
Submitted:
21 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Cerebral ischemic stroke poses a significant global cause of death, with ischemia-reperfusion injury contributing to neuronal cell death and tissue damage. However, significant therapeutic approaches with meaningful treatment effects for patients with ischemic stroke in actual clinical practice are lacking. This study investigated the potential neuroprotective effects of sildenafil, a phosphodiesterase-5 inhibitor, in a global cerebral ischemia model. We explored the impact of sildenafil on GFAP and AQP-4 expression, which are markers associated with astrocyte activation and water homeostasis, respectively. The immunofluorescence analysis revealed that the number of cells co-expressing the markers, which increased in the ischemia-induced group, was significantly reduced in sildenafil-treated groups, suggesting a potential mitigating effect on ischemia-induced astrocyte activation. Additionally, we conducted diverse behavioral tests, including the open-field test, novel object recognition, Barnes maze, Y-maze, and passive avoidance tests, to assess the effect of sildenafil on the cognitive function impaired by ischemia. Cognitive improvements were observed following sildenafil administration (20mg/kg) compared to the untreated group. Taken together, the results presented here suggest sildenafil's potential as a neuroprotective agent for alleviating delayed neuronal cell death and cognitive function impaired by ischemia.
Keywords:
global ischemia
; reperfusion
; sildenafil
; behavioral test
; neuroprotection
1. Introduction
Ischemic stroke, resulting from disrupted blood flow to the brain, causes brain cell death and diverse impairments, including physical, cognitive, and emotional challenges, and leads to death in severe cases [1,2,3,4,5,6,7,8]. Although numerous treatment methods for stroke have been suggested, they are not very effective in actual clinical practice, contrary to the promising results from preclinical studies [4,9,10,11]. Therefore, the only available treatment for ischemic stroke aims only to restore normal blood flow and involves the administration of thrombolytic agents [12]. However, this treatment is limited in that it should be administered within 4.5 hours after the onset of cerebral ischemia [13].
Sildenafil, a type of phosphodiesterase-5 (PDE5) inhibitor, was initially developed and approved as a medication for the treatment of erectile dysfunction (ED) in men. The inhibition of PDE5 by sildenafil increases the intracellular level of cyclic guanosine monophosphate (cGMP), which in turn, leads to vasodilation and increases in blood flow [14,15,16]. The increased blood flow by sildenafil is effective for improving symptoms such as pulmonary hypertension, heart failure, inflammatory response, and fibrin deposition [16,17,18]. In particular, sildenafil also has effect on the brain through neuroprotection, restoring neuronal development, improving brain injury, and enhancing cognitive function [19,20,21]. However, the effects of sildenafil on delayed neuronal cell death and cognitive impairment following global ischemia, as well as its therapeutic mechanisms, remain unknown.
Therefore, in this study, we evaluated the therapeutic effects of sildenafil on systemic ischemia. The use of sildenafil for the treatment of global ischemia has not been extensively studied, and this research will provide valuable information regarding its potential effects.
2. Materials and Methods
2.1. Experimental Animals
All experiments were conducted using 60-80g Mongolian gerbils (Meriones unguiculatus) obtained from the Animal Experiment Center at Soonchunhyang University (Cheonan, Republic of Korea). All animals were provided with a commercial diet and water ad libitum under controlled temperature, humidity, and lighting conditions (22 ± 2°C, 55 ± 5%, and 12:12 light/dark cycle, respectively). All animal protocols were approved by the Soonchunhyang University Animal Experimentation Panel (approval No. SCH22-0086). All efforts were made to minimize the number of animals used and their suffering.
2.2. Induction of Transient Global Cerebral Ischemia
The Mongolian gerbil, with an approximately 70% incomplete circle of Willis, is a useful animal model for studying the mechanisms of delayed neuronal death following global cerebral ischemia [22,23]. Therefore, it serves as a good screening model for evaluating neuroprotective drugs related to brain ischemia [24]. The gerbils were anesthetized with a mixture of 33% oxygen and 67% nitrous oxide containing isoflurane. Both common carotid arteries were isolated and occluded using atraumatic aneurysm clips. Complete occlusion was confirmed by visualizing the common carotid artery using an ophthalmoscope. The aneurysmal clips were removed after a 5-minute occlusion of the common carotid artery. Body (rectal) temperature was monitored using a rectal temperature probe and maintained at 36.7–37 ℃ with a heating pad equipped with a thermostat.
2.3. Drug Administration
The efficacy of sildenafil was evaluated by intraperitoneal injection of sildenafil citrate (SIL, 10 mg/kg or 20 mg/kg; Hanmi Parm. Co., Hwaseng, Republic of Korea) dissolved in 0.8% saline solution 30 minutes before and after the induction of transient global cerebral ischemia. We divided the gerbils into 6 groups as follows: Group 1, control (n = 15); Group 2, ischemic (n = 15); Group 3, SIL-pre (10 mg/kg, n = 15); Group 4, SIL-post (10 mg/kg, n = 15); Group 5, SIL-pre (20 mg/kg, n = 15); and group 6, SIL-post (20 mg/kg, n = 15).
2.4. Behavioral Tests
The animals were exposed to white noise consisting of a combination of 60 Hz frequency sounds for 1 hour before the behavioral tests to facilitate adaptation to various behavioral test environments. All experimental animals underwent behavioral testing 4 days after ischemia. The behavioral tests were recorded and analyzed using a PC-based video behavioral analysis system, EthoVision XT 14 (Noldus, Wageningen, Netherlands), with automated tracking software.
2.4.1. Open-Field Test
The open-field test (OFT) assesses general motor activity and anxiety. The maze arena consists of a 40 × 40 × 40 cm open-field apparatus with diffused lighting. The maze is virtually divided into a central zone and a peripheral zone. The gerbils were placed in the center of the zone and allowed to explore freely for 30 minutes. The total distance moved determined locomotor activity, and the time spent in the central zone was evaluated as the index of anxiety [25,26].
2.4.2. Novel Object Recognition
Object exploratory behavior was investigated using a novel object to exploit the basic exploration characteristics of rodent species. The induction and maintenance of activity vary depending on the difference in response (recognition) to a novel physical object in a familiar place. The novel object recognition (NOR) test used in the open-field test in this study was a white acrylic box. The animals were familiarized with the box for 30 minutes a day before the experiment. In the habituation phase the next day, all groups were placed in the same chamber with 2 similar objects and freely explored them for 10 minutes before returning to their cage. In the test phase 24 hours later, one of the objects was replaced with a new object with a different shape and color. Animals were placed back in the test chamber with the novel and previously explored (familiar) objects for 10 minutes. At each phase, the animals were placed in the center of the box, and their gaze was directed toward the opposite wall of the two objects. Exploration behavior was analyzed through various actions, such as approaching, smelling, licking, and direct contact with the forefoot. We evaluated the ability of the animals to remember familiar objects. Object exploration was defined as the time spent by animals in direct contact with the object (e.g. smelling the object or being less than 1 cm from the object) [27].
2.4.3. Barnes Maze
The Barnes maze consists of a circular platform (92 cm in diameter) with 20 holes evenly spaced around the circumference and located 105 cm above the floor. Four different visual signals were placed on the walls surrounding the platform in each quadrant for the animals to use as cues. For safety an escape chamber was located under one of the holes (escape hole). Bedding was added to the chamber to allow the animals to hide. A metronome (80 Hz noise) and high lighting (300 lux lighting at the center of the platform) were used to amplify anxiety and motivate the animals to find the escape hole. The gerbils interacted with the Barnes maze in 3 stages: habituation (1 day), training (2 - 4 days in a long-term training paradigm), and probe (5 days). During the four-day adaptation phase, animals were allowed to explore the platform from the center when a black cylinder was removed 10 seconds later. Their activities were recorded with a video camera. If the animals found the escape hole within 180 seconds, the hole was covered with a lid for 120 seconds to block light and stop electronic sounds. Otherwise, animals were carefully guided to the hole with the escape chamber to induce learning of the space. Each animal was tested 3 times at 15-minute intervals in the acquisition-training phase to improve spatial memory for finding the escape hole, and the time it took to find the escape hole on the fifth day was measured as the spatial acquisition phase (probe phase). The escape chamber was removed, and this stage was performed only once, unlike the 3 times in the adaptation stage [28,29].
2.4.4. Y-maze
Spontaneous alternation was assessed in the Y-maze test as previously described [27]. The Y-maze consisted of 3 arms (40 x 5 x 20 cm; length, width, height) marked A, B, and C, which were equidistant and converged at a 120° angle. Each animal was placed at the end of one arm and allowed to freely explore for 7 minutes. The order (e.g., ABC, BCA, and CAB) and the number of arm entries were documented manually for each animal during the 7-minute trial. Spontaneous alternation behavior was defined as consecutive selections of ABC, CAB, or BCA without choosing ABA and entering all 3 arms. The maze was thoroughly cleaned with 70% ethanol and distilled water between trials to eliminate residual odor. The percentage of spontaneous alternation behavior was calculated using the following equation: % alternation = [(number of alternations) / (total arm entries - 2)] × 100. The hippocampal-dependent spatial working memory Y-maze was located in a secluded room with specific cues on each arm. The remaining recording and analysis were conducted in the same manner as described above.
2.4.5. Passive Avoidance
The passive avoidance test was conducted using 2 chambers of equal size, one brightly lit and the other dark, separated by a door with an electric grid floor that allowed the electric current to pass through. During the acquisition trial phase, a gerbil was initially placed and allowed to explore the bright chamber for approximately 120 seconds. When the door to the dark chamber was opened, the animal moved, causing the door to close automatically. Training trials were conducted for 24 hours after the acquisition attempts. The gerbil was allowed to explore the bright chamber for about 120 seconds, after which the door was opened. As soon as the gerbil entered the dark chamber, the door closed automatically, and an electric shock (1mA for 2 seconds) was delivered through the grid floor. After 24 hours of training, the testing trial was performed. The retention time was estimated by measuring the time taken by the gerbil to enter the dark chamber after opening the door, within a 180-second time. If the gerbil did not enter the dark chamber within 180 seconds, a retention time of 180 seconds was recorded [30]
2.5. Tissue Processing and Cresyl Violet Staining
Anesthetized animals (with urethane 1.5g/kg) were perfused with saline via the heart, followed by an injection of 4% paraformaldehyde in 0.1M phosphate-buffer (PB). The brain was removed and post-fixed in the same fixative for 4 hours, then exchanged with 30% sucrose with PB for 2 days at 4 °C. The tissue was frozen and sectioned at 30µm thickness using a cryostat and collected onto 6-well plates containing phosphate-buffered saline (PBS) storage solution. Cresyl violet staining was used to investigate the effects of ischemia on cell death or damage, following the procedure described previously [21]. Tissue sections were mounted onto gelatin-coated microscope slides and stained with cresyl violet acetate (Sigma, St. Louis, MO, USA) for 20 minutes at room temperature. The slides were then washed with distilled water, dehydrated in 50%, 70%, 80%, 90%, 95%, and 100% ethanol sequentially for 5 minutes each at room temperature, and finally mounted with Canada balsam (Kanto, Tokyo Japan). After that, images were captured for the measurements of cell death or damage following ischemia using an Axiocam digital camera and ZEN 3.4 microscope digital camera software (Zeiss, Aalen, Germany).
2.6. Double Immunofluorescence
Double-immunofluorescence staining for aquaporin-4 (AQP-4) and glial fibrillary acidic protein (GFAP) was performed to assess morphological changes in the blood-brain barrier (BBB) induced by ischemia-reperfusion in the same hippocampal tissue and changes induced by sildenafil. Brain tissues were incubated overnight at 4°C in a mixture containing primary antibodies in PBS with 0.3% Triton X-100 for 48 hours: mouse anti-GFAP IgG (Millipore; diluted 1:500). After washing with PBS, the sections were incubated at room temperature for 3 hours in Cy2-conjugated secondary antibody diluted 1:200 (Jackson Immuno Research Labs, West Grove, PA, USA). Subsequently, same brain tissues were incubated with another set of primary antibodies overnight at 4°C for 48 hours: rabbit anti-AQP-4 IgG (Alomone Labs, Jerusalem, Israel; diluted 1:200). The tissues were then incubated with Cy3-conjugated secondary antibody for 3 hours at room temperature. After washing with PBS, the tissues were incubated with DAPI (Invitrogen, Waltham, MA, USA) diluted 1:500 for 10 minutes at room temperature, mounted on gelatin-coated slides, and cover-slipped with DPX (Sigma). All images were captured using a Fluoview FV10i microscope and FV10i software (Olympus, Tokyo, Japan).
2.7. Statistical Analysis
All data were analyzed using GraphPad Prism 8.0.1 (GraphPad Software, San Diego, CA, USA). The data are presented as the mean ± standard deviation. One and two-way analysis of variance (ANOVA) was used to analyze the statistical significance of the data. Bonferroni's test was employed for post-hoc comparisons. Statistical significance was defined as *p < 0.05, **p < 0.01, and ***p < 0.001.
3. Results
3.1. Neuroprotective Effects of Sildenafil on Neuronal Cell Death Following Transient Global Cerebral Ischemia-Reperfusion
First, we tested the neuroprotective effects of sildenafil on brains with ischemia. Cresyl violet staining was performed to assess neuronal cell death and damage in the hippocampus of the animal model after transient global cerebral ischemia-reperfusion (Figure 1). Responses to Nissl bodies in the control gerbils were predominantly detected in hippocampal CA1 neurons and various neuronal cell bodies, encompassing the majority of the neuronal cell populations (Figure 1A1,A2). Most neuronal populations in the hippocampal CA1 region of ischemic animals showed significant cell death (Figure 1B1,B2). Quantitative analysis of viable neurons yielded results consistent with the histological examination of cresyl violet staining (ischemia group, F1, 20 = 53.52, p < 0.001 by one-way ANOVA; Figure 1G). Sildenafil pre-treated (SIL-pre 10mg/kg) gerbils did not demonstrate sufficient changes in neuroprotection (F1, 20 = 3.858, p = 0.084 by one-way ANOVA; Figure 1C1,C2 and G). In contrast, the number of neurons in the hippocampal CA1 region increased in SIL-post (10mg/kg) gerbils compared to ischemic gerbils (F1, 20 = 8.918, p < 0.001 by one-way ANOVA; Figure 1D1,D2 and G). Both the SIL-pre and SIL-post (20mg/kg) gerbils showed significant increases in the expression of Nissl bodies compared to ischemic gerbils (F1, 20 = 23.38 and 28.8, p < 0.001 by one-way ANOVA; Figure 1E1-F2 and G).
3.2. Effects of Sildenafil on AQP4 and GFAP Expression Following Transient Global Cerebral Ischemia-Reperfusion
Double-staining for AQP4 and GFAP was performed to assess the functional changes induced by transient cerebral ischemia-reperfusion and sildenafil treatment in the hippocampus by BBB disruption (Figure 2). GFAP and AQP4-positive immunoreactivity were significantly increased in the hippocampal CA1 region of the ischemic gerbils compared to control gerbils (Figure 2A1-3 and B1-3). The number of double-labeled astrocytes and vessels, which express both AQP4 and GFAP, in the hippocampal CA1 region was increased in ischemic gerbils compared to control gerbils (ischemic group, F1,10 = 7.432, p < 0.001 by one-way ANOVA; Figure 2A4, B4 and G). In contrast, in all of the sildenafil pre- and post-treatment groups, the double-labeling of AQP4 and GFAP was significantly restored to levels comparable to control gerbils (SIL-pre and -post(10mg/kg), F1,10 = 5.23 and 4.129, p < 0.001; SIL-pre and -post(20mg/kg), F1,10 = 4.954 and 5.78, p < 0.001 by one-way ANOVA; Figure 2C4-F4 and G). AQP4 immunoreactivity was not decreased in any group (Figure 2A2-F2), whereas GFAP immunoreactivity was decreased in all groups administered sildenafil compared to ischemic gerbils (Figure 2B3-F3).
3.3. Hyper-Locomotion Activity Following Ischemia in Gerbils Treated with Sildenafil
The OFT was conducted on day 4 after ischemia-reperfusion to assess the changes in general locomotor activity observed in experimental animals after ischemic stroke and evaluate the impact of sildenafil (Figure 3). Representative tracings of each group are displayed in Figure 3A, and the movement tracks of each group (line) are indicated. The ischemic gerbils showed a significant increase in the total distance moved compared to control gerbils (ischemic group, F1,28 = 16.89, p < 0.001 by one-way ANOVA; Figure 3A,B). No change in the total distance moved was seen in SIL-pre (10 mg/kg) gerbils, whereas a significant decrease was observed in SIL-post (10 mg/kg) gerbils compared to ischemic ones [SIL-pre (10mg/kg), F1,28 = 1.395, p = 0.9209; SIL-post (10mg/kg), F1,28 = 10.26, p < 0.001 by one-way ANOVA; Figure 3A, B]. In contrast, both the SIL-pre and SIL-post (20 mg/kg) gerbil groups exhibited a decrease in the total distance moved to the level of the control gerbils [SIL-pre (20mg/kg), F1,28 = 10.37, p < 0.001; SIL-post (20mg/kg), F1,28 = 13.15, p < 0.001 by one-way ANOVA; Figure 3A and B).
3.4. Sildenafil Treatment Alleviates Deficits in Cognitive Behavior Following Ischemia in Gerbils
The NOR test was performed to evaluate cognitive function related to the exploration of novel objects [27]. During the habituation phase, no statistically significant differences were seen in exploration frequency or time percentage between gerbils in the control, ischemic, and sildenafil groups, indicating comparable adaptation patterns between two identical objects (Figure 4A1,A2). In the test phase, the control gerbils showed a significant increase in exploration frequency and time percentage for the novel object compared to the familiar object (control group, frequency, F1,28 = 27.1, p < 0.001; exploration time, F1,28 = 25.94, p < 0.001 by two-way ANOVA; Figure 4B1,B2). However, in the ischemic gerbils, no statistical differences were found in these measures, indicating similar preferences for the novel and familiar objects (ischemic group, frequency, F1,28 = 0.172, p = 0.999; exploration time, F1,28 = 0.902, p = 0.9368 by two-way ANOVA; Figure 4B1, B2). The frequency and exploration time percentage for the novel object were significantly increased in all sildenafil-treat gerbils, similar to the control gerbils. [SIL-pre and SIL-post (10 mg/kg), frequency, F1,28 = 13.7 and 10.1, p < 0.001; exploration time, F1,28 = 18.09 and 14.46, p < 0.001; SIL-pre and SIL-post (20 mg/kg), frequency, F1,28 = 16.8 and 22.5. p < 0.001; exploration time, F1,28 = 18.83 and 21.55, p < 0.001 by two-way ANOVA; Figure 4B1,B2]. In addition, the discrimination index between familiar and novel objects was reduced in ischemic gerbils, but significantly improved in all sildenafil-treated animals [ischemic group, F1,28 = 17.71, p < 0.001; SIL-pre and SIL-post (10 mg/kg), F1,23 = 13.44 and 10.63, p < 0.001; SIL-pre and SIL-post (20 mg/kg), F1,23 = 14.02 and 16.12, p < 0.001 by one-way ANOVA; Fig 4B3].
The Barnes maze test was performed to investigate spatial reference learning and memory (Figure 5). All groups showed improvements throughout the training phase in the number of errors and latency time in finding the escape hole through repeated attempts (Figure 5A and C). In the probe phase, the number of errors and latency time to the escape hole were significantly increased in ischemic gerbils compared to control gerbils (ischemic group, number of errors, F1,18 = 19.62, p < 0.001; latency, F1,18 = 32.38, p < 0.001 by one-way ANOVA; Figure 5B,D), indicating a pronounced learning deficit during this period. In contrast, regardless of the dosage, all sildenafil-treated groups showed a significant reduction in the number of errors [SIL-pre (10 mg/kg), F1,18 = 10.32, p < 0.001; SIL-post (10 mg/kg), F1,18 = 4.772, p < 0.05; SIL-pre (20 mg/kg), F1,18 = 13.91, p < 0.001; SIL-post (20 mg/kg), F1,18 = 17.76, p < 0.001 by one-way ANOVA; Figure 5B] and in latency time to the escape hole on the probe test day compared with ischemic gerbils [SIL-pre and SIL-post (10 mg/kg), F1,18 = 23.45 and 20.74, p < 0.001; SIL-pre and SIL-post (20 mg/kg), F1,18 = 20.93 and 27.58, p < 0.001 by one-way ANOVA; Figure 5D].
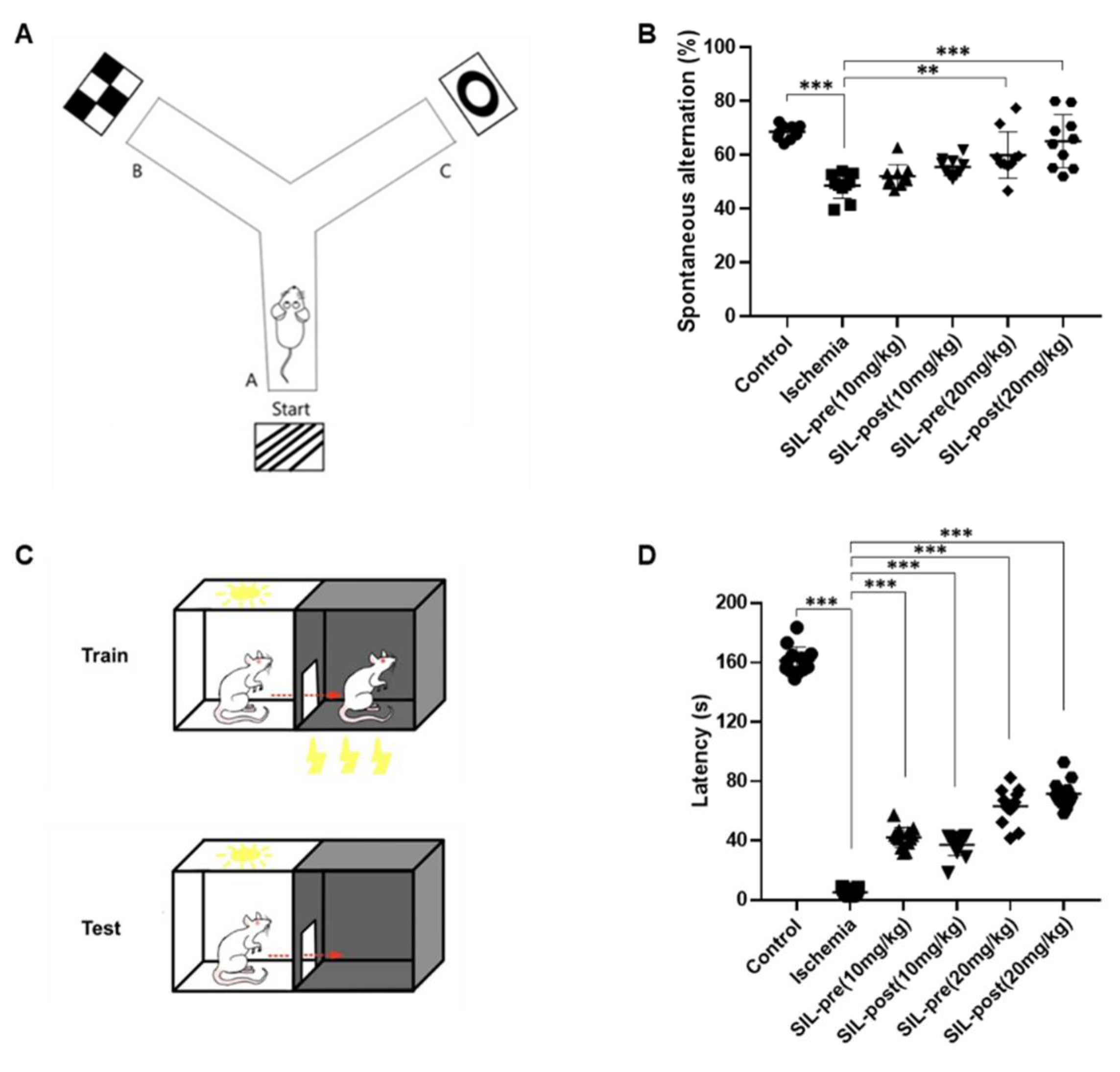
The Y-maze task was utilized to evaluate hippocampus-dependent spatial working memory, which is a form of short-term memory, and the passive avoidance test was used to evaluate emotional short-term memory (Figure 6A,C) [27,30]. The average percentage of spontaneous alternations in ischemic gerbils was significantly decreased compared to control gerbils (ischemic group, F1,18 = 10.2, p < 0.001 by one-way ANOVA; Figure 6B), indicating impaired memory and spatial working memory. Both sildenafil-treated (10 mg/kg) gerbils showed a tendency toward increased voluntary spontaneous alternations. However, this difference was not statistically significant compared to ischemic gerbils [SIL-pre (10 mg/kg), F1,18 = 1.793, p = 0.8009; SIL-post (10 mg/kg), F1,18 = 3.545, p = 0.1402 by one-way ANOVA; Figure 6B]. However, both the SIL-pre and SIL-post treated (20 mg/kg) groups showed an increase in the average percentage of spontaneous alternations compared to ischemic group [SIL-pre (20 mg/kg), F1,18 = 5.803, p < 0.01; SIL-post (20 mg/kg), F1,18 = 8.432, p < 0.001 by one-way ANOVA; Figure 6B]. The experimental results for passive avoidance indicated a significant increase in latency in the ischemic group compared to the control group (ischemic group, F1,24 = 68.84, p < 0.001 by one-way ANOVA; Figure 6D). An increase in latency was seen in all sildenafil-treated gerbils compared to ischemic gerbils [SIL-pre and SIL-post (10 mg/kg), F1,24 = 16.29 and 14.08, p < 0.001; SIL-pre and SIL-post (20 mg/kg), F1, 24 = 25.59 and 29.26, p < 0.001 by one-way ANOVA; Figure 6D]. As the sildenafil dosage increased, there was a tendency for transfer latency to increase. These results suggest that sildenafil may improve cognitive function in brains affected by ischemic damage.
4. Discussion
In this study, we confirmed the beneficial efficacy of sildenafil treatment in improving after-stroke symptoms. Brain edema is a serious complication of stroke, and previous studies reported that cellular edema in ischemia primarily affects astroglia, increasing glial K+ uptake [31]. Additionally, AQP4-knockout mice exhibited neuroprotection and improved inflammation due to reductions in brain edema in transient cerebral ischemia [31,32]. The double-immunofluorescence analysis of GFAP and AQP-4 co-expression provides evidence of astrocytic activation and water regulatory mechanisms in response to global ischemic insults. The administration of sildenafil significantly reduced the co-expression of GFAP and AQP-4 across all treatment groups. This finding aligns with previous research indicating the anti-inflammatory and neuroprotective properties of sildenafil [21]. Sildenafil inhibits microglia activation and the death of oligodendrocytes through the mitogen-activated protein kinase (MAPK) signaling pathway, while increases in the production of pro-inflammatory cytokines via PKG signaling affects the activation of astrocytes and microglia [20,33,34,35]. The downregulation of GFAP and AQP-4 co-expression implies mitigation of the astrocytic reaction and potential reductions in edema formation, supporting the hypothesis that sildenafil contributes to neuroprotection in the context of global cerebral ischemia. These findings contribute to our understanding of the complex cellular interactions involved in ischemic events and highlight the therapeutic potential of sildenafil in mitigating astrocyte-mediated damage.
Previous research reported that gerbils exhibited hyperactivity in the early stages following ischemia [24,36,37] and suggested that cell loss induced by global ischemia, along with the consequent loss of spatial mapping function, contributes to hyperlocomotion in the open-field test [24]. Improvements in hippocampal delayed neuronal death by neuroprotective agents has been claimed to ameliorate hyperactivity, suggesting a close association between hyperlocomotion and hippocampal delayed neuronal death [24]. Consistent with these results, the decrease in cell death caused by sildenafil prevented decreases in activity in the open-field test (Figure 1 and 3). Further investigations are needed to clarify the causal relationship of these changes.
We utilized the NOR test, Barnes maze, Y-maze with a special cue, and the passive avoidance test to assess hippocampal-dependent memory [27,38,39,40,41,42,43]. The Barnes maze test serves as a dry-land-based test for spatial learning and memory [40,41]. It represents a hippocampus-dependent task where animals learn the relationship between distal cues in the surrounding environment and a fixed escape location [41]. Spontaneous alternation behavior in the Y-maze test is commonly interpreted as a task evaluating working memory in a novel environment [38,39]. We conducted Y-maze tests using spatial cues to evaluate hippocampus-dependent spatial working memory [42]. The passive avoidance test was employed as a measure of aversive memory [45]. It is recognized for its ability to assess an animal's ability to remember and avoid unpleasant stimuli [44,45]. In our study, ischemic gerbils exhibited impairments in spatial learning and cognitive function similar to previous research findings (Figure 4-6). This observation aligns with existing knowledge that highlights the vulnerability of hippocampal-dependent tasks to ischemic damage [27,46,47]. In the present results, the cognitive impairment of novel objects in the ischemia group increased to the level of the control group regardless of the dose and timing of sildenafil administration (Figure 4). The group treated with sildenafil demonstrated a marked improvement in spatial learning, spatial memory, working memory, and aversive memory (Figure 5 and 6). This improvement suggests a potential ameliorative effect of sildenafil not only on spatial learning and memory but also on working and aversive memory following an ischemic insult. According to the current study results, spontaneous alternations in short-term spatial working memory were significantly impaired in the ischemic group, while the group treated with 20mg/kg of sildenafil demonstrated significant improvements compared to the ischemic group (Figure 6B). The inhibition of the PDE5 effect by sildenafil increases intracellular cGMP levels by breaking down the phosphodiester bond of cGMP and inhibiting the hydrolysis of cGMP to GMP [20]. Previous studies have proposed that sildenafil activates the PI3K/Akt pathway via the NO-cGMP-PKG signaling pathway, enhancing neurogenesis and synaptic plasticity [48,49]. Therefore, our results demonstrated the synaptic plasticity-enhancing effects of sildenafil administration in improving various hippocampal-dependent memory impairments following transient global cerebral ischemia. In particular, the NOR test involves various brain regions influencing memory, learning, preference for novelty, and recognition processes associated with the hippocampus and the perirhinal cortex [50,51]. Aversive emotional learning is regulated by neurotransmitters, such as acetylcholine, noradrenaline, and dopamine, in brain regions, including the amygdala, cortex, hippocampus, striatum, and medial prefrontal cortex [54]. Clinical studies revealed that theta oscillations originating from the amygdala temporarily modulate amygdala-hippocampal gamma power coherence, facilitating the encoding of aversive memories [53]. Therefore, our study findings suggest that sildenafil may contribute to positive effects in other brain regions besides the hippocampus, warranting further investigation.
5. Conclusions
The combined results of behavioral and neurobiological assessments support the notion that sildenafil mitigates ischemic damage, particularly highlighted in the attenuation of activated astrocytic responses, as indicated by the double-immunofluorescence analysis of GFAP and AQP-4. The neuroprotective properties of sildenafil in global cerebral ischemia are associated with its ability to regulate intracellular signaling pathways related to memory formation.
However, certain study limitations must be acknowldedged. The study primarily focused on short-term effects, and the long-term consequences of sildenafil treatment remain unclear. The observed dose-dependent response, with the higher dose exhibiting a more pronounced effect, raises questions about the optimal dosage for achieving cognitive benefits. This prompts the need for further exploration into the dose-response relationship and necessitates a nuanced understanding of the pharmacokinetics and pharmacodynamics of sildenafil in the context of cerebral ischemia. The distinctive performance between the pre-treated and post-treated groups introduces a temporal dimension to sildenafil's cognitive effects. The consistently superior performance of the post-treated groups suggests that the timing of sildenafil administration may be a critical determinant in harnessing its full cognitive benefits. This temporal aspect adds a layer of complexity to the design of therapeutic strategies, emphasizing the need for precise timing considerations in clinical applications.
Despite these limitations, this research provides a foundation for future investigations into sildenafil's therapeutic potential for cognitive impairments associated with cerebral ischemia. The multifaceted approach, combining behavioral tests with molecular analyses, contributes to a comprehensive understanding of sildenafil's impact on post-ischemic cognitive function. Overall, this study opens avenues for further exploration and emphasizes the need for clinical trials to validate these preclinical findings in a translational context.
Author Contributions
Conceptualization, Y.H.Y., D.-K.P. and D.-S.K.; methodology, G.W.K., Y.R.L., and D.-K.P.; formal analysis, Y.H.Y., G.W.K., and D.-S.K.; investigation, Y.H.Y. and Y.R.L.; data curation, Y.H.Y. and G.W.K. writing—original draft preparation, Y.H.Y., G.W.K., B.S.; writing—review and editing, Y.H.Y., G.W.K., and B.S.; visualization, Y.H.Y., G.W.K. and B.S.; supervision, B.S. and D.-S.K.; funding acquisition, Y.H.Y., G.W.K.and D.-S.K. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Soonchunhyang University and National Research Foundation of Korea: NRF-2017R1D1A1B05036195 and RS-2023-00245605.
Institutional Review Board Statement
All animal protocols were approved by the Administrative Panel on Laboratory Animal Care of Soonchunhyang University (permit No. SCH22-0086).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Nagakannan, P.; Shivasharan, B.D.; Thippeswamy, B.S.; Veerapur, V.P.; Bansal, P. Protective Effect of Hydroalcoholic Extract of Mimusops Elengi Linn. Flowers against Middle Cerebral Artery Occlusion Induced Brain Injury in Rats. J. Ethnopharmacol. 2012, 140. [Google Scholar] [CrossRef] [PubMed]
- Leys, D.; Hénon, H.; Mackowiak-Cordoliani, M.A.; Pasquier, F. Poststroke Dementia. Lancet Neurol. 2005, 4. [Google Scholar] [CrossRef] [PubMed]
- Román, G.C.; Tatemichi, T.K.; Erkinjuntti, T.; Cummings, J.L.; Masdeu, J.C.; Garcia, J.H.; Amaducci, L.; Orgogozo, J.M.; Brun, A.; Hofman, A.; et al. Vascular Dementia: Diagnostic Criteria for Research Studies - Report of the NINDS-AIREN International Workshop. Neurology 1993, 43. [Google Scholar] [CrossRef]
- Rost, N.S.; Brodtmann, A.; Pase, M.P.; van Veluw, S.J.; Biffi, A.; Duering, M.; Hinman, J.D.; Dichgans, M. Post-Stroke Cognitive Impairment and Dementia. Circ. Res. 2022, 130. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.L.; Murray, L.; Richards, L.; Zorowitz, R.D.; Bakas, T.; Clark, P.; Billinger, S.A. Comprehensive Overview of Nursing and Interdisciplinary Rehabilitation Care of the Stroke Patient: A Scientific Statement from the American Heart Association. Stroke 2010, 41. [Google Scholar] [CrossRef] [PubMed]
- Teasell, R.; Hussein, N. Clinical Consequences of Stroke Pg; 2016; Vol. 1;
- Aqueveque, P.; Ortega, P.; Pino, E.; Saavedra, F.; Germany, E.; Gómez, B. After Stroke Movement Impairments: A Review of Current Technologies for Rehabilitation. In Physical Disabilities - Therapeutic Implications; 2017.
- Hewetson, R.; Cornwell, P.; Shum, D. Social Participation Following Right Hemisphere Stroke: Influence of a Cognitive-Communication Disorder. Aphasiology 2018, 32. [Google Scholar] [CrossRef]
- Sun, J.H.; Tan, L.; Yu, J.T. Post-Stroke Cognitive Impairment: Epidemiology, Mechanisms and Management. Ann. Transl. Med. 2014, 2. [Google Scholar]
- Fure, B.; Wyller, T.B.; Engedal, K.; Thommessen, B. Emotional Symptoms in Acute Ischemic Stroke. Int. J. Geriatr. Psychiatry 2006, 21. [Google Scholar] [CrossRef] [PubMed]
- Ferro, J.M.; Caeiro, L.; Figueira, M.L. Neuropsychiatric Sequelae of Stroke. Nat. Rev. Neurol. 2016, 12. [Google Scholar] [CrossRef]
- Wechsler, L.R. Intravenous Thrombolytic Therapy for Acute Ischemic Stroke. N. Engl. J. Med. 2011, 364. [Google Scholar] [CrossRef]
- O’Collins, V.E.; Macleod, M.R.; Donnan, G.A.; Horky, L.L.; Van Der Worp, B.H.; Howells, D.W. 1,026 Experimental Treatments in Acute Stroke. Ann. Neurol. 2006, 59. [Google Scholar] [CrossRef] [PubMed]
- Andersson, K.E. PDE5 Inhibitors – Pharmacology and Clinical Applications 20 Years after Sildenafil Discovery. Br. J. Pharmacol. 2018, 175, 175. [Google Scholar] [CrossRef] [PubMed]
- Rutten, K.; De Vente, J.; Şik, A.; Markerink-Van Ittersum, M.; Prickaerts, J.; Blokland, A. The Selective PDE5 Inhibitor, Sildenafil, Improves Object Memory in Swiss Mice and Increases CGMP Levels in Hippocampal Slices. Behav. Brain Res. 2005, 164. [Google Scholar] [CrossRef] [PubMed]
- Moretti, R.; Leger, P.L.; Besson, V.C.; Csaba, Z.; Pansiot, J.; Di Criscio, L.; Gentili, A.; Titomanlio, L.; Bonnin, P.; Baud, O.; et al. Sildenafil, a Cyclic GMP Phosphodiesterase Inhibitor, Induces Microglial Modulation after Focal Ischemia in the Neonatal Mouse Brain. J. Neuroinflammation 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Jiang, Q.; Zhang, L.; Ding, G.; Gang Zhang, Z.; Li, Q.; Ewing, J.R.; Lu, M.; Panda, S.; Ledbetter, K.A.; et al. Angiogenesis and Improved Cerebral Blood Flow in the Ischemic Boundary Area Detected by MRI after Administration of Sildenafil to Rats with Embolic Stroke. Brain Res. 2007, 1132. [Google Scholar] [CrossRef]
- de Visser, Y.P.; Walther, F.J.; Laghmani, E.H.; Boersma, H.; van der Laarse, A.; Wagenaar, G.T.M. Sildenafil Attenuates Pulmonary Inflammation and Fibrin Deposition, Mortality and Right Ventricular Hypertrophy in Neonatal Hyperoxic Lung Injury. Respir. Res. 2009, 10. [Google Scholar] [CrossRef] [PubMed]
- Cuadrado-Tejedor, M.; Hervias, I.; Ricobaraza, A.; Puerta, E.; Pérez-Roldán, J.M.; García-Barroso, C.; Franco, R.; Aguirre, N.; García-Osta, A. Sildenafil Restores Cognitive Function without Affecting β-Amyloid Burden in a Mouse Model of Alzheimer’s Disease. Br. J. Pharmacol. 2011, 164. [Google Scholar] [CrossRef]
- Xiong, Y.; Wintermark, P. The Role of Sildenafil in Treating Brain Injuries in Adults and Neonates. Front. Cell. Neurosci. 2022, 16. [Google Scholar] [CrossRef]
- Yu, Y.H.; Kim, S.W.; Kang, J.; Song, Y.; Im, H.; Kim, S.J.; Yoo, D.Y.; Lee, M.R.; Park, D.K.; Oh, J.S.; et al. Phosphodiesterase-5 Inhibitor Attenuates Anxious Phenotypes and Movement Disorder Induced by Mild Ischemic Stroke in Rats. J. Korean Neurosurg. Soc. 2022, 65. [Google Scholar] [CrossRef]
- Martínez, N.S.; Machado, J.M.; Saad, H.P.; Antich, R.M.C.; Acosta, J.A.B.; Salgueiro, S.R.; Illera, G.G.; Alba, J.S.; del Barco Herrera, D.G. Global Brain Ischemia in Mongolian Gerbils: Assessing the Level of Anastomosis in the Cerebral Circle of Willis. Acta Neurobiol. Exp. (Wars). 2012, 72. [Google Scholar]
- Delbarre, G.; Delbarre, B.; Barrau, Y. A Suitable Method to Select Gerbils with Incomplete Circle of Willis: To the Editor. Stroke 1988, 19. [Google Scholar] [CrossRef] [PubMed]
- Katsuta, K.; Umemura, K.; Ueyama, N.; Matsuoka, N. Pharmacological Evidence for a Correlation between Hippocampal CA1 Cell Damage and Hyperlocomotion Following Global Cerebral Ischemia in Gerbils. Eur. J. Pharmacol. 2003, 467. [Google Scholar] [CrossRef] [PubMed]
- Tóthová, Ľ.; Bábíčková, J.; Borbélyová, V.; Filová, B.; Šebeková, K.; Hodosy, J. Chronic Renal Insufficiency Does Not Induce Behavioral and Cognitive Alteration in Rats. Physiol. Behav. [CrossRef]
- Gangadharan, G.; Shin, J.; Kim, S.W.; Kim, A.; Paydar, A.; Kim, D.S.; Miyazaki, T.; Watanabe, M.; Yanagawa, Y.; Kim, J.; et al. Medial Septal GABAergic Projection Neurons Promote Object Exploration Behavior and Type 2 Theta Rhythm. Proc. Natl. Acad. Sci. U. S. A. 6550. [Google Scholar] [CrossRef]
- Wappler, E.A.; Szilágyi, G.; Gál, A.; Skopál, J.; Nyakas, C.; Nagy, Z.; Felszeghy, K. Adopted Cognitive Tests for Gerbils: Validation by Studying Ageing and Ischemia. Physiol. Behav. 2009. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, T.P.; Brown, R.E. The Effects of Apparatus Design and Test Procedure on Learning and Memory Performance of C57BL/6J Mice on the Barnes Maze. J. Neurosci. Methods 2012. [Google Scholar] [CrossRef] [PubMed]
- Parent, J.M.; Elliott, R.C.; Pleasure, S.J.; Barbaro, N.M.; Lowenstein, D.H. Aberrant Seizure-Induced Neurogenesis in Experimental Temporal Lobe Epilepsy. Ann. Neurol. 2006. [Google Scholar] [CrossRef] [PubMed]
- Nasehi, M.; Piri, M.; Nouri, M.; Farzin, D.; Nayer-Nouri, T.; Zarrindast, M.R. Involvement of Dopamine D1/D2 Receptors on Harmane-Induced Amnesia in the Step-down Passive Avoidance Test. Eur. J. Pharmacol. 2010, 634. [Google Scholar] [CrossRef]
- Vella, J.; Zammit, C.; Di Giovanni, G.; Muscat, R.; Valentino, M. The Central Role of Aquaporins in the Pathophysiology of Ischemic Stroke. Front. Cell. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [PubMed]
- Hirt, L.; Fukuda, A.M.; Ambadipudi, K.; Rashid, F.; Binder, D.; Verkman, A.; Ashwal, S.; Obenaus, A.; Badaut, J. Improved Long-Term Outcome after Transient Cerebral Ischemia in Aquaporin-4 Knockout Mice. J. Cereb. Blood Flow Metab. 2017, 37. [Google Scholar] [CrossRef] [PubMed]
- Pifarré, P.; Gutierrez-Mecinas, M.; Prado, J.; Usero, L.; Roura-Mir, C.; Giralt, M.; Hidalgo, J.; García, A. Phosphodiesterase 5 Inhibition at Disease Onset Prevents Experimental Autoimmune Encephalomyelitis Progression through Immunoregulatory and Neuroprotective Actions. Exp. Neurol. 2014, 251. [Google Scholar] [CrossRef]
- de Santana Nunes, A.K.; Rapôso, C.; Björklund, U.; da Cruz-Höfling, M.A.; Peixoto, C.A.; Hansson, E. Sildenafil (Viagra®) Prevents and Restores LPS-Induced Inflammation in Astrocytes. Neurosci. Lett. 2016, 630. [Google Scholar] [CrossRef]
- Araújo, S.M. da R.; Duarte-Silva, E.; Marinho, C.G. de S.; Oliveira, W.H.; França, M.E.R. de; Lós, D.; Peron, G.; Tomaz, L.; Bonfanti, A.P.; Verinaud, L.; et al. Effect of Sildenafil on Neuroinflammation and Synaptic Plasticity Pathways in Experimental Autoimmune Encephalomyelitis. Int. Immunopharmacol. 2020, 85. [Google Scholar] [CrossRef] [PubMed]
- Karasawa, Y.; Araki, H.; Otomo, S. Changes in Locomotor Activity and Passive Avoidance Task Performance Induced by Cerebral Ischemia in Mongolian Gerbils. Stroke 1994, 25. [Google Scholar] [CrossRef] [PubMed]
- Katsumata, N.; Kuroiwa, T.; Ishibashi, S.; Li, S.; Endo, S.; Ohno, K. Heterogeneous Hyperactivity and Distribution of Ischemic Lesions after Focal Cerebral Ischemia in Mongolian Gerbils. Neuropathology 2006, 26. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, H.; Lee, Y.B.; Lee, B.; Lee, D. A Quantitative Analysis of Spontaneous Alternation Behaviors on a Y-Maze Reveals Adverse Effects of Acute Social Isolation on Spatial Working Memory. Sci. Rep. 2023, 13. [Google Scholar] [CrossRef] [PubMed]
- Hazim, A.I.; Mustapha, M.; Mansor, S.M. The Effects on Motor Behaviour and Short-Term Memory Tasks in Mice Following an Acute Administration of Mitragyna Speciosa Alkaloid Extract and Mitragynine. J. Med. Plant Res. 2011. [Google Scholar]
- Chachua, T.; Goletiani, C.; Maglakelidze, G.; Sidyelyeva, G.; Daniel, M.; Morris, E.; Miller, J.; Shang, E.; Wolgemuth, D.J.; Greenberg, D.A.; et al. Sex-Specific Behavioral Traits in the Brd2 Mouse Model of Juvenile Myoclonic Epilepsy. Genes, Brain Behav. [CrossRef]
- Pitts, M. Barnes Maze Procedure for Spatial Learning and Memory in Mice. BIO-PROTOCOL. [CrossRef]
- Yu, Y.H.; Kim, S.W.; Im, H.; Oh, S.W.; Cho, N.J.; Park, S.; Park, D.K.; Kim, D.S.; Gil, H.W. Cognitive Sequelae and Hippocampal Dysfunction in Chronic Kidney Disease Following 5/6 Nephrectomy. Brain Sci. 2022, 12. [Google Scholar] [CrossRef] [PubMed]
- Moriarty, O.; Roche, M.; McGuire, B.E.; Finn, D.P. Validation of an Air-Puff Passive-Avoidance Paradigm for Assessment of Aversive Learning and Memory in Rat Models of Chronic Pain. J. Neurosci. Methods 2012, 204. [Google Scholar] [CrossRef]
- Mendl, M.; Burman, O.; Laughlin, K.; Paul, E. Animal Memory and Animal Welfare. Anim. Welf. 2001, 10. [Google Scholar] [CrossRef]
- Steele, R.J.; Stewart, M.G.; Rose, S.P.R. Increases in NMDA Receptor Binding Are Specifically Related to Memory Formation for a Passive Avoidance Task in the Chick: A Quantitative Autoradiographic Study. Brain Res. 1995, 674. [Google Scholar] [CrossRef]
- Squire, L.R.; Zola, S.M. Ischemic Brain Damage and Memory Impairment: A Commentary. Hippocampus 1996, 6. [Google Scholar] [CrossRef]
- Lee, T.K.; Hong, J.; Lee, J.W.; Kim, S.S.; Sim, H.; Lee, J.C.; Kim, D.W.; Lim, S.S.; Kang, I.J.; Won, M.H. Ischemia-Induced Cognitive Impairment Is Improved via Remyelination and Restoration of Synaptic Density in the Hippocampus after Treatment with Cog-Up® in a Gerbil Model of Ischemic Stroke. Vet. Sci. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Peixoto, C.A.; Nunes, A.K.S.; Garcia-Osta, A. Phosphodiesterase-5 Inhibitors: Action on the Signaling Pathways of Neuroinflammation, Neurodegeneration, and Cognition. Mediators Inflamm. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Yazdani, A.; Howidi, B.; Shi, M.Z.; Tugarinov, N.; Khoja, Z.; Wintermark, P. Sildenafil Improves Hippocampal Brain Injuries and Restores Neuronal Development after Neonatal Hypoxia–Ischemia in Male Rat Pups. Sci. Rep. 2021, 11. [Google Scholar] [CrossRef] [PubMed]
- Antunes, M.; Biala, G. The Novel Object Recognition Memory: Neurobiology, Test Procedure, and Its Modifications. Cogn. Process. 2012, 13. [Google Scholar] [CrossRef]
- Kinnavane, L.; Amin, E.; Olarte-Sánchez, C.M.; Aggleton, J.P. Detecting and Discriminating Novel Objects: The Impact of Perirhinal Cortex Disconnection on Hippocampal Activity Patterns. Hippocampus 2016, 26. [Google Scholar] [CrossRef]
- Likhtik, E.; Johansen, J.P. Neuromodulation in Circuits of Aversive Emotional Learning. Nat. Neurosci. 2019, 22. [Google Scholar] [CrossRef]
- Costa, M.; Lozano-Soldevilla, D.; Gil-Nagel, A.; Toledano, R.; Oehrn, C.R.; Kunz, L.; Yebra, M.; Mendez-Bertolo, C.; Stieglitz, L.; Sarnthein, J.; et al. Aversive Memory Formation in Humans Involves an Amygdala-Hippocampus Phase Code. Nat. Commun. 2022, 13. [Google Scholar] [CrossRef]
Figure 1.
Sildenafil treatment increased cell populations in the CA1 region compared to transient global cerebral ischemia. In the cresyl violet staining, the number of neurons in the stratum pyramidal (SP, asterisk) of the CA1 field was reduced in ischemic gerbils compared to control gerbils (A1-B2 and G). SIL-pre (10 mg/kg) gerbils did not show significant recovery (C1-C2 and G). The number of neurons in SIL-post (10 mg/kg) gerbils was significantly increased compared to ischemic gerbils (D1-D2 and G, arrowhead). Especially, the number of neurons in the SP area in the SIL-treated (20 mg/kg) groups were significantly increased compared to the ischemic group (E1-F2 and G, arrowhead). Scale bar = A1-F1, 200μm; A2-F2, 50μm. Data are presented as means ± standard errors of the mean. ***p < 0.001 by one-way analysis of variance.
Figure 1.
Sildenafil treatment increased cell populations in the CA1 region compared to transient global cerebral ischemia. In the cresyl violet staining, the number of neurons in the stratum pyramidal (SP, asterisk) of the CA1 field was reduced in ischemic gerbils compared to control gerbils (A1-B2 and G). SIL-pre (10 mg/kg) gerbils did not show significant recovery (C1-C2 and G). The number of neurons in SIL-post (10 mg/kg) gerbils was significantly increased compared to ischemic gerbils (D1-D2 and G, arrowhead). Especially, the number of neurons in the SP area in the SIL-treated (20 mg/kg) groups were significantly increased compared to the ischemic group (E1-F2 and G, arrowhead). Scale bar = A1-F1, 200μm; A2-F2, 50μm. Data are presented as means ± standard errors of the mean. ***p < 0.001 by one-way analysis of variance.
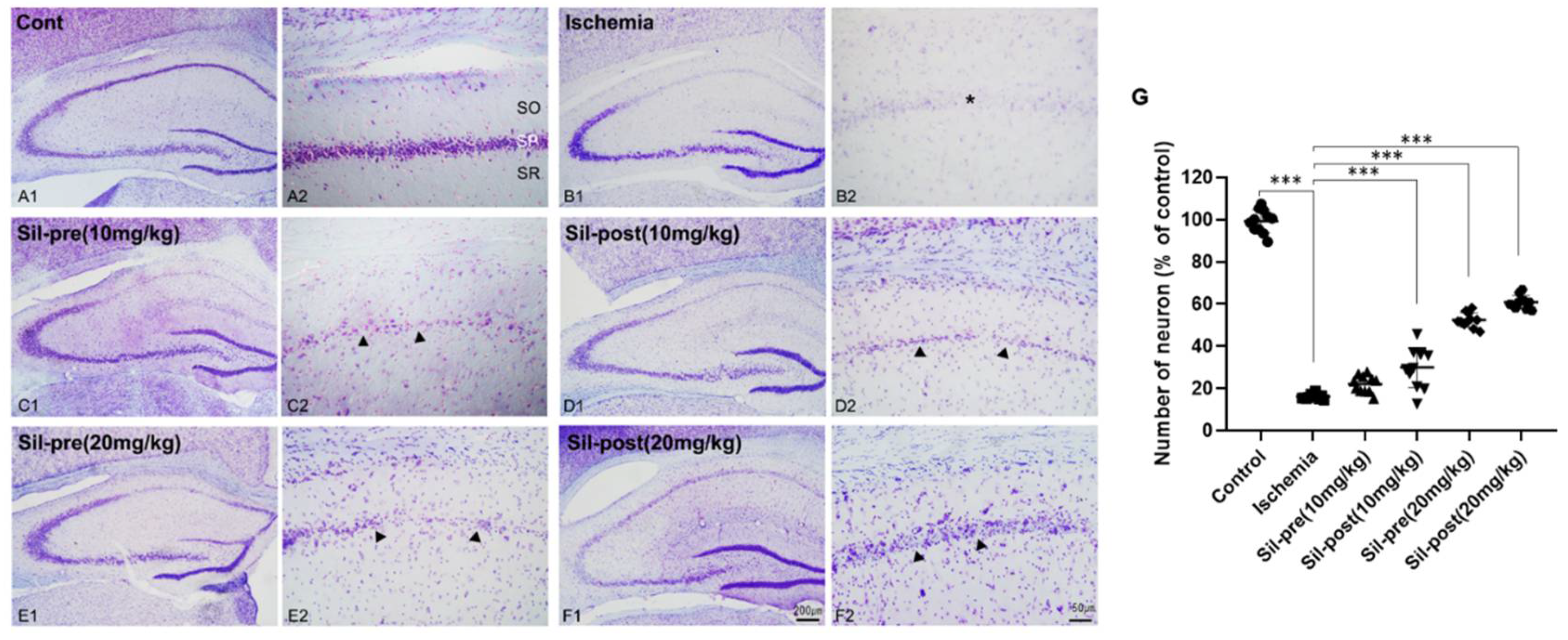
Figure 2.
Sildenafil treatment decreased AQP4 and GFAP co-expression in the CA1 region compared to transient global cerebral ischemia. The co-expression of AQP4 and GFAP was significantly increased in ischemic gerbils compared to control gerbils (A1-B4 and G, arrow). Co-expression was significantly decreased in sildenafil-treated gerbils compared to ischemic gerbils (C1-F4 and G, arrow). Scale bar = 20μm. Data are presented as means ± standard errors of the mean. ***p < 0.001 by one-way analysis of variance.
Figure 2.
Sildenafil treatment decreased AQP4 and GFAP co-expression in the CA1 region compared to transient global cerebral ischemia. The co-expression of AQP4 and GFAP was significantly increased in ischemic gerbils compared to control gerbils (A1-B4 and G, arrow). Co-expression was significantly decreased in sildenafil-treated gerbils compared to ischemic gerbils (C1-F4 and G, arrow). Scale bar = 20μm. Data are presented as means ± standard errors of the mean. ***p < 0.001 by one-way analysis of variance.
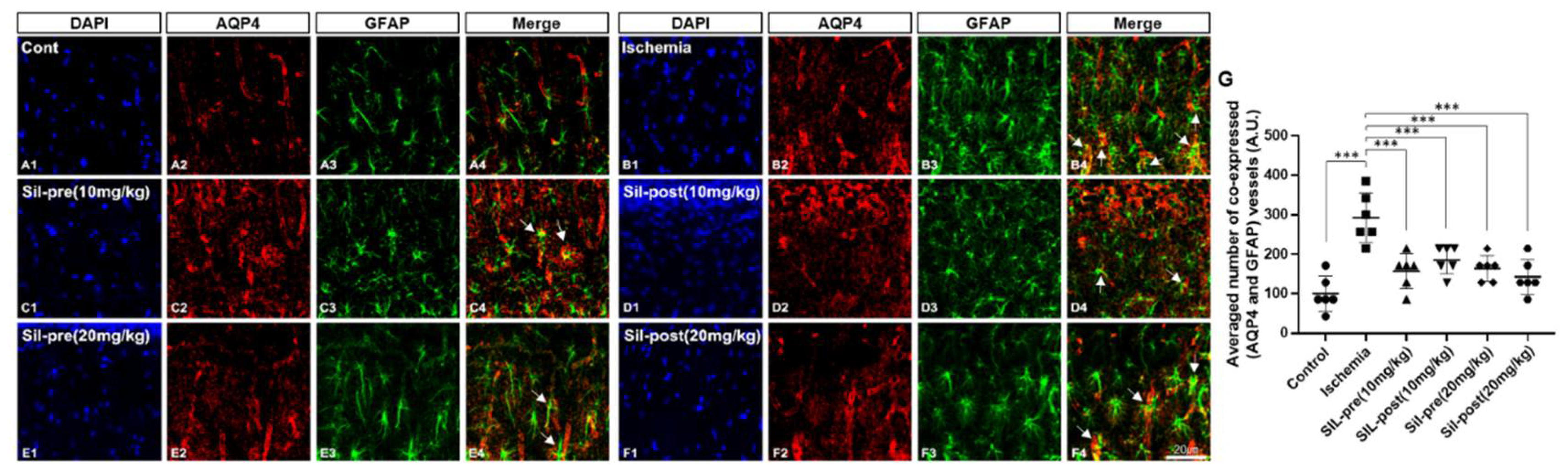
Figure 3.
Sildenafil treatment restored generalized locomotor activity in transient global cerebral ischemia. Representative cumulative traces of navigation pathways in the control, ischemic, SIL-pre, and SIL-post groups during exploration in the open-field test (A). The total distance moved by ischemic gerbils was increased compared to control gerbils (B). The total distance moved by sildenafil-treated gerbils was significantly reduced compared with ischemic gerbils, except for SIL-pre (10mg/kg) gerbils (B). Data are presented as means ± standard errors of the mean. *p < 0.05 by one-way analysis of variance.
Figure 3.
Sildenafil treatment restored generalized locomotor activity in transient global cerebral ischemia. Representative cumulative traces of navigation pathways in the control, ischemic, SIL-pre, and SIL-post groups during exploration in the open-field test (A). The total distance moved by ischemic gerbils was increased compared to control gerbils (B). The total distance moved by sildenafil-treated gerbils was significantly reduced compared with ischemic gerbils, except for SIL-pre (10mg/kg) gerbils (B). Data are presented as means ± standard errors of the mean. *p < 0.05 by one-way analysis of variance.
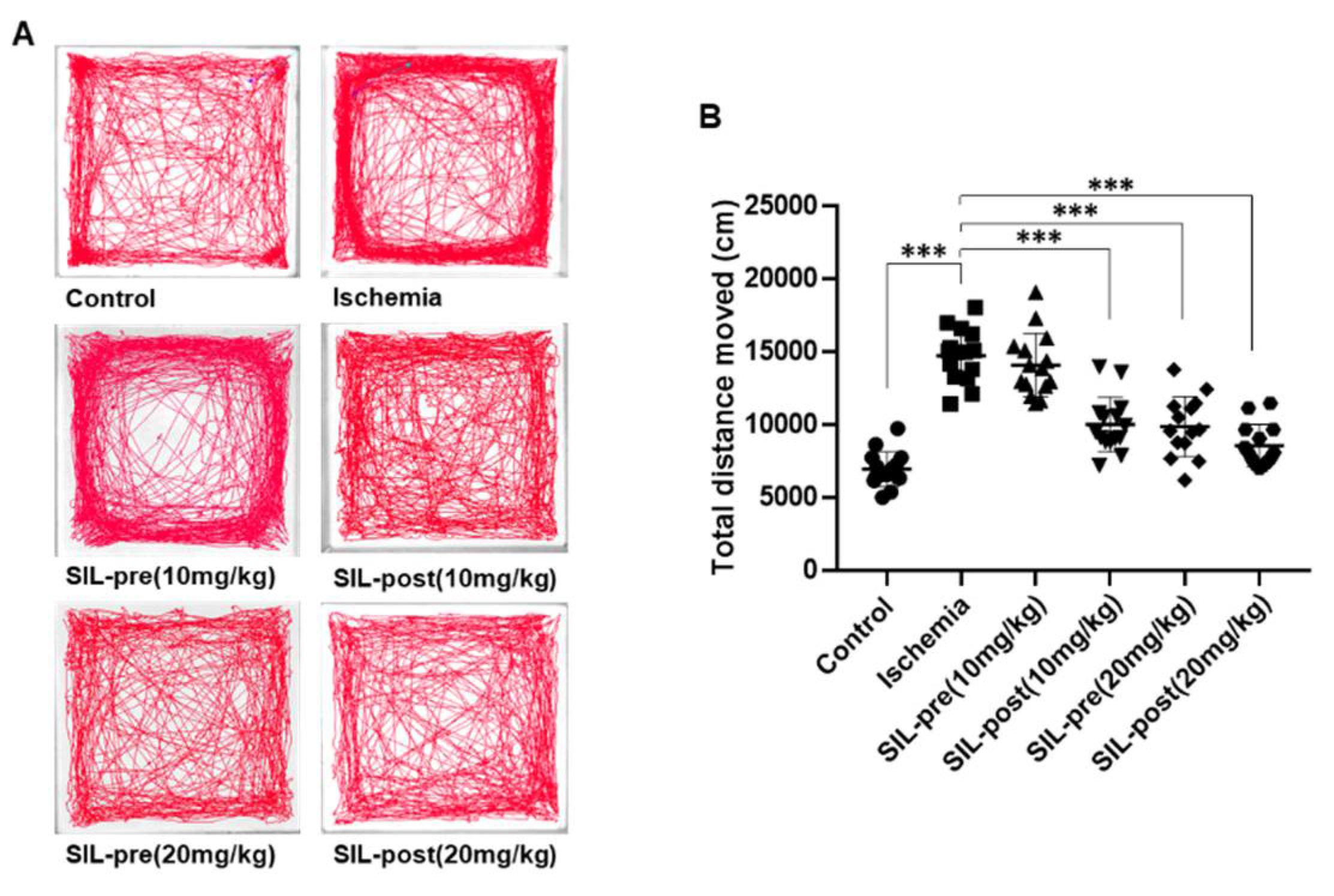
Figure 4.
Sildenafil treatment alleviated novel object recognition deficits induced by transient global cerebral ischemia. In the habituation phase, the frequency percentage of searching for the same two objects was similar in the control, ischemic, and sildenafil-treated groups (A1). The percentage of time spent on object exploration was similar in the control, ischemic, and sildenafil-treated groups (A2). In the test phase, the frequency and percentage of time spent exploring novel objects were decreased in ischemic gerbils compared with control gerbils (B1, B2). However, both the frequency and percentage of time spent exploring novel objects in sildenafil-treated gerbils were significantly increased as compared to ischemic gerbils (B1, B2). In particular, the discrimination index of the sildenafil-treated groups was notably different compared with the ischemic group (B3). Data are presented as means ± standard errors of the mean. ***p < 0.001 by two-way ANOVA analysis of variance.
Figure 4.
Sildenafil treatment alleviated novel object recognition deficits induced by transient global cerebral ischemia. In the habituation phase, the frequency percentage of searching for the same two objects was similar in the control, ischemic, and sildenafil-treated groups (A1). The percentage of time spent on object exploration was similar in the control, ischemic, and sildenafil-treated groups (A2). In the test phase, the frequency and percentage of time spent exploring novel objects were decreased in ischemic gerbils compared with control gerbils (B1, B2). However, both the frequency and percentage of time spent exploring novel objects in sildenafil-treated gerbils were significantly increased as compared to ischemic gerbils (B1, B2). In particular, the discrimination index of the sildenafil-treated groups was notably different compared with the ischemic group (B3). Data are presented as means ± standard errors of the mean. ***p < 0.001 by two-way ANOVA analysis of variance.
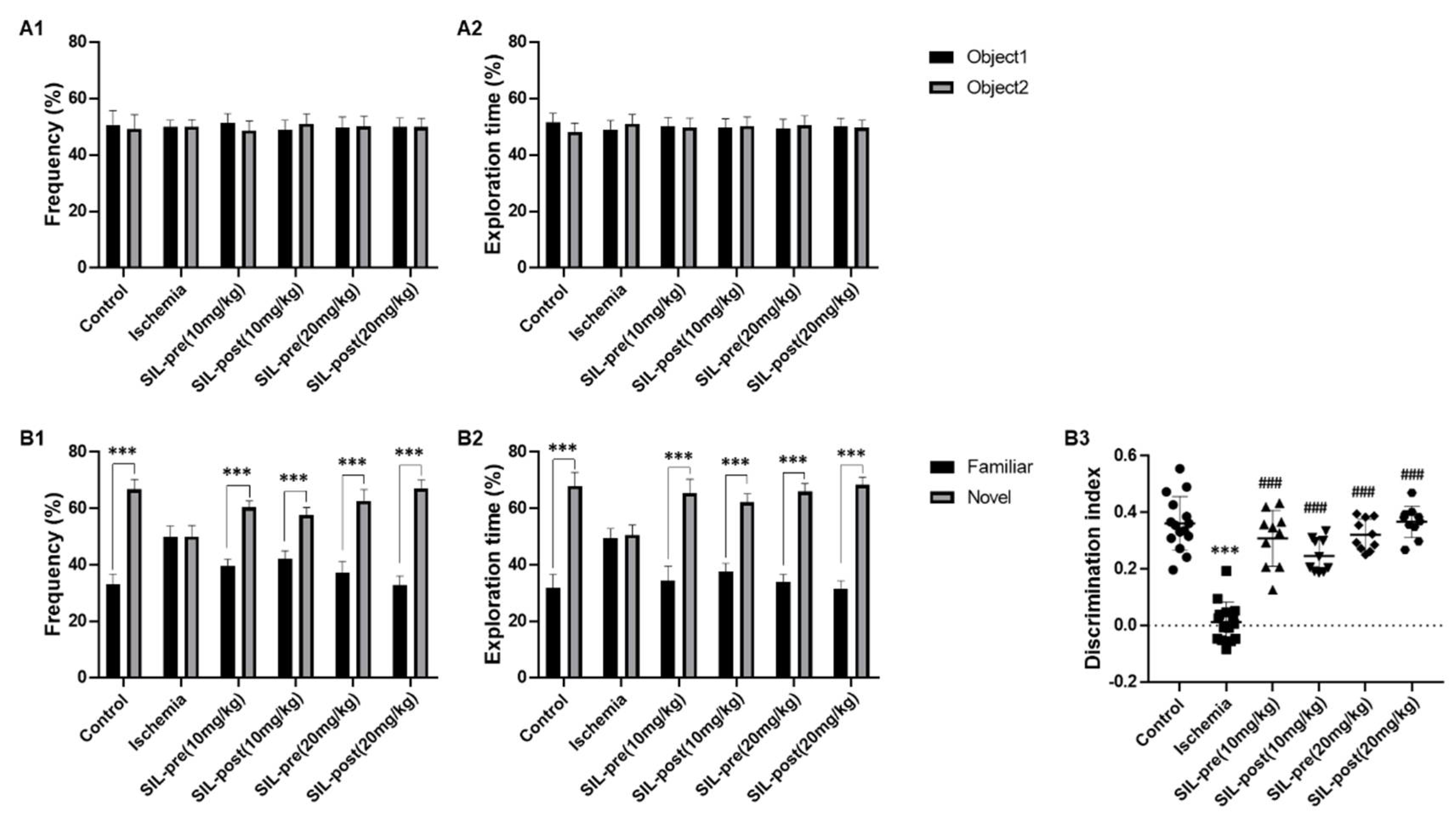
Figure 5.
Sildenafil treatment improved long-term memory, spatial learning, and memory impairment caused by transient global cerebral ischemia. In the Barnes maze test conducted for 5 days, the number of errors in all groups decreased (A). On the probe day, the number of errors made by ischemic gerbils increased significantly compared to control gerbils (B). The number of errors was significantly reduced on the probe test day in all sildenafil-treated groups as compared to ischemic gerbils (A and B). The latency time to the escape hole decreased in all groups over 5 days (C). On the probe day, the latency time to the escape hole was significantly increased in ischemic gerbils compared to control gerbils (D). In all sildenafil-treated groups, the latency time to the escape hole was significantly reduced compared to the ischemic group (C and D). Data are presented as means ± standard errors of the mean. * control group vs. ischemic group. # ischemic group vs. SIL-pre (10 mg/kg), $ ischemic group vs. SIL-post (10 mg/kg), + ischemic group vs. SIL-pre (20 mg/kg), ¶ ischemic group vs. SIL-post (20 mg/kg) (A and C). *p < 0.05, ***p < 0.001 by one-way analysis of variance.
Figure 5.
Sildenafil treatment improved long-term memory, spatial learning, and memory impairment caused by transient global cerebral ischemia. In the Barnes maze test conducted for 5 days, the number of errors in all groups decreased (A). On the probe day, the number of errors made by ischemic gerbils increased significantly compared to control gerbils (B). The number of errors was significantly reduced on the probe test day in all sildenafil-treated groups as compared to ischemic gerbils (A and B). The latency time to the escape hole decreased in all groups over 5 days (C). On the probe day, the latency time to the escape hole was significantly increased in ischemic gerbils compared to control gerbils (D). In all sildenafil-treated groups, the latency time to the escape hole was significantly reduced compared to the ischemic group (C and D). Data are presented as means ± standard errors of the mean. * control group vs. ischemic group. # ischemic group vs. SIL-pre (10 mg/kg), $ ischemic group vs. SIL-post (10 mg/kg), + ischemic group vs. SIL-pre (20 mg/kg), ¶ ischemic group vs. SIL-post (20 mg/kg) (A and C). *p < 0.05, ***p < 0.001 by one-way analysis of variance.
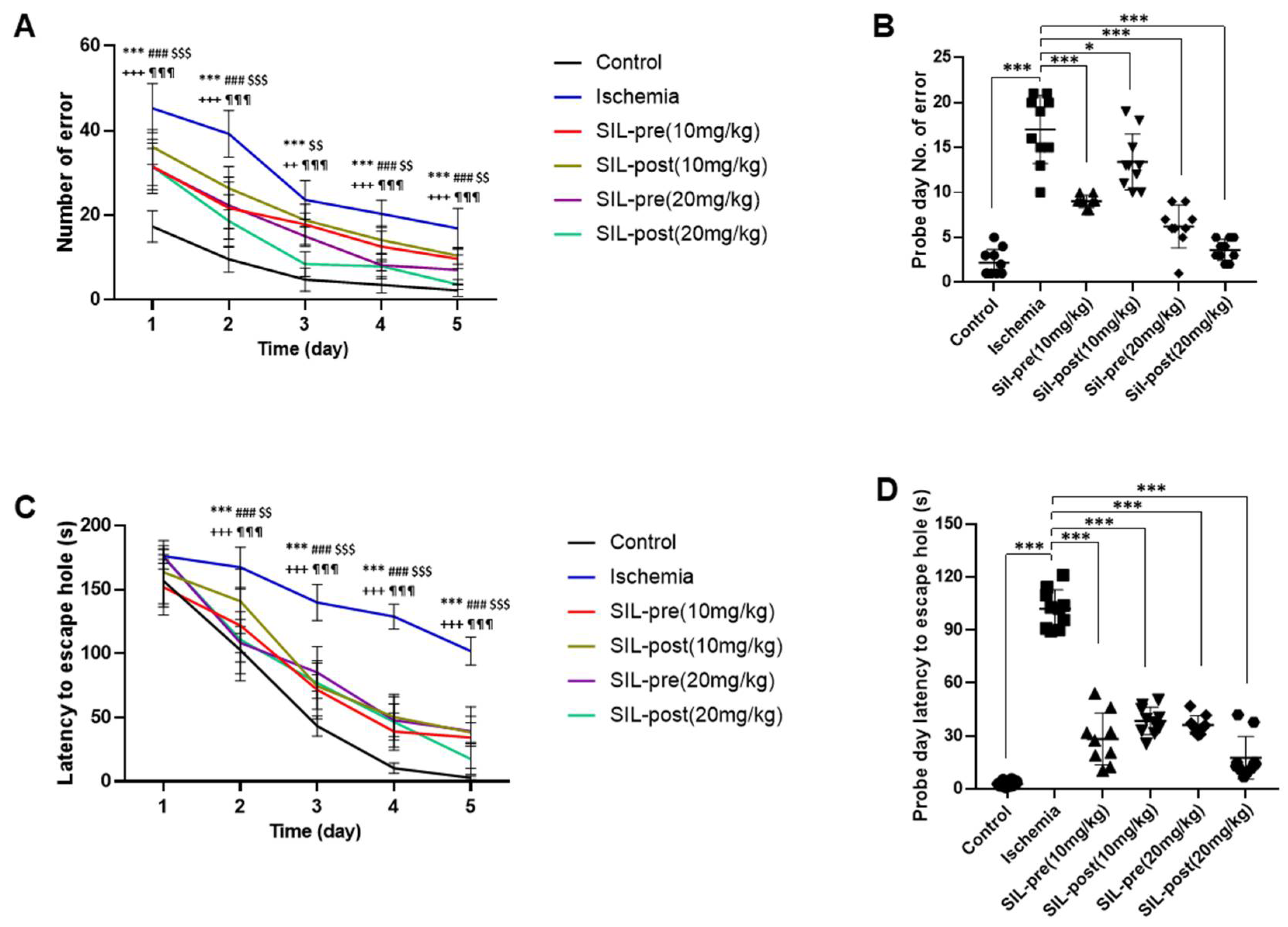
Figure 6.
Sildenafil treatment alleviated the spatial and emotional memory deficits induced by transient global cerebral ischemia. A schematic diagram showing special cues in the working memory test (A). The average percentage of spontaneous alternations in ischemic gerbils was noticeably reduced compared with control gerbils (B). However, in the sildenafil-treated (20 mg/kg) groups, spontaneous alternations were significantly increased compared to ischemic gerbils (B). A schematic diagram showing the passive avoidance test (C). The transfer time were significantly decreased in the ischemic group compared to the control group (D). In contrast, the latency time in all sildenafil-treated groups was increased compared to the ischemic group (D). Data are presented as means ± standard errors of the mean. **p < 0.01, ***p < 0.001 by one-way analysis of variance.
Figure 6.
Sildenafil treatment alleviated the spatial and emotional memory deficits induced by transient global cerebral ischemia. A schematic diagram showing special cues in the working memory test (A). The average percentage of spontaneous alternations in ischemic gerbils was noticeably reduced compared with control gerbils (B). However, in the sildenafil-treated (20 mg/kg) groups, spontaneous alternations were significantly increased compared to ischemic gerbils (B). A schematic diagram showing the passive avoidance test (C). The transfer time were significantly decreased in the ischemic group compared to the control group (D). In contrast, the latency time in all sildenafil-treated groups was increased compared to the ischemic group (D). Data are presented as means ± standard errors of the mean. **p < 0.01, ***p < 0.001 by one-way analysis of variance.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



