
Submitted:
19 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Cancers, a large and heterogeneous group of malignancies, are becoming an increasingly important cause of premature mortality worldwide. 2% of these are primary CNS tumors, of which 24% are gliomas. These tumors are diverse, but what they have in common is one of the most unfavorable prognoses among all cancers. A patient's future varies depending on the grade of the diagnosed glioma. To date, however, no standard treatment for grade IV has been established. All glioblastomas eventually undergo progression or recurrence. The current standard of treatment, which includes surgical intervention, radiation therapy and chemotherapy, is therefore far from sufficient. Work is constantly underway to discover a new, effective form of glioma therapy. Photodynamic therapy (PDT) may be one of them. It involves the local or systemic application of a photosensitive compound - a photosensitizer (PS), which accumulates in the affected tissues. Photosensitizer molecules absorb light of the appropriate wavelength, initiating activation processes leading to the formation of reactive oxygen species and selective destruction of inappropriate cells. Currently, this method has been approved for the treatment of several cancers. Research focusing on the effective use of PDT in glioma therapy is already underway, with promising results. In our work, we present molecular insights into PDT of glioma. Based on the available literature, we analyze and systematize the impact of various molecules, proteins, transporters and transmitters on the efficacy of PDT and the effect of PDT on their expression. In addition, we highlight gaps in current knowledge and point out directions for future research that may contribute to the efficacy of PDT glioma.
Keywords:
Photodynamic Therapy (PDT)
; glioma
; molecular insights
; treatment cancer
; oncology
1. Introduction
Cancers, a large and heterogeneous group of malignancies, are becoming an increasingly important cause of premature mortality worldwide [1,2]. Although primary tumors of the central nervous system (CNS) account for only 2% of all primary cancers, they cause 7% of cancer deaths in people under 70 years of age [3,4]. Gliomas account for 24% of all primary brain and CNS tumors [4]. These tumors are diverse in terms of cellular origin, genetic profile and morphology, but together have one of the most dismal prognoses of all cancers [5]. These tumors are diverse in terms of cellular origin, genetic profile and morphology, but together have one of the most dismal prognoses of all cancers [6]. However, there is no standard treatment for stage IV. All glioblastomas eventually progress or recur [7]. The current standard of therapy, which includes surgical intervention, radiotherapy and chemotherapy, is therefore far from sufficient [8]. Work is still underway to discover a new, effective form of glioma therapy [9,10]. One of them may be photodynamic therapy (PDT) [11]. It involves the topical or systemic application of a photosensitive compound, a photosensitizer (PS), which accumulates in the affected tissues. Photosensitizer molecules absorb light of the appropriate wavelength, initiating activation processes leading to the formation of reactive oxygen species and selective destruction of inappropriate cells [12]. Currently, this method has been approved for the treatment of head and neck cancer, esophageal cancer, pancreatic cancer, prostate cancer and esophageal squamous cell carcinoma. In addition, it is widely used in dermatology to combat precancerous and cancerous lesions [13]. Research focusing on the effective use of PDT in glioma therapy is already underway with promising results [14]. In our work, we present molecular insights into PDT of glioma. Based on the available literature, we analyze and systematize the influence of various molecules, proteins, transporters and transmitters on the efficacy of PDT and the effect of PDT on their expression. In addition, we highlight gaps in current knowledge and point out directions for future research that may contribute to the efficacy of PDT glioma.
2. Adenosine Triphosphate (ATP)-Binding Cassette Efflux Transporter G2
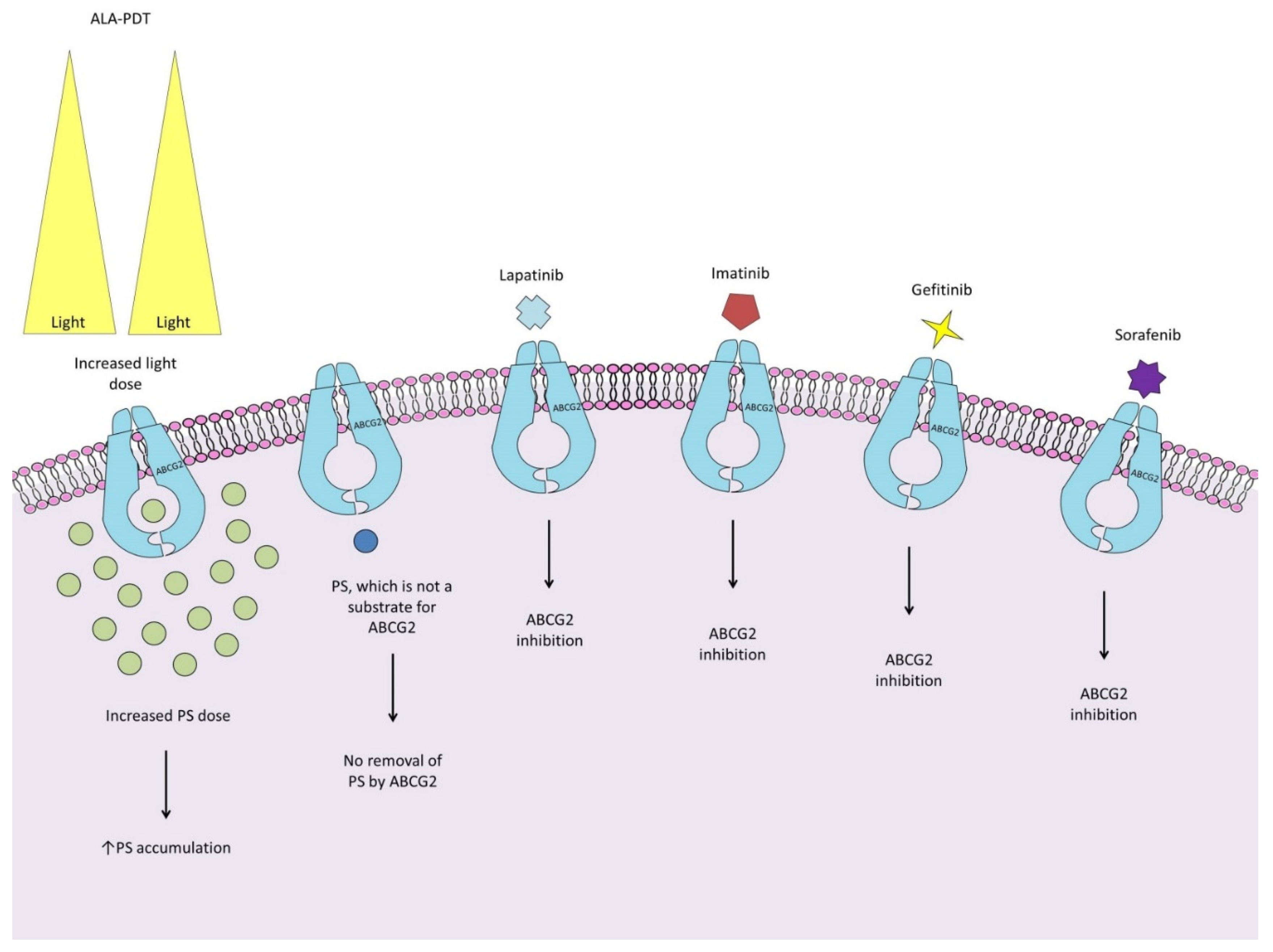
Adenosine triphosphate (ATP)-binding cassette efflux transporter G2 plays a central role in the efflux of various molecules from cells, providing protection against the adverse effects of xenobiotics [15]. Observations by Morgan et al. indicate that cells with ABCG2 overexpression are found only in a small group of U87 glioma side population (SP) cells, which also harbor resistant stem cells and may be a source of future tumor growth [16]. The involvement of ABCG2 in the response to PDT is not clear, and there are conflicting findings. Some reports indicate that glioma cells with high ABCG2 expression accumulate less photosensitizer, so ABCG2 may be responsible for the poor response to PDT [17,18]. In this work, the effect of PDT alone, and combined with the ABCG2 inhibitors KO143 and its anaologue fumitremorgin C, at concentrations of 1.5 μM and 5 µM, respectively, were compared. However, Weidner et al. demonstrate that KO143 at concentrations ≥1 μM also affects the transport activity of both ABCB1 and ABCC1, which may have influenced the outcome of these studies [19]. Wang et al. showed that the use of reserpine and verapamil, inhibitors of ABCG2, did not result in increased accumulation of protoporphyrin IX (PpIX) in both SP and non-SP cells. However, this may be due to their affinity for ABCB6 resulting in decreased synthesis of PpIX [20]. On the other hand, Abdel Gaber et al. restored PS Chlorin e6 (Ce6) accumulation using an anti-ABCG2 antibody, which would indicate a key role for this transporter in the removal of PS [21]. The presence of ABCG2 alone does not indicate that glioma cells are resistant to PDT. The work of Hamid et al. achieved PDT efficacy despite 6-fold reduced PS photolon accumulation [22]. Regardless of the exact role of ABCG2, improving the phototoxicity of cells with expression of this transporter is possible through several approaches. Some work indicates that ABCG2 saturation and improved PDT efficacy is provided by increasing light doses and PS [16,17]. Another of the strategies to solve this problem could be to use PSs that are not substrates for ABCG2, as demonstrated by Selbo et al. [23]. Another approach is to use ABCG2 inhibitors. Because of their ability to inhibit this transporter, clinically approved kinase inhibitors have been tested for this purpose. The results achieved vary depending on the inhibitor used. PDT with PS 5-aminolevulinic acid (ALA-PDT) in combination with imatinib was characterized by such high phototoxicity that the number of surviving SP cells was too small to measure [16]. Gefitinib, acting by reducing mRNA and plasma membrane protein ABCG2 expression, increased intracellular PpIX levels in a dose-dependent manner [24]. Lapatinib, also increased ALA-PpIX fluorescence in a dose-dependent manner, but was about 15% less effective than the KO143 inhibitor [25]. A similar effect was obtained using sorafenib, but after its use, Ce6 levels also failed to reach those achieved by KO143 or the anti-ABCG2 5D3 antibody [21]. However, kinase inhibitors do not selectively block ABCG2, so they may also increase the effectiveness of PDT by affecting other targets.
Figure 1.
Strategies to improve the efficacy of photodynamic therapy for ABCG2-expressing glioma cells: 1) ABCG2 saturation and improved PDT efficacy can be achieved by increasing light and PS doses [16,17]; 2) Increased PS concentration can be achieved by using Strongly amphiphilic photosensitizers that are not substrates for ABCG2 [23]; 3), 4), 5), 6): Association of PDT with ABCG2 inhibitors lapatinib, imatinib, gefitiniben, and sorafenib increases the efficacy of glioma PDT [16,21,24,25].
Figure 1.
Strategies to improve the efficacy of photodynamic therapy for ABCG2-expressing glioma cells: 1) ABCG2 saturation and improved PDT efficacy can be achieved by increasing light and PS doses [16,17]; 2) Increased PS concentration can be achieved by using Strongly amphiphilic photosensitizers that are not substrates for ABCG2 [23]; 3), 4), 5), 6): Association of PDT with ABCG2 inhibitors lapatinib, imatinib, gefitiniben, and sorafenib increases the efficacy of glioma PDT [16,21,24,25].
3. Glutathione
Acting as an antioxidant, glutathione (GSH) promotes tumor progression, and elevated levels correlate with temozolomide resistance and increased glioma metastasis [26,27]. High levels of intracellular GSH are also observed in PDT-resistant cells [28]. To address this issue and improve the efficacy of PDT, the blocker of reduced GSH synthesis, buthionine sulfoximine (BSO), was tested. Overnight treatment with 100 μM BSO followed by 4 hours of coincubation with ALA resulted in a nearly 100% reduction in intracellular GSH of the T98G and U87 lines [29]. However, lowering GSH did not always result in increased PDT efficacy. In the above study, a significant improvement in ALA-PDT efficacy was achieved only in the T98G line, reducing cell survival from 70 to 18% [29]. On the other hand, Jiang et al. report that BSO already at the lowest dose used (0.225 μM) significantly increased Photofrin-PDT (Ph-PDT) activity in U87 and U251n lines in vitro after 24-hour coincubation, and this effect was dose-dependent. BSO also potentiated the effect of Ph-PDT in U87 cells in vivo [30]. Thus, the data are disparate, and the different PS used, BSO dose, incubation time, and the fact that BSO itself exhibits cytotoxic activity make it difficult to interpret these differences and point to the need for further research [31]. However, the presence of GSH in glioma cells can also be used to improve the effect of PDT. Hwang et al. engineered PS ubiquinone-BODIPY, whose reduction by intracellular glutathione results in increased generation of singlet oxygen [32]. GSH also enables PS and chemotherapeutic agent release, which is being taken into account in the design of new nanoparticles [33,34,35].
4. Ferrochelatase
Ferrochelatase (FECH) is an enzyme that converts photochemically active PpIX into photochemically inactive heme by incorporating ferrous iron [36]. High expression and activity of this enzyme has been linked to glioma cell resistance to ALA-PDT [29,37]. Moreover, hypoxia was found to cause increased ferrochelatase gene expression in glioblastoma stem cells, which may further contribute to ALA-PDT resistance [38]. Therefore, iron chelation represents an attractive target that can enhance the efficacy of ALA-PDT. Reburn et al. synthesized an ester between ALA and the hydroxypyridinone iron chelating agent CP94 and observed significant enhanced effects on both PpIX accumulation and PDT cytotoxicity in glioma cells [39]. Also, coincubation of CP94 with ALA, methyl aminolevulinate (MAL) and hexyl aminolevulinate (HAL) increased PpIX accumulation. Blake et al. compared these results with coincubation with dexrazosan, a clinically approved iron chelating prodrug, and found that it had less efficacy than CP94 [40,41]. Another approved compound tested for this purpose is deferoxamine (DFO), but data on its efficacy are conflicting. A paper by Wang et al. showed that it effectively increased PpIX accumulation in glioma lateral population stem cells [20]. On the other hand, Mansi et al. found that the addition of DFO led to only a slight increase in PpIX fluorescence in all glioma cell lines, significantly reducing the viability of only A172 cells [25]. It is difficult to determine the reason for this difference. The incubation time was identical in both studies at 4 hours. A lower concentration of DFO (100 µM) was used in the first study than in the second (1 mM), and yet DFO was found to be effective. Different cell lines were used in both works. This may have played a role in the discrepancy, as Teng et al. showed that ferrochelatase levels were lowest in the U87 line and therefore did not use them to try to silence the enzyme [42]. However, this was the only common line also used by Mansi et al. In their work, the FECH activity measured in U87 cells was lower than in H4 cells, but despite this, no significant differences in fluorescence were observed after DFO treatment [25]. On the other hand, Blake et al. used the U-87 line and, as mentioned above, successfully inhibited FECH, but using compounds other than DFO [40]. The role of DFO in inhibiting glioma cell ferrochelatase is therefore unclear and requires further clarification.
5. Heme Oxygenase 1
Heme oxygenase 1 (HO-1) is an enzyme that catalyzes the oxidation of the heme group to form biliverdin, it also generates carbon monoxide and ferrous iron as reaction products. HO and its reaction products have protective effects against oxidative stress, anti-inflammatory and signaling effects [43]. Observations by Wang et al. indicate that cells of the lateral population of glioma show significantly higher expression of HO-1. Moreover, the expression of this enzyme is much higher in WHO grade IV GBM. Higher levels of HO-1 expression may accelerate the PpIX/hem metabolic pathway, leading to poor ALA-mediated PpIX accumulation and a low response to ALA-PDT [20]. The expression of heme oxygenase 1 increases significantly after both ALA administration alone and subsequent irradiation [20,29]. The reduction of HO-1 activity may therefore contribute to the efficacy of glioma therapy. Mastrangelopoulou et al. observed, however, that the synergistic effect of ALA PDT and the HO-1 inhibitor OB24 was observed only in T98G cells preincubated overnight. They did not find it in other cell lines, nor during hourly preincubation [29]. Since HO-1 inducibility can also occur in response to activation of growth factor receptor signal transduction pathways, including extracellular signal-regulated kinases ERK1 and ERK2, c-jun-NH2 kinase, and p38 kinase, in order to further reduce the negative effects of HO-1 on the efficacy of ALA-PDT glioma, it seems attractive to test molecules that affect these pathways [44,45,46].
6. Hypoxia-Inducible Factor 1 α
Hypoxia-inducible factor 1 α (HIF-1α) is a transcription factor that activates multiple glioma survival signaling pathways [47,48]. Its level is increased by an oxygen concentration of about 2.5% to 10%, found in most glioblastoma multiforme [49]. It has been observed that HIF-1 α expression increases after PDT in many cancers [50,51,52]. In the case of glioma, it increases with time since PDT application [53]. Moreover, sublethal PDT induces an increase in HIF-1α expression in healthy brain tissue as well, stimulating expression of the ADAM17-EGFR-PI3K-Akt pathway and increasing the invasion of subsequently implanted glioma cells [54,55]. HIF-1α may be responsible for PDT resistance in several types of cancer [56,57]. Thus, it can be speculated that there is a mechanism whereby glioma cells attenuate the PDT response through increased HIF-1α levels, and PDT further exacerbates this effect through additional induction of this factor induced by oxygen consumption, possibly ultimately leading to tumor proliferation, metastasis and invasion. Work on solving this problem is still ongoing [58]. There is a report indicating that selecting the optimal therapeutic regimen may not negatively affect HIF-1α expression. Li et al. showed that PDT with PS hematoprophyrin monomethyl ether (HMME-PDT) incubated 2 h in the dark with glioma cells and irradiated for 60 s with a 0.75 J/cm2 laser led to a decrease in the level of HIF-1α [59]. Another solution, is to use a molecule that does not induce an increase in HIF-1α levels. This was achieved by Xu et al. but the active photodynamic phthalocyanine-based molecular beacon they developed increased VEGF expression [60]. A third approach may be the association of PDT with agents that reduce HIF-1α expression. A liposome composed of the HIF-1α inhibitor vitexin and PS indocyanine green (IG) showed potent inhibition of tumor cells and suppression of their migration in a dose-dependent manner [61]. Association of Acriflavin, an inhibitor of HIF-1α/HIF-1β dimerization, with ALA-PDT reduced the expression of HIF-1α, GLUT-1, GLUT-3, and HK2, improved the efficacy of treatment of refractory GBM, and reduced tumor cell invasion and migration [62]. Molecules that do not directly target this transcription factor can also lead to HIF-1α downregulation. Association of PDT with endostar, a recombinant human endostatin, led to prolonged survival of U251 glioma mice, significantly reducing the expression of both VEGF-A and HIF-1α [53]. Since HIF-1α is induced by hypoxia, an entirely different strategy is to develop nanoparticles that reduce the problem of hypoxia, which advanced work on with favorable results is still underway [63,64,65,66].
7. Histone Deacetylases
Histone deacetylases (HDACs), are a group of enzymes that catalyze the acetylation of histones. They are involved in many processes, such as cancer cell growth and proliferation [67]. There are several different classes of HDACs, the expression of which, depending on the class, is increased or decreased in the glioma [68]. PDT affects the expression and activity of HDACs. Single reports show an increase in HDACs activity in A375 melanoma cells and healthy brain tissue [69,70]. Work is already underway to develop optimal inhibitors of HDACs; however, clinical trials have shown that they are currently ineffective in the treatment of GBM [71]. One of the inhibitors being tested is sodium butyrate (NaB). It has been shown that it can induce apoptosis, inhibit VEGF expression and stimulate medulloblastoma differentiation [72,73,74]. Association of sodium butyrte with ALA-PDT increased U373-MG and D54-MG astrocytoma cell deaths by 67.8% and 53.95%, respectively, relative to ALA-PDT alone [75]. Modified chromatin and NaB-induced genes are responsible for this effect [76]. A different approach was used by Wei et al. using HDAC6 to selectively activate their engineered molecule and enhance the effect of PDT [77]. In conclusion, the lack of knowledge about HDACs and PDT in the glioma is considerable, but available work points to the potential use of these molecules and their inhibitors to improve the efficacy of PDT of the glioma.
8. Protein Kinase C
Histone Protein kinase C (PKC) is a family of serine/threonine-specific protein kinases [78]. PKC activity was found to be increased in gliomas compared to astrocytes [79,80]. PKCs play a diverse role in glioma development, as the contribution of each PKC isoform depends on the phosphorylation of tyrosine residues, the presence of oncogenic mutations, the type of stimuli and the cellular environment [78]. Previous work has shown that PDT affects the PKC activity of glioma cells, both through the action of PS alone and subsequent irradiation. Huntusova et al. report that hypericin interacts directly with PKCα and increases PKCδ autophosphorylation in the glomus [81]. This issue needs clarification, as there are reports indicating that hypericin can inhibit PKC in other cells [82]. Data on the effects of PDT on PKC in the glioma are limited. Uzdensky et al. demonstrate, that sublethal ALA-PDT induces an increase in PKCγ and PKCβ [83]. The effect of PDT on PKC is confirmed by the work of Dzurov et al. who showed that Hypericin-PDT (Hyp-PDT) caused activation of PKCα [84]. On the other hand, Pevna et al. achieved a decrease in PKCα levels by Hyp-PDT [85]. Since both Dzurova et al. and Pevna et al. used U87, Hyp cells at a concentration of 500 nM, but different light doses (4 J/cm2 and 2 J/cm2, respectively), the light dose may be crucial for PKCα regulation in glioma. Given the involvement of PKCα and PKCβ in the survival, proliferation and migration of glioma cells, the potential possibility of their regulation by an appropriately selected light dose seems particularly attractive [78]. An approach being tested to improve the efficacy of glioma PDT is association with PKC inhibitors. Tamoxifen at a dose of 500 μg/ml and higher was shown to significantly increase the toxicity of the Ph-PDT response [86]. Work on using tamoxifen for this purpose is particularly promising, as it has shown potential in treating glioma [87,88]. Pretreatment of glioma cells with rottlerin before Hyp-PDT led to a significant increase in apoptosis [89]. However, recent work has challenged previous findings, the main function of rottlerin as a PKCδ inhibitor [90,91,92]. A completely different approach is the association of photobiomodulation with PDT. It was shown that photobiomodulation performed before Hyp-PDT with 500 nM Hyp led to a greater decrease in PKCα than Hyp-PDT alone [85]. In conclusion, PKC targeting may be an attractive form of improving the effectiveness of PDT, but there is still much to be clarified.
9. Epidermal Growth Factor Receptor
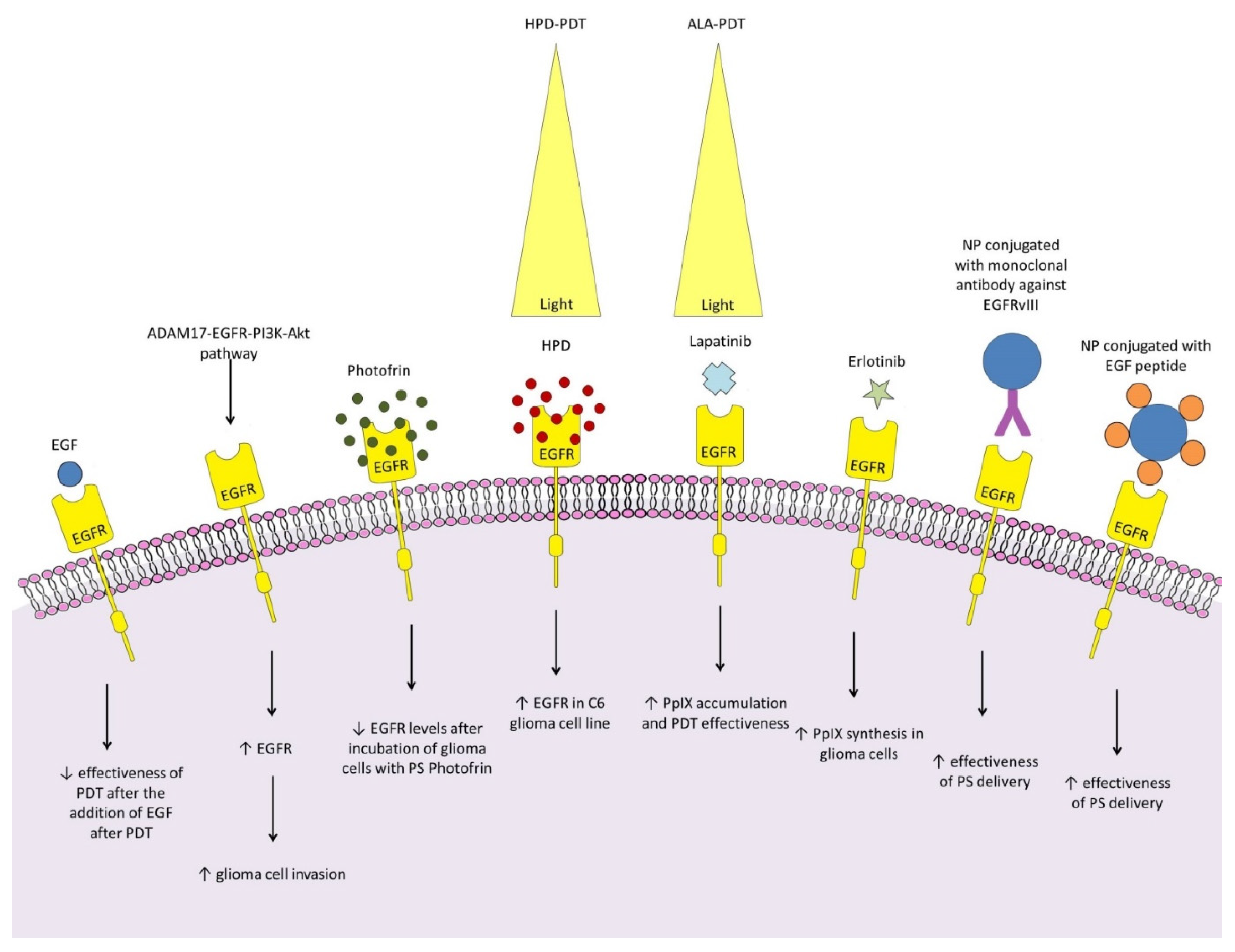
Epidermal growth factor receptor (EGFR) is a trans-membrane glycoprotein and belongs to the tyrosine kinase receptor superfamily [93]. High EGFR levels contribute to glioma development, progression and resistance in both kinase-dependent and -independent ways [94,95]. The exact effect of EGFR on the efficacy of glioma PDT has not yet been thoroughly investigated, however, available work suggests that EGFR may contribute to PDT resistance. It has been shown that Ph-PDT can induce the ADAM17-EGFR-PI3K-Akt pathway in healthy brain tissue, leading to an increase in EGFR in both healthy cells and subsequently implanted glioma cells, significantly increasing U87 cell invasion in nude mice [55]. The effect of EGF, an EGFR ligand, is also unexplained. Fanuel Barret et al. showed that it had no effect on PS haematoporphyrin derivative (HPD)-induced and laser-induced toxicity when added to cells before PDT, but added after it attenuated the cellular response to PDT [96]. Data on the direct effect of PDT on EGFR expression by glioma cells are inconclusive and disparate. Fanuel Barret et al. showed that incubation with PS HPD and subsequent laser irradiation had no effect on the increase of EGFR in T98G and U87 cells, but resulted in its increase in the C6 line [96]. On the other hand, in the work of Yang et al. incubation of T98G cells with PS Photofrin led to a decrease in EGFR levels [97]. It seems to be an interesting direction for future work to test the effect of ALA-PDT on EGFR expression by glioma cells, as testing other cancer cells has shown that it can lead to a reduction in EGFR levels [98]. Given the important involvement of EGFR in cancer pathogenesis, attempts to combine PDT with EGFR-targeting molecules are currently attracting considerable interest [99]. For glioma, the focus was on combining PDT with clinically accepted EGFR inhibitors. Erlotinib improved the selectivity of PpIX accumulation, as it increased PpIX synthesis in glioma cell lines, while not affecting PpIX synthesis in neuronal and astrocytic lines [100]. The association of lapatinib with ALA-PDT led to a significant increase in PpIX accumulation in glioma cells and induced stronger responses of two human glioma tumors in vivo, leading to increased survival in rats compared with lapatinib alone and PDT alone [101]. However, it should be emphasized that none of the EGFR-targeted therapies have shown promising results in clinical trials in patients with glioma, so these positive results of the work on the combination of PDT with EGFR inhibitors should be treated with caution and need to be confirmed clinically [102]. EGFR and EGF have also been successfully used as targets to improve PS delivery to glioma cells [103,104].
Figure 2.
Relationships between photodynamic therapy, glioma and EGFR: EGF had no effect on PS haematoporphyrin derivative (HPD)- and laser-induced toxicity when added to cells before PDT, but added after attenuated the cellular response to PDT [96]; 2) Ph-PDT can induce the ADAM17-EGFR-PI3K-Akt pathway in healthy brain tissue, leading to an increase in EGFR in glioma cells as well as healthy brain cells, significantly increasing the invasion of U87 cells in nude mice [55]; 3). incubation of T98G cells with PS Photofrin led to a decrease in EGFR levels [97]; 4) HPD-PDT increased EGFR in C6 cells, but not T98G and U87 cells [96]; 5) The association of lapatinib with ALA-PDT led to a significant increase in PpIX accumulation and induced stronger responses of two human glioma tumors in vivo, leading to increased survival in rats compared to lapatinib alone and PDT alone [101]; 6) Erlotinib improved the selectivity of PpIX accumulation as it increased PpIX synthesis in glioma cell lines [100]; 7), 8) Coupling NP with a monclonal antibody against EGFR, or EGF peptide improves the efficiency of PS delivery to the glioma [103,104].
Figure 2.
Relationships between photodynamic therapy, glioma and EGFR: EGF had no effect on PS haematoporphyrin derivative (HPD)- and laser-induced toxicity when added to cells before PDT, but added after attenuated the cellular response to PDT [96]; 2) Ph-PDT can induce the ADAM17-EGFR-PI3K-Akt pathway in healthy brain tissue, leading to an increase in EGFR in glioma cells as well as healthy brain cells, significantly increasing the invasion of U87 cells in nude mice [55]; 3). incubation of T98G cells with PS Photofrin led to a decrease in EGFR levels [97]; 4) HPD-PDT increased EGFR in C6 cells, but not T98G and U87 cells [96]; 5) The association of lapatinib with ALA-PDT led to a significant increase in PpIX accumulation and induced stronger responses of two human glioma tumors in vivo, leading to increased survival in rats compared to lapatinib alone and PDT alone [101]; 6) Erlotinib improved the selectivity of PpIX accumulation as it increased PpIX synthesis in glioma cell lines [100]; 7), 8) Coupling NP with a monclonal antibody against EGFR, or EGF peptide improves the efficiency of PS delivery to the glioma [103,104].
10. Fibroblast Growth Factors
Fibroblast growth factors (FGFs) are mitogens that regulate a wide range of cellular functions, including migration, proliferation, differentiation and survival [105]. Their receptors (FGFR) are overexpressed and mutated in many types of cancer [106]. High levels of FGF1 and FGF2 o in glioma tissue contribute to angiogenesis, growth, invasion and resistance to treatment of this cancer [106,107]. High levels of FGF1 and FGF2 o in glioma tissue contribute to angiogenesis, growth, invasion and resistance to treatment of this cancer [108,109]. There are isolated papers describing the relationship between glioma, FGF and PDT efficacy. Vilchez et al. showed that glioma cells resistant to PDT with methyl-5-aminolevulinic acid PS expressed higher mRNA levels of the fibroblastic growth factor receptor (FGFR), but this was not the only receptor up-regulated [110]. However, it does not appear that FGF2 alone is crucial to the efficacy of ALA-PDT, as FGF2 added to U-105MG glioma cells did not have a stimulating or inhibitory effect on final treatment outcomes [111]. However, this was an in vitro study, and since FGF2 induces angiogenesis by activating endothelial cell proliferation and migration, the exact effect would need to be studied in vivo [106]. A new strategy leading to, among other things, downregulation of the fibroblast growth factor receptor and increased apoptosis was developed by Chakrabarti et al. by associating Ph-PDT with transfection of miR-99a [112]. Some of the selective FGF/FGFR inhibitors have already been approved for therapy, and testing them in combination with PDT could be an interesting direction to potentially increase the efficacy of PDT and help further clarify the relationship between PDT, FGF/FGFR and glioma [113].
11. Hepatocyte Growth Factor
Hepatocyte growth factor (HGF), also called scatter factor, along with its receptor tyrosine kinase c-Met, are key determinants of brain tumor growth and angiogenesis [114]. HGF can affect tumor progression through various mechanisms, including modulating cell growth, migration and chemo-resistance to drugs [115]. There is one paper describing the effect of PDT on HGF. The study by Vogel et al. showed that ALA-PDT induced HGF expression in glioblastoma spheroids and thus stimulated migration of mesenchymal stem cells into the tumor. Moreover, they conclude that both the increase in HGF expression and MSC migration after PDT can have positive and negative effects on the ultimate efficacy of therapy [116]. Development and effective use of HGF inhibitor already underway [117].
12. Vascular Endothelial Growth Factor
Vascular endothelial growth factor (VEGF) is the most important mediator of angiogenesis in glioma [118]. The formation of abnormal tumor vascularization is one of the main factors responsible for the resistance of these tumors to treatment, and VEGF levels in glioma patients are higher than in healthy individuals [119,120]. Therefore, an increase in VEGF is particularly undesirable [120]. The effect of PDT on VEGF levels is adverse. The ability of PDT to induce VEGF expression in both tumor and healthy brain tissue in a dose-dependent manner of the light used is well documented in the literature [60,121,122,123,124,125,126]. A key approach to improving the efficacy of glioma PDT may therefore be to inhibit VEGF expression or block the adverse effects of its receptor. Combined treatment with Ce6-PDT and the humanized anti-VEGF monoclonal antibody bevacizumab prolonged the mean survival time of mice relative to PDT alone [127]. The association of Ph-PDT with anti-VEGFR-1 and anti-VEGFR-2 monoclonal antibodies reduced tumorigenesis more and prolonged mouse survival relative to PDT alone. Moreover, it also led to a significant reduction in the expression of both VEGF and von Willebrand Factor [128]. The association of PDT with endostatin, an endogenous VGFR-inhibiting angiogenesis inhibitor, seems to be an interesting approach. It was previously observed that PDT with PS Hypocrellin-A and -B induces VEGF and endostatin release in glioma cells, while reducing endostatin release in endothelial cells, and it has been shown that the predominance of VEGF over endostatin observed early after PDT, may contribute to enhanced angiogenesis [121,129]. This approach was tested by Zhan et al.. By associating endostatin with HMME-PDT, they enhanced the effects of PDT, leading to greater tumor shrinkage, prolonged mouse survival, and decreased expression of HIF-1α and VEGF-A [53]. However, the status of endostatin as an effective inhibitor of angiogenesis is debatable [130]. Important to the efficacy of VEGF-blocking treatment and PDT is the timing of administration of the inhibitor, as it has been shown that, when administered before PDT, it can reduce PS levels and blunt the effects of PDT [131]. However, it should be noted that the results of clinical trials of antiangiogenic drugs in patients with malignant gliomas have been generally disappointing [132]. Thus, a combination therapy with bevacizumab appears to be a particularly promising direction for future research, as its use has previously been shown to benefit patients with recurrent glioblastoma [132].
13. Neuropilin-1
Neuropilin-1 (NRP-1) is a transmembrane protein involved in glioma proliferation, invasion and migration, as well as tumor angiogenesis [133]. This molecule is overexpressed in the glioma [134,135]. NRP1 expression has been shown to be correlated with poor prognosis, glioma grade and associated with mesenchymal tumor subtype [136]. Previous work has shown that the association of PS with NRP-1-targeting molecules promotes the vascular effect of PDT, ultimately resulting in reduced tumor blood flow and delayed tumor growth [137,138,139,140,141,142]. This coupling has a higher efficiency than the PS itself [143]. Neuropilin-1 has also been successfully used as a target to improve PS delivery to glioma cells [144].

Table 1.
Molecular targets tested to improve photosensitizer delivery to the glioma. The first column presents the molecular target used. In the second, the molecule responsible for the targeted delivery, which was conjugated to either a photosensitizer or a photosensitizer-loaded nanoparticle.
Table 1.
Molecular targets tested to improve photosensitizer delivery to the glioma. The first column presents the molecular target used. In the second, the molecule responsible for the targeted delivery, which was conjugated to either a photosensitizer or a photosensitizer-loaded nanoparticle.
| Molecular target | Targeting method | Article |
|---|---|---|
| EGFR | Monoclonal antibody against EGFRvIII | Jamali et al. [103] |
| EGFR | EGF peptide | Meyers JD et al. [104] |
| Neuropilin-1 | ATWLPPR heptapeptide targeting NRP-1 | Thomas et al. [137] Tirand et al. [138] Bechet et al. [139] Tirand et al. [142] Thomas et al. [143] |
| Neuropilin-1 | KDKPPR peptide targeting NRP-1 | Gries et al. [140] Thomas et al. [141] |
| Neuropilin-1 Integrin αvβ3 Integrin αvβ5 |
RGD internalizing peptide targeting NRP1, INT αvβ3 and αvβ5 | Lu et al. [144] |
| Integrin αvβ3 | Modified monoclonal antibody against INT αvβ3 | Wei et al. [157] |
| Neuropeptide Y receptor 1 | Neuropeptide Y type D | He et al. [255] |
| Low-density lipoprotein receptor | Low density lipoprotein | Andreazza et al. [269] Huntosova et al. [270] |
| Extra domain A of fibronectin (ED A) | Ligand targeting epitope ED A small immunoprotein antibody F8 | Acker et al. [281] |
| US28 protein | Nanoprotein binding discontinuous epitope US28 with high affinity | De Groof et al. [282] |
| Nucleolin | Single-stranded DNA aptamer AS1411 with high affinity for nucleolin | Zhu et al. [283] |
| Transferrin receptor | Transferrin | Zhu et al. [283] |
| Cell nucleus | T-ag antigen variants of SV40 virus | Akhlynina et al. [284] |
14. Vascular Cell Adhesion Protein 1
Vascular cell adhesion protein 1 (VCAM-1, CD106) is a glycoprotein involved in angiogenesis and cancer metastasis [145]. VCAM-1 is highly expressed in both gliomas and surrounding normal brain tissue [146,147]. It has been shown that PDT can induce an increase in VCAM-1 levels in the glioma [148]. However, the available data on the effects of PDT on VCAM-1 expression in tumors are conflicting, so this issue requires additional clarification [149,150]. Due to its functions and the high expression of VCAM-1 in the glioma, targeting this molecule may be a strategy to increase the effectiveness of therapy. Zhan et al. showed that the combination of HMME-PDT with a monoclonal antibody against VCAM-1 induced apoptosis and prolonged mouse survival more than PDT alone [148].
15. Integrins
Integrins (INTs) are a large family of molecules and receptors on the cell surface, consisting of 18 α subunits and 8 β subunits, enabling the formation of 24 unique INTs [151]. They are important surface adhesion receptors that mediate interactions between the extracellular matrix and cells and are essential for cell migration and maintenance of tissue homeostasis. Abnormal activation of integrins promotes the initial formation, growth and metastasis of tumors [152]. INTs are among the major contributors to the invasive glioma phenotype, particularly abnormal expression of β1 subunit, α3β1 integrin, αvβ3, αvβ5 integrin [153]. The beneficial effects of PDT on integrin levels in tumors have already been fairly well studied [154]. A similar effect of PDT was demonstrated in the glioma. The administration of PS alone could reduce the level of integrin α3β1 in the glioma, and subsequent irradiation exacerbated this effect depending on the light dose, the PS applied, and the glioma cell line [155]. These observations are consistent with those of other cancers [154,156]. Integrins can also be used as useful targets to improve PS delivery and hit nanoparticles (NP) [144,157].
16. Matrix Metalloproteinases
Matrix metalloproteinases (MMPs) are a group of enzymes that catalyze the remodeling of the extracellular matrix [158]. They play a key role in the mechanisms of glioma invasion, and increased expression of some of them has been linked to poor prognosis and glioma recurrence [159,160]. PDT leads to decreased expression of MMPs (-2, -7, -8, -9) glioma and cell migration, which is well documented in the literature and consistent with observations from other cancers [155,161,162,163,164,165,166]. It has been shown that the association of MMP inhibitors with PDT can potentially enhance the clinical effect of PDT, so an interesting direction for future work seems to be the association of PDT with MMP inhibitors tested in glioma [167,168].
17. Glial Fibrillary Acidic Protein
Glial fibrillary acidic protein (GFAP), is an intermediate filament protein occurring in several isoforms, characteristic of astrocytes and neural stem cells and their malignant analogs in glioma [169,170]. The role of GFAP in development in glioma aggressiveness is complex. Cells with a high GFAPδ/α ratio can be highly malignant and more invasive [171]. Moreover, deprivation of any of these isoforms has also been shown to increase the migratory capacity of glioma cells [172]. The effect of PDT on GFAP expression has not yet been precisely established. Namatame et al., reported that PDT with PS talaporfin sodium decreased GFAP expression leading to disappearance of GFAP expression [173]. However, it is difficult to assess the effect of GFAP knockdown on the efficacy of PDT. On the one hand, this effect in the long term may be unfavorable, as GFAP-negative cells have been shown to have a higher average number of nucleolar organizer regions (Ag-NOR), an increase of which has been linked in a couple of papers to a worse prognosis in several cancers, including gliomas [174,175,176,177]. On the other hand, however, there are no reports on the effect of PDT on Ag-NOR, so it is difficult to infer whether a decrease in GFAP by PDT can lead to an increase in Ag-NOR. For obvious reasons, this inference also does not take into account the percentage of GFAP-negative cell death after PDT. Namatame et al. also did not determine the dynamics of changes between GFAP isoforms, which may be crucial in determining the benefit of PDT in this regard [171].
18. Na+ /H+ Exchanger Isoform 1
Na/H exchanger isoform 1 (NHE1) is a membrane Na+/H+ exchanger that maintains the alkaline intracellular pH of glioma cells and the acidic tumor microenvironment (TME) [178,179]. NHE-1 is highly expressed in both glioma cells and tumor-associated microglia cells and macrophages [179,180]. Its activity increases proliferation, invasion and migration of glioma cells and contributes to TME immunosuppression and treatment resistance [179,180,181]. The relationship between PDT, NHE1, and glioma has been little studied to date. It has been established that NHE1 can attenuate the PDT effect of glioma through increased extrusion of H+ [182]. It has been shown that ALA-PDT and HMME-PDT can reduce NHE-1 levels, and this effect is enhanced by associating them with dihydroartemisinin and temozolomide, respectively [182,183]. However, it should be noted that HIF-1α-dependent NHE1 expression is induced by hypoxia, so this issue needs further clarification [184]. An interesting, as yet unexplored approach appears to be the association of PDT with the NHE1 inhibitor 5-benzylglycinyl-amiloride, which has shown anti-tumor activity in glioma [185].
19. Nuclear Factor kB
Nuclear factor kB (NF-kB) comprises a family of transcription factors involved in the regulation of a wide variety of biological responses. NF-kB plays a well-known function in the regulation of the immune response and inflammation, as well as regulating the expression of genes involved in many key processes of oncogenesis [186]. One of the cancers whose NF-kB hyperactivation can promote development, progression and resistance to therapy is glioma [187,188]. NF-kB may have both positive and negative effects on the efficacy of PDT of glioma, however, this issue is not completely clarified. NF-kB has been shown to have pro-apoptotic and anti-necrotic effects after PDT [189]. However, earlier work on other cancers suggested both anti-apoptotic and anti-necrotic effects of NF-kB [190]. Thus, this issue requires further clarification. Moreover, PDT-induced NF-kB can also increase the migration of glioma cells [191]. On the other hand, NF-kB activation is important for the activation of the anti-tumor immune response by PDT [190]. PDT is not indifferent to NF-kB expression, either directly in the glioma or in other tissues. It has been shown that ALA-PDT can lead to both down-regulation of NF-kB and enhanced its activation in glioma cells [189,192,193]. Moreover, low-dose PDT with PS Porfimer sodium induces NF-kB of TNF-a/NF-kB pathway in cerebral vascular endothelial cells, and impairment of their function [194]. This effect may be important for the effectiveness of delivering other therapeutics to the tumor after PDT. Arguably, then, the goal of PDT in terms of its effect on NF-kB should be to modify therapy in such a way as to achieve an optimal, not overly high or low level of NF-kB expression. To achieve this, it is possible to test strategies based on both the selection of an appropriate PDT therapeutic regimen and the association of PDT with NF-kB inhibitors. Coupienne et al. showed that the association of ALA-PDT with an NF-kB inhibitor improves glioma cell death [189]. Work on the development and use of an effective NF-kB inhibitor in the glioma is still ongoing, creating the potential to test new combinations [195,196].
20. Transporter Associated with Antigen Processing 1
Transporter associated with antigen processing 1 (TAP1) is a molecule involved in the processing and presentation of antigens restricted to the major class I tissue compatibility complex, including tumor-associated antigens [197]. TAP1 is involved in tumor immunity and is abnormally expressed in many types of cancer, including glioma, which may be less immunogenic due to low levels of TAP1 expression [198,199,200]. Since it has been shown that high TAP1 expression resulted in better patient response to anti-PD-1, anti-PD-L1, anti-CTLA-4 immunotherapy in several cancers, increasing TAP1 expression presumably could be an attractive form of improving the hitherto unsatisfactory results of immune checkpoint blockade in glioma therapy [197,201]. The work of Zhang et al. showed that PDT may have this ability. They showed that HMME-PDT enables glioma cells to recover both the expression of functional TAP1 and the presentation of MHC class I surface antigens [202]. However, this is the only report in the field so far, so it needs to be confirmed in other works.
21. PD-L1/PD-1
The axis of programmed cell death protein 1 (PD-1) and programmed cell death ligand 1 (PD-L1) plays an important role in inhibiting the host immune response against cancer cells [203]. Studies have shown that the PD-1/PD-L1 pathway plays a key role in glioma progression and the effectiveness of immunotherapy [204]. There is a dearth of work to date describing the effects of PDT on PD-1/PD-L1 expression in tumors, and the available results indicating that PDT can both block and enhance PD-1/PD-L1 axis activity are contradictory [205,206,207]. However, none of the papers cited used glioma cells for the study. Recently, the ability of PDT to induce immunogenic cell death (ICD), activate a tumor-specific immune response, and induce a long-lasting and effective anti-tumor response has been of great interest [208,209,210]. Turubanova in et al. showed that repetitive PDT with PS photosens and photoditazine can achieve this goal [211]. Shibata et al. also induced ICD of glioma cells via liposomally formulated IG conjugated to phospholipids [212]. ICD is also desirable for glioma therapy and can improve the results of immune checkpoint blocker therapy [213,214]. The utility of the potential use of PDT for this purpose is confirmed by the work of Xu et al. They showed that the association of PS Ce6 with αPD-L1 resulted in greater infiltration of immune effector cells, and prolonged life of mice with orthotopic GL261 glioma than PDT or αPD-L1 alone [215]. PDT-induced ICD and downregulation of PD-1 and PD-L1 by deprivation of extracellular cholesterol in glioma TME also induced NP designed by Yin et al. [216]. The combination of PD-1/PD-L1 axis blocking method, PDT and dendritic cell (DC) therapy seems to be an interesting approach. The presence of PD-1 may adversely affect the efficacy of dendritic cells, which dysfunctionally limit antigen-specific T-cell responses in glioma [217,218]. Several works have shown that PDT of glioma with different PS can improve DC functionality [219,220,221,222]. Since anti-PD-1 treatment during DC maturation resulted in increased DC survival, the approach of combining PDT and a method that blocks the PD-1/PD-L1 axis could therefore be an attractive improvement on the hitherto suboptimal results of glioma dendritic cell therapy [217,223].
22. Nitric Oxide Synthase
Nitric oxide (NO), a free gas with numerous biological properties, at concentrations of 50-100 nM can promote glioma growth, neovascularization, immunosuppressive properties of TME and resistance to therapies [224,225]. NO is mainly produced by the oxidation of L-arginine by three isoforms of NO synthase: inducible (iNOS), neuronal (nNOS), and endothelial (eNOS) [226,227,228]. While nNOS is the main synthase in the central nervous system, iNOS is most often associated with cancers, including glioma [226,228,229]. This synthase has been shown to function as a critical regulator of glioma development and is essential for brain tumor stem cell proliferation and tumorigenesis, and its increased expression has been described as a hallmark of chemoresistance in gliomas [229,230]. iNOS also has a negative impact on the efficacy of glioma PDT, increasing resistance, survival and migration of surviving cells [191,231,232,233]. This effect is particularly important because it has been proven that in an NF-kB-dependent manner, PDT can increase iNOS levels in glioma cells by up to fourfold within 24 hours of therapy [231,232,233]. However, the effect of PDT on nNOS levels has not been demonstrated [231]. An investigated proposal to solve this problem is the association of PDT with iNOS inhibitors. It was shown that inhibition of iNOS activity closely matched the inhibition of glioma cell growth and invasion after PDT, and this effect was synergistic when the BET inhibitor (JQ1) was also used [232]. This approach is particularly attractive because in numerous papers, iNOS inhibitors have shown significant potential as treatment options for oncologic lesions and have had a safe toxicity profile in humans for other pathological conditions [234]. On the other hand, higher NO concentrations of >300 nM have been shown to induce a cytotoxic effect on cancer cells, so a potential solution to the adverse effects of PDT on NO synthesis could be the development of a PDT therapeutic regimen characterized by significantly increased NO production [224]. There is still a lack of data on the relationship between glioma PDT and eNOS, and given the importance of vascular effects in PDT efficacy, this issue needs clarification.
23. Glutamate
Glutamate (GLU) is an excitatory neurotransmitter (NS) that plays a central role in glioma malignancy [235,236]. It is produced and released from cells as a by-product of glutathione synthesis [235]. PDT can increase GLU levels. HpD-PDT has been shown to induce a sharp threefold increase in gluatminate levels that persisted for at least 16 hours [237]. GLU induces glioma malignancy through multiple mechanisms, including excitotoxicity to surrounding healthy neurons and paracrine and autocrine effects on glutamine receptors [235,236,238]. It was shown that HpD-PDT increased GluR1 and GluR2 AMPAR subunit expression, Ca2+ influx and apoptosis of C6 glioma cells [237,239]. This effect may be exacerbated by PDT-induced release of glutamate [237]. However, it should be emphasized that the aforementioned work was conducted in vitro, and therefore could not verify the adverse effects of PDT-increased glutamate concentrations on healthy brain tissue surrounding the tumor. This issue requires necessary clarification. Data on the effects of PDT on NMDA receptors are also lacking. GLU inhibitors tested in glioma could presumably be useful in regulating the effects of PDT on glutamate receptors [235].
24. Peripheral-Type Benzodiazepine Receptors
The peripheral-type benzodiazepine receptor (PBR) is a mitochondrial protein that is expressed at high levels in steroid-synthesizing tissues, including brain glial cells [240]. Basal PBR expression is up-regulated in gliomas [241,242]. It has been suggested that the relatively high binding density of peripheral-type benzodiazepine receptors is associated with increased tumorigenicity and rate of cell proliferation [243,244]. It is known that PDT is not indifferent to PBR expression in the glioma. Bisland et al. showed that ALA-PDT with low light levels increases PBR expression in a CNS-1 glioma model [245]. Regulation of PBR expression by PDT has implications for the efficacy of therapy, but there are few reports in this area. A study on mouse leukemia cells showed that the affinity of PpIX-configured PS for PBR can affect the efficacy of PDT [246]. ALA-PDT-induced increase in PBR facilitated increased production of PpIX and enhanced phototoxic effects against glioma cells [245]. The potential beneficial effect of the presence of PBR on the efficacy of glioma PDT was exploited by Sarissky et al. They tested the association of Hy-PDT with diazepam, a non-selective PBR ligand with established activity to inhibit glioma cell proliferation [247,248]. They showed that the combination enhanced cell apoptosis, but only of the U-87 line, with increased PBR expression relative to the U373MG line, which was resistant to the combination. However, they did not describe the effect of Hy-PDT on PBR expression levels [247]. Significantly, the association of diazepam with chemotherapeutics tested to date has resulted in both improved efficacy and reduced efficacy, and glioma cells with high nuclear PBR proliferate in response to PBR ligands [244,249,250]. Thus, further work is needed to precisely describe the relationships between PDT, the PS used and the light dose applied, PBR expression and the ultimate impact on the effectiveness of glioma therapy.
25. Neuropeptide Y Receptors
Neuropeptide Y receptors (NPYRs) are a group of several receptors (Y1R, Y2R and Y5R) that mediate the action of the multifunctional neurotransmitter neuropeptide Y. They are characterized by proliferative effects, but their role has not been completely elucidated [251]. NPYR expression in the glioma varies according to the type of this tumor. WHO grade IV gliomas are characterized by extremely high Y2R expression of the NPY receptor in both frequency and density; WHO grade I to III astrocytomas and oligodendrogliomas also show high Y2R frequency but low density, and medulloblastomas show Y1R and Y2R expression in moderate frequency and density [252]. The presence of NPYR on glioma cells can be effectively used to target gliomas, as demonstrated in several papers [253,254]. This advantage can also be used to improve PS delivery and PDT effects, as demonstrated by Hu et al. by equipping their designed NP with the Y1R ligand [255]. However, there is a lack of work on targeting Y2R, which could be particularly attractive in the treatment of WHO stage IV gliomas [252]. Since NPYRs are characterized by proliferative effects, it is also worth checking the effect of PDT on their expression levels and the relationship between expression and PDT efficacy.
Table 2.
Neurotransmitters in glioma photodynamic therapy. In this table we have collected available information on neurotransmitters described in photodynamic therapy of glioma. The second through fourth columns, respectively, locate data on the effect of PDT on the level of a neurotransmitter, the expression of its receptor, and the effect of a neurotransmitter on the efficacy of PDT. In the fifth column are the neurotransmitter-related approaches tested in PDT of gliomas that are related to the described neurotransmitters.
Table 2.
Neurotransmitters in glioma photodynamic therapy. In this table we have collected available information on neurotransmitters described in photodynamic therapy of glioma. The second through fourth columns, respectively, locate data on the effect of PDT on the level of a neurotransmitter, the expression of its receptor, and the effect of a neurotransmitter on the efficacy of PDT. In the fifth column are the neurotransmitter-related approaches tested in PDT of gliomas that are related to the described neurotransmitters.
| Neurotransmitter | NS levels | Receptor expression | Impact on the effectiveness of PDT | Tested therapeutic approach |
|---|---|---|---|---|
| NO | Increase [231,232,233] | No data available | Increased resistance, survival and migration of glioma cells after PDT [191,231,232,233] | Association of PDT with iNOS inhibitors reduces NO levels and glioma cell growth and invasion after PDT [232] |
| Glutaminian | Increase [237] | Increase in AMPAR expression [237,239] | Increase in apoptosis of glioma cells [237,239] | No data available |
| GABA | No data available | Increase in PBR expression [245] | Increased production of PpIX and phototoxic effect against glioma cells (via increase in PBR) [245] | Association of PDT with diazepam enhances apoptosis of glioma cells [247] |
| Neuropeptide Y | No data available | No data available | No data available | NP coupling with NPY improves targeting and PS delivery in the glioma [255] |
26. Receptor-Interacting Serine/Threonine Kinases
Receptor-interacting protein kinases 1 (RIP1) and 3 (RIP3) are regulators of programmed necrosis [256]. The exact function of the RIP1/RIP3 pathway in cancer development has not been elucidated, and both stimulation and inhibition of this pathway have been proposed for anti-cancer therapy [257]. Several of the compounds tested in glioma therapy have been shown to induce the RIP1/RIP3 pathway to induce necroptosis of its cells [258,259,260]. RIP1/RIP3 complex regulates programmed glioma necrosis even after high dose of radiation [261]. The effect of PDT on RIP1/RIP3 is not established. Coupienne et al. showed that ALA-PDT of glioblastoma cells activates the RIP3-dependent necrotic pathway, and the resulting necrosome is composed of RIP1, RIP3, and other unrecognized stubs [262]. Fettweis et al. characterized the RIP3/TSC2 complex, suggesting that RIP3 promotes glioma cell death by targeting the TSC2-dependent survival pathway after ALA-PDT [263]. Since work on drugs affecting the RIP1/RIP3 pathway is already underway, a thorough elucidation of its role in glioma PDT is essential for their successful association [257].
27. Low-Density Lipoprotein Receptor
Low-density lipoprotein receptor (LDLR) is highly expressed in blood brain barrier and glioma cells, while normal brain tissues and neurons have relatively low levels of it [264,265]. Previous work has shown that targeting LDLRs may be a promising strategy for improving drug delivery in glioma therapy [266]. This approach was also tested in PDT, associating LDL with hydrophobic PS [267,268,269,270]. PDT-induced phototoxicity of glioma cells with berberine and hypericin PSs was correspondingly higher when combined with LDL than when used alone [269,270]. However, the above results were described in vitro under conditions that do not test the ability to cross the BBB, so they would need to be confirmed in vivo. Since the efficacy of PS-LDL association has been demonstrated in vivo in subcutaneously implanted HepG2 cells in nude mice, and targeting LDLR is a promising strategy for overcoming BBB limitations, this line of future research seems particularly warranted [266,271].
28. GLUT Glucose Transporters
Two transporters are involved in glucose transport into cells. The first, GLUT mediates facilitated passive transport, while the second, SGLT, mediates active transport. The expression of GLUT and SGLT is tissue-dependent and depends on its role in carbohydrate metabolism [272,273]. The complex role of GLUTs in the glioma has not yet been completely elucidated. Specific functions are known to vary for different types of GLUTs, but it has been established that GLUTs may be responsible for proliferation, invasion of the tumor [274,275,276]. Liu et al. found that GLUT-3 expression in glioma was significantly correlated with the pathological grade of the tumor [274]. The effect of PDT on GLUT levels in the glioma is unclear, and available data are conflicting. Li et al. report that HMME-PDT inhibits the expression of GLUT-1 [59]. On the other hand, Ma et al. show that the expression of GLUT-1 and GLUT-3 was significantly increased after ALA-PDT [62]. This effect is presumably dependent on HIF-1a. In the first study, PDT led to a decrease in it, while in the second, an increase was observed. This is consistent with previous findings on the effect of HIF-1a on GLUT-1 expression [275]. In view of this, regulation of HIF-1a expression may be an attractive way to reduce GLUT expression. Because of their effect on GLUT tumor growth, they can be used to improve the effects of PDT. Zhang et al. achieved this by synthesizing a novel PS that effectively targeted the GLUT/hexokinase pathway [277]. Moreover, by modifying drugs with glucose, GLUTs can also be used to achieve improved crossing of biological barriers and targeted delivery of therapeutics [278]. This property has also been successfully exploited in association with PS, but not in glioma therapy, where the association of PS with other sugars has been tested [279,280]. Targeting GLUT, however, must be done with caution. Indeed, it has been shown that inhibition of GLUT-1 can lead to reduced intracellular glucose uptake, which would be particularly detrimental in the highly metabolically active malignant glioma [275].
29. Other Molecules and Pathways
In this section, we present the remaining isolated reports on PDT and molecular approaches in the glioma. Several papers point to potential molecules that improve PS delivery. Akcker et al. showed that the association of PS with an antibody targeting the additional A domain of fibronectin leads to microvascular dysfunction and reduced glioma growth within the first 48 hours after treatment with complete recovery 5 days after treatment [281]. De Groof indicate that improved PS selectivity can be achieved by coupling PS with NPs targeting a US28 protein [282]. This function can also be performed by nucleolin and the receptor for transferrin [283]. Akhlynina et al. showed that nuclear targeting of photosensitizing conjugates by incorporating a large-tumor antigen nuclear localization signal results in enhanced photodynamic activity than with free Ce6 [284]. Other work presents further molecules that affect PDT efficacy. Wu et al. report that the presence of intercellular communication through Connexin Cx43 improves the phototoxicity of PDT gliomas [285]. An et al. demonstrated that PDT with PS Sinoporphyrin sodium reduces protein phosphorylation of the PI3K/AKT/mTOR signaling pathway [286]. Lee et al. indicate that a tumor protein (TP53) is involved in mediating Ph-PDT resistance by binding to the ALKBH2 promoter [287]. Park et al. suggest that the key molecule responsible for glioma PDT resistance is C5α [288]. Higher APE1 endonuclease activity and increased expression and activation of the DNA damage kinase ATM may also serve these functions [289]. The effect of PDT on the immune response is highlighted by Li et al. demonstrating that glioma PDT also induces the release of inflammatory factors such as TNF-α and IFN-γ [290].
Author Contributions
Conceptualization, D.A.; P.W.;and D.B.- A.; methodology, D.A.; P.W.; and D.B.- A.; software, D.A.; P.W.; and D.B.- A.; validation, D.A.; P.W.; and D.B.- A.; formal analysis, D.A.; P.W.; and D.B.- A.; investigation, D.A.; P.W.; and D.B.- A.; resources, D.A.; P.W.; and D.B.- A.; data curation, D.A.; P.W.; and D.B.- A.; writing—original draft preparation, D.A.; P.W.; and D.B.- A.; writing—review and editing, D.A.; P.W.; and D.B.- A.; visualization, D.A.; P.W.; and D.B.- A.; supervision, D.A.; funding acquisition, D.A.;. All authors have read and agreed to the published version of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schwartz, S.M. Epidemiology of Cancer. Clinical Chemistry 2024, 70, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Bray, F.; Laversanne, M.; Weiderpass, E.; Soerjomataram, I. The ever-increasing importance of cancer as a leading cause of premature death worldwide. Cancer. 2021, 127, 3029–3030. [Google Scholar] [CrossRef] [PubMed]
- Vigneswaran, K.; Neill, S.; Hadjipanayis, C.G. Beyond the World Health Organization grading of infiltrating gliomas: Advances in the molecular genetics of glioma classification. Ann Transl Med. 2015, 3, 95. [Google Scholar] [PubMed]
- Davis, M.E. Epidemiology and Overview of Gliomas. Semin Oncol Nurs. 2018, 34, 420–429. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.M.; Englander, Z.K.; Miller, M.L.; Bruce, J.N. Malignant Glioma. Adv Exp Med Biol. 2023, 1405, 1–30. [Google Scholar] [PubMed]
- Jakola, A.S.; Myrmel, K.S.; Kloster, R.; Torp, S.H.; Lindal, S.; Unsgård, G.; Solheim, O. Comparison of a strategy favoring early surgical resection vs a strategy favoring watchful waiting in low-grade gliomas. JAMA. 2012, 308, 1881–1888. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; Di Meco, F.; Lieberman, F.; Zhu, J.J.; Stragliotto, G.; Tran, D.; Brem, S.; Hottinger, A.; Kirson, E.D.; Lavy-Shahaf, G.; Weinberg, U.; Kim, C.Y.; Paek, S.H.; Nicholas, G.; Bruna, J.; Hirte, H.; Weller, M.; Palti, Y.; Hegi, M.E.; Ram, Z. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs Maintenance Temozolomide Alone on Survival in Patients With Glioblastoma: A Randomized Clinical Trial. JAMA. 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Wee, C.W. Treatment of Adult Gliomas: A Current Update. Brain Neurorehabil. 2022, 15, e24. [Google Scholar] [CrossRef] [PubMed]
- Yang, K.; Wu, Z.; Zhang, H.; Zhang, N.; Wu, W.; Wang, Z.; Dai, Z.; Zhang, X.; Zhang, L.; Peng, Y.; Ye, W.; Zeng, W.; Liu, Z.; Cheng, Q. Glioma targeted therapy: Insight into future of molecular approaches. Mol Cancer. 2022, 21, 39. [Google Scholar] [CrossRef]
- Bush, N.A.; Chang, S.M.; Berger, M.S. Current and future strategies for treatment of glioma. Neurosurg Rev. 2017, 40, 1–14. [Google Scholar] [CrossRef]
- Vermandel, M.; et al. Standardized intraoperative 5-ALA photodynamic therapy for newly diagnosed glioblastoma patients: A preliminary analysis of the INDYGO clinical trial. Journal of neuro-oncology 2021, 152, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kędzierska, E.; Knap-Czop, K.; Kotlińska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic therapy - mechanisms, photosensitizers and combinations. Biomed Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Mansoori, B.; Mohammadi, A.; Amin Doustvandi, M.; Mohammadnejad, F.; Kamari, F.; Gjerstorff, M.F.; Baradaran, B.; Hamblin, M.R. Photodynamic therapy for cancer: Role of natural products. Photodiagnosis Photodyn Ther. 2019, 26, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Aebisher, D.; Przygórzewska, A.; Myśliwiec, A.; Dynarowicz, K.; Krupka-Olek, M.; Bożek, A.; Kawczyk-Krupka, A.; Bartusik-Aebisher, D. Current Photodynamic Therapy for Glioma Treatment: An Update. Biomedicines 2024, 12, 375. [Google Scholar] [CrossRef] [PubMed]
- Kukal, S.; Guin, D.; Rawat, C.; Bora, S.; Mishra, M.K.; Sharma, P.; Paul, P.R.; Kanojia, N.; Grewal, G.K.; Kukreti, S.; Saso, L.; Kukreti, R. Multidrug efflux transporter ABCG2: Expression and regulation. Cell Mol Life Sci. 2021, 78, 6887–6939. [Google Scholar] [CrossRef] [PubMed]
- Morgan, J.; Petrucci, C.M. The effect of ALA/PpIX PDT on putative cancer stem cells in tumor side populations. Kessel D, editor. 12TH WORLD CONGRESS OF THE INTERNATIONAL PHOTODYNAMIC ASSOCIATION: PHOTODYNAMIC THERAPY: BACK TO THE FUTURE. 1000 20TH ST, PO BOX 10, BELLINGHAM, WA 98227-0010 USA: SPIE-INT SOC OPTICAL ENGINEERING; 2009. (Proceedings of SPIE; vol. 7380).
- Mueller, P.; Gaber, S.A.A.; Zimmermann, W.; Wittig, R.; Stepp, H. ABCG2 influence on the efficiency of photodynamic therapy in glioblastoma cells. Vol. 210, JOURNAL OF PHOTOCHEMISTRY AND PHOTOBIOLOGY B-BIOLOGY. PO BOX 564, 1001 LAUSANNE, SWITZERLAND: ELSEVIER SCIENCE SA; 2020.
- Pan, L.; Lin, H.; Tian, S.; Bai, D.; Kong, Y.; Yu, L. The Sensitivity of Glioma Cells to Pyropheophorbide-αMethyl Ester-Mediated Photodynamic Therapy Is Enhanced by Inhibiting ABCG2. Vol. 49, LASERS IN SURGERY AND MEDICINE. 111 RIVER ST, HOBOKEN 07030-5774, NJ USA: WILEY; 2017. p. 719–26.
- Weidner, L.D.; Zoghbi, S.S.; Lu, S.; Shukla, S.; Ambudkar, S.V.; Pike, V.W.; Mulder, J.; Gottesman, M.M.; Innis, R.B.; Hall, M.D. The Inhibitor Ko143 Is Not Specific for ABCG2. J Pharmacol Exp Ther. 2015, 354, 384–393. [Google Scholar] [CrossRef] [PubMed]
- Wang W, Tabu K, Hagiya Y, Sugiyama Y, Kokubu Y, Murota Y, Ogura S ichiro, Taga T. Enhancement of 5-aminolevulinic acid-based fluorescence detection of side population-defined glioma stem cells by iron chelation. vol. 7, scientific reports. macmillan building, 4 crinan st, london n1 9xw, england: Nature publishing group; 2017.
- Abdel Gaber, S.A.; Müller, P.; Zimmermann, W.; Hüttenberger, D.; Wittig, R.; Abdel Kader, M.H.; Stepp, H. ABCG2-mediated suppression of chlorin e6 accumulation and photodynamic therapy efficiency in glioblastoma cell lines can be reversed by KO143. J Photochem Photobiol B. 2018, 178, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Hamid, S.A.; Zimmermann, W.; Huettenberger, D.; Wittig, R.; Kader, M.A.; Stepp, H. In vitro Study for Photodynamic Therapy using Fotolon® in Glioma Treatment. Lilge L, Sroka R, editors. MEDICAL LASER APPLICATIONS AND LASER-TISSUE INTERACTIONS VII. 1000 20TH ST, PO BOX 10, BELLINGHAM, WA 98227-0010 USA: SPIE-INT SOC OPTICAL ENGINEERING; 2015. (Proceedings of SPIE; vol. 9542).
- Selbo, P.K.; Weyergang, A.; Eng, M.S.; Bostad, M.; Maelandsmo, G.M.; Hogset, A.; Berg, K. Strongly amphiphilic photosensitizers are not substrates of the cancer stem cell marker ABCG2 and provides specific and efficient light-triggered drug delivery of an EGFR-targeted cytotoxic drug. Vol. 159, JOURNAL OF CONTROLLED RELEASE. RADARWEG 29, 1043 NX AMSTERDAM, NETHERLANDS: ELSEVIER; 2012. p. 197–203.
- Sun, W.; Kajimoto, Y.; Inoue, H.; Miyatake, S.I.; Ishikawa, T.; Kuroiwa, T. Gefitinib enhances the efficacy of photodynamic therapy using 5-aminolevulinic acid in malignant brain tumor cells. Vol. 10, PHOTODIAGNOSIS AND PHOTODYNAMIC THERAPY. RADARWEG 29, 1043 NX AMSTERDAM, NETHERLANDS: ELSEVIER; 2013. p. 42–50.
- Mansi, M.; Howley, R.; Chandratre, S.; Chen, B. Inhibition of ABCG2 transporter by lapatinib enhances 5-aminolevulinic acid-mediated protoporphyrin IX fluorescence and photodynamic therapy response in human glioma cell lines. Vol. 200, BIOCHEMICAL PHARMACOLOGY. THE BOULEVARD, LANGFORD LANE, KIDLINGTON, OXFORD OX5 1GB, ENGLAND: PERGAMON-ELSEVIER SCIENCE LTD; 2022.
- Bansal, A.; Simon, M.C. Glutathione metabolism in cancer progression and treatment resistance. J Cell Biol. 2018, 217, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Du, S.; Du, Y.; Ren, J.; Ying, G.; Yan, Z. Glutathione reductase mediates drug resistance in glioblastoma cells by regulating redox homeostasis. J Neurochem. 2018, 144, 93–104. [Google Scholar] [CrossRef]
- Edano, M.; Kanda, T.; Tarumoto, R.; Hamamoto, W.; Hasegawa, T.; Mae, Y.; Onoyama, T.; Takata, T.; Sugihara, T.; Isomoto, H. Intracellular glutathione levels affect the outcomes of verteporfin-mediated photodynamic therapy in esophageal cancer cells. Photodiagnosis Photodyn Ther. 2022, 40, 103090. [Google Scholar] [CrossRef]
- Mastrangelopoulou, M.; Grigalavicius, M.; Raabe, T.H.; Skarpen, E.; Juzenas, P.; Peng, Q.; Berg, K.; Theodossiou, T.A. Predictive biomarkers for 5-ALA-PDT can lead to personalized treatments and overcome tumor-specific resistances. Cancer Rep (Hoboken). 2022, 5, e1278. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Robin, A.M.; Katakowski, M.; Tong, L.; Espiritu, M.; Singh, G.; Chopp, M. Photodynamic therapy with photofrin in combination with Buthionine Sulfoximine (BSO) of human glioma in the nude rat. Lasers Med Sci. 2003, 18, 128–133. [Google Scholar] [CrossRef] [PubMed]
- dos Reis Oliveira, C.; Pereira, J.C.; Barros Ibiapina, A.; Roseno Martins, I.R.; de Castro e Sousa, J.M.; Ferreira, P.M.P.; Carneiro da Silva, F.C. Buthionine sulfoximine and chemoresistance in cancer treatments: A systematic review with meta-analysis of preclinical studies. Journal of Toxicology and Environmental Health, Part B 2023, 26, 417–441. [Google Scholar] [CrossRef] [PubMed]
- Hwang, B.; Kim, T.I.; Kim, H.; Jeon, S.; Choi, Y.; Kim, Y. Ubiquinone-BODIPY nanoparticles for tumor redox-responsive fluorescence imaging and photodynamic activity. J Mater Chem B. 2021, 9, 824–831. [Google Scholar] [CrossRef] [PubMed]
- An, R.; Liu, L.; Wei, S.; Huang, Z.; Qiu, L.; Lin, J.; Liu, H.; Ye, D. Controlling Disassembly of Paramagnetic Prodrug and Photosensitizer Nanoassemblies for On-Demand Orthotopic Glioma Theranostics. ACS Nano. 2022, 16, 20607–20621. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-Y.; Ye, Y.-X.; Zhang, Q.; Kang, Q.-J.; Xu, Z.-M.; Ren, S.-Z.; Lin, F.; Duan, Y.-T.; Xu, H.-J.; Hu, Z.-Y.; Yang, S.-S.; Zhu, H.-L.; Zou, M.-J.; Wang, Z.-C. , Multifunctional Protein Hybrid Nanoplatform for Synergetic Photodynamic-Chemotherapy of Malignant Carcinoma by Homologous Targeting Combined with Oxygen Transport. Adv. Sci. 2023, 10, 2203742. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Li, S.; Chen, W.; Lu, H.; Ye, L.; Min, Z.; Sun, S.; Teng, C.; Yin, H.; Zhang, Q.; He, W.; Wang, X.; Lv, W.; Lv, L.; Xin, H. Multifunctional Hybrid Hydrogel System Enhanced the Therapeutic Efficacy of Treatments for Postoperative Glioma. ACS Appl Mater Interfaces. 2022, 14, 27623–27633. [Google Scholar] [CrossRef]
- Obi, C.D.; Bhuiyan, T.; Dailey, H.A.; Medlock, A.E. Ferrochelatase: Mapping the Intersection of Iron and Porphyrin Metabolism in the Mitochondria. Front Cell Dev Biol. 2022, 10, 894591. [Google Scholar] [CrossRef] [PubMed]
- Briel-Pump, A.; Beez, T.; Ebbert, L.; Remke, M.; Weinhold, S.; Sabel, M.C.; Sorg, R.V. Accumulation of protoporphyrin IX in medulloblastoma cell lines and sensitivity to subsequent photodynamic treatment. J Photochem Photobiol B. 2018, 189, 298–305. [Google Scholar] [CrossRef]
- Ihata, T.; Nonoguchi, N.; Fujishiro, T.; Omura, N.; Kawabata, S.; Kajimoto, Y.; Wanibuchi, M. The effect of hypoxia on photodynamic therapy with 5-aminolevulinic acid in malignant gliomas. Photodiagnosis Photodyn Ther. 2022, 40, 103056. [Google Scholar] [CrossRef]
- Reburn, C.; Anayo, L.; Magnussen, A.; Perry, A.; Wood, M.; Curnow, A. Experimental findings utilising a new iron chelating ALA prodrug to enhance protoporphyrin IX-induced photodynamic therapy. Hasan T, editor. 17TH INTERNATIONAL PHOTODYNAMIC ASSOCIATION WORLD CONGRESS. 1000 20TH ST, PO BOX 10, BELLINGHAM, WA 98227-0010 USA: SPIE-INT SOC OPTICAL ENGINEERING; 2019. (Proceedings of SPIE; vol. 11070).
- Blake, E.; Curnow, A. The hydroxypyridinone iron chelator CP94 can enhance PpIX-induced PDT of cultured human glioma cells. Photochem Photobiol. 2010, 86, 1154–1160. [Google Scholar] [CrossRef] [PubMed]
- Blake, E.; Allen, J.; Curnow, A. An in vitro comparison of the effects of the iron-chelating agents, CP94 and dexrazoxane, on protoporphyrin IX accumulation for photodynamic therapy and/or fluorescence guided resection. Photochem Photobiol. 2011, 87, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Teng, L.; Nakada, M.; Zhao, S.G.; Endo, Y.; Furuyama, N.; Nambu, E.; Pyko, I.V.; Hayashi, Y.; Hamada, J.I. Silencing of ferrochelatase enhances 5-aminolevulinic acid-based fluorescence and photodynamic therapy efficacy. Br J Cancer. 2011, 104, 798–807. [Google Scholar] [CrossRef] [PubMed]
- Chau, L.Y. Heme oxygenase-1: Emerging target of cancer therapy. J Biomed Sci. 2015, 22, 22. [Google Scholar] [CrossRef] [PubMed]
- Kyriakis, J.M.; Avruch, J. Mammalian mitogen-activated protein kinase signal transduction pathways activated by stress and inflammation. Physiol Rev. 2001, 81, 807–869. [Google Scholar] [CrossRef] [PubMed]
- Sigaud, R.; Rösch, L.; Gatzweiler, C.; Benzel, J.; von Soosten, L.; Peterziel, H.; Selt, F.; Najafi, S.; Ayhan, S.; Gerloff, X.F.; Hofmann, N.; Büdenbender, I.; Schmitt, L.; Foerster, K.I.; Burhenne, J.; Haefeli, W.E.; Korshunov, A.; Sahm, F.; van Tilburg, C.M.; Jones, D.T.W.; Pfister, S.M.; Knoerzer, D.; Kreider, B.L.; Sauter, M.; Pajtler, K.W.; Zuckermann, M.; Oehme, I.; Witt, O.; Milde, T. The first-in-class ERK inhibitor ulixertinib shows promising activity in mitogen-activated protein kinase (MAPK)-driven pediatric low-grade glioma models. Neuro Oncol. 2023, 25, 566–579. [Google Scholar] [CrossRef] [PubMed]
- Haller, V.; Nahidino, P.; Forster, M.; Laufer, S.A. An updated patent review of p38 MAP kinase inhibitors (2014-2019). Expert Opin Ther Pat. 2020, 30, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Chédeville, A.L.; Madureira, P.A. The Role of Hypoxia in Glioblastoma Radiotherapy Resistance. Cancers (Basel). 2021, 13, 542. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, A.; Carico, E. Role of HIF-1 in Cancer Progression: Novel Insights. A Review. Curr Mol Med. 2018, 18, 343–351. [Google Scholar] [CrossRef]
- Albert, I.; Hefti, M.; Luginbuehl, V. Physiological oxygen concentration alters glioma cell malignancy and responsiveness to photodynamic therapy in vitro. Neurol Res. 2014, 36, 1001–1010. [Google Scholar] [CrossRef]
- Broekgaarden, M.; Weijer, R.; van Gulik, T.M.; Hamblin, M.R.; Heger, M. Tumor cell survival pathways activated by photodynamic therapy: A molecular basis for pharmacological inhibition strategies. Cancer Metastasis Rev. 2015, 34, 643–690. [Google Scholar] [CrossRef] [PubMed]
- Lamberti, M.J.; Pansa, M.F.; Vera, R.E.; Fernández-Zapico, M.E.; Rumie Vittar, N.B.; Rivarola, V.A. Transcriptional activation of HIF-1 by a ROS-ERK axis underlies the resistance to photodynamic therapy. PLoS ONE. 2017, 12, e0177801. [Google Scholar] [CrossRef] [PubMed]
- Weijer, R.; Broekgaarden, M.; van Golen, R.F.; Bulle, E.; Nieuwenhuis, E.; Jongejan, A.; Moerland, P.D.; van Kampen, A.H.; van Gulik, T.M.; Heger, M. Low-power photodynamic therapy induces survival signaling in perihilar cholangiocarcinoma cells. BMC Cancer. 2015, 15, 1014. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Q.; Yue, W.; Hu, S. Effect of photodynamic therapy and endostatin on human glioma xenografts in nude mice. Vol. 8, PHOTODIAGNOSIS AND PHOTODYNAMIC THERAPY. RADARWEG 29, 1043 NX AMSTERDAM, NETHERLANDS: ELSEVIER; 2011. p. 314–20.
- Sun, Y.; Zhang, X.; Liu, W. Effect of low energy ALA-PDT on angiogenesis and glioma growth in brain. Zhongguo Jiguang/Chinese Journal of Lasers [Internet]. 2012;39(8).
- Zheng, X.; Jiang, F.; Katakowski, M.; Zhang, X.; Jiang, H.; Zhang, Z.G.; Chopp, M. Sensitization of cerebral tissue in nude mice with photodynamic therapy induces ADAM17/TACE and promotes glioma cell invasion. Cancer Lett. 2008, 265, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Ji, Z.; Yang, G.; Shahzidi, S.; Tkacz-Stachowska, K.; Suo, Z.; Nesland, J.M.; Peng, Q. Induction of hypoxia-inducible factor-1alpha overexpression by cobalt chloride enhances cellular resistance to photodynamic therapy. Cancer Lett. 2006, 244, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.E.; Catrinacio, C.; Ropolo, A.; Rivarola, V.A.; Vaccaro, M.I. A novel HIF-1α/VMP1-autophagic pathway induces resistance to photodynamic therapy in colon cancer cells. Photochem Photobiol Sci. 2017, 16, 1631–1642. [Google Scholar] [CrossRef] [PubMed]
- Wan, Y.; Fu, L.H.; Li, C.; Lin, J.; Huang, P. Conquering the Hypoxia Limitation for Photodynamic Therapy. Adv Mater. 2021, 33, e2103978. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, D.; Zhang, Z.; Wang, Y.; Zhang, Z.; Liang, Z.; Liu, F.; Chen, L. Photodynamic therapy enhances the cytotoxicity of temozolomide against glioblastoma via reprogramming anaerobic glycolysis. Photodiagnosis Photodyn Ther. 2023, 42, 103342. [Google Scholar] [CrossRef]
- Xu, D.; Chen, X.; Peng, Y.; Chen, W.; Li, Y.; Wang, X.; Bui, B.; Chen, K.; Zhou, M.; Kawai, N.; Tamiya, T. Effects of Phthalocyanine-Based Molecular Beacon-Mediated Photodynamic Therapy on U251 Cells. Vol. 8, NANOSCIENCE AND NANOTECHNOLOGY LETTERS. 26650 THE OLD RD, STE 208, VALENCIA, CA 91381-0751 USA: AMER SCIENTIFIC PUBLISHERS; 2016. p. 714–22.
- Cao, X.; Liu, Q.; Adu-Frimpong, M.; Shi, W.; Liu, K.; Deng, T.; Yuan, H.; Weng, X.; Gao, Y.; Yu, Q.; Deng, W.; Yu, J.; Wang, Q.; Xiao, G.; Xu, X. Microfluidic Generation of Near-Infrared Photothermal Vitexin/ICG Liposome with Amplified Photodynamic Therapy. AAPS PharmSciTech. 2023, 24, 82. [Google Scholar] [CrossRef]
- Ma, S.; Wang, F.; Dong, J.; Wang, N.; Tao, S.; Du, J.; Hu, S. Inhibition of hypoxia-inducible factor 1 by acriflavine renders glioblastoma sensitive for photodynamic therapy. J Photochem Photobiol B. 2022, 234, 112537. [Google Scholar] [CrossRef]
- Lv, Z.; Jin, L.; Gao, W.; Cao, Y.; Zhang, H.; Xue, D.; Yin, N.; Zhang, T.; Wang, Y.; Zhang, H. Novel YOF-Based Theranostic Agents with a Cascade Effect for NIR-II Fluorescence Imaging and Synergistic Starvation/Photodynamic Therapy of Orthotopic Gliomas. ACS Appl Mater Interfaces. 2022, 14, 30523–30532. [Google Scholar] [CrossRef] [PubMed]
- Song, R.; Hu, D.; Chung, H.Y.; Sheng, Z.; Yao, S. Lipid-Polymer Bilaminar Oxygen Nanobubbles for Enhanced Photodynamic Therapy of Cancer. ACS Appl Mater Interfaces. 2018, 10, 36805–36813. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, J.M.; Yu, H.; Deng, K.; Zhou, W.; Wang, C.X.; Zhang, Y.; Li, K.H.; Zhuo, R.X.; Huang, S.W. Fluorinated polymeric micelles to overcome hypoxia and enhance photodynamic cancer therapy. Vol. 6, BIOMATERIALS SCIENCE. THOMAS GRAHAM HOUSE, SCIENCE PARK, MILTON RD, CAMBRIDGE CB4 0WF, CAMBS, ENGLAND: ROYAL SOC CHEMISTRY; 2018. p. 3096–107.
- Xu, H.; Han, Y.; Zhao, G.; Zhang, L.; Zhao, Z.; Wang, Z.; Zhao, L.; Hua, L.; Naveena, K.; Lu, J.; Yu, R.; Liu, H. Hypoxia-Responsive Lipid-Polymer Nanoparticle-Combined Imaging-Guided Surgery and Multitherapy Strategies for Glioma. Vol. 12, ACS APPLIED MATERIALS & INTERFACES. 1155 16TH ST, NW, WASHINGTON, DC 20036 USA: AMER CHEMICAL SOC; 2020. p. 52319–28.
- Rroji, O.; Kumar, A.; Karuppagounder, S.S.; Ratan, R.R. Epigenetic regulators of neuronal ferroptosis identify novel therapeutics for neurological diseases: HDACs, transglutaminases, and HIF prolyl hydroxylases. Neurobiol Dis. 2021, 147, 105145. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Li, Y.; Li, N.; Shen, L.; Li, Z. Comprehensive analysis of histone deacetylases genes in the prognosis and immune infiltration of glioma patients. Aging (Albany NY). 2022, 14, 4050–4068. [Google Scholar] [CrossRef] [PubMed]
- Li, P.T.; Tsai, Y.J.; Lee, M.J.; Chen, C.T. Increased Histone Deacetylase Activity Involved in the Suppressed Invasion of Cancer Cells Survived from ALA-Mediated Photodynamic Treatment. Int J Mol Sci. 2015, 16, 23994–24010. [Google Scholar] [CrossRef] [PubMed]
- Demyanenko, S.V.; Uzdensky, A.B.; Sharifulina, S.A.; Lapteva, T.O.; Polyakova, L.P. PDT-induced epigenetic changes in the mouse cerebral cortex: A protein microarray study. Biochim Biophys Acta. 2014, 1840, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhang, M.; Zhou, Y.; Guo, W.; Yi, M.; Zhang, Z.; Ding, Y.; Wang, Y. The application of histone deacetylases inhibitors in glioblastoma. J Exp Clin Cancer Res. 2020, 39, 138. [Google Scholar] [CrossRef] [PubMed]
- Sawa, H.; Murakami, H.; Ohshima, Y.; Sugino, T.; Nakajyo, T.; Kisanuki, T.; Tamura, Y.; Satone, A.; Ide, W.; Hashimoto, I.; Kamada, H. Histone deacetylase inhibitors such as sodium butyrate and trichostatin A induce apoptosis through an increase of the bcl-2-related protein Bad. Brain Tumor Pathol. 2001, 18, 109–114. [Google Scholar] [CrossRef]
- Sawa, H.; Murakami, H.; Ohshima, Y.; Murakami, M.; Yamazaki, I.; Tamura, Y.; Mima, T.; Satone, A.; Ide, W.; Hashimoto, I.; Kamada, H. Histone deacetylase inhibitors such as sodium butyrate and trichostatin A inhibit vascular endothelial growth factor (VEGF) secretion from human glioblastoma cells. Brain Tumor Pathol. 2002, 19, 77–81. [Google Scholar] [CrossRef]
- Nör, C.; Sassi, F.A.; de Farias, C.B.; Schwartsmann, G.; Abujamra, A.L.; Lenz, G.; Brunetto, A.L.; Roesler, R. The histone deacetylase inhibitor sodium butyrate promotes cell death and differentiation and reduces neurosphere formation in human medulloblastoma cells. Mol Neurobiol. 2013, 48, 533–543. [Google Scholar] [CrossRef]
- Bueno-Carrazco, J.; Castro-Leyva, V.; García-Gomez, F.; Solís-Paredes, M.; Ramon-Gallegos, E.; Cruz-Orea, A.; Eguía-Aguilar, P.; Arenas-Huertero, F. Sodium butyrate increases the effect of the photodynamic therapy: A mechanism that involves modulation of gene expression and differentiation in astrocytoma cells. Childs Nerv Syst. 2012, 28, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Flores-Ancona, R.M.; García-Gómez, F.Y.; Jiménez-Betanzos, A.M.; Solis-Paredes, M.; Castro-Leyva, V.; Cruz-Orea, A.; Arenas-Huertero, F.; Ramón-Gallegos, E. Effects of sodium butyrate on cell death induced by photodynamic therapy in U373-MG and D54-MG astrocytoma cell lines. Photochem Photobiol. 2009, 85, 1182–1188. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Huang, C.; Zhang, J.; Chen, Q.; Liu, Z.; Ren, X.; Gan, S.; Wu, P.; Wang, D.; Tang, B.Z.; Sun, H. HDAC6-Activatable Multifunctional Near-Infrared Probe for Glioma Cell Detection and Elimination. Anal Chem. 2024, 96, 2406–2414. [Google Scholar] [CrossRef] [PubMed]
- do Carmo, A.; Balça-Silva, J.; Matias, D.; Lopes, M.C. PKC signaling in glioblastoma. Cancer Biol Ther. 2013, 14, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Couldwell, W.T.; Uhm, J.H.; Antel, J.P.; Yong, V.W. Enhanced protein kinase C activity correlates with the growth rate of malignant gliomas in vitro. Neurosurgery. 1991, 29, 880–886. [Google Scholar] [CrossRef]
- Couldwell, W.T.; Antel, J.P.; Yong, V.W. Protein kinase C activity correlates with the growth rate of malignant gliomas: Part II. Effects of glioma mitogens and modulators of protein kinase C. Neurosurgery. 1992, 31, 717–724. [Google Scholar] [CrossRef]
- Huntosova, V.; Stroffekova, K. Hypericin in the Dark: Foe or Ally in Photodynamic Therapy? Cancers 2016, 8, 93. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, I.; Nakanishi, S.; Kobayashi, E.; Nakano, H.; Suzuki, K.; Tamaoki, T. Hypericin and pseudohypericin specifically inhibit protein kinase C: Possible relation to their antiretroviral activity. Biochem Biophys Res Commun. 1989, 165, 1207–1212. [Google Scholar] [CrossRef]
- Uzdensky, A.; Kristiansen, B.; Moan, J.; Juzeniene, A. Dynamics of signaling, cytoskeleton and cell cycle regulation proteins in glioblastoma cells after sub-lethal photodynamic treatment: Antibody microarray study. Biochim Biophys Acta. 2012, 1820, 795–803. [Google Scholar] [CrossRef]
- Dzurová, L.; Petrovajova, D.; Nadova, Z.; Huntosova, V.; Miskovsky, P.; Stroffekova, K. The role of anti-apoptotic protein kinase Cα in response to hypericin photodynamic therapy in U-87 MG cells. Photodiagnosis Photodyn Ther. 2014, 11, 213–226. [Google Scholar] [CrossRef]
- Pevna, V.; Wagnières, G.; Huntosova, V. Autophagy and Apoptosis Induced in U87 MG Glioblastoma Cells by Hypericin-Mediated Photodynamic Therapy Can Be Photobiomodulated with 808 nm Light. Biomedicines. 2021, 9, 1703. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Cho, K.K.; Mikkelse, T.; Tong, L.; Lew, Y.S.; Hochbaum, N.; Shargorodsky, J.; Chop, M. Tamoxifen increases photodynamic therapeutic response of U87 and U25ln human glioma cells. J Neurooncol. 2002, 56, 51–58. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Liu, R.; Yang, S.H.; Yuan, F. Chemotherapeutic effect of tamoxifen on temozolomide-resistant gliomas. Anticancer Drugs. 2015, 26, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.; DiBiase, S.; Meisenberg, B.; Flannery, T.; Patel, A.; Dhople, A.; Cheston, S.; Amin, P. Phase I clinical trial assessing temozolomide and tamoxifen with concomitant radiotherapy for treatment of high-grade glioma. Int J Radiat Oncol Biol Phys. 2012, 82, 739–742. [Google Scholar] [CrossRef] [PubMed]
- Misuth, M.; Horvath, D.; Miskovsky, P.; Huntosova, V. Synergism between PKCδ regulators hypericin and rottlerin enhances apoptosis in U87 MG glioma cells after light stimulation. Vol. 18, PHOTODIAGNOSIS AND PHOTODYNAMIC THERAPY. PO BOX 211, 1000 AE AMSTERDAM, NETHERLANDS: ELSEVIER SCIENCE BV; 2017. p. 267–74.
- Kim, D.C.; Kim, S.H.; Jeong, M.W.; Baek, N.I.; Kim, K.T. Effect of rottlerin, a PKC-delta inhibitor, on TLR-4-dependent activation of murine microglia. Biochem Biophys Res Commun. 2005, 337, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Song, K.S.; Kim, J.S.; Yun, E.J.; Kim, Y.R.; Seo, K.S.; Park, J.H.; Jung, Y.J.; Park, J.I.; Kweon, G.R.; Yoon, W.H.; Lim, K.; Hwang, B.D. Rottlerin induces autophagy and apoptotic cell death through a PKC-delta-independent pathway in HT1080 human fibrosarcoma cells: The protective role of autophagy in apoptosis. Autophagy. 2008, 4, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.H.; Park, J.W.; Choi, K.S.; Park, Y.B.; Kwon, T.K. Rottlerin induces apoptosis via death receptor 5 (DR5) upregulation through CHOP-dependent and PKC delta-independent mechanism in human malignant tumor cells. Carcinogenesis. 2009, 30, 729–736. [Google Scholar] [CrossRef] [PubMed]
- Saadeh, F.S.; Mahfouz, R.; Assi, H.I. EGFR as a clinical marker in glioblastomas and other gliomas. Int J Biol Markers. 2018, 33, 22–32. [Google Scholar] [CrossRef] [PubMed]
- Brennan, C.W.; Verhaak, R.G.; McKenna, A.; Campos, B.; Noushmehr, H.; Salama, S.R.; Zheng, S.; Chakravarty, D.; Sanborn, J.Z.; Berman, S.H.; et al. The somatic genomic landscape of glioblastoma. Cell. 2013, 155, 462–477. [Google Scholar] [CrossRef]
- Oprita, A.; Baloi, S.C.; Staicu, G.A.; Alexandru, O.; Tache, D.E.; Danoiu, S.; Micu, E.S.; Sevastre, A.S. Updated Insights on EGFR Signaling Pathways in Glioma. Int J Mol Sci. 2021, 22, 587. [Google Scholar] [CrossRef]
- Fanuel-Barret, D.; Patrice, T.; Foultier, M.T.; Vonarx-Coinsmann, V.; Robillard, N.; Lajat, Y. Influence of epidermal growth factor on photodynamic therapy of glioblastoma cells in vitro. Res Exp Med (Berl). 1997, 197, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.W.; Hung, M.C.; Hsieh, C.Y.; Tung, E.C.; Wang, Y.H.; Tsai, J.C.; Lee, J.M. The effects of Photofrin-mediated photodynamic therapy on the modulation of EGFR in esophageal squamous cell carcinoma cells. Lasers Med Sci. 2013, 28, 605–614. [Google Scholar] [CrossRef] [PubMed]
- Tsai, T.; Ji, H.T.; Chiang, P.C.; Chou, R.H.; Chang, W.S.; Chen, C.T. ALA-PDT results in phenotypic changes and decreased cellular invasion in surviving cancer cells. Lasers Surg Med. 2009, 41, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Ulfo, L.; Costantini, P.E.; Di Giosia, M.; Danielli, A.; Calvaresi, M. EGFR-Targeted Photodynamic Therapy. Pharmaceutics. 2022, 14, 241. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.J.; Niu, C.J.; Lai, B.; Chen, Y.; Kuta, V.; Lilge, L.D. Modulation of PPIX synthesis and accumulation in various normal and glioma cell lines by modification of the cellular signaling and temperature. Lasers Surg Med. 2013, 45, 460–468. [Google Scholar] [CrossRef] [PubMed]
- Fisher, C.; Obaid, G.; Niu, C.; Foltz, W.; Goldstein, A.; Hasan, T.; Lilge, L. Liposomal Lapatinib in Combination with Low-Dose Photodynamic Therapy for the Treatment of Glioma. J Clin Med. 2019, 8, 2214. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, S.M.B.; Kamel, A.; Ciubotaru, G.V.; Onose, G.; Sevastre, A.S.; Sfredel, V.; Danoiu, S.; Dricu, A.; Tataranu, L.G. An Overview of EGFR Mechanisms and Their Implications in Targeted Therapies for Glioblastoma. Int J Mol Sci. 2023, 24, 11110. [Google Scholar] [CrossRef]
- Jamali, Z.; Khoobi, M.; Hejazi, S.M.; Eivazi, N.; Abdolahpour, S.; Imanparast, F.; Moradi-Sardareh, H.; Paknejad, M. Evaluation of targeted curcumin (CUR) loaded PLGA nanoparticles for in vitro photodynamic therapy on human glioblastoma cell line. Photodiagnosis Photodyn Ther. 2018, 23, 190–201. [Google Scholar] [CrossRef] [PubMed]
- Meyers, J.D.; Cheng, Y.; Broome, A.M.; Agnes, R.S.; Schluchter, M.D.; Margevicius, S.; Wang, X.; Kenney, M.E.; Burda, C.; Basilion, J.P. Peptide-Targeted Gold Nanoparticles for Photodynamic Therapy of Brain Cancer. Part Part Syst Charact. 2015, 32, 448–457. [Google Scholar] [CrossRef]
- Xie, Y.; Su, N.; Yang, J.; Tan, Q.; Huang, S.; Jin, M.; Ni, Z.; Zhang, B.; Zhang, D.; Luo, F.; Chen, H.; Sun, X.; Feng, J.Q.; Qi, H.; Chen, L. FGF/FGFR signaling in health and disease. Signal Transduct Target Ther. 2020, 5, 181. [Google Scholar]
- Klimaschewski, L.; Claus, P. Fibroblast Growth Factor Signalling in the Diseased Nervous System. Mol Neurobiol. 2021, 58, 3884–3902. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Zhang, Z.; Lin, M.; Zhou, B.; Wang, Z. Bevacizumab has bidirectional regulatory effects on the secretion of basic fibroblast growth factor in glioma cells. Cytokine. 2020, 129, 155022. [Google Scholar] [CrossRef] [PubMed]
- Chai, N.; Stachon, T.; Berger, T.; Li, Z.; Seitz, B.; Langenbucher, A.; Szentmáry, N. Human corneal epithelial cell and fibroblast migration and growth factor secretion after rose bengal photodynamic therapy (RB-PDT) and the effect of conditioned medium. PLoS ONE. 2023, 18, e0296022. [Google Scholar] [CrossRef] [PubMed]
- LaMuraglia, G.M.; Adili, F.; Karp, S.J.; Statius van Eps, R.G.; Watkins, M.T. Photodynamic therapy inactivates extracellular matrix-basic fibroblast growth factor: Insights to its effect on the vascular wall. J Vasc Surg. 1997, 26, 294–301. [Google Scholar] [CrossRef]
- Vilchez, M.L.; Rodríguez, L.B.; Palacios, R.E.; Prucca, C.G.; Caverzán, M.D.; Caputto, B.L.; Rivarola, V.A.; Milla Sanabria, L.N. Isolation and initial characterization of human glioblastoma cells resistant to photodynamic therapy. Photodiagnosis Photodyn Ther. 2021, 33, 102097. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.; Hsiao, Y.; Teng, L.; Chen, C.; Kao, M. Comparative study on the ALA photodynamic effects of human glioma and meningioma cells. Vol. 24, LASERS IN SURGERY AND MEDICINE. DIV JOHN WILEY & SONS INC, 605 THIRD AVE, NEW YORK, NY 10158-0012 USA: WILEY-LISS; 1999. p. 296–305.
- Chakrabarti, M.; Banik, N.L.; Ray, S.K. Photofrin based photodynamic therapy and miR-99a transfection inhibited FGFR3 and PI3K/Akt signaling mechanisms to control growth of human glioblastoma In vitro and in vivo. PLoS ONE. 2013, 8, e55652. [Google Scholar] [CrossRef] [PubMed]
- Repetto, M.; Crimini, E.; Giugliano, F.; Morganti, S.; Belli, C.; Curigliano, G. Selective FGFR/FGF pathway inhibitors: Inhibition strategies, clinical activities, resistance mutations, and future directions. Expert Rev Clin Pharmacol. 2021, 14, 1233–1252. [Google Scholar] [CrossRef] [PubMed]
- Abounader, R.; Laterra, J. Scatter factor/hepatocyte growth factor in brain tumor growth and angiogenesis. Neuro Oncol. 2005, 7, 436–451. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.F.; Wang, X.B.; Tian, X.Y.; Li, Y.; Li, B.; Huang, Q.; Zhang, M.; Li, Z. Tumor-derived hepatocyte growth factor is associated with poor prognosis of patients with glioma and influences the chemosensitivity of glioma cell line to cisplatin in vitro. World J Surg Oncol. 2012, 10, 128. [Google Scholar] [CrossRef]
- Vogel, S.; Peters, C.; Etminan, N.; Börger, V.; Schimanski, A.; Sabel, M.C.; Sorg, R.V. Migration of mesenchymal stem cells towards glioblastoma cells depends on hepatocyte-growth factor and is enhanced by aminolaevulinic acid-mediated photodynamic treatment. Biochem Biophys Res Commun. 2013, 431, 428–432. [Google Scholar] [CrossRef]
- Matsumoto, K.; Umitsu, M.; De Silva, D.M.; Roy, A.; Bottaro, D.P. Hepatocyte growth factor/MET in cancer progression and biomarker discovery. Cancer Sci. 2017, 108, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Weathers, S.P.; de Groot, J. VEGF Manipulation in Glioblastoma. Oncology (Williston Park). 2015, 29, 720–727. [Google Scholar]
- Seyedmirzaei, H.; Shobeiri, P.; Turgut, M.; Hanaei, S.; Rezaei, N. VEGF levels in patients with glioma: A systematic review and meta-analysis. Rev Neurosci. 2020, 32, 191–202. [Google Scholar] [CrossRef] [PubMed]
- Onishi, M.; Ichikawa, T.; Kurozumi, K.; Date, I. Angiogenesis and invasion in glioma. Brain Tumor Pathol. 2011, 28, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Deininger, M.H.; Weinschenk, T.; Morgalla, M.H.; Meyermann, R.; Schluesener, H.J. Release of regulators of angiogenesis following Hypocrellin-A and -B photodynamic therapy of human brain tumor cells. Biochem Biophys Res Commun. 2002, 298, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Chen, X.; Chen, K.; Peng, Y.; Li, Y.; Ke, Y.; Gan, D. Tetra-sulfonate phthalocyanine zinc-bovine serum albumin conjugate-mediated photodynamic therapy of human glioma. J Biomater Appl. 2014, 29, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Ke, Y.; Jiang, X.; Cai, Y.; Peng, Y.; Li, Y. In vitro photodynamic therapy on human U251 glioma cells with a novel photosensitiser ZnPcS4-BSA. Br J Neurosurg. 2010, 24, 660–665. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Jiang, F.; Kalkanis, S.N.; Zhang, Z.; Hong, X.; Yang, H.; Chopp, M. Post-acute response of 9L gliosarcoma to Photofrin-mediated PDT in athymic nude mice. Lasers Med Sci. 2007, 22, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Zhang, Z.; Katakowski, M.; Robin, A.; Faber, M.; Zhang, F.; Chopp, M. Angiogenesis induced by photodynamic therapy in normal rat brain. Vol. 79, PHOTOCHEMISTRY AND PHOTOBIOLOGY. 111 RIVER ST, HOBOKEN 07030-5774, NJ USA: WILEY-BLACKWELL; 2004. p. 494–8.
- Zhang, X.; Jiang, F.; Zhang, Z.G.; Kalkanis, S.N.; Hong, X.; deCarvalho, A.C.; Chen, J.; Yang, H.; Robin, A.M.; Chopp, M. Low-dose photodynamic therapy increases endothelial cell proliferation and VEGF expression in nude mice brain. Lasers Med Sci. 2005, 20, 74–79. [Google Scholar] [CrossRef]
- Tzerkovsky, D.A.; Osharin, V.V.; Istomin, Y.P.; Alexandrova, E.N.; Vozmitel, M.A. Fluorescent diagnosis and photodynamic therapy for C6 glioma in combination with antiangiogenic therapy in subcutaneous and intracranial tumor models. Exp Oncol. 2014, 36, 85–89. [Google Scholar]
- Jiang, F.; Zhang, X.; Kalkanis, S.N.; Zhang, Z.; Yang, H.; Katakowski, M.; Hong, X.; Zheng, X.; Zhu, Z.; Chopp, M. Combination therapy with antiangiogenic treatment and photodynamic therapy for the nude mouse bearing U87 glioblastoma. Photochem Photobiol. 2008, 84, 128–137. [Google Scholar] [CrossRef] [PubMed]
- Tatar, O.; Shinoda, K.; Adam, A.; Eckert, T.; Eckardt, C.; Lucke, K.; Deuter, C.; Bartz-Schmidt, K.U.; Grisanti, S. Effect of verteporfin photodynamic therapy on endostatin and angiogenesis in human choroidal neovascular membranes. Br J Ophthalmol. 2007, 91, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Walia, A.; Yang, J.F.; Huang, Y.H.; Rosenblatt, M.I.; Chang, J.H.; Azar, D.T. Endostatin’s emerging roles in angiogenesis, lymphangiogenesis, disease, and clinical applications. Biochim Biophys Acta. 2015, 1850, 2422–2438. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.L.; Lin, H.C.; Chiang, W.L.; Shih, Y.H.; Chiang, P.F.; Luo, T.Y.; Cheng, C.C.; Shieh, M.J. Anti-angiogenic treatment (Bevacizumab) improves the responsiveness of photodynamic therapy in colorectal cancer. Photodiagnosis Photodyn Ther. 2018, 23, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.; Wen, P.Y. Antiangiogenic Therapy of High-Grade Gliomas. Prog Neurol Surg. 2018, 31, 180–199. [Google Scholar] [PubMed]
- Zhang, G.; Chen, L.; Sun, K.; Khan, A.A.; Yan, J.; Liu, H.; Lu, A.; Gu, N. Neuropilin-1 (NRP-1)/GIPC1 pathway mediates glioma progression. Tumour Biol. 2016, 37, 13777–13788. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.K.; Miao, W.; Tang, X.Y.; Zhang, H.P.; Wang, S.Y.; Luo, F.H.; et al. The expression and significance of neuropilin-1 (NRP-1) on glioma cell lines and glioma tissues. J Biomed Nanotechnol. 2013, 9, 559–563. [Google Scholar] [CrossRef]
- Chen, L.K.; Miao, W.; Tang, X.Y.; Zhang, H.P.; Wang, S.Y.; Luo, F.H.; et al. Inhibitory effect of neuropilin-1 monoclonal antibody (NRP-1 MAb) on glioma tumor in mice. J Biomed Nanotechnol. 2013, 9, 551–558. [Google Scholar] [CrossRef]
- Caponegro, M.D.; Moffitt, R.A.; Tsirka, S.E. Expression of neuropilin-1 is linked to glioma associated microglia and macrophages and correlates with unfavorable prognosis in high grade gliomas. Oncotarget. 2018, 9, 35655–35665. [Google Scholar] [CrossRef]
- Thomas, N.; Tirand, L.; Chatelut, E.; Plénat, F.; Frochot, C.; Dodeller, M.; Guillemin, F.; Barberi-Heyob, M. Tissue distribution and pharmacokinetics of an ATWLPPR-conjugated chlorin-type photosensitizer targeting neuropilin-1 in glioma-bearing nude mice. Photochem Photobiol Sci. 2008, 7, 433–441. [Google Scholar] [CrossRef]
- Tirand, L.; Frochot, C.; Vanderesse, R.; Thomas, N.; Trinquet, E.; Pinel, S.; Viriot, M.L.; Guillemin, F.; Barberi-Heyob, M. A peptide competing with VEGF165 binding on neuropilin-1 mediates targeting of a chlorin-type photosensitizer and potentiates its photodynamic activity in human endothelial cells. J Control Release. 2006, 111, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Bechet, D.; Auger, F.; Couleaud, P.; Marty, E.; Ravasi, L.; Durieux, N.; Bonnet, C.; Plénat, F.; Frochot, C.; Mordon, S.; Tillement, O.; Vanderesse, R.; Lux, F.; Perriat, P.; Guillemin, F.; Barberi-Heyob, M. Multifunctional ultrasmall nanoplatforms for vascular-targeted interstitial photodynamic therapy of brain tumors guided by real-time MRI. Nanomedicine. 2015, 11, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Gries, M.; Thomas, N.; Daouk, J.; Rocchi, P.; Choulier, L.; Jubréaux, J.; Pierson, J.; Reinhard, A.; Jouan-Hureaux, V.; Chateau, A.; Acherar, S.; Frochot, C.; Lux, F.; Tillement, O.; Barberi-Heyob, M. Multiscale Selectivity and in vivo Biodistribution of NRP-1-Targeted Theranostic AGuIX Nanoparticles for PDT of Glioblastoma. Int J Nanomedicine. 2020, 15, 8739–8758. [Google Scholar] [CrossRef] [PubMed]
- Thomas, E.; Colombeau, L.; Gries, M.; Peterlini, T.; Mathieu, C.; Thomas, N.; Boura, C.; Frochot, C.; Vanderesse, R.; Lux, F.; Barberi-Heyob, M.; Tillement, O. Ultrasmall AGuIX theranostic nanoparticles for vascular-targeted interstitial photodynamic therapy of glioblastoma. Int J Nanomedicine. 2017, 12, 7075–7088. [Google Scholar] [CrossRef] [PubMed]
- Tirand, L.; Bastogne, T.; Bechet, D.; Linder, M.; Thomas, N.; Frochot, C.; Guillemin, F.; Barberi-Heyob, M. Response surface methodology: An extensive potential to optimize in vivo photodynamic therapy conditions. Int J Radiat Oncol Biol Phys. 2009, 75, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Thomas, N.; Bechet, D.; Becuwe, P.; Tirand, L.; Vanderesse, R.; Frochot, C.; Guillemin, F.; Barberi-Heyob, M. Peptide-conjugated chlorin-type photosensitizer binds neuropilin-1 in vitro and in vivo. Vol. 96, JOURNAL OF PHOTOCHEMISTRY AND PHOTOBIOLOGY B-BIOLOGY. PO BOX 564, 1001 LAUSANNE, SWITZERLAND: ELSEVIER SCIENCE SA; 2009. p. 101–8.
- Lu, L.; Zhao, X.; Fu, T.; Li, K.; He, Y.; Luo, Z.; Dai, L.; Zeng, R.; Cai, K. An iRGD-conjugated prodrug micelle with blood-brain-barrier penetrability for anti-glioma therapy. Biomaterials. 2020, 230, 119666. [Google Scholar] [CrossRef] [PubMed]
- Kong, D.H.; Kim, Y.K.; Kim, M.R.; Jang, J.H.; Lee, S. Emerging Roles of Vascular Cell Adhesion Molecule-1 (VCAM-1) in Immunological Disorders and Cancer. Int J Mol Sci. 2018, 19, 1057. [Google Scholar] [CrossRef]
- Mäenpää, A.; Kovanen, P.E.; Paetau, A.; Jäáskeläinen, J.; Timonen, T. Lymphocyte adhesion molecule ligands and extracellular matrix proteins in gliomas and normal brain: Expression of VCAM-1 in gliomas. Acta Neuropathol. 1997, 94, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Cheng, V.W.T.; de Pennington, N.; Zakaria, R.; Larkin, J.R.; Serres, S.; Sarkar, M.; Kirkman, M.A.; Bristow, C.; Croal, P.; Plaha, P.; Campo, L.; Chappell, M.A.; Lord, S.; Jenkinson, M.D.; Middleton, M.R.; Sibson, N.R. VCAM-1-targeted MRI Improves Detection of the Tumor-brain Interface. Clin Cancer Res. 2022, 28, 2385–2396. [Google Scholar] [CrossRef]
- Zhan, Q.; Yue, W.; Hu, S. The inhibitory effect of photodynamic therapy and of an anti-VCAM-1 monoclonal antibody on the in vivo growth of C6 glioma xenografts. Vol. 44, BRAZILIAN JOURNAL OF MEDICAL AND BIOLOGICAL RESEARCH. FACULDADE MEDICINA, CASA 10, 14049 RIBEIRAO PRETO, RIBEIRAO PRETO, SP 14049, BRAZIL: ASSOC BRAS DIVULG CIENTIFICA; 2011. p. 489–96.
- Volanti, C.; Gloire, G.; Vanderplasschen, A.; Jacobs, N.; Habraken, Y.; Piette, J. Downregulation of ICAM-1 and VCAM-1 expression in endothelial cells treated by photodynamic therapy. Oncogene. 2004, 23, 8649–8658. [Google Scholar] [CrossRef]
- Kawczyk-Krupka, A.; Czuba, Z.P.; Kwiatek, B.; Kwiatek, S.; Krupka, M.; Sieroń, K. The effect of ALA-PDT under normoxia and cobalt chloride (CoCl2)-induced hypoxia on adhesion molecules (ICAM-1, VCAM-1) secretion by colorectal cancer cells. Photodiagnosis Photodyn Ther. 2017, 19, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Takada, Y.; Ye, X.; Simon, S. The integrins. Genome Biol. 2007, 8, 215. [Google Scholar] [CrossRef]
- Liu, F.; Wu, Q.; Dong, Z.; Liu, K. Integrins in cancer: Emerging mechanisms and therapeutic opportunities. Pharmacol Ther. 2023, 247, 108458. [Google Scholar] [CrossRef]
- Ferrer, V.P.; Moura Neto, V.; Mentlein, R. Glioma infiltration and extracellular matrix: Key players and modulators. Glia. 2018, 66, 1542–1565. [Google Scholar] [CrossRef] [PubMed]
- Di Venosa, G.; Perotti, C.; Batlle, A.; Casas, A. The role of cytoskeleton and adhesion proteins in the resistance to photodynamic therapy. Possible therapeutic interventions. Photochem Photobiol Sci. 2015, 14, 1451–1464. [Google Scholar] [CrossRef]
- Au, C.M.; Luk, S.K.; Jackson, C.J.; Ng, H.K.; Yow, C.M.N.; To, S.S.T. Differential effects of photofrin, 5-aminolevulinic acid and calphostin C on glioma cells. Journal of Photochemistry and Photobiology B: Biology. 2006, 85, 92–101. [Google Scholar] [CrossRef] [PubMed]
- Pacheco Soares, C.; Maftou Costa, M.; da Cunha Menezes Costa, C.G.; de Siqueira Silva, A.C.; Moraes, K.C. Evaluation of photodynamic therapy in adhesion protein expression. Oncology Letters 2014, 8, 714–718. [Google Scholar] [CrossRef]
- Wei, Y.; Zhou, F.; Zhang, D.; Chen, Q.; Xing, D. A graphene oxide based smart drug delivery system for tumor mitochondria-targeting photodynamic therapy. Nanoscale. 2016, 8, 3530–3538. [Google Scholar] [CrossRef]
- de Almeida, L.G.N.; Thode, H.; Eslambolchi, Y.; Chopra, S.; Young, D.; Gill, S.; Devel, L.; Dufour, A. Matrix Metalloproteinases: From Molecular Mechanisms to Physiology, Pathophysiology, and Pharmacology. Pharmacol Rev. 2022, 74, 712–768. [Google Scholar] [CrossRef]
- Zhou, W.; Yu, X.; Sun, S.; Zhang, X.; Yang, W.; Zhang, J.; Zhang, X.; Jiang, Z. Increased expression of MMP-2 and MMP-9 indicates poor prognosis in glioma recurrence. Biomed Pharmacother. 2019, 118, 109369. [Google Scholar] [CrossRef]
- Wang, L.; Yuan, J.; Tu, Y.; Mao, X.; He, S.; Fu, G.; Zong, J.; Zhang, Y. Co-expression of MMP-14 and MMP-19 predicts poor survival in human glioma. Clin Transl Oncol. 2013, 15, 139–145. [Google Scholar] [CrossRef]
- Chu, E.S.M.; Wong, T.K.S.; Yow, C.M.N. Photodynamic effect in medulloblastoma: Downregulation of matrix metalloproteinases and human telomerase reverse transcriptase expressions. Photochem Photobiol Sci. 2008, 7, 76–83. [Google Scholar] [CrossRef]
- Etminan, N.; Peters, C.; Ficnar, J.; Anlasik, S.; Bünemann, E.; Slotty, P.J.; Hänggi, D.; Steiger, H.J.; Sorg, R.V.; Stummer, W. Modulation of migratory activity and invasiveness of human glioma spheroids following 5-aminolevulinic acid-based photodynamic treatment. Laboratory investigation. J Neurosurg. 2011, 115, 281–288. [Google Scholar] [CrossRef]
- Li, J.; Tian, Y.; Zheng, T. A multifunctional nanoprobe for real-time SERS monitoring of invasion ability affected by photodynamic therapy. Chem Commun (Camb). 2022, 58, 6542–6545. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lin, H.; Chen, Q.; Yu, L.; Bai, D. MPPa-PDT suppresses breast tumor migration/invasion by inhibiting Akt-NF-κB-dependent MMP-9 expression via ROS. BMC Cancer. 2019, 19, 1159. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Shen, B.; Swinarska, J.T.; Li, W.; Xiao, K.; He, P. 9-Hydroxypheophorbide α-mediated photodynamic therapy induces matrix metalloproteinase-2 (MMP-2) and MMP-9 down-regulation in Hep-2 cells via ROS-mediated suppression of the ERK pathway. Photodiagnosis Photodyn Ther. 2014, 11, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Li, B.O.; Meng, C.; Zhang, X.; Cong, D.; Gao, X.; Gao, W.; Ju, D.; Hu, S. Effect of photodynamic therapy combined with torasemide on the expression of matrix metalloproteinase 2 and sodium-potassium-chloride cotransporter 1 in rat peritumoral edema and glioma. Oncol Lett. 2016, 11, 2084–2090. [Google Scholar] [CrossRef]
- Lee, E.J.; Kim, S.Y.; Hyun, J.W.; Min, S.W.; Kim, D.H.; Kim, H.S. Glycitein inhibits glioma cell invasion through down-regulation of MMP-3 and MMP-9 gene expression. Chem Biol Interact. 2010, 185, 18–24. [Google Scholar] [CrossRef]
- Wang, X.; Chen, X.; Sun, L.; Bi, X.; He, H.; Chen, L.; Pang, J. MicroRNA 34a inhibits cell growth and migration in human glioma cells via MMP 9. Mol Med Rep. 2019, 20, 57–64. [Google Scholar] [CrossRef]
- Middeldorp, J.; Hol, E.M. GFAP in health and disease. Prog Neurobiol. 2011, 93, 421–443. [Google Scholar] [CrossRef]
- van Asperen, J.V.; Robe, P.A.J.T.; Hol, E.M. GFAP Alternative Splicing and the Relevance for Disease - A Focus on Diffuse Gliomas. ASN Neuro. 2022, 14, 17590914221102065. [Google Scholar] [CrossRef]
- van Bodegraven, E.J.; van Asperen, J.V.; Robe, P.A.J.; Hol, E.M. Importance of GFAP isoform-specific analyses in astrocytoma. Glia. 2019, 67, 1417–1433. [Google Scholar] [CrossRef] [PubMed]
- Uceda-Castro, R.; van Asperen, J.V.; Vennin, C.; Sluijs, J.A.; van Bodegraven, E.J.; Margarido, A.S.; Robe, P.A.J.; van Rheenen, J.; Hol, E.M. GFAP splice variants fine-tune glioma cell invasion and tumour dynamics by modulating migration persistence. Sci Rep. 2022, 12, 424. [Google Scholar] [CrossRef] [PubMed]
- Namatame, H.; Akimoto, J.; Matsumura, H.; Haraoka, J.; Aizawa, K. Photodynamic therapy of C6-implanted glioma cells in the rat brain employing second-generation photosensitizer talaporfin sodium. Vol. 5, PHOTODIAGNOSIS AND PHOTODYNAMIC THERAPY. RADARWEG 29, 1043 NX AMSTERDAM, NETHERLANDS: ELSEVIER; 2008. p. 198–209.
- Hara, A.; Sakai, N.; Yamada, H.; Niikawa, S.; Ohno, T.; Tanaka, T.; Mori, H. Proliferative assessment of GFAP-positive and GFAP-negative glioma cells by nucleolar organizer region staining. Surg Neurol. 1991, 36, 190–194. [Google Scholar] [CrossRef]
- Hara, A.; Hirayama, H.; Sakai, N.; Yamada, H.; Tanaka, T.; Mori, H. Correlation between nucleolar organizer region staining and Ki-67 immunostaining in human gliomas. Surg Neurol. 1990, 33, 320–324. [Google Scholar] [CrossRef]
- Harmelin, A.; Zuckerman, A.; Nyska, A. Correlation of Ag-NOR protein measurements with prognosis in canine transmissible venereal tumour. J Comp Pathol. 1995, 112, 429–433. [Google Scholar] [CrossRef]
- Arden, K.C.; Pathak, S.; Frankel, L.S.; Zander, A. Ag-NOR staining in human chromosomes: Differential staining in normal and leukemic bone-marrow samples. Int J Cancer. 1985, 36, 647–649. [Google Scholar] [CrossRef]
- Schelling, J.R.; Abu Jawdeh, B.G. Regulation of cell survival by Na+/H+ exchanger-1. Am J Physiol Renal Physiol. 2008, 295, F625–F632. [Google Scholar] [CrossRef] [PubMed]
- Guan, X.; Hasan, M.N.; Begum, G.; Kohanbash, G.; Carney, K.E.; Pigott, V.M.; Persson, A.I.; Castro, M.G.; Jia, W.; Sun, D. Blockade of Na/H exchanger stimulates glioma tumor immunogenicity and enhances combinatorial TMZ and anti-PD-1 therapy. Cell Death Dis. 2018, 9, 1010. [Google Scholar] [CrossRef]
- Zhu, W.; Carney, K.E.; Pigott, V.M.; Falgoust, L.M.; Clark, P.A.; Kuo, J.S.; Sun, D. Glioma-mediated microglial activation promotes glioma proliferation and migration: Roles of Na+/H+ exchanger isoform 1. Carcinogenesis. 2016, 37, 839–851. [Google Scholar] [CrossRef]
- Hasan, M.N.; Luo, L.; Ding, D.; et al. Blocking NHE1 stimulates glioma tumor immunity by restoring OXPHOS function of myeloid cells. Theranostics. 2021, 11, 1295–1309. [Google Scholar] [CrossRef]
- Hou, K.; Liu, J.; Du, J.; Mi, S.; Ma, S.; Ba, Y.; Ji, H.; Li, B.; Hu, S. Dihydroartemisinin prompts amplification of photodynamic therapy-induced reactive oxygen species to exhaust Na/H exchanger 1-mediated glioma cells invasion and migration. J Photochem Photobiol B. 2021, 219, 112192. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.; Chen, L.; Chi, D.; Cong, D.; Zhou, P.; Jin, J.; Ji, H.; Liang, B.; Gao, S.; Hu, S. Photodynamic therapy combined with temozolomide inhibits C6 glioma migration and invasion and promotes mitochondrial-associated apoptosis by inhibiting sodium-hydrogen exchanger isoform 1. Photodiagnosis Photodyn Ther. 2019, 26, 405–412. [Google Scholar] [CrossRef]
- Shimoda, L.A.; Fallon, M.; Pisarcik, S.; Wang, J.; Semenza, G.L. HIF-1 regulates hypoxic induction of NHE1 expression and alkalinization of intracellular pH in pulmonary arterial myocytes. Am J Physiol Lung Cell Mol Physiol. 2006, 291, L941–L949. [Google Scholar] [CrossRef]
- Tamtaji, O.R.; Mirzaei, H.; Shamshirian, A.; Shamshirian, D.; Behnam, M.; Asemi, Z. New trends in glioma cancer therapy: Targeting Na⁺/H⁺ exchangers. J Cell Physiol. 2019, 235, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Dolcet, X.; Llobet, D.; Pallares, J.; Matias-Guiu, X. NF-kB in development and progression of human cancer. Virchows Arch. 2005, 446, 475–482. [Google Scholar] [CrossRef]
- Li, Y.; Zhou, Q.L.; Sun, W.; Chandrasekharan, P.; Cheng, H.S.; Ying, Z.; Lakshmanan, M.; Raju, A.; Tenen, D.G.; Cheng, S.Y.; Chuang, K.H.; Li, J.; Prabhakar, S.; Li, M.; Tergaonkar, V. Non-canonical NF-κB signalling and ETS1/2 cooperatively drive C250T mutant TERT promoter activation. Nat Cell Biol. 2015, 17, 1327–1338. [Google Scholar] [CrossRef]
- Hu, Y.H.; Jiao, B.H.; Wang, C.Y.; Wu, J.L. Regulation of temozolomide resistance in glioma cells via the RIP2/NF-κB/MGMT pathway. CNS Neurosci Ther. 2021, 27, 552–563. [Google Scholar] [CrossRef] [PubMed]
- Coupienne, I.; Bontems, S.; Dewaele, M.; Rubio, N.; Habraken, Y.; Fulda, S.; Agostinis, P.; Piette, J. NF-kappaB inhibition improves the sensitivity of human glioblastoma cells to 5-aminolevulinic acid-based photodynamic therapy. Biochem Pharmacol. 2011, 81, 606–616. [Google Scholar] [CrossRef]
- Piette, J. Signalling pathway activation by photodynamic therapy: NF-κB at the crossroad between oncology and immunology. Photochem Photobiol Sci. 2015, 14, 1510–1517. [Google Scholar] [CrossRef]
- Fahey, J.M.; Korytowski, W.; Girotti, A.W. Upstream signaling events leading to elevated production of pro-survival nitric oxide in photodynamically-challenged glioblastoma cells. Free Radic Biol Med. 2019, 137, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, S.; Banik, N.L.; Patel, S.J.; Ray, S.K. 5-Aminolevulinic acid-based photodynamic therapy suppressed survival factors and activated proteases for apoptosis in human glioblastoma U87MG cells. Neurosci Lett. 2007, 415, 242–247. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Alqawi, O.; Espiritu, M. Metronomic PDT and cell death pathways. Methods Mol Biol. 2010, 635, 65–78. [Google Scholar] [PubMed]
- He, Y.; Duan, L.; Wu, H.; Chen, S.; Lu, T.; Li, T.; He, Y. Integrated Transcriptome Analysis Reveals the Impact of Photodynamic Therapy on Cerebrovascular Endothelial Cells. Front Oncol. 2021, 11, 731414. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, M.; Candido, M.F.; Valera, E.T.; Brassesco, M.S. The multifaceted NF-kB: Are there still prospects of its inhibition for clinical intervention in pediatric central nervous system tumors? Cell Mol Life Sci. 2021, 78, 6161–6200. [Google Scholar] [CrossRef] [PubMed]
- Kumthekar, P.; Grimm, S.; Chandler, J.; et al. A phase II trial of arsenic trioxide and temozolomide in combination with radiation therapy for patients with malignant gliomas. J Neurooncol 2017, 133, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Tu, Z.; Li, K.; Ji, Q.; Huang, Y.; Lv, S.; Li, J.; Wu, L.; Huang, K.; Zhu, X. Pan-cancer analysis: Predictive role of TAP1 in cancer prognosis and response to immunotherapy. BMC Cancer. 2023, 23, 133. [Google Scholar] [CrossRef]
- Satoh, E.; Mabuchi, T.; Satoh, H.; Asahara, T.; Nukui, H.; Naganuma, H. Reduced expression of the transporter associated with antigen processing 1 molecule in malignant glioma cells, and its restoration by interferon-gamma and -beta. J Neurosurg. 2006, 104, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Žilionytė, K.; Bagdzevičiūtė, U.; Mlynska, A.; Urbštaitė, E.; Paberalė, E.; Dobrovolskienė, N.; Krasko, J.A.; Pašukonienė, V. Correction to: Functional antigen processing and presentation mechanism as a prerequisite factor of response to treatment with dendritic cell vaccines and anti-PD-1 in preclinical murine LLC1 and GL261 tumor models. Cancer Immunol Immunother. 2022, 71, 2701. [Google Scholar] [CrossRef]
- Yang, W.; Li, Y.; Gao, R.; Xiu, Z.; Sun, T. MHC class I dysfunction of glioma stem cells escapes from CTL-mediated immune response via activation of Wnt/beta-catenin signaling pathway. Oncogene. 2020, 39, 1098–1111. [Google Scholar] [CrossRef]
- Yasinjan, F.; Xing, Y.; Geng, H.; Guo, R.; Yang, L.; Liu, Z.; Wang, H. Immunotherapy: A promising approach for glioma treatment. Front Immunol. 2023, 14, 1255611. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Li, J.L.; Xu, X.K.; Zheng, M.G.; Wen, C.C.; Li, F.C. HMME-based PDT restores expression and function of transporter associated with antigen processing 1 (TAP1) and surface presentation of MHC class I antigen in human glioma. J Neurooncol. 2011, 105, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Liu, D.; Li, L. PD-1/PD-L1 pathway: Current researches in cancer. Am J Cancer Res. 2020, 10, 727–742. [Google Scholar] [PubMed]
- Xue, S.; Hu, M.; Iyer, V.; Yu, J. Blocking the PD-1/PD-L1 pathway in glioma: A potential new treatment strategy. J Hematol Oncol. 2017, 10, 81. [Google Scholar] [CrossRef] [PubMed]
- Gurung, P.; Lim, J.; Shrestha, R.; et al. Chlorin e6-associated photodynamic therapy enhances abscopal antitumor effects via inhibition of PD-1/PD-L1 immune checkpoint. Sci Rep 2023, 13, 4647. [Google Scholar] [CrossRef] [PubMed]
- Lobo, C.S.; Mendes, M.I.P.; Pereira, D.A.; et al. Photodynamic therapy changes tumour immunogenicity and promotes immune-checkpoint blockade response, particularly when combined with micromechanical priming. Sci Rep 2023, 13, 11667. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Shen, A.; Cheng, C.E.; Chen, J.; Powers, J.; Rayman, P.; Diaz, M.; Hasan, T.; Maytin, E.V. Combination of vitamin D and photodynamic therapy enhances immune responses in murine models of squamous cell skin cancer. Photodiagnosis Photodyn Ther. 2024, 45, 103983. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Tait, S.W.G. Targeting immunogenic cell death in cancer. Mol Oncol. 2020, 14, 2994–3006. [Google Scholar] [CrossRef] [PubMed]
- Nabrinsky, E.; Macklis, J.; Bitran, J. A Review of the Abscopal Effect in the Era of Immunotherapy. Cureus. 2022, 14, e29620. [Google Scholar] [CrossRef] [PubMed]
- Aebisher, D.; Woźnicki, P.; Bartusik-Aebisher, D. Photodynamic Therapy and Adaptive Immunity Induced by Reactive Oxygen Species: Recent Reports. Cancers 2024, 16, 967. [Google Scholar] [CrossRef]
- Turubanova, V.D.; Balalaeva, I.V.; Mishchenko, T.A.; Catanzaro, E.; Alzeibak, R.; Peskova, N.N.; Efimova, I.; Bachert, C.; Mitroshina, E.V.; Krysko, O.; Vedunova, M.V.; Krysko, D.V. Immunogenic cell death induced by a new photodynamic therapy based on photosens and photodithazine. J Immunother Cancer. 2019, 7, 350. [Google Scholar] [CrossRef]
- Shibata, S.; Shinozaki, N.; Suganami, A.; Ikegami, S.; Kinoshita, Y.; Hasegawa, R.; Kentaro, H.; Okamoto, Y.; Aoki, I.; Tamura, Y.; Iwadate, Y. Photo-immune therapy with liposomally formulated phospholipid-conjugated indocyanine green induces specific antitumor responses with heat shock protein-70 expression in a glioblastoma model. Oncotarget. 2019, 10, 175–183. [Google Scholar] [CrossRef] [PubMed]
- Pangal, D.J.; Yarovinsky, B.; Cardinal, T.; Cote, D.J.; Ruzevick, J.; Attenello, F.J.; Chang, E.L.; Ye, J.; Neman, J.; Chow, F.; Zada, G. The abscopal effect: Systematic review in patients with brain and spine metastases. Neurooncol Adv. 2022, 4, vdac132. [Google Scholar] [CrossRef] [PubMed]
- Lin, R.A.; Lin, J.K.; Lin, S.Y. Mechanisms of immunogenic cell death and immune checkpoint blockade therapy. Kaohsiung J Med Sci. 2021, 37, 448–458. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yu, S.; Wang, X.; Qian, Y.; Wu, W.; Zhang, S.; Zheng, B.; Wei, G.; Gao, S.; Cao, Z.; Fu, W.; Xiao, Z.; Lu, W. High Affinity of Chlorin e6 to Immunoglobulin G for Intraoperative Fluorescence Image-Guided Cancer Photodynamic and Checkpoint Blockade Therapy. ACS Nano. 2019, 13, 10242–10260. [Google Scholar] [CrossRef] [PubMed]
- Yin, N.; Wang, Y.; Liu, Y.; Niu, R.; Zhang, S.; Cao, Y.; Lv, Z.; Song, S.; Liu, X.; Zhang, H. A Cholesterol Metabolic Regulated Hydrogen-Bonded Organic Framework (HOF)-Based Biotuner for Antibody Non-Dependent Immunotherapy Tailored for Glioblastoma. Adv Mater. 2023, 35, e2303567. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Namkoong, H.; Doh, J.; Choi, J.C.; Yang, B.G.; Park, Y.; Chul Sung, Y. Negative role of inducible PD-1 on survival of activated dendritic cells. J Leukoc Biol. 2014, 95, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Friedrich, M.; Hahn, M.; Michel, J.; Sankowski, R.; Kilian, M.; Kehl, N.; Günter, M.; Bunse, T.; Pusch, S.; von Deimling, A.; Wick, W.; Autenrieth, S.E.; Prinz, M.; Platten, M.; Bunse, L. Dysfunctional dendritic cells limit antigen-specific T cell response in glioma. Neuro Oncol. 2023, 25, 263–276. [Google Scholar] [CrossRef] [PubMed]
- Etminan, N.; Peters, C.; Lakbir, D.; Bünemann, E.; Börger, V.; Sabel, M.C.; Hänggi, D.; Steiger, H.J.; Stummer, W.; Sorg, R.V. Heat-shock protein 70-dependent dendritic cell activation by 5-aminolevulinic acid-mediated photodynamic treatment of human glioblastoma spheroids in vitro. Br J Cancer. 2011, 105, 961–969. [Google Scholar] [CrossRef]
- Rothe, F.; Patties, I.; Kortmann, R.D.; Glasow, A. Immunomodulatory Effects by Photodynamic Treatment of Glioblastoma Cells In Vitro. Molecules. 2022, 27, 3384. [Google Scholar] [CrossRef]
- Vedunova, M.; Turubanova, V.; Vershinina, O.; Savyuk, M.; Efimova, I.; Mishchenko, T.; Raedt, R.; Vral, A.; Vanhove, C.; Korsakova, D.; Bachert, C.; Coppieters, F.; Agostinis, P.; Garg, A.D.; Ivanchenko, M.; Krysko, O.; Krysko, D.V. DC vaccines loaded with glioma cells killed by photodynamic therapy induce Th17 anti-tumor immunity and provide a four-gene signature for glioma prognosis. Cell Death Dis. 2022, 13, 1062. [Google Scholar] [CrossRef] [PubMed]
- Redkin, T.S.; Sleptsova, E.E.; Turubanova, V.D.; Saviuk, M.O.; Lermontova, S.A.; Klapshina, L.G.; Peskova, N.N.; Balalaeva, I.V.; Krysko, O.; Mishchenko, T.A.; Vedunova, M.V.; Krysko, D.V. Dendritic Cells Pulsed with Tumor Lysates Induced by Tetracyanotetra(aryl)porphyrazines-Based Photodynamic Therapy Effectively Trigger Anti-Tumor Immunity in an Orthotopic Mouse Glioma Model. Pharmaceutics. 2023, 15, 2430. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, L.; Jia, M.; Liao, Q.; Peng, G.; Luo, G.; Zhou, Y. Dendritic cell vaccines improve the glioma microenvironment: Influence, challenges, and future directions. Cancer Med. 2023, 12, 7207–7221. [Google Scholar] [CrossRef] [PubMed]
- Mazurek, M.; Rola, R. The implications of nitric oxide metabolism in the treatment of glial tumors. Neurochem Int. 2021, 150, 105172. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.I.; Yin, J.H.; Mishra, S.; Mishra, R.; Hsu, C.Y. NO-mediated chemoresistance in C6 glioma cells. Ann N Y Acad Sci. 2002, 962, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.J.; Li, F.; Zhu, D.Y. nNOS and Neurological, Neuropsychiatric Disorders: A 20-Year Story. Neurosci Bull. 2023, 39, 1439–1453. [Google Scholar] [CrossRef] [PubMed]
- Kolluru, G.K.; Siamwala, J.H.; Chatterjee, S. eNOS phosphorylation in health and disease. Biochimie. 2010, 92, 1186–1198. [Google Scholar] [CrossRef] [PubMed]
- Lechner, M.; Lirk, P.; Rieder, J. Inducible nitric oxide synthase (iNOS) in tumor biology: The two sides of the same coin. Semin Cancer Biol. 2005, 15, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Jahani-Asl, A.; Bonni, A. iNOS: A potential therapeutic target for malignant glioma. Curr Mol Med. 2013, 13, 1241–1249. [Google Scholar] [CrossRef] [PubMed]
- Maccallini, C.; Gallorini, M.; Cataldi, A.; Amoroso, R. Targeting iNOS As a Valuable Strategy for the Therapy of Glioma. ChemMedChem. 2020, 15, 339–344. [Google Scholar] [CrossRef]
- Fahey, J.M.; Emmer, J.V.; Korytowski, W.; Hogg, N.; Girotti, A.W. Antagonistic Effects of Endogenous Nitric Oxide in a Glioblastoma Photodynamic Therapy Model. Photochem Photobiol. 2016, 92, 842–853. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.M.; Stancill, J.S.; Smith, B.C.; Girotti, A.W. Nitric oxide antagonism to glioblastoma photodynamic therapy and mitigation thereof by BET bromodomain inhibitor JQ1. J Biol Chem. 2018, 293, 5345–5359. [Google Scholar] [CrossRef] [PubMed]
- Bazak, J.; Korytowski, W.; Girotti, A.W. Bystander Effects of Nitric Oxide in Cellular Models of Anti-Tumor Photodynamic Therapy. Cancers (Basel). 2019, 11, 1674. [Google Scholar] [CrossRef]
- Merenzon, M.A.; Hincapie Arias, E.; Bhatia, S.; Shah, A.H.; Higgins, D.M.O.; Villaverde, M.; Belgorosky, D.; Eijan, A.M. Nitric oxide synthase inhibitors as potential therapeutic agents for gliomas: A systematic review. Nitric Oxide. 2023, 138-139, 10–16. [Google Scholar] [CrossRef] [PubMed]
- de Groot, J.; Sontheimer, H. Glutamate and the biology of gliomas. Glia. 2011, 59, 1181–1189. [Google Scholar] [CrossRef]
- Pei, Z.; Lee, K.C.; Khan, A.; Erisnor, G.; Wang, H.Y. Pathway analysis of glutamate-mediated, calcium-related signaling in glioma progression. Biochem Pharmacol. 2020, 176, 113814. [Google Scholar] [CrossRef]
- Du, P.; Hu, S.; Cheng, Y.; Li, F.; Li, M.; Li, J.; Yi, L.; Feng, H. Photodynamic therapy leads to death of C6 glioma cells partly through AMPAR. Brain Res. 2012, 1433, 153–159. [Google Scholar] [CrossRef]
- Lyons, S.A.; Chung, W.J.; Weaver, A.K.; Ogunrinu, T.; Sontheimer, H. Autocrine glutamate signaling promotes glioma cell invasion. Cancer Res. 2007, 67, 9463–9471. [Google Scholar] [CrossRef]
- Hu, S.L.; Du, P.; Hu, R.; Li, F.; Feng, H. Imbalance of Ca2+ and K+ fluxes in C6 glioma cells after PDT measured with scanning ion-selective electrode technique. Lasers Med Sci. 2014, 29, 1261–1267. [Google Scholar] [CrossRef]
- Papadopoulos, V.; Lecanu, L.; Brown, R.C.; Han, Z.; Yao, Z.X. Peripheral-type benzodiazepine receptor in neurosteroid biosynthesis, neuropathology and neurological disorders. Neuroscience. 2006, 138, 749–756. [Google Scholar] [CrossRef]
- Starosta-Rubinstein, S.; Ciliax, B.J.; Penney, J.B.; McKeever, P.; Young, A.B. Imaging of a glioma using peripheral benzodiazepine receptor ligands. Proc Natl Acad Sci U S A. 1987, 84, 891–895. [Google Scholar] [CrossRef] [PubMed]
- Takada, A.; et al. Autoradiographic study of peripheral benzodiazepine receptors in animal brain tumor models and human gliomas. European Journal of Pharmacology: Environmental Toxicology and Pharmacology 1992, 228, 131–139. [Google Scholar]
- Veenman, L.; Levin, E.; Weisinger, G.; Leschiner, S.; Spanier, I.; Snyder, S.H.; Weizman, A.; Gavish, M. Peripheral-type benzodiazepine receptor density and in vitro tumorigenicity of glioma cell lines. Biochem Pharmacol. 2004, 68, 689–698. [Google Scholar] [CrossRef] [PubMed]
- Corsi, L.; Geminiani, E.; Baraldi, M. Peripheral benzodiazepine receptor (PBR) new insight in cell proliferation and cell differentiation review. Curr Clin Pharmacol. 2008, 3, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Bisland, S.K.; Goebel, E.A.; Hassanali, N.S.; Johnson, C.; Wilson, B.C. Increased expression of mitochondrial benzodiazepine receptors following low-level light treatment facilitates enhanced protoporphyrin IX production in glioma-derived cells in vitro. Lasers Surg Med. 2007, 39, 678–684. [Google Scholar] [CrossRef] [PubMed]
- Kessel, D.; Antolovich, M.; Smith, K.M. The Role of the Peripheral Benzodiazepine Receptor in the Apoptotic Response to Photodynamic Therapy. Photochemistry and Photobiology 2001, 74, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Sarissky, M.; Lavicka, J.; Kocanová, S.; Sulla, I.; Mirossay, A.; Miskovsky, P.; Gajdos, M.; Mojzis, J.; Mirossay, L. Diazepam enhances hypericin-induced photocytotoxicity and apoptosis in human glioblastoma cells. Vol. 52, NEOPLASMA. BAJZOVA 7, BRATISLAVA, 821 08, SLOVAKIA: AEPRESS SRO; 2005. p. 352–9.
- Chen, J.; Ouyang, Y.; Cao, L.; Zhu, W.; Zhou, Y.; Zhou, Y.; Zhang, H.; Yang, X.; Mao, L.; Lin, S.; Lin, J.; Hu, J.; Yan, G. Diazepam inhibits proliferation of human glioblastoma cells through triggering a G0/G1 cell cycle arrest. J Neurosurg Anesthesiol. 2013, 25, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Lavicka, J.; Sarisský, M.; Mirossay, A.; Sulla, I.; Mojzis, J.; Mirossay, L. Diazepam enhances etoposide-induced cytotoxicity in U-87 MG human glioma cell line. Fundam Clin Pharmacol. 2001, 15, 201–207. [Google Scholar] [CrossRef]
- Drljača, J.; Popović, A.; Bulajić, D.; Stilinović, N.; Vidičević Novaković, S.; Sekulić, S.; Milenković, I.; Ninković, S.; Ljubković, M.; Čapo, I. Diazepam diminishes temozolomide efficacy in the treatment of U87 glioblastoma cell line. CNS Neurosci Ther. 2022, 28, 1447–1457. [Google Scholar] [CrossRef]
- Czarnecka, M.; Lu, C.; Pons, J.; Maheswaran, I.; Ciborowski, P.; Zhang, L.; Cheema, A.; Kitlinska, J. Neuropeptide Y receptor interactions regulate its mitogenic activity. Neuropeptides. 2019, 73, 11–24. [Google Scholar] [CrossRef]
- Körner, M.; Reubi, J.C. Neuropeptide Y receptors in primary human brain tumors: Overexpression in high-grade tumors. J Neuropathol Exp Neurol. 2008, 67, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; He, X.; Wang, P.; Yuan, B.; Pan, Y.; Hu, X.; Lu, L.; Wu, A.; Li, J. A D-Y Shaped Neuropeptide Y Mimetic Peptide-Dye Self-Assembly with Maximal Emission Beyond 1300 nm and Glioma Mitochondrial Activity Modulation. Small. 2024, 20, e2308621. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Du, Y.; Jiang, Z.; Tian, Y.; Qiu, N.; Wang, Y.; Lqbal, M.Z.; Hu, M.; Zou, R.; Luo, L.; Du, S.; Tian, J.; Wu, A. Y1 receptor ligand-based nanomicelle as a novel nanoprobe for glioma-targeted imaging and therapy. Nanoscale. 2018, 10, 5845–5851. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Luo, Y.; Li, Y.; Pan, Y.; Kwok, R.T.K.; He, L.; Duan, X.; Zhang, P.; Wu, A.; Tang, B.Z.; Li, J. D-type neuropeptide decorated AIEgen/RENP hybrid nanoprobes with light-driven ROS generation ability for NIR-II fluorescence imaging-guided through-skull photodynamic therapy of gliomas. AGGREGATE. 111 RIVER ST, HOBOKEN 07030-5774, NJ USA: WILEY; 2023.
- Moriwaki, K.; Chan, F.K. RIP3: A molecular switch for necrosis and inflammation. Genes Dev. 2013, 27, 1640–1649. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, T.; Lei, T.; Zhang, D.; Du, S.; Girani, L.; Qi, D.; Lin, C.; Tong, R.; Wang, Y. RIP1/RIP3-regulated necroptosis as a target for multifaceted disease therapy (Review). Int J Mol Med. 2019, 44, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zheng, S.Y.; Jiang, R.L.; Wu, H.D.; Li, Y.A.; Lu, J.L.; Xiong, Y.; Han, B.; Lin, L. Necroptosis signaling and mitochondrial dysfunction cross-talking facilitate cell death mediated by chelerythrine in glioma. Free Radic Biol Med. 2023, 202, 76–96. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Wang, W.; Zhang, Y.; Fu, X.; Ping, K.; Zhao, J.; Lei, Y.; Mou, Y.; Wang, S. Synthesis and biological evaluation of celastrol derivatives as potential anti-glioma agents by activating RIP1/RIP3/MLKL pathway to induce necroptosis. Eur J Med Chem. 2022, 229, 114070. [Google Scholar] [CrossRef]
- Zhou, J.; Li, G.; Han, G.; Feng, S.; Liu, Y.; Chen, J.; Liu, C.; Zhao, L.; Jin, F. Emodin induced necroptosis in the glioma cell line U251 via the TNF-α/RIP1/RIP3 pathway. Invest New Drugs. 2020, 38, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Das, A.; McDonald, D.G.; Dixon-Mah, Y.N.; Jacqmin, D.J.; Samant, V.N.; Vandergrift, W.A., 3rd; Lindhorst, S.M.; Cachia, D.; Varma, A.K.; Vanek, K.N.; Banik, N.L.; Jenrette, J.M., 3rd; Raizer, J.J.; Giglio, P.; Patel, S.J. RIP1 and RIP3 complex regulates radiation-induced programmed necrosis in glioblastoma. Tumour Biol. 2016, 37, 7525–7534. [Google Scholar] [CrossRef]
- Coupienne, I.; Fettweis, G.; Rubio, N.; Agostinis, P.; Piette, J. 5-ALA-PDT induces RIP3-dependent necrosis in glioblastoma. Photochem Photobiol Sci. 2011, 10, 1868–1878. [Google Scholar] [CrossRef]
- Fettweis, G.; Di Valentin, E.; L’homme, L.; Lassence, C.; Dequiedt, F.; Fillet, M.; Coupienne, I.; Piette, J. RIP3 antagonizes a TSC2-mediated pro-survival pathway in glioblastoma cell death. Biochim Biophys Acta Mol Cell Res. 2017, 1864, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Maletínská, L.; Blakely, E.A.; Bjornstad, K.A.; Deen, D.F.; Knoff, L.J.; Forte, T.M. Human glioblastoma cell lines: Levels of low-density lipoprotein receptor and low-density lipoprotein receptor-related protein. Cancer Res. 2000, 60, 2300–2303. [Google Scholar] [PubMed]
- Han, L.; Jiang, C. Evolution of blood-brain barrier in brain diseases and related systemic nanoscale brain-targeting drug delivery strategies. Acta Pharm Sin B. 2021, 11, 2306–2325. [Google Scholar] [CrossRef] [PubMed]
- Pawar, S.; Koneru, T.; McCord, E.; Tatiparti, K.; Sau, S.; Iyer, A.K. LDL receptors and their role in targeted therapy for glioma: A review. Drug Discov Today. 2021, 26, 1212–1225. [Google Scholar] [CrossRef] [PubMed]
- Buzova, D.; Huntosova, V.; Kasak, P.; Petrovajova, D.; Joniova, J.; Dzurova, L.; Nadova, Z.; Sureau, F.; Miskovsky, P.; Jancura, D. Towards increased selectivity of drug delivery to cancer cells: Development of a LDL-based nanodelivery system for hydrophobic photosensitizers. Mohseni H, Agahi M, Razeghi M, editors. BIOSENSING AND NANOMEDICINE V. 1000 20TH ST, PO BOX 10, BELLINGHAM, WA 98227-0010 USA: SPIE-INT SOC OPTICAL ENGINEERING; 2012. (Proceedings of SPIE; vol. 8460).
- Huntosova, V.; Alvarez, L.; Bryndzova, L.; Nadova, Z.; Jancura, D.; Buriankova, L.; Bonneau, S.; Brault, D.; Miskovsky, P.; Sureau, F. Interaction dynamics of hypericin with low-density lipoproteins and U87-MG cells. Int J Pharm. 2010, 389, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Luiza Andreazza, N.; Vevert-Bizet, C.; Bourg-Heckly, G.; Sureau, F.; José Salvador, M.; Bonneau, S. Berberine as a photosensitizing agent for antitumoral photodynamic therapy: Insights into its association to low density lipoproteins. Int J Pharm. 2016, 510, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Huntosova, V.; Nadova, Z.; Dzurova, L.; Jakusova, V.; Sureau, F.; Miskovsky, P. Cell death response of U87 glioma cells on hypericin photoactivation is mediated by dynamics of hypericin subcellular distribution and its aggregation in cellular organelles. Photochem Photobiol Sci. 2012, 11, 1428–1436. [Google Scholar] [CrossRef]
- Song, L.; Li, H.; Sunar, U.; Chen, J.; Corbin, I.; Yodh, A.G.; Zheng, G. Naphthalocyanine-reconstituted LDL nanoparticles for in vivo cancer imaging and treatment. Int J Nanomedicine. 2007, 2, 767–774. [Google Scholar]
- Chen, L.Q.; Cheung, L.S.; Feng, L.; Tanner, W.; Frommer, W.B. Transport of sugars. Annu Rev Biochem. 2015, 84, 865–894. [Google Scholar] [CrossRef]
- Nabavizadeh, A.; Zhao, C.; Martinez, D.; Familiar, A.; Koptyra, M.; Ippolito, J.; Rubin, J.; Storm, P.B.; Resnick, A.C.; Santi, M. TMET-37. SODIUM-DEPENDENT GLUCOSE TRANSPORTER 2 (SGLT2) IS OVEREXPRESSED IN THE MAJORITY OF ATRTS AND A SUBSET OF PEDIATRIC PATIENTS WITH HIGH-GRADE GLIOMA; A POTENTIAL IMAGING AND THERAPEUTIC TARGET. Neuro Oncol. 2022 Nov 14;24(Suppl 7):vii270.
- Liu, Y.; Li, Y.M.; Tian, R.F.; Liu, W.P.; Fei, Z.; Long, Q.F.; Wang, X.A.; Zhang, X. The expression and significance of HIF-1alpha and GLUT-3 in glioma. Brain Res. 2009, 1304, 149–154. [Google Scholar] [CrossRef]
- Jensen, R.L.; Chkheidze, R. (2011). The Role of Glucose Transporter-1 (GLUT-1) in Malignant Gliomas. In: Hayat, M. (eds) Tumors of the Central Nervous System, Volume 1. Tumors of the Central Nervous System, vol 1. Springer, Dordrecht.
- Han, W.; Shi, J.; Cao, J.; Dong, B.; Guan, W. Emerging Roles and Therapeutic Interventions of Aerobic Glycolysis in Glioma. Onco Targets Ther. 2020, 13, 6937–6955. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, Z.; Blessington, D.; Li, H.; Busch, T.M.; Madrak, V.; Miles, J.; Chance, B.; Glickson, J.D.; Zheng, G. Pyropheophorbide 2-deoxyglucosamide: A new photosensitizer targeting glucose transporters. Bioconjug Chem. 2003, 14, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Guo, K.; Huang, B.; Lin, Z.; Cai, Z. Role of Glucose Transporters in Drug Membrane Transport. Curr Drug Metab. 2020, 21, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, M.; Kataoka, H.; Yano, S.; Ohi, H.; Moriwaki, K.; Akashi, H.; Taguchi, T.; Hayashi, N.; Hamano, S.; Mori, Y.; Kubota, E.; Tanida, S.; Joh, T. Antitumor effects in gastrointestinal stromal tumors using photodynamic therapy with a novel glucose-conjugated chlorin. Mol Cancer Ther. 2014, 13, 767–775. [Google Scholar] [CrossRef] [PubMed]
- Nishie, H.; Kataoka, H.; Yano, S.; Kikuchi, J.I.; Hayashi, N.; Narumi, A.; Nomoto, A.; Kubota, E.; Joh, T. A next-generation bifunctional photosensitizer with improved water-solubility for photodynamic therapy and diagnosis. Oncotarget. 2016, 7, 74259–74268. [Google Scholar] [CrossRef] [PubMed]
- Acker, G.; Palumbo, A.; Neri, D.; Vajkoczy, P.; Czabanka, M. F8-SIP mediated targeted photodynamic therapy leads to microvascular dysfunction and reduced glioma growth. J Neurooncol. 2016, 129, 33–38. [Google Scholar] [CrossRef] [PubMed]
- De Groof, T.W.M.; Mashayekhi, V.; Fan, T.S.; Bergkamp, N.D.; Sastre Toraño, J.; van Senten, J.R.; Heukers, R.; Smit, M.J.; Oliveira, S. Nanobody-Targeted Photodynamic Therapy Selectively Kills Viral GPCR-Expressing Glioblastoma Cells. Mol Pharm. 2019, 16, 3145–3156. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhou, H.; Liu, Y.; Wen, Y.; Wei, C.; Yu, Q.; Liu, J. Transferrin/aptamer conjugated mesoporous ruthenium nanosystem for redox-controlled and targeted chemo-photodynamic therapy of glioma. Acta Biomater. 2018, 82, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Akhlynina, T.V.; Jans, D.A.; Statsyuk, N.V.; Balashova, I.Y.; Toth, G.; Pavo, I.; Rosenkranz, A.A.; Naroditsky, B.S.; Sobolev, A.S. Adenoviruses synergize with nuclear localization signals to enhance nuclear delivery and photodynamic action of internalizable conjugates containing chlorin e6. Int J Cancer. 1999, 81, 734–740. [Google Scholar] [CrossRef]
- Wu, D.P.; Bai, L.R.; Lv, Y.F.; Zhou, Y.; Ding, C.H.; Yang, S.M.; Zhang, F.; Huang, J.L. A novel role of Cx43-composed GJIC in PDT phototoxicity: An implication of Cx43 for the enhancement of PDT efficacy. Int J Biol Sci. 2019, 15, 598–609. [Google Scholar] [CrossRef]
- An, Y.W.; Liu, H.Q.; Zhou, Z.Q.; Wang, J.C.; Jiang, G.Y.; Li, Z.W.; Wang, F.; Jin, H.T. Sinoporphyrin sodium is a promising sensitizer for photodynamic and sonodynamic therapy in glioma. Oncol Rep. 2020, 44, 1596–1604. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Luk, S.K.; Chuang, C.P.; Yip, S.P.; To, S.S.; Yung, Y.M. TP53 regulates human AlkB homologue 2 expression in glioma resistance to Photofrin-mediated photodynamic therapy. Br J Cancer. 2010, 103, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Oh, S.J.; Shim, J.K.; Ji, Y.B.; Moon, J.H.; Kim, E.H.; Huh, Y.M.; Suh, J.S.; Chang, J.H.; Lee, S.J.; Kang, S.G. C5α secreted by tumor mesenchymal stem-like cells mediates resistance to 5-aminolevulinic acid-based photodynamic therapy against glioblastoma tumorspheres. J Cancer Res Clin Oncol. 2023, 149, 4391–4402. [Google Scholar] [CrossRef] [PubMed]
- Shahmoradi Ghahe, S.; Kosicki, K.; Wojewódzka, M.; Majchrzak, B.A.; Fogtman, A.; Iwanicka-Nowicka, R.; Ciuba, A.; Koblowska, M.; Kruszewski, M.; Tudek, B.; Speina, E. Increased DNA repair capacity augments resistance of glioblastoma cells to photodynamic therapy. DNA Repair (Amst). 2021, 104, 103136. [Google Scholar] [CrossRef]
- Li, F.; Cheng, Y.; Lu, J.; Hu, R.; Wan, Q.; Feng, H. Photodynamic therapy boosts anti-glioma immunity in mice: A dependence on the activities of T cells and complement C3. J Cell Biochem. 2011, 112, 3035–3043. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



