
Submitted:
18 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Trichoderma has been widely studied for its potential as a biocontrol agent against plant pathogenic organisms. Trichoderma's biological control mechanisms include competition, modification of environmental conditions, antibiosis, induction of plant defensive mechanisms, and mycoparasitism. Trichoderma species are known to produce a variety of secondary metabolites that have antifungal activity. These metabolites include peptaibols, gliotoxin, and trichokonins. Trichoderma also produces chitinases and β-1,3-glucanases that can degrade the cell walls of fungal pathogens. In addition to direct antagonism against fungal pathogens, Trichoderma can also induce systemic or localised resistance in plants, which is achieved through the production of elicitors such as chitin oligosaccharides and β-glucans that activate plant defence responses. Trichoderma can also form mutualistic associations with plants. In these associations, Trichoderma colonises the roots of plants and promotes plant growth by increasing nutrient uptake and inducing systemic resistance. Using Trichoderma as a biocontrol agent has several advantages over conventional crop protection techniques based on applying synthetic pesticides.
Keywords:
Trichoderma
; fungi
; pathogen
; mode of action
; plant diseases
; biocontrol
; microbe-associated molecular pattern (MAMPs)
1. Introduction
Plant diseases are one of the most critical factors impacting food production, significantly lowering physical and economic output [1,2,3]. The pathogenic microorganisms known to cause plant diseases include bacteria, fungi, viruses, and nematodes, which can cause mild to severe physiological and morphological conditions [4]. These microbial agents are incredibly damaging and can cause widespread plant disease [2].
Plant diseases are responsible for more than 30% of crop losses, resulting in global economic problems [5,6,7]. Despite the arrays of synthetic chemical interventions provided, plant disease control that significantly improves agricultural productivity and ensures consumer food security remains a challenge [8]. Pesticides are chemical compounds used in various agricultural (agronomic) activities to eradicate or limit the spread of pests, insects, weeds, and pathogenic microorganisms [9]. Pesticides such as fungicides, bactericides, and nematicides control agricultural pathogenic microorganisms such as fungi, bacteria, and nematodes [10]. Tina et al. [11] and Pooja et al. [4] submitted that resistance to pesticides, natural microbiota modification or eradication, distortion of the natural habitats of plants, soil contamination, and plant bioaccumulation of hazardous chemicals are some of the negative impacts recorded due to the use of conventional pesticides. As a result, reliance on the use of synthetic pesticides for plant disease management must be discouraged.
Nonetheless, plant disease control has seen significant progress with the employment of other integrated plant disease management (IPDM) strategies, which involve the applications of physical, cultural, biotechnological, and biological control agents [12]. Some biological control agents (BCAs) (fungi and bacteria) have been identified as essential biocontrol agents for plant disease control because they are environmentally safe and may be cost-efficient [13,14,15]. Among the identified BCAs is Trichoderma, a soilborne fungus that has proven effective in controlling pathogenic microorganisms in plant tissues and rhizosphere using various mechanisms [16,17,18]. They are highly tolerant of soil contaminants, alter the rhizosphere to improve plant performance, use available soil micronutrients, maintain efficiency in severe conditions, and promote plant growth [19].
However, Trichoderma is under-exploited for managing plant diseases, especially in Africa, due to a lack of innovation strategies, financial constraints, technological limitations, and technical assistance [12], despite its efficient control level of plant pathogens [20]. Therefore, there is a need to explore Trichoderma's mechanisms of action against plant pathogenic microorganisms to enhance both the researchers' and farmers' knowledge [21].
2. Trichoderma as a Biocontrol Agent
Biological control is the process by which one or more organisms reduce the density of inoculums or the disease-causing activities of a pathogen or parasite in a dynamic or static state. This can be done naturally or by changing the environment, the host, or the antagonist [22].
Microorganisms have a long history of being used as biocontrol agents to control plant diseases caused by pathogenic fungi, bacteria, and nematodes. Biofungicides are natural, non-toxic and ecologically friendly compounds [2]. According to history, farmers' traditional and cultural values were linked to preparing and using plant-protection crops to boost food production before the advent and widespread of synthetic fungicides.
Synthetic fungicides have long been employed to control plant diseases caused by plant pathogenic fungi, even though this practice has been criticised for various reasons [23]. The overuse and incorrect management of synthetic fungicides can harm humans, the environment, and non-target creatures, ultimately leading to resistance and declining biodiversity [24,25,26] and negatively impacting the ozone layer [27]. In addition to ozone layer depletion, component molecules of synthetic fungicides have been associated, in either ingestion or exposure scenarios, with chronic human, animal, and aquatic biodiversity disorders due to their low and prolonged biodegradability and high tendency to persist in the environment [28].
However, advances in research and technology have helped improve safe plant disease management strategies by introducing Plant-incorporated protectants (PIPs), biochemical pesticides, and biofungicides [2]. Biofungicides are safer for human and animal health and have a lower environmental impact than synthetic fungicides, making them suitable for organic farming. Biofungicides can also lower the risk of pathogen resistance by acting against the pathogen in various ways, both directly and indirectly [12,28].
Trichoderma species is one of the most dominant species of soil fungi used as BCAs [29]. Due to their tremendous reproductive potential, ability to thrive in severe conditions, capacity to enhance plant growth, and ability to parasitise phytopathogens, Trichoderma are used as bioagents [30]. Trichoderma wards off pathogenic microfungi through mycoparasitism, antibiosis, resource competition, induction of resistance in plants, and the synthesis of cell wall-degrading enzymes such as glucanases and chitinases [31,32].
3. Mechanism of Action of Trichoderma
Trichoderma uses various biocontrol strategies, alone or combined, to inhibit plant diseases explicitly or implicitly. It is widely known that indirect processes cause morphological and biochemical changes in the host plant, which boost root development and increase systemic resistance, increasing plant tolerance to stressors [33]. The antagonist releases lytic enzymes, antibiotics, and several other toxic compounds during direct processes to combat diseases, which work to antagonise the pathogen. Biocontrol agents (BCAs) show most of these abilities to control the target disease effectively [34]. The following paragraphs provide a complete description of Trichoderma's strategies to defend against plant pathogens.
3.1. Mycoparasitism
In mycoparasitism, fungal plant pathogens are inhibited by beneficial fungi possessing biocontrol ability. The parasitising fungus (bio-agent) gains entry into the tissue of the plant pathogenic fungus using the hyphae and then releases metabolic substances, which result in structural compromise and nutrient absorption from the host (pathogenic) fungus [35]. Oyesola et al. [15] reported the in vitro efficacy of Trichoderma koningii (biocontrol agent) in inhibiting the mycelia growth of Aspergillus niger, A. nidulans, and A. flavus isolated from rotting tomato fruits, which suggested the possible application of Trichoderma in managing post-harvest diseases caused by pathogenic fungi. Several other researchers have also reported that Trichoderma have been effectively employed for the control of fungal plant pathogens due to their ability for rapid hyphal elongation, resulting in the overgrowth of the pathogenic fungi hyphae [20,21,22,36]
Trichoderma's ancestral mode of life is depicted by mycoparasitism, according to a comparative study of the genomes of three species of the fungus that are commonly utilised as biocontrol agents: T. virens, T. reesei, and T. atroviride. Trichoderma interacts with plants' roots, colonising them due to the increased quantity of fungal pathogens in the rhizosphere and the generation of nutrient-rich exudates [37]. Accordingly, their mode of operation changed from mycoparasitism to a more generalist plant-related one [35].
In order to initiate the formation of specialised structures and produce enzymes, Trichoderma mycoparasitism depends on several biochemical signals. Trichoderma initiates chemotropic growth by identifying oligochitins in the pathogen's cell wall. Trichoderma hyphae coil around the pathogen's hyphae upon contact, releasing enzymes that break down the cell wall, such as chitinases and β-1,3-glucanases. Trichoderma exploits the pores created by the breakdown of the fungal cell wall to draw nutrients from its meal [31,38,39].
3.2. Mycovirus-Mediated Cross-Protection (MMCP)
Biocontrol fungi employ mycovirus-mediated cross-protection (MMCP) to shield plants against disorders resulting from fungal infections [40]. MMCP is one of the five biocontrol techniques employed by helpful fungi, along with mycoparasitism, antibiosis, induced systemic resistance (ISR), and competition for resources and space with pathogens [41]. Mycoviruses, such as Cryphonectria parasitica hypovirus 1 (CHV1), are transferred from one fungal strain to another during MMCP, which causes the recipient strain to become less virulent. Due to the identical virulence pathways shared by different fungal pathogens, this weakening may result in cross-protection [37,41].
3.3. Synthesis of Cell Wall Degrading Compounds (CWDCs)
The generation of Cell Wall Degrading Compounds (CWDCs) is a crucial aspect of mycoparasitism's final phase [41]. Chitin, 1,6 -glucans, mannan, chitosan, galactomannan, and most fungal cell walls comprise proteins [38]. Trichoderma are well known for their ability to create a suite of extracellular enzymes that hydrolyse essential cell components, such as chitinases [(1,3;1,6)-β-glucanases] and proteases [42]. Chitinases are the lytic enzymes produced by Trichoderma that hydrolyse glycosidic bonds between the C1 and C4 carbons of two different molecules [43]. The glycoside hydrolase, the 18 (GH18) family of fungus chitinases, has 20 to 90 kDa of molecular weight [44]. The presence of genes in encoding chitinase synthesis, a vast range of binding biocontrol active against various fungal phytopathogens, results in Trichoderma chitinolytic activity [41].
Exo-glucanases and endo-glucanases are two glucanase enzymes that destroy the cell walls of pathogenic fungi [38]. When comparing the genetic properties of Trichoderma with other closely related fungi, the genes coding for synthesising their secondary metabolites are likely to be solely associated with the Trichoderma genome [45]. Excessive exposure to the gene encoding β-1,3-glucanase at the early stages of mycoparasitism was discovered during the expression of T. harzianum and Fusarium solani transcriptome interaction [38]. The capacity to regulate bio-T. virens by Pythium ultimum is also reduced when the Tv bgn-3-gene encoding the enzyme β-1,6-glucanase is removed [46]. Trichoderma soluble in fluid cultures having a wall of lyophilised cells of the pathogenic Fusarium culmorum strain as the only carbon source has been reported to have increased chitinolytic and glucanolytic activity [47]. Trichoderma displays intense enzymatic activity in chitin and the S-cell wall [41]. In fungal diseases, this shows increased hydrolytic enzyme synthesis [48,49].
Proteolytic enzymes enhance the hydrolysis of peptide bonds in proteins and play a significant part in mycoparasitism [50]. The Prb1 protease bound by Trichoderma was demonstrated to play an important role in biodiversity regulation. For example, cotton seedlings exposed to the T. virens gene, tvsp1, recorded enhanced resistance against fungi pathogens by 15-32% compared to the wild type. T. harzianum's aspartic protease considerably lowers the incidence of fruit grey fungus that inhibits seed germination and growth of the plant [51]. Proteases generated by the Trichoderma gene are hypothesised to control the expression or activity of other hydrolytic enzymes implicated in mycoparasitism [51].
3.4. Production of Antibiotics and Other Antifungal Compounds
Antibiosis is how biocontrol agents produce antimicrobial substances inhibiting phytopathogen growth and reproduction. Antibiosis was observed in Trichoderma, which synthesises a variety of antagonistic compounds that break down the cell wall, e.g. pectinase, xylanase, cellulase, lipase, amylase, glucanase and protease, as well as other flexible metabolites or volatile compounds (VOCs) like antibiotics such as trichodermin, gliotoxin, viridin, peptaibols [46]. These and several other secondary metabolites have been isolated and characterised, with over 370 belonging to different groups of potent antagonists [41,52].
Many Trichoderma species produce these VOCs, with peptaibols and polyketides being the most prevalent [53]. Trichoderma is rich in peptaibols, with over 80% stored in the "Peptaibiotics Database" [54]. Peptaibols are polypeptide antibiotics with molecular weights ranging from 500 to 2200 Da with high protein-free amino acids, particularly isovaline and alpha-aminoisobutyric acid. They are identified by acylated N-terminal and amino alcohols in the residual C-terminal end [55,56]. These antibiotics are produced in numerous Trichoderma strains. For example, T. longibrachiatum produces a variety of new peptaibols, most of which are related to trichobrachins, suzukacillins, trichoaureocins, and longibrachins [57]. Acylated N-termini and amino alcohols in the residual C-terminal region distinguish these biochemicals [58]. In Trichoderma, non-ribosomal peptide synthetases (NRPSs) generate peptaibols [45]. These peptaibols are produced in Trichoderma species such as T. citrinoviride, T. longibrachiatum, T. koningi and T. pseudokoningi, which produce Trichorzins, harzianins, trichotoxin, and trichokindins. These have also been implicated in T. harzianum [58]. T. atroviride also produces peptaibols like atroviridins A–C and neoatroviridins A–D, whereas T. viride produces trichotoxins A and B, trichodecenin, trichocellins and trichorovins [38].
Trichoderma produces polyketides (PKs), a class of architecturally varied biochemical compounds found in bacteria, fungi, and plants [59,60]. Polyketide synthases (PKSs) make PKs from basic units like acetyl-CoA and malonyl-CoA [61]. However, Trichoderma is implicated in high PKs encoding genes and polyketide biosynthesis; they have received minimal research attention. Most PKSs are found in fumonisin and represent orthologs in T. reesei, T. virens, and T. atroviride [62,63]. In addition, the T. reesei gene PKs 4 is essential for the composite cell wall's green colour and stability and its resistance to other fungi [64,65].
Secondary metabolites generated by Trichoderma include anthraquinones, pyrones, epipolythiodioxopiperazines (ETPs) and terpenoids [65]. Tetracyclic diterpenes, sesquiterpenes and triterpene viridin are among the terpenoids in Trichoderma [66]. T. asperellum generates 6-phenyl-a-pyrone (6PAP), effectively controlling Fusarium oxysporum [66]. Importantly, T. harzianum, T. aureoviride and T. viride produce anthraquinone pigment compounds such as pachybasin and emodin, which have high antifungal activities and mediate mycoparasitic fungi [21,67,68].
Gliotoxin is the most well-known ETP with immunosuppressive properties, and its antifungal activity also plays an essential role in biological control [69,70]. In addition, Trichoderma, particularly T. harzianum, synthesises harzianic acid with several properties, including antifungal, plant growth enhancing, and irrigation abilities [47,71].
3.5. Competition for Vital Nutrients and Space
Trichoderma possess effective strategies for colonising plants. Trichoderma competes for space and nutrition with pathogenic microorganisms by invading their natural habitats, such as the plant tissues, rhizospheres, or the phyllosphere [72,73]. Their rapid growth pattern significantly enhances their competition with plant pathogens for space and nutrition by producing biochemical substances to eliminate pathogens [74,75]. However, sugar and sucrose levels are high in Trichoderma [40] and are utilised as a carbon source [34]. Trichoderma has superior competition and nutrient absorption ability compared to other fungi genera [46]. This process is linked to gluconic, citric, and fumaric acid production, which reduces soil pH and increases phosphate and microelement (iron, manganese, and magnesium) synthesis [76].
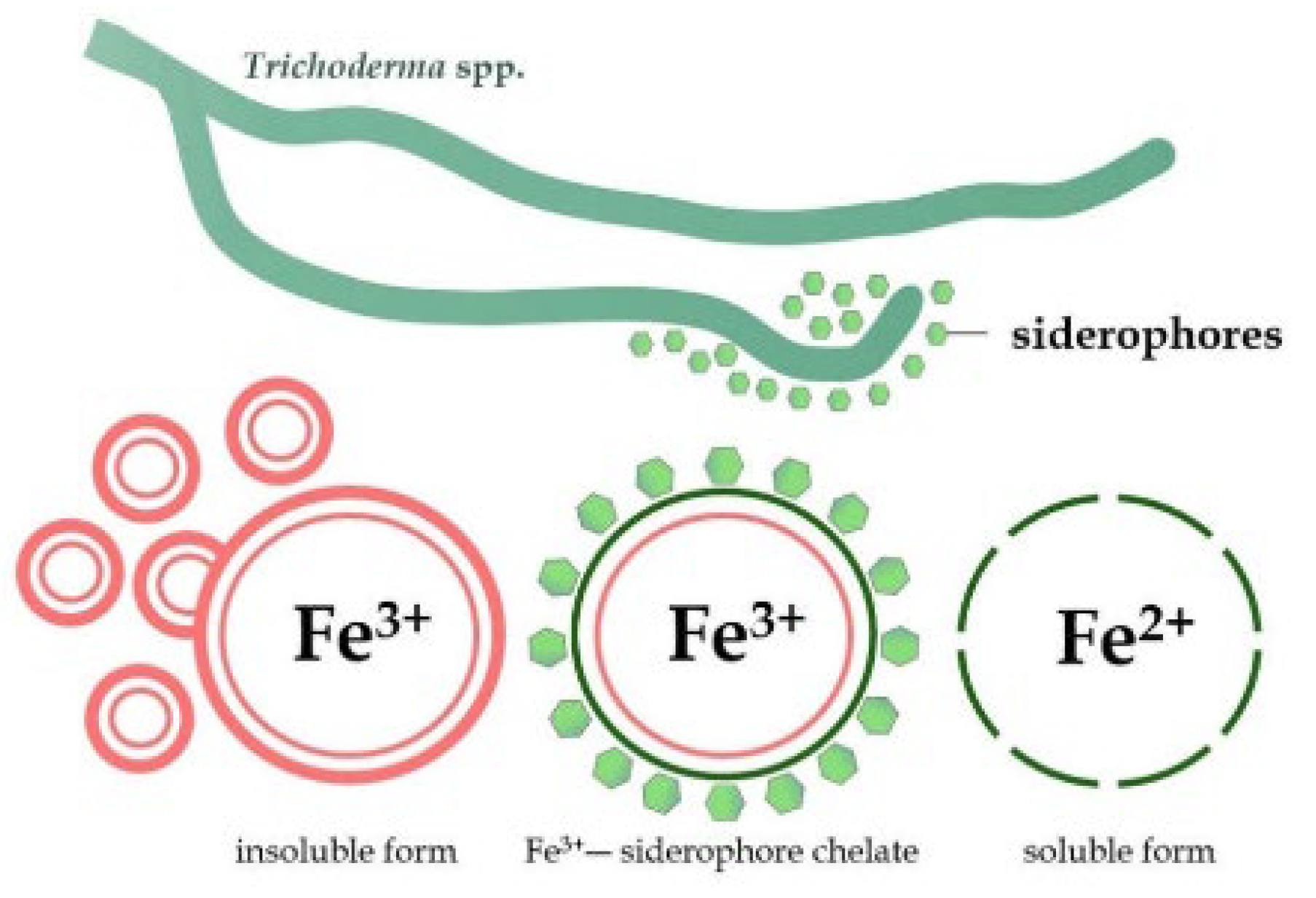
Siderophores is a low molecular weight (less than 10 KDa) chelator with high iron affinity (Fe). It is formed under the stress of iron deficit and significantly impacts the competition process in Trichoderma [77,78]. Fe ions, a cofactor, aid the development and growth of many fungal pathogens [79]. Microbial siderophores' chemical composition and iron-binding characteristics are commonly separated into catecholate, hydroxamate and carboxylate [80]. Coprogens, ferrichrome, and fusarinines are hydroxamate siderophores widely found in fungi and have a standard architectural arrangement of N5-acyl-N5-hydroxyornithine [81,82]. Figure 1 explains how siderophores bind to Fe3+ and convert it to Fe2+, a soluble form that plants and microbes may easily absorb [83].
Trichoderma inhibits the mycelia growth of soil pathogens by depleting iron from their natural sources [84,85]. A report from an in vitro experiment carried out by Oyesola et al. [2] showed that T. koninggi inhibited the mycelia growth of Aspergillus niger, A. flavus and A. nidulans, while T. harzianum controlled the growth of Fusarium acuminatum, Alternaria infectoria and Alternaria alternata [86]. In addition, one of the significant abilities of Trichoderma as a biocontrol agent is metal ion absorption [34]. Trichoderma has also been shown to compete effectively with complex carbon substrates and phytopathogens such as Colletotrichum, Botrytis, Verticillium, and Phytophthora. Trichoderma's decisive inhibitory action in Botrytis cinerea, Macrophomina phaseolina and Fusarium graminearum pathogens was also attributed to enzymes-mediate nutrient competition and environment [87,88].
3.6. Induction of Plant Resistance
The evolution of specific pattern recognition receptors to identify microbial-based signals called microbe-associated molecular patterns (MAMPs) or plant-associated molecular patterns (PAMPs) induced during pathogen invasion are examples of plant resistance molecules [41,89].
Trichoderma, a rhizosphere fungus, interacts with plant roots to induce resistance against diseases in the host plants. They indirectly induce plants' local or systemic immune systems for resistance against pathogens [38,59]. Plant resistance develops because of the activity of multiple elicitors from microbial cells (exoelicitors) and plant tissues (endoelicitors). There are two types of elicitors: race-specific elicitors that only promote gene-to-gene type defence in a variety of host cultivars, and gene-to-gene elicitors produced by pathogenic and non-pathogenic variants that activate non-race specific defences in both the host and pathogen [90,91].
Elicitors generate physical, chemical, and biological changes in plants, such as ion flow and the formation of reactive oxygen species (ROS), which operate as a barrier to limit phytopathogen spread and the production of numerous immunological chemicals such as phytoalexins, enzymes, and phytohormones [92,93]. In the presence of the non-pathogenic fungus Trichoderma, various monocotyledonous and dicotyledonous plant species display increased immunological activity [94]. The recognition of stored domains, such as microbe-related molecular patterns (MAMP) or pathogen-related molecular patterns (PAMP), is the foundation of plant defence [95].
Figure 2.
Plant defence response to the microbe-associated molecular pattern (MAMPs) molecules and effectors. MTI (MAMP-Triggered Immunity): This is activated when the plant detects specific molecules (MAMPs/PAMPs) from the fungus. ETI (Effector-Triggered Immunity): Triggered by the presence of fungal effectors. Intracellular signalling MAPK: Part of the process leading to defence response activation [96].
Figure 2.
Plant defence response to the microbe-associated molecular pattern (MAMPs) molecules and effectors. MTI (MAMP-Triggered Immunity): This is activated when the plant detects specific molecules (MAMPs/PAMPs) from the fungus. ETI (Effector-Triggered Immunity): Triggered by the presence of fungal effectors. Intracellular signalling MAPK: Part of the process leading to defence response activation [96].
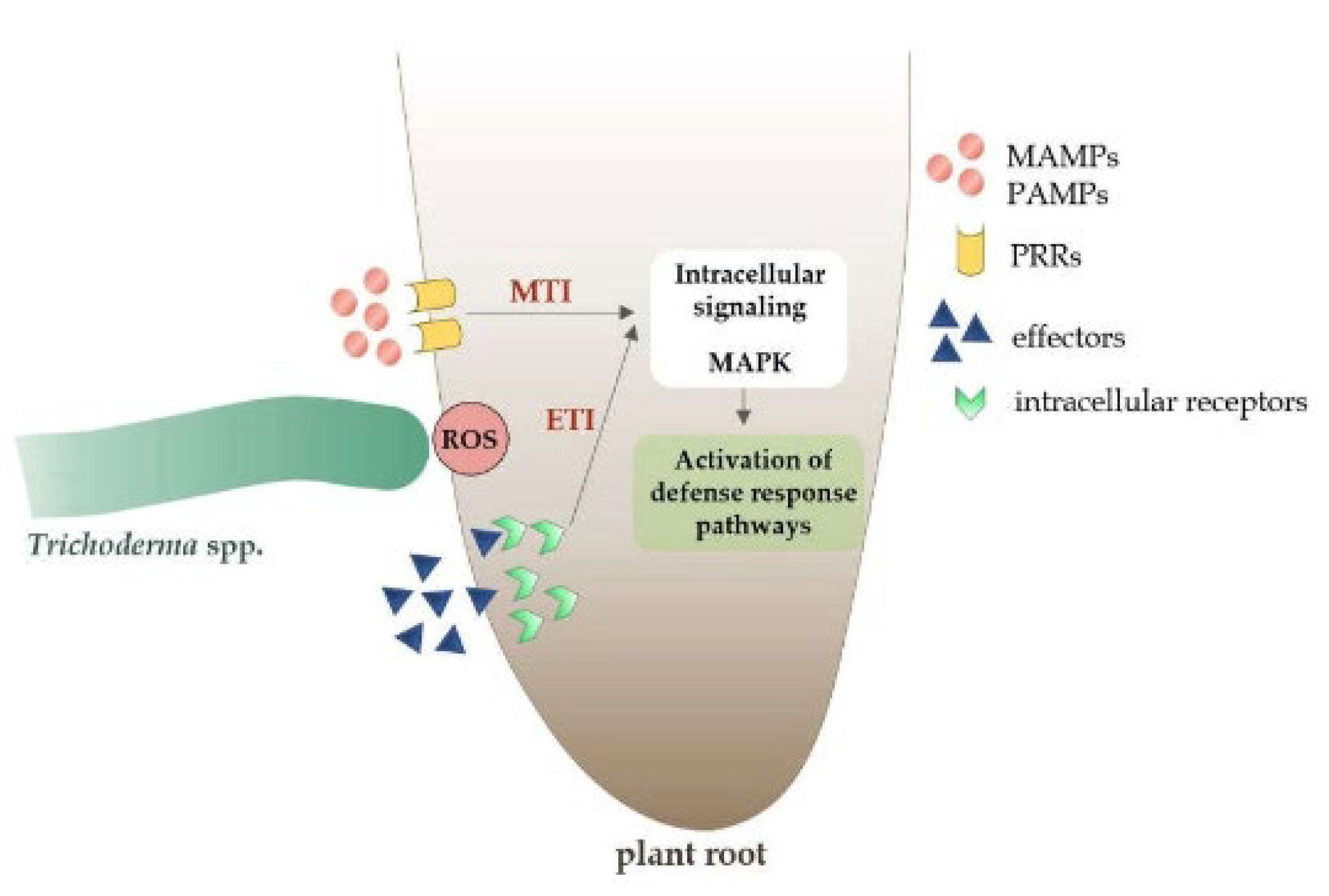
MAMP/PAMP and effector-triggered immunity (ETI) molecules are detected by both transmembrane pattern recognition receptors (PRRs) and intracellular receptors (IR), resulting in MTI/PTI (the microbe-associated molecular pattern (MAMPs)-triggered immunity/pattern-triggered immunity) or local ETI diseases, as depicted in Figure 3 [96]. An ETI-type immune response is usually more effective than MTI/PTI and results in systemic cell death due to hypersensitivity response (HR) activation [97]. Mitogen-activated protein kinases (MAPKs) convey information from receptors to plant cells, triggering a chain of immunological reactions throughout the body [97].
Trichoderma activation of the MTI and ETI immunological pathways results in systemic acquired resistance (SAR) against biotrophic diseases, induced systemic resistance (ISR) against necrotrophic diseases, and induced resistance (IR), which is an efficient defence against biotrophic and necrotrophic diseases and some abiotic stress factors [98].
The expression of pathogenesis-associated protein components (PR) and the generation of salicylic acid (SA) as a signalling molecule describe SAR [94]. Next, significant chemicals targeting the ISR type include jasmonic acid (JA) and ethylene (ET). Aminobutyric acid activates the IR defence, which uses abscisic acid (ABA) as a signalling molecule [99].
Increased enzymes and metabolites indicate the stimulation of signalling systems for the plant's immune response due to Trichoderma elicitor action. Introducing Trichoderma into host plants significantly boosts the activity of phenylalanine, tyrosinelyase, catalase, guaiacol peroxidase, glucanase and chitinase [40]. In a study, Capsicum annum treated with T. harzianum and T. asperellum species showed antifungal abilities to control fungal disease caused by Colletotrichum truncatum [100,101]. In another study, T. longibrachiatum was linked to cucumber plant resistance to B. cinerea challenge through signalling pathways related to plant hormones [102].
Trichoderma's role in efficiently controlling various diseases of plants is made possible by their different mechanisms of action against disease-causing organisms [103].
3.7. Plant Growth Promotion
The capacity of biocontrol agents (BCAs) to stimulate plant growth makes them beneficial to plants. Due to their inherent compatibility, plant growth-promoting fungi, such as Trichoderma, can provide this advantage through their endophytic interactions with the plant roots [104,105]. Trichoderma are rhizospheric organisms that are prominent in boosting plant growth [106]. Trichoderma use several methods to promote plant development; these include.
3.7.1. Mineral Solubilisation
Many microorganisms, particularly those connected to roots, can boost the development and yield of plants. It has been proposed in a few instances that this impact entails the solubilisation of mineral nutrients that would otherwise be inaccessible. The soil's microflora could determine the intricate dynamic balance of solubilisation and insolubilisation that macro- and micronutrients go through in soil, determining how easily plant roots can absorb the nutrients [107].
Thus, it is well known that microbial interactions with plant roots significantly impact the nutritional condition of plants and their resistance to pathogens [108]. Trichoderma species are among the most researched biocontrol microorganisms because they enhance plant development [109].
One of the earliest in vitro studies to investigate the mineral solubilisation ability of Trichoderma was conducted by Altomare et al. [107]. The result of the study revealed that Trichoderma harzianum solubilised Rock phosphate, MnO2, Zn, and Fe2O3 by chelation and reduction, which play a significant role in the biocontrol of plant pathogens. Li et al. [110] investigated the relationship between Trichoderma harzianum and tomato plant roots for phosphate and micronutrient solubilisation. The result of the experiment showed the capability of the Trichoderma strain (SQR-T037) to solubilise phytate, Fe2O3, CuO, and metallic Zn. In other experiments by Tandon et al., Bononi et al., Boat Bedine et al., and Song et al., [111,112,113,114] Trichoderma displayed the capability to solubilise sparingly soluble and insoluble soil micronutrients, which are of immense benefits to the economic plants with effect on biomass accumulation.
3.7.2. Biological Nitrogen Fixation
The biochemical process known as biological nitrogen fixation (BNF) involves particular bacteria (diazotrophs) that have the nitrogenase enzyme converting atmospheric N2 into ammonia. Nitrogen fixation is carried out by several "free-living," "associative," or "symbiotic" diazotrophs [115,116]. Trichoderma enhances photosynthetic carbon fixation capacity, increases nitrogen usage efficiency, and accelerates nitrogen absorption, assimilation, and buildup, significantly affecting plant development and protection against diseases [117].
3.7.3. Phytohormone Level Control
Plant development and defence stimulation are closely linked to Trichoderma's role in synthesising phytohormone [106].
Trichoderma and phytohormones have a complex relationship essential for improving plant health and preventing diseases. The phytohormones produced by these fungi include auxin (indole-3-acetic acid: IAA), gibberellins (GA), abscisic acid (ABA), salicylic acid (SA), and cytokinins (CK). Auxin seems to be especially important in promoting plant development in conjunction with Trichoderma [118].
The enzyme 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase in Trichoderma is significant because it modulates the plant's ethylene biosynthesis and reduces the effects of ethylene-induced stress [119,120]. Trichoderma increases plant resistance to diseases caused by fungi, bacteria, and viruses by modulating phytohormone levels [121]. Applying Trichoderma cultures has been linked to higher plant biomass accumulation, excellent water stress resistance, and improved antioxidant machinery. These fungi also cause systemic resistance, activating defensive systems in plants such as the hypersensitive response (HR) and pathogenesis-related (PR) proteins. Trichoderma's capacity to control phytohormones plays a significant role in plant disease management, making it an essential microfungi for crop management and sustainable agriculture [122].
3.7.4. Modification of Plant's Environmental Conditions
Trichoderma modifies soil pH to generate an environment that helps plants defend against diseases. Trichoderma lowers the pH of the soil by solubilising minerals and releasing organic acids, which inhibits the growth of pathogens and increases the availability of nutrients for plants. Further impeding the growth of pathogens, this acidic environment also initiates the synthesis of antibiotic chemicals [123]. Additionally, plant growth-promoting actions strengthen plant defence systems, whereas Trichoderma's mycoparasitic activity and resource competition weaken pathogens. This makes plants more disease-resistant and improves the soil ecosystem's balance, lessening the need for artificial fungicides and encouraging sustainable agricultural methods [124]. In addition, Trichoderma's capacity to alter the soil pH enhances soil structure, retains more water, and harbours beneficial microbes, all of which contribute to creating a soil environment that promotes plant protection and development [125]. We can create novel, environmentally friendly methods of managing plant diseases by utilising Trichoderma, which will help to ensure a more sustainable future for both the environment and agriculture.
4. Prospects of Trichoderma in Plant Disease Control
The prospect of Trichoderma is vast and multifaceted, holding great promise for sustainable agriculture and disease management, as this versatile fungus employs a range of modes to protect plants and promote their growth, including mycoparasitism, whereby it parasitises and kills plant pathogens, antibiosis, which involves the production of antibiotics to inhibit pathogen growth, competition, where it out-competes pathogens for resources, induced resistance, which triggers plant defence mechanisms, and plant growth promotion, through the production of plant hormones and nutrients [126,127].
These mechanisms collectively create a robust defence system against pathogens, enhance soil fertility and structure, and boost plant productivity and resilience, making Trichoderma an attractive biocontrol agent and plant growth promoter with potential applications in integrated pest management strategies, organic farming, and environmental remediation [128,129].
Further research into its mechanisms of action is likely to uncover even more innovative and effective ways to harness its benefits, such as developing novel formulations and delivery systems, elucidating its role in soil microbiome engineering, and exploring its potential in mitigating the impacts of climate change on agriculture, ultimately leading to a more sustainable food future and a reduced reliance on chemical pesticides.
5. Conclusions
Trichoderma, which has several modes of action via which it exerts biological control against fungi pathogens, is a potential biocontrol agent. The mode of control mechanisms includes competition for resources and space, alteration of environmental factors, antibiosis, activation of plant defence systems, mycoparasitism, and phytohormone synthesis modulation. These mechanisms collectively create a robust defence system against plant pathogens, enhance soil fertility and structure, and promote sustainable agricultural practices. As a biocontrol agent, Trichoderma offers several benefits over the conventional crop protection methods that rely on synthetic pesticides. It relies heavily on generating spore-based biopesticides through fermentation procedures and formulation development. However, more study is required to maximise its effectiveness as a biocontrol agent.
Author Contributions
Conceptualisation, O.L.O. and O.O.O.; Manuscript preparation, O.L.O.; Supervision, O.O.O. and R.T.K.; Funding Acquisition, O.L.O. and O.O.O.; Project Administration, O.O.O. and R.T.K.; Review and Proofreading, B.O.A. and T.O.S. All authors have read and agreed to the published version of the manuscript.
Funding
This project was funded by the Covenant University Center for Research, Innovation and Discovery (CUCRID), with grant number CSG./0116/24/01.
Acknowledgments
The authors acknowledged the Covenant University Center for Research, Innovation and Discovery (CUCRID) for the publication support of this project.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Abade, A.; Ferreira, P.A.; de Barros Vidal, F. Plant diseases recognition on images using convolutional neural networks: A systematic review. Computers and Electronics in Agriculture 2021, 185, 106125. [Google Scholar] [CrossRef]
- Oyesola, O.L.; Aworunse, O.S.; Oniha, M.I.; Obiazikwor, O.H.; Bello, O.; Atolagbe, O.M.; Obembe, O.O. Impact and Management of Diseases of Solanum tuberosum. IntechOpen 2021. [Google Scholar] [CrossRef]
- Ashwini, T.; Srinivas, A.; Mallikarjuna, G. CRISPR-Cas genome editing system: A versatile tool for developing disease resistant crops. Plant Stress 2022, 3, 100056. [Google Scholar] [CrossRef]
- Pooja, C.; Pooja, R.A.; Sumi, R.; Radhakrishnan, N.; Mehanathan, M. Molecular and metabolomic interventions for identifying potential bioactive molecules to mitigate diseases and their impacts on crop plants. Physiological and Molecular Plant Pathology 2021, 114, 101624. [Google Scholar] [CrossRef]
- Savary, S.; Willocquet, L.; Pethybridge, S.J.; Esker, P.; McRoberts, N.; Nelson, A. The global burden of pathogens and pests on major food crops. Natural Ecology and Evolution 2019, 3, 430–439. [Google Scholar] [CrossRef]
- van Esse, R.; van der, D.; van Esse, T.L.; Reuber, D. Genetic modification to improve disease resistance in crops. New Phytology 2020, 225, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases - A review. Ecological Complexity 2022, 49, 100978. [Google Scholar] [CrossRef]
- Menka, T.; Debasish, P.; Reecha, M.; Binod, B.S.; Prashant, S. The Impact of Microbes in Plant Immunity and Priming Induced Inheritance: A Sustainable Approach for Crop protection. Plant Stress 2022, 4, 100072. [Google Scholar] [CrossRef]
- Hassaan, M.A.; El Nemr, A. Pesticides pollution: Classifications, human health impact, extraction and treatment techniques. The Egyptian Journal of Aquatic Research 2020, 46, 207–220. [Google Scholar] [CrossRef]
- Saheem, R.; Tanveer, R.; Khalid, M.G. A review of interactions of pesticides within various interfaces of intrinsic and organic residue amended soil environment. Chemical Engineering Journal Advances 2022, 11, 100301. [Google Scholar] [CrossRef]
- Tina, R.; Anuradha, B.; Parshuram, J.S.; Sukanta, M.; Nitish, R.M.; Shariful, A.; Nirmalendu, D. Bio-effective disease control and plant growth promotion in lentil by two pesticide degrading strains of Bacillus sp. Biological Control 2018, 127, 55–63. [Google Scholar] [CrossRef]
- Ons, L.; Bylemans, D.; Thevissen, K.; Cammue, B. Combining biocontrol agents with chemical fungicides for integrated plant fungal disease control. Microorganisms 2020, 8, 1930. [Google Scholar] [CrossRef] [PubMed]
- Figlan, S.; Ntushelo, K.; Mwadzingeni, L.; Terefe, T.; Tsilo, T.J.; Shimelis, H. Breeding Wheat for Durable Leaf Rust Resistance in Southern Africa: Variability, Distribution, Current Control Strategies, Challenges, and Future Prospects. Frontier of Plant Science 2020, 11, 549. [Google Scholar] [CrossRef]
- Olowe, O.M.; Lidia, N.; Michael, D.A.; Akinlolu, O.A.; Olubukola, O.B. Trichoderma: Potential bio-resource for the management of tomato root rot diseases in Africa. Microbiological Research 2022, 257, 126978. [Google Scholar] [CrossRef]
- Oyesola, O.L.; Sobowale, A.A.; Obembe, O.O. Effectiveness of Trichoderma koningii Extract on Aspergillus Species Isolated from Rotting Tomato (Solanum lycopersicum Mill). Tropical Journal of Natural Products and Research 2020, 4, 961–965. [Google Scholar] [CrossRef]
- Kuzmanovska, B.; Rusevski, R.; Jankulovska, M.; Oreshkovikj, K. Antagonistic activity of Trichoderma asperellum and Trichoderma harzianum against genetically diverse Botrytis cinerea isolates. Chilean Journal of Agricultural Research 2018, 78, 391–399. [Google Scholar] [CrossRef]
- Filizola, P.R.B.; Luna, M.A.C.; de Souza, A.F.; Coelho, I.L.; Laranjeira, D.; Campos–Takaki, G.M. Biodiversity and phylogeny of novel Trichoderma isolates from mangrove sediments and potential of biocontrol against Fusarium strains. Microbial Cell Fact 2019, 18, 89. [Google Scholar] [CrossRef]
- Manandhar, S.; Pant, B.; Manandhar, C.; Baidya, S. In-vitro evaluation of biocontrol agents against soil borne plant pathogens. J. Nep. Agric. Res. Counc 2019, 5, 68–72. [Google Scholar] [CrossRef]
- Kohl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Frontier of Plant Science 2019, 10, 845. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; Kumar, A.; de Los Santos-Villalobos, S.; Parra-Cota, F.I.; Orozco-Mosqueda, M.D.C.; Fadiji, A.E.; Hyder, S.; Babalola, O.O.; Santoyo, G. Trichoderma Species: Our Best Fungal Allies in the Biocontrol of Plant Diseases-A Review. Plants (Basel, Switzerland) 2023, 12, 432. [Google Scholar] [CrossRef]
- Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Jaroszuk-Ściseł, J. Trichoderma: The Current Status of Its Application in Agriculture for the Biocontrol of Fungal Phytopathogens and Stimulation of Plant Growth. International journal of molecular sciences 2022, 23, 2329. [Google Scholar] [CrossRef]
- Asad, S.A. Mechanisms of action and biocontrol potential of Trichoderma against fungal plant diseases - A review. Ecological Complexity 2022, 49, 100978. [Google Scholar] [CrossRef]
- Deresa, E.M.; Diriba, T.F. Phytochemicals as alternative fungicides for controlling plant diseases: A comprehensive review of their efficacy, commercial representatives, advantages, challenges for adoption, and possible solutions. Heliyon 2023, 9, e13810. [Google Scholar] [CrossRef]
- Grovermann, C.; Schreinemachers, P.; Berger, T. Quantifying pesticide overuse from farmer and societal points of view: An application to Thailand. Crop Protection 2013, 53, 161–168. [Google Scholar] [CrossRef]
- Srinivasan, R.; Tamò, M.; Subramanian, S. The case for integrated pest management in Africa: Transition from a pesticide-based approach. Current Opinion in Insect Science 2022, 54, 100970. [Google Scholar] [CrossRef]
- Pimentão, A.R.; Cuco, A.P.; Pascoal, C.; Cássio, F.; Castro, B.B. Current trends and mismatches on fungicide use and assessment of the ecological effects in freshwater ecosystems. Environmental Pollution 2024, 347, 123678. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; He, Y.; Jiang, M.; You, Q.; Ma, X.; Xu, Z.; Bo, X. Unveiling the importance of VOCs from pesticides applicated in main crops for elevating ozone concentrations in China. Journal of Hazardous Materials 2024, 465, 133385. [Google Scholar] [CrossRef]
- Anuagasi, C.L.; Okigbo, R.N.; Anukwuorji, C.A.; Okereke, C.N. The Impact of Biofungicides on Agricultural Yields and Food Security in Africa. International Journal of Agricultural Technology 2017, 13, 953–978. [Google Scholar]
- Patrick, B.; Paul, E.; Gerardina, U. Incorporation of microorganisms to reduce chemical fungicide usage in black sigatoka control programs in Costa Rica by use of biological fungicides. Crop Protection 2021, 146, 105657. [Google Scholar] [CrossRef]
- Manzar, N.; Kashyap, A.S.; Goutam, R.S.; Rajawat, M.V.S.; Sharma, P.K.; Sharma, S.K.; Singh, H.V. Trichoderma: Advent of Versatile Biocontrol Agent, Its Secrets and Insights into Mechanism of Biocontrol Potential. Sustainability. 2022, 14, 12786. [Google Scholar] [CrossRef]
- Guzmán-Guzmán, P.; María, D.P.; Vianey, O.; Alfredo, H. Trichoderma species: versatile plant symbionts. Phytopathology 2019, 109, 6–16. [Google Scholar] [CrossRef] [PubMed]
- Fraceto, L.F.; Maruyama, C.R.; Guilger, M.; Mishra, S.; Keswani, C.; Singh, H.B.; de Lima, R. Trichoderma harzianum-based novel formulations: potential applications for management of Next-Gen agricultural challenges. Journal of Chemical Technology & Biotechnology 2018, 93, 2056–2063. [Google Scholar]
- Poveda, J. Trichoderma as biocontrol agent against pests: New uses for a mycoparasite. Biological Control 2021, 159, 104634. [Google Scholar] [CrossRef]
- Rauf, A.; Subhani, M.N.; Siddique, M.; Shahid, H.; Chattha, M.B.; Alrefaei, A.F.; Hasan Naqvi, S.A.; Ali, H.; Lucas, R.S. Cultivating a greener future: Exploiting trichoderma derived secondary metabolites for fusarium wilt management in peas. Heliyon 2024, 10, e29031. [Google Scholar] [CrossRef] [PubMed]
- Prasun, K.; Mukherjee, A.M.; Susanne, Z.; Benjamin, A.H. Mycoparasitism as a mechanism of Trichoderma-mediated suppression of plant diseases. Fungal Biology Reviews 2022, 39, 15–33. [Google Scholar] [CrossRef]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Frontiers of Microbiology 2023, 14, 1160551. [Google Scholar] [CrossRef] [PubMed]
- Hough, B.; Steenkamp, E.; Wingfield, B.; Read, D. Fungal Viruses Unveiled: A Comprehensive Review of Mycoviruses. Viruses 2023, 15, 1202. [Google Scholar] [CrossRef] [PubMed]
- Sood, M.; Kapoor, D.; Kumar, V.; Sheteiwy, M.S.; Ramakrishnan, M.; Landi, M.; Araniti, F.; Sharma, A. Trichoderma: The "secrets" of a multitalented biocontrol agent. Plants 2020, 9, 762. [Google Scholar] [CrossRef]
- Macías-Rodríguez, L.; Contreras-Cornejo, H.A.; Adame-Garnica, S.G.; Del-Val, E.; Larsen, J. The interactions of Trichoderma at multiple trophic levels: Inter-kingdom communication. Microbiological Research 2020, 240, 126552. [Google Scholar] [CrossRef]
- Pandit, M.A.; Kumar, J.; Gulati, S.; Bhandari, N.; Mehta, P.; Katyal, R.; Rawat, C.D.; Mishra, V.; Kaur, J. Major Biological Control Strategies for Plant Pathogens. Pathogens 2022, 11. [Google Scholar] [CrossRef]
- Ghorbanpour, M.; Omidvari, M.; Abbaszadeh-Dahaji, P.; Omidvar, R.; Kariman, K. Mechanisms underlying the protective effects of beneficial fungi against plant diseases. Biological Control 2018, 117, 147–157. [Google Scholar] [CrossRef]
- Ribeiro, M.S.; Voltan, A.R.; Carraro, C.B.; Steindorff, A.S.; Goldman, G.H.; Silva, R.N.; Ulhoa, C.J.; Monteiro, V.N. Endo-β-1,3-glucanase (GH16 Family) from Trichoderma harzianum Participates in Cell Wall Biogenesis but Is Not Essential for Antagonism Against Plant Pathogens. Biomolecules 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Loc, N.H.; Huy, N.D.; Quang, H.T.; Lan, T.T.; Ha, T.T.T. Characterization and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology 2020, 11, 38–48. [Google Scholar] [CrossRef] [PubMed]
- Goughenour, K.D.; Whalin, J.; Slot, J.C.; Rappleye, C.A. Diversification of Fungal Chitinases and Their Functional Differentiation in Histoplasma capsulatum. Molecular Biology and Evolution 2021, 38, 1339–1355. [Google Scholar] [CrossRef] [PubMed]
- Vicente, I.; Baroncelli, R.; Hermosa, R.; Monte, E.; Vannacci, G.; Sarrocco, S. Role and genetic basis of specialised secondary metabolites in Trichoderma ecophysiology. Fungal Biology Reviews 2022, 39, 83–99. [Google Scholar] [CrossRef]
- Volkov, P.; Rubtsova, E.; Rozhkova, A.; Sinitsyna, O.; Zorov, I.; Kondratyeva, E.; Sinitsyn, A. Properties of recombinant endo-β-1,6-glucanase from Trichoderma harzianum and its application in the pustulan hydrolysis. Carbohydrate Research 2020, 499, 108211. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Guo, H.; Zhang, K.; Zhao, M.; Ruan, J.; Chen, J. Trichoderma and its role in biological control of plant fungal and nematode disease. Frontiers of Microbiology 2023, 14, 1160551. [Google Scholar] [CrossRef] [PubMed]
- Gueye, N.; Kumar, G.K.; Ndiaye, M.; Sall, S.Y.D.; Ndiaye, M.A.F.; Diop, T.A.; Ram, M.R. Factors affecting the chitinase activity of Trichoderma asperellum isolated from agriculture field soils. Joural of Applied Biology and Biotechnology. 2020, 8, 41–44. [Google Scholar] [CrossRef]
- Olasehinde, G.I.; Okolie, Z.V.; Oniha, M.I.; Adekeye, B.T.; Ajayi, A.A. In vitro antibacterial and antifungal activities of Chrysophyllum albidum and Diospyros monbuttensis leaves. Journal of Pharmacognosy and Phytotherapy 2016, 8, 1–7. [Google Scholar] [CrossRef]
- De Zotti, M.; Sella, L.; Bolzonello, A.; Gabbatore, L.; Peggion, C.; Bortolotto, A.; Elmaghraby, I.; Tundo, S.; Favaron, F. Targeted amino acid substitutions in a Trichoderma peptaibol confer activity against fungal plant pathogens and protect host tissues from Botrytis cinerea infection. International Journal Molecular Sciences 2020, 21, 7521. [Google Scholar] [CrossRef]
- Deng, J.; Huang, W.; Li, Z.; Lu, D.; Zhang, Y.; Luo, X. Biocontrol activity of recombinant aspartic protease from Trichoderma harzianum against pathogenic fungi. Enzyme and Microbial Technology 2018, 112, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H.; Hirpara, D.G.; Savaliya, D.D.; Golakiya, B. Extracellular metabolomics of Trichoderma biocontroller for antifungal action to restrain Rhizoctonia solani Kuhn in cotton. Physiological and Molecular Plant Pathology 2020, 112, 101547. [Google Scholar] [CrossRef]
- Xiao, Z.; Zhao, Q.; Li, W.; Gao, L.; Liu, G. Strain improvement of Trichoderma harzianum for enhanced biocontrol capacity: Strategies and prospects. Frontiers in Microbiology 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.; Pablo, J. Peptaibol Production and Characterization from Trichoderma asperellum and Their Action as Biofungicide. Journal of Fungi 2022, 8, 1037. [Google Scholar] [CrossRef]
- Tamandegani, P.R.; Marik, T.; Zafari, D.; Balázs, D.; Vágvölgyi, C.; Szekeres, A.; Kredics, L. Changes in Peptaibol Production of Trichoderma Species during In Vitro Antagonistic Interactions with Fungal Plant Pathogens. Biomolecules 2020, 10. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Dias, L.; Oliveira-Pinto, P.R.; Fernandes, J.O.; Regalado, L.; Mendes, R.; Teixeira, C.; Mariz-Ponte, N.; Gomes, P.; Santos, C. Peptaibiotics: Harnessing the potential of microbial secondary metabolites for mitigation of plant pathogens. Biotechnology Advances 2023, 68, 108223. [Google Scholar] [CrossRef] [PubMed]
- Marik, T.; Tyagi, C.; Balázs, D.; Urbán, P.; Szepesi, Á.; Bakacsy, L.; Endre, G.; Rakk, D.; Szekeres, A.; Andersson, M.A.; Salonen, H.; Druzhinina, I.S.; Vágvölgyi, C.; Kredics, L. Structural Diversity and Bioactivities of Peptaibol Compounds From the Longibrachiatum Clade of the Filamentous Fungal Genus Trichoderma. Frontiers of Microbiology 2019, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Siddiquee, S. Fungal volatile organic compounds: Emphasis on their plant growth-promoting. In Volatiles and Food Security; Choudhary, D., Sharma, A., Agarwal, P., Varma, A., Tuteja, N., Eds.; Springer: Singapore, 2017; pp. 313–333. [Google Scholar]
- Zeilinger, S.; Gruber, S.; Bansal, R.; Mukherjee, P.K. Secondary metabolism in Trichoderma—Chemistry meets genomics. Fungal Biology Review 2016, 30, 74–90. [Google Scholar] [CrossRef]
- Cardoza, R.E.; McCormick, S.P.; Izquierdo-Bueno, I. Identification of polyketide synthase genes required for aspinolide biosynthesis in Trichoderma arundinaceum. Applied Microbiology and Biotechnology 2022, 106, 7153–7171. [Google Scholar] [CrossRef]
- Mary, L.S.; Russell, J.C. ; Molecular methods unravel the biosynthetic potential of Trichoderma species. RSC Advances 2021, 11, 6–3622. [Google Scholar] [CrossRef]
- Sobowale, A. Probable Effects of Dual Inoculation of Maize (Zea mays) Stem with Fusarium verticillioides and Certain Trichoderma Species on Fumonisin Content of Maize Seeds. American Journal of Plant Sciences 2019, 10, 752–759. [Google Scholar] [CrossRef]
- Deepa, N.; Achar, P.N.; Sreenivasa, M.Y. Current Perspectives of Biocontrol Agents for Management of Fusarium verticillioides and Its Fumonisin in Cereals—A Review. Journal of Fungi 2021, 7, 776. [Google Scholar] [CrossRef] [PubMed]
- Atanasova, L.; Knox, B.P.; Kubicek, C.P.; Druzhinina, I.S.; Baker, S.E. The polyketide synthase gene pks4 of Trichoderma reesei provides pigmentation and stress resistance. Eukaryotic Cell 2013, 12, 1499–1508. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Song, B.; Liu, M.; Qin, L.; Dong, Z. Understanding the Role of Trichoderma reesei Vib1 in Gene Expression during Cellulose Degradation. Journal of Fungi 2021, 7, 613. [Google Scholar] [CrossRef] [PubMed]
- Shenouda, M.L.; Ambilika, M.; Skellam, E.; Cox, R.J. Heterologous Expression of Secondary Metabolite Genes in Trichoderma reesei for Waste Valorization. Journal of Fungi 2022, 8, 355. [Google Scholar] [CrossRef] [PubMed]
- Sehim, A.E.; Hewedy, O.A.; Altammar, K.A.; Alhumaidi, M.S.; Abd Elghaffar, R.Y. Trichoderma asperellum empowers tomato plants and suppresses Fusarium oxysporum through priming responses. Frontiers in microbiology 2023, 14, 1140378. [Google Scholar] [CrossRef]
- Yusuf, M.A.; Singh, B.N.; Sudheer, S.; Kharwar, R.N.; Siddiqui, S.M.A.; Fernandes Fraceto, L.; Dashora, K.; Gupta, V.K. Chrysophanol: A Natural Anthraquinone with Multifaceted Biotherapeutic Potential. Biomolecules 2019, 9, 68. [Google Scholar] [CrossRef] [PubMed]
- Kubiak, A.; Pilarska, A.A.; Niewiadomska, A. Fungi of the Trichoderma Genus: Future Perspectives of Benefits in Sustainable Agriculture. Applied Sciences 2022, 13, 6434. [Google Scholar] [CrossRef]
- Scharf, D.H.; Brakhage, A.A.; Mukherjee, P.K. Gliotoxin--bane or boon? Environmental microbiology 2016, 18, 1096–1109. [Google Scholar] [CrossRef]
- Kumar, V.; Koul, B.; Taak, P.; Yadav, D.; Song, M. Journey of Trichoderma from Pilot Scale to Mass Production: A Review. Agriculture 2023, 13, 2022. [Google Scholar] [CrossRef]
- Zaid, R.; Koren, R.; Kligun, E.; Gupta, R.; Leibman-Markus, M.; Mukherjee, P.K.; Kenerley, C.M.; Bar, M.; Horwitz, B.A. Gliotoxin, an Immunosuppressive Fungal Metabolite, Primes Plant Immunity: Evidence from Trichoderma virens-Tomato Interaction. Microbiology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Halifu, S.; Deng, X.; Song, X.; Song, R. Effects of Two Trichoderma Strains on Plant Growth, Rhizosphere Soil Nutrients, and Fungal Community of Pinus sylvestris var. mongolica Annual Seedlings. Forests 2019, 10, 758. [Google Scholar] [CrossRef]
- Oszust, K.; Cybulska, J.; Frąc, M. How Do Trichoderma Genus Fungi Win a Nutritional Competition Battle against Soft Fruit Pathogens? A Report on Niche Overlap Nutritional Potentiates. International Journal of Molecular Sciences 2019, 21, 4235. [Google Scholar] [CrossRef] [PubMed]
- Martinez, Y.; Ribera, J.; Schwarze, F.W.M.R. Biotechnological development of Trichoderma-based formulations for biological control. Appl Microbiol Biotechnol 2023, 107, 5595–5612. [Google Scholar] [CrossRef] [PubMed]
- Vinale, F.; Marra, R.; Scala, F.; Ghisalberti, E.L.; Lorito, M.; Sivasithamparam, K. Major secondary metabolites produced by two commercial Trichoderma strains active against different phytopathogens. Letter of Applied Microbiology 2006, 43, 143–148. [Google Scholar] [CrossRef]
- Zhao, L.; Wang, Y.; Kong, S. Effects of Trichoderma asperellum and its siderophores on endogenous auxin in Arabidopsis thaliana under iron-deficiency stress. International Microbiology 2020, 23, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, M.; Shafiq, M.; Noman, F.; Abbas, S.A.A.A.; Ali, N. Structure and potential applications of bacterial siderophores. Bacterial Secondary Metabolites 2023, 159–175. [Google Scholar] [CrossRef]
- Robinson, J.R.; Isikhuemhen, O.S.; Anike, F.N. Fungal–Metal Interactions: A Review of Toxicity and Homeostasis. Journal of Fungi 2021, 7. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.K.; Bera, T.; Chakrabarty, A.M. Microbial siderophore – A boon to agricultural sciences. Biological Control 2020, 144, 104214. [Google Scholar] [CrossRef]
- Chowdappa, S.; Jagannath, S.; Konappa, N.; Udayashankar, A.C.; Jogaiah, S. Detection and Characterization of Antibacterial Siderophores Secreted by Endophytic Fungi from Cymbidium aloifolium. Biomolecules 2020, 10, 1412. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, P.; Zeng, G.; Wu, G.; Qi, L.; Chen, G.; Fang, W.; Yin, W.-B. Transcriptional Differences Guided Discovery and Genetic Identification of Coprogen and Dimerumic Acid Siderophores in Metarhizium robertsii. Frontiers of Microbiology 2021, 12, 783609. [Google Scholar] [CrossRef] [PubMed]
- Pecoraro, L.; Wang, X.; Shah, D.; Song, X.; Kumar, V.; Shakoor, A.; Tripathi, K.; Ramteke, P.W.; Rani, R. Biosynthesis Pathways, Transport Mechanisms and Biotechnological Applications of Fungal Siderophores. Journal of Fungi 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Deb, L.; Pandey, A.K. Trichoderma- from lab bench to field application: Looking back over 50 years. Frontiers in Agronomy 2022, 4, 932839. [Google Scholar] [CrossRef]
- Lahlali, R.; Ezrari, S.; Radouane, N.; Kenfaoui, J.; Esmaeel, Q.; Hamss, H.E.; Belabess, Z.; Barka, E.A. Biological Control of Plant Pathogens: A Global Perspective. Microorganisms 2022, 10. [Google Scholar] [CrossRef] [PubMed]
- Patkowska, E.; Mielniczuk, E.; Jamiołkowska, A. The Influence of Trichoderma harzianum Rifai T-22 and Other Biostimulants on Rhizosphere Beneficial Microorganisms of Carrot. Agronomy 2020, 10, 1637. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Wang, M. Isolation and molecular identification of Trichoderma species from wetland soil and their antagonistic activity against phytopathogens. Physiological and Molecular Plant Pathology 2019, 109, 101458. [Google Scholar] [CrossRef]
- López-Coria, M.; Guzmán-Chávez, F.; Carvente-García, R.; Muñoz-Chapul, D.; Sánchez-Sánchez, T.; Arciniega-Ruíz, J.M.; King-Díaz, B.; Sánchez-Nieto, S. Maize plant expresses SWEET transporters differently when interacting with Trichoderma asperellum and Fusarium verticillioides, two fungi with different lifestyles. Frontiers in Plant Science 2023, 14. [Google Scholar] [CrossRef] [PubMed]
- Abdul Malik, N.A.; Kumar, I.S.; Nadarajah, K. Elicitor and Receptor Molecules: Orchestrators of Plant Defense and Immunity. International Journal of Molecular Sciences 2020, 21. [Google Scholar] [CrossRef] [PubMed]
- Zehra, A.; Raytekar, N.A.; Meena, M.; Swapnil, P. Efficiency of microbial bio-agents as elicitors in plant defense mechanism under biotic stress: A review. Current Research in Microbial Sciences 2021, 2. [Google Scholar] [CrossRef]
- Köhl, J.; Kolnaar, R.; Ravensberg, W.J. Mode of Action of Microbial Biological Control Agents Against Plant Diseases: Relevance Beyond Efficacy. Frontiers in Plant Science 2019, 10, 845. [Google Scholar] [CrossRef]
- Aljbory, Z.; Chen, M.S. Indirect plant defense against insect herbivores: A review. Journal of Insect Science 2018, 25, 2–23. [Google Scholar] [CrossRef] [PubMed]
- Juan, C.A.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. International Journal of Molecular Sciences 2021, 22. [Google Scholar] [CrossRef] [PubMed]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species—Opportunistic, avirulent plant symbionts. Nature Review Microbiology 2021, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.; Dangl, J. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Jaroszuk-Ściseł, J.; Tyśkiewicz, R.; Nowak, A.; Ozimek, E.; Majewska, M.; Hanaka, A.; Tyśkiewicz, K.; Pawlik, A.; Janusz, G. Phytohormones (Auxin, Gibberellin) and ACC Deaminase In Vitro Synthesized by the Mycoparasitic Trichoderma DEMTkZ3A0 Strain and Changes in the Level of Auxin and Plant Resistance Markers in Wheat Seedlings Inoculated with this Strain Conidia. International Journal of Molecular Sciences 2019, 20. [Google Scholar] [CrossRef] [PubMed]
- De Schutter, K.; Van Damme, E.J.M. Protein-carbohydrate interactions as part of plant defense and animal immunity. Molecules 2015, 20, 9029–9053. [Google Scholar] [CrossRef] [PubMed]
- Jagodzik, P.; Tajdel-Zielinska, M.; Ciesla, A.; Marczak, M.; Ludwikow, A. Mitogen-activated protein kinase cascades in plant hormone signalling. Frontiers in Plant Science 2018, 9, 1387. [Google Scholar] [CrossRef] [PubMed]
- Boccardo, N.A.; Segretin, M.E.; Hernandez, I.; Mirkin, F.G.; Cha ́con, O.; Lopez, Y.; Borrás-Hidalgo, O.; Bravo-Almonacid, F.F. Expression of pathogenesis-related proteins in transplastomic tobacco plants confers resistance to filamentous pathogens underfield trials. Scientific Reports 2019, 9, 2791. [Google Scholar] [CrossRef] [PubMed]
- Hartman, G.L.; Pawlowski, M.L.; Chang, H.-X.; Hill, C.B. Successful technologies and approaches used to develop and manage resistance against crop diseases and pests. In Woodhead Publishing Series in Food Science, Technology and Nutrition. Emerging Technologies for Promoting Food Security; Madramootoo, C., Ed.; Woodhead Publishing: Sawston, UK 2016. [Google Scholar]
- Kim, S.H.; Lee, Y.; Balaraju, K.; Jeon, Y. Evaluation of Trichoderma atroviride and Trichoderma longibrachiatum as biocontrol agents in controlling red pepper anthracnose in Korea. Frontiers Plant Science 2023, 14, 1201875. [Google Scholar] [CrossRef]
- Yadav, M.; Divyanshu, K.; Dubey, M.K.; Rai, A.; Kumar, S.; Tripathi, Y.N.; Shukla, V.; Upadhyay, R.S. Plant growth promotion and differential expression of defense genes in chilli pepper against Colletotrichum truncatum induced by Trichoderma asperellum and T. Harzianum. BMC Microbiology 2023, 23. [Google Scholar] [CrossRef]
- Yuan, M.; Huang, Y.; Ge, W.; Jia, Z.; Song, S.; Zhang, L.; Huang, Y. Involvement of jasmonic acid, ethylene and salicylic acid signaling pathways behind the systemic resistance induced by Trichoderma longibrachiatumH9 in cucumber. BMC Genomics 2019, 20, 144. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, O.C.; Babalola, O.O. Productivity and quality of horticultural crops through co-inoculation of arbuscular mycorrhizal fungi and plant growth promoting bacteria. Microbiological Research 2020, 239, 126569. [Google Scholar] [CrossRef]
- Ayub, J.; Tahir, A.; Iqbal, U.; Ayub, H.; Hyder, M.Z.; Kiyani, A.; Hafeez, F.Y.; Ilyas, M.K.; Ghafoor, A.; Yasmin, T. Effective biological control of Lentil (Lens culinaris) Fusarium wilt and plant growth promotion through native Rhizobacteria. Physiological and Molecular Plant Pathology 2023, 129, 102203. [Google Scholar] [CrossRef]
- Contreras-Cornejo, H.A.; Schmoll, M.; Esquivel-Ayala, B.A.; González-Esquivel, C.E.; Rocha-Ramírez, V.; Larsen, J. Mechanisms for plant growth promotion activated by Trichoderma in natural and managed terrestrial ecosystems. Microbiological Research 2024, 281, 127621. [Google Scholar] [CrossRef]
- Altomare, C.; Norvell, W.A.; Björkman, T.; Harman, G.E. Solubilization of Phosphates and Micronutrients by the Plant-Growth-Promoting and Biocontrol Fungus Trichoderma harzianum Rifai 1295-22. Applied and Environmental Microbiology 1999, 65, 2926–2933. [Google Scholar] [CrossRef]
- Amoo, A.E.; Olanrewaju, O.S.; Babalola, O.O.; Ajilogba, C.F.; Chukwuneme, C.F.; Ojuederie, O.B.; Omomowo, O.I. The functionality of plant-microbe interactions in disease suppression. Journal of King Saud University – Science 2023, 35, 102893. [Google Scholar] [CrossRef]
- Alshammari, W.; Bairum, R.; Sulieman, A.M.; Alshammari, N.; Elamin, H. In vitro and in vivo Study of Antagonistic and Biocontrol of Trichoderma harzianum Strains Against Wood Decay Pathogens. Polish Journal of Environmental Studies 2024, 33, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Li, R.-X.; Cai, F.; Pang, G.; Shen, Q.-R.; Li, R.; Chen, W. Solubilisation of Phosphate and Micronutrients by Trichoderma harzianum and Its Relationship with the Promotion of Tomato Plant Growth. PLoS ONE 2015, 10, e0130081. [Google Scholar] [CrossRef]
- Tandon, A.; Fatima, T.; Shukla, D.; Tripathi, P.; Srivastava, S.; Singh, P.C. Phosphate solubilisation by Trichoderma koningiopsis (NBRI-PR5) under abiotic stress conditions. Journal of King Saud University – Science 2019, 32, 791–798. [Google Scholar] [CrossRef]
- Bononi, L.; Chiaramonte, J.B.; Pansa, C.C.; Moitinho, M.A.; Melo, I.S. Phosphorus-solubilizing Trichoderma spp. From Amazon soils improve soybean plant growth. Scientific Reports 2020, 10, 1–13. [Google Scholar] [CrossRef]
- Boat Bedine, M.A.; Iacomi, B.; Tchameni, S.N.; Sameza, M.L.; Fekam, F.B. Harnessing the phosphate-solubilising ability of Trichoderma strains to improve plant growth, phosphorus uptake and photosynthetic pigment contents in common bean (Phaseolus vulgaris). Biocatalysis and Agricultural Biotechnology 2022, 45, 102510. [Google Scholar] [CrossRef]
- Song, M.; Wang, X.; Xu, H.; Zhou, X.; Mu, C. Effect of Trichoderma viride on insoluble phosphorus absorption ability and growth of Melilotus officinalis. Scientific Reports 2023, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rascio, N.; La Rocca, N. Biological Nitrogen Fixation. Encyclopedia of Ecology (Second Edition) 2012, 264–279. [Google Scholar] [CrossRef]
- Shirvani, M.; Yahaghi, Z. Role of Pb-solubilizing and plant growth-promoting bacteria in Pb uptake by plants. Advances in Microbe-Assisted Phytoremediation of Polluted Sites 2021, 231–270. [Google Scholar] [CrossRef]
- Mei, H.M.; Ruan, Y.N.; Zhang, J.X.; Cui, J.X.; Yan, K.; Dong, X.Y.; Bian, L.X.; Sun, Y.H. Effects of Trichoderma on nitrogen absorption and use efficiency in Lycium chinense roots under saline stress. Ying yong sheng tai xue bao = The journal of applied ecology 2022, 33, 2539–2546. [Google Scholar] [CrossRef] [PubMed]
- Stewart, A.; Hill, R. Applications of Trichoderma in Plant Growth Promotion. Biotechnology and Biology of Trichoderma 2013, 415–428. [Google Scholar] [CrossRef]
- Rauf, M.; Awais, M.; Ud-Din, A.; Ali, K.; Gul, H.; Rahman, M.M.; Hamayun, M.; Arif, M. Molecular Mechanisms of the 1-Aminocyclopropane-1-Carboxylic Acid (ACC) Deaminase Producing Trichoderma asperellum MAP1 in Enhancing Wheat Tolerance to Waterlogging Stress. Frontiers in plant science 2021, 11, 614971. [Google Scholar] [CrossRef] [PubMed]
- Jha, C.K.; Sharma, P.; Shukla, A.; Parmar, P.; Patel, R.; Goswami, D.; Saraf, M. Microbial enzyme, 1-aminocyclopropane-1-carboxylic acid (ACC) deaminase: An elixir for plant under stress. Physiological and Molecular Plant Pathology 2021, 115, 101664. [Google Scholar] [CrossRef]
- Isabel, M.; Rubén, S. Plant Defensive Responses Triggered by Trichoderma spp. As Tools to Face Stressful Conditions. Horticulturae 2022, 8, 1181. [Google Scholar] [CrossRef]
- Bahadur, A.; Dutta, P. Trichoderma Spp.: Their Impact in Crops Diseases Management. IntechOpen, 2022. [Google Scholar] [CrossRef]
- Esparza-Reynoso, S.; Pelagio-Flores, R.; López-Bucio, J. Mechanism of plant immunity triggered by Trichoderma. New and Future Developments in Microbial Biotechnology and Bioengineering 2019, 57–73. [Google Scholar] [CrossRef]
- Abbas, A.; Mubeen, M.; Zheng, H.; Sohail, M.A.; Shakeel, Q.; Solanki, M.K.; Iftikhar, Y.; Sharma, S.; Kashyap, B.K.; Hussain, S.; Romano, C.Z.; Moya-Elizondo, E.A.; Zhou, L. Trichoderma spp. Genes Involved in the Biocontrol Activity Against Rhizoctonia solani. Frontiers in Microbiology 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Alfiky, A.; Weisskopf, L. Deciphering Trichoderma–Plant–Pathogen Interactions for Better Development of Biocontrol Applications. Journal of Fungi 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Akbari, S.I.; Prismantoro, D.; Permadi, N.; Rossiana, N.; Miranti, M.; Mispan, M.S.; Mohamed, Z.; Doni, F. Bioprospecting the roles of Trichoderma in alleviating plants' drought tolerance: Principles, mechanisms of action, and prospects. Microbiological Research 2024, 283, 127665. [Google Scholar] [CrossRef] [PubMed]
- Barman, S.; Gorai, P.S.; Mandal, N.C. Trichoderma spp.—Application and future prospects in agricultural industry. Recent Advancement in Microbial Biotechnology 2020, 49–70. [Google Scholar] [CrossRef]
- Khan, M.R.; Mohiddin, F.A. Trichoderma: Its Multifarious Utility in Crop Improvement. Crop Improvement Through Microbial Biotechnology 2017, 263–291. [Google Scholar] [CrossRef]
- Kapoor, D.; Sharma, M.M.M.; Yadav, S.; Sharma, P. Applications of Trichoderma virens and biopolymer-based biostimulants in plant growth and productions. Biostimulants in Plant Protection and Performance 2023, 349–367. [Google Scholar] [CrossRef]
Figure 1.
The insoluble iron (Fe3+) transformation into a soluble and easily assimilated (Fe2+) form. Trichoderma releases molecules called siderophores. These siderophores bind to the insoluble iron (Fe³⁺), forming a siderophore chelate complex. The chelate makes the iron soluble, converting it into Fe²⁺ [83].
Figure 1.
The insoluble iron (Fe3+) transformation into a soluble and easily assimilated (Fe2+) form. Trichoderma releases molecules called siderophores. These siderophores bind to the insoluble iron (Fe³⁺), forming a siderophore chelate complex. The chelate makes the iron soluble, converting it into Fe²⁺ [83].
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



