
Submitted:
17 May 2024
Posted:
17 May 2024
You are already at the latest version
Abstract
Background: Carbapenemase-producing Enterobacter cloacae complex (ECC) are increasingly identified in hospital-acquired infections. They usually belong to four main multilocus sequence types (STs) named ST114, ST93, ST90, and ST78. Instead, ST182 has been sporadically reported, and recently outbreaks of blaNDM-producing ST182 strains have emerged. Herein, we aimed to investigate the presence of ST182 and explore its evolution and modes of blaNDM acquisition. Methods: A phylogenetic analysis of 646 MLST STs identified among 4,685 E. hormaechei WGS assemblies deposited in public repositories was performed, as well as an in silico comparative and phylogenomic analyses for 55 WGS assemblies of ST182. blaNDM-harboring contigs were also compared to published plasmid sequences. Results: ST182 E. hormaechei strains were recovered from patients in five continents during 2011-2021. They were divided into three major genomic clusters, comprising a separate clonal complex with six other STs. In 30 out of 55 ST182 WGS assemblies, blaNDM-harboring structures were identified similar to plasmids predominant in Gram-negative bacteria, harboring resistance genes to multiple antibiotic classes and virulence genes. No associations between the genomic clusters and the country/continent of isolation, the presence and the plasmid types of the blaNDM-harboring contigs were observed. Conclusions: Our findings show that ST182 E. hormaechei strains were identified the past decade worldwide; 54.5% of them carried diverse blaNDM genetic structures, suggesting recent acquisition of the blaNDM alleles. Thus, blaNDM-producing ST182 is an emerging multidrug-resistant and virulent lineage in ECC that requires close monitoring.
Keywords:
Enterobacter cloacae complex
; NDM carbapenemase
; MLST
; WGS
1. Introduction
Enterobacter cloacae complex (ECC) species are often recognised as the causative agents of hospital-acquired infections, such as pneumonia, urinary tract and soft-tissue infections, septicaemia, and meningitis [1]. Among them, E. cloacae and E. hormaechei are the most frequently identified in clinical specimens from hospitalized patients [1,2]. Enterobacter species are considered members of the ESKAPE (Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa and Enterobacter species) group of pathogens, exhibiting resistance to most common antibacterial treatments [3]. Multidrug-resistant ECC isolates are often reported worldwide [1, 3-7]. More specifically, ECC isolates are intrinsically resistant to first- and second-generation cephalosporins due to the presence of chromosomally-encoded class C β-lactamases, whereas resistance to multiple antibiotic classes has been associated mostly with overexpression of efflux pumps and the acquisition via mobile gene elements (MGEs) of resistance genes from other species; e.g. extended spectrum β-lactamase (ESBL), carbapenemase, aminoglycoside and quinolone resistance genes.
Carbapenemase-producing ECC strains are now reported in all WHO health regions [1,3,4,5,6,7] and may contain class A and class D carbapenemases, which are serine carbapenemases, and class B metallo-β-lactamases (MBLs) [1,4]. In particular, MDR blaNDM-producing ECC have recently emerged causing nosocomial outbreaks in several countries [7]. NDM is a MBL type carbapenemase, which is able to hydrolyse a wide range of β-lactams, including carbapenems, but not monobactams [5]. Furthermore, NDM is not inactivated by most of the recently developed β-lactamase inhibitors. Among carbapenemase-producing ECC, the most common lineage ECC ST114 is globally distributed and associated not only with NDM-type carbapenemases, but also with several other carbapenemases, such as VIM-1 MBL, class A KPC-2, and class D OXA-48 [1,3,4,5,6,7]. ST78 and ST171 have also been reported as two emerging lineages of carbapenem-resistant ECC, but strains of these lineages usually produce KPC rather than NDM carbapenemases [4]. Nonetheless, these STs comprise only a minority of STs found in ECC strains [4,5,6]. ECC has a very diverse clonal population structure, and there are neither well-defined international clones nor obvious associations with NDM-positive ECC strains.
Instead, ST182 ECC isolates have been sporadically reported worldwide from clinical specimens [4,5,6,7.8,9]. In Europe, ST182 was detected for the first time in a blaNDM-4-producing isolate from the Czech Republic in 2012 [10]. Later on, in 2016, ST182 has caused an outbreak of blaNDM-4-producing ECC in the same country [11]. Additionally, a recent study of multidrug-resistant ECC isolates from Lebanon has shown that ST182 was the second most frequent ST accounting for 10.4% of the ECC isolates [12]. Lately, the largest European dissemination of ECC NDM-producers has been reported in Greece and the outbreak was caused by a ST182 clonal strain [13]. Whole-genome taxonomic analysis of two blaNDM-1-producing strains recovered during the outbreak (EC-ML559 of MLST ST182 and EC-ML621 of ST2143, a single locus variant of ST182) revealed that both strains were assigned as E. hormaechei [14]. In silico prediction of components of the bacterial cell surface and genomic islands showed the presence of various virulence factors and resistance genes to several antimicrobial classes, as well as differences in the plasmids carrying β-lactamase genes [14].
In the present study, we aimed to investigate the presence of WGS assemblies of ST182 E. hormaechei in public databases and explore their characteristics, geographic distribution and evolution. For this purpose, we have compared in silico the WGS assemblies of ECC isolates of ST182, including plasmid types, and antimicrobial resistance and virulence genes. Furthermore, the blaNDM-harbouring contigs of the WGS assemblies of the isolates were compared with published plasmid sequences, so as to explore the plausible modes of acquiring blaNDM alleles.
2. Results
2.1. Bacterial strains, whole genome sequences and phylogenetic analysis of ECC isolates
The WGS assemblies retrieved from public databases have been obtained from E. hormaecheiisolates(n=4,685),which belonged to 646 MLST STs. In this dataset,the most prevalent STs were ST171 (n=396), ST93 (n=244), ST78 (n=220) and ST114 (n=208), whereas ST182 (n=55) ranked at position 16 (Figure 1a). By implementing the goeBURST algorithm and PHYLOViz analysis based on the MLST allelic profiles, the possible phylogenetic relationships between STs were obtained. Of the 646 MLST STs, 400MLST STs (3,953 isolates) were clustered into73 CCs, whereas the remaining 246 STs (732 isolates) were singletons (i.e., each group was comprised of one ST) [data not shown]. ST182 comprised a separate lineage in the phylogenetic tree, being in the same CC with ST98, ST710, ST1611, ST1752, ST2143 and ST2608 (Figure 1b).
Among the nucleotide sequences of the 55 ST182 E. hormaechei WGS assemblies, there were 4,554 SNPs, and the overall mean distance was 0.1013 (Supplemental Figure S1). Phylogenomic analysis of the WGS assemblieshas revealed that the strains were distributed into three genomic clusters (sublineages); cluster A (n=37), cluster B (n=10) and cluster C (n=8) [Figure 2; Figures S1 and S2; Table 1; Table S1].The 55 ST182 WGS assemblieswere collected mainly from Asia (n=17), Europe (n=17), North America (n=14), but also from Africa (n=4) and South America (n=3),while 30 of them carriedblaNDM genes (Table 1; Table S1). The first WGS assemblies of MLST ST182 strains with no blaNDM genes were identified inthe United Kingdom collected in 2002 and 2006, which belonged to cluster B. No blaNDM genes were identified in nine out of ten cluster B isolates, whereas one isolate collected from India carried blaNDM-1. Thereafter, both blaNDM carriers and strains with no blaNDM genes were recovered annually from 2011 and onwards (Figure S3).
2.2. In silico identification of plasmids, antimicrobial resistance and virulence genes of the ECC NDM-producing isolates
The predictions for the presence of plasmids, antimicrobial resistance and virulence genes of the 30 blaNDM-carrying WGS assemblies of ST182 are shown in Supplemental Table S3. All isolates were predicted to carry plasmids predominant in Gram-negative antibiotic resistant strains, belonging to several incompatibility groups, such as IncX3, IncFII/IncFIB, IncHI2, IncHI2A, IncL, IncM,IncN, IncN3, IncR,IncX5, Col440I, Col440II replicon typeplasmids. No associations were observed between the country/continent of isolation, or the presence of the blaNDM plasmid types and the genetic clusters (Figure 3, Figure S2, Table 2).
All isolates were predicted to harbour antimicrobial resistance genes to multiple antibiotic classes and virulence genes. In more detail, four blaNDM variants were identified; blaNDM-1 (n=24), blaNDM-4 (n=2), blaNDM-5 (n=2) and blaNDM-7 (n=2). Besides blaNDM, all carried the chromosomal blaACT-16and several acquired β-lactamase genes, including blaTEM-1, blaTEM-104,blaOXA-1, blaOXA-9,blaOXA-10,blaOXA-48, blaCTX-M-3, blaCTX-M-9, blaCTX-M-14, blaCTX-M-15, blaDHA-1, blaKPC-2, blaSHV-12, blaLAP-2blaSFO-1 andblaGES-5.In addition to the β-lactamase genes, acquired genes conferring resistance to various antimicrobial classes were identified, including: aminoglycosides [aac(3)-IIa, aac(3)-Id, aac(3)-IId, aac(6′)-Ib,aac(6′)-Ib3, aac(6′)-IIc,aac(6′)-Ib-cr, aadA1, aadA2, aadA2b, aadA16, aph(3′)-Ia, aph(3′′)-Ib, aph(6)-Id, ant(2′′)-Ia, armA, rmtB, rmtC], quinolones (qnrA1, qnrB1, qnrB4, qnrB6, qnrB19, qnrS1, OqxA, OqxB), chloramphenicol (catA2, catB3), trimethoprim (dfrA12, dfrA14, dfrA19, dfrA27), sulphonamides (sul1, sul2), macrolide–lincosamide–streptogramin B (MLS) [mph(A), mph(E), ere(A), msr(E)], tetracyclines [tet(A,), tet(D], polymyxins (mcr-9), fosfomycin (fosA) and rifampicin (ARR-3). Moreover, resistance genes for quaternary ammonium compounds (qacE) and formaldehyde (formA) were also predicted.
Of the 30 blaNDM-harbouring strains, 23 strains were predicted to harbour plasmidic sequences similar tothe 139kb E. cloacae subsp. cloacae ATCC13047 plasmid pECL_A, which carries several virulence factor genes, such as two clusters of Type IV secretion system (T4SS) genes, associated with pathogenesis in plants and mammalian bacterial pathogens, and also multiple heavy metal resistance operons for copper, tellurium and mercury that are not conserved with other Enterobacter species, but share notably high homology to Cronobacter sakazakii, K.pneumoniae and E.coli [26]. Additionally, the virulence genes nlpI, terC, traT and mrkA, shiB, kpsM_K11 and astA were identified. All WGS assemblies possessed the virulence gene nlpI, encoding the lipoprotein NlpI, which is involved in the cell division, virulence, and bacterial interaction with eukaryotic host cells [27]. All but one isolate (strain PEER1096 from India) possessed terC, which is one of the key proteins of the tellurite resistance gene operon (ter) involved in tellurite resistance phage inhibition, colicine resistance, and pathogenicity [28].Four strains also harboured the traT gene encoding the TraT protein, a cell-surface-exposed, outer membrane lipoprotein associated with resistance to the bactericidal activities of serum and prevention of self-mating of cells carrying identical or closely related conjugative plasmids [29]. The mrkA adhesion gene, which has been associated with biofilm formation in carbapenemase-producing K. pneumoniae [30], was present in three strains. The astA and kpsM_K11 genes, which have been previously associated with virulence in pathogenic E. coli strains [31,32], were also predicted in two (BiosamplesSAMN25161196 and SAMN25161198 from the United States) and one (Biosample SAMN15904743 from Hong Kong) blaNDM-1- carriers, respectively. Finally, the shiB gene, which has been found previously in the pathogenicity island SHI-2 (Shigellaisland 2) of Shigella flexneri [33], was predicted in a blaNDM-1 carrier (Biosample SAMEA8581547 from Pakistan).
2.3. Genetic background of blaNDM and plasmid analysis
In three ST182 strains (M515, MY196, and AZ 664), a blaNDM-1 gene was located on two different contigs of the WGS assemblies. BlastN comparisons of the blaNDM-harbouring contigs revealed the presence of genetic structures showing 100% identities with regions of sixdifferent plasmid types; an IncX3 (pNDM-HN380), three different IncFII (pKOX_NDM-1, pGUE-NDM, pKPX-1), an IncA/C (pM214_AC2) and an IncN2 (pJN24NDM) (Table 2; Figure 3, Figures S3 and S4) [34,35,36,37]. The most prevalent plasmidic sequences were found in 16 strains and distributed into clusters A and C, which were similar to the IncX3 replicon type K. pneumoniae pNDM-HN380 from China [34] (Table 2). The blaNDM-4-encoding plasmid pEncl-922cz of the incompatibility group IncX3 fromtheCzech Republic has been published previously [10]. pEncl-922cz was identical to the respective sequences of blaNDM-4-encoding plasmids recovered in the same hospital during 2016 (such as strain Encl-44578 included in the present study) [10], but differed by the insertion of a Tn3-like transposon downstream of the topB gene compared with pNDM-HN380 and other IncX3 replicon types, such as the blaNDM-5-producing K. pneumoniae pNDM-MGR194 from India [38].
The blaNDM-carrying pKOX_NDM-1 strains (n=8) were distributed into cluster A (Table 2). The genome sequence and the blaNDM-1-harbouring plasmid of strain P1 from Iran has been published previously [8]. blaNDM-1 was carried on a pKOX_NDM1-like plasmid, which is a non-transferable IncFIIYtype plasmid first reported in Taiwan [34, 39],and later in other E. cloacae complex, K. pneumoniae, K. oxytoca and Serratiamarcescens isolates recovered in Romania [40].Different evolutionary events, including single nucleotide level change, indels and recombination events were observed among pKOX_NDM-1-like plasmids. The blaNDM-carrying pGUE-NDM strains (n=3) were distributed into clusters A and B (Table 2). The IncFII-type plasmid pGUE-NDM (IncFII) was first described in an E. coli MLST ST131 isolate from France [41]and plasmids from other Enterobacterales [42]. The EC-ML-559 strain from Greece (cluster C) carried a blaNDM-1-harbouring structure found in Klebsiellapneumoniae subsp. pneumoniae strain KPX plasmid pKPX-1from Taiwan [14,43] and Enterobacter hormaechei subsp. xiangfangensis strain ST114 plasmid pLAU_ENM30_NDM1 from Lebanon [12].Finally, blaNDM-1-harboring contigs of strain RIVM_C015180 from the Netherlands and strain E472 from Singapore, were similar to plasmids pM214_AC2 (IncA/C) [36] and pJN24NDM (IncN2) [37] types, respectively, which have been previously described in blaNDM-harbouring plasmids of E. coli (Figure S4).
The conjugative regions (oriT, relaxase gene, T4CP gene and T4SS gene cluster) of the self-transmissible MGEs were characterised for the ST182strains that have caused the outbreaks in the Czech Republic and Greece. A conjugative plasmid must possess all the conjugative regions, whereas a transmissible plasmid must possess at a minimum an oriT and usually a relaxase, but this can be provided in trans [24]. In the blaNDM-1-harbouring strain EC-ML559 from Greece (Biosample SAMN33955250), all four conjugative regions were predicted; the oriT(region: 13705-13786), the relaxase gene (region: 14142-16070), the T4CP gene (region: 19453-21645), and T4SS gene cluster (region: 19453-44454). In plasmid pEncl-922cz from the Czech Republic (Biosample SAMN08436979), no oriT region was predicted, but a relaxase gene (region 34117-35277), the gene encoding type IV coupling protein (T4CP, region: 21075-22910), the gene cluster for bacterial type IV secretion system (T4SS, region: 20284-33022) were predicted.
3. Discussion
The ECC mainly comprises of six Enterobacter species (E. asburiae, E. cloacae, E. hormaechei, E. kobei, E. ludwigii and E. nimipressuralis); however, the accurate identification of species/subspecies of the genus Enterobacterby routine identification techniques, as well as 16S rRNA and housekeeping genes has often been inconsistent [1, 26].Thus, reclassification of species and subspecies of the genus Enterobacter by phylogenetic studies based on whole genome DNA-DNA hybridizations and sequencing is challenging and ongoing [1,26,44]. A global study of carbapenemase-producing ECC isolates collected during 2008-2014 revealed that the most common identified carbapenemase was VIM MB, followed by NDM MBL, class A KPC, class D OXA-48, and IMP MBL [7]. As observed with other carbapenemase-producing ECC, blaNDM-producing ECC were also found to mainly belong to four STs, named ST114, ST93, ST90, and ST78.In the present assay, we performed phylogenetic analysis for 646 STs identified among all 4,685 E. hormaechei WGS assemblies deposited in public databases, which revealed that ST182 is an emerging lineage in ECC. ECC ST182 strains were predicted in silicoto harbour plasmids commonly found among multidrug-resistant bacteria, which have acquired antimicrobial resistance and virulence genes, whereas different blaNDM-harboring plasmid types among ST182 ECC were distributed in all sublineages.
NDM-1 was first reported in a K.pneumoniae strain recovered from a urinary culture on 9 January 2008 of a Swedish patient with a history of hospitalization in New Delhi, India[45].Thereafter, it has spread rapidly and 24 NDM variants have been identified in various species of Gram-negative bacteria, such as Enterobacterales, Acinetobacter and Pseudomonas from clinical specimens worldwide [1,5]. It has been suggested that global travel has facilitated the rapid spread of NDM from its initial emergence in India to all continents, since importation of NDM producers has been associated with patients having a history of travel [3-7, 12, 45]. A recent study from Israel has also shown that most of the blaNDM-harboring Enterobacterales possessed nine different MGE modules, variably distributed across species and hospitals [45]. In another study, the role of mobile genetic elements in the global dissemination of the blaNDM was investigated and it was estimated that blaNDM emerged on a Tn125 transposon before 1985, but only reached global prevalence around a decade after its first recorded observation in 2008 [47]. The global dissemination of the blaNDM gene was primarily driven by successive between-plasmid transposon jumps. In K. pneumoniae, different trajectories have been shown for the spread of carbapenemase genes, including via one plasmid/multiple lineages (blaOXA-48-like), multiple plasmids/multiple lineages (blaVIM, blaNDM), and multiple plasmids/one lineage (blaKPC) [48]. The findings of the current study revealed that E. hormeacei ST182 WGS assemblies deposited in public databases were collected from 2002 to 2021, and during this period we have identified both WGS assemblies carrying blaNDM and WGS assemblies with no blaNDM genes. No clustering over time was observed for the two groups of strains or the different blaNDM plasmid types, suggesting that strains without blaNDM genes have been distributed globally, and then blaNMD genes were diffused in different genomic clusters.
In a previous study, a common blaNDM genetic structure on plasmid pNDM-U.S. was identified in 14 different ECC clones obtained from six countries spanning four continents[5]. Moreover, in some cases certain mobile genetic elements with carbapenemase genes were found associated with the geographic distribution of clades, clones and species, suggesting that these mobile elements have the ability to move between clones and clades of ECC on a global scale. Several surveys have shown that the blaNDM genes were distributed across a large number of STs in the most prevalent species of Enterobacterales (E. coli, K. pneumoniae and Enterobacter spp.), with no predominant lineages, suggesting that there are no obvious high-risk clones of blaNDM-producing strains [3,4,6]. In the current study, blaNDM-harbouring contigs showed similarities with six different plasmid types. The most prevalent IncX3 replicon type pNDM-HN380-like structures were found in four continents (Asia, Europe, North America and Africa) and diffused into genomic clusters (sublineages) A and C. Similarly, the three different IncFII-type genetic structures were also distributed into different continents and/or genomic clusters; the pKOX_NDM-1 in Asia, Europe, North and South America (cluster A), the pGUE-NDM-like structures in Asia (clusters A and B), and the pKPX-1 in Europe (cluster C). Thus, different blaNDM-carrying plasmids were diffused among strains of the same genomic cluster (sublineage), and on the other hand, the same blaNDM-carrying plasmid could be found in strains belonging to different sublineages of ST182. Therefore, no associations were observed between the genetic clusters and the country/continent of isolation, the presence of the blaNDM alleles and the plasmid types.
A limitation of the present study is that plasmid reconstruction was not performed due to short sequencing reads. It should be noted that plasmids are difficult to reconstruct from WGS data. NGS assembly programs tend to return short contigs of heterogeneous origins. On the other hand, alignment-based tools tend to miss diverged plasmids, while learning-based tools often have lower precision. In some studies, the combination of short and long sequencing read WGS strategies has been used [47,48]. Another limitation of the present descriptive survey is that it included only sequenced ECC isolates in the NCBI and PubMLST public repositories, which are deposited randomly by users and there may be a bias towards multidrug-resistant strains; thus, they do not represent the global molecular epidemiology of blaNDM-producing ECC isolates. Further epidemiological and molecular surveillance studies at a global scale would define the prevalence of the ST182 lineage in ECC.
4. Materials and Methods
4.1. Bacterial isolates, genome sequences and phylogenetic analysis
A total of 4,685 WGS assemblies of E. hormaechei isolates with available MLST profiles were analysed. We have retrieved WGS assemblies from the Pathogenwatch database [15] andthe PubMLST Enterobacter cloacae database (available at: https://pubmlst.org/organisms/enterobacter-cloacae; last accessed 15/1/2024) [16], which include WGS assemblies from public repositories, such as the European Nucleotide Archive (ENA) and NCBI. Additionally, we have searched the PubMLST database for the presence of the alleles of ST182 (dnaA-49, fusA-20, gyrB-19, leuS-44, pyrG-90, rplB-24, rpoB-32) in other MLST profiles, which were found in 62, 174, 89, 220, 9, 27 and 125 MLST profiles (STs), respectively. Since alleles pyrG-90 and rplB-24 are present in the fewer MLST profiles (9 and 27 profiles, respectively) compared with the other MLST alleles of ST182, we have also searched the NCBI database for these alleles (pyrG-90 and rplB-24) and the MLST 2.0 tool (available at: https://cge.food.dtu.dk/services/MLST/,Center for Genomic Epidemiology, Technical University of Denmark) was used to define the MLST STs, so as to retrieve any additional WGS assemblies of ST182.In the final dataset, a total of 55 WGS assemblies of ST182 were included. The genetic relationships and groups of STS were formed by linking all STs that were single locus variants (SLVs), known as Clonal Complexes (CCs), by using the goeBURST and the PHYLOViZ software (available at http://www.phyloviz.net/) [17].
The phylogenomic analysis of the WGS assemblies of ST182 strains was performedusing the Reference sequence Alignment based Phylogeny (REALPHY) tool (available at: https://realphy.unibas.ch/realphy/) [18].The WGS assembly of the type strain E.cloacae subsp. cloacae NCTC9394 (GenBank accession no.: FP929040.1) was used as a reference sequence. Of note, the taxonomic classification of the type strain NCTC9394 was updated on 04/08/2020 from Enterobacter cloacae to Enterobacter hormaechei (https://www.ncbi.nlm.nih.gov/nuccore/NC_021046.1?report=genbank). Single- nucleotide polymorphisms (SNPs) were extracted using the Galaxy Server (available at https://usegalaxy.org/) from the aligned set of orthologous sites obtained by REALPHY [18] and phylogenetic analysis was performedusing theMEGA version 11 software [19]. Visualization of the timeline of the isolation, the geographical distribution and the phylogenetic tree of the 55 ST182 ECC strains were performed by using the Microreact software [20].
4.2. Identification of MGEs, antimicrobial resistance genes and virulence factors, and plasmid analysis
BLASTN (available at: https://blast.ncbi.nlm.nih.gov/Blast.cgi), the KmerResistance 2.2 tooland the Mobile Element Finder tool (available at: https://cge.food.dtu.dk/services/; last accessed 25/02/2024) were used to query the sequence assemblies for identification of the blaNDM-harbouring contigs, MGEs, plasmids and their relation to antimicrobial resistance genes and virulence factors on the genomes[21, 22].k-mer alignment examines the co-occurrence of k-mers between the WGS data and a database of resistance genes, and scales well for large redundant databases[21].The GC content of the WGS assemblies was calculated by using the GC-profile (available at: http://tubic.tju.edu.cn/GC-Profile/) [23],and the GCdraw (available at: http://www.endmemo.com/bio/gcdraw.php) online tools. The blaNDM-harbouring contigs of the isolates were analysed with the oriTfinder tool [24], so as to explore the presence of conjugative regions of the self-transmissible MGEs: the origin of transfer site (oriT), the relaxase gene, the gene encoding type IV coupling protein (T4CP) and the gene cluster for bacterial type IV secretion system (T4SS).BlastN comparisons of the blaNDM-harbouring contigs with plasmid sequences retrieved from the NCBI was performed by using the BLAST Ring Image Generator (BRIG) version 0.95 software (available at: https://brig.sourceforge.net/) [25].
5. Conclusions
In the present survey, we have shown via phylogenetic analysis that the multidrug-resistant ST182 is an emerging lineage in ECC, representing a distinct clonal complex among blaNDM-producing ECC. ST182 strains retrieved from public databases were distributed into three genomic clusters (sublineages), which contained strains recovered from five different continents and both strains that harbour and did not harbour blaNDM genes. Different plasmid types have been spread among the genetic clusters of ECC ST182, whereas no associations were observed between the genetic clusters and plasmid types. The diversity of the blaNDM-harbouring genetic structures identified among ECC ST182 isolates denotes different routes of blaNDM acquisition into the ECC ST182 clusters worldwide. These findings suggest that ST182 strains without blaNDM genes were emerged and spread initially, and acquired later on the blaNDM genetic structures via horizontal gene transfer from other bacteria in the recent past. Furthermore, ECC ST182 has already caused outbreaks in the Czech Republic and Greece and, therefore it has the potential of causing outbreaks worldwide. Vigilance and continuous molecular-typing based surveillance seems mandatory among ECC strains in order to understand the further expansion of the emerged ST182 ECC and restrain its dissemination.
Supplementary Materials
The following supporting information can be downloaded at: www.mdpi.com/xxx/s1, Figure S1: Evolutionary relationships of ST182 E.hormaechei WGS assemblies.; Figure S2: Genomic clusters, blaNDM variants, country, continent and year of isolation of 55 ST182 E. hormaechei WGS assemblies; Figure S3: Yearly distribution of ST182 E. hormaechei (a) blaNDM variants and (b) plasmid types of blaNDM-carrying WGS assemblies during 2011-2021; Figure S4: BlastN comparisons of the nucleotide sequences of blaNDM-harbouring contigs; Table S1: Characteristics of 55 ST182 E. hormaechei WGS assemblies; Table S2: Characteristics of 30 ST182 E. hormaecheiblaNDM-carryingWGS assemblies; Table S3: In silico prediction of contig Inc types, antimicrobial resistance and virulence genes in 30 blaNDM-harbouring ST182 E. hormaechei WGS assemblies.
Author Contributions
Conceptualization, A.M and A.T.; methodology and software E.F. and A.M; formal analysis and investigation, writing, A.M; original draft preparation, A.M.; writing—review and editing, supervision, A.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Davin-Regli A, Lavigne JP, Pagès JM. Enterobacter spp.: Update on taxonomy, clinical aspects, and emerging antimicrobial resistance. ClinMicrobiol Rev. 2019; 32:e00002-19. [CrossRef]
- Paauw A, Caspers MPM, Schuren FHJ, Leverstein-van Hall MA, Delétoile A, Montijn RC, et al. Genomic diversity within the Enterobacter cloacae complex. PLoS One 2008; 3: e3018. [CrossRef]
- De Oliveira DMP, Forde BM, Kidd TJ, Harris PNA, Schembri MA, Beatson SA, Paterson, DL, Walker MJ. Antimicrobial resistance in ESKAPE pathogens. Clin Microbiol Rev 2020; 33: e00181-19. [CrossRef]
- Annavajhala MK, Gomez-Simmonds A, Uhlemann AC. Multidrug-resistant Enterobacter cloacae Complex emerging as a global, diversifying threat. Front Microbiol. 2019; 10: 44. [CrossRef]
- Wu W, Feng Y, Tang G, Qiao F, McNally A, Zong Z. NDM Metallo-β-lactamases and their bacterial producers in health care settings. ClinMicrobiol Rev 2019; 32: e00115-18. [CrossRef]
- Izdebski R, Baraniak A, Herda M, Fiett J, Bonten MJ, Carmeli Y, Goossens H, Hryniewicz W, Brun-Buisson C, Gniadkowski M; MOSAR WP2, WP3 and WP5 Study Groups. MLST reveals potentially high-risk international clones of Enterobacter cloacae. J AntimicrobChemother 2015; 70:48-56. [CrossRef]
- Peirano G, Matsumura Y, Adams MD, Bradford P, Motyl M, Chen L, et al. Genomic epidemiology of global carbapenemase-producing Enterobacter spp., 2008–2014. Emerg Infect Dis 2018; 24:1010-19. [CrossRef]
- Bolourchi, N., Giske, C. G., Nematzadeh, S., Mirzaie, A., Abhari, S. S., Solgi, H., &Badmasti, F. Comparative resistome and virulome analysis of clinical NDM-1–producing carbapenem-resistant Enterobacter cloacae complex. J Glob Antimicrob Resist 2022; 28: 254-63. [CrossRef]
- Sugawara Y, Akeda Y, Hagiya H, Sakamoto N, Takeuchi D, Shanmugakani RK, Motooka D, Nishi I, Zin KN, Aye MM, MyintT,Tomono K, Hamada S. Spreading patterns of NDM-producing Enterobacteriaceae in clinical and environmental settings in Yangon, Myanmar. Antimicrob Agents Chemother. 2019; 63: e01924-18. [CrossRef]
- Papagiannitsis, CC, Studentova V, Chudackova E. et al. Identification of a New Delhi metallo-β-lactamase-4 (NDM-4)-producing Enterobacter cloacae from a Czech patient previously hospitalized in Sri Lanka. Folia Microbiol 2013; 58: 547–9. [CrossRef]
- Paskova V, Medvecky M, Skalova A, Chudejova K, Bitar I, Jakubu V, Bergerova T, Zemlickova H, Papagiannitsis CC, Hrabak J. Characterization of NDM-encoding plasmids from Enterobacteriaceae recovered from Czech hospitals. Front Microbiol. 2018; 9:1549. [CrossRef]
- Merhi G, Amayri S, Bitar I, Araj GF, Tokajian S. Whole genome-based characterization of multidrug-resistant Enterobacterand Klebsiellaaerogenes isolates from Lebanon. MicrobiolSpectr 2023; 1: e0291722. [CrossRef]
- Gartzonika K, Politi L, Mavroidi A, Tsantes AG, Spanakis N, Priavali E, Vrioni G, Tsakris A. High prevalence of clonally related ST182 NDM-1-producing Enterobacter cloacae complex clinical isolates in Greece. Int J Antimicrob Agents 2023; 62: 106837. [CrossRef]
- Mavroidi, A., Gartzonika, K., Spanakis, N., Froukala, E., Kittas, C., Vrioni, G., Tsakris, A. Comprehensive analysis of virulence determinants and genomic islands of blaNDM-1-producing Enterobacterhormaechei clinical isolates from Greece. Antibiotics 2023; 12: 1549. [CrossRef]
- Argimón S, David S, Underwood A, Abrudan M, Wheeler NE, Kekre M, Abudahab K, Yeats CA, Goater R, Taylor B, Harste H, Muddyman D, Feil EJ, Brisse S, Holt K, Donado-Godoy P, Ravikumar KL, Okeke IN, Carlos C, Aanensen DM; NIHR Global Health Research Unit on Genomic Surveillance of Antimicrobial Resistance. 2021. Rapid genomic characterization and global surveillance of Klebsiellausing Pathogenwatch. Clin Infect Dis. 2021; 73 (Suppl_4): S325-S335. [CrossRef] [PubMed]
- Jolley KA, Bray JE, Maiden MCJ. Open-access bacterial population genomics: BIGSdb software, the PubMLST.org website and their applications. WellcomeOpenRes 2018; 3:124. [CrossRef]
- Nascimento M, Sousa A, Ramirez M, Francisco AP, Carriço JA, Vaz C, PHYLOViZ 2.0: providing scalable data integration and visualization for multiple phylogenetic inference methods.Bioinformatics 2017; 33: 128–9. [CrossRef]
- Bertels F, Silander OK, Pachkov M, Rainey PB, van Nimwegen E. Automated reconstruction of whole genome phylogenies from short sequence reads. MolBiol Evol 2014; 31:1077-88.
- Tamura K., Stecher G., and Kumar S. MEGA 11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol 2021; 38:3022-27. [CrossRef]
- Argimón S, Abudahab K, Goater RJE, Fedosejev A, Bhai J, Glasner C, Feil EJ, Holden MTG, Yeats CA, Grundmann H. Spratt BG, Aanensen DM. Microreact: Visualizing and sharing data for genomic epidemiology and phylogeography. MicrobGenom 2016; 2: e000093. [CrossRef]
- Clausen PT, Zankari E, Aarestrup FM, Lund O. Benchmarking of methods for identification of antimicrobial resistance genes in bacterial whole genome data. J AntimicrobChemother 2016;71:2484-8. [CrossRef]
- Johansson MHK, Bortolaia V, Tansirichaiya S, Aarestrup FM, Roberts AP, Petersen TN. Detection of mobile genetic elements associated with antibiotic resistance in Salmonella enterica using a newly developed web tool: MobileElementFinder. J AntimicrobChemother 2021;76:101-9. [CrossRef]
- Gao F, Zhang CT. GC-Profile: a web-based tool for visualizing and analyzing the variation of GC content in genomic sequences. Nucleic Acids Res 2006; 34 (Web Server issue): W686-91. [CrossRef]
- Li X, Xie Y, Liu M, Tai C, Sun J, Zixin Deng, Ou H-Y. oriTfinder: a web-based tool for the identification of origin of transfers in DNA sequences of bacterial mobile genetic elements, Nucleic Acids Research 2018; 46: W229–34. [CrossRef]
- Alikhan NF, Petty NK, Ben Zakour NL, Beatson SA. BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons, BMC Genomics 2011, 12: 402. [CrossRef]
- Liu W-Y, Wong C-F, Chung KM-K, Jiang J-W, Leung FC-C. Comparative genome analysis of Enterobacter cloacae. PLoS One, 2013; 8: e74487. [CrossRef]
- Tao J, Sang Y, Teng Q, Ni J, Yang Y, Tsui SK, Yao YF. Heat shock proteins IbpA and IbpB are required for NlpI-participated cell division in Escherichia coli. Front Microbiol 2015; 6:51. [CrossRef]
- L. Turkovicova, R. Smidak, G. Jung, J. Turna, G. Lubec, J. Aradska, Proteomic analysis of the TerCinteractome: Novel links to tellurite resistance and pathogenicity, J Proteomics 2016; 136:167-73. [CrossRef]
- Sukupolvi S, O′Connor CD. TraT lipoprotein, a plasmid-specified mediator of interactions between gram-negative bacteria and their environment. MicrobiolRev 1990; 54:331-41. [CrossRef]
- Gual-de-Torrella A, Delgado-Valverde M, Pérez-Palacios P, Oteo-Iglesias J, Rojo-Molinero E, Macià MD, Oliver A, Pascual Á, Fernández-Cuenca F. Prevalence of the fimbrial operon mrkABCD, mrkA expression, biofilm formation and effect of biocides on biofilm formation in carbapenemase-producing Klebsiellapneumoniae isolates belonging or not belonging to high-risk clones. Int J Antimicrob Agents 2022; 60: 106663. [CrossRef]
- Wang J, Huang Y, Guan C, Li J, Yang H, Zhao G, Liu C, Ma J, Tang B. Characterization of an Escherichia coli isolate coharboring the virulence gene astA and tigecycline resistance gene tet(X4) from a dead piglet. Pathogens 2023; 12: 903.
- BingbingZong, Wugang Liu, Yanyan Zhang, Xiangru Wang, Huanchun Chen, Chen Tan, Effect of kpsM on the virulence of porcine extraintestinal pathogenic Escherichia coli, FEMS Microbiology Letters 2016; 363: fnw232.
- Moss JE, Cardozo TJ, Zychlinsky A, Groisman EA. The selC-associated SHI-2 pathogenicity island of Shigellaflexneri. MolMicrobiol 1999; 33; 74-83.
- Pak-Leung Ho, Zhen Li, Wai-U Lo, Yuk-Yam Cheung, Chi-Ho Lin, Pak-Chung Sham, Vincent Chi-Chung Cheng, Tak-Keung Ng, Tak-Lun Que & Kin-Hung Chow (2012). Identification and characterization of a novel incompatibility group X3 plasmid carrying blaNDM-1 in Enterobacteriaceae isolates with epidemiological links to multiple geographical areas in China. Emerg Microbes Infect 2012; 1: e39. [CrossRef]
- Wailan AM, Paterson DL, Kennedy K, Ingram PR, Bursle E, Sidjabat HE. Genomic characteristics of NDM-producing Enterobacteriaceae isolates in Australia and their blaNDM genetic contexts. Antimicrob Agents Chemother. 2015; 60: 136-41. [CrossRef]
- Sugawara Y, Akeda Y, Sakamoto N, Takeuchi D, Motooka D, Nakamura S, et al. Genetic characterization of blaNDM-harboring plasmids in carbapenem-resistant Escherichia coli from Myanmar. PLoS ONE 2017; 12: e0184720. [CrossRef]
- Hao Y, Shao C, Geng X, Bai Y, Jin Y, Lu Z. Genotypic and phenotypic characterization of clinical Escherichia coli Sequence Type 405 carrying IncN2 plasmid harboring blaNDM-1. Front Microbiol. 2019;10:788. [CrossRef]
- Krishnaraju M, Kamatchi C, Jha AK, Devasena N, Vennila R, Sumathi G, Vaidyanathan R. Complete sequencing of an IncX3 plasmid carrying blaNDM-5 allele reveals an early stage in the dissemination of the blaNDM gene. Indian J Med Microbiol 2015, 33: 30-8. [CrossRef]
- Huang TW, Wang JT, Lauderdale TL, Liao TL, Lai JF, Tan MC, Lin AC, Chen YT, Tsai SF, Chang SC. Complete sequences of two plasmids in a blaNDM-1-positive Klebsiellaoxytoca isolate from Taiwan. Antimicrob Agents Chemother 2013; 57: 4072-6. [CrossRef]
- Phan HTT, Stoesser N, Maciuca IE, Toma F, Szekely E, Flonta M, Hubbard ATM, Pankhurst L, Do T, Peto TEA, Walker AS, Crook DW, Timofte D. Illumina short-read and MinION long-read WGS to characterize the molecular epidemiology of an NDM-1 Serratiamarcescens outbreak in Romania. J AntimicrobChemother. 2018; 73: 672-679. [CrossRef]
- Bonnin RA, Poirel L, Carattoli A, Nordmann P. Characterization of an IncFII plasmid encoding NDM-1 from Escherichia coli ST131. PLoS One. 2012; 7: e34752. [CrossRef]
- Shin J, Baek JY, Cho SY, Huh HJ, Lee NY, Song JH, Chung DR, Ko KS. blaNDM-5-bearing IncFII-type plasmids of Klebsiellapneumoniae sequence type 147 transmitted by cross-border transfer of a patient. Antimicrob Agents Chemother. 2016; 60:1932-4. Retraction in: Antimicrob Agents Chemother 2023; 67: e0133022. [CrossRef]
- Huang TW, Chen TL, Chen YT, Lauderdale TL, Liao TL, Lee YT, Chen CP, Liu YM, Lin AC, Chang YH, Wu KM, Kirby R, Lai JF, Tan MC, Siu LK, Chang CM, Fung CP, Tsai SF. Copy number change of the NDM-1 sequence in a multidrug-resistantKlebsiellapneumoniae clinical isolate. PLoS One 2013; 8: e62774.
- Ciufo S, Kannan S, Sharma S, Badretdin A, Clark K, Turner S, Brover S, Schoch CL, Kimchi A, DiCuccio M. Using average nucleotide identity to improve taxonomic assignments in prokaryotic genomes at the NCBI. Int J SystEvolMicrobiol. 2018; 68:2386-2392. [CrossRef]
- Yong D, Toleman MA, Giske CG, Cho HS, Sundman K, Lee K, Walsh TR. Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob Agents Chemother. 2009; 53: 5046-54. [CrossRef]
- Adler A, Ghosh H, Gross A, Rechavi A, Lasnoy M, Assous MV, Geffen Y, Darawshe B, Wiener-Well Y, Grundmann H, Reuter S. Molecular features and transmission of NDM-producing Enterobacterales in Israeli hospitals. J Antimicrob Chemother. 2023; 78: 719-23. [CrossRef]
- Acman M, Wang R, van Dorp L, Shaw LP, Wang Q, Luhmann N, Yin Y, Sun S, Chen H, Wang H, Balloux F. Role of mobile genetic elements in the global dissemination of the carbapenem resistance gene blaNDM. Nat Commun 2022; 13:1131. [CrossRef]
- David S, Cohen V, Reuter S, Sheppard AE, Giani T, Parkhill J; European Survey of Carbapenemase-Producing Enterobacteriaceae (EuSCAPE) Working Group; ESCMID Study Group for Epidemiological Markers (ESGEM); Rossolini GM, Feil EJ, Grundmann H, Aanensen DM. Integrated chromosomal and plasmid sequence analyses reveal diverse modes of carbapenemase gene spread among Klebsiella pneumoniae. Proc Natl AcadSci U S A. 2020; 117:25043-25054. [CrossRef]
Figure 1.
(a) Phylogenetic relationships of 646 MLST STs (n=4,685 isolates) retrieved by the Phyloviz software. The numbers of the allelic differences are shown on the lines of the branches of the phylogenetic tree. (b) Assignment of ST182 E. hormaechei into a clonal complex with ST98, ST710, ST1611, ST1752, ST2143 and ST2608 by using the goeBURST algorithm.
Figure 1.
(a) Phylogenetic relationships of 646 MLST STs (n=4,685 isolates) retrieved by the Phyloviz software. The numbers of the allelic differences are shown on the lines of the branches of the phylogenetic tree. (b) Assignment of ST182 E. hormaechei into a clonal complex with ST98, ST710, ST1611, ST1752, ST2143 and ST2608 by using the goeBURST algorithm.
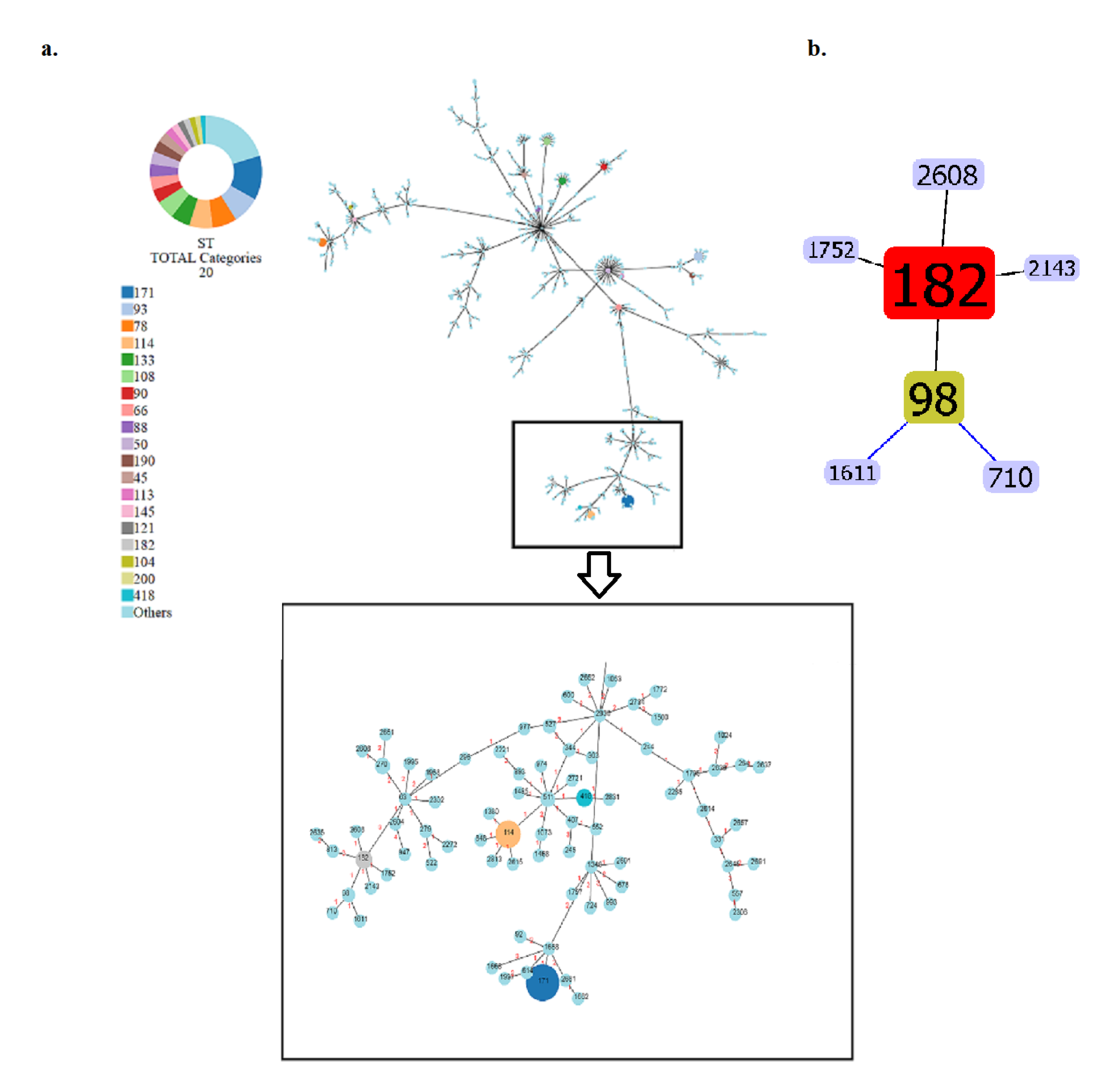
Figure 2.
The geographical distribution, phylogenomic analysis and timeline of isolation of the 55 ST182 E. hormaecheiobtained with the Microreact software.
Figure 2.
The geographical distribution, phylogenomic analysis and timeline of isolation of the 55 ST182 E. hormaecheiobtained with the Microreact software.
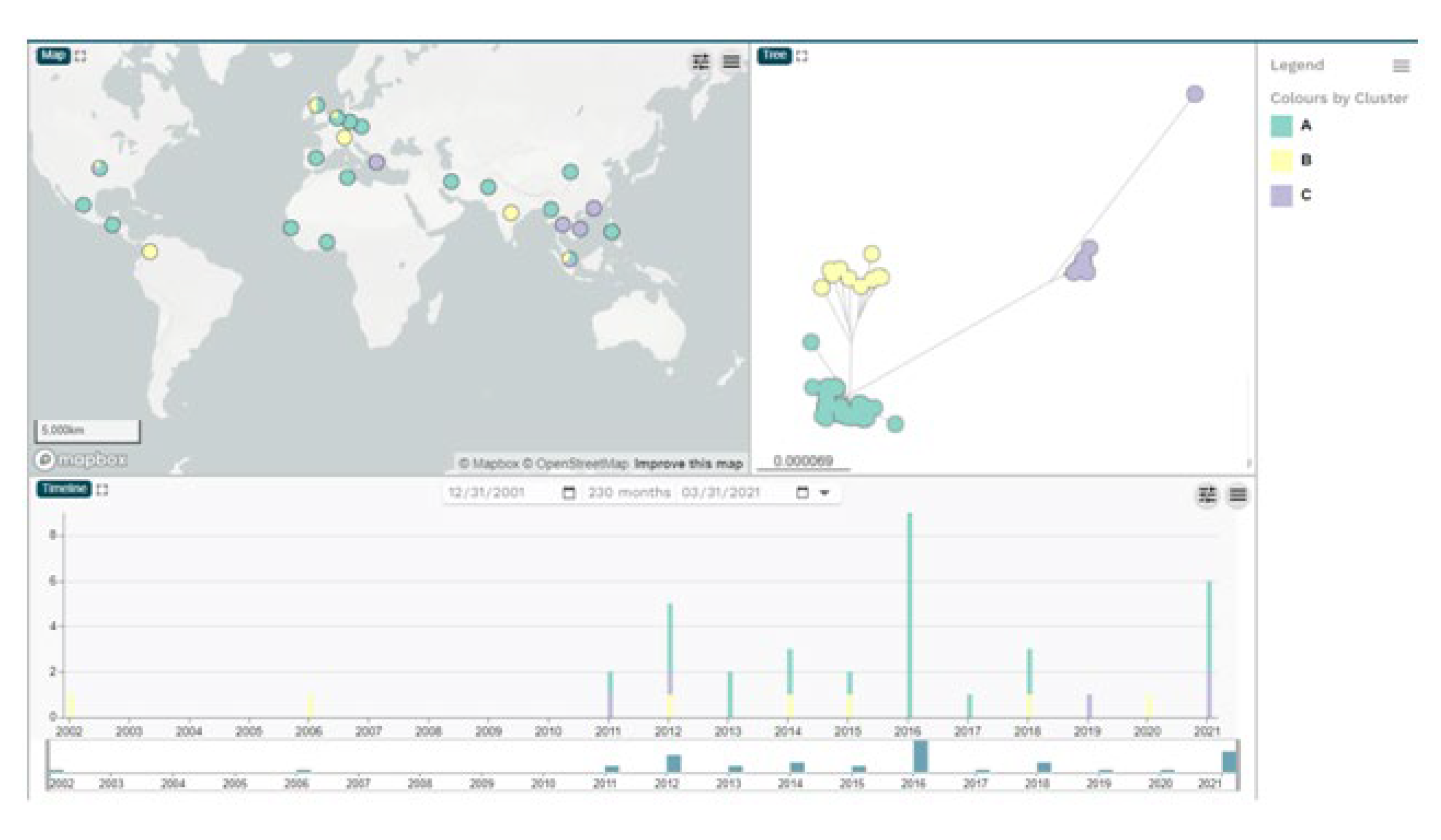
Figure 3.
(a) The geographical distribution of the genomic clusters; (b) blaNDM variants, and (c) blaNDM plasmid types of 30 ST182 E.hormaechei blaNDM carriers.
Figure 3.
(a) The geographical distribution of the genomic clusters; (b) blaNDM variants, and (c) blaNDM plasmid types of 30 ST182 E.hormaechei blaNDM carriers.
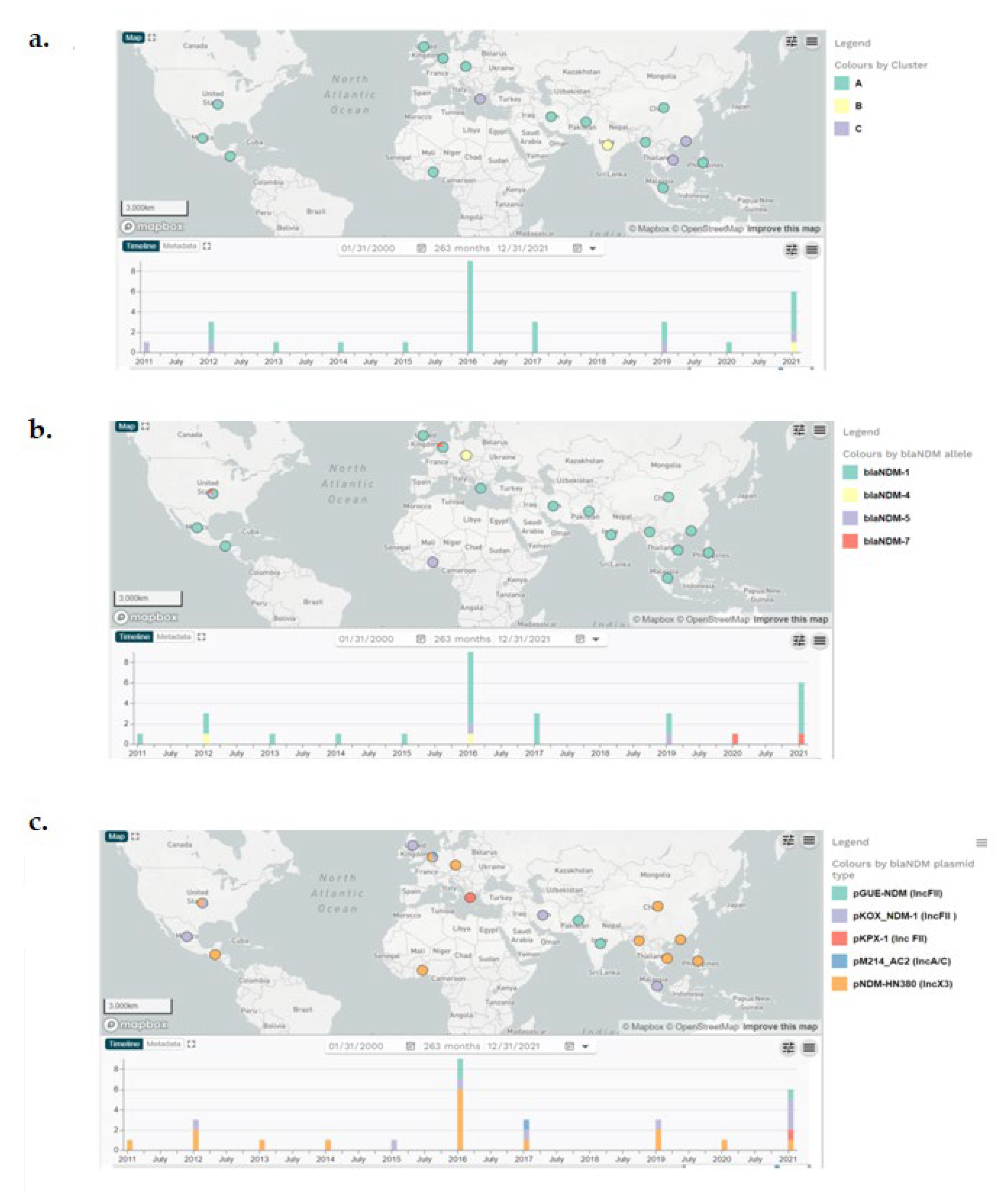

Table 1.
Distribution of 55 ST182 E. hormaechei isolates into genomic clusters (sublineages), continent/country of isolationand blaNDM variants.
Table 1.
Distribution of 55 ST182 E. hormaechei isolates into genomic clusters (sublineages), continent/country of isolationand blaNDM variants.
| Genomic clusters |
Continent/country of isolation |
blaNDM variants | Total | ||||
|---|---|---|---|---|---|---|---|
| blaNDM-1 | blaNDM-4 | blaNDM-5 | blaNDM-7 | None | |||
| Cluster A | 19 | 2 | 1 | 2 | 13 | 37 | |
| Africa | 1 | 3 | 4 | ||||
| Senegal | 2 | 2 | |||||
| Togo | 1 | 1 | |||||
| Tunisia | 1 | 1 | |||||
| Asia | 10 | 1 | 11 | ||||
| China | 2 | 1 | 3 | ||||
| Iran | 1 | 1 | |||||
| Myanmar | 3 | 3 | |||||
| Pakistan | 2 | 2 | |||||
| Philippines | 1 | 1 | |||||
| Singapore | 1 | 1 | |||||
| Europe | 4 | 2 | 1 | 5 | 12 | ||
| CzechRepublic | 2 | 2 | |||||
| Germany | 1 | 1 | |||||
| Netherlands | 3 | 1 | 4 | ||||
| Spain | 3 | 3 | |||||
| United Kingdom | 1 | 1 | 2 | ||||
| North America | 4 | 1 | 4 | 9 | |||
| Guatemala | 1 | 1 | |||||
| USA | 3 | 1 | 4 | 8 | |||
| SouthAmerica | 1 | 1 | |||||
| Mexico | 1 | 1 | |||||
| Cluster B | 1 | 9 | 10 | ||||
| Asia | 1 | 1 | 2 | ||||
| India | 1 | 1 | |||||
| Singapore | 1 | 1 | |||||
| Europe | 4 | 4 | |||||
| Netherlands | 1 | 1 | |||||
| Switzerland | 1 | 1 | |||||
| United Kingdom | 2 | 2 | |||||
| North America | 2 | 2 | |||||
| USA | 2 | 2 | |||||
| SouthAmerica | 2 | 2 | |||||
| Colombia | 2 | 2 | |||||
| Cluster C | 4 | 1 | 3 | 8 | |||
| Asia | 3 | 1 | 4 | ||||
| HongKong | 1 | 1 | |||||
| Singapore | 1 | 1 | |||||
| Thailand | 1 | 1 | |||||
| VietNam | 1 | 1 | |||||
| Europe | 1 | 1 | |||||
| Greece | 1 | 1 | |||||
| North America | 1 | 2 | 3 | ||||
| USA | 1 | 2 | 3 | ||||
| Total | 24 | 2 | 2 | 2 | 25 | 55 | |
Table 2.
Distribution of the 30 blaNDM-producing ST182 E. hormaechei isolates into plasmidtypes, blaNDM variants, continent of isolation and genomic clusters (sublineages).
Table 2.
Distribution of the 30 blaNDM-producing ST182 E. hormaechei isolates into plasmidtypes, blaNDM variants, continent of isolation and genomic clusters (sublineages).
| Plasmid types, blaNDM variants and continent of isolation |
Genomic clusters (sublineages) |
Number of isolates | ||
|---|---|---|---|---|
| A | B | C | ||
| pNDM-HN380 (IncX3) | 13 | 3 | 16 | |
| blaNDM-1 | 8 | 2 | 10 | |
| Asia | 6 | 2 | 8 | |
| Europe | 1 | 1 | ||
| North America | 1 | 1 | ||
| blaNDM-4 | 2 | 2 | ||
| Europe | 2 | 2 | ||
| blaNDM-5 | 1 | 1 | 2 | |
| Africa | 1 | 1 | ||
| North America | 1 | 1 | ||
| blaNDM-7 | 2 | 2 | ||
| Europe | 1 | 1 | ||
| North America | 1 | 1 | ||
| pKOX_NDM-1 (IncFII ) | 8 | 8 | ||
| blaNDM-1 | 8 | 8 | ||
| Asia | 2 | 2 | ||
| Europe | 2 | 2 | ||
| North America | 3 | 3 | ||
| SouthAmerica | 1 | 1 | ||
| pGUE-NDM (IncFII) | 2 | 1 | 3 | |
| blaNDM-1 | 2 | 1 | 3 | |
| Asia | 2 | 1 | 3 | |
| pKPX-1 (Inc FII) | 1 | 1 | ||
| blaNDM-1 | 1 | 1 | ||
| Europe | 1 | 1 | ||
| pJN24NDM1 (IncN2) | 1 | 1 | ||
| blaNDM-1 | 1 | 1 | ||
| Asia | 1 | 1 | ||
| pM214_AC2 (IncA/C) | 1 | 1 | ||
| blaNDM-1 | 1 | 1 | ||
| Europe | 1 | 1 | ||
| Total | 24 | 1 | 5 | 30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



