Submitted:
19 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Hydrogen sulfide (H2S) is a novel gasotransmitter, sucrose (SUC) is both cellular energy and signaling molecule. However, their signaling interaction in maize thermotolerance is poorly known. In this study, using maize seedlings as materials, metabolic and functional interaction of H2S and SUC in maize thermotolerance was investigated. Data show that under heat stress survival rate and tissue viability were increased by exogenous SUC, while malondialdehyde content and electrolyte leakage were decreased by SUC, indicating SUC could upraise maize thermotolerance. Also, SUC-upraised thermotolerance was enhanced by H2S, while weakened by inhibitor (propargylglycine) and scavenger (hypotaurine) of H2S and SUC-transport inhibitor (N-ethylmaleimide). To figure out underlying mechanism of H2S-SUC interaction-upraised thermotolerance, redox parameters in mesocotyls of maize seedlings were measured before and after heat stress. The data display that activity and gene expression of H2S-metabolizing enzymes were up-regulated by SUC, whereas H2S had no significant effect on activity and gene expression of SUC-metabolizing enzymes. Besides, activity and gene expression of catalase, glutathione reductase, ascorbate peroxidase, peroxidase, dehydroascorbate reductase, monodehydroascorbate reductase, and superoxide dismutase were reinforced by H2S, SUC, and their combination under non-heat and heat conditions in varying degrees. Similarly, content of ascorbic acid, flavone, carotenoid, and polyphenol was increased by H2S, SUC, and their combination, whereas production of superoxide radical and hydrogen peroxide level were impaired by these treatments to different extent. These results imply that the metabolic and functional interaction of H2S and sucrose exists in the formation of maize thermotolerance through redox homeodynamics.
Keywords:
Hydrogen sulfide
; maize seedlings
; redox homeodynamics
; sucrose signaling
; thermotoleranceugh MG-scavenging system
; Calcium
; Maize seedling
; Methylglyoxal
; Methylglyoxal-scavenging system
; Thermotolerance
1. Introduction
Hydrogen sulfide (H2S) is a novel gasotransmitter, which participates in many physiological processes from seed germination to organ senescence [1,2]. H2S, similar to other signaling molecules, has a dual role of signaling and cytotoxin in a dose-dependent manner [2,3]. Therefore, action of H2S is strictly controlled by its homeostasis system, which includes enzymatic and non-enzymatic pathways. In plants, enzymatic pathways for H2S biosynthesis are involved in L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), O-acetyl-serine (thiol) lyase (OAS-TL), carbonic anhydrase (CA), mercaptopyruvate sulfur transferase (MST), sulfite reductase (SiR), -cyanoalanine synthase (CAS), and nitrogenase Fe-S cluster-like (NFS) [2,4]. These enzymes use L-/D-cysteine, carbonyl sulfide, and sulfite as substances to synthesize H2S. In non-enzymatic pathways for H2S production, H2S can be released from organic and inorganic compounds, such as persulfides and polysulfides, under the present of reducing agents like glutathione (GSH) [2,5]. Recent years, substantial studies reported that H2S as a signaling molecule modulates seed germination, stomatal movement, floral induction, organ senescence, and plant stress response [2,4,5,6]. Our studies also found that signaling molecule H2S was able to upraise maize thermotolerance, which is implicated in signaling crosstalk of H2S with calcium, nitric oxide, methylglyocal, and abscisic acid [2,7,8]. However, whether H2S-upraised maize thermotolerance is related to sucrose signaling remains unclear.
Sucrose (SUC) is a disaccharide, which is both cellular energy and signaling molecule, regulating the whole life processes [9,10,11]. In general, SUC can exert its signaling role in plants by multiple pathways, which are involved in hexose kinase-dependent and hexose kinase-independent signaling [12,13,14]. Also, SUC, as non-structural sugar, has a high solubility, exhibiting multiple physiological functions, such as energy, nutrient, osmotic adjustment substance, and reactive oxygen species (ROS) scavenger [15,16,17]. In addition, SUC, as a short- and long-distance signaling molecule, which regulates seed germination, seedling establishing, plant growth and development, as well as response and adaptation to environmental stresses [1,13,14,16,17]. In potato and rice plants, SUC treatment could boost thermotolerance by activating ROS-scavengering system [18,19], but the relationship of SUC-upraised thermotolerance with H2S signaling is little known. Also, in model plant Arabidopsis, Aroca et al. [20], using proteomics approach, found that at least 5% (approximately 2330 proteins) of the entire Arabidopsis proteome was able to be persulfidated. These persulfidated proteins are mainly involved in the enzymes related to SUC and starch metabolism, such as glycolysis and calvin cycle [20]. This study, from a point of view, hints the underlying crosstalk of SUC and H2S in plants.
Maize is the third food crop worldwide after rice and wheat, it, especially seedling stage, is sensitive to heat stress [7,21,22,23,24]. An increase in average 1℃ globally could lead to a reduction of maize yield by 7.4% [21,22]. With the exacerbation of global warming, heat stress has become a major stress factor that limits maize growth, development, and reproduction [25,26]. The underlying mechanism of plant thermotolerance has to be settled urgently to develop climate-resilient crops for sustainable agriculture. Therefore, in this paper, In this study, using maize seedlings as materials, the metabolic and functional interaction of H2S and SUC in maize thermotolerance was investigated. The purpose of this paper is to figure out the interaction of H2S and SUC in the development of maize thermotolerance by redox homeodynamics. molecular mechanism.
2. Results
2.1. H2S and SUC Upraises Thermotolerance
After heat stress and recovery, the survival rate of the maize seedlings was calculated and results were shown in Figure 1. Compared to the control, the survival rate of the seedlings irrigated with NaHS, SUC, and their combination after heat stress was significantly increased (Figure 1A, B), while MTL treatment had no significant effect on the survival rate (Figure 1B). Also, the survival rate was markedly improved by NEM combined with NaHS (Figure 1A), but NEM alone worsen an increase in the survival rate (Figure 1A) compared with the control. In addition, the increased SR by SUC was abolished by PAG + SUC and HT + SUC, and deteriorated by PAG and HT alone (Figure 1C). Similarly, compared with the control, under non-heat stress conditions (at 26℃), NaHS and SUC alone or in combination had no significant effect on MDA accumulation, electrolyte leakage, and tissue viability (TTC reduction) in maize seedlings (Figure 1D, E, F). Under heat stress conditions (at 46℃), an increase in MDA level and electrolyte leakage, as well as a decrease in tissue viability in maize seedlings were alleviated by NaHS and SUC alone or in combination compared to the control (Figure 1D, E, F).
2.2. SUC Upraises H2S Level
To study the possible effect of SUC on H2S level, an endogenous H2S and its metabolizing enzymes in maize seedlings were analyzed. The results shown in Figure 2, under non-heat stress conditions, compared with the control, LCD, DCD, and OAS-TL activities in maize seedlings were enhanced by SUC (Figure 2A, B, C). Also, LCD and OAS-TL activities were increased by NaHS alone or combined with SUC, which had no significant effect on DCD activity (Figure 2A, B, C). Similarly, a gene expression of ZmLCD1 and ZmOAS-TL1 in maize seedlings was up-regulated by SUC, while ZmOAS-TL1 expression was also enhanced by NaHS alone or combined with SUC, which was not significant impact on ZmLCD1 expression compared with the control (Figure 2D, E). In addition, compared with the control, an endogenous H2S level in maize seedlings was obviously enhanced by SUC and NaHS alone or in combination (Figure 2F).
Similarly, under heat stress conditions, compared to the control, LCD and OAS-TL activities in maize seedlings were enhanced by SUC, NaHS, and SUC + NaHS, which were not significant impact on DCD activity (Figure 2A, B, C). Moreover, a gene expression of ZmLCD1 in maize seedlings was significantly up-regulated by SUC, NaHS, and SUC + NaHS, which were not significant effect on ZmOAS-TL1 expression (Figure 2D, E). Analogously, compared to the control, an endogenous H2S level in maize seedlings was markedly increased by SUC, NaHS, and SUC + NaHS (Figure 2F).
2.3. H2S Modulates SUC Level
To further explore the effect of H2S on endogenous SUC level, SUC and its metabolizing enzymes in maize seedlings were estimated. The results show that, under non-heat stress conditions, SUS activity in maize seedlings was significant increased by SUC + NaHS, which was no significant impact on SPS activity compared with the control (Figure 3A, B). Also, compared to the control, a significant increase in SPS activity and SUC content in maize seedlings by SUC and NaHS alone or in combination was not observed (Figure 3A, C). Similarly, SUC and NaHS alone had no significant effect on SPS activity (Figure 3B). Moreover, SUN and NaHS alone or in combination had no significant impact on gene expression of ZmSPS1 and ZmSUS6 in maize seedlings compared with the control (Figure 3D, E).
Also, under heat stress conditions, compared with the control, SPS and SUS activities in maize seedlings were markedly reduced by SUC and SUC + NaHS, while NaHS alone had no significant effect on enzyme activity (Figure 3A, B). Similar to non-heat stress conditions, compared with the control, a significant up-regulation of gene expression of ZmSPS1 and ZmSUS6 in maize seedlings was not saw (Figure 3D, E). Besides, an endogenous SUC content in maize seedlings was enhanced by exogenous SUS, whereas NaHS and NaHS + SUC were not significant effect on endogenous SUC level compared with the control (Figure 3C).
2.4. H2S-SUC Interaction Enhances Antioxidant Capacity
To further illustrate the underlying mechanism of H2S-SUC interaction-upraised maize thermotolerance, cellular redox parameters in maize seedlings was tested. Under non-heat stress conditions, compared with the control, CAT and GR activities in maize seedlings were significant increased by exogenous SUC (Figure 4A, B). Similarly, CAT activity in maize seedlings was significant enhanced by NaSH (Figure 4A), while SUN and NaHS alone or in combination had no significant effect on APX, POD, DHAR, MDHAR, and SOD activities in maize seedlings (Figure 4C and Figure 5A–C). Also, compared with the control, a gene expression of ZmDHAR1 and ZmMDHAR1 in maize seedlings was significant up-regulated by SUN, NaHS, and SUC + NaHS (Fig. 5D, E), which had no significant effect on the expression of ZmCAT1, ZmGR1, and ZmSOD4 in maize seedlings (Figure 4D,E and Figure 5F).
Similarly, under heat stress, compared with the control, GR and DHAR activities in maize seedlings were increased by NaHS (Figure 4A and Figure 5B), which was not significant effect on CAT, APX, POD, and SOD activities (Figure 4A,C,F and Figure 5C), but reduced DHAR activity (Figs. 5A). Also, SUC alone or combined with NaHS had no significant impact on CAT, GR, APX, POD, DHAR, MDHAR, and SOD activities in maize seedlings compared with the control (Figure 4A–C,F and Figure 5B,C), except for combined treatment reduced DHAR activity (Figure 5A). Similarly, a gene expression of ZmMDHAR1 in maize seedlings was up-regulated by SUC, NaHS, and SUC + NaHS, which were not significant influence on ZmCAT1, ZmGR1, ZmDHAR1, and ZmSOD4 in maize seedlings compared with the control (Figure 4D,E and Figure 5D,F).
For antioxidant and secondary metabolites, under non-heat stress conditions, compared with the control, AsA and flavone contents in maize seedlings were enhanced by NaHS and NaHS + SUC (Figure 6A, D), which had no significant influence on total phenol content (Figure 6E). Similarly, compared to the control, flavone and carotenoid contents in maize seedlings were markedly increased by SUC (Figure 6D, B), and the later level was also enhanced by SUC + NaHS (Figure 6B). Also, SUC alone had no significant effect on AsA and total phenol contents in maize seedlings (Figure 6A, E), while the effect of NaHS on carotenoid and total phenol contents was able to observe similar results (Figure 6B, E).
Under heat stress conditions, compared to the control, AsA content in maize seedlings was significant enhanced by SUC and NaHS alone or in combination (Figure 6A), which had no significant effect on total phenol content, but maintained a high total phenol level (Figure 6A). Analogously, flavone and carotenoid contents in maize seedlings were obviously increased by SUC and SUC + NaHS, respectively (Figure 6B, D). However, compared to the control, a significant increase in flavone level in maize seedlings treated with NaHS and SUC + NaHS was not noted (Figure 6D), similar results were able to be recorded in maize seedlings treated with NaHS, but SUC reduced carotenoid level (Figure 6B).
2.5. H2S-SUC Interaction Regulates ROS Level
As stated above, H2S-SUC interaction was able to enhance cellular antioxidant capacity in maize seedlings under both non-heat and heat stress conditions (Figure 4, Figure 5 and Figure 6). To further figure out the influence of H2S-SUC interaction on the production of superoxide radical (O2.-) and hydrogen peroxide (H2O2) level in maize seedlings under non-heat and heat stress conditions, their production and level were measured. The results shown as Figure 7, under non-heat stress conditions, compared with the control, the production rate of superoxide radical and hydrogen peroxide level in maize seedlings were significantly reduced by SUC alone, and the later was also lowered by SUN + NaHS. However, NaHS alone had no significant effect on superoxide radical production and hydrogen peroxide level in maize seedlings (Figure 7A, B), similar results related to superoxide radical production was able to be noted in maize seedlings treated with SUC + NaHS (Figure 7A). Under heat stress conditions, superoxide radical production and hydrogen peroxide level in maize seedlings were reduced by SUC + NaHS (Figure 7A, B), and the former was also lowered by SUC alone (Figure 7A), while NaHS alone was not significant influence on superoxide radical production and hydrogen peroxide level (Figure 7A, B). Similarly, a significant difference of SUC alone on hydrogen peroxide level in maize seedlings was not observed (Figure 7B).
3. Discussion
In plants, H2S and SUC both are signaling molecules, which regulate many kinds of stress responses including heat stress [2,6,7,27]. However, whether exists in crosstalk of H2S and SUC in the formation of maize thermotolerance remains unclear. Therefore, in this study, using physiological, biochemical, and molecular approaches, the metabolic and functional interaction of H2S with SUC in the development of thermotolerance in maize seedlings was found (Figure 1). Their interactions are involved in the mutual regulation on the activity and gene expression of metabolic enzymes (Figure 2 and Figure 3) and antioxidant enzymes (Figure 4 and Figure 5), as well as on the content of non-enzymatic antioxidants and secondary metabolites (Figure 6).
In general, the acquirement of plant thermotolerance is an intricately physiological, biochemical, and molecular process, involving in the crosstalk among signaling pathways [2,25,28,29,30]. In this study, under non-heat stress conditions, SUC treatment up-regulated gene expression of ZmLCD1 and ZmOATL1 (Figure 2D, E), which in turn increased the activity of LCD, DCD, and OAS-TL (Figure 2A, B, C), thus accumulating endogenous H2S in maize seedlings (Figure 2F). Similarly, under heat stress conditions, gene expression of ZmLCD1 (Figure 2D), activity of LCD and OAS-TAL (Figure 2A, C), as well as endogenous H2S level (Figure 2F) were up-regulated by SUC. On the other hand, H2S treatment had no significant effect on gene expression of ZmSPS1 and ZmSUS6, activity of SPS and SUS, as well as endogenous SUC content in maize seedlings under both non-heat and heat stress conditions (Figure 3). Also, Pearson correlation analysis shows that H2S content was significantly positive relationship with SUC level and SUS activity (Table 1). Adversely, SUC content was also significantly positive correlation to LCD and OAS-TL activities and endogenous H2S level (Table 1). These data indicate that SUC was able to trigger H2S signaling in maize seedlings under both non-heat and heat stress conditions, that is, H2S might exert its signaling role in the downstream of SUC signaling.
Substantial studies have shown that heat stress can lead to multiple damages, such as protein denaturation, biomembrane damage, osmotic stress, and oxidative stress [2,7,30]. Among damages, oxidative stress is a major determinant, which can further cause other damages [2,25,30,31]. Therefore, the acquirement of plant thermotolerance is closely associated with the mitigation of oxidative stress [16,27,32,33,34,35]. In this study, to further explore the underlying mechanism of SUC-H2S interaction-upraised thermotolerance in maize seedlings, antioxidant enzymes, non-enzymatic antioxidants, and secondary metabolites were analyzed. Under non-heat stress conditions, compared with the control, SUC treatment increased CAT and GR activities (Figure 4A, B), up-regulated ZmDHAR1 and ZmMDHAR1 expression (Figure 5A, B), accumulated flavone and carotenoids (Figure 6D, B), thus reducing the production rate of superoxide radical in maize seedlings (Figure 6C, F). Similarly, CAT activity (Figure 4A), ZmMDHAR1 expression (Figure 5B) as well as AsA and flavone levels (Figure 6A, D) in maize seedlings were elevated by H2S. Also, MDHAR activity, ZmMDHAR1 expression, as well as AsA, flavone, and carotenoid contents were upraised by SUC in combination with H2S in maize seedlings. Taken together, before heat stress, the enhanced antioxidant enzymes, non-enzymatic antioxidants, and secondary metabolites by SUC and H2S alone or in combination lay the physiological, biochemical, and molecular foundation for the development of subsequent thermotolerance in maize seedlings.
Under heat stress condition, ZmMDHAR1 gene expression (Figure 5B) as well as AsA and flavone contents (Figure 6A, D) in maize seedlings were increased by SUC, which in turn decreased the production rate of superoxide radical (Figure 6C). Analogously, GR and MDHAR activity (Figure 4B and Figure 5B), ZmMDHAR1 expression (Figure 5B), and AsA content (Figure 6A) in maize seedlings were augmented by H2S. Moreover, SUN in combination with H2S up-regulated ZmMDHAR1 expression (Figure 5B), and increased AsA and carotenoid contents (Figure 6A,B) in maize seedlings, thus decreasing superoxide radical production (Figure 6C) and hydrogen peroxide accumulation (Figure 6F). In addition, Pearson correlation analysis indicates that the survival rate was markedly positive correlation with CAT, GR, DHAR, and MDHAR activities (Table 2). Similarly, the survival rate was significantly positive relationship with AsA, flavone, carotenoid, and hydrogen peroxide levels, while negative correlation to the production of superoxide radical (Table 3). These data imply that the maize seedlings treated with SUC and H2S alone or in combination maintained a high antioxidant capacity under heat stress conditions, thus reducing oxidative damage and improving maize thermotolerance (Figure 1). In rice seedlings, foliar-sprayed with SUC increased CAT activity, as well as SUC, total soluble sugar, and NAD+ contents, which in turn reduced MDA and hydrogen peroxide accumulation, thus improving photosynthetic efficiency and increasing thermotolerance in rice seedlings [18]. Also, in potato plants, SUC treatment increased SOD, POD, CAT, and APX activities, as well as SUC, total sugar, proline, and soluble protein contents, followed by decrease in hydrogen peroxide level, thus alleviating oxidative damage and osmotic stress caused by heat stress [19]. These studies further support our current hypothesis that metabolic and functional interaction of H2S and sucrose exists in maize thermotolerance through redox homeodynamics.
4. Materials and Methods
4.1. Seed Germination and Seedling Treatment
In this work, materials maize (Zea mays L., cv. Diyu No. 401) seeds were purchased from Diyu Seed Company, China, and then immersed in 5% sodium hypochlorite solution for 10 min to sterilize. The sterilized seeds were imbibed in distilled water at 26℃ for 12 h after washing. The imbibed seeds were germinated for 60 h (namely 2.5 d) at 26℃on the eight-layer filter papers (which were wetted with distilled water) in trays with covers (approximately 250 seeds per tray). After germination, the 2.5 d-old seedlings were classified into 11 groups, which were irrigated with 90 mL of solutions listed as follows for 12 h, respectively. (1) distilled water (control, CK), (2) 500 M NaHS (NaHS), (3) 25 mM sucrose (SUC), (4) 25 mM mannitol (MTL), (5) 300 M N-ethylmaleimide (NEM), (6) 300 M NEM + 500 M NaHS (NEM + NaHS), (7) 25 mM SUC + 500 M NaHS (SUC + NaHS), (8) 300 M propargylglycine + 25 mM SUC (PAG + SUC), (9) 300 M hydroxylamine + 25 mM SUC (HA + SUC), (10) 300 M PAG (PAG), (11) 300 M HT (HT).
Based on the previous reports, NaHS, PAG, and HT are donor, inhibitor, and scavenger of H2S [36,37], while NEM is a SUC transport inhibitor [38]. An appropriate concentration of these pharmacological reagents was selected from the preliminary experiments and previous reports [36,39,40]. After treatment with reagents for 12 h, the treated seedlings were executed heat stress at 46℃ for 16 h. After heat stress, the seedling mesocotyls (the most sensitive organ to heat stress [41]) were cut and used to estimate the following parameters.
4.2. Estimation of Thermotolerance Indexes
To illustrate the effect of H2S-SUC interaction on maize thermotolerance, after heat stress and recovery at 26℃ for a week, thermotolerance indexes (survival rate, tissue viability, malondialdehyde (MDA) content, and electrolyte leakage) were estimated as per the report by Wang et al. [7]. Survival rate and electrolyte leakage were indicated by %, whereas MDA content and tissue viability were expressed by nmol g-1 fresh weight (FW) and A485, respectively.
4.3. Measurement of Metabolizing Enzymes and Endogenous Content of H2S
To study the effect of SUC on the metabolizing enzyme activity and endogenous level of H2S, the maize seedlings were irrigated with SUC and NaHS alone or in combination, and then exposed to heat stress. H2S content and LCD, DCD, and OAS-TL activities in mesocotyls of maize seedlings were measured based on the methods of Ye et al. [8] and Sun et al. [41]. Enzyme activity was indicated by nmol min-1 g-1 FW, while H2S content was expressed by mol g-1 FW. Aslo, gene expression of ZmLCD1 and ZmOAS-TL was quantified by qRT-PCR and Zea mays beta-5 tubulin (ZmTUB) was used as internal reference. Expression level was counted using the 2-∆∆CT [7] and gene primers were listed in supplement 1.
4.4. Analysis of Metabolizing Enzymes and Endogenous Content of SUC
Also, to investigate the effect of H2S on the metabolizing enzyme activity and endogenous content of H2S, which were analyzed after the maize seedlings were irrigated with SUC and NaHS alone or in combination and subsequently subjected to heat stress. Extraction and analysis of SPS, SUS, and SUC in mesocotyls of maize seedlings refer to the procedures of Zhu et al. [42]. SUC level and SPS activity was analyzed as resorcinol methods, the later was determined by measuring the formation of sucrose (plus sucrose-6-P) in reaction mixture. Also, analysis of SUS activity was in accordance with SPS except that pH in reaction buffer was changed to 8.5, and fructose was replaced by fructose-6-P. Enzyme activity and SUC content were expressed by mg min-1 g-1 FW and mg g-1 FW, respectively. Also, gene expression of ZmSPS1 and ZmSUS6 was quantified and calculated according to the procedures mentioned above.
4.5. Evaluation of Antioxidant Capacity
To further figure out the underlying mechanism of H2S-SUC interaction-upraised maize thermotolerance, antioxidant enzymes, non-enzymatic antioxidants, and secondary metabolites were evaluated before and after heat stress. Antioxidant enzymes (catalase, CAT; glutathione reductase, GR; ascorbate peroxidase, APX; guaiacol peroxidase, POD; dehydroascorbate reductase, DHAR; monodehydro- ascorbate reductase, MDHAR; and superoxide dismutase, SOD) activities were evaluated according to the extraction and analysis procedures of Wang et al. [7] and Sun et al. [41]. An extinction coefficient (40 nm mM-1 cm-1 at 240 nm, 6.2 nm mM-1 cm-1 at 340 nm, 2.8 nm mM-1 cm-1 at 290 nm, 26.6 nm mM-1 cm-1 at 470 nm, 14.0 nm mM-1 cm-1 at 265 nm, 14.0 nm mM-1 cm-1 at 265 nm, and 21.6 nm mM-1 cm-1 at 560 nm) was used to calculate the activity of corresponding enzymes (CAT, GR, APX, POD, DHAR, MDHAR, and SOD), respectively. Enzyme activity was expressed by nmol min-1 g-1 FW.
Also, gene expression of ZmCAT1, ZmGR1, ZmDHAR1, ZmMDHAR1, and ZmSOD4 was quantified and calculated according to the methods stated above. In addition, ascorbic acid (AsA), flavonone, carotenoids, and total phenol in mesocotyls of maize seedlings were evaluated based on the reports by Wang et al. [7]. Their contents were indicated by nmol g-1 FW and g g-1 FW, respectively.
4.6. Assay for ROS
To explore the effect of H2S-SUC interaction on the production rate of superoxide radical (O2.-) and hydrogen peroxide (H2O2) level in mesocotyls of maize seedlings, before and after heat stress, their production and level were assayed. Superoxide radical (O2.-) production and hydrogen peroxide (H2O2) level were separately determined using Na,3-[1-[(phenylamino)-carbonyl]-3, 4-tetrazolium] (4-methoxy-6- nitro) benzene sulfonic acid hydrate method [41] and titanium sulfate method [7]. An extinction coefficient (21.6 mM-1 cm-1 at 470 nm and 0.28 mM-1 cm-1 at 410 nm) was used to calculated superoxide radical production and hydrogen peroxide level, which were expressed by nmol min-1 g-1 FW and nmol g-1 FW, respectively.
4.7. Statistical Analysis
Experiments were carried out based on a random design, and data analysis was executed as per one-way analysis of variance (ANOVA). Duncan multiple-range test was used to estimate significant difference among data, and the same letters on the bars in figures denote no significant difference, whereas the different letters denote significant difference. The data in figures indicate means ± standard error (SE). In addition, Pearson correlation analysis was performed using sigmaplot 15, and the asterisk (*) and double asterisks (**) in tables denote significant (p ˂ 0.05) and very significant (p ˂ 0.01) difference.
5. Conclusions
Taken together, the findings were shown in Figure 8, heat stress leads to a decrease in the survival rate of maize seedlings under heat stress. However, SUC and H2S alone or in combination increased the survival rate of maize seedlings under heat stress conditions, and SUC-increased survival rate was weakened by SUC transport inhibitor (NEM) and inhibitor (PAG) and scavenger (HT) of H2S, indicating the interaction of H2S and SUC in the development of maize thermotolerance. Further, SUC treatment, to different extent, up-regulated LCD, DCD, and OAS-TL activities and corresponding gene expression, as well as endogenous H2S level in maize seedlings under non-heat and heat stress conditions, further supporting the fact that H2S interacted with SUC. Also, SUC and H2S alone or in combination, to varying degrees, enhanced antioxidant enzyme activity and their gene expression, as well as non-enzymatic antioxidant level including secondary metabolites in maize seedlings under non-heat and heat stress conditions. These data illustrate that the metabolic and functional interaction of H2S and SUC exists in the formation of maize thermotolerance by cellular redox homeodynamics.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
Conceptualization, Z.G.L. and X.E.L.; methodology, X.E.L.; validation, X.E.L., and H.D.Z.; formal analysis, X.E.L.; investigation, X.E.L.; data curation, X.E.L.; writing—original draft preparation, Z.G.L.; writing—review and editing, Z.G.L., and X.E.L.; funding acquisition, Z.G.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (32160065).
Informed Consent Statement
Not applicable.
Data Availability Statement
The study did not require ethical approval.
Data Availability Statement
All data are displayed in the manuscript and Supplementary Materials.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Choudhary, A.; Kumar, A.; Kaur, N.; Kaur, H. Molecular cues of sugar signaling in plants. Physiol. Plant. 2022, 174, e13630. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. G.; Fang, J. R.; Bai, S. J. Hydrogen sulfide signaling in plant response to temperature stress. Front. Plant Sci. 2024, 15, 1337250. [Google Scholar] [CrossRef] [PubMed]
- Lee, Z.; Lim, J.; Harikrishna, J.; Islam, T.; Rahim, M. H. A.; Yaacob, J. S. Regulation of plant responses to temperature stress: A key factor in food security and for mitigating effects of climate change. Int. J. Plant Prod. 2024, 18. In press. [Google Scholar] [CrossRef]
- Yi, B.; Liu, Y.; Wu, Z.; Zheng, Y.; Chen, H.; Jin, P. Hydrogen sulfide alleviates chilling injury of zucchini fruit by regulating antioxidant capacity, endogenous hydrogen sulfide, proline, and polyamine metabolism. Postharv. Biol. Technol. 2024, 208, 112638. [Google Scholar] [CrossRef]
- Alvi, A. F.; Khan, S.; Khan, N. A. Hydrogen sulfide and ethylene regulate sulfur-mediated stomatal and photosynthetic responses and heat stress acclimation in rice. Plant Physiol. Biochem. 2024, 207, 108437. [Google Scholar] [CrossRef] [PubMed]
- Corpas, F. J.; Muñoz-Vargas, M. A.; González-Gordo, S.; Rodríguez-Ruiz, M.; Palma, J. M. Nitric oxide (NO) and hydrogen Sulfide (H2S): New potential biotechnological tools for postharvest storage of horticultural crops. J. Plant Growth Regul. 2024, 43. In press. [Google Scholar] [CrossRef]
- Wang, J. Q.; Xiang, R. H.; Li, Z. G. The essential role of H2S-ABA crosstalk in maize thermotolerance through the ROS-scavenging system. Int. J. Mol. Sci. 2023, 24, 12264. [Google Scholar] [CrossRef] [PubMed]
- Ye, X. Y.; Qiu, X. M.; Sun, Y. Y.; Li, Z. G. Interplay between hydrogen sulfide and methylglyoxal initiates thermotolerance in maize seedlings by modulating reactive oxidative species and osmolyte metabolism. Protoplasma 2020, 257, 1415–1432. [Google Scholar] [CrossRef] [PubMed]
- Hajibehzad, S. S.; Romanowski, A.; Pierik, R. Plant signaling: The sugar-coated story of root growth. Curr. Biol. 2023, 33, R805–R808. [Google Scholar] [CrossRef]
- Asad, M. A. U.; Yan, Z.; Zhou, L.; Guan, X.; Cheng, F. How abiotic stresses trigger sugar signaling to modulate leaf senescence? Plant Physiol. Biochem. 2024, 210, 108650. [Google Scholar] [CrossRef]
- Yamada, K.; Mine, A. Sugar coordinates plant defense signaling. Sci. Adv. 2024, 10, eadk4131. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, I. Z. Role of sugars in abiotic stress signaling in plants. In: Khan, M. I. R.; Reddy, P. S.; Ferrante, A.; Khan, N. A (Eds.) Plant Signaling Molecules, Woodhead Publishing, 2019, pp. 207-217.
- Raza, A.; Bhardwaj, S.; Rahman, M. A.; García-Caparrós, P.; Habib, M.; Saeed, F.; Charagh, S.; Foyer, C. H.; Siddique, K. H. M.; Varshney, R. K. Trehalose: A sugar molecule involved in temperature stress management in plants. Crop J. 2024, 12, 1–16. [Google Scholar] [CrossRef]
- Zou, W.; Yu, Q.; Ma, Y.; Sun, G.; Feng, X.; Ge, L. Pivotal role of heterotrimeric G protein in the crosstalk between sugar signaling and abiotic stress response in plants. Plant Physiol. Biochem. 2024, 210, 108567. [Google Scholar] [CrossRef]
- Mishra, B. S.; Sharma, M.; Laxmi, A. Role of sugar and auxin crosstalk in plant growth and development. Physiol. Plant. 2022, 174, e13546. [Google Scholar] [CrossRef] [PubMed]
- Misra, V.; Mall, A. K.; Ansari, S. A.; Ansari, M. I. Sugar transporters, sugar-metabolizing enzymes, and their interaction with phytohormones in sugarcane. J. Plant Growth Regul. 2023, 42, 4975–4988. [Google Scholar] [CrossRef]
- Sharma, P.; Kapoor, N.; Dhiman, S.; Kour, J.; Singh, A. D.; Sharma, A.; Bhardwaj, R. Role of sugars in regulating physiological and molecular aspects of plants under abiotic stress. In: Sharma, A.; Pandey, S.; Bhardwaj, R.; Zheng, B.; Tripathi, D. K. (Eds.) The Role of Growth Regulators and Phytohormones in Overcoming Environmental Stress. Academic Press, 2023, pp.355-374.
- Chen, Q. Q. The effects of exogenous sucrose on tolerance in potato ender heat and chilling. Lanzhou University of Technology, 2017.
- Cisse, A. Role of sucrose in heat response in rice plants. Chinese Academy of Agricultural Sciences, 2020.
- Aroca, A.; Benito, J. M.; Gotor, C.; Romero, L. C. Persulfidation proteome reveals the regulation of protein function by hydrogen sulfide in diverse biological processes in Arabidopsis. J. Exp. Bot. 2017, 68, 4915–4927. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Howell, S. H. Heat stress responses and thermotolerance in maize. Int. J. Mol. Sci. 2021, 22, 948. [Google Scholar] [CrossRef] [PubMed]
- Chandra, A. K.; Joshi, A.; Tripathi, A.; Kumar, A.; Pandey, S.; Singh, A.; Lal, D.; Bharati, A.; Adhikari, S.; Dinkar, V. Climate-resilience maize: Heat stress, signaling, and molecular interventions. J. Plant Growth. Regul. 2023, 42, 6349–6366. [Google Scholar] [CrossRef]
- Djalovic, I.; Kundu, S.; Bahuguna, R. N.; Pareek, A.; Raza, A.; Singla-Pareek, S. L.; Prasad, P. V. V.; Varshney, R. K. Maize and heat stress: Physiological, genetic, and molecular insights. Plant Genome 2024, 17, e20378. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, Y.; Wu, W.; Lyu, L.; Li, W. A review of changes at the phenotypic, physiological, biochemical, and molecular levels of plants due to high temperatures. Planta 2024, 259, 57. [Google Scholar] [CrossRef]
- Fortunato, S.; Lasorella, C.; Dipierro, N.; Vita, F.; de Pinto, M. C. Redox signaling in plant heat stress response. Antioxidants 2023, 12, 605. [Google Scholar] [CrossRef] [PubMed]
- Sehar, Z.; Mir, I. R.; Khan, S.; Masood, A.; Khan, N. A. Nitric oxide and proline modulate redox homeostasis and photosynthetic metabolism in wheat plants under high temperature stress acclimation. Plants 2023, 12, 1256. [Google Scholar] [CrossRef]
- Sharma, G.; Sharma, N.; Ohri, P. Harmonizing hydrogen sulfide and nitric oxide: A duo defending plants against salinity stress. Nitric Oxide 2024, 144, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, R.; Rajput, V. D.; Ghodake, G.; Ahmad, F.; Meena, M.; Rehman, R.; Prasad, R.; Sharma, R. K.; Singh, R.; Seth, C. S. Comprehensive journey from past to present to future about seed priming with hydrogen peroxide and hydrogen sulfide concerning drought, temperature, UV and ozone stresses- a review. Plant Soil 2024, 498. In press. [Google Scholar] [CrossRef]
- Huang, Y.; An, J.; Sircar, S.; Bergis, C.; Lopes, C. D.; He, X.; Costa, B. D.; Tan, F. Q.; Bazin, J.; Antunez-Sanchez, J.; Mammarella, M. F.; Devani, R. S. HSFA1a modulates plant heat stress responses and alters the 3D chromatin organization of enhancer-promoter interactions. Nat. Commun. 2023, 14, 469. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. G. Sugar signaling in plants under physiological and stress conditions. In: Hasanuzzaman, M.; Hawrylak-Nowak, B.; Islam, T.; Fujita, M. (Eds.) Biostimulants for Crop Production and Sustainable Agriculture. CABI, Wallingford, 2022, pp. 372-385.
- Hernandez-Leon, S. G.; Valenzuela-Soto, E. M. Glycine betaine is a phytohormone-like plant growth and development regulator under stress conditions. J. Plant Growth Regul. 2023, 42, 5029–5040. [Google Scholar] [CrossRef]
- Raza, A.; Charagh, S.; Abbas, S.; Hassan, M. U.; Saeed, F.; Haider, S.; Sharif, R.; Anand, A.; Corpas, F. J.; Jin, W.; Varshney, R. K. Assessment of proline function in higher plants under extreme temperatures. Plant Biol. 2023, 25, 379–395. [Google Scholar] [CrossRef]
- Alvarez, M. E.; Savouré, A.; Szabados, L. Proline metabolism as regulatory hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Vuković, M.; Kutnjak, M.; Vitko, S.; Tkalec, M.; Vidaković-Cifrek, Z. Heat priming modifies heat stress response in BPM1-overexpressing Arabidopsis thaliana (L.) Heynh. J. Plant Growth Regul. 2024, 43, in, in press. [Google Scholar] [CrossRef]
- Li, Z. G.; Yang, S. Z.; Long, W. B.; Yang, G. X.; Shen, Z. Z. Hydrogen sulfide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize (Zea mays L.) seedlings. Plant Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef] [PubMed]
- Gautam, H.; Fatma, M.; Sehar, Z.; Mir, I. R.; Khan, N. A. Hydrogen sulfide, ethylene, and nitric oxide regulate redox homeostasis and protect photosynthetic metabolism under high temperature stress in rice plants. Antioxidants 2022, 11, 1478. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Pan, W.; Xin, Y.; Wu, J.; Li, R.; Shi, J.; Long, S.; Qu, L.; Yang, Y.; Yi, M.; Wu, J. Regulating bulb dormancy release and flowering in lily through chemical modulation of intercellular communication. Plant Methods 2023, 19, 136. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z. H.; Wang, Y.; Ye, X. Y.; Li, Z. G. Signaling molecule hydrogen sulfide improves seed germination and seedling growth of maize (Zea mays L.) under high temperature by inducing antioxidant system and osmolyte biosynthesis. Front. Plant Sci. 2018, 9, 1288. [Google Scholar] [CrossRef] [PubMed]
- Li, Z. G.; Xie, L. R.; Li, X. J. Hydrogen sulfide acts as a downstream signal molecule in salicylic acid-induced heat tolerance in maize (Zea mays L.) seedlings. J. Plant Physiol. 2015, 177, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y. Y.; Wang, J. Q.; Xiang, R. H.; Li, Z. G. Key role of reactive oxygen species-scavenging system in nitric oxide and hydrogen sulfide crosstalk-evoked thermotolerance in maize seedlings. Front. Plant Sci. 2022, 13, 967968. [Google Scholar] [CrossRef]
- Zhu, J.; Qi, J.; Fang, Y.; Xiao, X.; Li, J.; Lan, J.; Tang, C. Characterization of sugar contents and sucrose metabolizing enzymes in developing leaves of Hevea brasiliensis. Front. Plant Sci. 2018, 9, 58. [Google Scholar] [CrossRef]
Figure 1.
Effect of pretreatment with NaHS, sucrose (SUC), mannitol (MTL), N-ethylmaleimide (NEM), propargylglycine (PAG), hydroxylamine (HT) alone or in combination on the survival rate (A,B,C, %), malondialdehyde (MDA, D), electrolyte leakage (E), and tissue viability (F) of maize seedlings under heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 1.
Effect of pretreatment with NaHS, sucrose (SUC), mannitol (MTL), N-ethylmaleimide (NEM), propargylglycine (PAG), hydroxylamine (HT) alone or in combination on the survival rate (A,B,C, %), malondialdehyde (MDA, D), electrolyte leakage (E), and tissue viability (F) of maize seedlings under heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
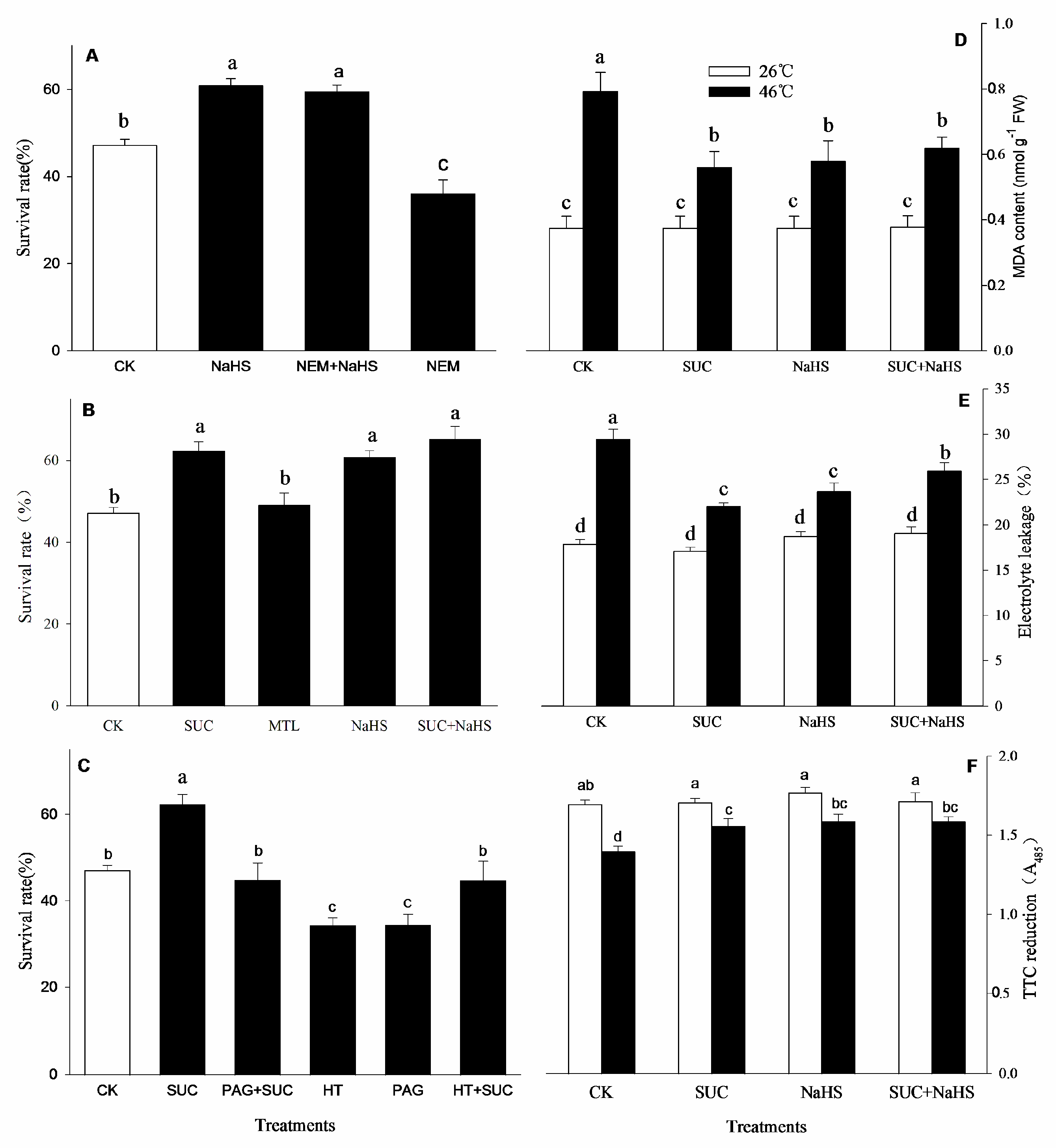
Figure 2.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on L-cysteine desulfhydrase (LCD, A), D-cysteine desulfhydrase (DCD, B), and O-acetyl-serine (thiol) lyase (OAS-TL, C) activities, gene expression of ZmLCD1 (D) and ZmOAS-TL (E), and endogenous H2S content (F) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 6), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 2.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on L-cysteine desulfhydrase (LCD, A), D-cysteine desulfhydrase (DCD, B), and O-acetyl-serine (thiol) lyase (OAS-TL, C) activities, gene expression of ZmLCD1 (D) and ZmOAS-TL (E), and endogenous H2S content (F) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 6), the same and different letters on the bars indicate no significant and significant difference, respectively.
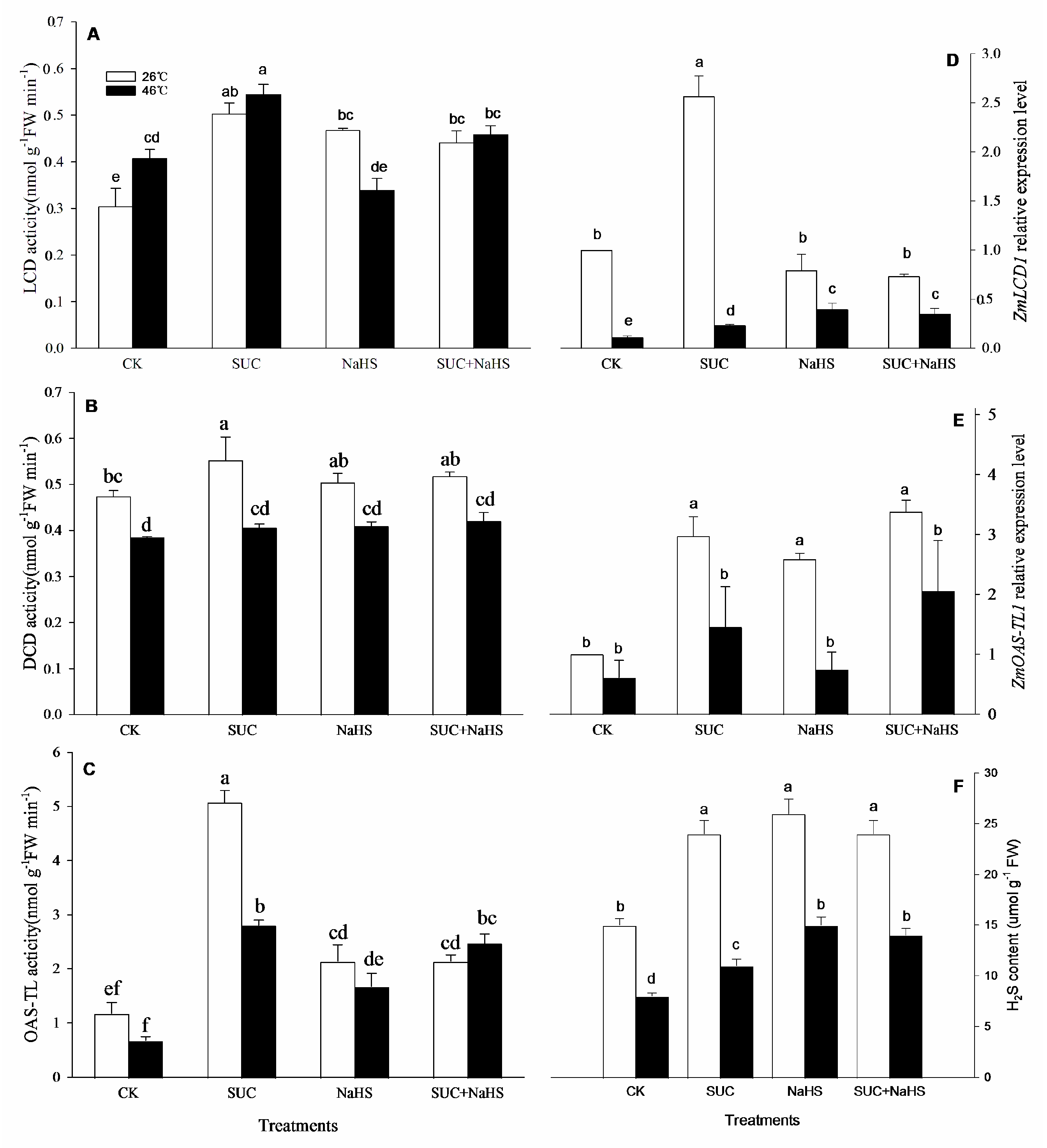
Figure 3.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on sucrose-P synthase (SPS, A) and sucrose synthase (SUS, B) activities, SUC content (C), and gene expression of ZmSPS1 (D) and ZmSUS6 (E) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 5), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 3.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on sucrose-P synthase (SPS, A) and sucrose synthase (SUS, B) activities, SUC content (C), and gene expression of ZmSPS1 (D) and ZmSUS6 (E) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 5), the same and different letters on the bars indicate no significant and significant difference, respectively.
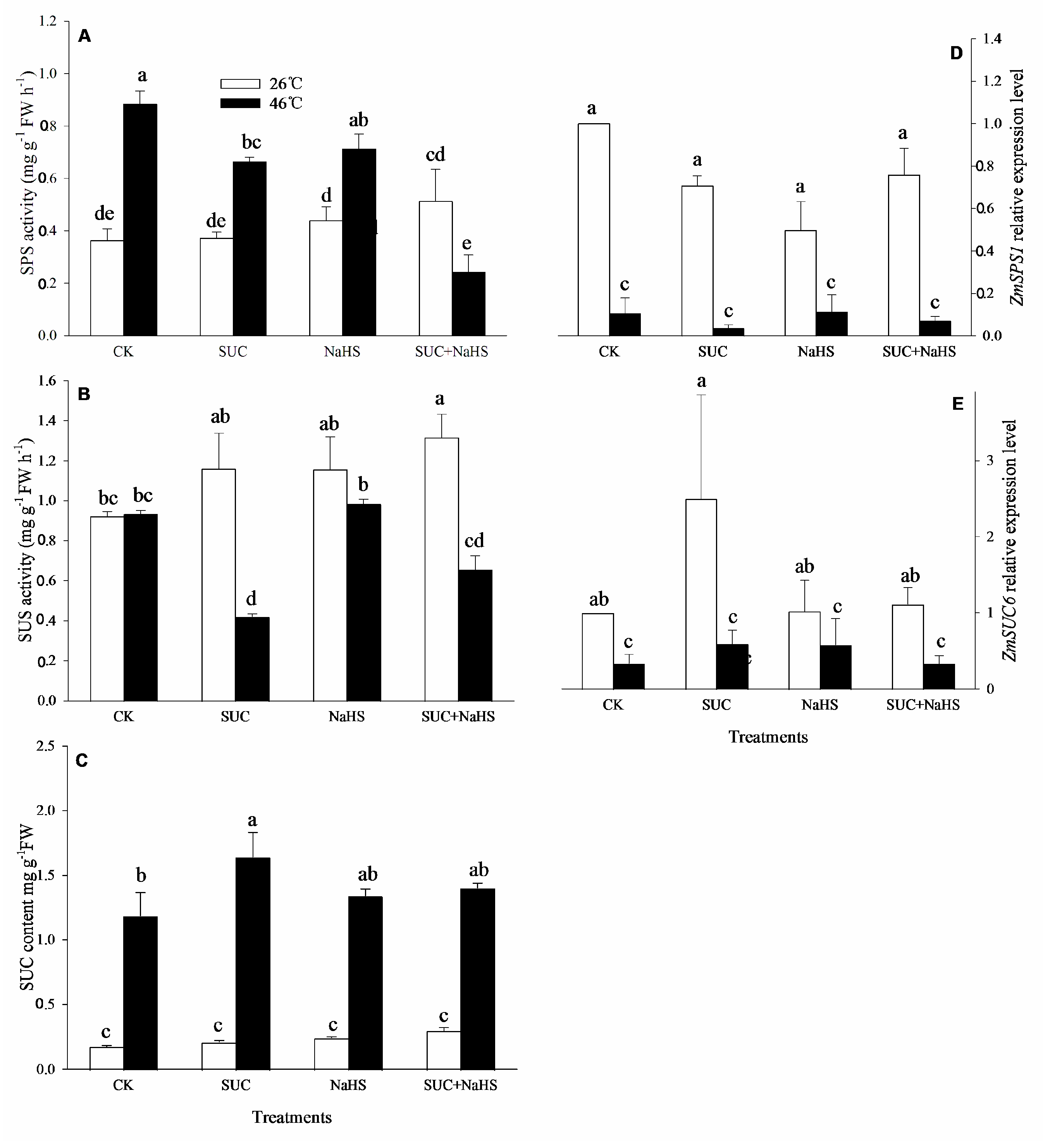
Figure 4.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on catalase (CAT, A), glutathione reductase (GR, B), ascorbate peroxidase (APX, C), and peroxidase (POD, F) activities and gene expression of ZmCAT (D) and ZmGR1 (E) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 5), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 4.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on catalase (CAT, A), glutathione reductase (GR, B), ascorbate peroxidase (APX, C), and peroxidase (POD, F) activities and gene expression of ZmCAT (D) and ZmGR1 (E) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 5), the same and different letters on the bars indicate no significant and significant difference, respectively.
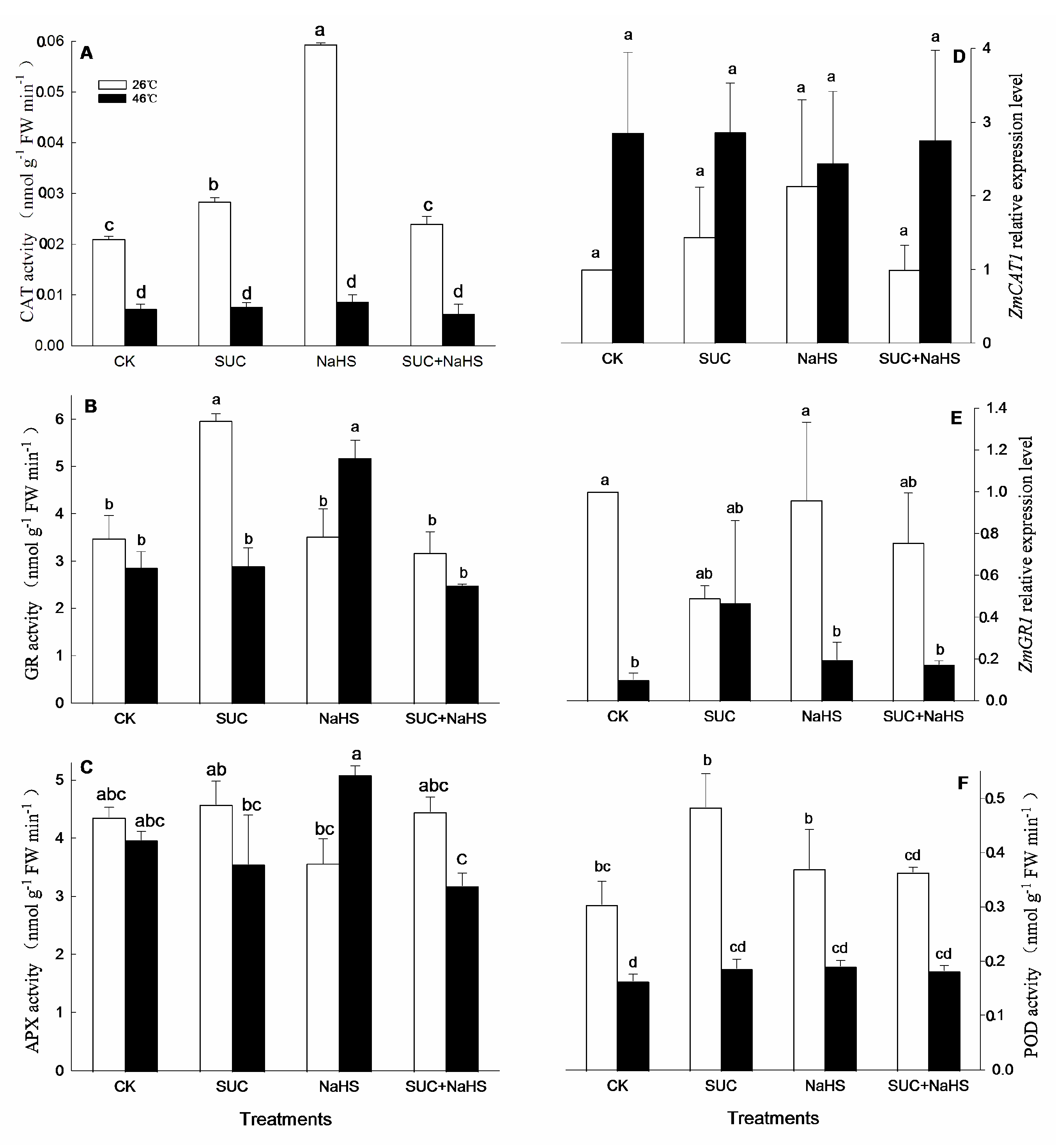
Figure 5.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on dehydroascorbate reductase (DHAR, A), monodehydroascorbate reductase (MDHAR, B), and superoxide dismutase (SOD, C) activities and gene expression of ZmDHAR1 (D), ZmMDHAR1 (E), and ZmSOD4 (F) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 5.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on dehydroascorbate reductase (DHAR, A), monodehydroascorbate reductase (MDHAR, B), and superoxide dismutase (SOD, C) activities and gene expression of ZmDHAR1 (D), ZmMDHAR1 (E), and ZmSOD4 (F) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
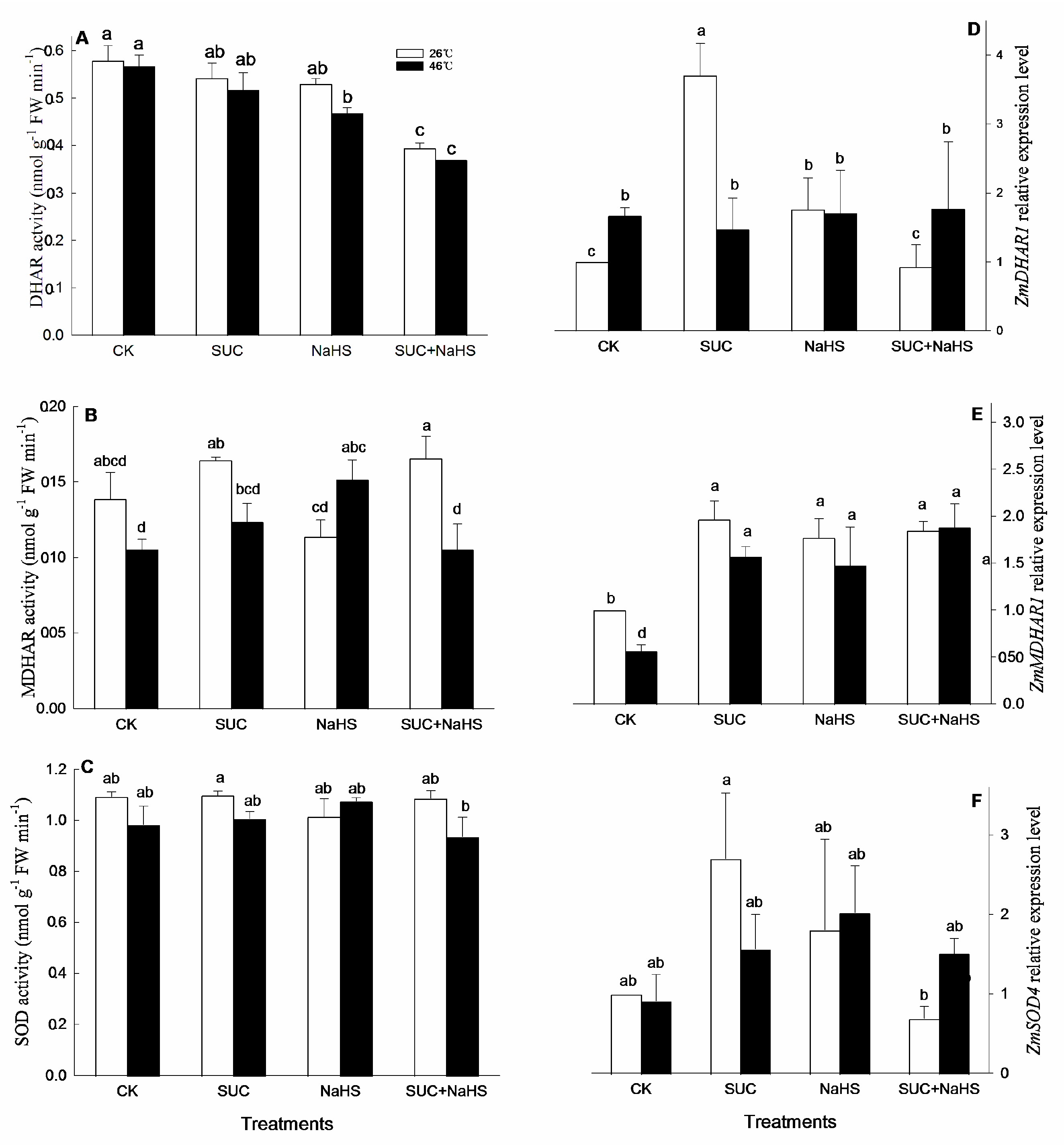
Figure 6.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on ascorbic acid (AsA, A), carotenoid (B), flavone (C), and total phenol (D) contents in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 6.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on ascorbic acid (AsA, A), carotenoid (B), flavone (C), and total phenol (D) contents in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 4), the same and different letters on the bars indicate no significant and significant difference, respectively.
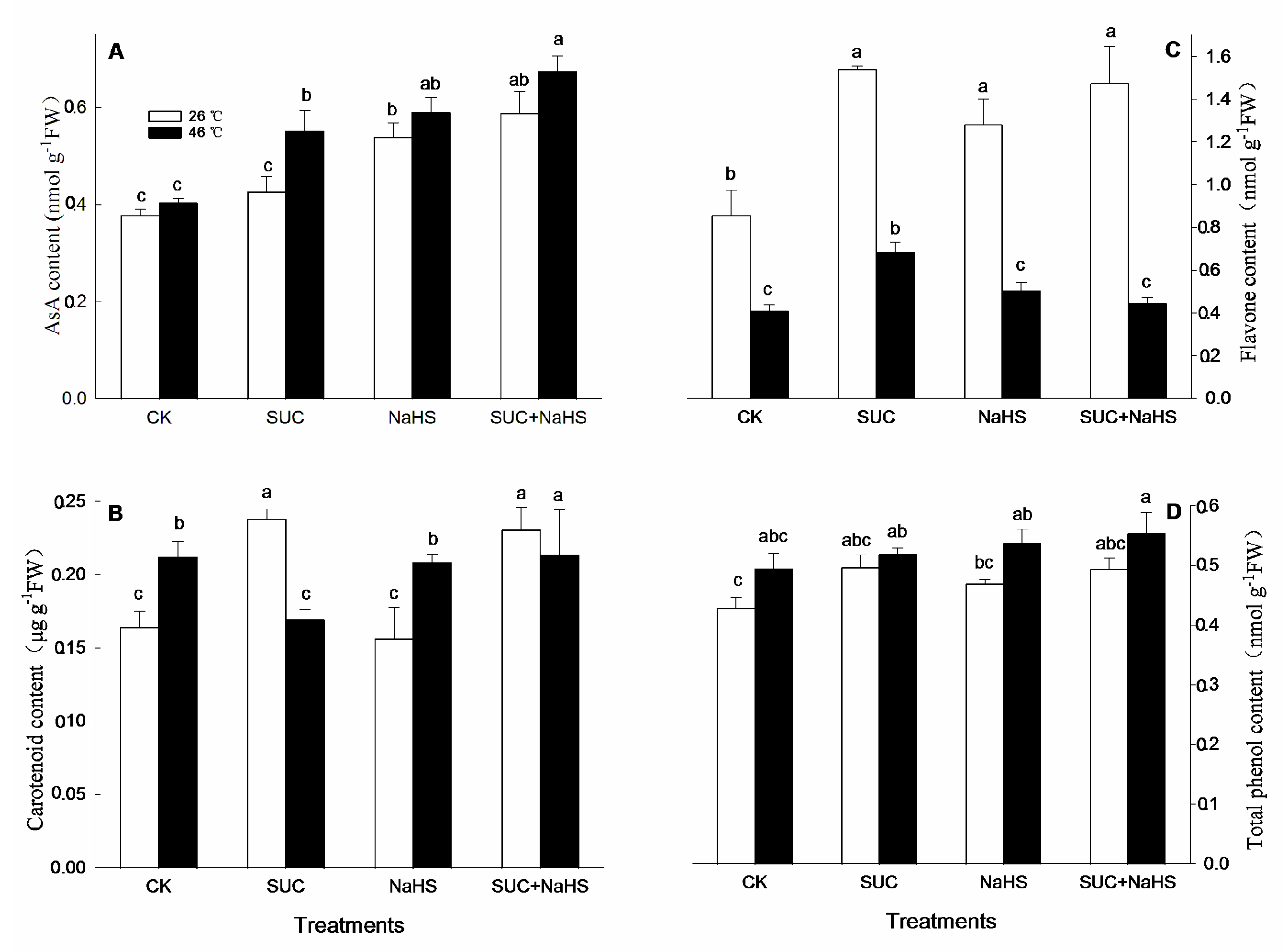
Figure 7.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on the production of superoxide radical (O2.-, A) and hydrogen peroxide level (B) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 6), the same and different letters on the bars indicate no significant and significant difference, respectively.
Figure 7.
Effect of pretreatment with sucrose (SUC) and NaHS alone or in combination on the production of superoxide radical (O2.-, A) and hydrogen peroxide level (B) in maize seedlings under non-heat and heat stress conditions. Significance between treatments was evaluated as Duncan multiple-range test, and the data in figures denote means ± standard error (SE, n = 6), the same and different letters on the bars indicate no significant and significant difference, respectively.
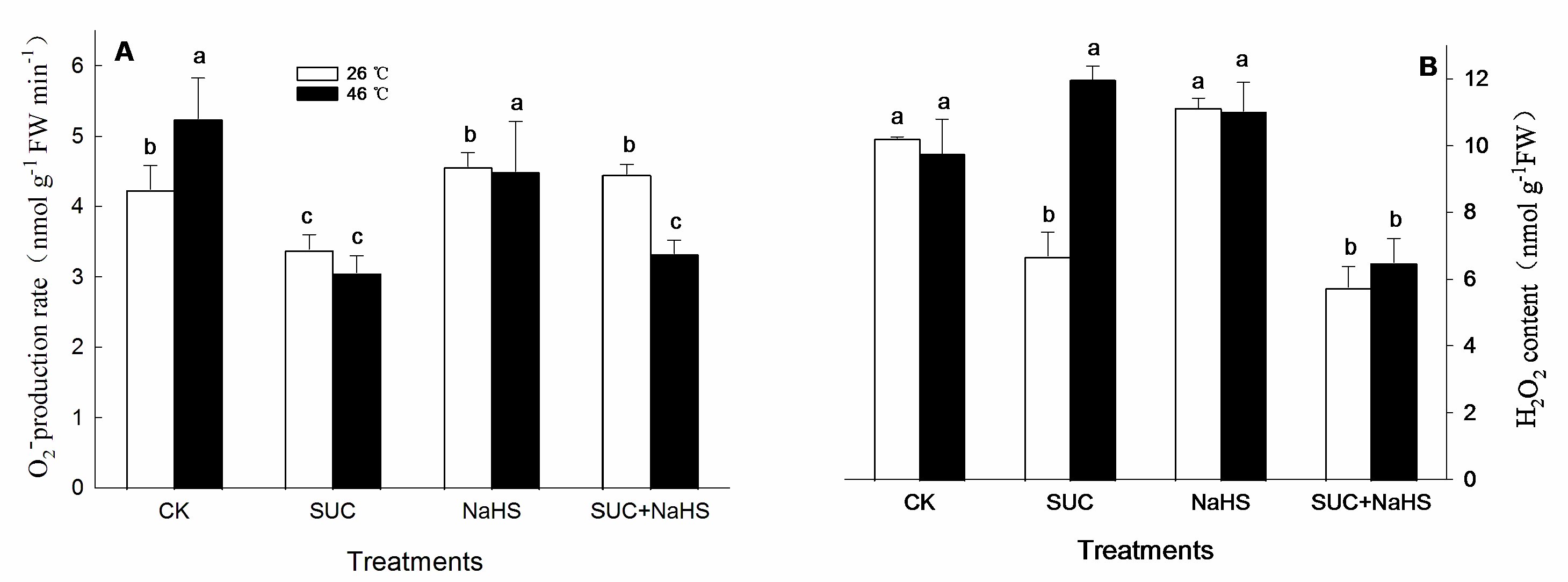
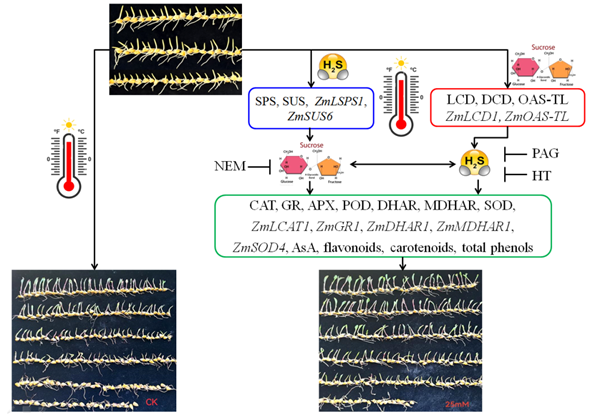
Figure 8.
The underlying mechanism of hydrogen sulfide (H2S) and sucrose interaction-upraised maize thermotolerance. H2S signaling could be triggered by exogenous sucrose via activating L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), O-acetyl-serine (thiol) lyase (OAS-TL) activities and corresponding gene expression of ZmLCD1 and ZmOAS-TL. Also, sucrose signaling could be modulated by H2S via regulating sucrose-P synthase (SPS) and sucrose sucrose synthase (SUS) activities and corresponding gene expression of ZmSPS1 and ZmSUS6. Therefore, the interaction of H2S and sucrose signaling upraised maize thermotolerance by enhancing antioxidant enzymes, non-enzymatic antioxidants, and secondary metabolites.
Figure 8.
The underlying mechanism of hydrogen sulfide (H2S) and sucrose interaction-upraised maize thermotolerance. H2S signaling could be triggered by exogenous sucrose via activating L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), O-acetyl-serine (thiol) lyase (OAS-TL) activities and corresponding gene expression of ZmLCD1 and ZmOAS-TL. Also, sucrose signaling could be modulated by H2S via regulating sucrose-P synthase (SPS) and sucrose sucrose synthase (SUS) activities and corresponding gene expression of ZmSPS1 and ZmSUS6. Therefore, the interaction of H2S and sucrose signaling upraised maize thermotolerance by enhancing antioxidant enzymes, non-enzymatic antioxidants, and secondary metabolites.
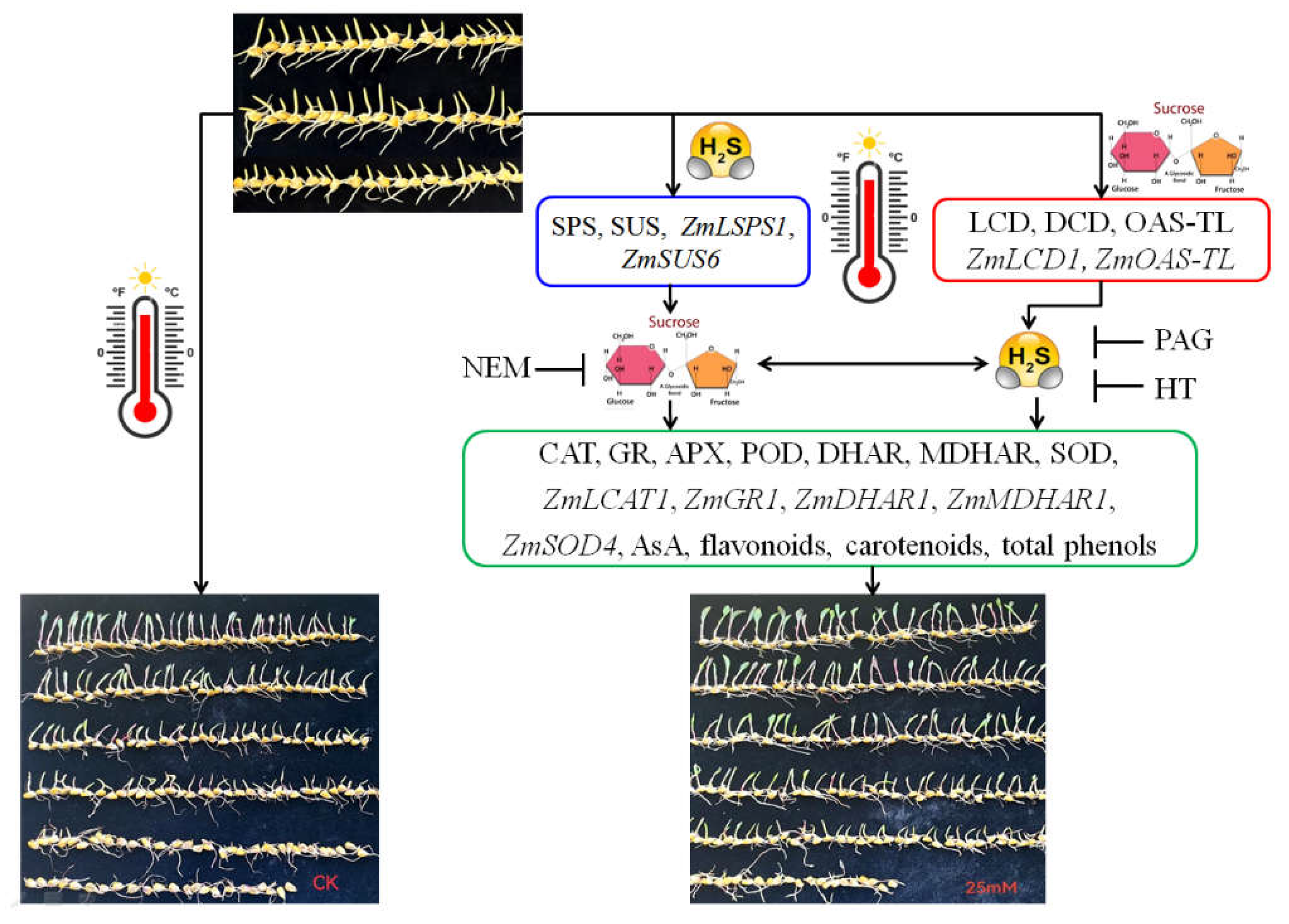

Table 1.
Pearson correlation analysis between H2S and its metabolic enzymes and SUC and its metabolic enzymes among L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), O-acetyl-serine (thiol) lyase (OAS-TL), sucrose (SUC), sucrose-P synthase (SPS), and sucrose synthase (SUS). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
Table 1.
Pearson correlation analysis between H2S and its metabolic enzymes and SUC and its metabolic enzymes among L-cysteine desulfhydrase (LCD), D-cysteine desulfhydrase (DCD), O-acetyl-serine (thiol) lyase (OAS-TL), sucrose (SUC), sucrose-P synthase (SPS), and sucrose synthase (SUS). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
| r | H2S | LCD | DCD | OAS-TL |
|---|---|---|---|---|
| SUC | 0.712* | 0.804** | 0.126 | 0.421* |
| SPS | 0.520 | 0.623* | 0.201 | 0.512* |
| SUS | 0.651* | 0.530* | 0.321 | 0.450* |
Table 2.
Pearson correlation analysis among survival rate, catalase (CAT), glutathione reductase (GR), ascorbate peroxidase (APX), peroxidase (POD), dehydroascorbate reductase (DHAR), monodehydroascorbate reductase (MDHAR), and superoxide dismutase (SOD). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
Table 2.
Pearson correlation analysis among survival rate, catalase (CAT), glutathione reductase (GR), ascorbate peroxidase (APX), peroxidase (POD), dehydroascorbate reductase (DHAR), monodehydroascorbate reductase (MDHAR), and superoxide dismutase (SOD). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
| r | Survival rate | CAT | GR | APX | POD | DHAR | MDHAR | SOD |
|---|---|---|---|---|---|---|---|---|
| Survival rate | 1 | |||||||
| CAT | 0.731** | 1 | ||||||
| GR | 0.654** | 0.320* | 1 | |||||
| APX | 0.321 | 0.412 | 0.437* | 1 | ||||
| POD | 0.452 | 0.214 | 0.213 | 0.234 | 1 | |||
| DHAR | 0.641** | 0.426* | 0.512* | 0.423* | 0.254 | 1 | ||
| MDHAR | 0.536* | 0.402** | 0.342* | 0.312* | 0. 315 | 0.452* | 1 | |
| SOD | 0.132 | 0.204 | 0.206 | 0.217 | 0.218 | 0.256 | 0.235 | 1 |
Table 3.
Pearson correlation analysis among survival rate, ascorbic acid (AsA), flavone, carotenoid, total phenol, hydrogen peroxide (H2O2), and superoxide radical (O2.-). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
Table 3.
Pearson correlation analysis among survival rate, ascorbic acid (AsA), flavone, carotenoid, total phenol, hydrogen peroxide (H2O2), and superoxide radical (O2.-). R (numbers in table) denotes correlation coefficient, whereas asterisk (*, p < 0.05) and double asterisks (**, p < 0.05) indicate significant and very significant difference.
| r | Survival rate | AsA | Flavone | Carotenoid | Total phenol | H2O2 | O2·ˉ |
|---|---|---|---|---|---|---|---|
| Survival rate | 1 | ||||||
| AsA | 0.601* | 1 | |||||
| Flavone | 0.352* | 0.321 | 1 | ||||
| Carotenoid | 0.314* | 0.432* | 0.410* | 1 | |||
| Total phenol | 0.215 | 0.284 | 0.256* | 0.413* | 1 | ||
| H2O2 | 0.412* | 0.402* | 0.426* | -0.246* | 0.622* | 1 | |
| O2·ˉ | -0.523* | -0.346 | -0.532* | -0.420* | -0.516* | -0.421* | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



