
Submitted:
20 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
In southern South America, zonal flora, defined by macroclimatic conditions, is strongly influenced by an aridity filter near 29°S. Cushion bogs, high-altitude wetlands along the Andes, display homogeneous flora with cushion species dominance, contrasting with zonal vegetation. Despite being influenced by microclimatic conditions, these ecosystems experience varied environmental effects. This study identifies environmental filters affecting bog communities along a broad-scale latitudinal gradient from 15°S to 42°S.
We analyzed 420 bogs and 284 species across three macroclimatic regions with distinct summer, winter, and transitional arid rainfall regimes. Using variance partitioning and membership-based regionalization models, we examined the impacts of climatic, edaphic, and spatial variables on beta diversity. We also assessed species' niche overlap and the influence of environmental filters on the communities' phylogenetic diversity.
Results show that species turnover and niche overlap vary with macroclimatic differences, delineating three distinct regions. Notably, phylogenetic clustering in the driest part of the gradient (23°S —24°S) highlights the impact of the environmental filter. Aridity and temperature variations at a broad scale serve as environmental filters shaping the composition of bog communities across southern South America.
Keywords:
bogs
; environmental filters
; Andes
; phylogenetic diversity
1. Introduction
The uplift of the Andes represents one of the most significant events in the biogeographic history of South America [1], serving as a vicariant barrier, a biological corridor, and a source of new ecosystems and conditions in extra-Andean areas [2,3,4,5]. The influence of this mountain range on the climatic and biotic history is partly attributed to its impact on atmospheric circulation [6], affecting the distribution of precipitation along the latitudinal gradient [3].
Macroclimatic conditions, alongside diverse histories of colonization and in situ speciation, shape the zonal vegetation associated with the Andes, influencing its structure and composition [2,7]. In southern South America, a biogeographic break is observed near 27°S in the Andes, which divides the ecosystems of the Puna to the north and the Andean steppe to the south [8]. Each is influenced by different precipitation regimes: summer rainfall from easterly winds and winter rainfall from westerly winds, respectively (Figure 1) [3,8]. The aridity resulting from the shift between winter and summer precipitation regimes acts as an environmental filter that limits north-south dispersal, particularly around 29°S [9]. This phenomenon correlates with a turnover of species, a decrease in richness, and changes in life forms [9,10,11].
Additionally, microclimatic conditions emerge in the context of the interaction between Andean topography and the local environment [12,13], conducive to establishing azonal ecosystems such as Cushion Bogs [8,9]. These high Andean wetlands, also known as “Bofedales”, “Vegas”, or “Mallines” [14,15], develop near the hydrological and altitudinal limits for plant life in the Andes [14] from Colombia/Venezuela to Patagonia [15,16,17]. They are dominated by “bogs-forming” plants, which grow in compact cushions, capable of forming peat, retaining moisture, altering local hydrological conditions, and creating conditions for the colonization of other species [15,18,19]. These plants, mainly Juncaceae like Oxychloe, Distichia, and Patosia, along with Cyperaceae such as Zameiocirpus [15], show variations in their distribution. For instance, Distichia dominates bogs with a tropical and subtropical distribution from Colombia to northern Chile and Argentina. At the same time, Oxychloe is found in subtropical and extratropical distributions from southern Peru to central-southern Chile and Argentina [20,21].
Bogs represent a complex of different species interlocked with each other [22,23,24]. Generally, the flora composing these ecosystems is characterized by rapid vegetative reproduction, high seed production, and both endozoochoric and epizoochoric dispersion [15], as is the case with the dominant Juncaceae, which exhibit clear adaptations for dispersion by birds [9]. Moreover, many of these species demonstrate an anemophilous pollination strategy, and a significant number are considered autogamous [9]. It is proposed that these reproductive characteristics have played a crucial role in the homogeneity of bog flora compared to zonal flora along the latitudinal gradient, regardless of macroclimatic conditions [9,10]. However, bogs are affected by small-scale climatic conditions; for instance, the influence of elevation and temperature on the change in dominance of Juncaceae species has been documented [15,25,26,27,28], as well as the effect of the physicochemical characteristics of associated waters [9,15,16]. Moreover, significant impacts of human activity on floristic composition have been identified [15,22,23], but above all, the aridity-humidity gradients have been proposed as the main factor shaping these communities’ composition [9,10,15,25,26,27,29,30].
The “Environmental Filters” hypothesis proposes that environmental factors restrict communities by selecting species through a hierarchical process from large-scale filters to microclimatic factors and biotic interactions [31,32], leaving a mark on the species composition of communities [33,34], that is, on beta diversity, a measure of the differences in species composition between communities [35,36]. Beta diversity can be broken down into two components: (1) turnover, which is the replacement of species between sites, and (2) nestedness, which refers to the loss of species from one site, transforming into a subset of another [37,38]. These components provide insights into the underlying processes driving differences in species composition. Turnover is generally associated with niche diversity along the study gradient, while nestedness is related to spatial variables and dispersal [38,39,40].
Environmental filters also impact the phylogenetic structure of communities, manifesting in patterns of phylogenetic clustering, where community members are more closely related due, for instance, to shared traits for persistence in a particular environment [41,42]. In contrast, in the absence of environmental filters, a pattern of phylogenetic overdispersion emerges, where individuals are not closely related, and interactions predominate [41,43,44,45].
These backgrounds on the biogeographic patterns of zonal and azonal vegetation in the Andes, as well as the various environmental effects that influence the structure of communities, guide the research question in this work: What environmental filters explain the floristic composition of bogs on a large scale along the latitudinal gradient of southern South America? We hypothesize that the variation in precipitation patterns, particularly between the northern macroclimatic areas, dominated by summer rains, southern areas, dominated by winter rains, and the transition between both precipitation regimes, differentiates the bog communities along the latitudinal gradient of their distribution.
Accordingly, it is predicted: (1) An increase in beta diversity towards the extremes of the latitudinal gradient, primarily due to species turnover or rotation; (2) Clustering of communities, based on beta diversity, that differentiates the macroclimatic regions north, transition, and south of the precipitation regimes; (3) Greater overlap of climatic niches among species within regions than between macroclimatic regions; and (4) Phylogenetic clustering in communities within each macroclimatic region. This study aims to determine the influence of environmental filters on the differentiation on a large scale of bog communities along the latitudinal gradient of the Andes.
2. Results
2.1. Beta Diversity and Environment
2.1.1. Variation Partitioning
The variation in the community matrix along the gradient is explained by 36% (Figure 2a), attributed to the following factors: (1) pure spatial (10%) and (2) pure environment (5%). Breaking down the environmental factors (Figure 2b), 35% of the variation is explained by: (1) pure spatial (10%), (2) pure temperature (2%), (3) pure precipitation (1%), and (4) pure soil (1%).
2.1.2. Beta Diversity and Its Components
Total beta diversity (BDtotal) indicates a dissimilarity of 0.37, suggesting low species variation along the gradient, with contributions of 68% by the turnover component and 32% by nestedness. Mantel tests (Figure 3) showed that dissimilarity between sites increases with geographic and environmental distance similarly observed for the turnover component; however, the explanatory power of nestedness was very low (R2 of 0.03 and 0.02, respectively; Figure 3c,f).
2.1.3. Clustering and Regionalization
The Sorensen index (S) identified nine distinct groups, two consisting of only one site each (Figure 4a). Clusters at the geographical extremes exhibit more significant dissimilarity between them. Certain groups are endemic to specific geographic zones: groups 9 and 6 are exclusively associated with the northern area, while groups 4 and 5 are characteristic of the southern zone. The remaining clusters are heterogeneously grouped in the transition zone.
The regionalization by the Grade of Membership model (Figure 4b) delineates three macro zones, hereafter referred to as bioregions, moving the boundaries of each proposed operational zone northward around 22°S and 32°S.
2.2. Niche Overlap
Greater intra-regional than inter-regional niche overlap was observed, with all comparisons statistically significant (Figure 5). Species in the northern bioregion show an average overlap of 57%, transitional species 66%, while southern species exhibit an average overlap of 27%; however, this low value is due to one species having a limited environmental niche but is almost completely nested within the other two (see “South” in Figure S2).
The lowest average overlap (0.05%) occurs between species from the northern and southern bioregions, differentiated by: (1) Isothermality (Bio3), (2) Annual Temperature Range (Bio7), and (3) Precipitation of the Coldest Quarter (Bio19). The transition zone shows a climatic affinity with the north of 22% average overlap, differentiated by (1) Mean Annual Precipitation (Bio12), (2) Isothermality (Bio3), and (3) Annual Temperature Range (Bio7). Between the southern and transitional species, the average overlap is 16%, with main differences in (1) Isothermality (Bio3), (2) Mean Annual Precipitation (Bio12), and (3) Precipitation of the Coldest Quarter (Bio19).
2.3. Phylogenetic Diversity
2.3.1. Bioregions
Observed phylogenetic diversity (PD) shows a north-south increasing pattern (Figure 6), indicating that southern communities are more heterogeneous regarding evolutionary histories from a phylogenetic perspective. Comparing PD metrics, MPD (mean pairwise distance) and MNTD (mean nearest taxon distance) against the null model show no significant difference indicating clustering or overdispersion at the scale of these bioregions (Table 1).
2.3.2. Latitudinal Bands
The increasing observed PD north-south pattern no longer presents; instead, there is a fluctuation along the latitudinal gradient (Figure 7). Comparing PD, MPD, and MNTD against the null model reveals a pattern of overdispersion from 22°S northwards and clustering towards the south, except for the southernmost band. However, it is only significant in the 23°S-24°S band for PD (Table 2), indicating that the species within these communities are more phylogenetically related to each other than by chance, suggesting an environmental filter that restricts them.
3. Discussion
3.1. Beta Diversity and Environment
3.1.1. Variation Partitioning
The results from the variation partitioning analysis are enlightening in several respects. Firstly, they indicate that spatial factors understood as limitations to dispersion [46,47], contribute the most on their own, while the isolated contribution from the environment is very low. This suggests that structuring the floristic community in bogs is more influenced by landscape configuration and habitat connectivity than by macro-environmental conditions, contrary to studies in other mountainous regions for zonal vegetation [48]. However, their contributions appear more balanced when considering the synergistic effects of environmental and spatial factors. This indicates that although spatial factors are crucial, their interaction with the environment cannot be ignored, reflecting that while species may disperse across the landscape, environmental conditions ultimately determine their establishment and persistence.
Secondly, the detailed analysis of environmental factors revealed that, even though the isolated effects of environmental factors such as temperature, precipitation, and edaphic variables have a minor or negligible contribution to the structuring of these communities compared to spatial factors, their importance is significantly magnified when considered in interaction with other factors. These results suggest that structuring floristic communities in bogs is not governed exclusively by a single environmental or spatial factor but rather by the complex interaction of multiple factors.
Furthermore, 64% - 65% of the variance remains unexplained in both models, suggesting the influence of unconsidered factors such as biological interactions, the sources and physicochemical characteristics of the waters associated with bogs, topography (slope), and anthropogenic impact variables, all of which have evidenced their impact on floristic composition [14,15,30,49].
3.1.2. Beta Diversity and Its Components
Despite the low BDtotal value in the results, which could be because of under-sampling, either from original data collection or from species ‘cleaning’ for analysis, it is consistent with previous studies in bogs of South America showing a significant number of species common along the latitudinal gradient [9,10,16,25,29]. This directly impacts the Sorensen dissimilarity index due to the duplication in the importance of shared species, consequently increasing floristic similarity along the gradient. Moreover, the turnover component emerges as the main contributor to beta diversity in the study area, indicating a mechanism of species replacement in response to environmental changes. This aligns with studies by Casagranda & Izquierdo [28] and Méndez [27], which show a turnover in dominant species under colder conditions and at higher altitudes. Arroyo et al. [9] also described species turnover in Andean zonal vegetation, observing that under an aridity gradient, species are replaced by plants more adapted to those conditions. They also noted that bogs, when facing a reduction in area due to aridity, lose fewer species than zonal vegetation, probably due to the constant reintroduction of species through birds and livestock. This explains the low contribution observed by the nestedness component, suggesting that species loss occurs at smaller scales, under gradients of physicochemical water conditions [15,16], or even under human activity, which strongly influences species loss due to the effects of livestock, water extraction, or peat harvesting [25,30].
These explanations also help understand the positive relationships between the dissimilarity of each component (S, turnover, and nestedness) and geographic and environmental distance, a pattern that aligns with other ecosystems, such as tropical forests [50] or relic forests of Chile [51]. The Mantel tests were significant; however, the nestedness (R²) explanatory power was very low, revealing no clear pattern. In contrast, the turnover component showed a pattern like S, so it was decided to continue working with the latter to avoid overestimating the environmental effect.
3.1.3. Clustering and Regionalization
The hierarchical clustering analysis identified 9 groups based on the Sorensen dissimilarity index, structured similarly to the three proposed operational zones. However, due to the sensitivity of this type of analysis to very homogeneous communities, noisy data, and outliers [52], situations like the one observed occurred, where two groups were formed by a single site each. Additionally, Ruthsatz [29] clarifies that the floristic lists for Chile are incomplete, mainly due to limited time and season, as many plants were observed in the vegetative stage without characteristic organs allowing identification. [53,54]. This study used a membership grade model to transform clusters into bioregions. These models offer a more flexible and detailed way to analyze regions and their biotic transitions, particularly useful in cases of gradual transitions and for studies aiming to understand the influence of various factors on regional differences (see [55]). This result, in part, concurs with the hierarchical clustering and with the operational zones defined for this study, with the difference that it pushes the boundaries slightly northward but still coincides with the macroclimatic conditions. It is interesting to observe how the transition bioregion, associated with greater aridity, extends up to northern Chile on the western slope of the Andes, which makes sense since that area corresponds to the “dry Puna,” with a rain shadow effect from the mountain range on the easterly winds [3,8,14].
This grouping into the three bioregions provides evidence of the environmental effect on large-scale community assembly and strongly aligns with the three phytogeographic districts proposed by Biganzoli et al. [56] for the Andean Province of Southern South America, which in turn coincide with the macroclimates present in Chile that extend to Argentina [57].
3.2. Niche Overlap
The analysis of niche overlap suggests a significant separation between communities at the extremes of the gradient (North-South). This differentiation is primarily due to factors such as Isothermality, Annual Temperature Range, and Precipitation of the Coldest Quarter. Northern communities are more adapted to stable temperature conditions throughout the year, while southern communities show adaptations to greater temperature variations and more intense precipitation during the coldest month, which are characteristic of the respective macroclimates for the bioregions [8,14]. These findings support the hypothesis that macroclimatic conditions influence these azonal ecosystems, consistent with previous research [15,25,29,30].
The comparison between the transition bioregion and the northern and southern bioregions reveals substantial differences in climatic niches. These differences are mainly attributed to a decrease in annual precipitation in the transition zone. This pattern aligns with multiple studies examining the effects of aridity on the composition of bogs and zonal vegetation [9,10,15,25,29,30,58]. In the transition bioregion, the versatility of species in terms of their climatic niches is notable when compared to those of the north and south. They show a greater affinity for aridity and include temperature variations that separate northern and southern species, likely due to the presence of species from these bioregions in the transition zone, which aligns with observed floristic affinities (Figure 4). This diversity allows them to occupy a broad spectrum within the climatic gradient, consistent with studies that include transition zones in South America [59].
3.3. Phylogenetic Diversity
The results from phylogenetic diversity metrics derived from using the three bioregions indicate an observed PD pattern that increases from north to south, consistent with studies on the flora of Chile [60]. However, this pattern is not observed in the analysis conducted by latitudinal bands. This discrepancy is due to high PD between 27°S and 30° (Figure 7), sites that, according to previous hierarchical clustering and the membership grade model (Figure 4), contain species more related to the communities of the north and south, so greater sampling resolution affects the large-scale PD pattern. The significant standardized effect of PD indicates phylogenetic clustering around 23°S and 24°S, a signal of phylogenetic conservatism in niche preference [60], likely associated with the arid conditions at that latitude, functioning as an environmental filter. Although other results are not significant, both PD, MPD, and MNTD show a clear tendency towards phylogenetic overdispersion north of 23°S and phylogenetic clustering to the south (Table 2), results that contradict observations for the zonal flora of Chile [60] but are consistent with global patterns [61].
These findings should be approached with caution, as many of the species included in the study were not present in the mega-tree used for analysis and were added by inserting the species at the basal node of the genus. If a family or genus has only one representative in the tree, the branch is split to represent taxonomic levels and new species are added at corresponding points of this split (see [62]), reducing the PD. Therefore, it is advisable to work on more detailed phylogenies for high Andean species, such as the phylogeny proposed by Brozova et al. [63] for Juncaceae and Cyperaceae, a phylogeny that has been criticized by some authors (see [64]).
Overall, the results obtained, both in niche overlap and PD analysis, reinforce the predominance of the environmental filter over the role of stochasticity suggested by neutral theory. This theory attributes a fundamental role to ecological drift, random dispersion, and speciation in determining community composition [65]. However, our findings strongly support the niche theory, where environmental factors are key in community assembly [66,67], through the joint effects of the variables studied. Among these variables, dispersal capacity stands out, influenced by spatial factors and determined by mechanisms such as endozoochoric and epizoochoric dispersal, linked to birds and their migratory routes, as well as the movement of livestock and native ungulates like guanacos and vicuñas [9,15]. Other significant elements are aridity and rainfall regimes, which, while contributing modestly to the variation of taxonomic diversity, are decisive in the climatic characteristics of communities along the latitudinal gradient in southern South America and in the restriction of phylogenetic communities.
In relation to the niche conservatism hypothesis [68], the notable divergence in climatic niches, as well as trends in dispersion and phylogenetic clustering observed in the north and south, respectively, support hypotheses by Cleef [16] and Ruthsatz [25] about different colonization histories for bogs plants, with contributions from Antarctic austral origins that advanced as suitable conditions were generated alongside the rise of the Andes, finding taxa in tropical environments with influences from Holarctic species [2,16]. Subsequently, speciation processes with specific adaptations [69] contributed new elements to the mixing zone in the arid conditions of the gradient.
4. Materials and Methods
4.1. Study area
The study area includes the eastern and western slopes of the Andes, spanning from 15°S to 42°S across Peru, Bolivia, Argentina, and Chile. The latitudinal gradient was divided into three operational zones (macroclimatic regions) based on rainfall regimes (Figure 8): (1) North, from 15°S to 28°S, influenced by summer rains from the easterly winds; (2) Transition, from 29°S to 35°S, an area between summer and winter rainfall with varying precipitation distribution annually and interannually; and (3) South, from 35°S to 41°S, affected by winter rains from the westerly winds.
4.2. Data Preparation
A presence/absence matrix (1 and 0) was constructed using floristic lists from Ruthsatz [15,25,26,29,30], selecting only angiosperms and excluding uncertain identifications marked as “spec”, “cf”, “aff”, and those with two epithets (e.g., Luzula racemosa/vulcanica). Varieties and subspecies were recorded under the species of the first epithet, and “sl” (sensu lato) species were retained as such. Nomenclature was updated using the Vascular Plants Catalogue of the Southern Cone [70], the Vascular Plants Catalogue of Chile [20], and the Vascular Plants Catalogue of Bolivia [71]}, resulting in a total of 420 sites and 284 species (Table S1).
4.3. Environmental Variables
Climatic data included the 19 bioclimatic variables from CHELSA 2.1 [72] for the period 1981-2010, proven effective in mountainous regions [73]. Elevation data was extracted from CHELSA 2.1 [72], and 9 edaphic variables at a depth of 60-100 cm were obtained from ISRIC-World Soil Information [74]. All variables were used at a 30” resolution (Table 3).
4.4. Data analysis
4.4.1. Variation Partitioning
Using the software R 4.3.2 [75], the environmental variables (climatic + edaphic) set was optimized by excluding those with a correlation coefficient >0.75, preferring those with the highest contribution in a principal components analysis. Geographic distance between sites was calculated using the distm function from the geosphere 1.5-18 library [76], and Moran’s Eigenvalues (MEM) were computed for each site. From the complete set of variables (environmental + MEM + elevation), those with a significant effect were selected using the forward selection method, applied to the raw presence/absence matrix via the forward.sel function of the adespatial library (with 999 permutations and alpha = 0.5) [77].
All variable values were normalized to ensure comparability (Table 4). This allowed for a variance partitioning analysis, decomposing the contribution of environmental (temperature + precipitation + soil) and spatial components (geographic distance (MEM) + elevation) to the original presence/absence matrix [78]. The contribution of each environmental and spatial variable was evaluated using the varpart function of the vegan library [79], with significance tested through ANOVA.
4.4.2. Beta Diversity and Its Components
Beta diversity among sites was assessed using the Sorensen dissimilarity index from the presence/absence matrix. Total beta diversity (BDtotal) corresponding to the total variance of the community matrix (see [80]) was calculated and partitioned into its components of turnover (ReplS) and nestedness (RichDiffS), as shown in Table 5, using Podani family indices [38](Legendre, 2014) with the beta.div.com function of the adespatial 0.3-23 library [77].
The relationship between beta diversity and the environment was analyzed through a permutation-based forward selection analysis using non-correlated environmental variables (climatic and edaphic), as shown in Table 6, transforming them into Euclidean distance matrices via the vegan 2.6-4 library [79]. A Mantel test was then used to analyze the correlation between environmental and geographic distance with beta diversity and its components, using the mantel function of the vegan library [79].
4.4.3. Clustering and Regionalization
Based on beta diversity, a hierarchical clustering analysis was conducted on sites to identify clusters along the latitudinal gradient. The dendrogram was constructed using the UPGMA clustering algorithm, with an optimal number of clusters set to nine. The best clustering algorithm was selected using the select_linkage function based on the cofenetic correlation coefficient of Sokal and Rohlf [81], and the optimal number of clusters was determined using the “elbow” method (see [82,83]) from the function optimal_phyloregion, both from the phyloregion 1.0.8 library [84].
A membership grade model was then developed to regionalize the sites into three areas (based on the proposed macroclimatic regions). This process involved fitting the model to the original presence/absence matrix and assigning bioregions based on the taxonomic diversity probability of each site belonging to each bioregion (see [55]). These steps were carried out using the fitgom function of the phyloregion library [84].
4.4.4. Niche Overlap
To measure niche overlap, three representative species from each bioregion were selected using the “Kullback-Leibler divergence” method (Table 7) with the indicators function from the phyloregion library [84]. Occurrences for each species were obtained from the Global Biodiversity Information Facility (GBIF)[85], with anomalous data, country centroids, and museum or research center records filtered using the CoordinateCleaner 3.0.1 library [86].
In the analysis, one occurrence per climatic grid (~1km²) was filtered. The 19 bioclimatic variables and 9 edaphic variables (Table 1) were used, removing those with a correlation coefficient >0.75, prioritizing the most influential according to a principal component analysis (Table 8). Niche overlap was calculated using Schoener’s D index [87] with the ecospat 4.0.0 library [88]. The significance of the results was assessed through a niche equivalence test, based on the D index, comparing the observed value with a distribution of values obtained from random samples. Comparisons were made between the three species from each bioregion and with those from other bioregions, generating three D values for each zone (north-north, transition-transition, and south-south) and nine between each comparison (north-transition, north-south, and south-transition).
4.4.5. Phylogenetic Diversity
To assess phylogenetic diversity metrics [89], the mega-tree included in the V.PhyloMaker2 library [90] was pruned. Two scales were established for analysis: the first corresponds to the three bioregions resulting from the membership grade model, and the second involves latitudinal bands of 2°. The standardized effect of phylogenetic diversity (PD), mean pairwise distance (MPD), and mean nearest taxon distance (MNTD) was analyzed in both contexts. The observed metrics were compared with a null model that randomizes species composition in the community using the Picante 1.8.2 library [91]. Positive and significant values indicate phylogenetic overdispersion, while negative and significant values indicate phylogenetic clustering. The main distinction between MPD and MNTD lies in the fact that the analysis encompasses deeper levels of the tree (orders and families) and towards the branch tips (genera and species), respectively [41].
5. Conclusions
This study provided a comprehensive assessment of how macro-environmental variables function as filters in the differentiation of communities in bogs along the Andes (15°S - 41°S). The findings indicate a low total beta diversity across this gradient, primarily influenced by dispersal limitations and macro-environmental conditions. Three distinct bioregions were identified based on taxonomic diversity, corresponding to the macroclimates of Chile and the phytogeographic districts of the high Andean province of southern South America.
Notably, species at the extremes of the north-south gradient exhibited significant differences in their climatic niches, with a broader niche width in the transition zone. Phylogenetic metric analyses indicate clustering between rainfall regimes in the arid transition zone, reflecting phylogenetic conservatism in niche preference.
In conclusion, this study validates the proposed hypothesis by identifying a clear separation of communities in the transition of rainfall regimes. However, it adds temperature variation as an influential factor in community formation. Significantly, macro-environmental conditions exert a considerable effect on the biodiversity of azonal flora in the Southern Andes of South America, playing a critical role in shaping these unique communities.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: Biplot of environmental variables in Principal Components Analysis on which niche overlap was performed; Figure S2: Principal Components Analysis. In green, the environmental niche of the first species mentioned in the title of each box; in red, the environmental niche of the second species mentioned in the title of each box; and in blue, the environmental niche shared by both species. The red arrow indicates the movement in the environmental space of the niche centroids for each species. The shaded areas correspond to the highest concentration of species occurrences, and the solid and dashed lines represent 100% and 75% of the total climatic envelope of the studied gradient, respectively. The species codes (e.g., N1) are according to Table 7. Table S1: Complete list of high Andean bogs flora of southern South America used in this study.
Author Contributions
Conceptualization, Felipe Figueroa-Ponce and Luis Felipe Hinojosa; Data curation, Felipe Figueroa-Ponce; Formal analysis, Felipe Figueroa-Ponce; Funding acquisition, Luis Felipe Hinojosa; Investigation, Felipe Figueroa-Ponce and Luis Felipe Hinojosa; Methodology, Felipe Figueroa-Ponce and Luis Felipe Hinojosa; Project administration, Felipe Figueroa-Ponce; Resources, Luis Felipe Hinojosa; Software, Felipe Figueroa-Ponce; Supervision, Luis Felipe Hinojosa; Validation, Felipe Figueroa-Ponce and Luis Felipe Hinojosa; Visualization, Felipe Figueroa-Ponce and Luis Felipe Hinojosa; Writing – original draft, Felipe Figueroa-Ponce; Writing – review & editing, Felipe Figueroa-Ponce and Luis Felipe Hinojosa. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Fondecyt 1221214.
Data Availability Statement
The data presented in this study are available on GBIF (https://www.gbif.org/), and CHELSA (https://chelsa-climate.org/downloads/).
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hoorn, C.; Wesselingh, F.P.; ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartín, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia Through Time: Andean Uplift, Climate Change, Landscape Evolution, and Biodiversity. Science 2010, 330, 927-931. [CrossRef]
- Simpson, B.B. An Historical Phytogeography of the High Andean Flora. Rev Chil Hist Nat 1983, 56, 109-122.
- Garreaud, R.D. The Andes climate and weather. Adv Geosci 2009, 22, 3-11. [CrossRef]
- Weir, J.T.; Price, M. Andean uplift promotes lowland speciation through vicariance and dispersal in Dendrocincla woodcreepers. Mol Ecol 2011, 20, 4550-4563. [CrossRef]
- Dibán, M.J.; Hinojosa, L.F. Testing the Tropical Niche Conservatism Hypothesis: Climatic Niche Evolution of Escallonia Mutis ex L. F. (Escalloniaceae). Plants 2024, 13, 133. [CrossRef]
- Graham, A.; Gregory-Wodzicki, K.M.; Wright, K.L. Studies in Neotropical Paleobotany. XV. A Mio-Pliocene palynoflora from the Eastern Cordillera, Bolivia: Implications for the uplift history of the Central Andes. Am J Bot 2001, 88, 1545-1557. [CrossRef]
- Pérez-Escobar, O.A.; Zizka, A.; Bermúdez, M.A.; Meseguer, A.S.; Condamine, F.L.; Hoorn, C.; Hooghiemstra, H.; Pu, Y.; Bogarín, D.; Boschman, L.M.; et al. The Andes through time: Evolution and distribution of Andean floras. Trends Plant Sci 2022, 27, 364-378. [CrossRef]
- Arroyo, M.T.K.; Cavieres, L. High-Elevation Andean Ecosystems. In Encyclopedia of Biodiversity; 2013, pp. 96-110.
- Arroyo, M.T.K.; Squeo, F.A.; Armesto, J.J.; Villagran, C. Effects of Aridity on Plant Diversity in the Northern Chilean Andes: Results of a Natural Experiment. Annals of the Missouri Botanical Garden 1988, 75, 55-78. [CrossRef]
- Villagrán, C.; Arroyo, M.T.K.; Marticorena, C. Effectos de la desertización en la distribución de la flora andina de Chile. Rev Chil Hist Nat 1983, 56, 137-157.
- Martínez-Carretero, E. La puna Argentina: Delimitación general y división en distritos florísticos. Bol Soc Argent Bot 1995, 31, 27-40.
- Alatalo, J.M.; Jägerbrand, A.K.; Molau, U. Impacts of different climate change regimes and extreme climatic events on an alpine meadow community. Sci Rep 2016, 6, 21720. [CrossRef]
- Scherrer, D.; Körner, C. Topographically controlled thermal-habitat differentiation buffers alpine plant diversity against climate warming. J Biogeogr 2011, 38, 406-416. [CrossRef]
- Squeo, F.A.; Warner, B.G.; Aravena, R.; Espinoza, D. Bofedales: High altitude peatlands of the central Andes. Rev Chil Hist Nat 2006, 79, 245-255. [CrossRef]
- Ruthsatz, B. Vegetation and ecology of the high Andean peatlands of Bolivia. Phytocoenologia 2012, 42, 133-179. [CrossRef]
- Cleef, A.M. The vegetation of the páramos of the Colombian Cordillera Oriental. Mededelingen Bot Museum Herb Rijksuniversiteit Utrecht 1981, 481, 1-320.
- Ruthsatz, B.; Villagran, C. Vegetation pattern and soil nu trien ts of a Magellanic moorland on the Cordillera de Piuchué, Chiloé Island, Chile. Rev Hist Chil Nat 1991, 64, 461-478.
- Badano, E.I.; G. Jones, C.; A. Cavieres, L.; P. Wright, J. Assessing impacts of ecosystem engineers on community organization: A general approach illustrated by effects of a high-Andean cushion plant. Oikos 2006, 115, 369-385. [CrossRef]
- Izquierdo, A.E.; Aragón, M.R.; Navarro, C.J.; Casagranda, M.E. Humedales de la Puna: Principales proveedores de servicios ecosistémicos de la región. In Puna Argentina: Naturaleza y cultura, 1st ed.; Grau, H.R., Babot, M.J., Izquierdo, A.E., Grau, A., Eds.; Fundación Miguel Lillo: Tucumán, Argentina, 2018; pp. 96–111.
- Rodriguez, R.; Marticorena, C.; Alarcón, D.; Baeza, C.; Cavieres, L.; Finot, V.L.; Fuentes, N.; Kiessling, A.; Mihoc, M.; Pauchard, A.; et al. Catálogo de las plantas vasculares de Chile. Gayana Bot 2018, 75, 1-430. [CrossRef]
- Carilla, J.; Grau, A.; Cuello, A.S. Vegetación de la Puna Argentina. In Puna Argentina: Naturaleza y cultura, 1st ed.; Grau, H.R., Babot, M.J., Izquierdo, A.E., Grau, A., Eds.; Fundación Miguel Lillo: Tucumán, Argentina, 2018; pp. 146–156.
- Maldonado-Fonkén, M.S. An introduction to the bofedales of the Peruvian High Andes. Mires and Peat 2014, 15, 1-13.
- Polk, M.H.; Young, K.R.; Cano, A.; León, B. Vegetation of Andean wetlands (bofedales) in Huascarán National Park, Peru. Mires and Peat 2019, 24, 1-26. [CrossRef]
- Izquierdo, A.E.; Carilla, J.; Nieto, C.; Osinaga Acosta, O.; Martin, E.; Grau, H.R.; Reynaga, M.C. Multi-taxon patterns from high Andean peatlands: Assessing climatic and landscape variables. Community Ecology 2020, 21, 317-332. [CrossRef]
- Ruthsatz, B. Die Hartpolstermoore der Hochanden und ihre Artenvielfalt, Ber.Reinh.-Tüxen-Ges. 2000, 12, 351-371.
- Ruthsatz, B. Hartpolstermoore der Hochanden NW-Argentiniens als Indikatoren für Klimagradienten. Mitteilungen der Arbeitsgemeinschaft für Geobotanik Schleswig-Holstein 2008, 65, 209-238.
- Méndez, E. La vegetación de los Altos Andes II: Las Vegas del flanco oriental del Cordón del Plata (Mendoza, Argentina). Bol Soc Argent Bot 2007, 42, 273-294.
- Casagranda, M.E.; Izquierdo, A.E. Modeling the potential distribution of floristic assemblages of high Andean wetlands dominated by Juncaceae and Cyperaceae in the Argentine Puna. Vegetation Classification and Survey 2023, 4, 47-58. [CrossRef]
- Ruthsatz, B. Flora und ökologische Bedingungen hochandiner Moore Chiles zwischen 18°00’ (Arica) und 40°30’ (Osorno) suPdl. Br. Phytocoenologia 1993, 23, 157-199.
- Ruthsatz, B.; Schittek, K.; Backes, B. The vegetation of cushion peatlands in the Argentine Andes and changes in their floristic composition across a latitudinal gradient from 39°S to 22°S. Phytocoenologia 2020, 50, 249-278. [CrossRef]
- Woodward, F.I.; Diament, A.D. Functional Approaches to Predicting the Ecological Effects of Global Change. Funct Ecol 1991, 5, 202-212. [CrossRef]
- Weiher, E.; Keddy, P.A. Assembly Rules, Null Models, and Trait Dispersion: New Questions from Old Patterns. Oikos 1995, 74, 159-164. [CrossRef]
- Zobel, M. The relative of species pools in determining plant species richness: An alternative explanation of species coexistence? Trends Ecol Evol 1997, 12, 266-269. [CrossRef]
- Fang, J.; Wang, X.; Tang, Z. Local and regional processes control species richness of plant communities: The species pool hypothesis. Biodiv Sci 2009, 17, 605-612. [CrossRef]
- Whittaker, R.H. Evolution and Measurement of Species Diversity. Taxon 1972, 21, 213-251. [CrossRef]
- Whittaker, R.H. Vegetation of the Siskiyou Mountains, Oregon and California. Ecol Monogr 1960, 30, 279-338. [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Global Ecol Biogeogr 2010, 19, 134-143. [CrossRef]
- Legendre, P. Interpreting the replacement and richness difference components of beta diversity. Global Ecol Biogeogr 2014, 23, 1324-1334. [CrossRef]
- Dobrovolski, R.; Melo, A.S.; Cassemiro, F.A.S.; Diniz-Filho, J.A.F. Climatic history and dispersal ability explain the relative importance of turnover and nestedness components of beta diversity. Global Ecol Biogeogr 2011, 21, 191-197.
- Hu, D.; Jiang, L.; Hou, Z.; Zhang, J.; Wang, H.; Lv, G. Environmental filtration and dispersal limitation explain different aspects of beta diversity in desert plant communities. Global Ecol Conserv 2022, 33, e01956. [CrossRef]
- Webb, C.O. Exploring the Phylogenetic Structure of Ecological Communities: An Example for Rain Forest Trees. The American Naturalist 2000, 156, 145-155. [CrossRef]
- Kellar, P.R.; Ahrendsen, D.L.; Aust, S.K.; Jones, A.R.; Pires, J.C. Biodiversity comparison among phylogenetic diversity metrics and between three North American prairies. App Plant Sci 2015, 3, 1400108. [CrossRef]
- Horner-Devine, M.C.; Bohannan, B.J.M. Phylogenetic Clustering and Overdispersion in Bacterial Communities. Ecology 2006, 87, S100-S108. [CrossRef]
- Cavender-Bares, J.; Ackerly, D.D.; Baum, D.A.; Bazzaz, F.A. Phylogenetic Overdispersion in Floridian Oak Communities. The American Naturalist 2004, 163, 823-843. [CrossRef]
- Verdú, M.; Pausas, J.G. Fire drives phylogenetic clustering in Mediterranean Basin woody plant communities. J Ecol 2007, 95, 1316-1323. [CrossRef]
- Jones, M.M.; Tuomisto, H.; Borcard, D.; Legendre, P.; Clark, D.B.; Olivas, P.C. Explaining variation in tropical plant community composition: Influence of environmental and spatial data quality. Oecologia 2008, 155, 593-604. [CrossRef]
- Blundo, C.; González-Espinosa, M.; Malizia, L.R. Relative contribution of niche and neutral processes on tree species turnover across scales in seasonal forests of NW Argentina. Plant Ecol 2016, 217, 359-368. [CrossRef]
- Zheng, J.; Arif, M.; He, X.; Ding, D.; Zhang, S.; Ni, X.; Li, C. Plant community assembly is jointly shaped by environmental and dispersal filtering along elevation gradients in a semiarid area, China. Front Plant Sci 2022, 13. [CrossRef]
- Ross, A.C.; Mendoza, M.M.; Drenkhan, F.; Montoya, N.; Baiker, J.R.; Mackay, J.D.; Hannah, D.M.; Buytaert, W. Seasonal water storage and release dynamics of bofedal wetlands in the Central Andes. Hydrol Processes 2023, 37, e14940. [CrossRef]
- Guo, Y.; Xiang, W.; Wang, B.; Li, D.; Mallik, A.U.; Chen, H.Y.H.; Huang, F.; Ding, T.; Wen, S.; Lu, S.; et al. Partitioning beta diversity in a tropical karst seasonal rainforest in Southern China. Sci Rep 2018, 8, 17408. [CrossRef]
- Villagrán, C.; Armesto, J.J.; Hinojosa, L.F.; Cuvertino, J.; Pérez, C.; Medina, C. El enigmático origen del bosque relicto de Fray Jorge. In Historia Natural del Parque Nacional Bosque Fray Jorge, 1st ed.; Squeo, F.A., Gutiérrez, J.R., Hernández, I.R., Eds.; Ediciones Universidad de La Serena, La Serena, Chile, 2004; Volume 1, pp. 3-43.
- Cheng, D.; Zhu, Q.; Huang, J.; Wu, Q.; Yang, L. A local cores-based hierarchical clustering algorithm for data sets with complex structures. Neural Computing and Applications 2019, 31, 8051-8068. [CrossRef]
- Almeida, J.A.S.; Barbosa, L.M.S.; Pais, A.A.C.C.; Formosinho, S.J. Improving hierarchical cluster analysis: A new method with outlier detection and automatic clustering. Chemom Intell Lab Syst 2007, 87, 208-217. [CrossRef]
- Carta, A.; Peruzzi, L.; Ramírez-Barahona, S. A global phylogenetic regionalization of vascular plants reveals a deep split between Gondwanan and Laurasian biotas. New Phytol 2022, 233, 1494-1504. [CrossRef]
- Li, Q.; Sun, H.; Boufford, D.E.; Bartholomew, B.; Fritsch, P.W.; Chen, J.; Deng, T.; Ree, R.H. Grade of Membership models reveal geographical and environmental correlates of floristic structure in a temperate biodiversity hotspot. New Phytol 2021, 232, 1424-1435. [CrossRef]
- Biganzoli, F.; Oyarzabal, M.; Teillier, S.; Zuloaga, F.O. Fitogeografía de la provincia Altoandina del Cono Sur de Sudamérica. Darwiniana 2022, 10, 537-574. [CrossRef]
- Moreira-Muñoz, A. Plant Geography of Chile; Springer: Santiago, Chile, 2011; pp. 0-351.
- Lörch, M.; Mutke, J.; Weigend, M.; Luebert, F. Historical biogeography and climatic differentiation of the Fulcaldea-Archidasyphyllum-Arnaldoa clade of Barnadesioideae (Asteraceae) suggest a Miocene, aridity-mediated Andean disjunction associated with climatic niche shifts. Global Planet Change 2021, 201, 103495. [CrossRef]
- Luebert, F.; Lörch, M.; Acuña, R.; Mello-Silva, R.; Weigend, M.; Mutke, J. Clade-Specific Biogeographic History and Climatic Niche Shifts of the Southern Andean-Southern Brazilian Disjunction in Plants. In Neotropical Diversification: Patterns and Processes, Rull, V., Carnaval, A.C., Eds.; Springer International Publishing: Cham, 2020; pp. 661-682.
- Scherson, R.A.; Thornhill, A.H.; Urbina-Casanova, R.; Freyman, W.A.; Pliscoff, P.A.; Mishler, B.D. Spatial phylogenetics of the vascular flora of Chile. Mol Phylogenet Evol 2017, 112, 88-95. [CrossRef]
- Qian, H.; Zhang, J.; Jiang, M. Global patterns of taxonomic and phylogenetic diversity of flowering plants: Biodiversity hotspots and coldspots. Plant Diversity 2023, 45, 265-271. [CrossRef]
- Qian, H.; Jin, Y. An updated megaphylogeny of plants, a tool for generating plant phylogenies and an analysis of phylogenetic community structure. J Plant Ecol 2016, 9, 233-239. [CrossRef]
- Brožová, V.; Proćków, J.; Záveská Drábková, L. Toward finally unraveling the phylogenetic relationships of Juncaceae with respect to another cyperid family, Cyperaceae. Mol Phylogenet Evol 2022, 177, 107588. [CrossRef]
- Elliott, T.L.; Larridon, I.; Barrett, R.L.; Bruhl, J.J.; Costa, S.M.; Escudero, M.; Hipp, A.L.; Jiménez-Mejías, P.; Kirschner, J.; Luceño, M.; et al. Addressing inconsistencies in Cyperaceae and Juncaceae taxonomy: Comment on Brožová et al. (2022). Mol Phylogenet Evol 2023, 179, 107665. [CrossRef]
- Jr, E.G.L.; Rosindell, J.; Etienne, R.S. Unified neutral theory of biodiversity and biogeography. Scholarpedia 2010, 5, 8822. [CrossRef]
- Chase, J.M. Community assembly: When should history matter? Oecologia 2003, 136, 489-498. [CrossRef]
- De Bello, F.; Lavergne, S.; Meynard, C.N.; Lepš, J.; Thuiller, W. The partitioning of diversity: Showing Theseus a way out of the labyrinth. J Veg Sci 2010, 21, 992-1000. [CrossRef]
- Wiens, J.J. Speciation and ecology revisited: Phylogenetic niche conservatism and the origin of species. Evolution 2004, 58, 193-197. [CrossRef]
- Luebert, F.; Weigend, M. Phylogenetic insights into Andean plant diversification. Front Ecol Evol 2014, 2. [CrossRef]
- Zuloaga, F.O.; Belgrano, M.J.; Zanotti, C.A. Actualización del Catálogo de las Plantas Vasculares del Cono Sur. Darwiniana 2019, 7, 208-278. [CrossRef]
- Jørgensen, P.M.; Nee, M.; Beck, S.G.; Arrázola, S.; Saldias, M.; Hirth, S.; Swift, V.; Penagos, J.C.; Romero, C. Catálogo de las plantas vasculares de Bolivia; Missouri Botanical Garden Press: 2014.
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Data from: Climatologies at high resolution for the earth’s land surface areas. EnviDat 2018. [CrossRef]
- Karger, D.N.; Conrad, O.; Böhner, J.; Kawohl, T.; Kreft, H.; Soria-Auza, R.W.; Zimmermann, N.E.; Linder, H.P.; Kessler, M. Climatologies at high resolution for the earth’s land surface areas. Scientific Data 2017, 4, 170122. [CrossRef]
- Poggio, L.; De Sousa, L.M.; Batjes, N.H.; Heuvelink, G.B.M.; Kempen, B.; Ribeiro, E.; Rossiter, D. SoilGrids 2.0: Producing soil information for the globe with quantified spatial uncertainty. SOIL 2021, 7, 217-240. [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria, 2023. https://www.R-project.org/.
- Hijmans J., R.; Karney, C.; Williams, E.; Vennes, C. geosphere: Spherical Trigonometry. R package version 1.5-18, 2022.
- Dray, S.; Bauman, D.; Blanchet, G.; Borcard, D.; Clappe, S.; Guenard, G.; Jombart, T.; Larocque, G.; Legendre, P.; Madi, N.; et al. Adespatial: Multivariate Multiscale Spatial Analysis. R package version 0.3-23, 2023.
- Legendre, P.; Fortin, M.-J. Comparison of the Mantel test and alternative approaches for detecting complex multivariate relationships in the spatial analysis of genetic data. Mol Ecol Resour 2010, 10, 831-844. [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’hara, R.B.; Simpson, G.L.; Solymos, P. vegan: Community Ecology Package. R package version 2.5-6; 2019.
- Legendre, P.; De Cáceres, M. Beta diversity as the variance of community data: Dissimilarity coefficients and partitioning. Ecol Lett 2013, 16, 951-963. [CrossRef]
- Sokal, R.R.; Rohlf, F.J. The Comparison of Dendrograms by Objective Methods. Taxon 1962, 11, 33-40. [CrossRef]
- Salvador, S.; Chan, P. Determining the number of clusters/segments in hierarchical clustering/segmentation algorithms. In Proceedings of the 16th IEEE International Conference on Tools with Artificial Intelligence, 2004; pp. 576-584.
- Vavrek, M.J. A comparison of clustering methods for biogeography with fossil datasets. PeerJ 2016, 4, e1720. [CrossRef]
- Daru, B.H.; Karunarathne, P.; Schliep, K. phyloregion: R package for biogeographical regionalization and macroecology. Methods Ecol Evol 2020, 11, 1483-1491. [CrossRef]
- GBIF.org. GBIF Home Page, 2023.
- Zizka, A.; Silvestro, D.; Andermann, T.; Azevedo, J.; Duarte Ritter, C.; Edler, D.; Farooq, H.; Herdean, A.; Ariza, M.; Scharn, R.; et al. CoordinateCleaner: Standardized cleaning of occurrence records from biological collection databases. Methods Ecol Evol 2019, 10, 744-751. [CrossRef]
- Schoener, T.W. The Anolis Lizards of Bimini: Resource Partitioning in a Complex Fauna. Ecology 1968, 49, 704-726. [CrossRef]
- Broennimann, O.; Di Cola, V.; Petitpierre, B.; Breiner, F.; Scherrer, D.; D’Amen, M.; Randin, C.; Engler, R.; Hordijk, W.; Mod, H.; et al. ecospat: Spatial Ecology Miscellaneous Methods. R package version 4.0.0, 2023.
- Faith, D.P. Conservation evaluation and phylogenetic diversity. Biol Conserv 1992, 61, 1-10. [CrossRef]
- Jin, Y.; Qian, H. V.PhyloMaker2: An updated and enlarged R package that can generate very large phylogenies for vascular plants. Plant Diversity 2022, 44, 335-339. [CrossRef]
- Kembel, S.W.; Cowan, P.D.; Helmus, M.R.; Cornwell, W.K.; Morlon, H.; Ackerly, D.D.; Blomberg, S.P.; Webb, C.O. Picante: R tools for integrating phylogenies and ecology. Bioinformatics 2010, 26, 1463-1464. [CrossRef]
Figure 1.
Map of the precipitation regimes in southern South America represented by Precipitation of Warmer Quarter (red lines) and Precipitation of Coldest Quarter (blue lines). Dotted lines divide the Puna (PU) and southern Andean steppe (SA) ecosystems.
Figure 1.
Map of the precipitation regimes in southern South America represented by Precipitation of Warmer Quarter (red lines) and Precipitation of Coldest Quarter (blue lines). Dotted lines divide the Puna (PU) and southern Andean steppe (SA) ecosystems.
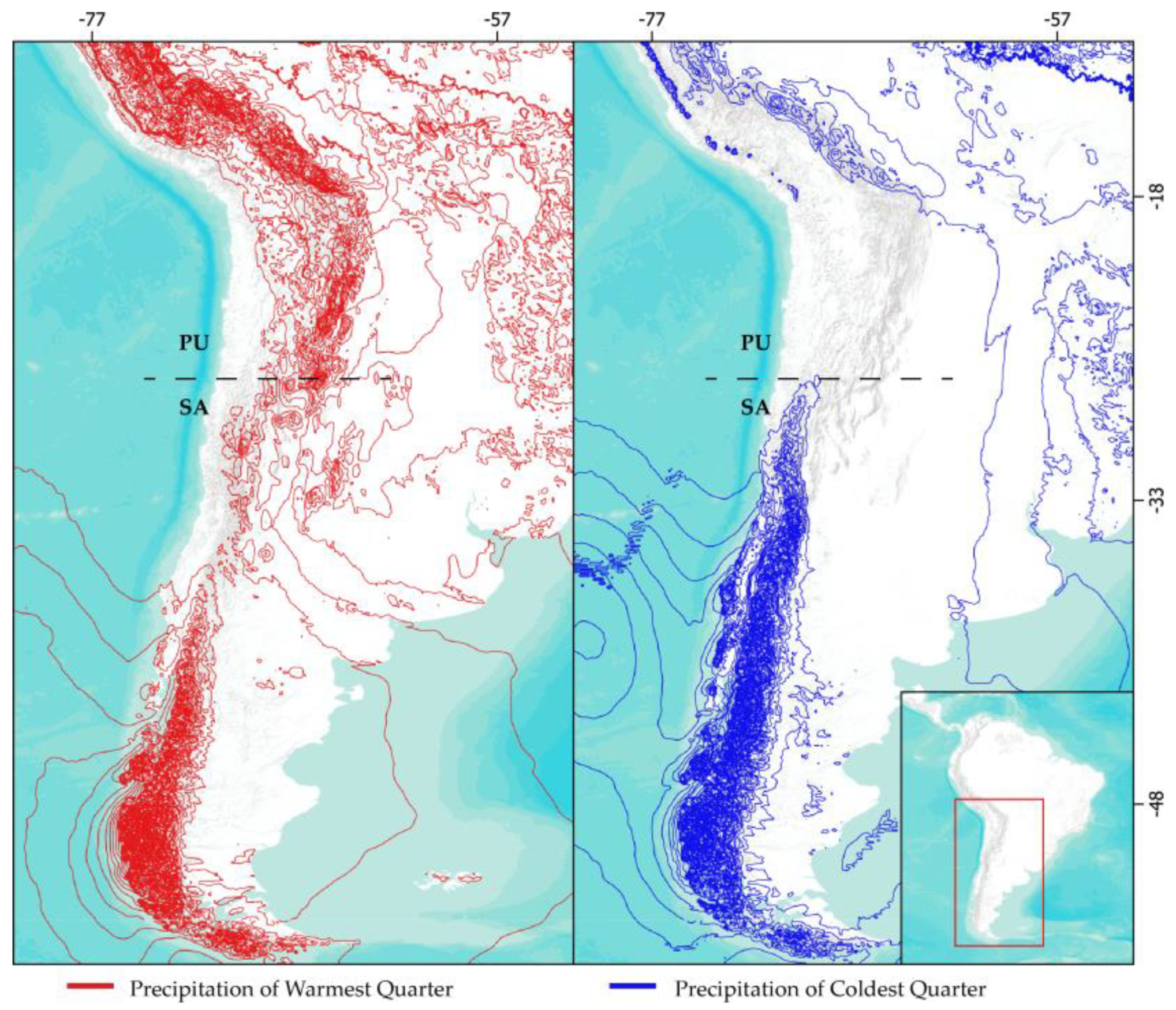
Figure 2.
Venn diagram of the variance partitioning analysis between environmental and spatial factors (a) and between each environmental component with spatial factors (b). The values inside each circle/oval indicate the individual contribution of that variable. The values in the small white circles indicate the contribution of the variable considering a synergistic effect with the others. The values in the small circles colored according to the intersection correspond to the contribution of the interaction of those two variables, considering the synergistic effect with the others. ENV = environment; SPA = spatial; T = temperature; P = precipitation; S = soil.
Figure 2.
Venn diagram of the variance partitioning analysis between environmental and spatial factors (a) and between each environmental component with spatial factors (b). The values inside each circle/oval indicate the individual contribution of that variable. The values in the small white circles indicate the contribution of the variable considering a synergistic effect with the others. The values in the small circles colored according to the intersection correspond to the contribution of the interaction of those two variables, considering the synergistic effect with the others. ENV = environment; SPA = spatial; T = temperature; P = precipitation; S = soil.
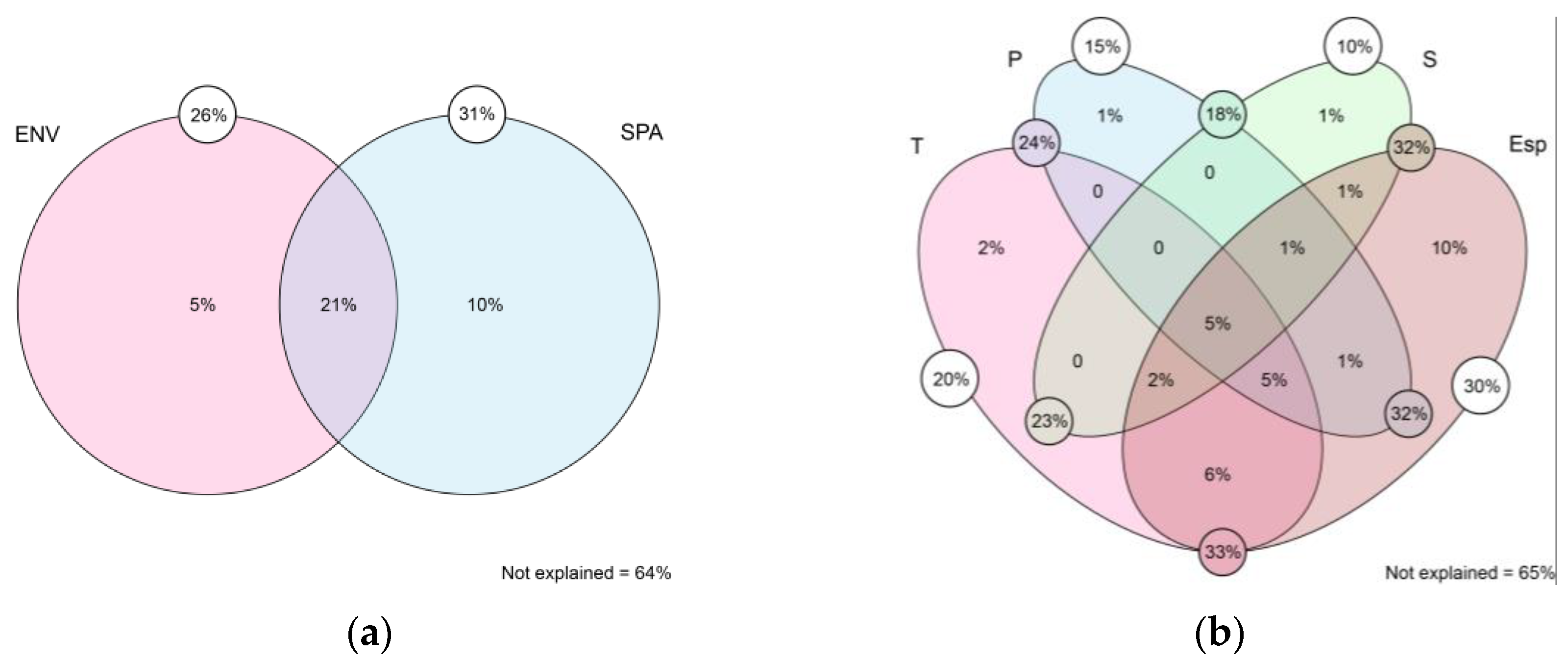
Figure 3.
The first row (a, b, c) shows the relationships between beta diversity and its components with geographic distance. The second row (d, e, f) shows the relationships between beta diversity and its components with environmental distance.
Figure 3.
The first row (a, b, c) shows the relationships between beta diversity and its components with geographic distance. The second row (d, e, f) shows the relationships between beta diversity and its components with environmental distance.
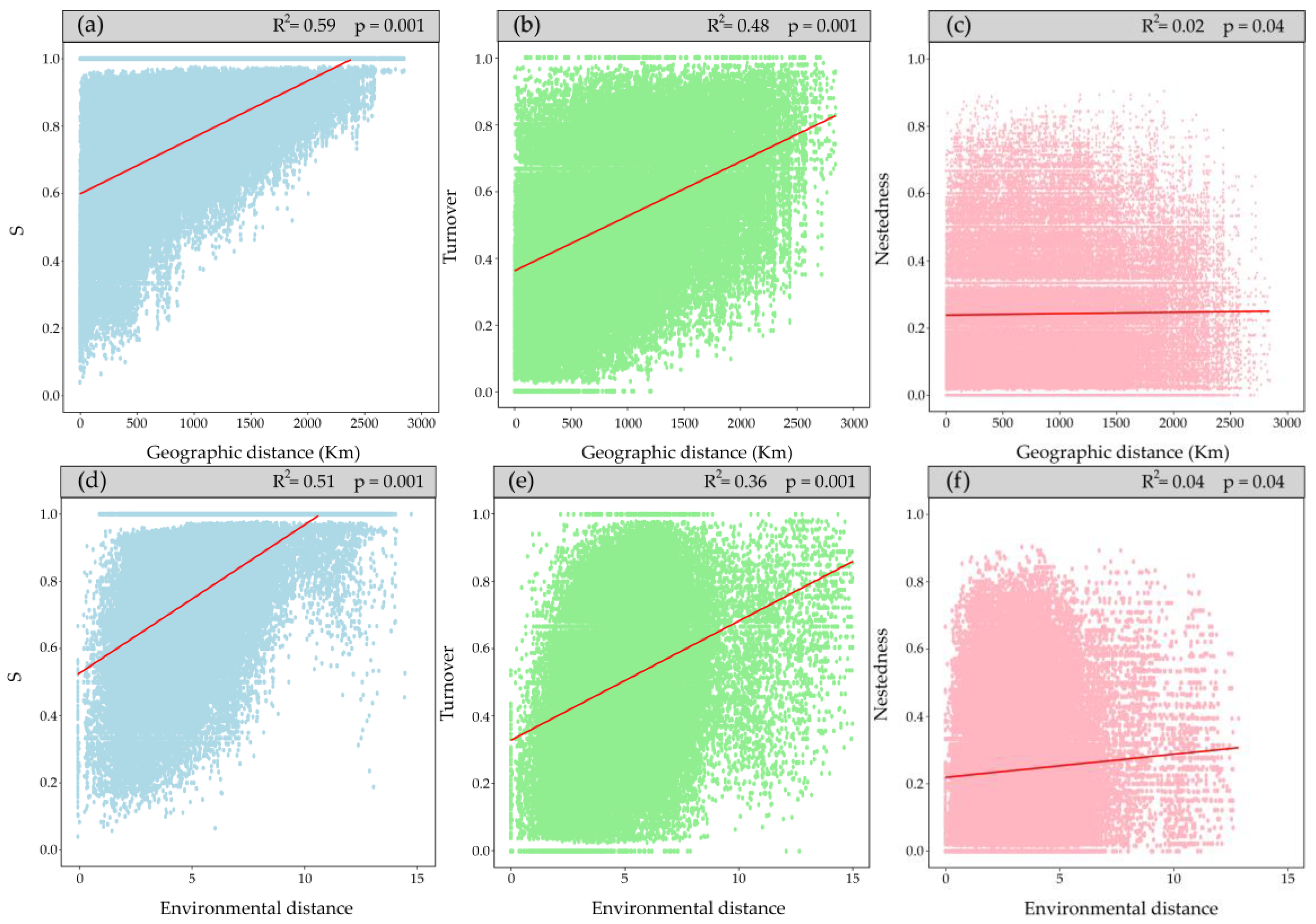
Figure 4.
Dendrogram of floristic affinities based on the Sorensen dissimilarity index with its projection onto geographic space in (a). Regionalization by membership grade model in (b). The dotted red lines correspond to the operational macrozones defined in this study.
Figure 4.
Dendrogram of floristic affinities based on the Sorensen dissimilarity index with its projection onto geographic space in (a). Regionalization by membership grade model in (b). The dotted red lines correspond to the operational macrozones defined in this study.
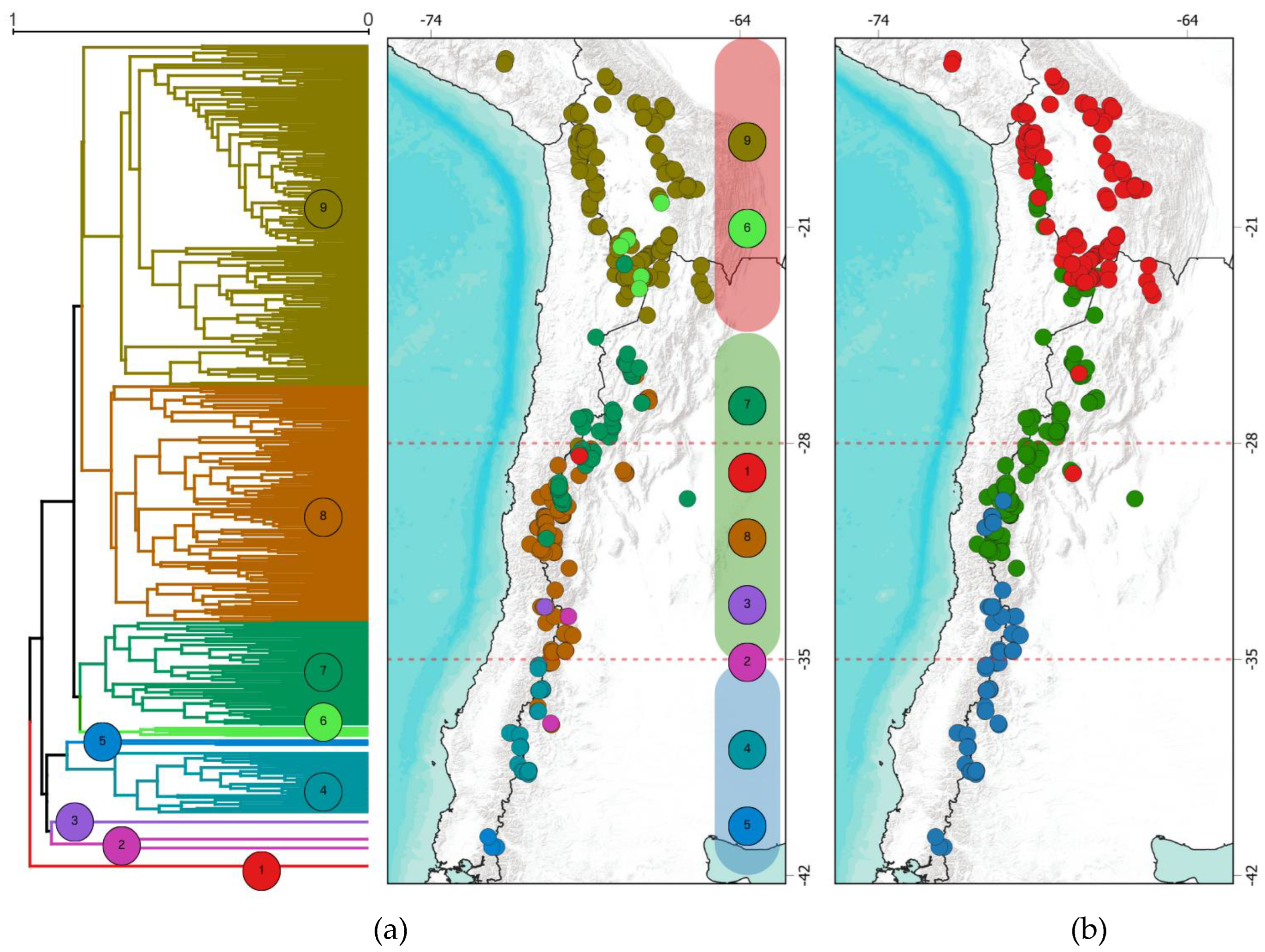
Figure 5.
Average “D” Overlap Index as a percentage for species from each bioregion. “N” represents the average overlap among species within the northern bioregion; “T” indicates the average overlap among species within the transition bioregion; “S” refers to the average overlap among species within the southern bioregion. “N-S” denotes the average overlap between species from the north and south bioregions; “N-T” refers to the average between species from the northern and transition bioregions; “S-T” shows the average between species from the southern and transition bioregions.
Figure 5.
Average “D” Overlap Index as a percentage for species from each bioregion. “N” represents the average overlap among species within the northern bioregion; “T” indicates the average overlap among species within the transition bioregion; “S” refers to the average overlap among species within the southern bioregion. “N-S” denotes the average overlap between species from the north and south bioregions; “N-T” refers to the average between species from the northern and transition bioregions; “S-T” shows the average between species from the southern and transition bioregions.
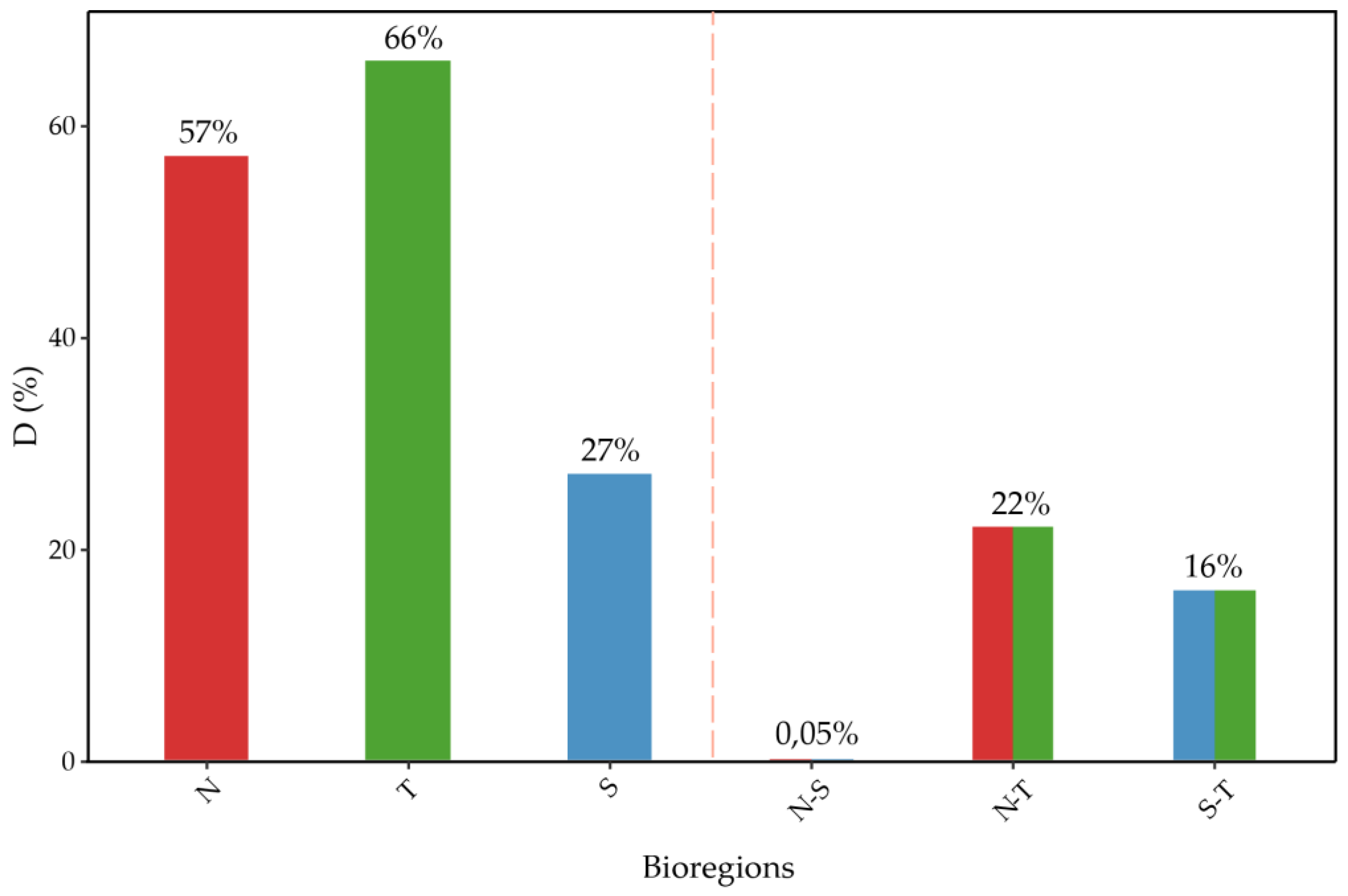
Figure 6.
Observed Phylogenetic Diversity (PD) for each bioregion (blue) and expected PD according to the null model (gray).
Figure 6.
Observed Phylogenetic Diversity (PD) for each bioregion (blue) and expected PD according to the null model (gray).
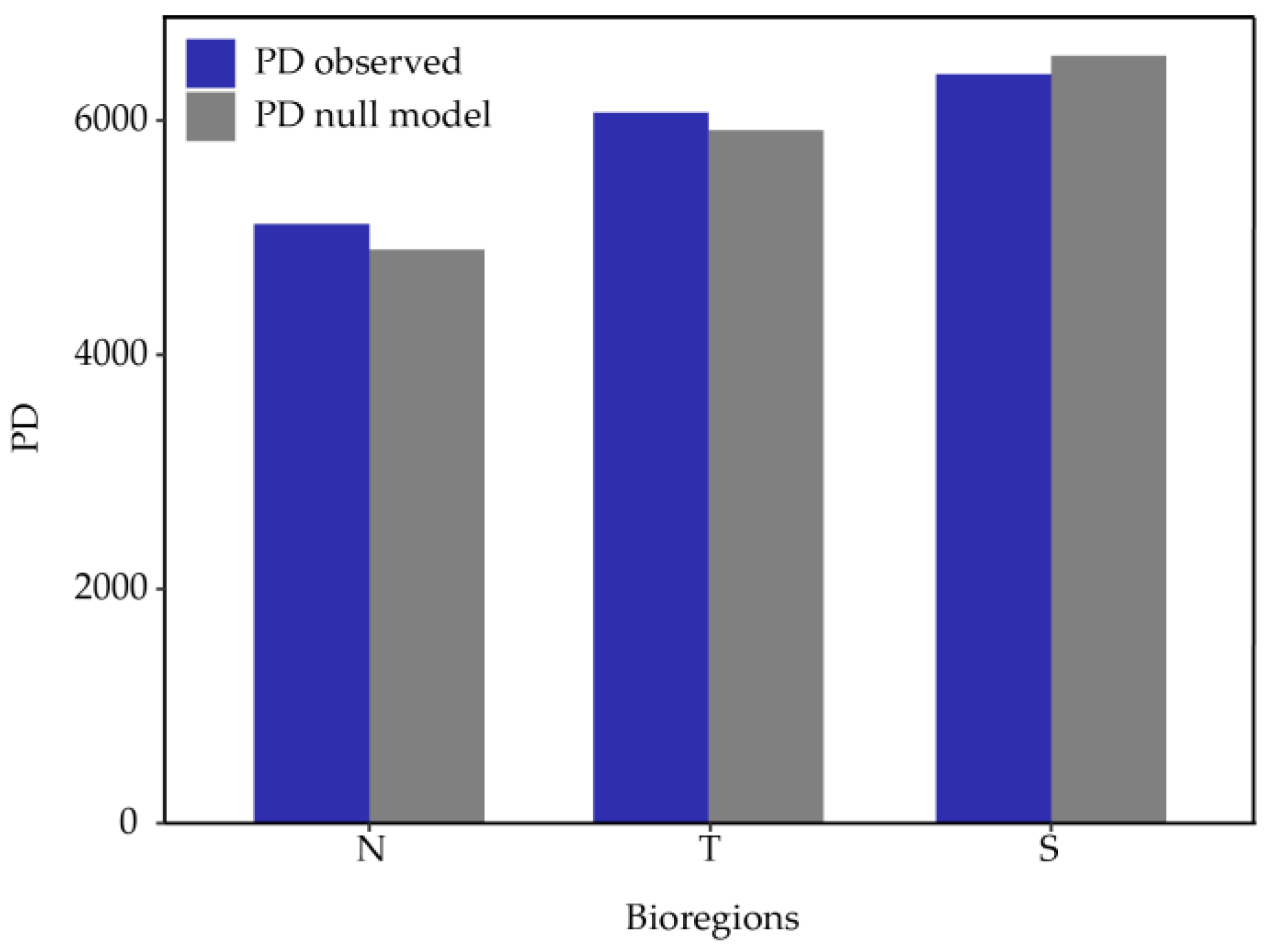
Figure 7.
Observed Phylogenetic Diversity (PD) for each 2° latitudinal band (blue) and the expected PD according to the null model (gray).
Figure 7.
Observed Phylogenetic Diversity (PD) for each 2° latitudinal band (blue) and the expected PD according to the null model (gray).

Figure 8.
Distribution map of the 420 bogs in southern South America with the macroclimatic regions defined for this study: North (N), Transition (T), and South (S).
Figure 8.
Distribution map of the 420 bogs in southern South America with the macroclimatic regions defined for this study: North (N), Transition (T), and South (S).
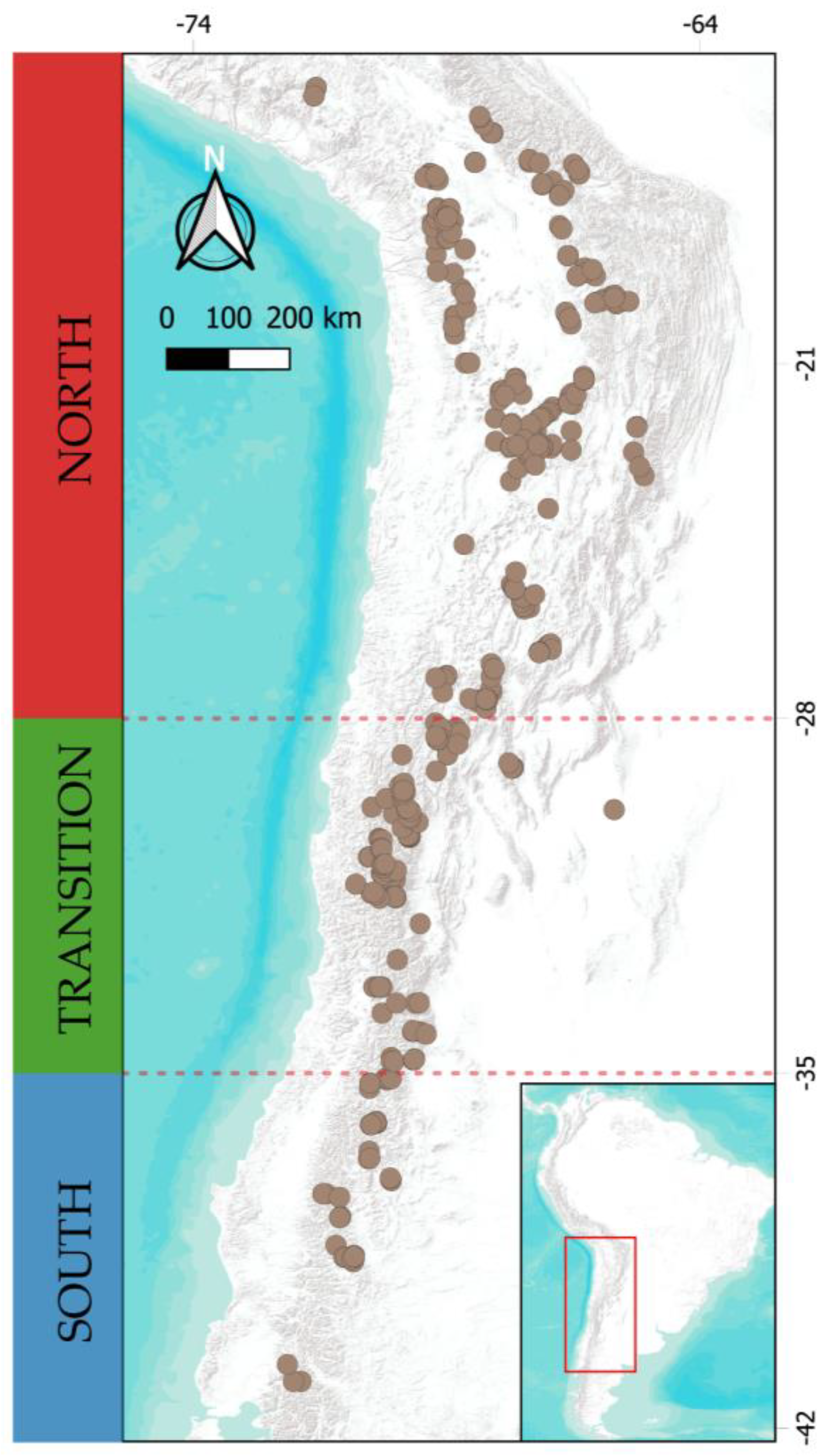

Table 1.
Standardized effects of PD, MPD, and MNTD compared to the null model (z-score) for each bioregion with statistical significance (α).
Table 1.
Standardized effects of PD, MPD, and MNTD compared to the null model (z-score) for each bioregion with statistical significance (α).
| Bioregion | PD z-score | αPD | MPD z-score | αMPD | MNTD z-score | αMNTD |
|---|---|---|---|---|---|---|
| N | 0.771 | 0.773 | 1.899 | 0.979 | 0.462 | 0.678 |
| T | 0.523 | 0.683 | 1.430 | 0.949 | 0.275 | 0.591 |
| S | -0.596 | 0.277 | -1.310 | 0.142 | -0.503 | 0.305 |
Table 2.
Standardized PD, MPD, and MNTD effects relative to the null model (z-score) for each 2° latitudinal band with statistical significance (α).
Table 2.
Standardized PD, MPD, and MNTD effects relative to the null model (z-score) for each 2° latitudinal band with statistical significance (α).
| Band | PD z-score | αPD | MPD z-score | αMPD | MNTD z-score | αMNTD |
| 15-16 | 0.386 | 0.658 | 1.507 | 0.859 | 0.343 | 0.659 |
| 17-18 | 1.041 | 0.836 | 1.740 | 0.931 | 1.291 | 0.879 |
| 18-19 | 1.738 | 0.953 | 2.141 | 0.982 | 1.681 | 0.943 |
| 21-22 | 0.189 | 0.608 | 1.866 | 0.932 | 0.243 | 0.598 |
| 23-24 | -1.786 | 0.022 | -0.912 | 0.126 | -1.408 | 0.078 |
| 25-26 | -0.783 | 0.231 | -0.452 | 0.371 | -0.546 | 0.304 |
| 27-28 | -0.570 | 0.309 | -0.558 | 0.305 | -0.721 | 0.249 |
| 29-30 | -1.452 | 0.064 | -0.266 | 0.515 | -1.517 | 0.059 |
| 31-32 | -0.904 | 0.186 | -0.805 | 0.179 | -0.292 | 0.408 |
| 33-34 | -0.562 | 0.295 | -0.090 | 0.609 | -0.809 | 0.215 |
| 35-36 | -0.887 | 0.189 | -0.587 | 0.322 | -1.319 | 0.089 |
| 37-38 | -0.344 | 0.376 | -0.736 | 0.265 | 0.232 | 0.594 |
| 40-41 | 0.504 | 0.713 | -0.435 | 0.388 | 0.637 | 0.747 |
Table 3.
Environmental Variables and Their Abbreviations Used in This Study.
| Type | Variable (abbreviation) | |
| Climate | Annual Mean Temperature (Bio1) | Mean Temperature of Coldest Quarter (Bio11) |
| Mean Diurnal Range (Bio2) | Annual Precipitation (Bio12) | |
| Isothermality (Bio3) | Precipitation of Wettest Month (Bio13) | |
| Temperature Seasonality (Bio4) | Precipitation of Driest Month (Bio14) | |
| Max Temperature of Warmest Month (Bio5) | Precipitation Seasonality (Bio15) | |
| Min Temperature of Coldest Month (Bio6) | Precipitation of Wettest Quarter (Bio16) | |
| Temperature Annual Range (Bio7) | Precipitation of Driest Quarter (Bio17) | |
| Mean Temperature of Wettest Quarter (Bio8) | Precipitation of Warmest Quarter (Bio18) | |
| Mean Temperature of Driest Quarter (Bio9) | Precipitation of Coldest Quarter (Bio19) | |
| Mean Temperature of Warmest Quarter (Bio10) | ||
| Edaphic | Soil organic carbon in fine earth (Soc) | Total nitrogen (Nitrogen) |
| Bulk density of the fine earth fraction (Bdod) | Vol. fraction of coarse fragments (> 2 mm) (Cfvo) | |
| pH H2O (Phh2o) | Densidad de carbono orgánico (Ocd) | |
| Silt (Silt) | Sand (Sand) | |
| Clay (Clay) | ||
| Elevación | Digital elevation model (Elev) | |
Table 4.
Environmental variables after correlation analysis and Forward Selection used for Variation Partitioning.
Table 4.
Environmental variables after correlation analysis and Forward Selection used for Variation Partitioning.
| Type | Variables | |||
| Environment | Temperature | Bio2$$Bio7$$Bio9$$Bio10$$Bio11 | ||
| Precipitation | Bio15$$Bio18 | |||
| Edaphic | Bdod$$Phh2o$$Cfvo | |||
| Spatial | Elevation | Elev | ||
| MEM | 4 | 1 | 3 | |
| 5 | 9 | 8 | ||
| 2 | 12 | 51 | ||
| 19 | 17 | 10 | ||
| 32 | 59 | 28 | ||
| 11 | 37 | 6 | ||
| 16 | 40 | 24 | ||
| 23 | 20 | 33 | ||
| 22 | 52 | 42 | ||
| 31 | 36 | |||
Table 5.
Equations Used for Calculating Beta Diversity. In (1), (3), and (4), a = species shared by the compared sites; b = species present exclusively in one site; and c = species present exclusively in the other site. In (2), n = sites in the dissimilarity matrix, created from equation (1); D2hi = √S between each site.
Table 5.
Equations Used for Calculating Beta Diversity. In (1), (3), and (4), a = species shared by the compared sites; b = species present exclusively in one site; and c = species present exclusively in the other site. In (2), n = sites in the dissimilarity matrix, created from equation (1); D2hi = √S between each site.
| Index | Equation | Source | |
| S (Sorensen) | (Chao et al., 2006) | (1) | |
| BDtotal | (Legendre, 2013) | (2) | |
| Turnover(ReplS) | (Legendre, 2014) | (3) | |
| Nestedness(RichDiffS) | (Legendre, 2014) | (4) |
Table 6.
Environmental Variables After Correlation Analysis and Forward Selection Used for Mantel Tests.
Table 6.
Environmental Variables After Correlation Analysis and Forward Selection Used for Mantel Tests.
| Type | S | Turnover | Nestedness |
| Temperature | Bio2 | Bio2 | Bio11 |
| Bio7 | Bio7 | ||
| Bio9 | Bio9 | ||
| Bio10 | Bio10 | ||
| Bio11 | Bio11 | ||
| Precipitation | Bio15 | Bio14 | Bio16 |
| Bio16 | Bio15 | ||
| Bio18 | Bio16 | ||
| Edaphic | Bdod | Bio18 | Silt |
| Phh2o | Bdod | Bdod | |
| Silt | Phh2o | Soc | |
| Sand | Silt | ||
| Cfvo | Cfvo | ||
| Elevation | Elev | Elev |
Table 7.
Selected Species for Niche Overlap Analysis.
| North (N) | Transition (T) | South (S) | |
| 1 | Plantago tubulosa | Deschampsia eminens | Ochetophila nana |
| 2 | Distichia muscoides | Cinnagrostis velutina | Ranunculus peduncularis |
| 3 | Hypochaeris taraxacoides | Eleocharis pseudoalbibracteata | Hordeum comosum |
Table 8.
Environmental variables used in niche overlap analysis after correlation and Forward Selection analysis.
Table 8.
Environmental variables used in niche overlap analysis after correlation and Forward Selection analysis.
| Climate | Edaphic | |
| Bio2 | Bio11 | Bdod |
| Bio3 | Bio12 | Cfvo |
| Bio5 | Bio15 | Clay |
| Bio7 | Bio19 | Nitrogen |
| Bio9 | Silt | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



