, W. N. (1967). Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. The Journal of laboratory and clinical medicine, 70(1), 158-169.
- H.A. Ells, H.N. H.A. Ells, H.N. Kirkman, A colorimetric method for assay of erythrocytic glucose-6-phosphate dehydrogenase. Proc. Soc. Exp. Biol. Med. 106 (1961) 607-609.
- Habig, W. H. , Pabst, M. J., & Jakoby, W. B. (1974). Glutathione S-transferases: the first enzymatic step in mercapturic acid formation. Journal of biological Chemistry, 249(22), 7130-7139.
- <b>29. </b>H. Jia, T. H. Jia, T. Yamashita, X. Li, H. Kato, Laurel attenuates dexamethasone-induced skeletal muscle atrophy in vitro and in a rat model, Nutrients 14 (2022) 2029.
- S.P. Kamani, J.K. S.P. Kamani, J.K. Waguia, D. Miaffo, M. Nchouwet, C.D. Kadji, M.W. Kamgaing, R. D. Djimeli, J.M. Ngnitedem, A. Kamanyi, S.W. Ngnokam, Efficacy of Emilia coccinea aqueous extract on inhibition of α-amylase enzyme activity and insulin resistance in dexamethasone treated-rats, Metab. Open (2022), 100193.
- Santos CL, Rafacho A, Bosqueiro JR (2007) Efeitos da administração de dexametasona in vivo sobre glicemia, insulinemia e substrates circulantes são dependentes do tempo de tratamento. Biosci J 23(3):101–110.
- Rafacho A, Giozzet VA, Boschero AC, Bosqueiro JR (2008) Functional alterations in endocrine pancreas of rats with diVerent degrees of dexamethasone-induced insulin resistance. Pancreas 36(3):284–293.
- V.S. Kumar, M.N. V.S. Kumar, M.N. Inamdar, G.L. Viswanatha, Protective effect of lemongrass oil against dexamethasone induced hyperlipidemia in rats: possible role of decreased lecithin cholesterol acetyl transferase activity, Asian Pac. J. Trop. Med. 4 (2012) 658–660.
- Hasona, N.A. , 2018. Grape seed extract attenuates dexamethasone-induced testicular and thyroid dysfunction in male albino rats. Andrologia, 50(5), p.e13002.
- Khorsandi L, Mirhoseini M, Mohamadpour M, Orazizadeh M, Khaghani S. Effect of curcumin on dexamethasone induced testicular toxicity in mice. Pharm Biol. 2013;51(2):206-212. [CrossRef]
- Nkono, B. L. N. Y., Sokeng, S. D., Désiré, D. D. P., & Kamtchouing, P. (2014). Antihyperglycemic and antioxydant properties of alstonia boonei De wild. (Apocynaceae) stem bark aqueous extract in dexamethasone-induced hyperglycemic rats. Int. J. Diabetes Res, 3(3), 27-35.
- Patel, S. , & Rai, S. (2023). Glucocorticoid-Induced Dose-Dependent Reproductive Impairments in Male Albino Rat. Journal of Endocrinology and Reproduction, 27(2), 99–108. [CrossRef]
- Gao HB, Tong MH, Hu YQ, et al. Mechanisms of glucocorticoid-induced Leydig cell apoptosis. Mol Cell Endocrinol. 2003;199(1-2):153-163. [CrossRef]
- Ing, N. H., Brinsko, S. P., Curley, K. O., Forrest, D. W., Love, C. C., Hinrichs, K., … Welsh, T. H. (2015). Dexamethasone acutely regulates endocrine parameters in stallions and subsequently affects gene expression in testicular germ cells. Animal Reproduction Science, 152, 47–54. [CrossRef] [PubMed]
- Chen, Y., Wang, Q., Wang, F. F., Gao, H. B., & Zhang, P. (2012). Stress induces glucocorticoid-mediated apoptosis of rat Leydig cells in vivo. Stress, 15, 74–84.
- Jeje SO, Akindele OO, Balogun ME, Raji Y. Maternal treatment with dexamethasone during lactation delays male puberty and disrupts reproductive functions via hypothalamic-pituitary-gonadal axis alterations. Pathophysiology. 2016;23(1):43-49.
- Yun, H. J., Ji-Yeon, L., & Myoung, H. K. (2016). Prenatal exposure to dexamethasone disturbs sex-determining gene expression and fetal testosterone production in male embryos. Biochemical and Biophysical Research Communications, 471, 149–155.
- Hu GX, Lian QQ, Lin H, et al. Rapid mechanisms of glucocorticoid signaling in the Leydig cell. Steroids 2008; 73(9–10): 1018–1024.
- Arab Dolatabadi A, Rezaei Zarchii S. The effect of prescription of different Dexamethasone doses on reproductive system. Biomed Res. 2015;26(4):656-660.
- Mohamadpour M, Mollajani R, Sarabandi F, Hosseini F, Mohsenkia M, Erfanizadeh M, Soleimani A, Kargar E, Mosallaei H, Yavarahmadi M. Protective effect of grape seed extract on dexamethasone-induced testicular toxicity in mice. Crescent Journal of Medical and Biological Sciences. 2020;7(1):59-65.
- Wang FF, Wang Q, Chen Y, Lin Q, Gao HB, Zhang P. Chronic stress induces ageing-associated degeneration in rat Leydig cells. Asian J Androl. 2012;14(4):643-648. [CrossRef]
- Sadeghzadeh, F.; Mehranjani, M.S.; Mahmoodi, M. Vitamin C ameliorates the adverse effects of dexamethasone on sperm motility, testosterone level, and spermatogenesis indexes in mice. Hum. Exp. Toxicol. 2019, 38, 409–418. [Google Scholar] [CrossRef] [PubMed]
- El-Sokkary GH, Abdel-Rahman GH, Kamel ES (2005) Melatonin protects against lead-induced hepatic and renal toxicity in male rats. Toxicology 213: 25–33.
- Kumi-Diaka J, Townsend J. Effects of genistein isoflavone (4′, 5′, 7-trihydroxyisoflavone) and dexamethasone on functional characteristics of spermatozoa. J Med Food. 2001;4(1):39-47. [CrossRef]
- Mukherjee A, Haldar C, Vishwas DK. Melatonin prevents dexamethasone-induced testicular oxidative stress and germ cell apoptosis in golden hamster, M esocricetus auratus. Andrologia. 2015 Oct;47(8):920-31.
- Tang HY, Ho HY, Wu PR, Chen SH, Kuypers FA, Cheng ML, Chiu DT. Inability to maintain GSH pool in G6PD-deficient red cells causes futile AMPK activation and irreversible metabolic disturbance. Antioxid Redox Signal. 2015 Mar 20;22(9):744-59. [CrossRef] [PubMed]
- Rizk SM, Zaki HF, Mina MA. Propolis attenuates doxorubicin-induced testicular toxicity in rats. Food and chemical toxicology. 2014 May 1;67:176-86.
- Othman AI, Edrees GM, El-Missiry MA, Ali DA, Aboel-Nour M, Dabdoub BR. Melatonin controlled apoptosis and protected the testes and sperm quality against bisphenol A-induced oxidative toxicity. Toxicology and Industrial Health. 2016 Sep;32(9):1537-49.
- Reiter RJ, Tan DX, Osuna C, et al. (2000) Actions of melatonin in the reduction of oxidative stress. Journal of Biomedical Science 7: 444–458.
- Zhu LJ, Hardy MP, Inigo IV, Huhtaniemi I, Bardin CW, Moo-Young AJ (2000) Effects of androgen-on-androgen receptor expression in rat testicular and epididymal cells: a quantitative immunohistochemical study. Biol Reprod 63:368–376.
- Wang L, Su DZ, Wang WS (1994) Studies on the aluminum content in wheat flour products. Biomed Environ Sci 7(1):91–99.
- Blok LJ, Themmen AP, Peters AH, Trapman J, Baarends WM, Hoogerbrugge JW, Grootegoed JA (1992) Transcriptional regulation of androgen receptor gene expression in Sertoli cells and other cell types. Mol Cell Endocrinol 88:153–164.
- Sun H, Hu C, Jia L, Zhu Y, Zhao H, Shao B, Wang N, Zhang Z, Li Y. Effects of aluminum exposure on serum sex hormones and androgen receptor expression in male rats. Biological trace element research. 2011 Dec;144:1050-8.
- Dubocovich ML, Markowska M. Functional MT 1 and MT 2 melatonin receptors in mammals. Endocrine. 2005 Jul;27:101-10.
- Izzo, G.; Francesco, A.; Ferrara, D.; Campitiello, M.R.; Serino, I.; Minucci, S.; d’Istria, M. Expression of melatonin (MT1, MT2) and melatonin-related receptors in the adult rat testes and during development. Zygote 2010, 3, 257–264 [CrossRef] [PubMed]. [Google Scholar] [CrossRef] [PubMed]
- Rocha, C.S.; Martins, A.D.; Rato, L.; Silva, B.M.; Oliveira, P.F.; Alves, M.G. Melatonin alters the glycolytic profile of Sertoli cells: Implications for male fertility. Mol. Hum. Reprod. 2014, 20, 1067–1076. [Google Scholar] [CrossRef] [PubMed]
- McLachlan, R.I.; Wreford, N.G.; O’Donnell, L.; de Kretser, D.M.; Robertson, D.M. The endocrine regulation of spermatogenesis: Independent roles for testosterone and FSH. J. Endocrinol. 1996, 148, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Heindel, J.J.; Jackson, F.L.; Berkowitz, A.S. Role of the pineal in the alteration of hamster Sertoli cell responsiveness to FSH during testicular regression. J. Androl. 1984, 5, 211–215 [PubMed]. [Google Scholar] [CrossRef] [PubMed]
- Zhang H, Zhang Y. Melatonin: a well-documented antioxidant with conditional pro-oxidant actions. Pineal Res 2014; 57: 131-146.
- Gupta S, Haldar C. Nycthemeral variation in melatonin receptor expression in the lymphoid organs of a tropical seasonal breeder Funambulus pennanti. Journal of Comparative Physiology A. 2014 Dec; 200:1045-55.
Figure 1.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on body weight of male rats. The graph represents Mean + SE, N=6. Histogram represents ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/DEX+MEL and # DEX vs DEX+MEL.
Figure 1.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on body weight of male rats. The graph represents Mean + SE, N=6. Histogram represents ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/DEX+MEL and # DEX vs DEX+MEL.
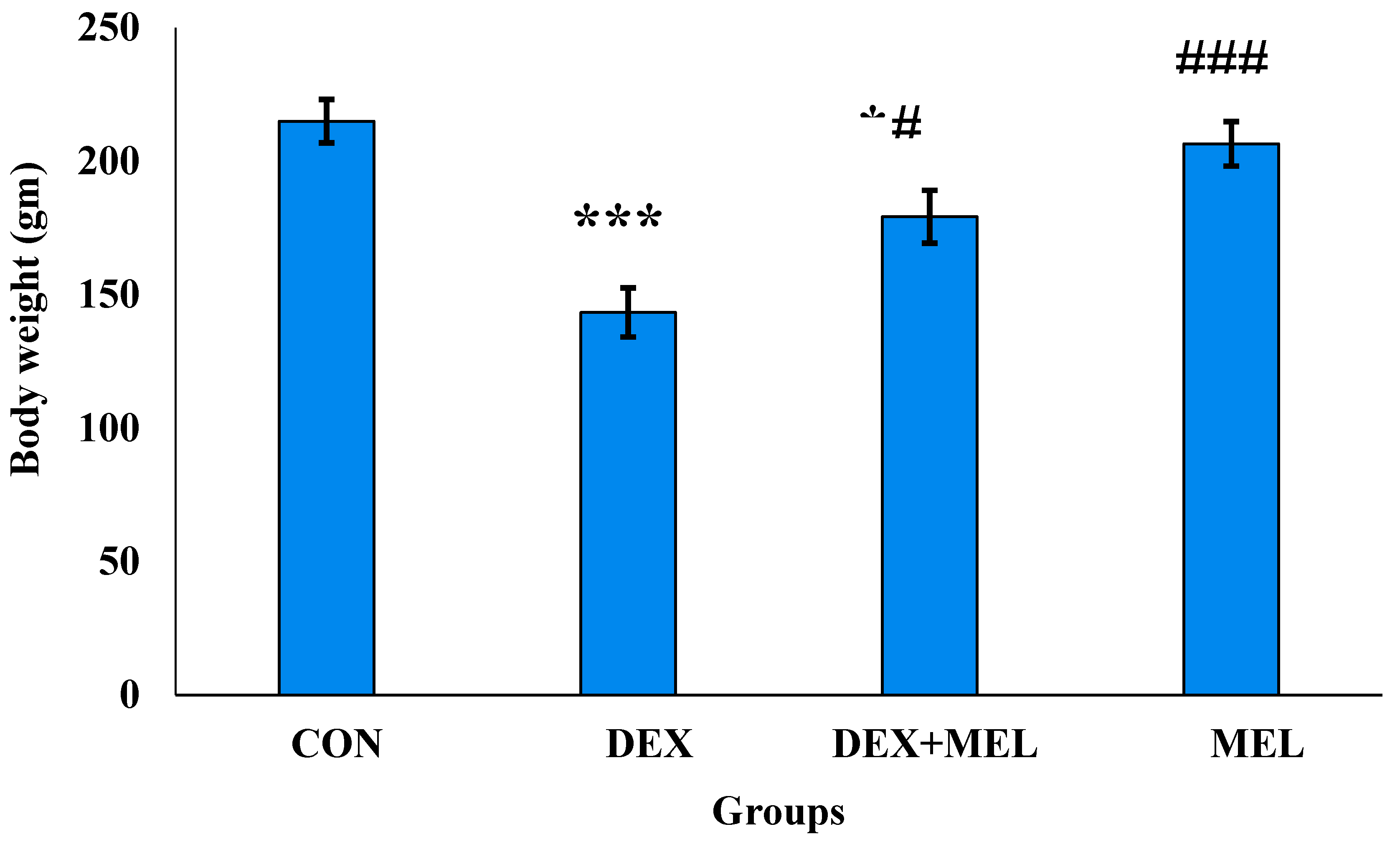
Figure 2.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on weight of testes. The graph represents Mean + SE, N=6. Histogram represents ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/DEX+MEL and # DEX vs DEX+MEL.
Figure 2.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on weight of testes. The graph represents Mean + SE, N=6. Histogram represents ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/DEX+MEL and # DEX vs DEX+MEL.
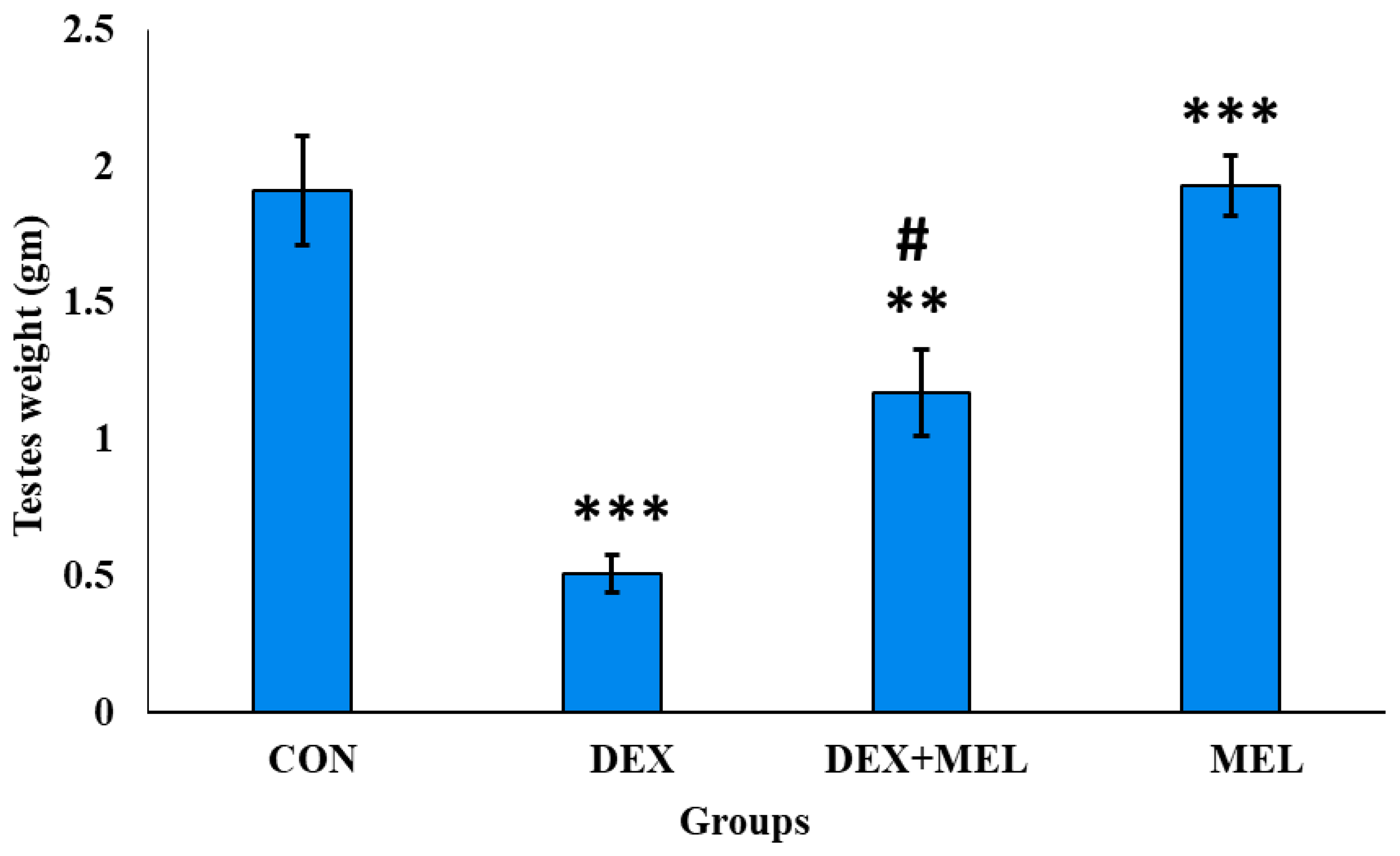
Figure 3.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on the morphology of testes. The melatonin-treated group showed no change in morphology as compared with the control group, whereas DEX treated group showed shrinkage as compared to the control. Melatonin treatment along with DEX showed recovery in morphology as compared with the DEX-treated group.
Figure 3.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on the morphology of testes. The melatonin-treated group showed no change in morphology as compared with the control group, whereas DEX treated group showed shrinkage as compared to the control. Melatonin treatment along with DEX showed recovery in morphology as compared with the DEX-treated group.
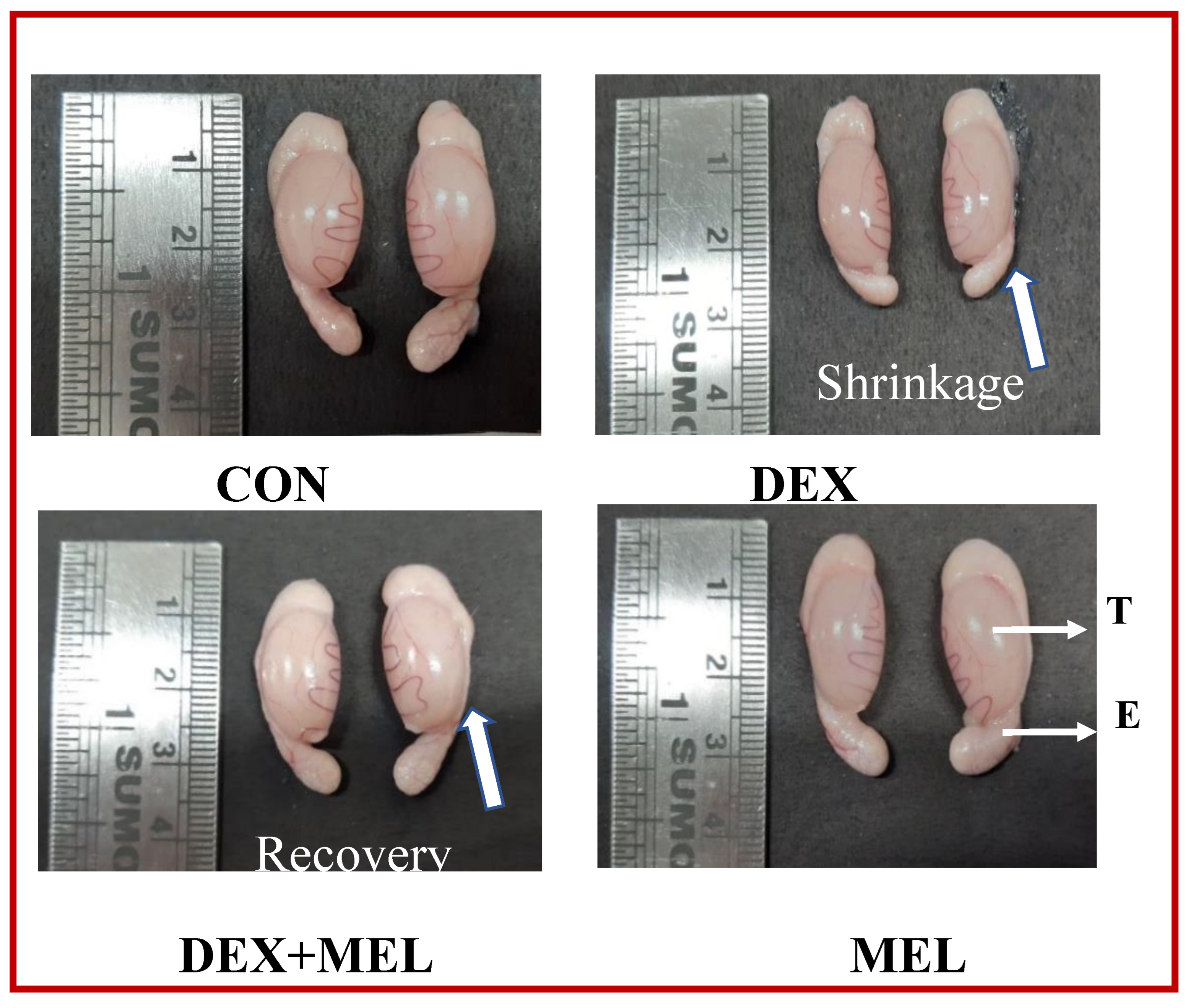
Figure 4.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of AR (Androgen receptor) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
Figure 4.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of AR (Androgen receptor) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
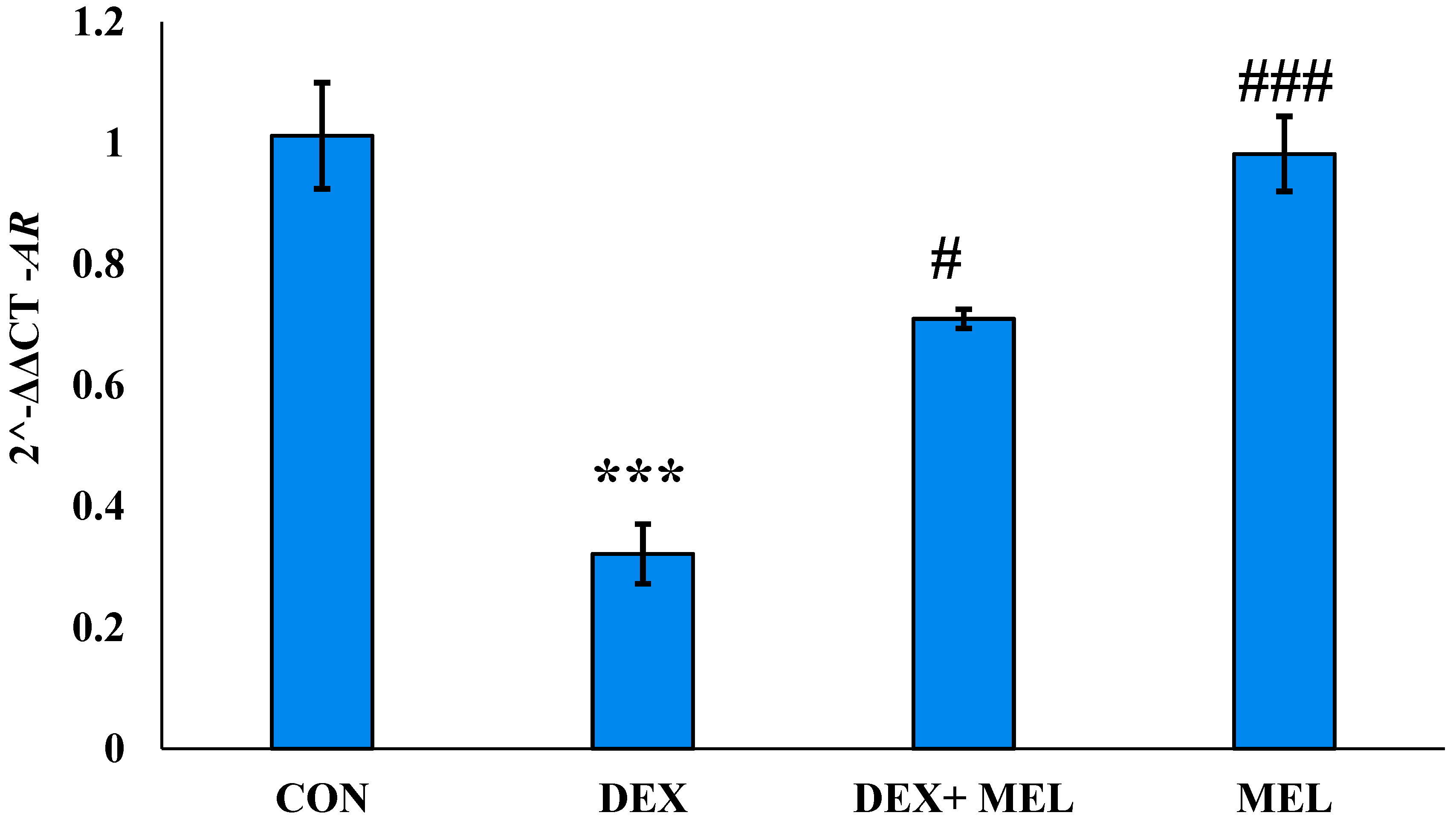
Figure 5.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of Mt1 (Melatonin receptor-1) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
Figure 5.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of Mt1 (Melatonin receptor-1) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
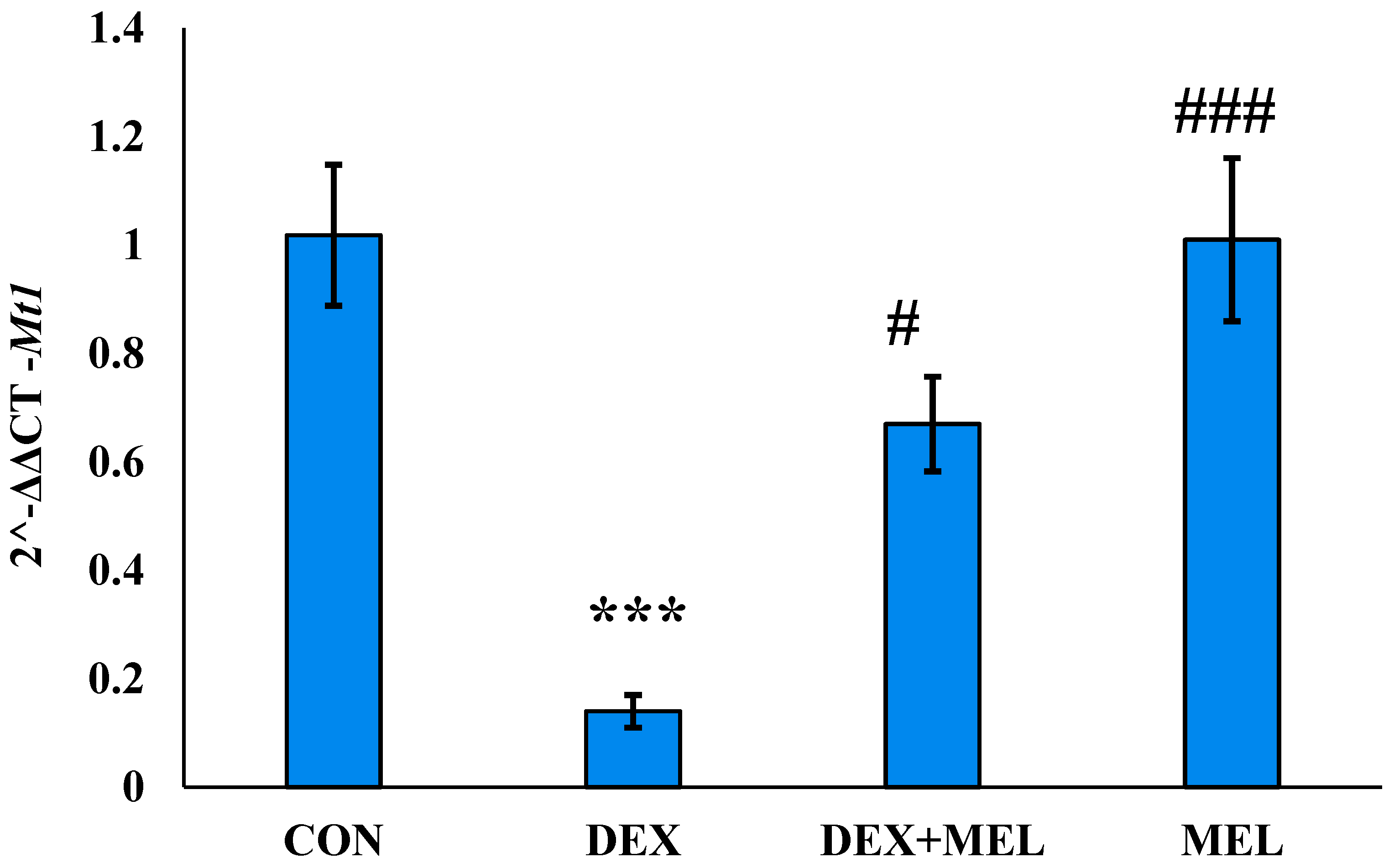
Figure 6.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of Mt2 (Melatonin receptor-2) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
Figure 6.
Histogram represents the relative gene expression showing fold change (2^-ΔΔCT) of Mt2 (Melatonin receptor-2) in testes of DEX-induced rat model. Mean ± SE; N=6; */#P≤0.05, **/##P≤0.01, ***/###P≤0.001; * represents CON vs DEX; CON vs DEX+MEL; CON vs MEL: # represents DEX vs DEX+MEL; DEX vs MEL. CON= Control; DEX= Dexamethasone; DEX+MEL= Dexamethasone+ Melatonin; MEL=Melatonin.
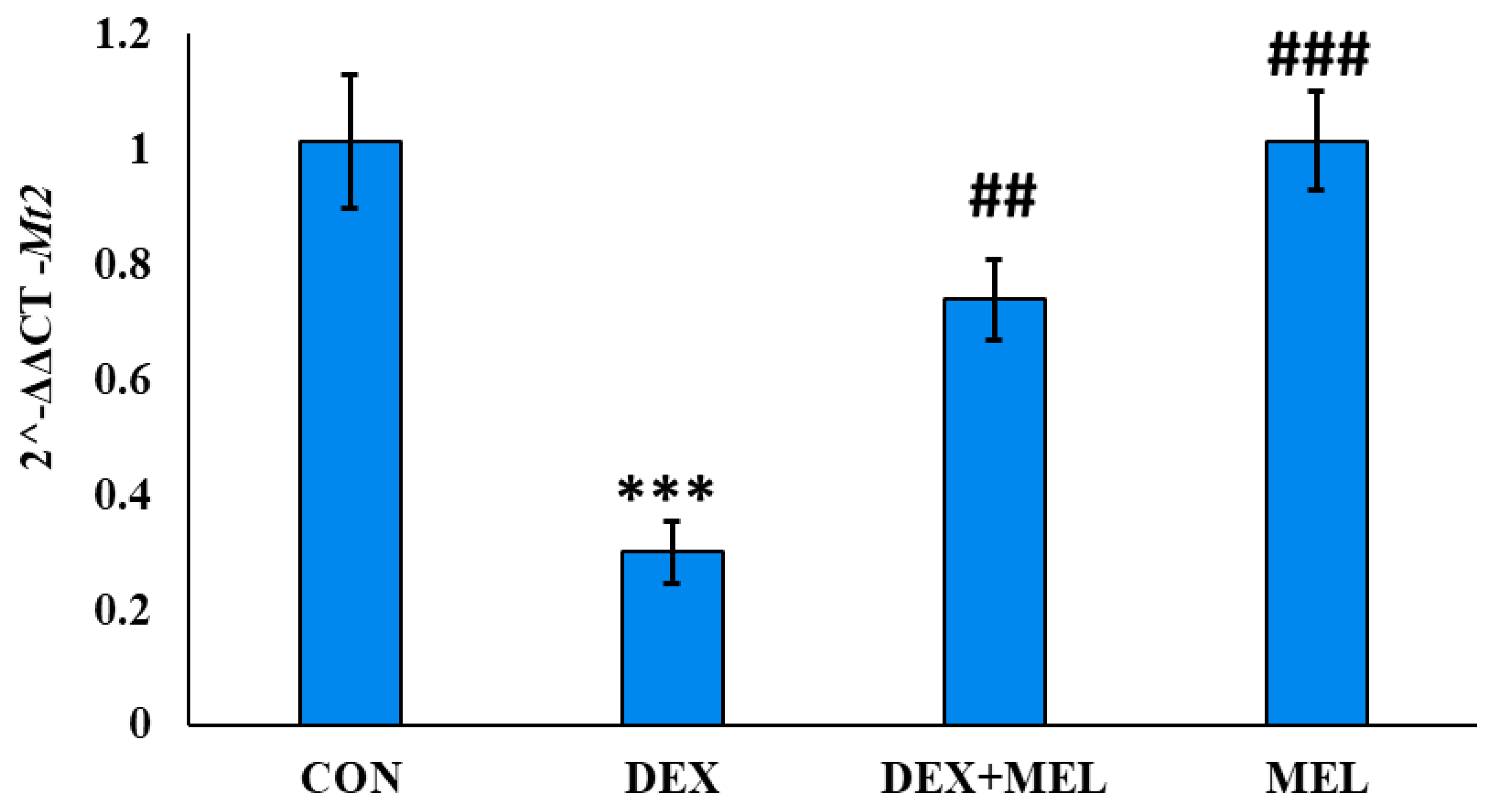
Figure 7.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum LH level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 7.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum LH level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
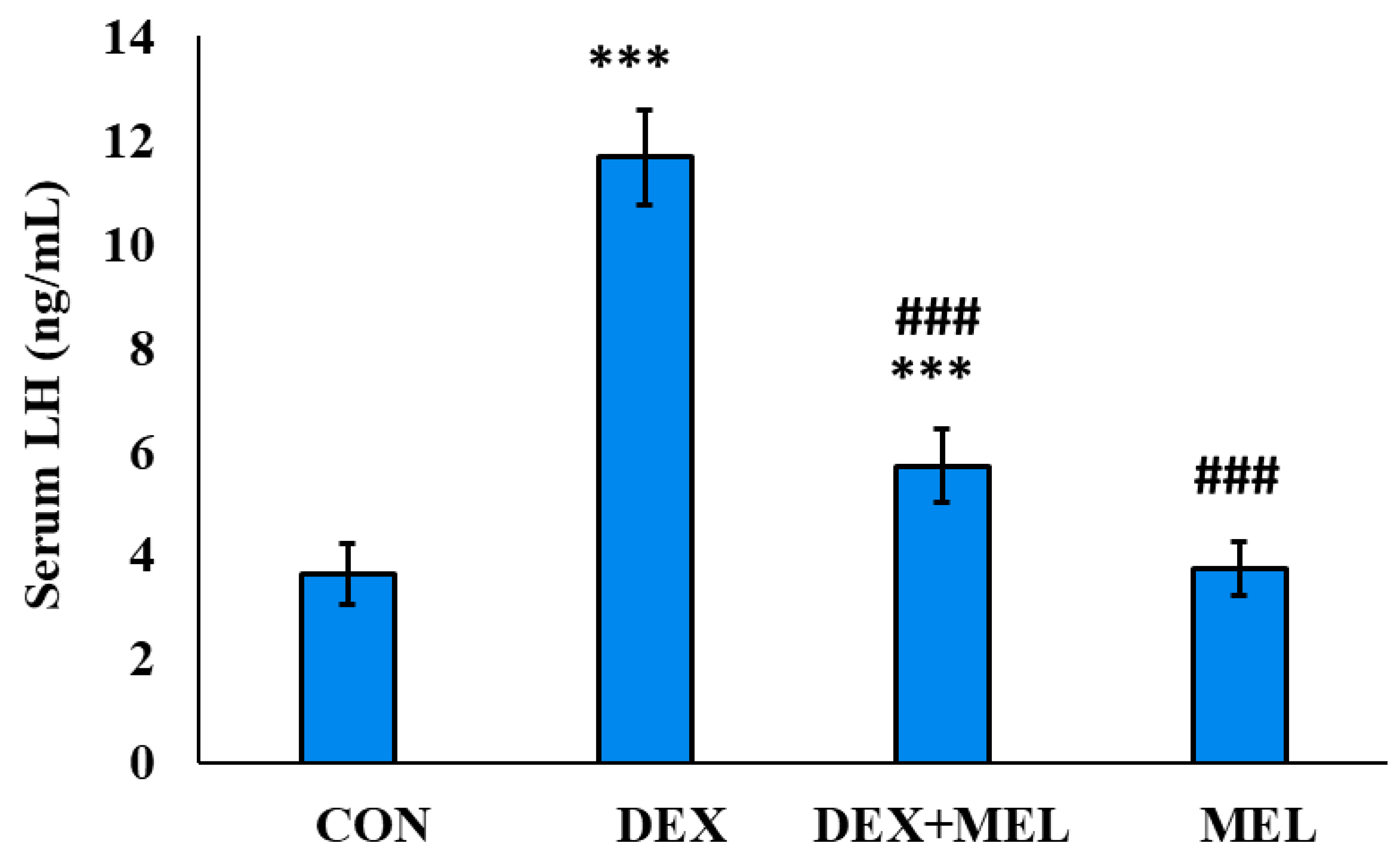
Figure 8.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum FSH level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 8.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum FSH level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.

Figure 9.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum testosterone level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 9.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum testosterone level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
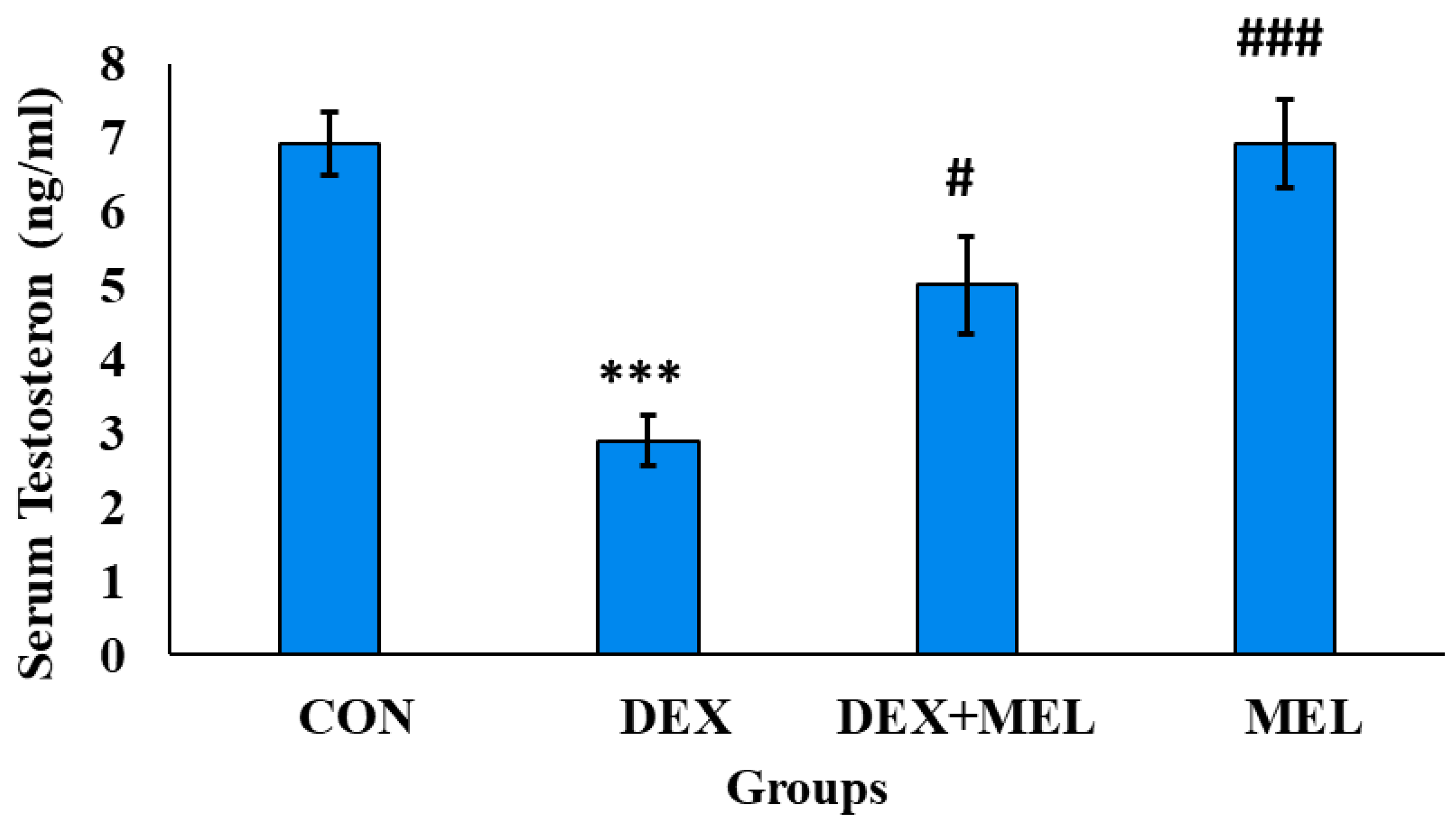
Figure 10.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum corticosterone level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 10.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum corticosterone level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
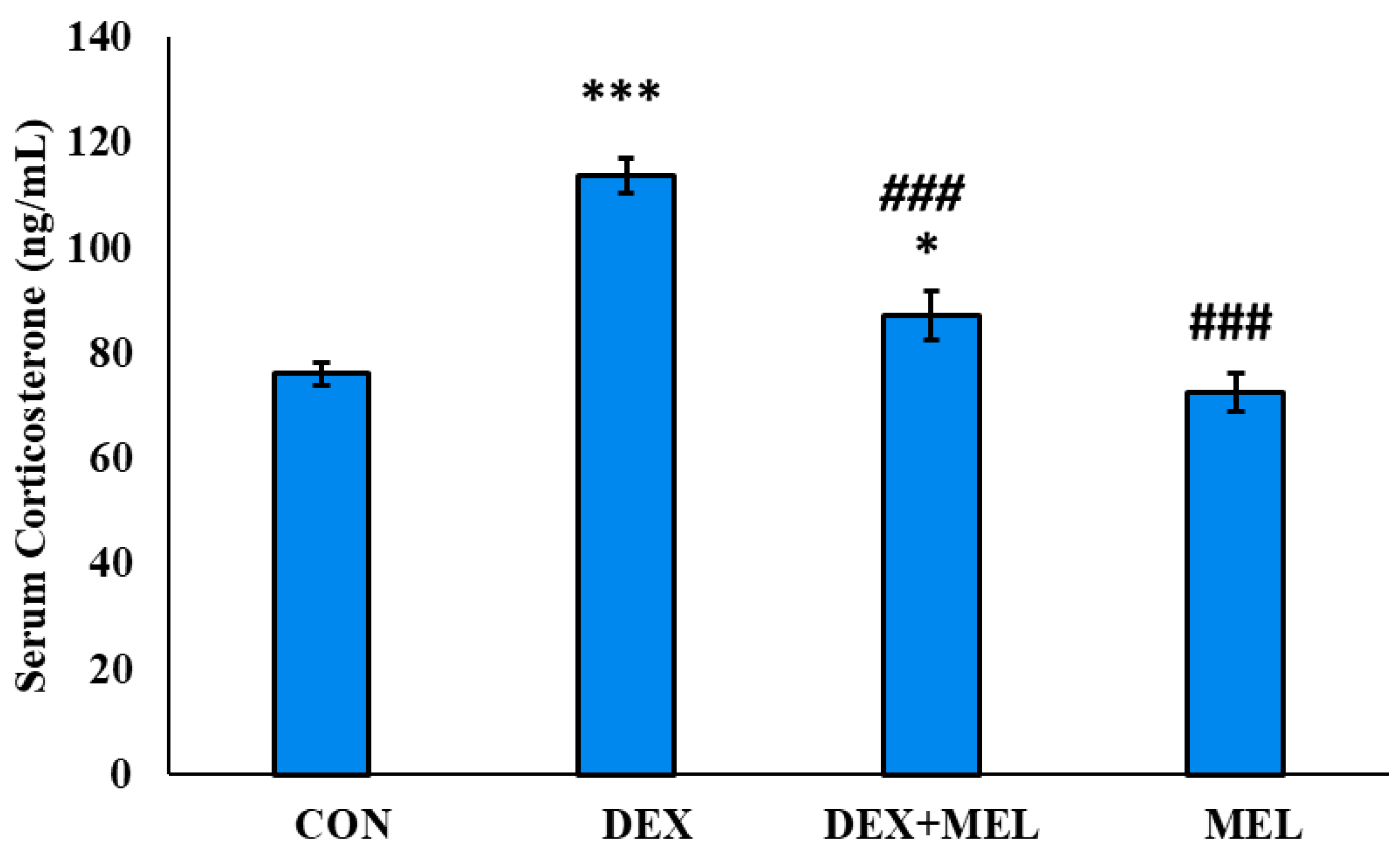
Figure 11.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum melatonin level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 11.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on serum melatonin level of the rat. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
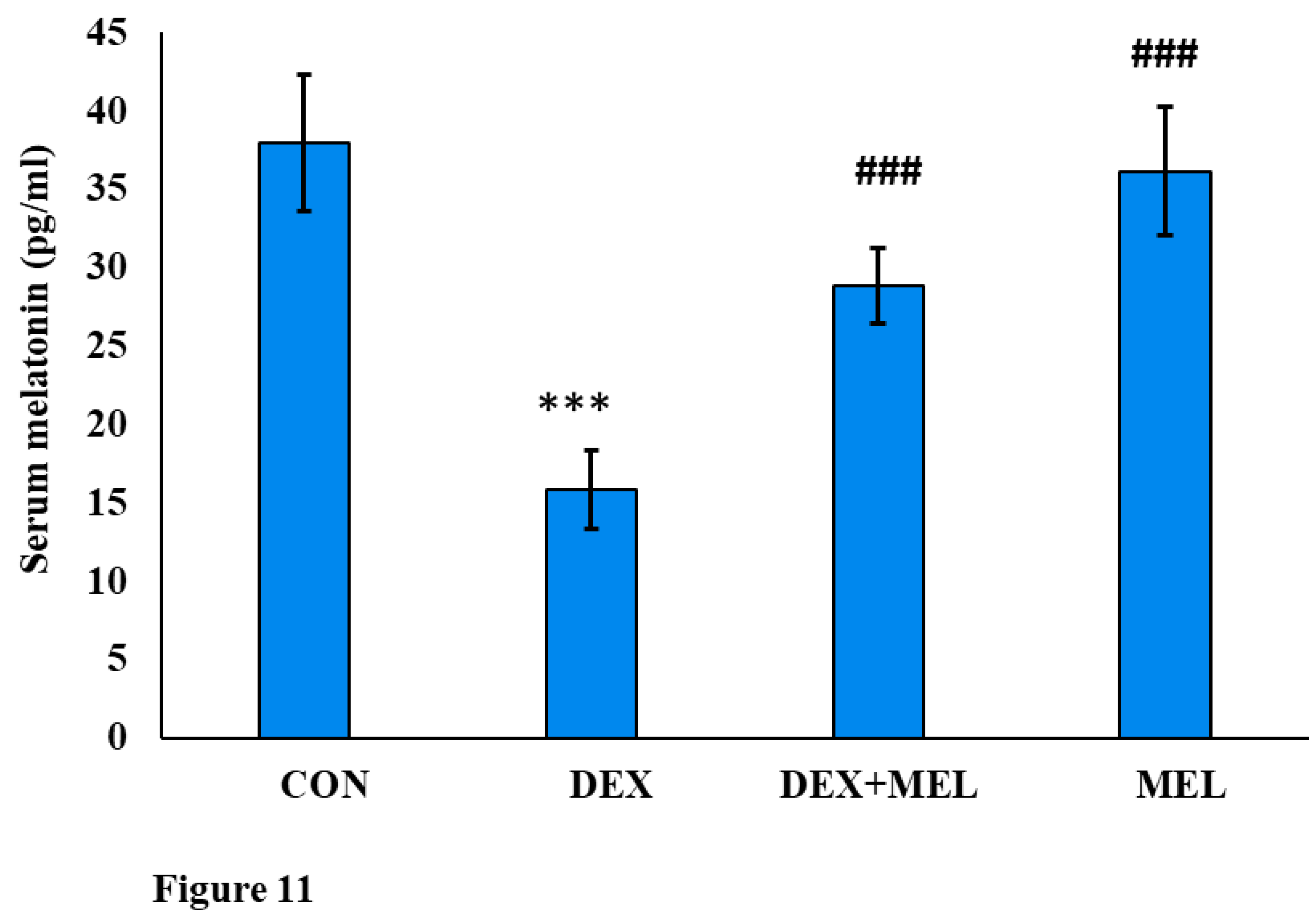
Figure 12.
TS of Testes. The histological assessment showed normal histoarchitecture (H&E staining) of seminiferous tubule and seminiferous epithelium and exhibited active spermatogenesis in the control group. Melatonin treatment (alone) showed no significant alterations in testicular histology as evident from the normal histoarchitecture of the seminiferous epithelium. DEX treatment showed varying degrees of germ cell degenerative changes characterized by the presence of degenerating cells, germinal epithelium disruption, and loosening and detachment of germinal epithelium (exfoliation). Melatonin treatment along with DEX decreased the degenerative changes in the seminiferous tubule. Magnification of upper panel-100X, lower panel-200X. CON=Control, DEX=Dexamethasone, SET= Seminiferous tubules, SZ=Spermatozoa, SC=Spermatocytes, ST=Spermatids, LC=Leydig Cells, D=Degeneration, R=Recovery.
Figure 12.
TS of Testes. The histological assessment showed normal histoarchitecture (H&E staining) of seminiferous tubule and seminiferous epithelium and exhibited active spermatogenesis in the control group. Melatonin treatment (alone) showed no significant alterations in testicular histology as evident from the normal histoarchitecture of the seminiferous epithelium. DEX treatment showed varying degrees of germ cell degenerative changes characterized by the presence of degenerating cells, germinal epithelium disruption, and loosening and detachment of germinal epithelium (exfoliation). Melatonin treatment along with DEX decreased the degenerative changes in the seminiferous tubule. Magnification of upper panel-100X, lower panel-200X. CON=Control, DEX=Dexamethasone, SET= Seminiferous tubules, SZ=Spermatozoa, SC=Spermatocytes, ST=Spermatids, LC=Leydig Cells, D=Degeneration, R=Recovery.
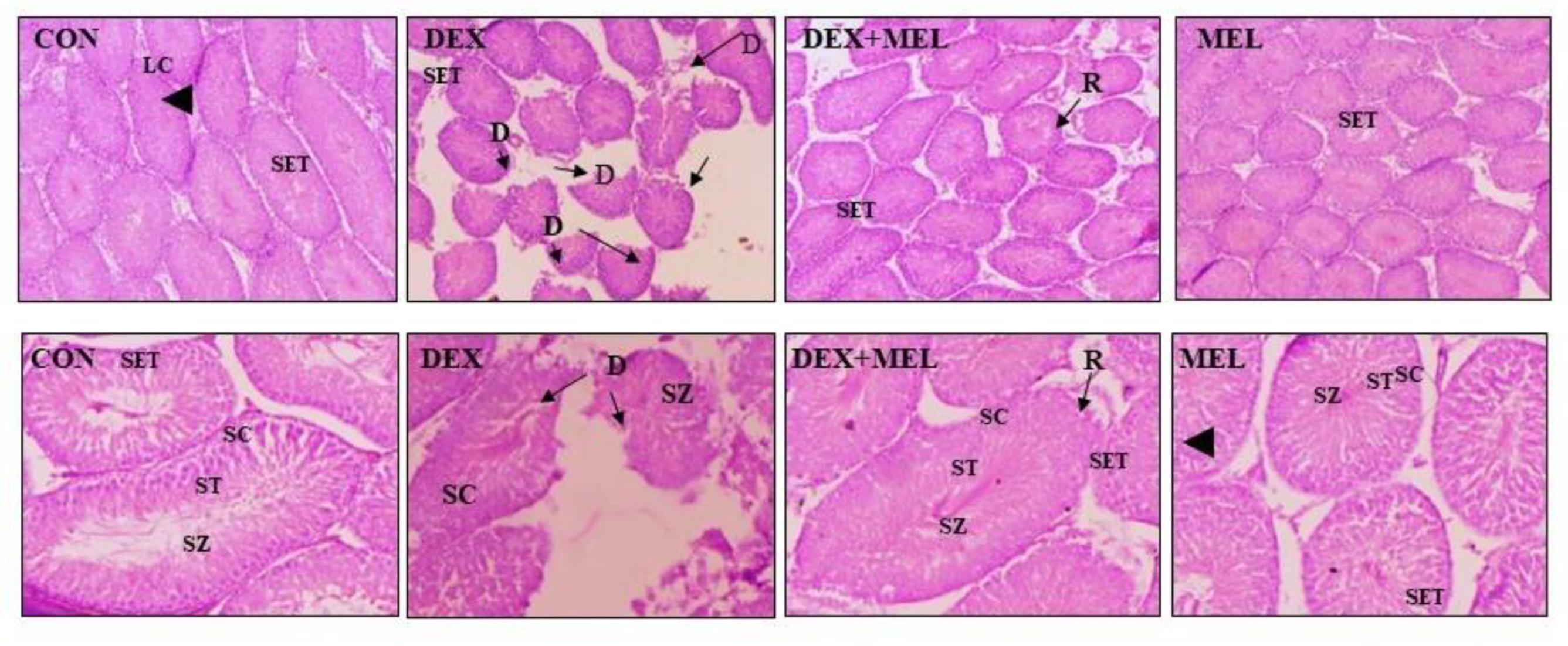
Figure 13.
Fluorescent photomicrograph of the rat testes after dexamethasone and melatonin treatment as determined by single gel electrophoresis with Ethidium bromide (EtBr) stain. A-CON-Control group presenting intact DNA. B-DEX-showing damage DNA represented by tail marked with the blue arrow. C- presented very little damage DNA marked with the green arrow, D- MEL group presented the same as the control group marked with the white arrow. Magnification X 200.
Figure 13.
Fluorescent photomicrograph of the rat testes after dexamethasone and melatonin treatment as determined by single gel electrophoresis with Ethidium bromide (EtBr) stain. A-CON-Control group presenting intact DNA. B-DEX-showing damage DNA represented by tail marked with the blue arrow. C- presented very little damage DNA marked with the green arrow, D- MEL group presented the same as the control group marked with the white arrow. Magnification X 200.
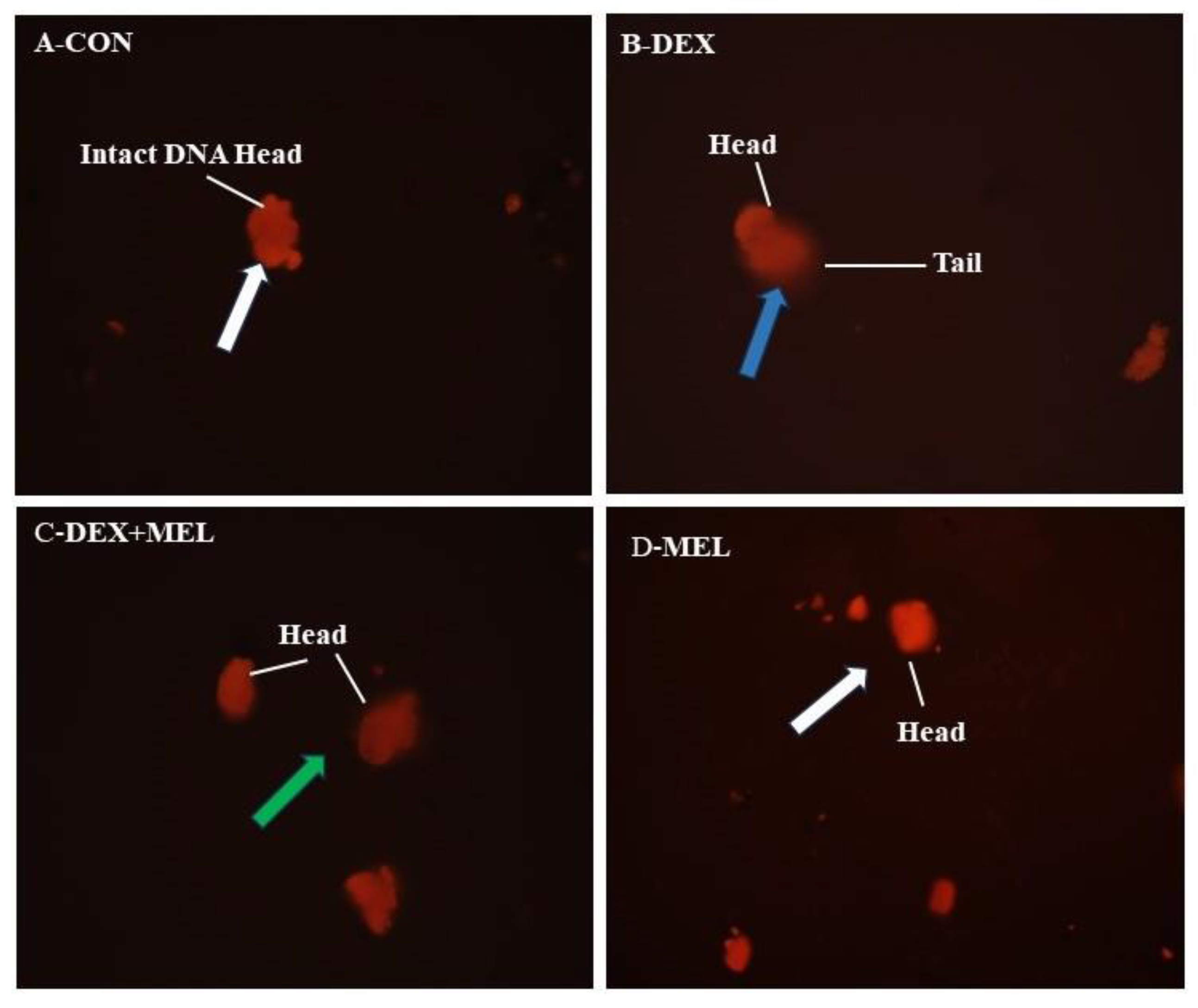
Figure 14.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on lipid peroxidation level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 14.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on lipid peroxidation level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
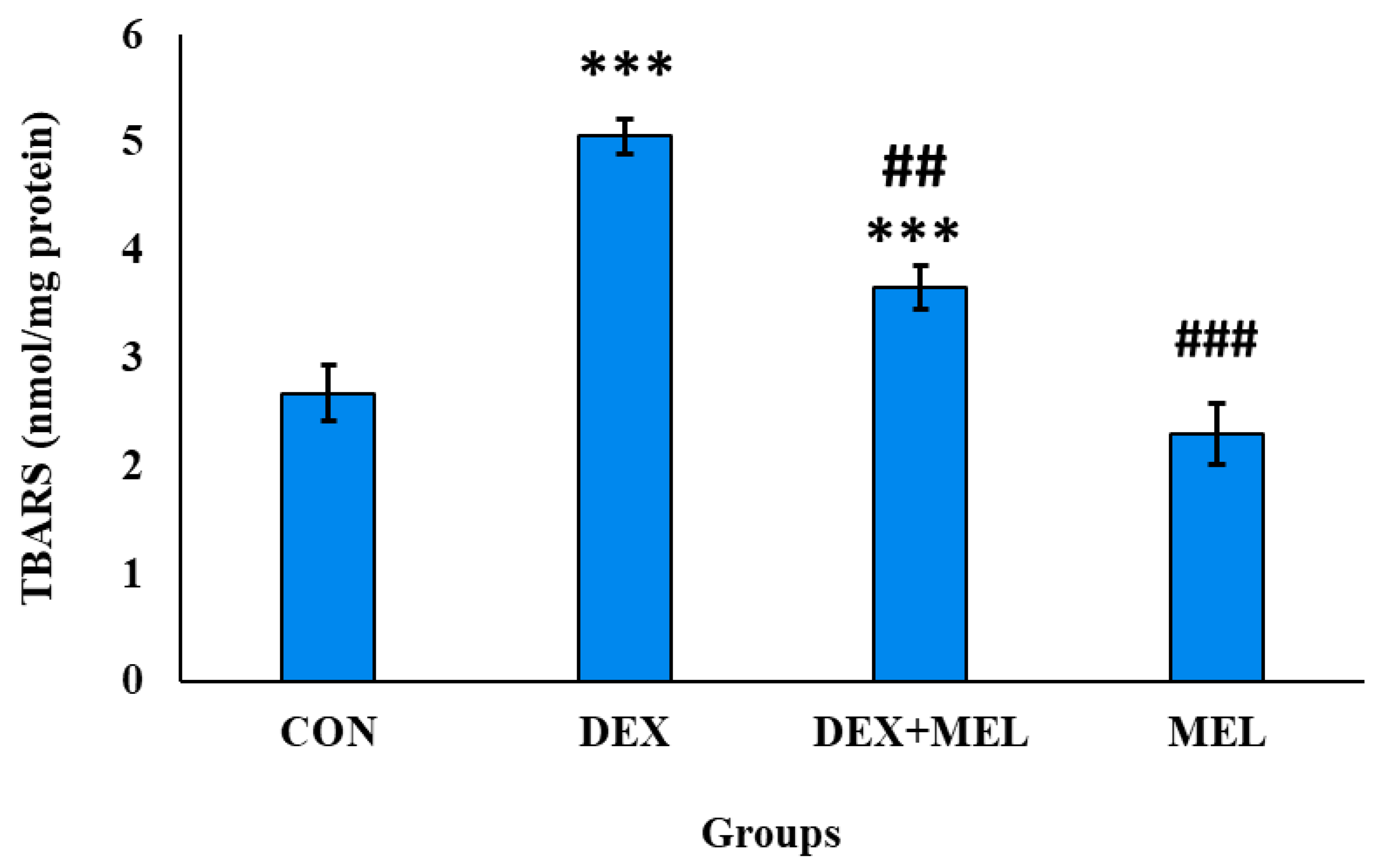
Figure 15.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on total protein content of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 15.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on total protein content of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
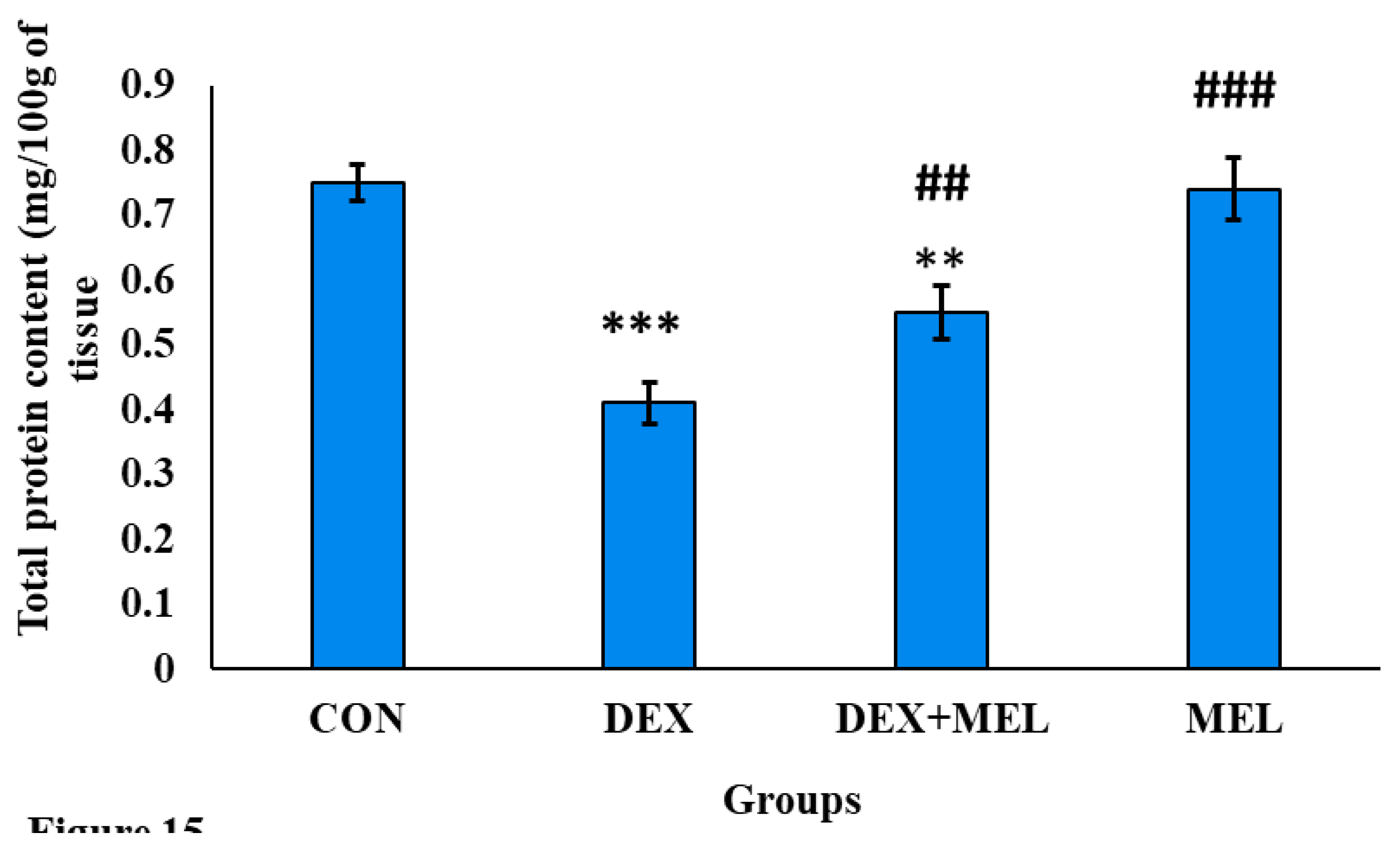
Figure 16.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on superoxide dismutase (SOD) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 16.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on superoxide dismutase (SOD) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
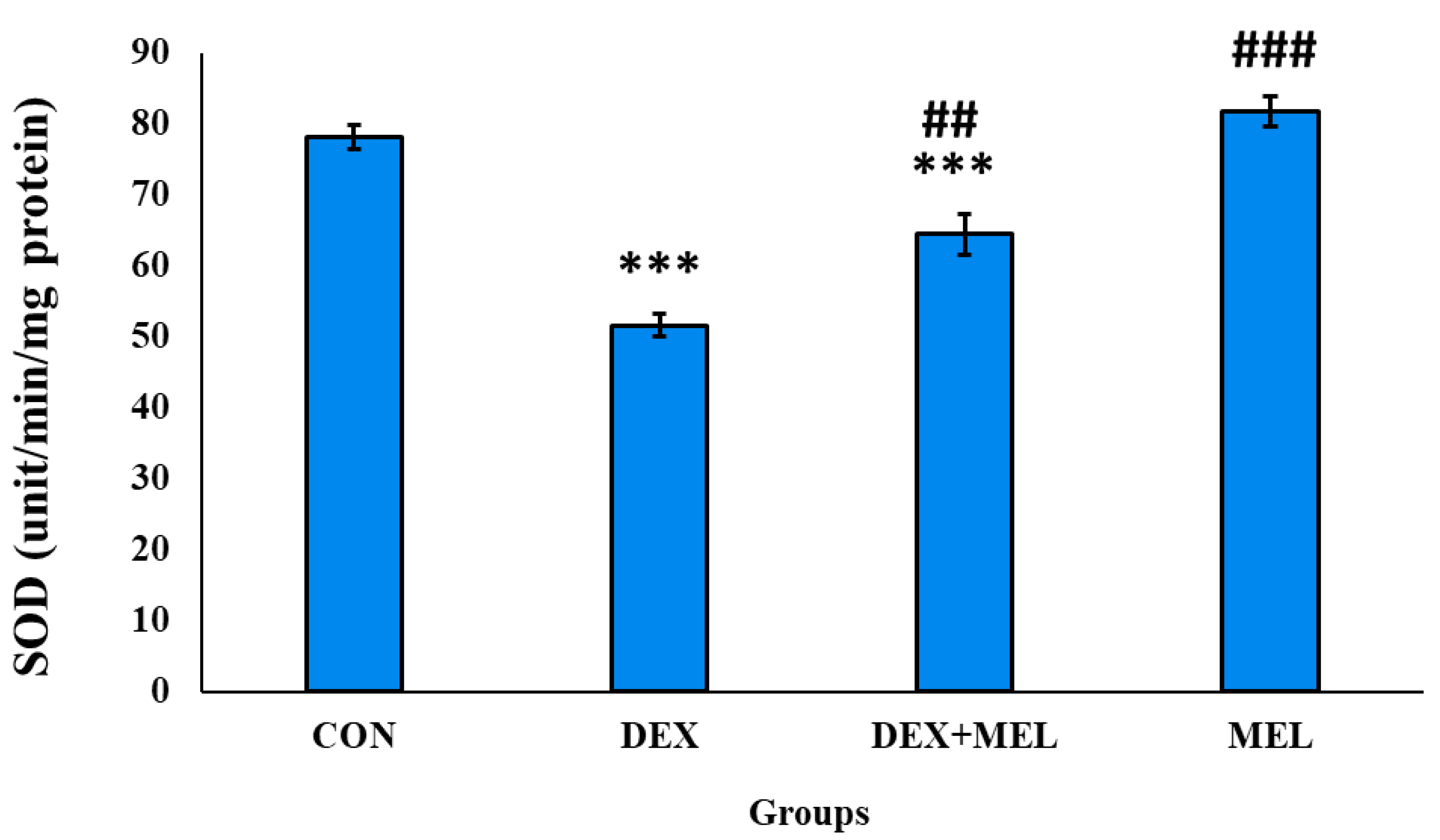
Figure 17.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on catalase (CAT) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 17.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on catalase (CAT) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
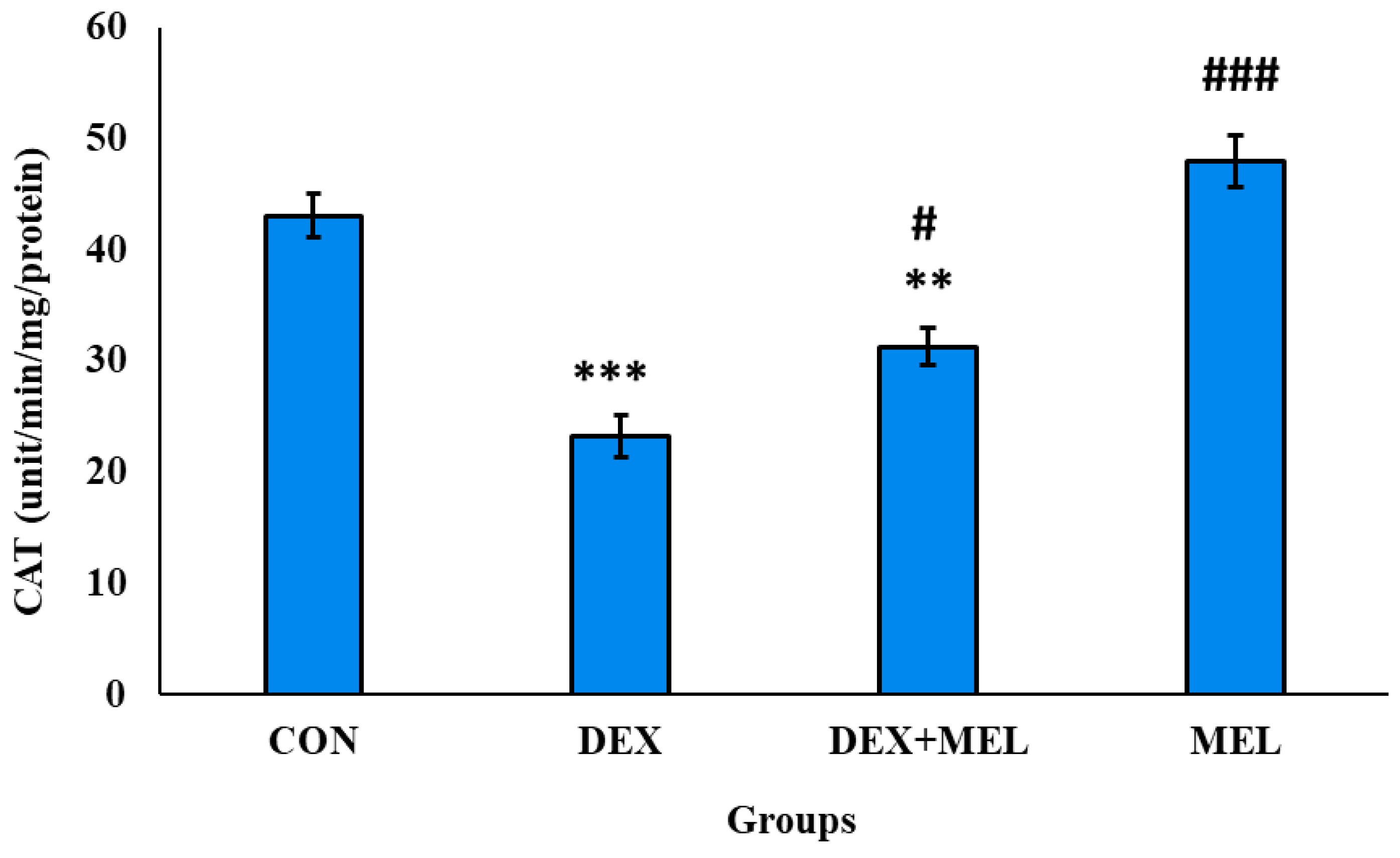
Figure 18.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on reduced glutathione (GSH) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 18.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on reduced glutathione (GSH) level of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
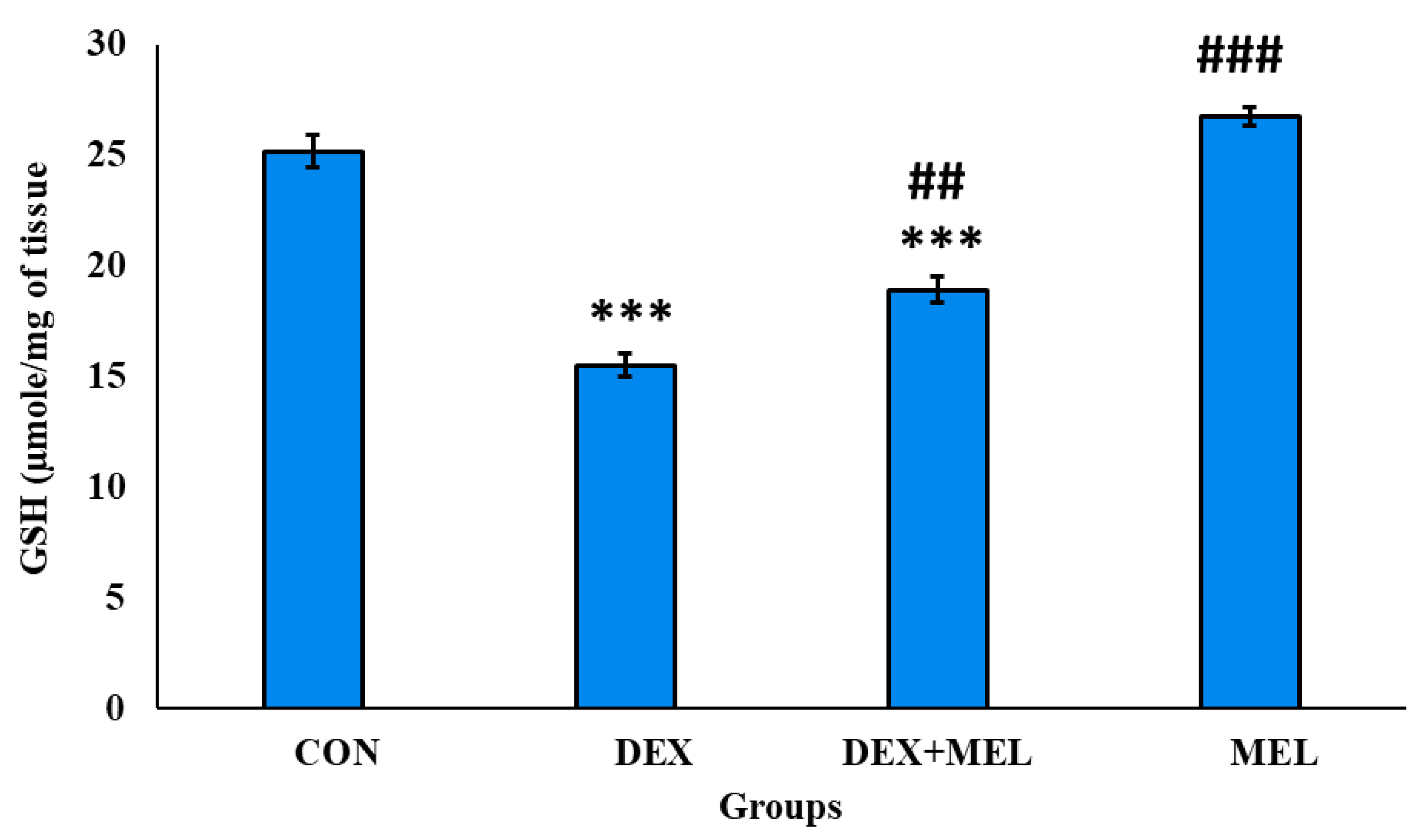
Figure 19.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on glutathione reductase (GR) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 19.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on glutathione reductase (GR) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
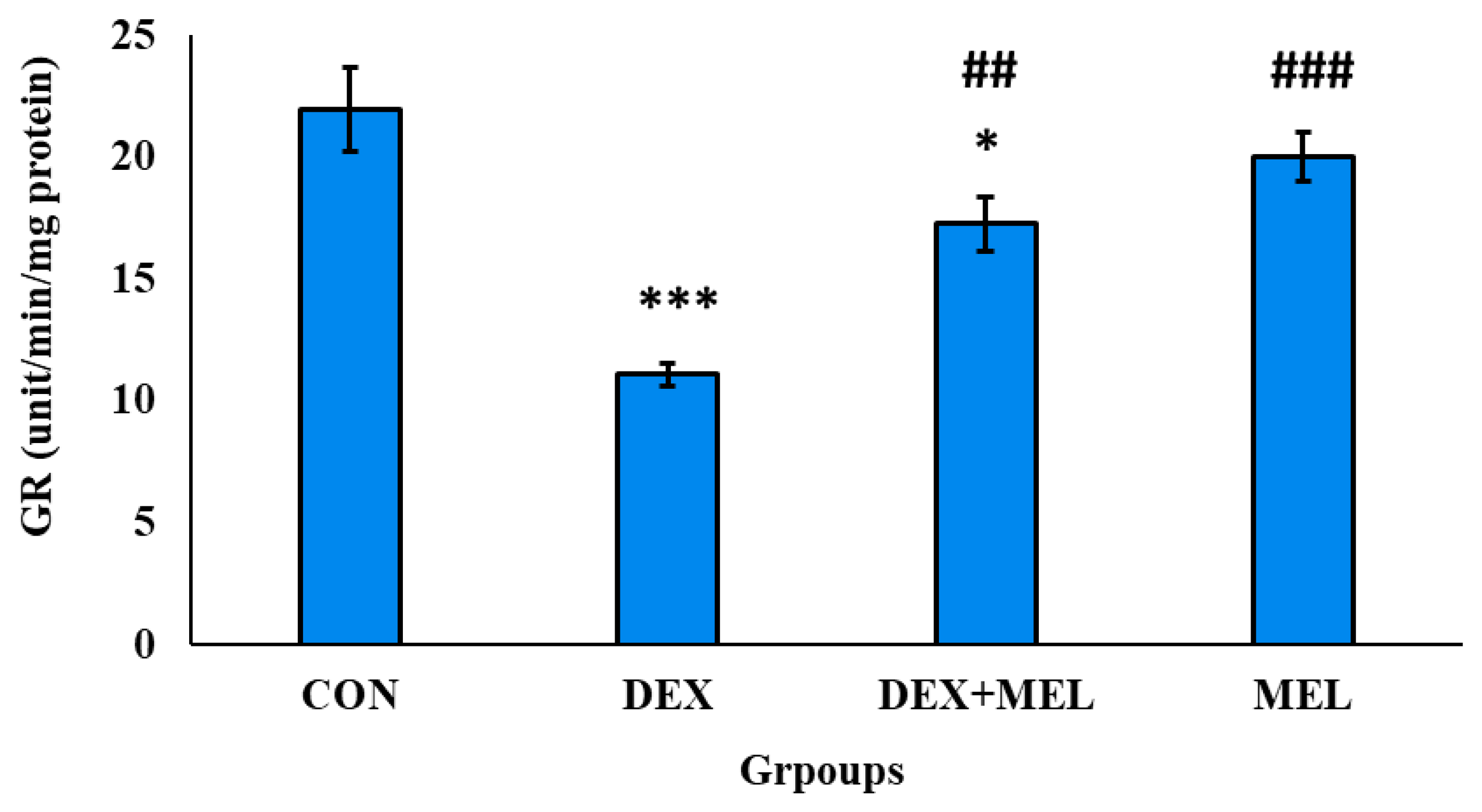
Figure 20.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on glutathione peroxidase (GPX) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 20.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on glutathione peroxidase (GPX) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
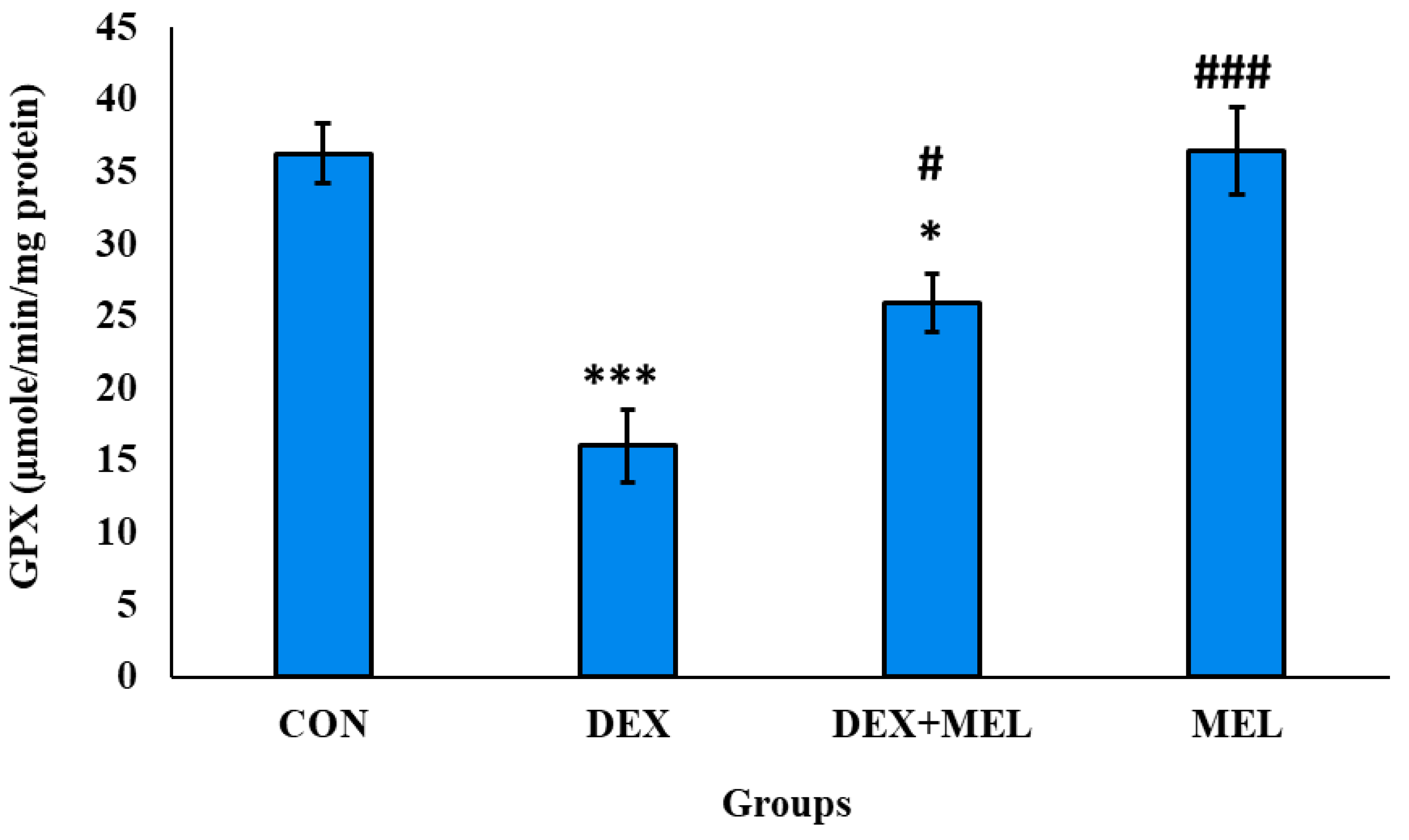
Figure 21.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on G6PDH of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
Figure 21.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on G6PDH of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.01, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL/MEL.
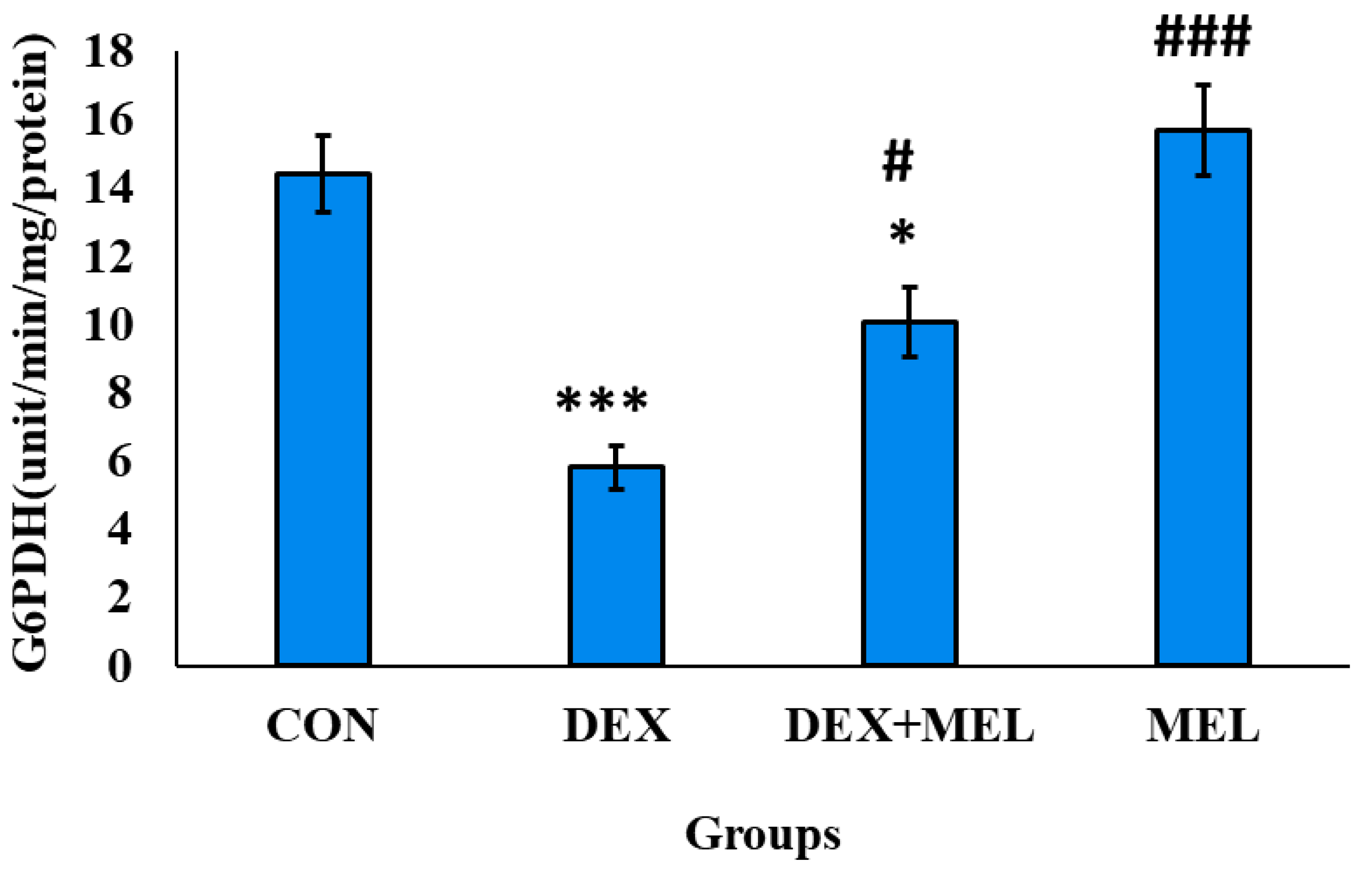
Figure 22.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on total glutathione-S-transferase (GST) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Figure 22.
Effect of melatonin (MEL) and dexamethasone (DEX) treatment on total glutathione-S-transferase (GST) of testes. The graph represents Mean + SE, N=6. ***/###p≤0.001, **/##p≤0.05, */#p≤0.05, where * CON vs DEX/ DEX+MEL and # DEX vs DEX+MEL.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



