Submitted:
18 May 2024
Posted:
20 May 2024
You are already at the latest version
Abstract
Due to the host immunological disease brought on by infection is sepsis, multiple organ dysfunction syndrome (MODS), which has a high morbidity and mortality rate, could be the result. With advances in pathogen detection technologies, breakthroughs in anti-infection drugs, and the creation of organ function support, the clinical diagnosis and treatment of sepsis have advanced significantly. Pathogen-induced abnormal immunological responses, which can range from excessive inflammation to immunosuppression, are recognized to play a significant role in the high death rate. Unfortunately, no medications that are specifically intended to treat sepsis have been licensed. In order to offer a theoretical foundation for the management of sepsis, we will now discuss current advancements in research on immunological responses in sepsis. The prognosis of sepsis patients may be improved by developing and optimizing a dynamic immune system treatment plan based on anti-infection therapy, fluid replacement, support for organ function, and timely application of immunomodulatory therapies
Keywords:
diagnosis
; treatment
; flavonoids
; sepsis
; inflammatory
1. Introduction
Sepsis is a dangerous and potentially fatal medical emergency that linked to dysregulated host immunological reactions to infection (Singer et al., 2016). It develops when your immune system overreacts to an infection. It produces widespread inflammation throughout your body, which can result in tissue damage, organ failure, and even death ( Chen & Wei, 2021). Sepsis stands as a critical medical emergency characterized by dysregulated host immunological reactions to infection (Singer et al., 2016). Epidemiological data illuminates the extensive impact and seriousness of sepsis as a global health challenge.
The global Burden on the Sepsis affects over 30 million individuals worldwide annually, with approximately 6 million sepsis-related deaths reported globally each year (Fleischmann-Struzek & Rudd, 2023). In the United States alone, sepsis accounts for over 1.7 million hospitalizations annually, making it one of the most common reasons for hospital admission (Rhee et al., 2019).
Based on modeling studies or meta-analyses of prospective studies, global annual sepsis incidence was found to be 276–678/100,000 persons. Case fatality ranged from 22.5 to 26.7%(Fleischmann-Struzek & Rudd, 2023). The Mortality Rates recorded on Sepsis remains a leading cause of mortality in hospitals worldwide, with mortality rates varying widely but ranging from 15% to 55% in critically ill patients admitted to intensive care units (Singer et al., 2016). It significantly contributes to multiple organ dysfunction syndromes (MODS) in acute care settings, leading to substantial morbidity and mortality among affected individuals (Robba et al., 2020).
This call for the Public Health Concern on the burden of sepsis which extends beyond individual health outcomes to affect healthcare systems and public health infrastructure globally. It poses considerable challenges in terms of resource allocation, healthcare delivery, and patient management (Prescott & Angus, 2018). Despite advancements in critical care and sepsis management, sepsis-related mortality remains a significant concern, particularly among vulnerable populations and in resource-limited settings (Fleischmann-Struzek & Rudd, 2023
The Incidence and Prevalence on the epidemiological studies indicate a rising incidence of sepsis worldwide, attributed to factors such as an aging population, increased prevalence of chronic diseases, antimicrobial resistance, and healthcare-associated infections(Cristina et al., 2021).it imposes a substantial economic burden on healthcare systems and societies. The costs associated with sepsis-related hospitalizations, intensive care unit stays, and long-term sequelae contribute significantly to healthcare expenditures globally (Schlapbach et al., 2020).
The Geographical Disparities in sepsis incidence, outcomes, and access to care exist across different regions and populations, highlighting the need for targeted interventions and healthcare policy reforms to address these inequities (Brinkworth & Shaw, 2022). Some Studies have shown variations in sepsis incidence and outcomes based on geographical location, socioeconomic status, and access to healthcare services, emphasizing the need for tailored interventions and resource allocation strategies (Seymour et al., 2019; Fleischmann et al., 2016). These epidemiological insights underscore the urgent need for enhanced awareness, early recognition, and effective management strategies to mitigate the burden of sepsis on individuals, healthcare systems, and society as a whole.
According to the information provided in, the late-onset anti-inflammatory response establishes a link between sepsis and an immunosuppressive condition that may last for a long time (weeks or months) after the onset of SIRS (Figure 1) For patients with severe sepsis, maintaining a balance between the pro- and anti-inflammatory responses is essential (Sheth et al., 2019). Proinflammatory cytokines such IL-1, TNF- α and IL-6 are thought to be released in response to SIRS (Sheth et al., 2019). On the contrary, the pharmacological actions of anti-inflammatory mediators such as IL-1ra, IL10 and sTNFR-1 may be responsible for CARS (Sheth et al., 2019). The worst prognostic outcomes in septic patients are correlated with higher levels of IL-1β, IL-6, IL-8 and TNF- α (Sheth et al., 2019)
The severity of the illness is consequently predicted by the anti-inflammatory chemicals that are produced in larger quantities during sepsis. In addition, it represents the overall clinical and survival results. As a result, the imbalance between pro-inflammatory and anti-inflammatory mediators in the septic state reveals how severe the body's defences against the infection were (Gabarin et al., 2021). In addition, it causes excessive cytokine production and stimulation, which can have negative consequences in individuals with serious and life-threatening illnesses.
Long before flavonoids were identified as the active ingredients, these natural products were well-known for their positive health effects (Panche et al., 2016). Flavonoids come in more than 4000 different types and many of them are what give flowers, fruits, and leaves their appealing hues (Perez-Vizcaino & Fraga, 2018). Red wine's flavonoids are at least partially to blame for this result (Antoce & Stockley, 2019). Furthermore, epidemiological research points to dietary flavonoids' preventive effects of dietary flavonoids against coronary heart disease (Micek et al., 2021). Up until about 50 years ago, nothing was known about how flavonoids functioned (Kumar & Pandey, 2013). When it was discovered that this substance was a flavonoid (rutin), a flurry of research was conducted to try and separate the different flavonoids and understand how they work. Early advances in floral genetics, according to Jorgensen (Jorgensen, 1995), were mainly the result of mutation techniques that have an effect on flower colours produced from flavonoids (Morita & Hoshino, 2018). It has been demonstrated that the subclass of plant polyphenols known as flavonoids has numerous physiological advantages that are health-promoting (Rupasinghe, 2020). It is clear that study advances our understanding of dietary flavonoids' beneficial physiological effects and their therapeutic potential. New information is provided regarding the potential use of flavonoid derivatives as potent therapeutics to treat specific cancers, in addition to their function as biologically active plant food molecules (Slika et al., 2022). Additionally, flavonoids show promise in the treatment of diseases linked to inflammation and obesity as well as the prevention and treatment of infectious diseases like COVID-19 (Berretta et al., 2020).
In clinical contexts, the term "nutrition evaluation" is frequently used and an human body serves as the evaluation object. There is a wide range of research on evaluating the nutritional qualities of foods, as well as on how the idea is understood and defined across disciplines. For example, food scientists typically assess nutritional value based on the type and amount of nutrients it contains (De Souza et al., 2017; Hong et al., 2020; Nowak et al., 2016; Pereira et al., 2019; Popovi-Djordjevi et al., 2022; Shannon & Abu-Ghannam, 2019; Wang et al., 2022). Therefore, addressing the optimal dietary requirements of various groups while also taking into account the nutritional characteristics of foodstuffs will be a key area of nutrition development in the future. The precise food matrix in which a vitamin appears determines its true bioavailability, although nutrition science still pays little attention to this. It is beginning to become clear how flavonoids affect the gut microbiota and how their microbial metabolites affect optimal health. Flavonoid molecules' intricate physiological modulations are caused by the variety of their structures. However, some flavonoids are poorly absorbed, and the use of structural changes and nanotechnology uses, such as encapsulation, could increase their bioavailability ( Zhao et al., 2019).
Whether or not ultra-processed foods (UPFs) play a significant role in the nutritional quality of modern diets is still up for debate (Martnez Steele et al., 2017). The primary focus of nutrition science is the impact of nutrients on human health. It breaks down complex dietary requirements into achievable suggestions in order to prevent disease. According to conventional nutrition, the type and quantity of nutrients in a food represent its nutritional value. Dietary components are correlated with one another and consumed in combination (Neelakantan et al., 2018). Today, nutrition provides us with more than just nutrients; it also offers the opportunity to combat illness and maintain good health. Consumers have a difficult time telling what food is actually healthy and nourishing (Ditlevsen et al., 2019). To assess the nutritional worth of meals, different specialists have adopted various points and methodologies. Following is the structure of flavonoids and its classes.
1.1. The Study Inclusion/Exclusion Criteria
The literature review included randomised clinical trials, randomized quasi-trials in both people and animals, as well as in vitro and in vivo research. Research, such as unpublished theses, review newspapers, editorial documents, and memos, in which the data provided is insufficient to draw reliable conclusions, was excluded as shown in Figure 1.
2. Treatment of Sepsis Flavonoids
The potential use of various flavonoid classes in the treatment of sepsis is discussed in this section. The classification that is frequently used based on the kind and arrangement of substituents, flavones, flavonols, and flavanones, was investigated (DGI & Role, 2019; Kumar & Pandey, 2013).
2.1. Flavanoids
2.1.1. Baicalin
Baicalin (shown in Figure 2) is the primary flavonoid glucoside of Scutellaria baicalensis Georgi, a member of the Lamiaceae family, which exhibits antiviral, bacteriostatic, anticancer, and antioxidant properties (Lin et al., 2007; Shi et al., 2016). It is known for a Chinese herbal remedy, containing an active component (Deng et al., 2023). It functions as a non-steroidal anti-inflammatory drug, an inhibitor of the enzyme prolyl oligopeptidase, a prodrug, a plant metabolite, a ferroptosis inhibitor, a neuroprotective agent, an antineoplastic agent, a cardioprotective agent, an antiatherosclerotic agent, an antioxidant, an inhibitor of the enzyme RNA-directed RNA polymerase, an antibacterial agent, and an inhibitor of the enzyme (Dinda et al., 2017).
The main active component of baicalin (5,6,7-trihydroxyflavone), a common plant in traditional Chinese medicine used to cure fever, is produced from the dried root of scutellaria (Kumagai et al., 2007). Small-molecule monomer baicalin has antibacterial, anti-oxidant, anti-apoptotic, and anti-inflammatory effects (Bao et al., 2022). Baicalin reduced inflammation and preserved many organ functions. Baicalin suppressed the production of pro-inflammatory cytokines, nitric oxide (NO), nuclear factor-B (NF-B), caspase-3 activity, and reversed organ harm brought on by endotoxic shock in vivo while decreasing LPS-stimulated macrophage activation in vitro (Shahcheraghi et al., 2023). Additionally, baicalin can be used as an adjuvant therapy for both ex vivo methicillin-resistant Staphylococcus aureus (MRSA) and animal meningitis caused by Escherichia coli (E. coli) (Divyakolu et al., 2019; dos Santos Ramos et al., 2020; Rani et al., 2021).
Baicalin is viewed as the therapeutic chemical marker for S. baicalensis root quality control (Li et al., 2021). To create the TCM formula, many herbs are typically blended to enhance the medicinal effects. The pharmacokinetics may change due to interactions between the herbs in the prescription. S. baicalensis is frequently recommended with herbs such as Bupleuri radix and Coptidis rhizoma as a key ingredient in TCM formulae (Yin et al., 2021).
Many studies concentrating on baicalin's pharmacological properties have been conducted as awareness of its antiviral effects has grown ( Song et al., 2020). We have outlined the mechanism of action of baicalin as it relates to several cytokines and cell pathways ( Chen et al., 2020; K. Li et al., 2021). More discussion is also conducted on the pharmacological importance of baicalin for the creation of a broad-spectrum antiviral drug. Furthermore, pharmacologically, baicalin has been shown to protect against Sepsis. Induce liver damage and increase survival in mice suffering from polymicrobial sepsis (Achuthan & Werschler, 2022; Vo et al., 2019; von Knethen et al., 2020).
2.1.2. Scutellarin
A flavone, or phenolic chemical compound, is scutellarin. Scutellaria barbata and S. lateriflora, two plants used in conventional medicine, contain it. Guido Goldschmiedt needed a lot of time to figure out the structure of scutellarin; after the first article on the subject was published in 1901, it wasn't until 1910 that he was able to gather enough information to begin more extensive research. Figure 3 shows its chemical structure. However, studies have demonstrated that scutellarin can provide neuroprotection by suppressing microglial activation induced by increased serum (TNF–α, IL–β, and IL-6), lactate dehydrogenase activities, and tissue glutathione levels. Scutellarin has been shown to cause ovarian and breast tumour cells to undergo apoptosis in vitro. Furthermore, scutellarin exhibits protective qualities for oestrogen-affected nerve cells (Zhu et al., 2007). It has been demonstrated by (Wang et al., 2014), that scutellarin may be used to cure diabetic retinopathy, hence preventing diabetic blindness.
Scutellarin has many pharmacological benefits, including anti-inflammatory (Chledzik et al., 2018; Niu et al., 2015; Peng et al., 2020; Tan et al., 2007) and antioxidant activity, anti-diabetic, anti-ischemic, anti-cancer impact, as demonstrated by systematic investigations in modern medicine (Peng et al., 2020; Wang et al., 2020). As a result, scutellarin's multifaceted effectiveness leads to the conclusion that it may have clinical uses for the treatment of a variety of illnesses, including tissues I/R injury. It is still unknown how scutellarin affects hepatic I/R damage. As a result, in the current investigation, we looked at how scutellarin affected in vitro hepatic hypoxia/reoxygenation (H/R) injury (Toklu et al., 2008).
3.1.3. Silymarin
Silymarin is a polyphenol that belongs to the flavone group. It is a natural herbal preparation derived from fruits and milky seeds. The milk thistle plant Silybum marianum L. Gaertn, which is widely cultivated in Europe and Asia, including India, produces silymarin, which is made up of many flavonoid-like substances. Kenguil seeds are small, hard fruits from which silymarin is extracted. The chemical, which belongs to the class of drugs known as flavonolignans. It is believed that silymarin, which is composed of the flavonolignan isomers silybin (70-80%), isosilybin (0.5%), silydianin (10%) and silychristin (20%) is what gives the extract its advantageous liver-protective qualities (Brent & Shao-Nong, 2013; Lee & Liu, 2003; Scott, 1998, p. 1). Clinical and pharmacotoxicological research on silybin, the main ingredient, is extensive (Fan et al., 2018; Q. Liang et al., 2015). The chemical structure of silymarin is shown in Figure 4. A chromone fragment in its chemical structure, which enables donor-acceptor interactions with base, gives it certain acidic characteristics. The molecule has a strong antioxidant activity because to the presence of polyphenol hydroxyls and its propensity to form complexes with other metal ions and transition metal ions at the 3,4, or 4,5-positions. Several studies have demonstrated that both people and animals can tolerate its active component at very high concentrations (Tvrd et al., 2021).
In fact, the oral 50% lethal dose for rats is 10,000 mg/kg, while the greatest tolerable dose for dogs is 300 mg/kg (Abenavoli et al., 2010). The exceptionally low water solubility of silybin (430 mg/L), which is symptomatic of its lipophilic nature, as evidenced by a logarithmic P value of 1.41 (Parveen et al., 2011), where P is the drug's partitioning coefficient, places certain restrictions on its therapeutic efficacy. Silymarin is classified as a compound in class II of the biopharmaceutical categorisation system (BCS), which includes chemicals that are totally or just very slightly soluble in water. Due to this, the drug has a low bioavailability in oral formulations and has poor GIT absorption (20–50%) ( Wu et al., 2007).Chemical modifications that made silybin more water soluble often reduced its antioxidant (antiradical) (Gažák et al., 2004). Due to its antioxidant activity and ability to stabilise membranes, silymarin and its main component, silybin, have hepatoprotective qualities that prevent or slow the process of lipid peroxidation (Kurkin et al., 2009).
Because of its tendency to discourage neutrophil and ROS infiltration, silymarin treatment was observed to reverse the biochemical parameter and minimize damage to distant organs caused by sepsis. It also regulates the release of inflammatory mediators (Yang et al., 2014). As the study by (Chan et al., 2015) discovered that silymarin inhibits the formation of superoxide amino acid radicals and nitrates. Silymarin therapy also reduced the amount of serum segregated by sepsis. Silymarin was discovered in a study by (Al-Kadi et al., 2020) to protect against sepsis in laboratory mice using the cecal ligature and puncture method. In animal models induced by sepsis, silymarin antioxidants and anti-inflammatory effects prevent liver and kidney damage and reduce levels of IL-6, NO and TNF– α levels.
3.1.4. Luteolin
Luteolin is a member of the flavonoids class represented in Figure 5 and is mostly present in fruits, vegetables, and herbs. It has many therapeutic benefits, such as antioxidant, anti-inflammatory, and anticancer properties (Chuammitri et al., 2017). By enhancing contractile function and lowering apoptosis, it lowers ROS-activated MAPK pathways in ischemia healing in rats, works as an anti-inflammatory, and guards against excessive carbohydrate and fat consumption (Pan et al., 2022).
3.2. Flavonol
3.2.1. Quercetin
Quercetin is a flavonoid from the polyphenol flavonoid group, as shown in Figure 6. It has a wide range of pharmaceutical effects, including anti-tuberculosis (Hasan et al., 2022), anti-inflammatory (Lesjak et al., 2018), anti-proliferation, (Luo et al., 2016) and antioxidant protection (Karuppagounder et al., 2016). In RAW264.7 macrophages, it reduces LPS-induced TNF- release of TNF–α induced by LPS and the production of IL-1β, thereby reducing inflammatory reactions (Chuammitri et al., 2016; Yadav et al., 2012).
A study by ( Huang et al., 2015) indicated the efficacy of quercetin pre-treatment on sepsis by lowering lung pathology, inflammatory cytokines, and increasing IL-10 secretions in LPS-Induced Acute Lung Injury. According to a study by (Maalik et al., 2014) research on the effects of quercetin on septic-induced mice, the antioxidant may be able to correct lung damage by lowering blood levels of NO and MDA and enhancing the activity of antioxidant enzymes. A similar finding has been revealed in (Gerin et al., 2016; Huang et al., 2015). A study by (Park et al., 2018) revealed that Quercetin suppresses the expression of HMGB1, greatly reduces oxidative stress, and activates the enzymatic antioxidant system, successfully healing sepsis and lung cells. According to study by (Cui et al., 2019) Quercetin lowers the possibility of tissue edema, boosts alveolar capacity, and safeguards the lungs of Septic Mice. Quercetin is discovered to suppress LPS-stimulated macrophages' production of TNF- α and IL-1 β as well as NF-κB activation (Dai et al., 2013; Vickers, 2017). IKK and IκBα phosphorylation is inhibited by quercetin, which prevents NF-κB from activating.
A study by (Bharrhan et al., 2012) revealed that the induced animals had much lower levels of oxidative enzyme production, as did the inflammatory pathways. (NF-κB) were negatively regulated. A study conducted by (Wang et al., 2014) revealed that Quercetin significantly reduces NOS, COX-2, NF-κB p65 phosphorylation, malondialdehyde and HMGB1 levels and further suggested the lengthens lung life time.
3.2.2. Isorhamnetin
Isorhamnetin, with the chemical structure illustrated in Figure 7, has been shown in research findings to have a broad spectrum of pharmaceutical impacts on cardiovascular disease (Zhao & Liu, 2008) and a wide range of tumors ( Li et al., 2011), as well as the potential to protect cardiocerebral vessels and nerves (Gong et al., 2020)and neurodegenerative diseases (Ishola et al., 2019) such as Alzheimer's disease (Xu et al., 2021). It also exhibits antihyperuricemia and anti-pulmonary fibrosis pharmacodynamics (Adachi et al., 2019) and pulmonary fibrosis (Zheng et al., 2019). Several investigations have shown that Isomemtrin protects the heart and cerebrovascular system (Wang et al., 2014), prevents obesity (González-Arceo et al., 2022), is an antioxidant (Jiang et al., 2019), and has antitumor ( Kim et al., 2011) and anti-inflammatory properties (Gong et al., 2020). Isorhamnetin significantly decreases AST, ALT, and BUN levels in mice with E-coli-induced sepsis. It has a pharmacological actions are connected to its control of NF- κB, PI3K/ AKT, MAPK, and other signaling pathways and their downstream variables. The pharmacological activity and mechanism of isorhamnetin are currently being studied in depth. According to (Dong et al., 2015), isorhamnetin is cytotoxic for H9C2 cardiomyocytes and mouse primary hepatocytes (Liang et al., 2017) and promotes DNA damage in HepG2 cells (Grollino et al., 2017).
3.2.3. Fisetin
Figure 8 demonstrates that strawberries, kiwi, onions, tea, grapes, oranges, peaches, and wine are high in fisetin, a flavonoid (Qian et al., 2019). Fisetin has the potential to cause apoptosis, carcinogenesis, antioxidant, anti-tumorigenic, anti-inflammation, antiangiogenic, antidiabetic and cardioprotective properties(Abotaleb et al., 2018; Mehta et al., 2018; Pal et al., 2016; Sharif et al., 2020; Yang & Yang, 2021; Yarla et al., 2016). It can used in reducing the protein and mRNA expression of inducible iNOS, COX-2, and COX-2 (Proinflammatory mediators) in LPS-stimulated cells (Bak et al., 2012; Lee et al., 2016; Zhang et al., 2020). An Austrian chemist, (Herzig, 1891; Sahu et al., 2014) revealed its chemical formula for the first time. The biological activity of Fisetin has been examined in numerous laboratory experiments; like other polyphenols, it has a wide range of actions (Herzig, 1891; Lodyga-Chrusciska et al., 2018; Maher, 2015; Naeimi & Alizadeh, 2017).
A study by Lee et al.(2014) discovered that Fisetin can inhibit LPS-induced HMGB1 release and CAM expression, which reduce EPCR shedding significantly. The finding revealed as an effective in the treatment of vascular inflammatory diseases in sepsis-infected rats. It has been successfully in preventing action that successfully alleviates CLP-induced liver, kidney, and lung damage (H. Zhang et al., 2020). The finding of the study revealed that fisetin can be used to minimize lung damage in patients with sepsis. Fisetin was discovered to minimize lung damage in sepsis patients. It greatly reduces lung verdoperoxidase levels as well as cell proliferation for inflammatory cytokines. Many investigations have found that fisetin is a promising new antioxidant (Grynkiewicz & Demchuk, 2019).
3.2.4. Myricetin
Myricetin is a polyphenolic molecule with antioxidant characteristics that belongs to the flavonoid class (Ong & Khoo, 1997). Typical food sources (Khan et al., 2021) include red wine, tea, nuts, berries, vegetables (including tomatoes), fruits (including oranges), tea, and nuts (Naeimi & Alizadeh, 2017). Fisetin, luteolin, and quercetin share structural similarities with myricetin, and both of these flavonoids are said to have many of the same properties ( Hollman & Katan, 1999). Myricetin's reported daily average consumption varies depending on diet, although it has been shown to be 23 mg on average per day in the Netherlands ( Hollman & Katan, 1999). Myricetin is created from the parent component taxifolin through the intermediate (+) -dihydromyricetin and it can then be processed to create laricitrin and syringetin, both of which are flavonoids that belong to the flavonol class (Nwachukwu et al., 2019). Dihydromyricetin is a supplement that is widely available for sale. It is used to treat alcohol use disorder and has a contentious role as a partial GABAA receptor potentiator (AUD). An alternative method is to make myricetin directly from kaempferol, a different flavonol (Felice et al., 2022).
Natural sources of the flavonoid myricetin include plants of the Primulaceae, Leguminosae, Rosaceae, Vitaceae, Ericaceae and Fagaceae families, as well as fruits, vegetables, tea, and honey (Taheri et al., 2020). Its chemistry is shown in Figure 9. Myricetin controls IB/NFB through reducing Nrf2 in NRCM. It has been shown by pharmacology studies that , myricetin can served as anti-diabetic potential (Gupta et al., 2020). It blocks the expression of the proteins protein kinase (PKB), GLUT-2 and GLUT-4, insulin receptors 1 and 2 as well as insulin receptor 3 (IRS-3) (Lalitha et al., 2020). A study by Ozcan et al. (2012), Myricetin can be used in decreasing the Malondialdehyde and protein carbonyl levels in diabetic erythrocytes.The study further revealed, it can also be used to reduce the amount of urine, BUN, and protein excretion in diabetic albinos. As anti-inflammatory, myricetin prevents the sepsis-induced animal from producing nitric oxide, iNOS, TNF- α, IL-6, or IL-12. Furthermore, it reduces the degradation of IκBα,, the translocation of the NF- αB subunit p65, and DNA binding activity in RAW264.7 macrophages, which decreases NF- αB stimulation (Imran et al., 2021). Moreover, it prevents the growth of leukemia-causing inosine 5'-monophosphate dehydrogenase and liver cancer cells ( Jiang et al., 2019). Reduces PIM1 and interferes with interactions between PMI and CXCR4 to prevent the invasion of malignant prostate cancer ( Huang et al., 2013; Imran et al., 2019; Song et al., 2021; Wang et al., 2019).
3.3. Flavan
3.3.1. Epigallocatechin-3-gallate (EGCG)
Green tea extracts contain EGEG, a powerful catechin that is widely distributed. Its chemistry is shown in Figure 10. It has cancer prevention, bacterial resistance, anti-inflammatory, and immune stimulating qualities (Almatroodi et al., 2020). According to reported by Wheeler et al. (2007) , supported by Xin et al. (2016), revealed that, EGCG can improved the hemodynamics via lowering CL2P activity and NOS2 gene expression. When used in rodent models of polymicrobial sepsis, it increases survival rates. A drop in ALT, AST, MDA, and TNF-, as well as an improvement in hepatic expression of Nrf-2 and HO-1, were linked to a rise in survival rates in sepsis rats given EGCG taken as a pill or capsule ( Yang et al., 2017). Similarly, Hu et al. (2018) recommended that daily EGCG intake be 338 mg, although drinking tea is safe at 704 mg. Green tea has approximately 70.2 mg of EGCG per 100 mL. (about 165 mg per cup). EGEG, a potent and widely used catechin, is present in green tea extracts. In Figure 10, its chemistry is displayed. It has anti-inflammatory, antibacterial, anticancer, and immune-stimulating properties (Almatroodi et al., 2020). By reducing CL2P activity and NOS2 gene expression, EGCG improved hemodynamics. Improves survival rates in rodent models of polymicrobial sepsis. In sepsis rats fed EGCG, a decrease in ALT, AST, MDA, and TNF-, as well as an improvement in Nrf-2 and HO-1 liver expression, was associated with a rise in survival rate.
The most prevalent catechin in tea, EGCG, is a polyphenol that is being investigated for its potential to impact health and disease in people. A variety of dietary supplements contain EGCG. Even with a daily intake of 8 to 16 cups of green tea, orally administered EGCG has limited absorption, which could result in unpleasant side effects such as nausea or heartburn (Chow et al., 2003). Within 1.7 hours of intake, EGCG blood levels reach their peak (Lee et al., 2002). Although most unmodified EGCG is eliminated from the urine over a period of 0 to 8 hours, the half-life of plasma absorbed is 5 hours. Methylated metabolites are present in plasma at levels 8–25 times higher than those of unmetabolized EGCG and seem to have longer half-lives (Williamson & Manach, 2005). EGCG has a wide range of biological effects in laboratory experiments and is well researched in basic science (Fürst & Zündorf, 2014; Granja et al., 2017; Riegsecker et al., 2013; Wu et al., 2012). According to study by the European Food Safety Authority, in 2011, there is no evidence of a causal connection between tea catechins and the preservation of normal blood LDL cholesterol levels (Prawira-Atmaja et al., 2022). Similarly, in the 2016 assessment, human individuals that took high daily doses (107 to 856 mg/day) for four to 14 weeks saw a slight decrease in LDL cholesterol. According to a 2018 review, consuming too much EGCG can be harmful to the liver (Momose et al., 2016).
3.4. Immune Response in Sepsis
3.4.1. Natural Killer (NK)
Sepsis is a life-threatening condition marked by a dysregulated immune response to infection, resulting in widespread inflammation, tissue damage, and organ dysfunction(Nedeva, 2021). This complex syndrome poses a significant challenge to healthcare providers worldwide due to its high mortality rates and profound impact on patient outcomes(Van Der Poll et al., 2021).
To grasp the intricacies of sepsis pathophysiology and devise effective treatment strategies, it is imperative to explore the intricate interplay of various components of the immune system. Among these components, natural killer (NK) cells, T cells, B cells, and cytokines play pivotal roles in orchestrating the immune response to infection and modulating inflammation levels within the body(Gálvez et al., 2020; Y. Wu et al., 2017).
Natural killer (NK) cells, an integral component of the innate immune system, play a pivotal role in the body's defense against pathogens and tumors. NK cells are known for their ability to recognize and eliminate infected cells and tumor cells without the need for prior sensitization(Pahl & Cerwenka, 2017). This unique capacity allows them to act as the first line of defense against invading pathogens.
However, in the context of sepsis, the function of NK cells can become compromised, leading to significant immune dysregulation and impairment of host defense mechanisms(Nedeva, 2021). Studies have shown that during sepsis, there is a reduction in NK cell numbers and altered NK cell phenotype and function (Guo et al., 2018). These alterations in NK cell activity contribute to the dysregulated immune response observed in septic patients, making them more susceptible to secondary infections and worsening clinical outcomes (Cao et al., 2023).
Therefore, understanding the role of NK cells in sepsis pathophysiology is crucial for developing targeted therapies aimed at restoring NK cell function and bolstering host immunity during sepsis management.
Available treatments for targets such as NK cells in sepsis are currently under investigation and development. Some potential therapeutic strategies aim to modulate NK cell function and restore immune balance during sepsis(Guo et al., 2018; van der Poll et al., 2017)
- Cytokine Therapy: Administration of cytokines such as interleukin-15 (IL-15) and interleukin-2 (IL-2) may help enhance NK cell activity and promote their proliferation and function in sepsis (Carson et al., 1995).
- Immunomodulatory Agents: Drugs that target immune checkpoints, such as programmed cell death protein 1 (PD-1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) inhibitors, could potentially enhance NK cell-mediated immune responses during sepsis (Chen et al., 2013).
- Adoptive NK Cell Therapy: Infusion of ex vivo expanded and activated NK cells may offer a promising approach to augment NK cell function and improve immune surveillance in septic patients (Romee et al., 2016).
- Targeted Immunotherapy: Monoclonal antibodies directed against inhibitory receptors on NK cells, such as killer cell immunoglobulin-like receptors (KIRs), may unleash the cytotoxic potential of NK cells and enhance their antitumor and antiviral activities (Vey et al., 2012).
- Immunomodulatory Strategies: Adjunctive therapies that modulate the balance between pro-inflammatory and anti-inflammatory cytokines, such as interferon-gamma (IFN-γ) and tumor necrosis factor-alpha (TNF-α), could help restore immune homeostasis and improve outcomes in septic patients (Hotchkiss et al., 2013).
Further research is needed to evaluate the safety, efficacy, and optimal dosing strategies of these treatments in clinical settings.
Therapeutic interventions to enhance NK cell function in sepsis remain an area of active research. While no specific NK cell-targeted therapies are currently approved for sepsis management, strategies such as cytokine therapy (e.g., interleukin-15) and adoptive NK cell transfer show promise in preclinical studies(Alrubayyi et al., 2020; Godfrey et al., 2018).
3.4.2. T Cells
T cells, fundamental constituents of the adaptive immune system, wield significant influence over immune responses and inflammation regulation. In the context of sepsis, T cell dysfunction and exhaustion frequently manifest, precipitating immune suppression and heightened vulnerability to subsequent infections (ElTanbouly & Noelle, 2021). Gaining insights into the intricate dynamics of T cell responses during sepsis is paramount for devising precise immunomodulatory interventions aimed at reinstating immune equilibrium.
Research indicates that T cell dysfunction in sepsis often involves impaired effector functions and dysregulated cytokine production, contributing to immune paralysis and prolonged critical illness (Boomer et al., 2011; Hotchkiss et al., 2013). Additionally, sepsis-induced T cell exhaustion, characterized by the upregulation of inhibitory receptors like programmed cell death protein 1 (PD-1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4), further exacerbates immune dysregulation and compromises host defense mechanisms (Chang et al., 2014; Hotchkiss et al., 2013).
Given the multifaceted role of T cells in orchestrating immune responses, therapeutic strategies aimed at restoring T cell function hold significant promise in mitigating sepsis-associated morbidity and mortality. Targeted immunomodulatory approaches, such as checkpoint blockade therapy targeting PD-1 and CTLA-4, have shown potential for rejuvenating T cell responses and augmenting host immunity in preclinical and clinical studies (Chang et al., 2014; Hotchkiss et al., 2013; Matsuo et al., 2021).
Furthermore, emerging immunotherapeutic modalities, including adoptive T cell transfer and cytokine-based therapies, offer novel avenues for bolstering T cell-mediated immune surveillance and combating sepsis-induced immune dysfunction (Jackie Oh et al., 2016; Napolitano, 2018; Vellani et al., 2023). However, comprehensive understanding of the complex interplay between T cell subsets, cytokine signaling pathways, and immunoregulatory networks in sepsis remains pivotal for the development of efficacious and targeted therapeutic interventions(Bhan et al., 2016; van der Poll et al., 2017; Y. Zhang & Ning, 2021).
Available treatments for targeting T cells in sepsis are an area of active research and development. Some potential therapeutic strategies aim to modulate T cell function and mitigate immune dysfunction during sepsis. These treatments include:
- Immune Checkpoint Inhibitors: Drugs targeting immune checkpoint molecules such as programmed cell death protein 1 (PD-1) and cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) may help alleviate T cell exhaustion and restore their effector functions in sepsis (Hotchkiss et al., 2013).
- Cytokine Therapy: Administration of cytokines such as interleukin-7 (IL-7) and interleukin-15 (IL-15) can promote T cell proliferation, survival, and effector functions, potentially enhancing immune responses in septic patients (Zagorulya, 2023).
- Adoptive T Cell Therapy: Infusion of ex vivo expanded and activated T cells, such as cytotoxic T lymphocytes (CTLs) or T cell receptor (TCR)-engineered T cells, may augment T cell-mediated immune responses and improve host defense mechanisms during sepsis (Lynn et al., 2019).
- Targeted Immunomodulation: Therapies targeting specific T cell subsets, such as regulatory T cells (Tregs) or memory T cells, could help rebalance the immune response and mitigate excessive inflammation in sepsis(Duffy & Crown, 2019)
- Immunomodulatory Agents: Small molecules or biologics that modulate T cell signaling pathways, such as Janus kinase (JAK) inhibitors or nuclear factor-kappa B (NF-κB) inhibitors, may offer novel approaches to regulate T cell activation and function in septic patients(Angioni et al., 2021).
- Supportive Therapies: Supportive care measures, including early and appropriate antibiotic therapy, fluid resuscitation, and organ support, remain crucial in managing sepsis and preventing further T cell dysfunction and immune compromise (Shankar-Hari et al., 2016).
Immunomodulatory therapies aimed at restoring T cell function in sepsis include cytokine-based therapies, such as interleukin-7 (IL-7) administration, which has shown potential in preclinical and early clinical studies to reverse T cell depletion and improve immune function in septic patients(Francois et al., 2018; Venet et al., 2008).
3.4.3. B Cells
B cells, crucial components of the adaptive immune system, play a pivotal role in antibody production and antigen presentation. In sepsis, dysregulation of B cell function can lead to abnormal antibody production and compromised humoral immunity, exacerbating the inflammatory response and tissue damage(Romero-Ramírez et al., 2019).
Studies have highlighted the significance of B cells in sepsis pathophysiology. For instance, research by Boomer et (Boomer et al., 2011) demonstrated alterations in B cell phenotypes and functions in septic patients, suggesting impaired B cell responses during the disease course. Additionally, a study by Venet et al., 2008) revealed reduced B cell counts and dysfunctional antibody production in septic shock patients, correlating with disease severity and poor outcomes.
Furthermore, B cell dysfunction in sepsis has been associated with impaired pathogen clearance and increased susceptibility to secondary infections. Studies by Pene et al. (2008) and Xiao et al., (2011) highlighted the role of B cell depletion in exacerbating sepsis progression and impairing host defense mechanisms against microbial pathogens.
Understanding the intricate role of B cells in sepsis pathogenesis is critical for developing targeted therapeutic interventions. Strategies aimed at modulating B cell responses, such as B cell-targeted therapies and immunomodulatory agents, hold promise in mitigating immune dysregulation and improving patient outcomes in sepsis (Shankar-Hari et al., 2016; Singer et al., 2016; Venet et al., 2008). Unraveling the complexities of B cell-mediated immunity in sepsis offers insights into potential therapeutic targets and treatment strategies to attenuate the detrimental effects of immune dysregulation in this life-threatening condition.
Available treatments targeting B cells in sepsis remain an area of active research and development. While specific therapies directly targeting B cells in sepsis are limited, several strategies aim to modulate B cell responses and mitigate immune dysregulation in this condition.
- Immunomodulatory Therapies: Immunomodulatory agents, such as corticosteroids, intravenous immunoglobulins (IVIG), and monoclonal antibodies, are being investigated for their potential to modulate B cell function and attenuate the inflammatory response in sepsis. For example, IVIG therapy has shown promise in improving outcomes in septic patients by enhancing antibody-mediated immunity and mitigating immune dysregulation (Alejandria et al., 2013).
- B Cell-Targeted Therapies: Emerging therapies targeting B cells, such as anti-CD20 monoclonal antibodies (e.g., rituximab), aim to deplete B cell populations and suppress aberrant antibody production in sepsis. These targeted approaches hold potential for modulating B cell responses and restoring immune homeostasis in septic patients (Fowler et al., 2014).
- Cytokine Modulation: Therapies aimed at modulating cytokine levels and inflammatory signaling pathways may indirectly influence B cell function and immune responses in sepsis. For instance, agents targeting pro-inflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α) and interleukin-6 (IL-6), may attenuate B cell activation and dampen the systemic inflammatory response associated with sepsis(Cohen et al., 2015).
- Supportive Care: In addition to targeted therapies, supportive care measures play a crucial role in managing sepsis-associated complications and optimizing patient outcomes. Supportive interventions, including fluid resuscitation, vasopressor therapy, and mechanical ventilation, aim to stabilize hemodynamics, maintain organ perfusion, and support vital organ function during the acute phase of sepsis(Rhodes et al., 2017).
Currently, no specific B cell-targeted therapies are approved for sepsis treatment. However, strategies to restore B cell function and antibody production, such as intravenous immunoglobulin (IVIG) therapy, have been explored in septic patients with mixed results(Netea et al., 2017; Stawicki et al., 2020) .
While ongoing research continues to explore novel therapeutic approaches targeting B cells and immune dysregulation in sepsis, the current treatment strategies primarily focus on mitigating systemic inflammation, restoring immune function, and providing supportive care to improve patient survival and outcomes.
3.4.4. Cytokines
Cytokines, pivotal signaling molecules produced by diverse immune cells, orchestrate immune responses and regulate inflammation. In the context of sepsis, dysregulated cytokine production can precipitate a phenomenon termed cytokine storm, characterized by uncontrolled inflammation and tissue damage. Addressing specific cytokines implicated in sepsis pathogenesis presents a promising avenue for ameliorating the adverse effects of hyperinflammation and enhancing patient outcomes(Reddy et al., 2024).
Studies have underscored the significance of cytokine modulation in sepsis management(Schulte et al., 2013). For instance, research by Tanaka et al., (2016) highlighted the role of interleukin-6 (IL-6) blockade in attenuating cytokine storm and improving survival outcomes in septic patients. Additionally, trials investigating the use of tumor necrosis factor-alpha (TNF-α) inhibitors, such as infliximab and etanercept, have shown potential in dampening inflammatory responses and mitigating organ dysfunction in sepsis(A. K. Mehta et al., 2018; Saad et al., 2008) (Mehta et al., 2018).
Moreover, targeted therapies aimed at interleukin-1 (IL-1) blockade have demonstrated efficacy in modulating inflammatory cascades and improving clinical outcomes in septic patients. Clinical studies by Shakoory et al., (2016); Tisoncik et al., (2012) evaluated the use of IL-1 receptor antagonists, such as anakinra, in attenuating cytokine storm and reducing mortality rates among septic individuals.
Furthermore, emerging immunomodulatory agents, including toll-like receptor (TLR) antagonists and Janus kinase (JAK) inhibitors, hold promise for selectively modulating cytokine responses and restoring immune homeostasis in sepsis (Davis et al., 2020; van der Poll et al., 2017)(van der Poll et al., 2017; Riva et al., 2020). These innovative therapeutic approaches target key components of the cytokine signaling pathway, offering potential avenues for precision medicine in sepsis management.
Available treatments targeting the immune system components involved in sepsis pathophysiology aim to modulate immune responses and mitigate inflammation, thereby improving patient outcomes. Here are some treatment approaches targeting specific immune system components:
Targeted cytokine therapies represent a promising approach for managing cytokine storm in sepsis. Anti-cytokine agents, including monoclonal antibodies targeting interleukin-6 (IL-6) or tumor necrosis factor-alpha (TNF-α), have shown efficacy in dampening hyperinflammatory responses and improving clinical outcomes in septic patients(Alhazzani et al., 2020; Remy et al., 2020).
While progress has been made in understanding the immune response in sepsis and developing targeted therapies, further research is needed to optimize treatment efficacy, safety, and clinical applicability. By elucidating the complex mechanisms underlying immune dysregulation in sepsis, researchers and clinicians can pave the way for innovative treatment strategies aimed at restoring immune balance and improving patient survival.
4. Conclusions and Future Prospects
This study review into the diverse biological functions of flavonoids, aiming to illuminate their potential health benefits. Flavonoids, organic compounds abundant in fruits, vegetables, tea, and wine, have garnered significant attention since the emergence of the French paradox, which highlighted the intriguing cardiovascular benefits associated with red wine consumption despite a high saturated fat diet in Mediterranean populations. Subsequent research has unveiled additional therapeutic properties of flavonoids, prompting an exploration of their mechanisms of action, functions, and potential applications.
Evidence from numerous studies has underscored the anti-inflammatory, anticancer, antituberculosis, antidiabetic, antibacterial, and neuroprotective effects of flavonoids. Human participation in clinical trials has corroborated the efficacy of flavonoids in managing various inflammatory conditions such as metabolic syndrome, rheumatoid arthritis, urosepsis, and tuberculosis. However, it's noteworthy that the majority of studies relied on animal models, indicating a gap in clinical research. While animal models offer insights, they may not fully replicate human sepsis, highlighting the necessity for more robust clinical investigations.
The pharmacological profile and safety profile of flavonoids position them as promising candidates for clinical studies across diverse therapeutic areas. However, further research is imperative to elucidate their full potential and establish their efficacy and safety in human subjects. Continued exploration of flavonoids' pharmaceutical applications holds promise for the development of novel therapeutic interventions to prevent and treat chronic diseases. Thus, future endeavors should prioritize clinical investigations to harness the therapeutic potential of flavonoids for the benefit of human health.
Author Contributions
Sara Osman: Drafting an introduction, performing a literature review, and evaluating the review. Ahmad Zorin Sahlan: Concept development, study design, data collection, first draft writing, and formal analysis.
Acknowledgments
No specific grant from funding organizations in the public, private or not-for-profit sectors supported this research.
Conflicts of Interest
We certify that the publication of this material does not involve conflicts of interest.
References
- Abenavoli, L.; Capasso, R.; Milic, N.; Capasso, F. Milk thistle in liver diseases: Past, present, future. Phytotherapy Research 2010, 24, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Abotaleb, M.; Samuel, S.M.; Varghese, E.; Varghese, S.; Kubatka, P.; Liskova, A.; Büsselberg, D. Flavonoids in cancer and apoptosis. Cancers 2018, 11, 28. [Google Scholar] [CrossRef] [PubMed]
- Achuthan, M.; Werschler, N. A Novel Approach to CFTR Gene Editing through CRISPR-Cas9 Technology in Cystic Fibrosis. 2022. [Google Scholar]
- Adachi, S.; Kondo, S.; Sato, Y.; Yoshizawa, F.; Yagasaki, K. Anti-hyperuricemic effect of isorhamnetin in cultured hepatocytes and model mice: Structure–activity relationships of methylquercetins as inhibitors of uric acid production. Cytotechnology 2019, 71, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Alejandria, M.M.; Lansang, M.A. D.; Dans, L.F.; Mantaring III, J.B. Intravenous immunoglobulin for treating sepsis, severe sepsis and septic shock. Cochrane Database of Systematic Reviews, 2013; 9, https://www.cochranelibrary.com/cdsr/doi/10.1002/14651858.CD001090.pub2/abstract. [Google Scholar]
- Alhazzani, W.; Møller, M.H.; Arabi, Y.M.; Loeb, M.; Gong, M.N.; Fan, E.; Oczkowski, S.; Levy, M.M.; Derde, L.; Dzierba, A. Surviving Sepsis Campaign: Guidelines on the management of critically ill adults with Coronavirus Disease 2019 (COVID-19). Intensive Care Medicine 2020, 46, 854–887. [Google Scholar] [CrossRef] [PubMed]
- Al-Kadi, A.; Ahmed, A.-S.; El-Tahawy, N.F. G.; Khalifa, M.M. A.; El-Daly, M. Silymarin protects against sepsis-induced acute liver and kidney injury via anti-inflammatory and antioxidant mechanisms in the rat. Journal of Advanced Biomedical and Pharmaceutical Sciences 2020, 3, 190–197. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Almatroudi, A.; Khan, A.A.; Alhumaydhi, F.A.; Alsahli, M.A.; Rahmani, A.H. Potential Therapeutic Targets of Epigallocatechin Gallate (EGCG), the Most Abundant Catechin in Green Tea, and Its Role in the Therapy of Various Types of Cancer. Molecules 2020, 25, Article 14. [Google Scholar] [CrossRef]
- Alrubayyi, A.; Ogbe, A.; Moreno Cubero, E.; Peppa, D. Harnessing natural killer cell innate and adaptive traits in HIV infection. Frontiers in Cellular and Infection Microbiology 2020, 10, 395, https://www.frontiersin.org/articles/10.3389/fcimb.2020.00395/full. [Google Scholar] [CrossRef]
- Angioni, R.; Sánchez-Rodríguez, R.; Viola, A.; Molon, B. TGF-β in cancer: Metabolic driver of the tolerogenic crosstalk in the tumor microenvironment. Cancers 2021, 13, 401, https://www.mdpi.com/2072-6694/13/3/401. [Google Scholar] [CrossRef]
- Antoce, A.O.; Stockley, C. An overview of the implications of wine on human health, with special consideration of the wine-derived phenolic compounds. AgroLife Sci J 2019, 8, 21–34. [Google Scholar]
- Bak, M.-J.; Hong, S.-G.; Lee, J.-W.; Jeong, W.-S. Red ginseng marc oil inhibits iNOS and COX-2 via NFκB and p38 pathways in LPS-stimulated RAW 264.7 macrophages. Molecules 2012, 17, 13769–13786. [Google Scholar] [CrossRef]
- Bao, M.; Ma, Y.; Liang, M.; Sun, X.; Ju, X.; Yong, Y.; Liu, X. Research progress on pharmacological effects and new dosage forms of baicalin. Veterinary Medicine and Science 2022, 8, 2773–2784. [Google Scholar] [CrossRef] [PubMed]
- Berretta, A.A.; Silveira, M.A. D.; Capcha, J.M. C.; De Jong, D. Propolis and its potential against SARS-CoV-2 infection mechanisms and COVID-19 disease: Running title: Propolis against SARS-CoV-2 infection and COVID-19. Biomedicine & Pharmacotherapy 2020, 131, 110622. [Google Scholar]
- Bhan, C.; Dipankar, P.; Chakraborty, P.; Sarangi, P.P. Role of cellular events in the pathophysiology of sepsis. Inflammation Research 2016, 65, 853–868. [Google Scholar] [CrossRef]
- Bharrhan, S.; Chopra, K.; Arora, S.K.; Toor, J.S.; Rishi, P. Down-regulation of NF-κB signalling by polyphenolic compounds prevents endotoxin-induced liver injury in a rat model. Innate Immunity 2012, 18, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Boomer, J.S.; To, K.; Chang, K.C.; Takasu, O.; Osborne, D.F.; Walton, A.H.; Bricker, T.L.; Jarman, S.D.; Kreisel, D.; Krupnick, A.S. Immunosuppression in patients who die of sepsis and multiple organ failure. Jama 2011, 306, 2594–2605, https://jamanetwork.com/journals/jama/article-abstract/1104735. [Google Scholar] [CrossRef] [PubMed]
- Brent, F.J.; Shao-Nong, C. (2013). HiFSA Fingerprinting Applied to Isomers with Near-Identical NMR Spectra: The Silybin/Isosilybin Case.
- Brinkworth, J.F.; Shaw, J.G. On race, human variation, and who gets and dies of sepsis. American Journal of Biological Anthropology 2022, 178, 230–255. [Google Scholar] [CrossRef]
- Cao, M.; Wang, G.; Xie, J. Immune dysregulation in sepsis: Experiences, lessons and perspectives. Cell Death Discovery 2023, 9, 1–11. [Google Scholar] [CrossRef]
- Chang, K.; Svabek, C.; Vazquez-Guillamet, C.; Sato, B.; Rasche, D.; Wilson, S.; Robbins, P.; Ulbrandt, N.; Suzich, J.; Green, J.; Patera, A.C.; Blair, W.; Krishnan, S.; Hotchkiss, R. Targeting the programmed cell death 1: Programmed cell death ligand 1 pathway reverses T cell exhaustion in patients with sepsis. Critical Care 2014, 18, R3. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wei, H. Immune intervention in sepsis. Frontiers in Pharmacology 2021, 12, 718089. [Google Scholar] [CrossRef]
- Chen, K.-H.; Wang, S.-F.; Wang, S.-Y.; Yang, Y.-P.; Wang, M.-L.; Chiou, S.-H.; Chang, Y.-L. Pharmacological development of the potential adjuvant therapeutic agents against coronavirus disease 2019. Journal of the Chinese Medical Association 2020, 83, 817. [Google Scholar] [CrossRef]
- Chow, H.S.; Cai, Y.; Hakim, I.A.; Crowell, J.A.; Shahi, F.; Brooks, C.A.; Dorr, R.T.; Hara, Y.; Alberts, D.S. Pharmacokinetics and safety of green tea polyphenols after multiple-dose administration of epigallocatechin gallate and polyphenon E in healthy individuals. Clinical Cancer Research 2003, 9, 3312–3319. [Google Scholar] [PubMed]
- Chuammitri, P.; Srikok, S.; Saipinta, D.; Boonyayatra, S. The effects of quercetin on microRNA and inflammatory gene expression in lipopolysaccharide-stimulated bovine neutrophils. Veterinary World 2017, 10, 403. [Google Scholar] [CrossRef] [PubMed]
- CİHAN, B.; YARAT, A.; AKBAY, T.T.; ŞENER, G. Investigation of the effect of silymarin on the liver in experimental sepsis. Marmara Pharmaceutical Journal 2015, 19, 52–59. [Google Scholar] [CrossRef]
- Cohen, J.; Vincent, J.-L.; Adhikari, N.K.; Machado, F.R.; Angus, D.C.; Calandra, T.; Jaton, K.; Giulieri, S.; Delaloye, J.; Opal, S. Sepsis: A roadmap for future research. The Lancet Infectious Diseases 2015, 15, 581–614, https://www.thelancet.com/journals/laninf/article/PIIS1473-3099(15)70112-X/fulltext. [Google Scholar] [CrossRef]
- Cristina, M.L.; Spagnolo, A.M.; Giribone, L.; Demartini, A.; Sartini, M. Epidemiology and prevention of healthcare-associated infections in geriatric patients: A narrative review. International Journal of Environmental Research and Public Health 2021, 18, 5333. [Google Scholar] [CrossRef] [PubMed]
- Cui, W.; Hu, G.; Peng, J.; Mu, L.; Liu, J.; Qiao, L. Quercetin exerted protective effects in a rat model of sepsis via inhibition of reactive oxygen species (ROS) and downregulation of high mobility group box 1 (HMGB1) protein expression. Medical Science Monitor: International Medical Journal of Experimental and Clinical Research 2019, 25, 5795. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Ding, Y.; Zhang, Z.; Cai, X.; Li, Y. Quercetin and quercitrin protect against cytokine-induced injuries in RINm5F β-cells via the mitochondrial pathway and NF-κB signaling. International Journal of Molecular Medicine 2013, 31, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.S.; Ferreira, D.; Paige, E.; Gedye, C.; Boyle, M. Infectious Complications of Biological and Small Molecule Targeted Immunomodulatory Therapies. Clinical Microbiology Reviews 2020, 33, e00035–19. [Google Scholar] [CrossRef] [PubMed]
- De Souza, R.G. M.; Schincaglia, R.M.; Pimentel, G.D.; Mota, J.F. Nuts and human health outcomes: A systematic review. Nutrients 2017, 9, 1311. [Google Scholar] [CrossRef]
- DGI, G.R. B. R. C.; Role, J.P. K. Z. P. Of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef]
- Dinda, B.; Dinda, S.; DasSharma, S.; Banik, R.; Chakraborty, A.; Dinda, M. Therapeutic potentials of baicalin and its aglycone, baicalein against inflammatory disorders. European Journal of Medicinal Chemistry 2017, 131, 68–80. [Google Scholar] [CrossRef]
- Ditlevsen, K.; Sandøe, P.; Lassen, J. Healthy food is nutritious, but organic food is healthy because it is pure: The negotiation of healthy food choices by Danish consumers of organic food. Food Quality and Preference 2019, 71, 46–53. [Google Scholar] [CrossRef]
- Divyakolu, S.; Chikkala, R.; Ratnakar, K.S.; Sritharan, V. Hemolysins of Staphylococcus aureus—An update on their biology, role in pathogenesis and as targets for anti-virulence therapy. Advances in Infectious Diseases 2019, 9, 80–104. [Google Scholar] [CrossRef]
- Dong, X.; Sun, G.; Luo, Y. Protective effect of isorhamnetin on oxidative stress induced by H2O2 in H9C2 cells. Chin. Pharmacol. Bull 2015, 3, 853–860. [Google Scholar]
- dos Santos Ramos, M.A.; Dos Santos, K.C.; da Silva, P.B.; de Toledo, L.G.; Marena, G.D.; Rodero, C.F.; de Camargo, B.A. F.; Fortunato, G.C.; Bauab, T.M.; Chorilli, M. Nanotechnological strategies for systemic microbial infections treatment: A review. International Journal of Pharmaceutics 2020, 589, 119780. [Google Scholar] [CrossRef]
- Duffy, M.J.; Crown, J. Biomarkers for predicting response to immunotherapy with immune checkpoint inhibitors in cancer patients. Clinical Chemistry 2019, 65, 1228–1238, https://academic.oup.com/clinchem/article-abstract/65/10/1228/5608308. [Google Scholar] [CrossRef] [PubMed]
- ElTanbouly, M.A.; Noelle, R.J. Rethinking peripheral T cell tolerance: Checkpoints across a T cell’s journey. Nature Reviews Immunology 2021, 21, 257–267, https://www.nature.com/articles/s41577-020-00454-2. [Google Scholar] [CrossRef] [PubMed]
- Fan, D.; Guo, Q.; Shen, J.; Zheng, K.; Lu, C.; Zhang, G.; Lu, A.; He, X. The effect of triptolide in rheumatoid arthritis: From basic research towards clinical translation. International Journal of Molecular Sciences 2018, 19, 376. [Google Scholar] [CrossRef]
- Felice, M.R.; Maugeri, A.; De Sarro, G.; Navarra, M.; Barreca, D. Molecular Pathways Involved in the Anti-Cancer Activity of Flavonols: A Focus on Myricetin and Kaempferol. International Journal of Molecular Sciences 2022, 23, Article 8. [Google Scholar] [CrossRef]
- Fleischmann-Struzek, C.; Rudd, K. Challenges of assessing the burden of sepsis. Medizinische Klinik - Intensivmedizin Und Notfallmedizin 2023, 118, 68–74. [Google Scholar] [CrossRef]
- Fowler, R.A.; Mittmann, N.; Geerts, W.; Heels-Ansdell, D.; Gould, M.K.; Guyatt, G.; Krahn, M.; Finfer, S.; Pinto, R.; Chan, B. Cost-effectiveness of dalteparin vs unfractionated heparin for the prevention of venous thromboembolism in critically ill patients. Jama 2014, 312, 2135–2145, https://jamanetwork.com/journals/jama/article-abstract/1921813. [Google Scholar] [CrossRef] [PubMed]
- Francois, B.; Jeannet, R.; Daix, T.; Walton, A.H.; Shotwell, M.S.; Unsinger, J.; Monneret, G.; Rimmelé, T.; Blood, T.; Morre, M. Interleukin-7 restores lymphocytes in septic shock: The IRIS-7 randomized clinical trial. JCI Insight, 2018; 3, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5922293/. [Google Scholar]
- Fürst, R.; Zündorf, I. Plant-derived anti-inflammatory compounds: Hopes and disappointments regarding the translation of preclinical knowledge into clinical progress. Mediators of Inflammation 2014, 2014. [Google Scholar] [CrossRef] [PubMed]
- Gabarin, R.S.; Li, M.; Zimmel, P.A.; Marshall, J.C.; Li, Y.; Zhang, H. Intracellular and extracellular lipopolysaccharide signaling in sepsis: Avenues for novel therapeutic strategies. Journal of Innate Immunity 2021, 13, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Gálvez, N.M. S.; Bohmwald, K.; Pacheco, G.A.; Andrade, C.A.; Carreño, L.J.; Kalergis, A.M. Type I Natural Killer T Cells as Key Regulators of the Immune Response to Infectious Diseases. Clinical Microbiology Reviews 2020, 34. [Google Scholar] [CrossRef] [PubMed]
- Gažák, R.; Svobodová, A.; Psotová, J.; Sedmera, P.; Přikrylová, V.; Walterová, D.; Křen, V. Oxidised derivatives of silybin and their antiradical and antioxidant activity. Bioorganic & Medicinal Chemistry 2004, 12, 5677–5687. [Google Scholar]
- Gerin, F.; Sener, U.; Erman, H.; Yilmaz, A.; Aydin, B.; Armutcu, F.; Gurel, A. The effects of quercetin on acute lung injury and biomarkers of inflammation and oxidative stress in the rat model of sepsis. Inflammation 2016, 39, 700–705. [Google Scholar] [CrossRef]
- Godfrey, D.I.; Le Nours, J.; Andrews, D.M.; Uldrich, A.P.; Rossjohn, J. Unconventional T cell targets for cancer immunotherapy. Immunity 2018, 48, 453–473, https://www.cell.com/immunity/pdf/S1074-7613(18)30085-2.pdf. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Guan, Y.-Y.; Zhang, Z.-L.; Rahman, K.; Wang, S.-J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomedicine & Pharmacotherapy 2020, 128, 110301. [Google Scholar]
- González-Arceo, M.; Gomez-Lopez, I.; Carr-Ugarte, H.; Eseberri, I.; González, M.; Cano, M.P.; Portillo, M.P.; Gómez-Zorita, S. Anti-Obesity Effects of Isorhamnetin and Isorhamnetin Conjugates. International Journal of Molecular Sciences 2022, 24, 299. [Google Scholar] [CrossRef]
- Granja, A.; Frias, I.; Neves, A.R.; Pinheiro, M.; Reis, S. Therapeutic potential of epigallocatechin gallate nanodelivery systems. BioMed Research International 2017, 2017. [Google Scholar] [CrossRef]
- Grollino, M.G.; Raschellà, G.; Cordelli, E.; Villani, P.; Pieraccioli, M.; Paximadas, I.; Malandrino, S.; Bonassi, S.; Pacchierotti, F. Cytotoxicity, genotoxicity and gene expression changes elicited by exposure of human hepatic cells to Ginkgo biloba leaf extract. Food and Chemical Toxicology 2017, 109, 486–496. [Google Scholar] [CrossRef] [PubMed]
- Grynkiewicz, G.; Demchuk, O.M. New perspectives for fisetin. Frontiers in Chemistry 2019, 7, 697. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Patil, N.K.; Luan, L.; Bohannon, J.K.; Sherwood, E.R. The biology of natural killer cells during sepsis. Immunology 2018, 153, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.; Siddiqui, M.A.; Khan, M.M.; Ajmal, M.; Ahsan, R.; Rahaman, M.A.; Ahmad, M.A.; Arshad, M.; Khushtar, M. Current pharmacological trends on myricetin. Drug Research 2020, 70, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Hasan, M.M.; Khan, Z.; Chowdhury, M.S.; Khan, M.A.; Moni, M.A.; Rahman, M.H. In silico molecular docking and ADME/T analysis of Quercetin compound with its evaluation of broad-spectrum therapeutic potential against particular diseases. Informatics in Medicine Unlocked 2022, 29, 100894. [Google Scholar] [CrossRef]
- Herzig, J. Studies on Quercetin and its derivatives, treatise VII. Monatshefte Für Chemie 1891, 12, 177–190. [Google Scholar] [CrossRef]
- Hollman, P.C.; Katan, M.B. Health effects and bioavailability of dietary flavonols. Free Radical Research 1999, 31, 75–80. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.H.; Katan, M.B. Dietary flavonoids: Intake, health effects and bioavailability. Food and Chemical Toxicology 1999, 37, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Hong, J.; Mu, T.; Sun, H.; Richel, A.; Blecker, C. Valorization of the green waste parts from sweet potato (Impoea batatas L.): Nutritional, phytochemical composition, and bioactivity evaluation. Food Science & Nutrition 2020, 8, 4086–4097. [Google Scholar]
- Hotchkiss, R.S.; Monneret, G.; Payen, D. Immunosuppression in sepsis: A novel understanding of the disorder and a new therapeutic approach. The Lancet Infectious Diseases 2013, 13, 260–268, https://www.thelancet.com/journals/a/article/PIIS1473-3099(13)70001-X/fulltext. [Google Scholar] [CrossRef]
- Hu, J.; Webster, D.; Cao, J.; Shao, A. The safety of green tea and green tea extract consumption in adults–results of a systematic review. Regulatory Toxicology and Pharmacology 2018, 95, 412–433. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Zhong, T.; Wu, H. Experimental research quercetin protects against lipopolysaccharide-induced acute lung injury in rats through suppression of inflammation and oxidative stress. Archives of Medical Science 2015, 11, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.-W.; Tsai, S.-C.; Peng, S.-F.; Lin, M.-W.; Chiang, J.-H.; Chiu, Y.-J.; Fushiya, S.; Tseng, M.T.; Yang, J.-S. Kaempferol induces autophagy through AMPK and AKT signaling molecules and causes G2/M arrest via downregulation of CDK1/cyclin B in SK-HEP-1 human hepatic cancer cells. International Journal of Oncology 2013, 42, 2069–2077. [Google Scholar] [CrossRef] [PubMed]
- Imran, M.; Saeed, F.; Hussain, G.; Imran, A.; Mehmood, Z.; Gondal, T.A.; El-Ghorab, A.; Ahmad, I.; Pezzani, R.; Arshad, M.U. Myricetin: A comprehensive review on its biological potentials. Food Science & Nutrition 2021, 9, 5854–5868. [Google Scholar]
- Imran, M.; Salehi, B.; Sharifi-Rad, J.; Aslam Gondal, T.; Saeed, F.; Imran, A.; Shahbaz, M.; Tsouh Fokou, P.V.; Umair Arshad, M.; Khan, H. Kaempferol: A key emphasis to its anticancer potential. Molecules 2019, 24, 2277. [Google Scholar] [CrossRef] [PubMed]
- Ishola, I.O.; Osele, M.O.; Chijioke, M.C.; Adeyemi, O.O. Isorhamnetin enhanced cortico-hippocampal learning and memory capability in mice with scopolamine-induced amnesia: Role of antioxidant defense, cholinergic and BDNF signaling. Brain Research 2019, 1712, 188–196. [Google Scholar] [CrossRef] [PubMed]
- Jackie Oh, S.; Han, S.; Lee, W.; Lockhart, A.C. Emerging immunotherapy for the treatment of esophageal cancer. Expert Opinion on Investigational Drugs 2016, 25, 667–677. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Yamashita, Y.; Nakamura, A.; Croft, K.; Ashida, H. Quercetin and its metabolite isorhamnetin promote glucose uptake through different signalling pathways in myotubes. Scientific Reports 2019, 9, 2690. [Google Scholar] [CrossRef] [PubMed]
- Jiang, M.; Zhu, M.; Wang, L.; Yu, S. Anti-tumor effects and associated molecular mechanisms of myricetin. Biomedicine & Pharmacotherapy 2019, 120, 109506. [Google Scholar]
- Jorgensen, R.A. Cosuppression, flower color patterns, and metastable gene expression states. Science 1995, 268, 686–691. [Google Scholar] [CrossRef]
- Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Sreedhar, R.; Giridharan, V.V.; Watanabe, K. Molecular targets of quercetin with anti-inflammatory properties in atopic dermatitis. Drug Discovery Today 2016, 21, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Deb, P.K.; Priya, S.; Medina, K.D.; Devi, R.; Walode, S.G.; Rudrapal, M. Dietary flavonoids: Cardioprotective potential with antioxidant effects and their pharmacokinetic, toxicological and therapeutic concerns. Molecules 2021, 26, 4021. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Lee, D.-E.; Lee, K.W.; Son, J.E.; Seo, S.K.; Li, J.; Jung, S.K.; Heo, Y.-S.; Mottamal, M.; Bode, A.M. Isorhamnetin Suppresses Skin Cancer through Direct Inhibition of MEK1 and PI3-KInhibition of Skin Cancer by Isorhamnetin. Cancer Prevention Research 2011, 4, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, T.; Müller, C.I.; Desmond, J.C.; Imai, Y.; Heber, D.; Koeffler, H.P. Scutellaria baicalensis, a herbal medicine: Anti-proliferative and apoptotic activity against acute lymphocytic leukemia, lymphoma and myeloma cell lines. Leukemia Research 2007, 31, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. The Scientific World Journal 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Kurkin, V.A.; Ryzhov, V.M.; Biryukova, O.V.; Mel’nikova, N.B.; Selekhov, V.V. Interaction of milk-thistle-fruit flavanonols with Langmuir monolayers of lecithin and bilayers of liposomes. Pharmaceutical Chemistry Journal 2009, 43, 101–109. [Google Scholar] [CrossRef]
- Lalitha, N.; Sadashivaiah, B.; Ramaprasad, T.R.; Singh, S.A. Anti-hyperglycemic activity of myricetin, through inhibition of DPP-4 and enhanced GLP-1 levels, is attenuated by co-ingestion with lectin-rich protein. PloS One 2020, 15, e0231543. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.Y.-W.; Liu, Y. Molecular structure and stereochemistry of silybin a, silybin B, isosilybin a, and isosilybin B, isolated from Silybum m arianum (milk thistle). Journal of Natural Products 2003, 66, 1171–1174. [Google Scholar] [CrossRef]
- Lee, M.-J.; Maliakal, P.; Chen, L.; Meng, X.; Bondoc, F.Y.; Prabhu, S.; Lambert, G.; Mohr, S.; Yang, C.S. Pharmacokinetics of tea catechins after ingestion of green tea and (-)-epigallocatechin-3-gallate by humans: Formation of different metabolites and individual variability. Cancer Epidemiology Biomarkers & Prevention 2002, 11, 1025–1032. [Google Scholar]
- Lee, S.-H.; Kwak, C.-H.; Lee, S.-K.; Ha, S.-H.; Park, J.; Chung, T.-W.; Ha, K.-T.; Suh, S.-J.; Chang, Y.-C.; Chang, H.W. Anti-inflammatory effect of ascochlorin in LPS-stimulated RAW 264.7 macrophage cells is accompanied with the down-regulation of iNOS, COX-2 and proinflammatory cytokines through NF-κB, ERK1/2, and p38 signaling pathway. Journal of Cellular Biochemistry 2016, 117, 978–987. [Google Scholar] [CrossRef]
- Lee, W.; Ku, S.-K.; Bae, J.-S. Vascular barrier protective effects of orientin and isoorientin in LPS-induced inflammation in vitro and in vivo. Vascular Pharmacology 2014, 62, 3–14. [Google Scholar] [CrossRef]
- Lesjak, M.; Beara, I.; Simin, N.; Pintać, D.; Majkić, T.; Bekvalac, K.; Orčić, D.; Mimica-Dukić, N. Antioxidant and anti-inflammatory activities of quercetin and its derivatives. Journal of Functional Foods 2018, 40, 68–75. [Google Scholar] [CrossRef]
- Li, J.; Wang, G.; Du, S. Research progress on antitumor effect and mechanism of isorhamnetin, Shanxi Med. J 2011, 40, 1215–1217. [Google Scholar]
- Li, K.; Liang, Y.; Cheng, A.; Wang, Q.; Li, Y.; Wei, H.; Zhou, C.; Wan, X. Antiviral Properties of Baicalin: A Concise Review. Revista Brasileira De Farmacognosia 2021, 31, 408–419. [Google Scholar] [CrossRef]
- Liang, Q.; Wang, C.; Li, B.; Zhang, A. Metabolic fingerprinting to understand therapeutic effects and mechanisms of silybin on acute liver damage in rat. Pharmacognosy Magazine 2015, 11, 586. [Google Scholar] [CrossRef]
- Liang, R.; Chen, J.; Zhi, D.; Fan, Y.; Liu, W.; HE, X. Effects of isorhamnetin on human liver microsomes CYPs and rat primary hepatocytes. Drug Evaluation Research 2017, 627–632. [Google Scholar]
- Lin, H.-Y.; Shen, S.-C.; Lin, C.-W.; Yang, L.-Y.; Chen, Y.-C. Baicalein inhibition of hydrogen peroxide-induced apoptosis via ROS-dependent heme oxygenase 1 gene expression. Biochimica et Biophysica Acta (BBA)-Molecular Cell Research 2007, 1773, 1073–1086. [Google Scholar] [CrossRef]
- \Lodyga-Chruscińska, E.; Pilo, M.; Zucca, A.; Garribba, E.; Klewicka, E.; Rowińska-Żyrek, M.; Symonowicz, M.; Chrusciński, L.; Cheshchevik, V.T. Physicochemical, antioxidant, DNA cleaving properties and antimicrobial activity of fisetin-copper chelates. Journal of Inorganic Biochemistry 2018, 180, 101–118. [Google Scholar] [CrossRef]
- Luo, C.; Liu, Y.; Wang, P.; Song, C.; Wang, K.; Dai, L.; Zhang, J.; Ye, H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomedicine & Pharmacotherapy 2016, 82, 595–605. [Google Scholar]
- Lynn, R.C.; Weber, E.W.; Sotillo, E.; Gennert, D.; Xu, P.; Good, Z.; Anbunathan, H.; Lattin, J.; Jones, R.; Tieu, V. C-Jun overexpression in CAR T cells induces exhaustion resistance. Nature 2019, 576, 293–300, https://idp.nature.com/authorize/casa?redirect_uri=https://www.nature.com/articles/s41586-019-1805-z&casa_token=VJ3gEm0iZVIAAAAA:_k2zyBOOMzkCOZuP0_iNOOTsKKXO05NrSOH1SPaP3NzP9lS93Gad7qwrQ3-DyLc0ZmqzF_d7SSwdQn03Zw. [Google Scholar] [CrossRef]
- Maalik, A.; Khan, F.A.; Mumtaz, A.; Mehmood, A.; Azhar, S.; Atif, M.; Karim, S.; Altaf, Y.; Tariq, I. Pharmacological applications of quercetin and its derivatives: A short review. Tropical Journal of Pharmaceutical Research 2014, 13, 1561–1566. [Google Scholar] [CrossRef]
- Maher, P. Fisetin acts on multiple pathways to reduce the impact of age and disease on CNS function. Frontiers in Bioscience (Scholar Edition) 2015, 7, 58. [Google Scholar] [CrossRef]
- Martínez Steele, E.; Popkin, B.M.; Swinburn, B.; Monteiro, C.A. The share of ultra-processed foods and the overall nutritional quality of diets in the US: Evidence from a nationally representative cross-sectional study. Population Health Metrics 2017, 15, 1–11. [Google Scholar] [CrossRef]
- Matsuo, K.; Yoshie, O.; Nakayama, T. Multifaceted roles of chemokines and chemokine receptors in tumor immunity. Cancers 2021, 13, 6132. [Google Scholar] [CrossRef]
- Mehta, A.K.; Gracias, D.T.; Croft, M. TNF activity and T cells. Cytokine 2018, 101, 14–18, https://www.sciencedirect.com/science/article/pii/S1043466616304501. [Google Scholar] [CrossRef]
- Mehta, P.; Pawar, A.; Mahadik, K.; Bothiraja, C. Emerging novel drug delivery strategies for bioactive flavonol fisetin in biomedicine. Biomedicine & Pharmacotherapy 2018, 106, 1282–1291. [Google Scholar]
- Micek, A.; Godos, J.; Del Rio, D.; Galvano, F.; Grosso, G. Dietary flavonoids and cardiovascular disease: A comprehensive dose–response meta-analysis. Molecular Nutrition & Food Research 2021, 65, 2001019. [Google Scholar]
- Momose, Y.; Maeda-Yamamoto, M.; Nabetani, H. Systematic review of green tea epigallocatechin gallate in reducing low-density lipoprotein cholesterol levels of humans. International Journal of Food Sciences and Nutrition 2016, 67, 606–613. [Google Scholar] [CrossRef]
- Morita, Y.; Hoshino, A. Recent advances in flower color variation and patterning of Japanese morning glory and petunia. Breeding Science 2018, 68, 128–138. [Google Scholar] [CrossRef]
- Naeimi, A.F.; Alizadeh, M. Antioxidant properties of the flavonoid fisetin: An updated review of in vivo and in vitro studies. Trends in Food Science & Technology 2017, 70, 34–44. [Google Scholar]
- Napolitano, L.M. Sepsis 2018: Definitions and Guideline Changes. Surgical Infections 2018, 19, 117–125. [Google Scholar] [CrossRef]
- Nedeva, C. Inflammation and Cell Death of the Innate and Adaptive Immune System during Sepsis. Biomolecules 2021, 11, Article 7. [Google Scholar] [CrossRef]
- Neelakantan, N.; Koh, W.-P.; Yuan, J.-M.; van Dam, R.M. Diet-quality indexes are associated with a lower risk of cardiovascular, respiratory, and all-cause mortality among Chinese adults. The Journal of Nutrition 2018, 148, 1323–1332. [Google Scholar] [CrossRef]
- Netea, M.G.; Balkwill, F.; Chonchol, M.; Cominelli, F.; Donath, M.Y.; Giamarellos-Bourboulis, E.J.; Golenbock, D.; Gresnigt, M.S.; Heneka, M.T.; Hoffman, H.M. A guiding map for inflammation. Nature Immunology 2017, 18, 826–831, https://www.nature.com/articles/ni.3790. [Google Scholar] [CrossRef]
- Nowak, V.; Du, J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chemistry 2016, 193, 47–54. [Google Scholar] [CrossRef]
- Nwachukwu, J.N.; Durojaye, O.A.; Njoku, U.O.; Tanze, M.M.; Christian, N.; Orum, T.G.; Cosmas, S. Improving the Drug Bioavailability Property of Myricetin through a Structural Monosubstitution Modification Approach: An In-Silico Pharmacokinetics Study. African Journal of Biomedical Research 2019, 22, 347–352. [Google Scholar]
- Ong, K.C.; Khoo, H.-E. Biological effects of myricetin. General Pharmacology: The Vascular System 1997, 29, 121–126. [Google Scholar] [CrossRef]
- Ozcan, F.; Ozmen, A.; Akkaya, B.; Aliciguzel, Y.; Aslan, M. Beneficial effect of myricetin on renal functions in streptozotocin-induced diabetes. Clinical and Experimental Medicine 2012, 12, 265–272. [Google Scholar] [CrossRef]
- Pahl, J.; Cerwenka, A. Tricking the balance: NK cells in anti-cancer immunity. Immunobiology 2017, 222, 11–20. [Google Scholar] [CrossRef]
- Pal, H.C.; Pearlman, R.L.; Afaq, F. Fisetin and its role in chronic diseases. Anti-Inflammatory Nutraceuticals and Chronic Diseases, 2016; 213–244. [Google Scholar]
- Pan, Q.; Liu, Y.; Ma, W.; Kan, R.; Zhu, H.; Li, D. Cardioprotective Effects and Possible Mechanisms of Luteolin for Myocardial Ischemia-Reperfusion Injury: A Systematic Review and Meta-Analysis of Preclinical Evidence. Frontiers in Cardiovascular Medicine 2022, 898. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. Journal of Nutritional Science 2016, 5, e47. [Google Scholar] [CrossRef]
- Park, H.J.; Lee, S.J.; Cho, J.; Gharbi, A.; Han, H.D.; Kang, T.H.; Kim, Y.; Lee, Y.; Park, W.S.; Jung, I.D. Tamarixetin exhibits anti-inflammatory activity and prevents bacterial sepsis by increasing IL-10 production. Journal of Natural Products 2018, 81, 1435–1443. [Google Scholar] [CrossRef]
- Parveen, R.; Baboota, S.; Ali, J.; Ahuja, A.; Vasudev, S.S.; Ahmad, S. Oil based nanocarrier for improved oral delivery of silymarin: In vitro and in vivo studies. International Journal of Pharmaceutics 2011, 413, 245–253. [Google Scholar] [CrossRef]
- Pene, F.; Zuber, B.; Courtine, E.; Rousseau, C.; Ouaaz, F.; Toubiana, J.; Tazi, A.; Mira, J.-P.; Chiche, J.-D. Dendritic cells modulate lung response to Pseudomonas aeruginosa in a murine model of sepsis-induced immune dysfunction. The Journal of Immunology 2008, 181, 8513–8520. [Google Scholar] [CrossRef]
- Peng, L.; Wen, L.; Shi, Q.-F.; Gao, F.; Huang, B.; Meng, J.; Hu, C.-P.; Wang, C.-M. Scutellarin ameliorates pulmonary fibrosis through inhibiting NF-κB/NLRP3-mediated epithelial–mesenchymal transition and inflammation. Cell Death & Disease 2020, 11, 978. [Google Scholar]
- Pereira, E.; Encina-Zelada, C.; Barros, L.; Gonzales-Barron, U.; Cadavez, V.; Ferreira, I.C. Chemical and nutritional characterization of Chenopodium quinoa Willd (quinoa) grains: A good alternative to nutritious food. Food Chemistry 2019, 280, 110–114. [Google Scholar] [CrossRef]
- Perez-Vizcaino, F.; Fraga, C.G. Research trends in flavonoids and health. Archives of Biochemistry and Biophysics 2018, 646, 107–112. [Google Scholar] [CrossRef]
- Popović-Djordjević, J.B.; Kostić, A. Ž. , Rajković, M.B.; Miljković, I.; Krstić, \DJur\dja, Caruso, G.; Siavash Moghaddam, S.; Brčeski, I. Organically vs. conventionally grown vegetables: Multi-elemental analysis and nutritional evaluation. Biological Trace Element Research 2022, 200, 426–436. [Google Scholar] [CrossRef]
- Prawira-Atmaja, M.I.; Sita, K.; Setyaningtyas, S.S. W.; Maulana, H.; Setiadi, D.; Priatni, S.; Sulaswatty, A. Perspective regulatory framework on health claim of tea-mini review. Turkish Journal of Agriculture-Food Science and Technology 2022, 10, 555–561. [Google Scholar] [CrossRef]
- Prescott, H.C.; Angus, D.C. Enhancing recovery from sepsis: A review. Jama 2018, 319, 62–75, https://jamanetwork.com/journals/jama/article-abstract/2667727. [Google Scholar] [CrossRef]
- Qian, R.; Liang, M.; Fu, P. FP297 Anti-Inflammatory Flavonoid Fisetin Attenuates LPS-Induced Acute Kidney Injury via Inhibiting TLR4/NF-kappaB Pathway. Nephrology Dialysis Transplantation 2019, 34 (Suppl. 1), gfz106–FP297. [Google Scholar] [CrossRef]
- Rani, N.N. I. M.; Hussein, Z.M.; Mustapa, F.; Azhari, H.; Sekar, M.; Chen, X.Y.; Amin, M.C. I. M. Exploring the possible targeting strategies of liposomes against methicillin-resistant Staphylococcus aureus (MRSA). European Journal of Pharmaceutics and Biopharmaceutics 2021, 165, 84–105. [Google Scholar] [CrossRef]
- Reddy, H.; Javvaji, C.K.; Malali, S.; Kumar, S.; Acharya, S.; Toshniwal, S.; Malali Jr, S.; Toshniwal, S.S. Navigating the Cytokine Storm: A Comprehensive Review of Chemokines and Cytokines in Sepsis. Cureus, 2024; 16, https://www.cureus.com/articles/219659-navigating-the-cytokine-storm-a-comprehensive-review-of-chemokines-and-cytokines-in-sepsis.pdf. [Google Scholar]
- Remy, K.E.; Mazer, M.; Striker, D.A.; Ellebedy, A.H.; Walton, A.H.; Unsinger, J.; Blood, T.M.; Mudd, P.A.; Daehan, J.Y.; Mannion, D.A. Severe immunosuppression and not a cytokine storm characterizes COVID-19 infections. JCI Insight, 2020; 5, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7526441/. [Google Scholar]
- Rhee, C.; Jones, T.M.; Hamad, Y.; Pande, A.; Varon, J.; O’Brien, C.; Anderson, D.J.; Warren, D.K.; Dantes, R.B.; Epstein, L. Prevalence, underlying causes, and preventability of sepsis-associated mortality in US acute care hospitals. JAMA Network Open 2019, 2, e187571–e187571, https://jamanetwork.com/journals/jamanetworkopen/article-abstract/2724768. [Google Scholar] [CrossRef]
- Rhodes, A.; Evans, L.E.; Alhazzani, W.; Levy, M.M.; Antonelli, M.; Ferrer, R.; Kumar, A.; Sevransky, J.E.; Sprung, C.L.; Nunnally, M.E.; Rochwerg, B.; Rubenfeld, G.D.; Angus, D.C.; Annane, D.; Beale, R.J.; Bellinghan, G.J.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.; … Dellinger, R.P. Surviving Sepsis Campaign: International Guidelines for Management of Sepsis and Septic Shock: 2016. Intensive Care Medicine 2017, 43, 304–377. [Google Scholar] [CrossRef]
- Riegsecker, S.; Wiczynski, D.; Kaplan, M.J.; Ahmed, S. Potential benefits of green tea polyphenol EGCG in the prevention and treatment of vascular inflammation in rheumatoid arthritis. Life Sciences 2013, 93, 307–312. [Google Scholar] [CrossRef]
- Robba, C.; Battaglini, D.; Pelosi, P.; Rocco, P.R. M. Multiple organ dysfunction in SARS-CoV-2: MODS-CoV-2. Expert Review of Respiratory Medicine 2020, 14, 865–868. [Google Scholar] [CrossRef]
- Romero-Ramírez, S.; Navarro-Hernandez, I.C.; Cervantes-Díaz, R.; Sosa-Hernández, V.A.; Acevedo-Ochoa, E.; Kleinberg-Bild, A.; Valle-Rios, R.; Meza-Sánchez, D.E.; Hernández-Hernández, J.M.; Maravillas-Montero, J.L. Innate-like B cell subsets during immune responses: Beyond antibody production. Journal of Leukocyte Biology 2019, 105, 843–856, https://academic.oup.com/jleukbio/article-abstract/105/5/843/6935465. [Google Scholar] [CrossRef]
- Rupasinghe, H.V. Special Issue “flavonoids and their disease prevention and treatment potential”: Recent advances and future perspectives. Molecules 2020, 25, 4746. [Google Scholar] [CrossRef]
- Saad, A.A.; Symmons, D.P.; Noyce, P.R.; Ashcroft, D.M. Risks and benefits of tumor necrosis factor-alpha inhibitors in the management of psoriatic arthritis: Systematic review and metaanalysis of randomized controlled trials. The Journal of Rheumatology 2008, 35, 883–890, https://www.jrheum.org/content/35/5/883.short. [Google Scholar]
- Sahu, B.D.; Kalvala, A.K.; Koneru, M.; Mahesh Kumar, J.; Kuncha, M.; Rachamalla, S.S.; Sistla, R. Ameliorative effect of fisetin on cisplatin-induced nephrotoxicity in rats via modulation of NF-κB activation and antioxidant defence. PloS One 2014, 9, e105070. [Google Scholar] [CrossRef]
- Schlapbach, L.J.; Kissoon, N.; Alhawsawi, A.; Aljuaid, M.H.; Daniels, R.; Gorordo-Delsol, L.A.; Machado, F.; Malik, I.; Nsutebu, E.F.; Finfer, S.; Reinhart, K. World Sepsis Day: A global agenda to target a leading cause of morbidity and mortality. American Journal of Physiology-Lung Cellular and Molecular Physiology 2020, 319, L518–L522. [Google Scholar] [CrossRef]
- Schulte, W.; Bernhagen, J.; Bucala, R. Cytokines in sepsis: Potent immunoregulators and potential therapeutic targets—an updated view. Mediators of Inflammation, 2013; 2013, https://www.hindawi.com/journals/mi/2013/165974/. [Google Scholar]
- Scott Luper, N.D. A review of plants used in the treatment of liver disease: Part 1. Alternative Medicine Review 1998, 3, 410–421. [Google Scholar]
- Shahcheraghi, S.H.; Salemi, F.; Small, S.; Syed, S.; Salari, F.; Alam, W.; Cheang, W.S.; Saso, L.; Khan, H. Resveratrol regulates inflammation and improves oxidative stress via Nrf2 signaling pathway: Therapeutic and biotechnological prospects. Phytotherapy Research. 2023. [CrossRef]
- Shakoory, B.; Carcillo, J.A.; Chatham, W.W.; Amdur, R.L.; Zhao, H.; Dinarello, C.A.; Cron, R.Q.; Opal, S.M. Interleukin-1 receptor blockade is associated with reduced mortality in sepsis patients with features of the macrophage activation syndrome: Re-analysis of a prior Phase III trial. Critical Care Medicine 2016, 44, 275. [Google Scholar] [CrossRef]
- Shankar-Hari, M.; Phillips, G.S.; Levy, M.L.; Seymour, C.W.; Liu, V.X.; Deutschman, C.S.; Angus, D.C.; Rubenfeld, G.D.; Singer, M. Developing a new definition and assessing new clinical criteria for septic shock: For the Third International Consensus Definitions for Sepsis and Septic Shock (Sepsis-3). Jama 2016, 315, 775–787, https://jamanetwork.com/journals/jama/article-abstract/2492876. [Google Scholar] [CrossRef]
- Shannon, E.; Abu-Ghannam, N. Seaweeds as nutraceuticals for health and nutrition. Phycologia 2019, 58, 563–577. [Google Scholar] [CrossRef]
- Sharif, H.; Akash, M.S. H.; Rehman, K.; Irshad, K.; Imran, I. Pathophysiology of atherosclerosis: Association of risk factors and treatment strategies using plant-based bioactive compounds. Journal of Food Biochemistry 2020, 44, e13449. [Google Scholar] [CrossRef]
- Sheth, M.; Benedum, C.M.; Celi, L.A.; Mark, R.G.; Markuzon, N. The association between autoimmune disease and 30-day mortality among sepsis ICU patients: A cohort study. Critical Care 2019, 23, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Ren, K.; Lv, B.; Zhang, W.; Zhao, Y.; Tan, R.X.; Li, E. Baicalin from Scutellaria baicalensis blocks respiratory syncytial virus (RSV) infection and reduces inflammatory cell infiltration and lung injury in mice. Scientific Reports 2016, 6, 35851. [Google Scholar] [CrossRef]
- Singer, M.; Deutschman, C.S.; Seymour, C.W.; Shankar-Hari, M.; Annane, D.; Bauer, M.; Bellomo, R.; Bernard, G.R.; Chiche, J.-D.; Coopersmith, C.M. The third international consensus definitions for sepsis and septic shock (Sepsis-3). Jama 2016, 315, 801–810. [Google Scholar] [CrossRef]
- Slika, H.; Mansour, H.; Wehbe, N.; Nasser, S.A.; Iratni, R.; Nasrallah, G.; Shaito, A.; Ghaddar, T.; Kobeissy, F.; Eid, A.H. Therapeutic potential of flavonoids in cancer: ROS-mediated mechanisms. Biomedicine & Pharmacotherapy 2022, 146, 112442. [Google Scholar]
- Song, J.-W.; Long, J.-Y.; Xie, L.; Zhang, L.-L.; Xie, Q.-X.; Chen, H.-J.; Deng, M.; Li, X.-F. Applications, phytochemistry, pharmacological effects, pharmacokinetics, toxicity of Scutellaria baicalensis Georgi. and its probably potential therapeutic effects on COVID-19: A review. Chinese Medicine 2020, 15, 102. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A review of the most recent research. Biomedicine & Pharmacotherapy 2021, 134, 111017. [Google Scholar]
- Stawicki, S.P.; Jeanmonod, R.; Miller, A.C.; Paladino, L.; Gaieski, D.F.; Yaffee, A.Q.; De Wulf, A.; Grover, J.; Papadimos, T.J.; Bloem, C. The 2019–2020 novel coronavirus (severe acute respiratory syndrome coronavirus 2) pandemic: A joint american college of academic international medicine-world academic council of emergency medicine multidisciplinary COVID-19 working group consensus paper. Journal of Global Infectious Diseases 2020, 12, 47, https://www.ncbi.nlm.nih.gov/pmc/articles/PMC7384689/. [Google Scholar] [CrossRef]
- Taheri, Y.; Suleria, H.A. R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S. Myricetin bioactive effects: Moving from preclinical evidence to potential clinical applications. BMC Complementary Medicine and Therapies 2020, 20, 1–14. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. Immunotherapeutic implications of IL-6 blockade for cytokine storm. Immunotherapy 2016, 8, 959–970. [Google Scholar] [CrossRef]
- Tisoncik, J.R.; Korth, M.J.; Simmons, C.P.; Farrar, J.; Martin, T.R.; Katze, M.G. Into the Eye of the Cytokine Storm. Microbiology and Molecular Biology Reviews 2012, 76, 16–32. [Google Scholar] [CrossRef]
- Toklu, H.Z.; Akbay, T.T.; Velioglu-Ogunc, A.; Ercan, F.; Gedik, N.; Keyer-Uysal, M.; Sener, G. Silymarin, the antioxidant component of Silybum marianum, prevents sepsis-induced acute lung and brain injury. Journal of Surgical Research 2008, 145, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Tvrdỳ, V.; Pourová, J.; Jirkovskỳ, E.; Křen, V.; Valentová, K.; Mladěnka, P. Systematic review of pharmacokinetics and potential pharmacokinetic interactions of flavonolignans from silymarin. Medicinal Research Reviews 2021, 41, 2195–2246. [Google Scholar] [CrossRef]
- Van Der Poll, T.; Shankar-Hari, M.; Wiersinga, W.J. The immunology of sepsis. Immunity 2021, 54, 2450–2464. [Google Scholar] [CrossRef]
- van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nature Reviews Immunology 2017, 17, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Vellani, S.D.; Nigro, A.; Varatharajan, S.; Dworkin, L.D.; Creeden, J.F. Emerging Immunotherapeutic and Diagnostic Modalities in Carcinoid Tumors. Molecules 2023, 28, 2047, https://www.mdpi.com/1420-3049/28/5/2047. [Google Scholar] [CrossRef] [PubMed]
- Venet, F.; Chung, C.-S.; Monneret, G.; Huang, X.; Horner, B.; Garber, M.; Ayala, A. Regulatory T cell populations in sepsis and trauma. Journal of Leucocyte Biology 2008, 83, 523–535, https://academic.oup.com/jleukbio/article-abstract/83/3/523/6975520. [Google Scholar] [CrossRef] [PubMed]
- Vickers, N.J. Animal communication: When i’m calling you, will you answer too? Current Biology 2017, 27, R713–R715. [Google Scholar] [CrossRef] [PubMed]
- Vo, Q.V.; Nam, P.C.; Thong, N.M.; Trung, N.T.; Phan, C.-T. D.; Mechler, A. Antioxidant motifs in flavonoids: O–H versus C–H bond dissociation. ACS Omega 2019, 4, 8935–8942. [Google Scholar] [CrossRef] [PubMed]
- von Knethen, A.; Heinicke, U.; Weigert, A.; Zacharowski, K.; Brüne, B. Histone deacetylation inhibitors as modulators of regulatory T cells. International Journal of Molecular Sciences 2020, 21, 2356. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wang, L.; Gu, J.; Yang, H.; Liu, N.; Lin, Y.; Li, X.; Shao, C. Scutellarin Inhibits High Glucose–induced and Hypoxia-mimetic Agent–induced Angiogenic Effects in Human Retinal Endothelial Cells through Reactive Oxygen Species/Hypoxia-inducible Factor-1α/Vascular Endothelial Growth Factor Pathway. Journal of Cardiovascular Pharmacology 2014, 64, 218–227. [Google Scholar] [CrossRef]
- Wang, L.; Chen, J.; Wang, B.; Wu, D.; Li, H.; Lu, H.; Wu, H.; Chai, Y. Protective effect of quercetin on lipopolysaccharide-induced acute lung injury in mice by inhibiting inflammatory cell influx. Experimental Biology and Medicine 2014, 239, 1653–1662. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Huang, J.; Sun, J.; Liu, R.; Jiang, T.; Sun, G. Evaluating the Nutritional Properties of Food: A Scoping Review. Nutrients 2022, 14, 2352. [Google Scholar] [CrossRef]
- Wang, X.; Yang, Y.; An, Y.; Fang, G. The mechanism of anticancer action and potential clinical use of kaempferol in the treatment of breast cancer. Biomedicine & Pharmacotherapy 2019, 117, 109086. [Google Scholar]
- Wang, Y.; Fan, X.; Fan, B.; Jiang, K.; Zhang, H.; Kang, F.; Su, H.; Gu, D.; Li, S.; Lin, S. Scutellarin Reduce the Homocysteine Level and Alleviate Liver Injury in Type 2 Diabetes Model. Frontiers in Pharmacology 2020, 11. https://www.frontiersin.org/articles/10.3389/fphar.2020.538407. [Google Scholar]
- Wheeler, D.S.; Lahni, P.M.; Hake, P.W.; Denenberg, A.G.; Wong, H.R.; Snead, C.; Catravas, J.D.; Zingarelli, B. The green tea polyphenol epigallocatechin-3-gallate improves systemic hemodynamics and survival in rodent models of polymicrobial sepsis. Shock 2007, 28, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Williamson, G.; Manach, C. Bioavailability and bioefficacy of polyphenols in humans. II. Review of 93 intervention studies. The American Journal of Clinical Nutrition 2005, 81, 243S–255S. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Wang, J.; Pae, M.; Meydani, S.N. Green tea EGCG, T cells, and T cell-mediated autoimmune diseases. Molecular Aspects of Medicine 2012, 33, 107–118. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.-W.; Lin, L.-C.; Hung, S.-C.; Chi, C.-W.; Tsai, T.-H. Analysis of silibinin in rat plasma and bile for hepatobiliary excretion and oral bioavailability application. Journal of Pharmaceutical and Biomedical Analysis 2007, 45, 635–641. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Tian, Z.; Wei, H. Developmental and Functional Control of Natural Killer Cells by Cytokines. Frontiers in Immunology 2017, 8. [Google Scholar] [CrossRef]
- Xiao, W.; Mindrinos, M.N.; Seok, J.; Cuschieri, J.; Cuenca, A.G.; Gao, H.; Hayden, D.L.; Hennessy, L.; Moore, E.E.; Minei, J.P. A genomic storm in critically injured humans. Journal of Experimental Medicine 2011, 208, 2581–2590, https://rupress.org/jem/article-abstract/208/13/2581/41049. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Li, A.; Gao, X.; Wang, W.; Xu, X.; Yu, L.; Ju, X. (-)-Epigallocatechin-3-gallate (EGCG), a polyphenol, protects against sepsis-induced acute liver injury in rats. International Journal of Clinical & Experimental Medicine, 2016; 9, 177. [Google Scholar]
- Xu, H.; Zhou, Q.; Liu, B.; Cheng, K.-W.; Chen, F.; Wang, M. Neuroprotective potential of mung bean (Vigna radiata L.) polyphenols in Alzheimer’s disease: A review. Journal of Agricultural and Food Chemistry 2021, 69, 11554–11571. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.-C.; Yang, C.-M. Chinese herbs and repurposing old drugs as therapeutic agents in the regulation of oxidative stress and inflammation in pulmonary diseases. Journal of Inflammation Research 2021, 14, 657. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Li, Y.; Shi, N. Attenuation of sepsis-induced rat liver injury by epigallocatechin gallate via suppression of oxidative stress-related inflammation. Tropical Journal of Pharmaceutical Research 2017, 16, 2877–2884. [Google Scholar] [CrossRef]
- Yang, Z.; Zhuang, L.; Lu, Y.; Xu, Q.; Chen, X. Effects and tolerance of silymarin (milk thistle) in chronic hepatitis C virus infection patients: A meta-analysis of randomized controlled trials. BioMed Research International 2014, 2014. [Google Scholar] [CrossRef]
- Yarla, N.S.; Bishayee, A.; Sethi, G.; Reddanna, P.; Kalle, A.M.; Dhananjaya, B.L.; Dowluru, K.S.; Chintala, R.; Duddukuri, G.R. Targeting arachidonic acid pathway by natural products for cancer prevention and therapy. Seminars in Cancer Biology 2016, 40, 48–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, B.; Li, W.; Qin, H.; Yun, J.; Sun, X. The Use of Chinese Skullcap (Scutellaria baicalensis) and Its Extracts for Sustainable Animal Production. Animals : An Open Access Journal from MDPI 2021, 11, 1039. [Google Scholar] [CrossRef] [PubMed]
- Zagorulya, M. (2023). Dendritic cell dysfunction restrains cytotoxic T cell responses against cancer [PhD Thesis]. Massachusetts Institute of Technology. https://dspace.mit.edu/handle/1721.1/150065.
- Zhang, H.; Zhang, H.; Wu, X.; Guo, Y.; Cheng, W.; Qian, F. Fisetin alleviates sepsis-induced multiple organ dysfunction in mice via inhibiting p38 MAPK/MK2 signaling. Acta Pharmacologica Sinica 2020, 41, 1348–1356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Ning, B. Signaling pathways and intervention therapies in sepsis. Signal Transduction and Targeted Therapy 2021, 6, 407. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Yang, J.; Xie, Y. Improvement strategies for the oral bioavailability of poorly water-soluble flavonoids: An overview. International Journal of Pharmaceutics 2019, 570, 118642. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Liu, Y. Cardiovascular protective effect of isorhamnetin, Med. Recapitulate 2008, 15, 2321–2323. [Google Scholar]
- Zheng, Q.; Tong, M.; Ou, B.; Liu, C.; Hu, C.; Yang, Y. Isorhamnetin protects against bleomycin-induced pulmonary fibrosis by inhibiting endoplasmic reticulum stress and epithelial-mesenchymal transition. International Journal of Molecular Medicine 2019, 43, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.T.; Choi, R.C.; Chu, G.K.; Cheung, A.W.; Gao, Q.T.; Li, J.; Jiang, Z.Y.; Dong, T.T.; Tsim, K.W. Flavonoids possess neuroprotective effects on cultured pheochromocytoma PC12 cells: A comparison of different flavonoids in activating estrogenic effect and in preventing β-amyloid-induced cell death. Journal of Agricultural and Food Chemistry 2007, 55, 2438–2445. [Google Scholar] [CrossRef]
Figure 1.
The Structure of Flavonoids and Classes.
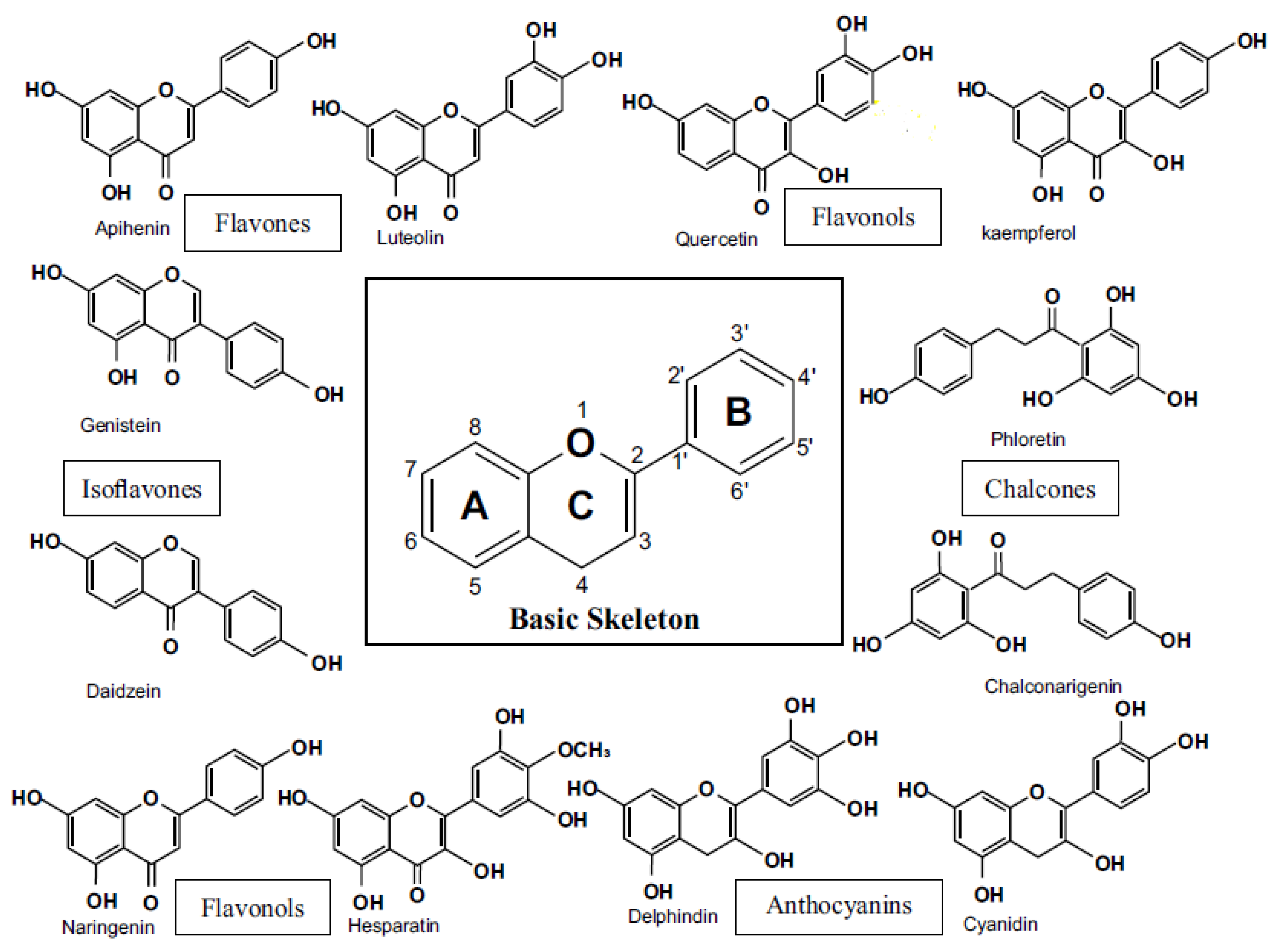
Figure 2.
Structure of Baicalin.
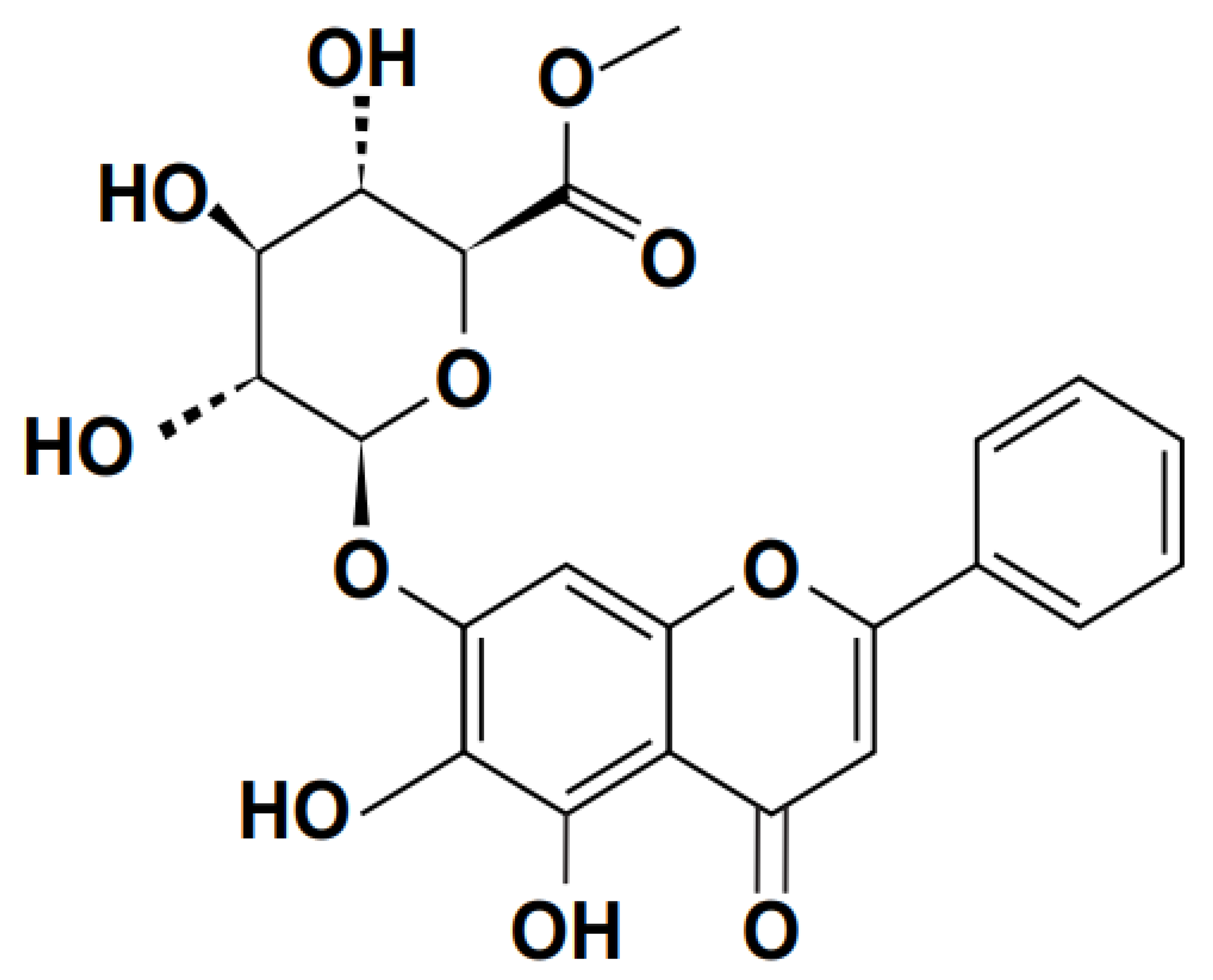
Figure 3.
The Structure of Scutellarin.
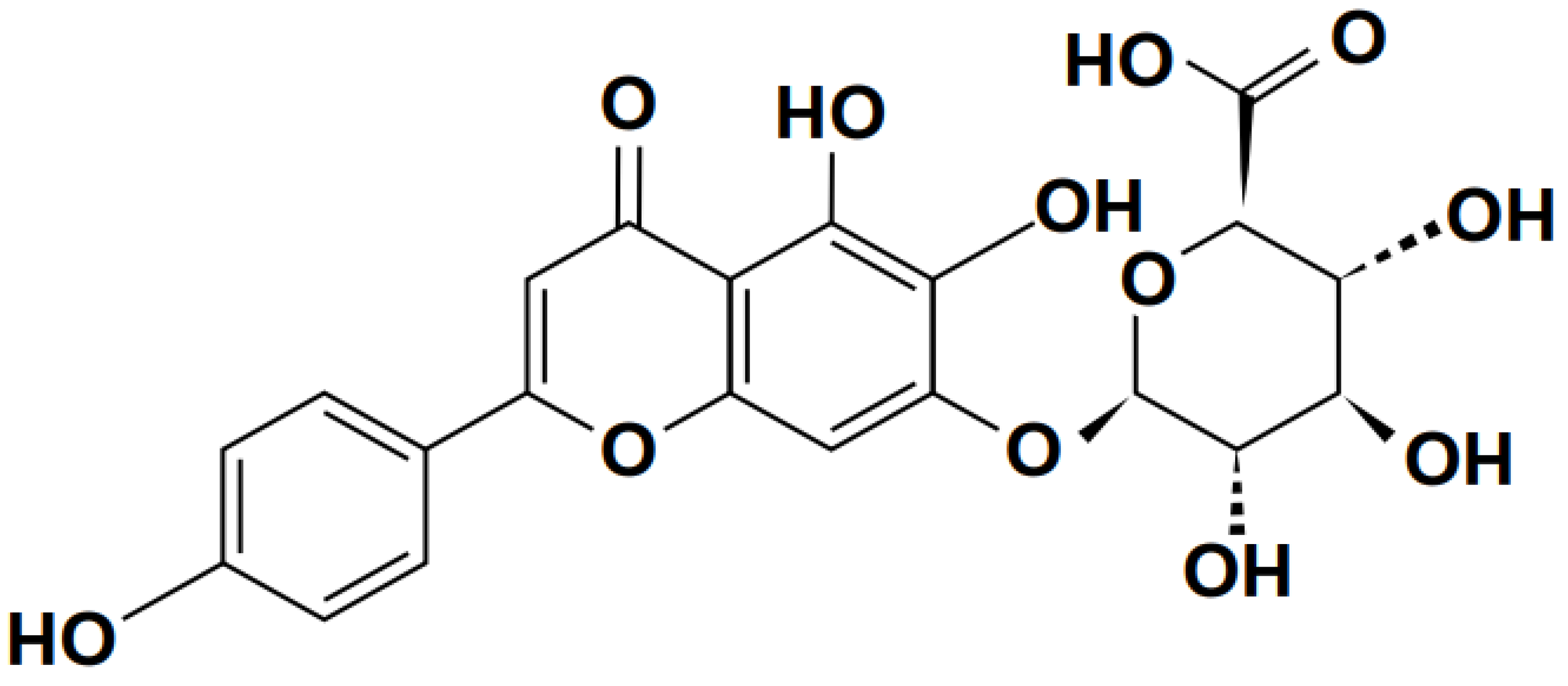
Figure 4.
The Structure of Silymarin.
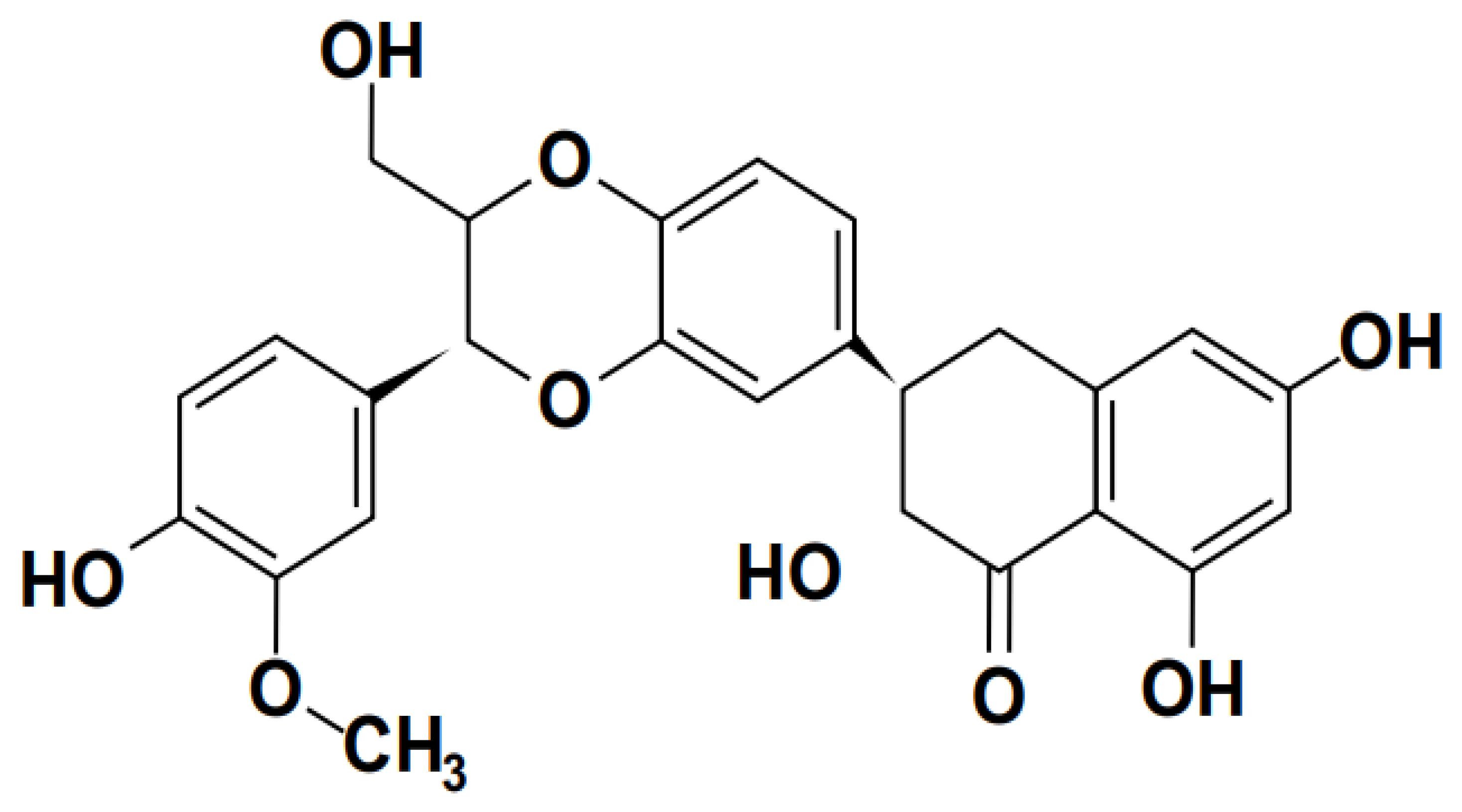
Figure 5.
The Structure of Luteolin.
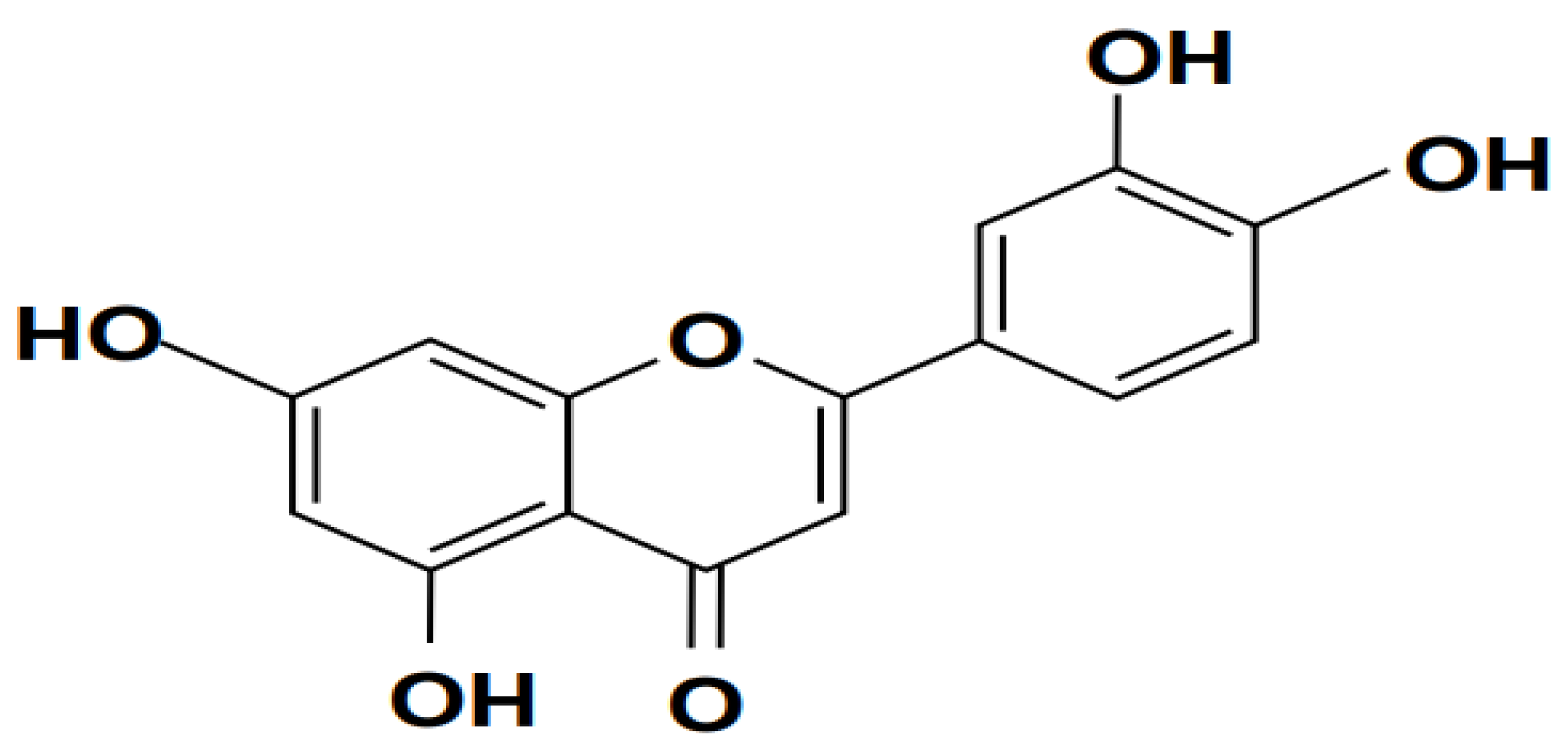
Figure 6.
The Structure of Quercetin.
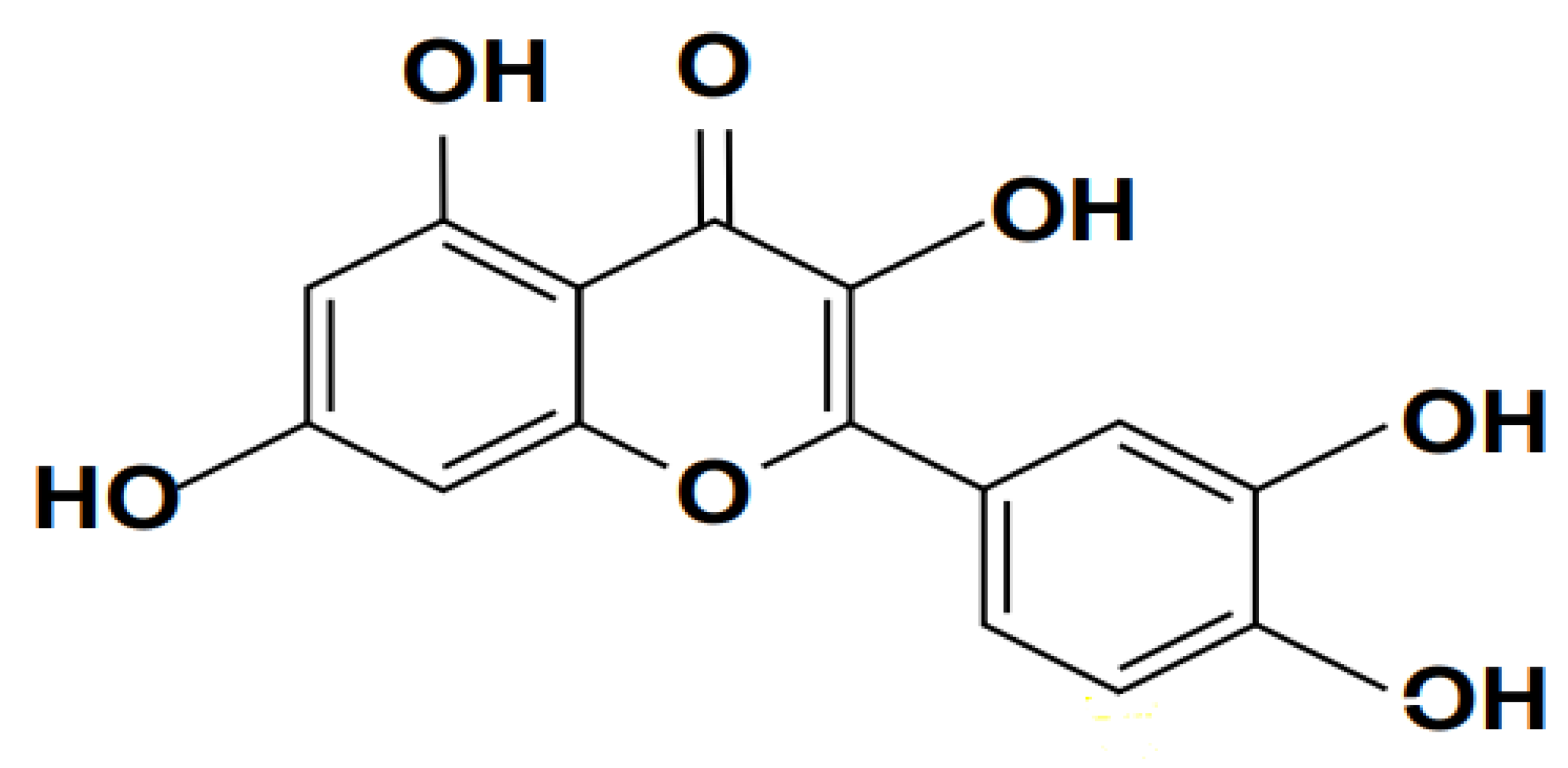
Figure 7.
The Structure of Isorhamnetin.
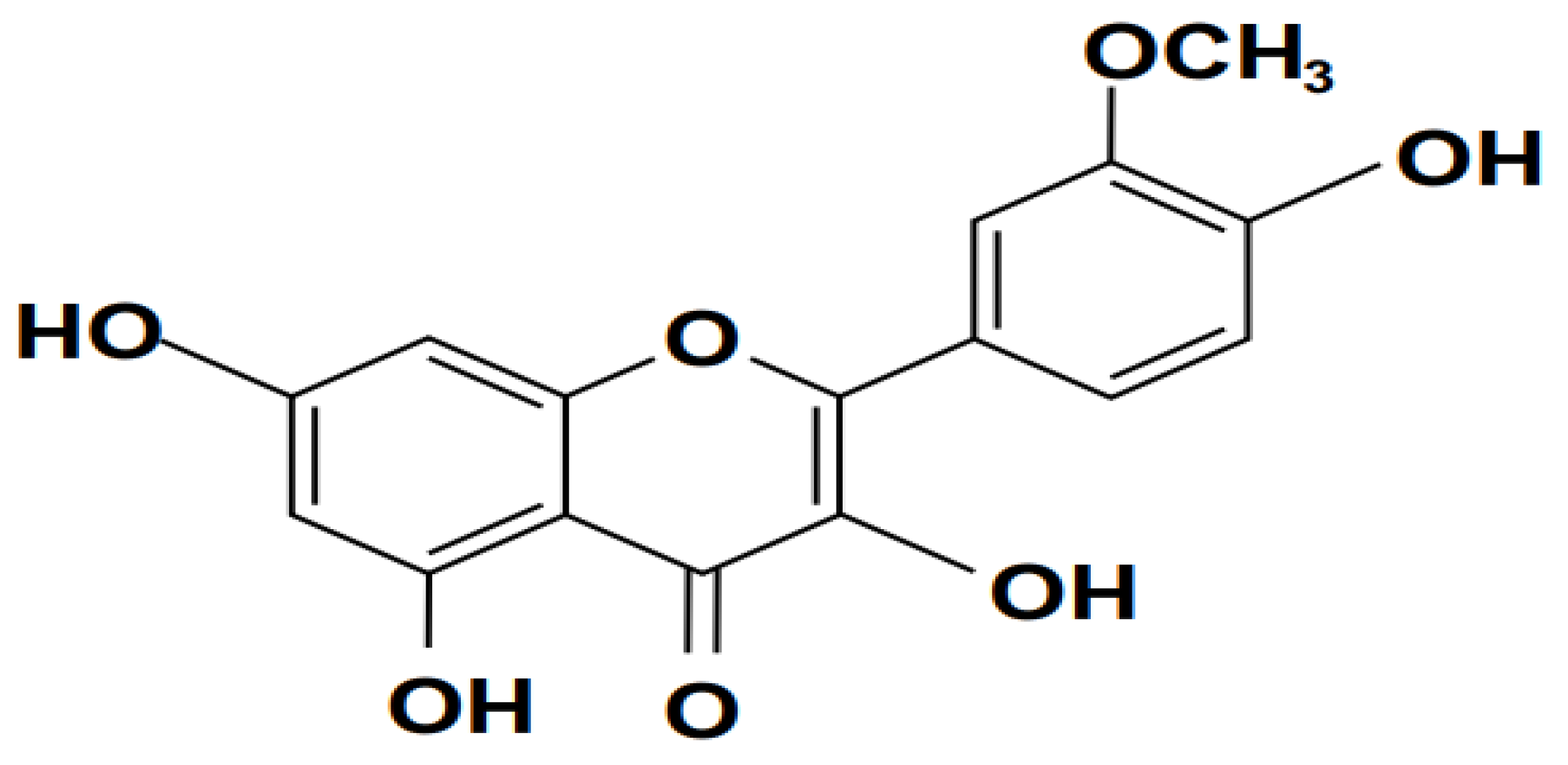
Figure 8.
The Structure of Fisetin.
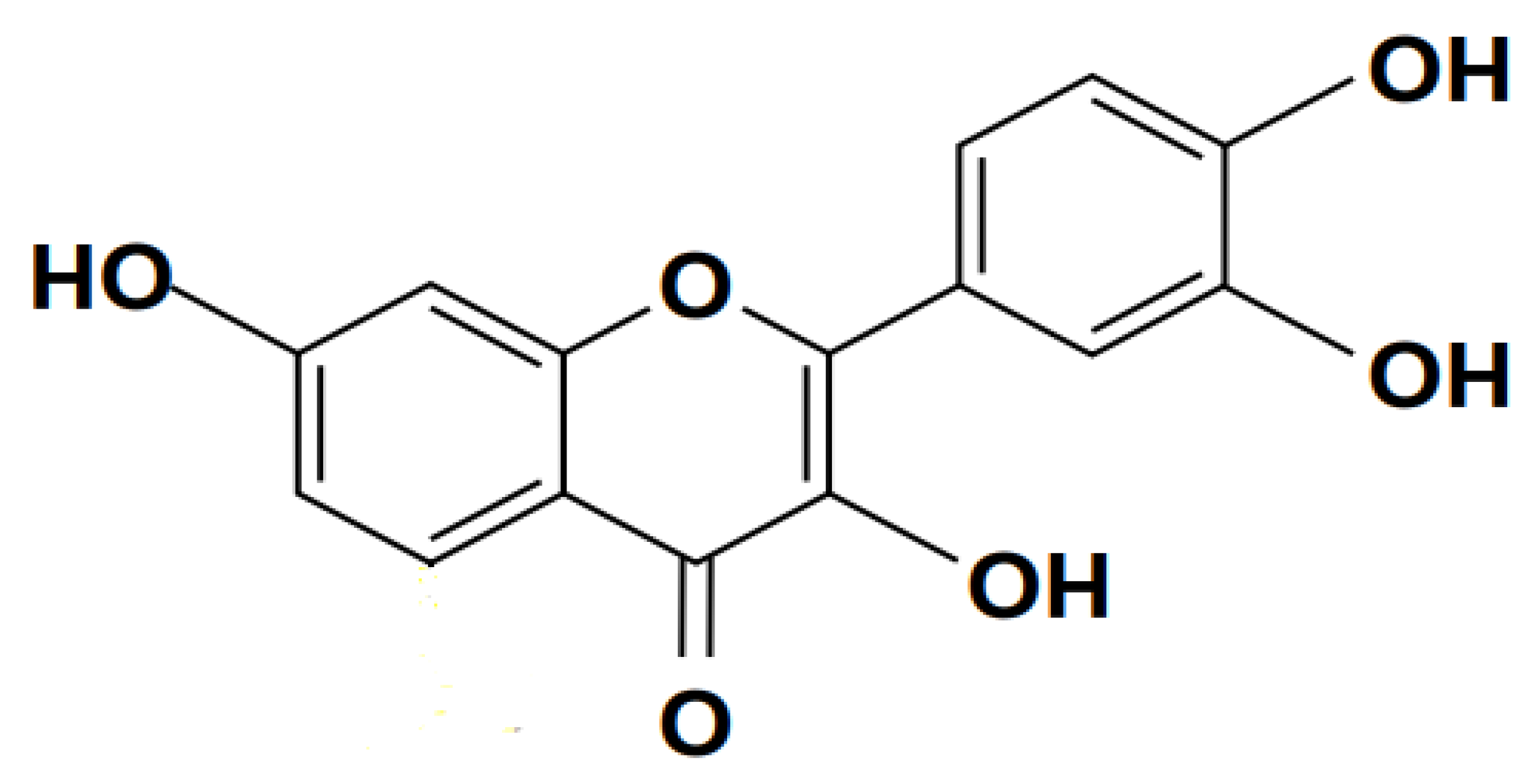
Figure 9.
The Composition of Myricetin.
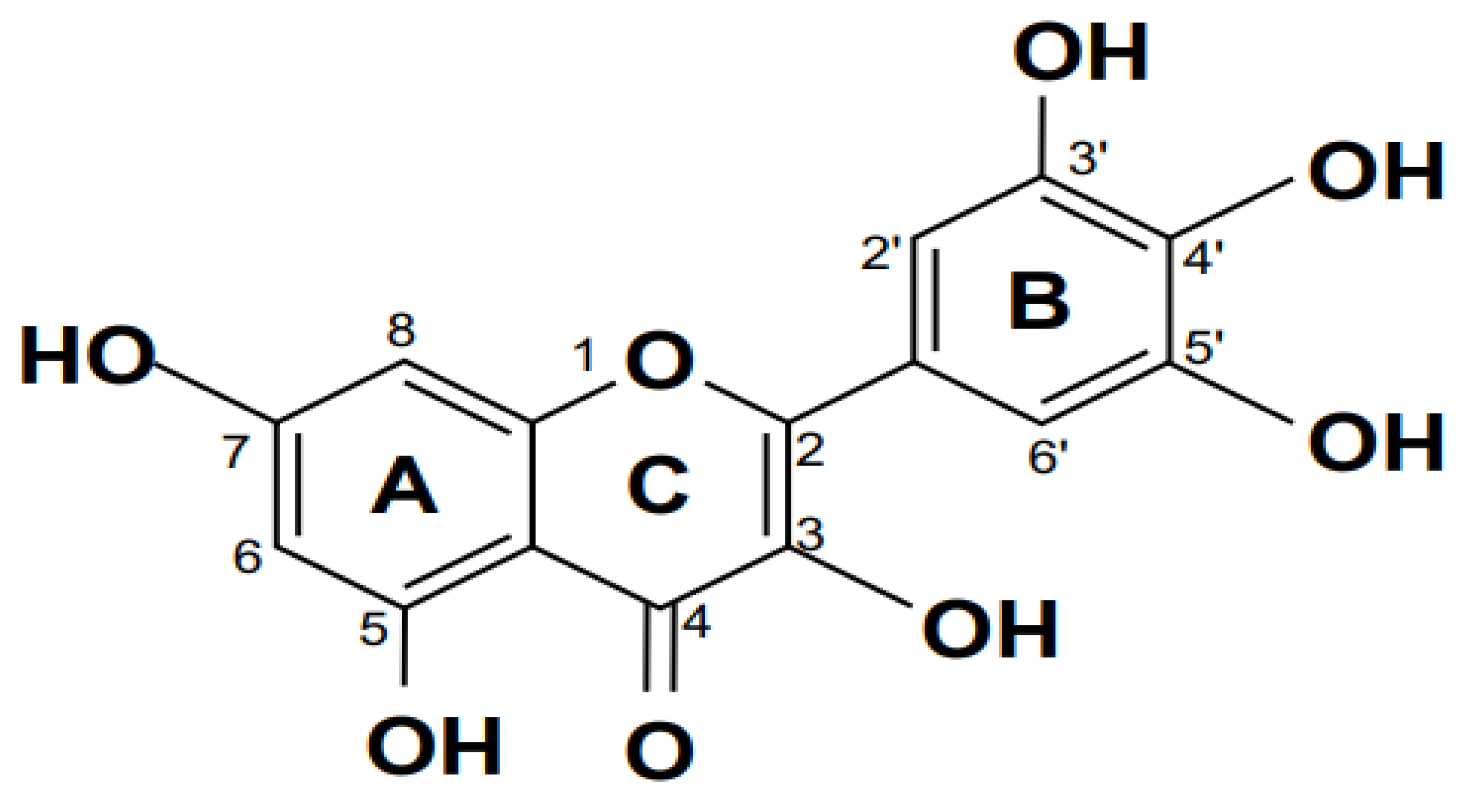
Figure 10.
The Structure of Epigallocatechin-3-gallate (EGCG).
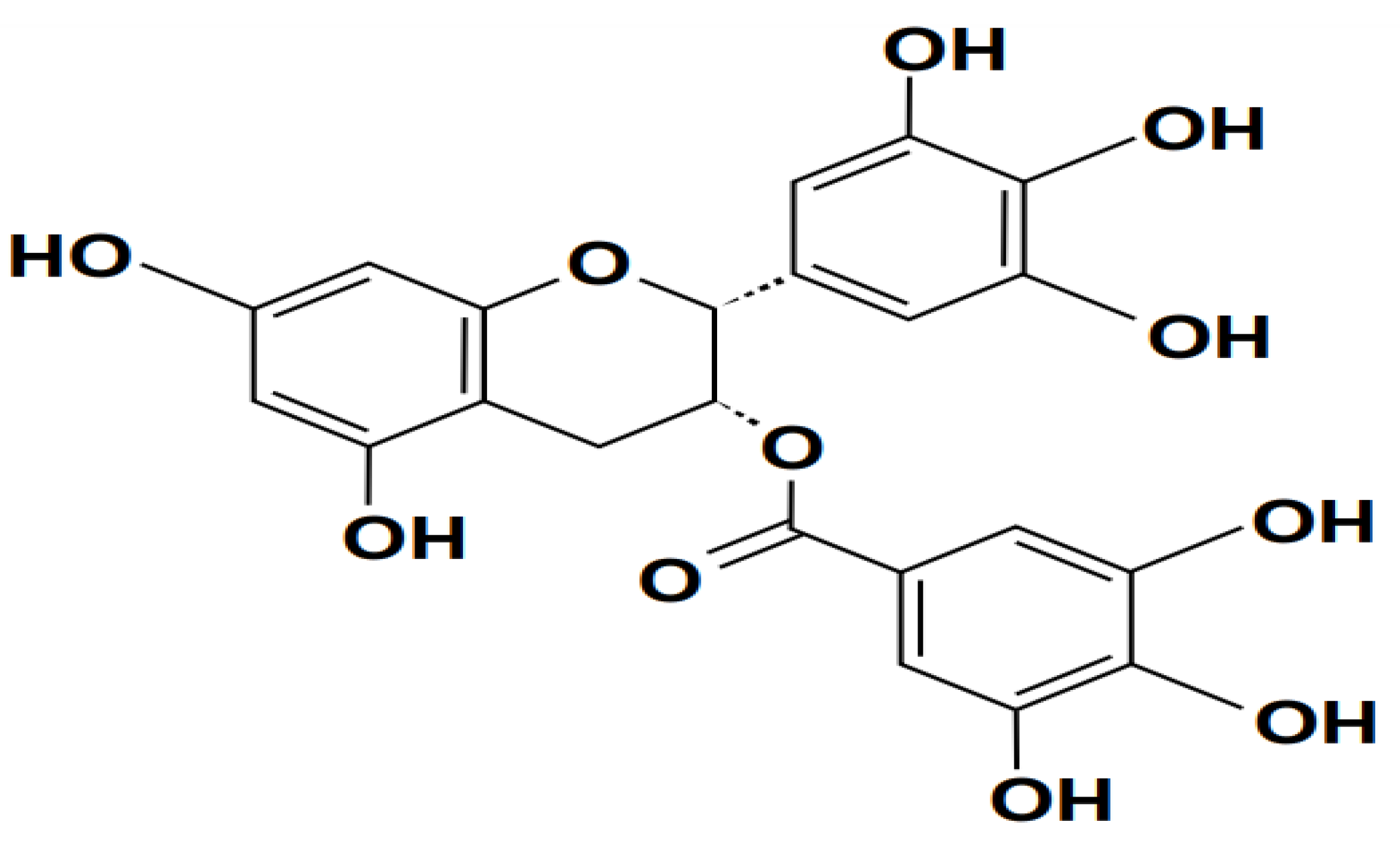

Table 1.
Table of review.
| Year | Author(s) | Title | Purpose/Objectives | Methods | Findings/Results |
|---|---|---|---|---|---|
| 2016 | Panche et al. | Flavonoids: An overview | To provide an overview of flavonoids, including their health benefits and potential therapeutic applications. | Review of existing literature | Flavonoids offer preventive effects against coronary heart disease, anti-inflammatory properties, and therapeutic potential for cancer and infectious diseases. |
| 2018 | Perez-Vizcaino & Fraga | Flavonoids in cardiovascular health: A review | To review the role of flavonoids in cardiovascular health, including their effects on endothelial function, blood pressure, and oxidative stress. | Review of existing literature | Flavonoids exhibit cardiovascular benefits by improving endothelial function, reducing blood pressure, and mitigating oxidative stress. |
| 2019 | Antoce & Stockley | The role of polyphenols in cardiovascular disease risk factors: A review | To examine the role of polyphenols, including flavonoids found in red wine, in mitigating cardiovascular disease risk factors. | Review of existing literature | Polyphenols, including flavonoids, found in red wine, contribute to the reduction of cardiovascular disease risk factors such as inflammation and oxidative stress. |
| 2021 | Micek et al. | Flavonoids - A Review of Their Potential Role in the Treatment of Diabetes Mellitus | To evaluate the potential role of flavonoids in the treatment of diabetes mellitus, including their effects on glucose metabolism and insulin sensitivity. | Review of existing literature | Flavonoids demonstrate potential in managing diabetes mellitus by improving glucose metabolism and enhancing insulin sensitivity. |
| 2017 | De Souza et al. | Methods for evaluation of food and nutrient intake | To review methods for evaluating food and nutrient intake, with a focus on different approaches used in nutrition research. | Review of existing literature | Various methods, such as dietary recalls, food frequency questionnaires, and biomarkers, are employed to evaluate food and nutrient intake in nutrition research. |
| 2020 | Hong et al. | Nutritional evaluation of food: A review | To provide an overview of nutritional evaluation methods used in assessing the nutritional quality of food products. | Review of existing literature | Nutritional evaluation methods include assessing nutrient content, bioavailability, and bioactivity to determine the overall quality of food products. |
| 2016 | Martnez Steele et al. | Ultra-processed foods and added sugars in the US diet: Evidence from a nationally representative cross-sectional study | To investigate the consumption of ultra-processed foods and added sugars in the US diet and their impact on health outcomes. | Cross-sectional study using national dietary data | Consumption of ultra-processed foods and added sugars is associated with poor dietary quality and increased risk of obesity and chronic diseases. |
| 2019 | Neelakantan et al. | Association between ultra-processed food consumption and risk of mortality among middle-aged adults in France | To examine the association between ultra-processed food consumption and risk of mortality among middle-aged adults in France. | Prospective cohort study using dietary data and mortality records | Higher consumption of ultra-processed foods is associated with an increased risk of mortality among middle-aged adults in France. |
| Year | Author(s) | Title | Objectives | Methods | Findings/Results |
| 2007 | Lin et al. | Baicalin: A natural compound with potent pharmacological activities | To investigate the pharmacological activities of baicalin, the primary flavonoid glucoside of Scutellaria baicalensis Georgi. | Review of existing literature | Baicalin exhibits antiviral, bacteriostatic, anticancer, and antioxidant properties, and functions as a non-steroidal anti-inflammatory drug and neuroprotective agent. |
| 2016 | Shi et al. | Baicalin: A review of its anti-cancer effects and mechanisms in hepatocellular carcinoma | To review the anti-cancer effects and mechanisms of baicalin in hepatocellular carcinoma. | Review of existing literature | Baicalin demonstrates anti-cancer effects in hepatocellular carcinoma through various mechanisms, including inhibition of cell proliferation and induction of apoptosis. |
| 2023 | Deng et al. | Pharmacological Properties of Baicalin: A Review | To provide an overview of the pharmacological properties of baicalin, including its role as an anti-inflammatory, neuroprotective, and anticancer agent. | Review of existing literature | Baicalin exhibits a wide range of pharmacological properties, including anti-inflammatory, neuroprotective, and anticancer effects, contributing to its therapeutic potential. |
| 2022 | Bao et al. | Baicalin as a natural product: Anti-oxidant, anti-apoptotic, and anti-inflammatory properties | To investigate the antioxidant, anti-apoptotic, and anti-inflammatory properties of baicalin. | Review of existing literature | Baicalin possesses antioxidant, anti-apoptotic, and anti-inflammatory properties, indicating its potential therapeutic value in various conditions. |
| 2023 | Shahcheraghi et al. | Therapeutic potential of Baicalin in inflammation and organ damage | To explore the therapeutic potential of baicalin in inflammation and organ damage. | Review of existing literature | Baicalin exhibits therapeutic potential in inflammation and organ damage by suppressing pro-inflammatory cytokines, nitric oxide production, and caspase-3 activity. |
| 2019 | Divyakolu et al. | Baicalin as an adjuvant therapy for methicillin-resistant Staphylococcus aureus infection | To investigate the adjuvant therapy potential of baicalin in methicillin-resistant Staphylococcus aureus infection. | Experimental study using animal models | Baicalin shows promise as an adjuvant therapy for methicillin-resistant Staphylococcus aureus infection. |
| 2020 | Song et al. | Baicalin: A potent compound with multiple pharmacological activities | To explore the pharmacological activities of baicalin and its potential as a broad-spectrum antiviral drug. | Review of existing literature | Baicalin exhibits multiple pharmacological activities, including antiviral effects, suggesting its potential as a broad-spectrum antiviral drug. |
| 2020 | Chen et al. | Mechanisms of action of baicalin: A review | To elucidate the mechanisms of action of baicalin, particularly its effects on cytokines and cell pathways. | Review of existing literature | Baicalin modulates various cytokines and cell pathways, contributing to its pharmacological effects and potential as a therapeutic agent. |
| 2021 | Li et al. | Baicalin as a potential therapy for sepsis: Mechanisms and implications | To investigate the potential therapeutic effects of baicalin in sepsis and its underlying mechanisms. | Review of existing literature | Baicalin demonstrates potential as a therapy for sepsis by protecting against liver damage and increasing survival in mice with polymicrobial sepsis. |
| Year | Author(s) | Title | Purpose/Objectives | Methods | Findings/Results |
| 2007 | Zhu et al. | Neuroprotective Effects of Scutellarin | Investigate neuroprotection mechanisms of Scutellarin | Suppressed microglial activation, measured serum (TNF–α, IL–β, IL-6), lactate dehydrogenase activities, tissue glutathione levels. | Scutellarin suppresses microglial activation, reduces serum levels of inflammatory cytokines, and enhances tissue glutathione levels, indicating neuroprotective potential. |
| 2014 | Wang et al. | Scutellarin as a Potential Treatment for Diabetic Retinopathy | Assess efficacy of Scutellarin in diabetic retinopathy treatment | Experimental studies on diabetic animal models, evaluation of retinal histopathology and biochemical markers. | Scutellarin shows promise in treating diabetic retinopathy by mitigating retinal histopathological changes and biochemical markers associated with the condition. |
| 2018 | Chledzik et al. | Anti-inflammatory Properties of Scutellarin | Examine anti-inflammatory effects of Scutellarin | In vitro and in vivo studies, assessment of inflammatory markers, histological analysis. | Scutellarin demonstrates significant anti-inflammatory effects both in vitro and in vivo by modulating inflammatory markers and improving tissue histology. |
| 2015 | Niu et al. | Antioxidant Activity of Scutellarin | Investigate antioxidant potential of Scutellarin | In vitro assays evaluating antioxidant capacity, measurement of ROS levels, assessment of lipid peroxidation. | Scutellarin exhibits potent antioxidant activity by scavenging ROS, reducing lipid peroxidation, and enhancing cellular antioxidant defenses. |
| 2007 | Tan et al. | Apoptotic Effects of Scutellarin on Ovarian and Breast Tumor Cells | Explore apoptotic mechanisms induced by Scutellarin in cancer cells | In vitro cell culture experiments, assessment of apoptosis markers, caspase activation assays. | Scutellarin induces apoptosis in ovarian and breast tumor cells through caspase activation, suggesting potential as an anticancer agent. |
| 2013 | Brent & Shao-Nong | Hepatoprotective Effects of Silymarin | Investigate hepatoprotective qualities and mechanisms of Silymarin | Animal models of liver injury, evaluation of liver histology, biochemical analysis of liver function parameters. | Silymarin exhibits hepatoprotective effects by preserving liver histology, reducing biochemical markers of liver injury, and inhibiting neutrophil infiltration. |
| 2018 | Fan et al. | Pharmacotoxicological Research on Silybin | Examine pharmacotoxicological aspects of Silybin, a major component of Silymarin | Pharmacokinetic studies in animals, assessment of toxicity profiles, evaluation of drug tolerance levels. | Silybin demonstrates favorable pharmacotoxicological profiles, indicating high tolerability and low toxicity levels in animals. |
| 2015 | Chan et al. | Antioxidant and Anti-inflammatory Effects of Silymarin | Investigate antioxidant and anti-inflammatory properties of Silymarin | In vitro assays measuring ROS levels, assessment of inflammatory mediators, animal models of sepsis. | Silymarin exhibits potent antioxidant and anti-inflammatory effects by scavenging ROS, reducing inflammatory mediator levels, and protecting against sepsis-induced organ damage. |
| 2020 | Al-Kadi et al. | Protective Effects of Silymarin Against Sepsis | Explore protective effects of Silymarin against sepsis-induced organ damage | Animal models of sepsis, assessment of liver and kidney function, measurement of inflammatory cytokines. | Silymarin protects against sepsis-induced liver and kidney damage by reducing inflammatory cytokine levels and preserving organ function in animal models. |
| 2017 | Chuammitri et al. | Therapeutic Properties of Luteolin | Investigate therapeutic benefits of Luteolin, including antioxidant, anti-inflammatory, and anticancer properties | In vitro and animal studies, assessment of antioxidant activity, evaluation of anti-inflammatory effects, measurement of tumor growth inhibition. | Luteolin exhibits diverse therapeutic properties, including antioxidant, anti-inflammatory, and anticancer effects, suggesting potential in various disease conditions. |
| 2022 | Pan et al. | Protective Effects of Luteolin Against Ischemia Injury | Evaluate protective effects of Luteolin against ischemia-induced tissue damage | Animal models of ischemia, assessment of tissue damage markers, measurement of ROS levels. | Luteolin protects against ischemia-induced tissue damage by reducing ROS levels, mitigating tissue damage markers, and improving tissue function in animal models. |
| 2022 | Hasan et al. | Anti-Tuberculosis Effects of Quercetin | Investigate the anti-tuberculosis properties of Quercetin | In vitro assays evaluating antimicrobial activity against Mycobacterium tuberculosis. | Quercetin exhibits anti-tuberculosis effects by inhibiting the growth of Mycobacterium tuberculosis. |
| 2018 | Lesjak et al. | Anti-inflammatory Properties of Quercetin | Examine the anti-inflammatory effects of Quercetin | In vitro studies using RAW264.7 macrophages, assessment of inflammatory marker expression, cytokine production. | Quercetin reduces LPS-induced inflammatory responses by suppressing TNF-α release and IL-1β production in RAW264.7 macrophages. |
| 2016 | Luo et al. | Anti-proliferative Effects of Quercetin | Investigate the anti-proliferative effects of Quercetin on cancer cells | In vitro cell culture experiments, assessment of cell viability, proliferation assays. | Quercetin inhibits cancer cell proliferation and induces apoptosis in cancer cells. |
| 2016 | Karuppagounder et al. | Antioxidant Protection by Quercetin | Evaluate the antioxidant protective effects of Quercetin | In vitro and in vivo assays measuring antioxidant enzyme activity, lipid peroxidation levels, ROS scavenging activity. | Quercetin demonstrates potent antioxidant activity by enhancing antioxidant enzyme levels and reducing oxidative stress markers in vitro and in vivo. |
| 2015 | Huang et al. | Quercetin Pre-treatment in Sepsis | Assess the efficacy of Quercetin pre-treatment in sepsis-induced lung injury | Animal models of sepsis, evaluation of lung pathology, measurement of inflammatory cytokines, IL-10 secretion levels. | Quercetin pre-treatment attenuates lung pathology, reduces inflammatory cytokine levels, and increases IL-10 secretion in sepsis-induced lung injury. |
| 2014 | Maalik et al. | Quercetin Effects on Septic-induced Lung Damage | Investigate the effects of Quercetin on lung damage in septic-induced mice | Animal models of sepsis, assessment of lung histopathology, measurement of blood NO, MDA levels, evaluation of antioxidant enzyme activity. | Quercetin reduces lung damage in septic mice by attenuating lung histopathological changes, lowering NO and MDA levels, and enhancing antioxidant enzyme activity. |
| 2018 | Park et al. | Quercetin Suppresses HMGB1 Expression in Sepsis | Examine the effects of Quercetin on HMGB1 expression and oxidative stress in sepsis | In vivo studies using sepsis-induced animal models, assessment of HMGB1 expression, measurement of oxidative stress markers. | Quercetin suppresses HMGB1 expression, reduces oxidative stress, and improves survival outcomes in sepsis-induced animal models. |
| 2019 | Cui et al. | Quercetin Alleviates Lung Damage in Septic Mice | Investigate the protective effects of Quercetin on lung damage in septic mice | Animal models of sepsis, evaluation of lung edema, alveolar capacity, histological analysis. | Quercetin alleviates lung damage in septic mice by reducing lung edema, preserving alveolar capacity, and protecting lung tissues. |
| 2013 | Dai et al. | Quercetin Inhibition of NF-κB Activation | Examine the inhibitory effects of Quercetin on NF-κB activation | In vitro studies using LPS-stimulated macrophages, assessment of NF-κB activation, measurement of inflammatory mediator production. | Quercetin inhibits NF-κB activation and reduces the production of inflammatory mediators in LPS-stimulated macrophages. |
| 2012 | Bharrhan et al. | Quercetin Regulation of Oxidative Enzymes and NF-κB | Investigate the regulatory effects of Quercetin on oxidative enzymes and NF-κB signaling pathways | In vivo studies using animal models, assessment of oxidative enzyme levels, NF-κB expression analysis. | Quercetin regulates oxidative enzyme production and suppresses NF-κB signaling pathways in vivo, reducing oxidative stress and inflammation. |
| 2014 | Wang et al. | Quercetin Reduces Lung Damage and Inflammatory Markers | Evaluate the effects of Quercetin on lung damage and inflammatory markers in sepsis-induced animals | Animal models of sepsis, measurement of inflammatory markers, histological analysis of lung tissues. | Quercetin reduces lung damage and inflammatory markers in sepsis-induced animals, improving survival outcomes and lung function. |
| 2012 | Ozcan et al. | Myricetin Effects on Diabetic Erythrocytes | Investigate the effects of Myricetin on oxidative stress and kidney function in diabetic erythrocytes | In vitro and in vivo studies, assessment of oxidative stress markers, measurement of kidney function parameters. | Myricetin reduces oxidative stress and improves kidney function in diabetic erythrocytes, suggesting potential therapeutic benefits in diabetes. |
| 2021 | Imran et al. | Anti-inflammatory Effects of Myricetin | Examine the anti-inflammatory effects of Myricetin on sepsis-induced inflammation | In vitro and in vivo studies using sepsis-induced animal models, assessment of inflammatory markers, NF-κB signaling analysis. | Myricetin attenuates sepsis-induced inflammation by suppressing NF-κB signaling and reducing inflammatory mediator production in vitro and in vivo. |
| 2013 | Lee et al. | Fisetin Inhibition of HMGB1 Release in Vascular Inflammation | Investigate the inhibitory effects of Fisetin on HMGB1 release and vascular inflammation | In vivo studies using animal models of vascular inflammation, measurement of HMGB1 levels, assessment of vascular function. | Fisetin inhibits HMGB1 release, reduces vascular inflammation, and improves vascular function in animal models of vascular inflammation. |
| 2020 | Zhang et al. | Fisetin Protection Against CLP-induced Organ Damage | Assess the protective effects of Fisetin against organ damage induced by CLP in sepsis | Animal models of sepsis, evaluation of organ pathology, measurement of inflammatory markers, assessment of oxidative stress. | Fisetin protects against CLP-induced organ damage in sepsis by attenuating organ pathology, reducing inflammatory markers, and alleviating oxidative stress. |
| 2019 | Grynkiewicz & Demchuk | Fisetin as a Promising Antioxidant | Investigate the antioxidant properties of Fisetin | In vitro assays evaluating antioxidant capacity, measurement of ROS levels, assessment of lipid peroxidation. | Fisetin exhibits potent antioxidant activity by scavenging ROS, reducing lipid peroxidation, and enhancing cellular antioxidant defenses. |
| 2016 | Almatroodi et al. | Pharmacological Activities of Epigallocatechin-3-gallate | Explore the pharmacological activities of EGCG | In vitro and in vivo studies, assessment of anti-inflammatory, antibacterial, anticancer effects. | EGCG demonstrates diverse pharmacological activities including anti-inflammatory, antibacterial, and anticancer effects in vitro and in vivo. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



