
Submitted:
21 May 2024
Posted:
21 May 2024
You are already at the latest version
Abstract
Pathogens that adapt to environmental stress can develop an increased tolerance to some physical or chemical antimicrobial treatments. The main objective of this study was to determine if acid adaptation increased the tolerance of Escherichia coli O157:H7 to high voltage atmospheric cold plasma (HVACP) in raw pineapple juice. Samples (10 ml) of juice were inoculated with non-acid-adapted (NAA) or acid adapted (AA) E. coli to obtain a viable count of ~ 7.00 log10 CFU/ml. The samples were exposed to HVACP (70 kV) for 1 to 7 min with inoculated non-HVACP-treated juice serving as control. Juice samples were analyzed for survivors at 0.1 h, and after 24 h of refrigeration (4 °C). Samples analyzed after 24 h exhibited significant decreases in viable NAA cells with sub-lethal injury detected in both NAA and AA survivors (P<0.05). No NAA survivor in juice exposed to HVACP for 5 or 7 min was detected after 24h. However, numbers of AA survivors were 3.33 and 3.09 log10 CFU/ml in juice treated for 5 and 7 min, respectively (P<0.05). These results indicate that acid adaptation increases the tolerance of E. coli to HVACP in pineapple juice. The potentially higher tolerance of AA E. coli O157:H7 to HVACP should be considered in developing safe juice processing parameters for this novel non-thermal technology.
Keywords:
Acid adaptation
; Escherichia coli O157:H7
; cold plasma
; pineapple juice
; sub-lethal injury
1. Introduction
The global market for fresh fruit and juices has grown consistently throughout the past two decades. For fresh fruit, the market was valued at 551.1 billion US dollars in 2021 and has a projected compound annual growth rate (CAGR) of 3.6% from 2022 to 2028 [1]. Likewise, the fresh juice market is also growing with a higher forecasted CAGR of 8.55% between 2022 and 2027, and a projected increase in market size to 104.21 billion dollars [2]. Part of the increase in the popularity of fresh fruits and juices is the plethora of health benefits that consumers derive from their consumption [3]. While there is an increasing demand for minimally processed fruits and fruit juices, these nutritious products may contain pathogenic bacteria if they are not properly handled during harvesting or in food processing establishments [4].
Unpasteurized fruit juices can harbor pathogenic bacteria such as E. coli O157:H7 in apple juices and Salmonella spp. in orange juices [5]. Fruits are mainly grown in orchards where they are exposed to many sources of microbial contamination such as water, windblown dust, insects, birds, and feral animals [6,7]. While whole fresh fruits are cleaned and cooled using water baths and dump tanks, these methods can sometime lead to whole fruit being infiltrated by bacteria in the wash water [8,9]. Pathogenic bacteria enter the fruit juices when the contaminated fruit is pressed to release the juices. Another mode of microbial access to fresh juices is via transfer of microorganisms from the surfaces of whole fruit to the flesh during peeling and slicing, or from the processing environment [10]. Enteric pathogens may survive long enough in fruit juice to pose a food safety risk to consumers. For example, E. coli O157:H7 and Salmonella spp. survived in refrigerated (4.4 ᵒC) pineapple juice for over 42 days [11].
Deadly outbreaks of E. coli 0157:H7 in unpasteurized apple juice in the 1990s resulted in the U.S Food and Drug Administration (FDA) issuing a juice HACCP (Hazard Analysis Critical Control Points) regulation. According to that regulation, juice manufacturers must implement a kill step in juice processing that produces in a 5-log (99.999%) reduction of the most pertinent microorganism [5,12]. Thermal pasteurization can achieve that 5-log reduction; however, thermal treatments can destroy heat-sensitive nutrients and bioactive components in juices [13]. Moreover, thermal processing can alter quality attributes of juices, such as color and flavor [14]. Due to these challenges, juice manufacturers developed a keen interest in non-thermal technologies to treat juices. Such technologies include high-pressure processing, pulsed electric fields, ultraviolet light, ultrasound, and cold plasma [15,16,17,18].
Atmospheric cold plasma (ACP) is a novel non-thermal technology that utilizes the fourth state of matter (plasma) to inactivate microorganisms in foods [18,19]. Gaseous plasma consists of a mixture of electrons, positive and negative ions, excited atoms and molecules, gas atoms, free radicals, UV photons and visible light [20]. Those reactive species interact with the oxygen and nitrogen gas in air to form several reactive oxygen and nitrogen species (RONS) such as atomic oxygen, hydroxyl radical (∙OH), nitric oxide (NO), and nitrogen dioxide (NO2). These RONS in plasma are largely responsible for the antimicrobial effect of cold plasma [21,22]. One advantage of ACP is that it can inactivate microorganisms without high temperatures [21]. The antimicrobial efficacy of ACP against foodborne microorganisms is well documented [9,23,24,25]. However, antimicrobial efficacy may be lessened following prior exposure of microorganisms to environmental stress [26].
Foodborne microorganisms inevitably encounter environmental stresses during food production, manufacturing, storage, distribution, and preparation. Pathogens response to stress may cause stress adaptation and an increase pathogen tolerance to single or multiple stresses [27,28]. For example, acid adaptation can enhance survival of pathogens in fruit juices to pose a food safety risk to consumers. More importantly, acid-adaption may cross-protect pathogens against various processing treatments such as heating, and non-thermal treatments [29,30,31,32,33]. The increased tolerance of stress-adapted pathogens to certain non-thermal processing treatments raises concerns about the overestimation of antimicrobial treatments especially when non-stress-adapted organisms are used in process validation studies. Moreover, non-thermal physical treatments, depending on their severity, can cause sub-lethal injury in pathogen survivors [34].
Sub-lethally injured foodborne pathogens pose an insidious problem for food processors. If they are undetected in foods, they may resuscitate under suitable conditions and regain their pathogenicity [35]. Traditionally, the plating of diluted food samples on both selective and non-selective agar has been used to evaluate the extent of sublethal injury in pathogen survivors of an antimicrobial treatment [35,36,37,38]. Selective agar media allow growth of non-injured pathogens but inhibit resuscitation of sub-lethally injured pathogens thus preventing their detection. Nonselective agar permits the enumeration of both non-injured and sub-lethally injured pathogens [35]. In this regard the difference in bacterial colony counts on non-selective and selective media is used in evaluating the percent sub-lethal injury in the survivors.
While there is a growing body of knowledge on the effectiveness of non-thermal technologies for inactivating foodborne pathogens, published reports on tolerance of stress-adapted pathogens to those technologies are scarce. To our knowledge, except for two reports [15,39], there is no published research on the tolerance of AA pathogens to ACP. Accordingly, the main objective of the present research was to evaluate the tolerance of AA E. coli O157:H7 to HVACP in pineapple juice. A secondary objective was to determine the extent of sub-lethal injury in both NAA and AA survivors of that pathogen following HVACP treatment of pineapple juice.
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
Shiga-toxin-producing Escherichia coli O157:H7 (FRIK 125), isolated from an outbreak linked to apple cider was obtained from Dr. Charles Kaspar, University of Wisconsin. Frozen stock cultures (−80 ᵒC) in brain heart infusion (BHI) broth (Difco; Becton Dickinson, Sparks, MD) with added glycerol (10% v/v) were thawed under cold running water and activated in tryptic soy broth supplemented with 6.0 g/L yeast extract (TSBYE; pH 7.2; Difco; Becton Dickinson) at 35 ᵒC. Working cultures were held at 4 ᵒC until used in experiments. Two consecutive 24-h transfers of a working culture in tryptic soy broth without dextrose (TSB -G) and in TSB supplemented with 7.5 g dextrose per liter (TSB +G) (35 oC) were performed to obtain non-acid adapted (NAA) and acid-adapted (AA) cells respectively [29].
2.2. Preparation of Inocula
For NAA and AA E. coli O157: H7, equal volumes (1.5 ml) of each cell type were aseptically transferred to 2-ml microcentrifuge tubes. Cells were harvested by centrifugation (10,000 x g, 10 min, 22 ᵒC) Beckman Coulter Microfuge 16 Centrifuge (Beckman Coulter, Inc, Brea, CA). The pelleted cells were suspended in 1.5 ml of NaCl (8.5 g/L; saline) to yield a concentration of ~9.0 log10 colony forming units (CFU)/mL as verified by plate counts on tryptic soy agar supplemented with 6 g/L yeast extract (TSAYE). Colony counts of NAA and AA E. coli O157:H7 were determined by serially diluting (10-fold) cell suspensions in saline and surface plating appropriate dilutions on selective agar [sorbitol MacConkey agar (SMAC)] and non-selective agar (TSAYE). Bacterial colonies were counted after aerobic incubation (35 ᵒC) of TSAYE and SMAC agar for 24 and 48 h, respectively.
2.3. Preparation and Inoculation of Pineapple Juice
Whole pineapples (Del Monte Gold® Extra Sweet) from the same production lot were purchased from a local grocery store in Ames, Iowa. The pineapples were rinsed with distilled water, then the outside rind, top, bottom, and cores were removed using a clean knife and polypropylene cutting board. The flesh of the pineapples was cut into smaller chunks, which were used to extract juice. The pineapple juice was extracted using a juice extractor (Model #67608Z, Hamilton Beach Big Mouth Pro Juice Extractor, Glen Allen, Virginia). Particulates in the juice were removed by filtering the juice through two double layers of cheese cloth clamped with five 2-inch metal binder clips over a stainless-steel strainer. Two-hundred ml of the filtered juice were transferred to a sterile 250-ml Erlenmeyer flask. Ten ml of juice were aseptically transferred to appropriately labeled sterile Petri dishes (60 mm x 15 mm) and inoculated with 100 µL of either NAA or AA E. coli O157:H7 to give an initial viable count of ~ 7.0 log10 CFU/mL. The inoculated juice samples with lids on were held at 22 ± 1 ᵒC for no more than 0.5 h before exposing them (with lids off) to HVACP.
2.4. Treatment of Juice Samples with HVACP
A schematic of the dielectric barrier discharge (DBD) HVACP system for generating atmospheric plasma is shown in Figure 1. That system produces low temperature atmospheric plasma for in-package plasma treatment [40]. Input voltage of 120 V (AC) at 60 Hz is amplified by a step-up transformer (Phenix Technologies, MD, USA). Just before applying the HVACP, the lids were removed from the petri dishes to allow direct contact of the plasma with the juice. The uncovered samples were placed in the approximate center of a rigid polypropylene box. The boxes closed and placed in separate 35 cm x 27 cm high-barrier polypropylene Cryovac bags (B2630, Cryovac Sealed Air Corp., Duncan, SC, USA). All bags with boxes of samples were heat-sealed to retain air at atmospheric pressure. For each experiment, the relative humidity of the air was recorded. Each bag containing a box was placed between two 15.2 cm diameter aluminum electrodes with a discharge distance of 5.1 cm between the electrodes. A layer of plexiglass was placed under the top electrode whereas, a polypropylene layer (38.5 cm x 27.3 cm) was placed above the ground electrode. The plexiglass and polypropylene layers served as dielectric barriers to prevent arching and sparks discharge [41]. The samples of juice were treated with HVACP (70 kV) for 0 (control), 1, 3, 5, and 7 min. One group of samples was analyzed at ~ 6.0 min (0.1 h) after HVACP treatment, while the other group was analyzed after 24 h of post-treatment storage at 4°C. The control samples (no HVACP treatment) were handled in the same way as the other samples. For physicochemical tests (pH and degrees Brix), non-inoculated samples were treated with HVACP (70 kV) for 0 (control), 3, and 7 min. Each experiment was replicated at least three times.
2.5. Microbial Analysis of Juice Samples
At 0.1 h and 24 h after HVACP treatment of the juice, the bags were cut open, and Petri dishes containing the inoculated juice were removed from the boxes. Each sample was gently swirled to mix it, and 1.0 ml of juice was serially diluted (10-fold) in double-strength (2x) buffered peptone water (BPW, Difco). Appropriate dilutions were surface plated in duplicate on sorbitol MacConkey (SMAC) and on thin agar layer (TAL) medium (SMAC overlaid with 14 ml of TSAYE). The inoculated agar plates were incubated at 35°C for 24 hours (TAL) and 48 hours (SMAC) before colonies were counted. In instances when numbers of E. coli survivors were lower that the detection limit (10 CFU/mL) of the plating method, juice samples were enriched in TSBYE with added selective cefexime-tellurite (CT) supplement for 24 h then streak plated on SMAC with CT supplement to determine the presence or absence of the pathogen [42].
2.6. Calculation of D-Values
The death rate of NAA and AA E. coli O157:H7 in pineapple juice during HVACP treatment was determined by calculating D-values. Each D-value, the HVACP treatment time (minutes) that results in 1.0 log (90%) reduction in a population of viable E. coli O157:H7 cells, was determined by plotting numbers of survivors (log10 CFU/mL) versus HVACP exposure time (minutes) using Microsoft Excel 16.0 (Microsoft Excel, Seattle, WA). The regression curve was evaluated using linear regression analysis. Each D- value was derived from the negative reciprocal of the slope of the respective regression curve.
2.7. Determination of Sub-Lethal Injury
For each type of agar medium (SMAC agar and TAL medium) the numbers of E. coli O157:H7 survivors after each HVACP treatment time were used to calculate the reduction factor (RF). For each treatment time, the RF was calculated by dividing the viable count (CFU/mL) of NAA or AA cells in the juice before HVACP treatment by the CFU/mL in the juice after treatment. The log (RF) is expressed as shown in the following equation [43]:
Log RF = log [CFU/mL before HVACP treatment ÷ CFU/mL after HVACP treatment]
For all treatment times the log RFs for the selective medium (SMAC; y-axis) versus the log RFs for the TAL medium (x-axis) were plotted. Linear regression lines were fitted through the data points, and sub-lethal injury was determined by comparing the slopes to 1 and the y-intercepts to 0 [43].
2.8. pH Evaluation of Pineapple Juice
The pH of the pineapple juice was measured using a pH meter (Accumet Basic AB15 pH meter, Thermo Fisher Scientific Inc., Waltham, MA, USA). Prior to each measurement, the samples in the tubes were mixed by vortexing to prevent separation. For each replicate experiment, two pH measurements were performed on control and treated juice samples.
2.9. Measurement of Degrees Brix of Pineapple Juice
Degrees Brix (ᵒBrix) of the juice was measured using a refractometer (Atago PAL 1, Atago Co., Ltd., Tokyo, Japan). Distilled water was used to represent a blank liquid sample. Samples of pineapple juice in tubes were homogenized by vortexing, and a separate Pasteur pipette was used to add a small amount of each the juice to the refractometer well for measurement. For each replicate experiment, two ᵒBrix measurements were performed on control and treated juice samples.
2.10. Statistical Analysis
For microbiological analysis and quality evaluation tests (pH, and ᵒBrix), at least three replications of each experiment were performed. The JMP Pro statistical software version 16 (SAS Institute, Inc., Cary NC) was used to analyze the average numbers of survivors as a function of HVACP treatment time. To determine the significant differences, analysis of variance (ANOVA) was performed with a p-value < 0.05. Tukey’s honestly significant difference (HSD) test was performed to identify means that are significantly different from each other.
3. Results
3.1. Survivors of E. coli in Pineapple Juice at 0.1h and 24 h after HVACP Treatment
Survivors of NAA and acid AA E. coli O157:H7 in pineapple juice based on colony counts on SMAC agar are shown in Figure 2A,B. When juice samples were analyzed at 0.1 h after HVACP treatment, viable counts of both NAA and AA cells did not differ from counts in control samples irrespective of treatment time (P>0.05). Significant reductions in viable counts of both NAA and AA cells were observed in juice treated with HVACP for 1 and 3 min and held at 4 °C for 24 h (P<0.05). However, the numbers AA survivors exceeded those of NAA survivors. Following exposure to HVACP for 5 or 7 min and subsequently holding the juice at 4 °C for 24 h, the NAA cells in juice were completely inactivated (negative enrichment test) (Figure 2A). In contrast, the numbers of viable AA cells (log10 CFU/ml) in juice treated for 5 and 7 min, and held 4 °C for 24 h, were 3.03 (5 min) and 2.82 (7 min) (Figure 2B).
As previously described for SMAC agar, a similar trend in viability of NAA and AA cells in control and HVACP-treated juice was observed based on bacterial counts on TAL medium (Figure 3A,B). Control and treated juice samples analyzed at 0.1 h after HVACP treatment exhibited high viable counts of both NAA and AA cells with no significant differences among those counts (P>0.05). Significant decreases in NAA cells were evident in juices analyzed at 24 h with no survivors detected in juice treated for 5 and 7 min (Figure 3A). Irrespective of duration of HVACP treatment, AA cells of the pathogen were consistently detected in juice analyzed at 24 h (Figure 3B).
3.2. Effect of Physiological State on E. coli O157:H7 Tolerance of HVACP in Juice
A direct comparison of the effect of physiological state (NAA and AA) on E. coli O157:H7 survivors in juice analyzed at 24 h after HVACP treatment is shown in Figure 4A,B. Based on bacterial counts on SMAC agar (Figure 4A) and on TAL medium (Figure 4B), the numbers of both NAA and AA survivors in control juice were not significantly different (P>0.05). The NAA survivors in refrigerated juice treated with HVACP for 1 and 3 min were 3.46 and 0.59 log10 CFU/mL, respectively, whereas numbers of AA survivors were higher - 6.51 (1 min) and 5.49 (3 min) (P<0.05). Very similar results were observed when TAL medium was used to recover E. coli survivors (Figure 4B). For both recovery media numbers of NAA survivors of 5- and 7- minute HVACP treatments were beyond detection (negative enrichment test); however, AA survivors were consistently detected with average viable counts ranging from 2.82 to 3.33 Log10 CFU/mL (Figure 4A,B).
3.3. D-Values of NAA and AA E. coli in Pineapple Juice
The resistance to HVACP (70kV) based on decimal reduction times (D-values) for NAA and AA E. coli O157:H7 recovered from pineapple juice at 24 h after HVACP treatment is shown in Table 1. Larger D-values were obtained for AA cells compared to NAA cells; however, differences were not statistically significant (P>0.05). Moreover, for each physiological state (NAA or AA) there were no significant differences in D-values irrespective of the type of agar medium used to enumerate survivors (P>0.05).
3.4. Sub-Lethal Injury of NAA and AA E. coli in Pineapple Juice
Sub-lethal injury (expressed by linear regression parameters) in NAA and AA E. coli O157:H7 survivors in HVACP-treated juice is shown in Table 2. Based on the method of Wuytack et al. [43] we observed that sub-lethal injury in NAA and AA survivors was not detected when juice samples were analyzed at 0.1 h after HVACP treatment. However, significant (P<0.05) sub-lethal injury was detected in both NAA and AA survivors in juice analyzed 24 h following HVACP (70 kV) treatment (Table 2).
3.5. pH and Degrees Brix of Pineapple Juice
The effects of HVACP treatment on the pH and ᵒBrix of pineapple juice analyzed at 0.1 h and 24 h after HVACP treatment are shown in Table 3. The initial pH of the pineapple juice (pH 3.36) decreased with increased exposure to HVACP; however, those decreases were not significant (P>0.05). No differences were noted between pH of the juice taken at 0.1 h compared to 24 h post-treatment (P>0.05). Significant increases in oBrix were observed in juice that was exposed to HVACP for 3 or 7 min and tested at 0.1 h after treatment (P<0.05). No increases in oBrix were observed in treated samples that were stored at 4 ᵒC and tested after 24 h (P>0.05). Juice evaluated after 24 h of storage (4 ᵒC) had higher oBrix values compared to juice evaluated at 0.1 h after HVACP treatment (P<0.05).
4. Discussion
4.1. Stress Adaptation in Foodborne Microorganisms
Foodborne microorganisms encounter a myriad of stresses in food processing environments, and survivors may become stress adapted thus increasing their tolerance to subsequent chemical or physical food processes [44,45,46]. Typical stresses imposed by food processes include, but are not limited to drying, high salt, shifts in temperatures, low water activity, exposure antimicrobial food preservatives, ultraviolet radiation, chemical cleaners and sanitizers, low pH, and organic acids [13,46,47,48]. Of those stresses, low pH and type of acid frequently impact bacterial survival and growth because organic acids and acid cleaners are widely used in the food industry. In this regard, foodborne bacteria have developed physiological mechanisms to enhance their survival by adapting to acid stress [13,47]. Moreover, acid adaptation in foodborne pathogens can cross-protect them against stresses imposed by subsequently applied food processes [13,31]. Based on this concept, we hypothesized that AA E. coli O157:H7 cells are more tolerant than NAA cells to HVACP in pineapple juice.
4.2. Survivors of E. coli O157:H7 in Juice after HVACP Treatment
When juice samples were exposed to HVACP for 1 to 7 min then transferred to nutrient agar within 0.1 h, no significant loss in viability of NAA or AA E. coli O157:H7 occurred (Figures 2A,B and 3A,B). By analyzing juice samples within such a short time after exposure to HVACP, it seemed that the treatments had no lethal effect on the pathogen. Our results are consistent with those of Hartanto [49] who reported significantly higher viable counts of E. coli O157:H7 in HVACP (80 kV)-treated organic pineapple juice analyzed within 1 h as opposed to 24 h after treatment. Our observation that HVACP-treated NAA and AA cells exhibited significantly lower viable counts in juice after 24 h suggests increased sensitivity of those cells to the juice environment. That sensitivity is likely due to sub-lethal structural and/or metabolic injury initiated by HVACP that was exacerbated by inhibitory conditions in the juice. The hostile environment of the pineapple juice (pH 3.36) likely precluded cellular repair, caused further injury, and decreased E. coli O157:H7 survivors in the juice held at 4 ͦ C for 24 h. Also, pineapple juice has phenolic compounds and organic acids such as malic acid, citric acid, ascorbic acid, and isocitric acid [50,51] that can prevent repair processes in sub-lethally injured cells [35]. Moreover, Yadav and Roopesh [52] demonstrated synergistically higher bacterial inactivation resulting from combining ACP and organic acids.
Results of the present study further suggest that sub-lethal injuries in both NAA and AA cells were repaired when cells were transferred to nutritious agar media within 0.1 h after HACP treatment. This may explain our earlier mentioned observation of no significant loss of viability of NAA cells in HVACP-treated juice, which was plated on agar media within 0.1h after treatment.
4.3. Effect of Physiological States on Numbers of E. coli Survivors
Figure 4A,B shows the results of a direct comparison of effects of physiological state (NAA vs AA) on the numbers of E. coli O157:H7 survivors in HVACP-treated juice for 24 h (Figure 4A,B. For all treatment times (1 to 7 min) irrespective of the type of recovery medium, the numbers of AA survivors in juice were significantly higher (P<0.05) than those of NAA survivors. Very low numbers of NAA cells (0.49 Log10 CFU/mL) survived the 3.0-min HVACP treatment of juice Figure 4B). This result demonstrates greater than 6.0 Log10 CFU/mL reduction of the initial viable count of the NAA pathogen. Based on the use of NAA cells, that HVACP (70 kV, 3 min) process would fully comply with the FDA juice HACCP regulation requiring a 5.0 Log10 CFU/mL reduction of the pertinent pathogen [12]. In contrast, the high numbers of AA cells (5.49 Log10 CFU/mL) that survived those same processing parameters represented only a 1.03 Log10 CFU/mL reduction (Figure 4B). Considering that both NAA and AA cells in control (0 kV) juice survived in high numbers (6.56 to 6.61 log10 CFU/mL) for 24 h, the very low numbers of NAA in HVACP-treated juice suggest that AA E. coli O157:H7 were more tolerant to HVACP. Therefore, to ensure microbial safety of acidic juices treated with cold plasma technology, AA cells of the pertinent pathogen should be used in process validation studies.
Tonsul and Gonul [53] demonstrated that acid shock of E. coli O157:H7 for 1 h in TSB (pH 4.5 to 5.5) significantly increased its acid tolerance in TSB at pH 2.5 to 3.0. In the present study, there was a 1 h interval between inoculation of the juice samples with the pathogen and HVACP treatment. Therefore, the similarity in survival of NAA and AA cells in control juice is likely due to NAA cells triggering an acid shock response [54], which protected them against the acidic conditions in the juice. Unlike the NAA cells, far higher numbers of AA cells that survived after 24 h in treated juice (Figure 4A,B) suggest that acid adaptation exerted a protective effect against HVACP. Wang et al. [47] reported that acid adaptation, apart from inducing bacterial tolerance to low pH, may also trigger physiological and genetic systems that aid in cross protecting cells against different stresses.
Acid-adaption can be induced by exposure of microbial cells to a gradual decrease in pH. This situation simulates environmental stress conditions in a fermented product in which acids are gradually produced by the natural microflora or by acid-producing bacteria that are added to the food products. In the present study, we cultured E. coli O157:H7 in TSB (with added 7.5 g dextrose) at 35 oC for 24 h to obtain AA cells [29]. Our results are consistent those of Liao et al. (2018), who reported that long-term (24 h) acid adaptation of Staphylococcus aureus at pH 4.5 resulted its higher tolerance to cold plasma compared to NAA cells of that same organism. Those findings are inconsistent with results obtained from short-term acid adaptation (4 h) of S. aureus, which exhibited no increased tolerance to the plasma [33]. Similarly, short term (2 h) acid adaptation induced with different acids including hydrochloric, ascorbic, acetic, citric, lactic, and malic at pH (6.4, 5.4, and 4.5), had very little effect on tolerance of Salmonella Typhimurium and Salmonella Enteritidis to cold plasma in a model system [15]. Based on those research findings and results of the present study, it seems that foodborne bacteria require a prolonged exposure to low pH conditions to develop cross-protection against cold plasma.
4.3. D-Values for E. coli O157:H7 in Pineapple Juice
In the present study the rate of inactivation of NAA and AA E. coli O157:H7 by HVACP was expressed as the D-value (decimal reduction time) to describe resistance of the pathogen to HVACP (Table 1). The D-values for AA cells were consistently higher than those of NAA cells; however, the differences were not statistically significant (P > 0.05) suggesting that the rate of inactivation by direct HVACP (70 kV) was similar for both physiological states of the pathogens in pineapple juice. These results are likely due to the relatively large standard deviations in the data that decrease the statistical significance of the results.
4.3. Sub-Lethal Injury of NAA and AA E. coli in Pineapple Juice
To determine sub-lethal injury in surviving populations of NAA and AA E. coli O157:H7, we used the method described by Wuytack et al. [43] which is based on the reduction factor (RF) concept as previously described in section 2.7. For each treatment time, the Log RF for the selective medium (SMAC; y-axis) versus the Log RF for the TAL medium (x-axis) were plotted. Linear regression lines through the data points were used to determine sub-lethal injury by comparing the slopes to 1 and the y-intercepts to 0 [43]. There is no sub-lethal injury when the slope of the line is 1.0 and intercept is 0. In this instance, the same decrease in viability occurs on the SMAC agar and on the TAL medium. A slope that is significantly greater than 1.0 or an intercept that is significantly greater than 0, suggests sub-lethal injury (P<0.05). This is based on the larger viability reduction on SMAC agar than on TAL medium. Moreover, one can assume that a larger deviation of the slope from 1.0 or a larger deviation of the intercept from 0 reflects a greater extent of sub-lethal injury.
Our results show that the intercept of the regression line for viability reduction in NAA and AA E. coli O157:H7 in HVACP-treated pineapple juice held at 4 ͦ C for 24 h, was significantly larger than 0 (Table 2). These results suggest that sub-lethal injury occurred among NAA and AA survivors in juice held at 4 oC for 24h but was not detected in NAA and AA survivors in juice samples that were plated within 0.1 h after HVACP treatment.
A factor that likely caused sub-lethal injury in NAA and AA survivors in the juice is the residual reactive species generated by the cold plasma. Short-lived reactive species such as free electrons (e-), singlet oxygen (O), hydrogen atom (H), hydroxyl radical (OH•), hydroperoxyl radical (HOO•), and superoxide anion (-O2) impinge on the surface of liquids and react with water molecules to form more long-lived secondary species [55,56]. Long-lived secondary species such as ozone (O3), hydrogen peroxide (H2O2) nitrous acid (HNO2), and peroxynitrous acid [57] continue to exert an antimicrobial effect even after the HVACP treatment has stopped [58]. Han et al. [24] published scanning electron microscopy images of S. aureus and E. coli cells that were exposed to cold plasma treatment at 80 kV and held for 24 h after treatment. The cells treated with indirect plasma showed signs of structural damage in comparison to their non-treated counter parts, indicating damage via residual reactive oxygen and nitrogen species. All those species can cause further lesions and prevent the resuscitation of sub-lethally injured bacteria [35,59]. Therefore, in the present study, the stress imposed by residual long lived plasma species and the intrinsic factors of the pineapple juice (ie. low pH, organic acids, phenolic compounds), contributed to sub-lethal injury in the NAA and AA E. coli cells recovered from the juice held at 4 °C for 24 h.
Determination of sub-lethal injury in foodborne pathogens after their exposure to chemical or physical food processes is important for two main reasons: i) failure to detect sub-lethally injured pathogens in a processed food or beverage product can overestimate the antimicrobial effectiveness of the applied process. This is also important when selective media are utilized to recover pathogen survivors because, depending on the extent of injury, sub-lethally injured cells may not grow on selective media. ii) levels of antimicrobial treatments that inflict sub-lethal injury in pathogens provide opportunities for their use in combination with other interventions to prevent cellular repair to ultimately inactivate those pathogens.
4.3. pH and Degrees Brix of Pineapple Juice
The pH and ᵒBrix values of juice treated with HVACP (Table 3) are similar to those stated in the published literature on pineapple juice [51]. Our results show a slight decrease in pH of the juice with increased exposure to HVACP; however, it was not significantly different from pH of control juice (Table 3). Several studies that used the DBD method for applying cold plasma to various juices, demonstrated that pH decreased with increasing exposure to HVACP [32,39,49]. One likely reason for the pH decrease is the formation of nitrous acid (HNO2) or nitric acid (HNO3) generated from reactive nitrogen species during and after HVACP treatment [60].
Our results indicate that HVACP significantly increased the ᵒBrix of the juice samples which were analyzed within 0.1 h after treatment (P<0.05). Both Hartanto et al. [49] and Liao et al. [39] also found that increasing HVACP treatment times increased the ᵒBrix of juice. The increase in ᵒBrix could be attributed to ROS such as ozone causing the depolymerization of polysaccharides via oxidation thus creating shorter chain polysaccharides and smaller soluble units (sugars). Additionally, ozone species could cleave the glycosidic bonds and oxidize the functional groups of polysaccharides leading to the development of lactones, carbon dioxide, hydroperoxides, carbonyl, and carboxyl compounds [61,62]. The stabilization of the ᵒBrix in juice analyzed after 24 h is likely due to a completion of further depolymerization reaction by residual reactive plasma species during the 24-h refrigeration (4 °C) of the juice.
5. Conclusions
Direct application of HVACP (70 kV) for 3, 5 or 7 min significantly reduces populations of NAA E. coli O157:H7 by greater 5.00 log10 CFU/mL in pineapple juice during post-treatment storage (4 ᵒC) for 24 h. However, those same HVACP treatments yield significantly lower reductions of AA E. coli that fail to comply with the FDA juice HACCP regulation for killing E. coli O157:H7. Acid adaptation can cross-protect E. coli against the bactericidal effect of HVACP in pineapple juice and should be considered when designing protocols for ensuring the microbial safety of juices treated with cold plasma technology.
Author Contributions
Conceptualization, methodology, supervision, writing – review and editing, AM; Investigation, formal analysis, writing - original draft preparation, AL and BL; Resources, writing – review and editing, JD and PD; Supervision, resources, review and editing, TB; Writing – review and editing, ETP and SC.
Funding
This research received no external funding.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
The authors thank Griffin Lind for assistance in performing microbiological analyses of the juice samples. This work was supported by the Iowa Agriculture and Home Economics Experiment Station, Ames, Iowa. Project No. IOW04202 is sponsored by Hatch Act and State of Iowa funds.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- 1Grand View Research. (2023). Fresh Fruits Market Size & Share| Industry Report. In Market Analysis Report. https://www.grandviewresearch.com/industry-analysis/fresh-fruits-market-report.
- Technavio. (2022, December). Juices Market - Size, Share, Growth, Trends, Industry Analysis 2027. https://www.technavio.com/report/juices-market-industry-analysis.
- Pem, D., & Jeewon, R. (2015). Fruit and Vegetable Intake: Benefits and Progress of Nutrition Education Interventions- Narrative Review Article. Iranian Journal of Public Health, 44(10), 1309. /pmc/articles/PMC4644575/.
- Dewanti-Hariyadi, R.; Maria, A.; Sandoval, P. (2013). Microbiological quality and safety of fruit juices. Food Rev. Int, 1, 54-57.
- Danyluk, M. D., Goodrich-Schneider, R. M., Schneider, K. R., Harris, L. J.; Worobo, R. W. (2012). Outbreaks of Foodborne Disease Associated with Fruit and Vegetable Juices, 1922–2010: FSHN12-04/FS188. https://journals.flvc.org/edis/article/download/119658/117576.
- Jajere, S. M. (2019). A review of Salmonella enterica with particular focus on the pathogenicity and virulence factors, host specificity and antimicrobial resistance including multidrug resistance. Veterinary world, 12(4), 504. [CrossRef]
- Salfinger, Y. (Ed.), & Tortorello, M. L. (Ed.). (2015). Compendium of Methods for the Microbiological Examination of Foods (Y. Salfinger & M. Lou Tortorello, Eds.; 5th ed.). American Public Health Association.
- Macarisin, D., Wooten, A., De Jesus, A., Hur, M., Bae, S., Patel, J., Evans, P., Brown, E., Hammack, T., & Chen, Y. (2017). Internalization of Listeria monocytogenes in cantaloupes during dump tank washing and hydrocooling. International Journal of Food Microbiology, 257, 165–175. [CrossRef]
- Mendonca, A. (2005). Bacterial infiltration and internalization in fruits and vegetables. In Produce degradation: Pathways and prevention (pp. 441–462). CRC Press, Boca Raton, FL. 10.1201/9781420039610.ch14.
- FDA. (2008, February). Guidance for Industry: Guide to Minimize Microbial Food Safety Hazards of Fresh-cut Fruits and Vegetables | FDA. https://www.fda.gov/regulatory-information/search-fda-guidance-documents/guidance-industry-guide-minimize-microbial-food-safety-hazards-fresh-cut-fruits-and-vegetables.
- Thomas-Popo, E., Mendonca, A., Dickson, J., Shaw, A., Coleman, S., Daraba, A., Jackson-Davis, A., & Woods, F. (2019). Isoeugenol significantly inactivates Escherichia coli O157:H7, Salmonella enterica, and Listeria monocytogenes in refrigerated tyndallized pineapple juice with added Yucca schidigera extract. Food Control, 106, 106727. [CrossRef]
- FDA. (2001). Hazard Analysis and Critical Control Point (HAACP); Procedures for the Safe and Sanitary Processing and Importing of Juice. https://www.federalregister.gov/documents/2001/01/19/01-1291/hazard-analysis-and-critical-control-point-haacp-procedures-for-the-safe-and-sanitary-processing-and.
- Wu, W., Xiao, G., Yu, Y., Xu, Y., Wu, J., Peng, J., & Li, L. (2021). Effects of high pressure and thermal processing on quality properties and volatile compounds of pineapple fruit juice. Food Control, 130, 108293. [CrossRef]
- Roobab, U., Aadil, R. M., Madni, G. M., & Bekhit, A. E. D. (2018). The Impact of Nonthermal Technologies on the Microbiological Quality of Juices: A Review. Comprehensive Reviews in Food Science and Food Safety, 17(2), 437–457. [CrossRef]
- Calvo, T., Alvarez-Ordóñez, A., Prieto, M., Bernardo, A., López, M. (2017). Stress adaptation has a minor impact on the effectivity of Non-Thermal Atmospheric Plasma (NTAP) against Salmonella spp. Food Research International, 102, 519–525. [CrossRef]
- Van Impe, J., Smet, C., Tiwari, B., Greiner, R., Ojha, S., Stulić, V., Vukušić, T., & Režek Jambrak, A. (2018). State of the art of nonthermal and thermal processing for inactivation of micro-organisms. Journal of Applied Microbiology, 125(1), 16–35. [CrossRef]
- Nonglait, D. L., Chukkan, S. M., Arya, S. S., Bhat, M. S., & Waghmare, R. (2022). Emerging non-thermal technologies for enhanced quality and safety of fruit juices. International Journal of Food Science & Technology, 57(10), 6368-6377. [CrossRef]
- Yepez, X., Illera, A. E., Baykara, H., & Keener, K. (2022). Recent advances and potential applications of atmospheric pressure cold plasma technology for sustainable food processing. Foods, 11(13), 1833.
- Ozen, E., & Singh, R. K. (2020). Atmospheric cold plasma treatment of fruit juices: A review. Trends in Food Science & Technology, 103, 144–151. [CrossRef]
- Misra, N. N., Pankaj, S. K., Frias, J. M., Keener, K. M., & Cullen, P. J. (2015). The effects of nonthermal plasma on chemical quality of strawberries. Postharvest Biology and Technology, 110, 197–202. [CrossRef]
- Bourke, P., Ziuzina, D., Han, L., Cullen, P. J., & Gilmore, B. F. (2017). Microbiological interactions with cold plasma. Journal of Applied Microbiology, 123(2), 308-324. [CrossRef]
- Nicol, M. K. J., Brubaker, T. R., Honish, B. J., Simmons, A. N., Kazemi, A., Geissel, M. A., Whalen, C. T., Siedlecki, C. A., Bilén, S. G., Knecht, S. D., & Kirimanjeswara, G. S. (2020). Antibacterial effects of low-temperature plasma generated by atmospheric-pressure plasma jet are mediated by reactive oxygen species. Scientific Reports 2020 10:1, 10(1), 1–11. [CrossRef]
- Niemira, B. A. (2012). Cold plasma decontamination of foods. Annual Review of Food Science and Technology, 3(1), 125–142. [CrossRef]
- Han, J. Y., Song, W. J., Eom, S., Kim, S. B., Kang, D. H. (2020). Antimicrobial efficacy of cold plasma treatment against food-borne pathogens on various foods. Journal of Physics D: Applied Physics, 53(20), 204003. [CrossRef]
- Ucar, Y., Ceylan, Z., Durmus, M., Tomar, O., & Cetinkaya, T. (2021). Application of cold plasma technology in the food industry and its combination with other emerging technologies. Trends in Food Science & Technology, 114, 355–371. [CrossRef]
- Liao, X., Ma, Y., Daliri, E. B. M., Koseki, S., Wei, S.; Liu, D., Ye, X., Chen, S., Ding, T. (2020). Interplay of antibiotic resistance and food-associated stress tolerance in foodborne pathogens. Trends in Food Science & Technology, 95, 97–106. [CrossRef]
- Begley, M.; Hill, C. (2015). Stress adaptation in foodborne pathogens. Annual review of food science and technology, 6, 191-210.
- Wu, R. A., Yuk, H. G., Liu, D., & Ding, T. (2022). Recent advances in understanding the effect of acid-adaptation on the cross-protection to food-related stress of common foodborne pathogens. Critical Reviews in Food Science and Nutrition, 62(26), 7336–7353. [CrossRef]
- Buchanan, R. L., Edelson, S. G., Snipes, K., Boyd, G. (1998). Inactivation of Escherichia coli O157:H7 in apple juice by irradiation. Applied and Environmental Microbiology, 64(11), 4533–4535. [CrossRef]
- Yuk, H.-G., & Marshall, D. L. (2005). Influence of Acetic, Citric, and Lactic Acids on Escherichia coli O157:H7 Membrane Lipid Composition, Verotoxin Secretion, and Acid Resistance in Simulated Gastric Fluid. In Journal of Food Protection (Vol. 68, Issue 4). http://meridian.allenpress.com/jfp/article-pdf/68/4/673/1676337/0362-028x-68_4_673.pdf.
- Haberbeck, L. U., Wang, X., Michiels, C., Devlieghere, F., Uyttendaele, M., Geeraerd, A. H. (2017). Cross-protection between controlled acid-adaptation and thermal inactivation for 48 Escherichia coli strains. International Journal of Food Microbiology, 241, 206–214. [CrossRef]
- Xu, L., Garner, A. L., Tao, B., & Keener, K. M. (2017). Microbial Inactivation and Quality Changes in Orange Juice Treated by High Voltage Atmospheric Cold Plasma. Food and Bioprocess Technology, 10(10), 1778–1791. [CrossRef]
- Liao, X., Li, J.; Suo, Y., Ahn, J., Liu, D., Chen, S., Hu, Y., Ye, X., Ding, T. (2018). Effect of preliminary stresses on the resistance of Escherichia coli and Staphylococcus aureus toward non-thermal plasma (NTP) challenge. Food Research International, 105, 178–183. [CrossRef]
- Wu, D., Forghani, F., Daliri, E. B. M., Li, J., Liao, X., Liu, D., ... & Ding, T. (2020). Microbial response to some nonthermal physical technologies. Trends in Food Science & Technology, 95, 107-117. [CrossRef]
- Wesche, A. M., Gurtler, J. B., Marks, B. P., & Ryser, E. T. (2009). Stress, sublethal injury, resuscitation, and virulence of bacterial foodborne pathogens. Journal of Food Protection, 72(5), 1121–1138. [CrossRef]
- Ray, B. (1989). Injured index and pathogenic bacteria: occurence and detection in foods, water and feeds. CRC Press, Boca Raton FL.
- Busch, S. V., & Donnelly, C. W. (1992). Development of a repair-enrichment broth for resuscitation of heat-injured Listeria monocytogenes and Listeria innocua. Applied and environmental microbiology, 58(1), 14-20.
- Espina, L., García-Gonzalo, D., & Pagán, R. (2016). Detection of thermal sublethal injury in Escherichia coli via the selective medium plating technique: mechanisms and improvements. Frontiers in microbiology, 7, 208323. [CrossRef]
- Liao, X., Li, J., Muhammad, A. I., Suo, Y., Chen, S., Ye, X.; Liu, D., Ding, T. (2018). Application of a Dielectric Barrier Discharge Atmospheric Cold Plasma (Dbd-Acp) for Eshcerichia coli Inactivation in Apple Juice. Journal of Food Science, 83(2), 401–408. [CrossRef]
- Yepez, X. V., & Keener, K. M. (2016). High-voltage atmospheric cold plasma (HVACP) hydrogenation of soybean oil without trans-fatty acids. Innovative Food Science & Emerging Technologies, 38, 169-174.
- Deepak, G. D., Joshi, N. K., Prakash, R. (2020). The modelling and characterization of dielectric barrier discharge-based cold plasma sets. https://www.cambridgescholars.com/product/978-1-5275-4539-7.
- Rodriguez, E., Arques, J. L., Nunez, M., Gaya, P., & Medina, M. (2005). Combined effect of high-pressure treatments and bacteriocin-producing lactic acid bacteria on inactivation of Escherichia coli O157: H7 in raw-milk cheese. Applied and Environmental Microbiology, 71(7), 3399-3404.
- Wuytack, E. Y., Phuong, L. D. T., Aertsen, A., Reyns, K. M. F., Marquenie, D., De Ketelaere, B., Masschalck, B., Van Opstal, I., Diels, A. M. J., & Michiels, C. W. (2003). Comparison of sublethal injury induced in Salmonella enterica serovar typhimurium by heat and by different nonthermal treatments. Journal of Food Protection, 66(1), 31–37. [CrossRef]
- Guillén, S., Nadal, L., Álvarez, I., Mañas, P., Cebrián, G. (2021). Impact of the resistance responses to stress conditions encountered in food and food processing environments on the virulence and growth fitness of non-typhoidal Salmonellae. Foods. Mar 14;10(3):617.
- Ding, T., Xinyu, L., Feng, J. The importance of understanding the stress response in foodborne pathogens along the food production chain. In Stress Responses of Foodborne Pathogens, Cham: Springer International Publishing, 2022, pp. 3-31. https://link.springer.com/chapter/10.1007/978-3-030-90578-1_1.
- Singh, A., & Yemmireddy, V. (2022). Pre-Growth Environmental Stresses Affect Foodborne Pathogens Response to Subsequent Chemical Treatments. Microorganisms, 10(4). [CrossRef]
- Wang, Q., Buchanan, R. L., & Tikekar, R. V. (2019). Evaluation of adaptive response in E. coli O157:H7 to UV light and gallic acid based antimicrobial treatments. Food Control, 106, 106723. [CrossRef]
- Hu, S., Yu, Y., Lv, Z., Shen, J., Ke, Y., Xiao, X. (2020). Proteomics study unveils ROS balance in acid-adapted Salmonella Enteritidis. Food Microbiology, 92, 103585. [CrossRef]
- Hartanto, V. (2022). Application of atmospheric cold plasma for inactivation of foodborne enteric pathogens on raw and dry roasted pistachio kernels and in pineapple juice M.S. Thesis, Iowa State University, Ames, Iowa 50011, May 2022. https://www.proquest.com/docview/2679879590?pq-origsite=gscholar&fromopenview=true&sourcetype=Dissertations%20&%20Theses.
- Couto, D. S., Cabral, L. M. C., da Matta, V. M.; Deliza, R., de Grandi Castro Freitas, D. (2011). Concentration of pineapple juice by reverse osmosis: physicochemical characteristics and consumer acceptance. Food Science and Technology, 31(4), 905–910. [CrossRef]
- Hounhouigan, M. H., Linnemann, A. R., Soumanou, M. M., Van Boekel, M. A. J. S. (2014). Effect of Processing on the Quality of Pineapple Juice. Food Reviews International, 30(2), 112–133. [CrossRef]
- Yadav, B., Roopesh, M. S. Synergistically enhanced Salmonella Typhimurium reduction by sequential treatment of organic acids and atmospheric cold plasma and the mechanism study. Food Microbiology 104 (2022): 103976.
- Tosun, H., and Gonul, S. A. (205). The effect of acid adaptation conditions on acid tolerance response of Escherichia coli O157: H7.” Turkish Journal of Biology 29, no. 4:197-202. https://journals.tubitak.gov.tr/biology/vol29/iss4/2.
- Foster, J. W. 2000. Microbial responses to acid stress, p. 99–115. In G. Storz and R. Hengge-Aronis (ed.), Bacterial stress responses. ASM Press, Washington, DC.
- Adamovich, I., Baalrud, S.D., Bogaerts, A.; Bruggeman, P.J.; Cappelli, M. et al. (2017). The 2017 Plasma Roadmap: Low temperature plasma science and technology J. Phys. D: Appl. Phys. 50: 323001 (46pp). [CrossRef]
- Kondeti, V.S., Santosh K., Bruggeman, P.J. (2020). The interaction of an atmospheric pressure plasma jet with liquid water: dimple dynamics and its impact on crystal violet decomposition. Journal of Physics D: Applied Physics 54, no. 4: 045204.
- Ikawa, S., Kitano, K., Hamaguchi, S. (2010). Effects of pH on bacterial inactivation in aqueous solutions due to low-temperature atmospheric pressure plasma application. Plasma Processes and Polymers. Jan 14;7(1):33-42. [CrossRef]
- Misra, N. N., Ximena Yepez, Lei Xu, and Kevin Keener. (2019). In-package cold plasma technologies.” Journal of Food Engineering 244: 21-31. [CrossRef]
- Ghate, V., Kumar, A., Zhou, W., Yuk, H. G. (2015). Effect of organic acids on the photodynamic inactivation of selected foodborne pathogens using 461 nm LEDs. Food Control, 57, 333–340. [CrossRef]
- Oehmigen, K., Hähnel, M., Brandenburg, R., Wilke, C., Weltmann, K. D., & Von Woedtke, T. (2010). The Role of Acidification for Antimicrobial Activity of Atmospheric Pressure Plasma in Liquids. Plasma Processes and Polymers, 7(3–4), 250–257. [CrossRef]
- Wang, Y., Hollingsworth, R. I., Kasper, D. L. (1999). Ozonolytic depolymerization of polysaccharides in aqueous solution. Carbohydrate Research, 319(1–4), 141–147. [CrossRef]
- Ben’Ko, E. M., Manisova, O. R., Lunin, V. V. (2013). Effect of ozonation on the reactivity of lignocellulose substrates in enzymatic hydrolyses to sugars. Russian Journal of Physical Chemistry A, 87(7), 1108–1113. [CrossRef]
Figure 1.
Schematic of the components of the dielectric barrier, high voltage atmospheric cold plasma (HVACP) system used to treat pineapple juice.
Figure 1.
Schematic of the components of the dielectric barrier, high voltage atmospheric cold plasma (HVACP) system used to treat pineapple juice.
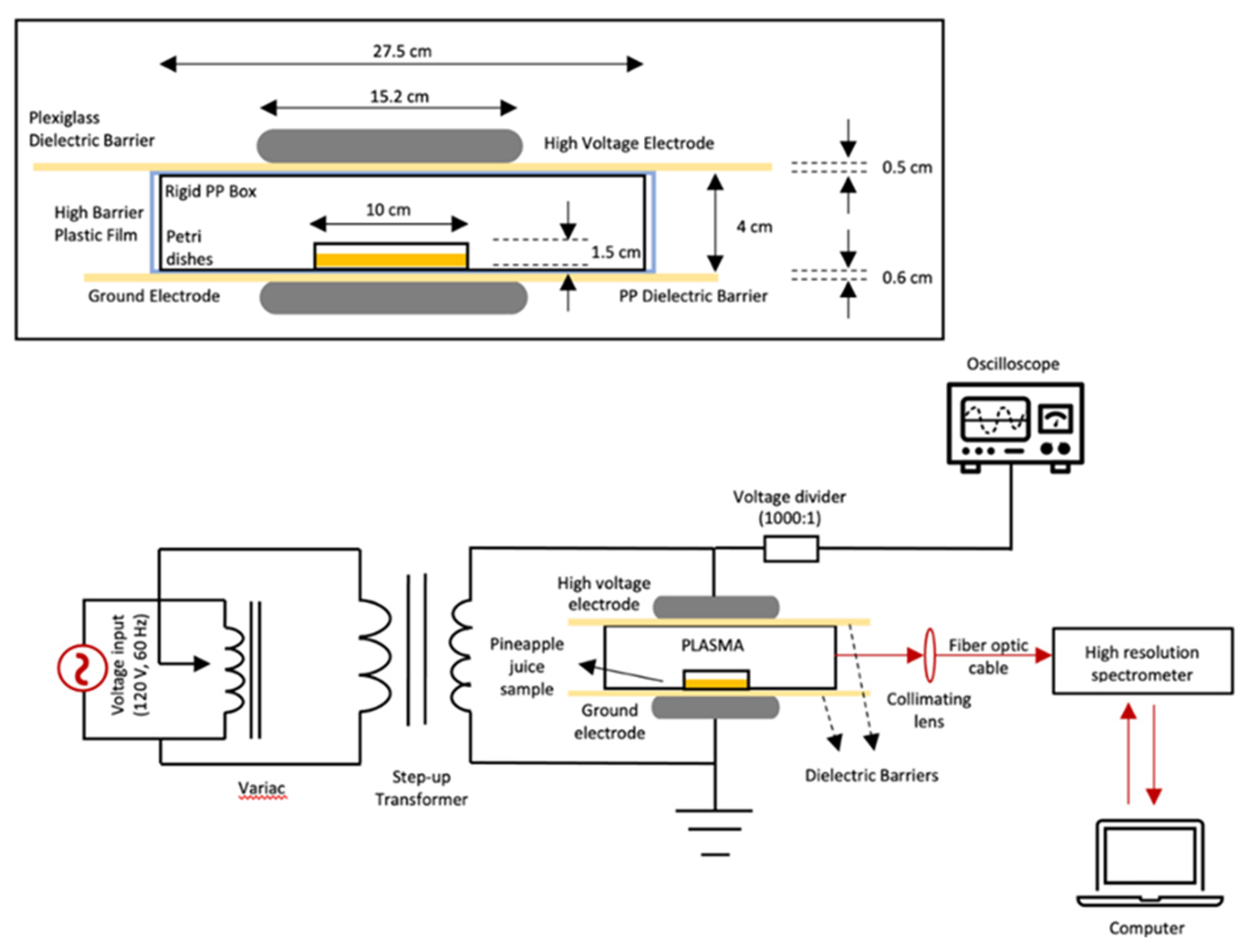
Figure 2.
Effect of holding time (0.1 h and 24 h) following HVACP (70 kV) treatment on viability of non-acid-adapted (A) and acid adapted (B) Escherichia coli O157:H7 survivors in pineapple juice. Survivors were recovered on sorbitol MacConkey (SMAC) agar. For each treatment time, different letters (A or B) over the bars indicate a significant difference in numbers of survivors at 0.1 h and 24 h post-treatment (P<0.05).
Figure 2.
Effect of holding time (0.1 h and 24 h) following HVACP (70 kV) treatment on viability of non-acid-adapted (A) and acid adapted (B) Escherichia coli O157:H7 survivors in pineapple juice. Survivors were recovered on sorbitol MacConkey (SMAC) agar. For each treatment time, different letters (A or B) over the bars indicate a significant difference in numbers of survivors at 0.1 h and 24 h post-treatment (P<0.05).
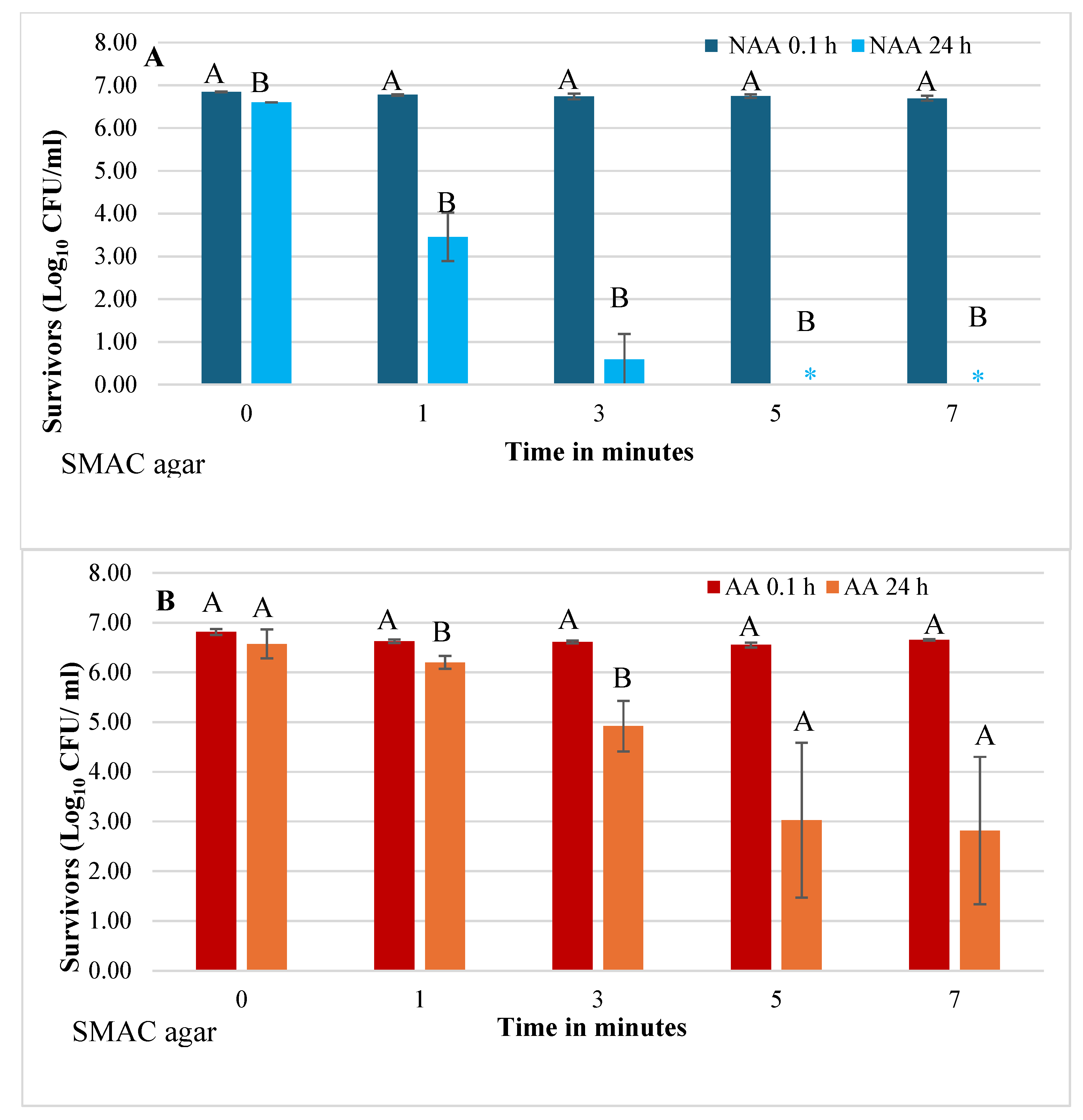
Figure 3.
Effect of holding time (0.1 h and 24 h) following HVACP (70 kV) treatment on viability of non-acid-adapted (A) and acid adapted (B) Escherichia coli O157:H7 survivors in pineapple juice. Survivors were recovered on thin agar layer (TAL) medium. For each treatment time, different letters (A or B) over the bars indicate a significant difference in numbers of survivors at 0.1 h and 24 h post-treatment (P<0.05).
Figure 3.
Effect of holding time (0.1 h and 24 h) following HVACP (70 kV) treatment on viability of non-acid-adapted (A) and acid adapted (B) Escherichia coli O157:H7 survivors in pineapple juice. Survivors were recovered on thin agar layer (TAL) medium. For each treatment time, different letters (A or B) over the bars indicate a significant difference in numbers of survivors at 0.1 h and 24 h post-treatment (P<0.05).
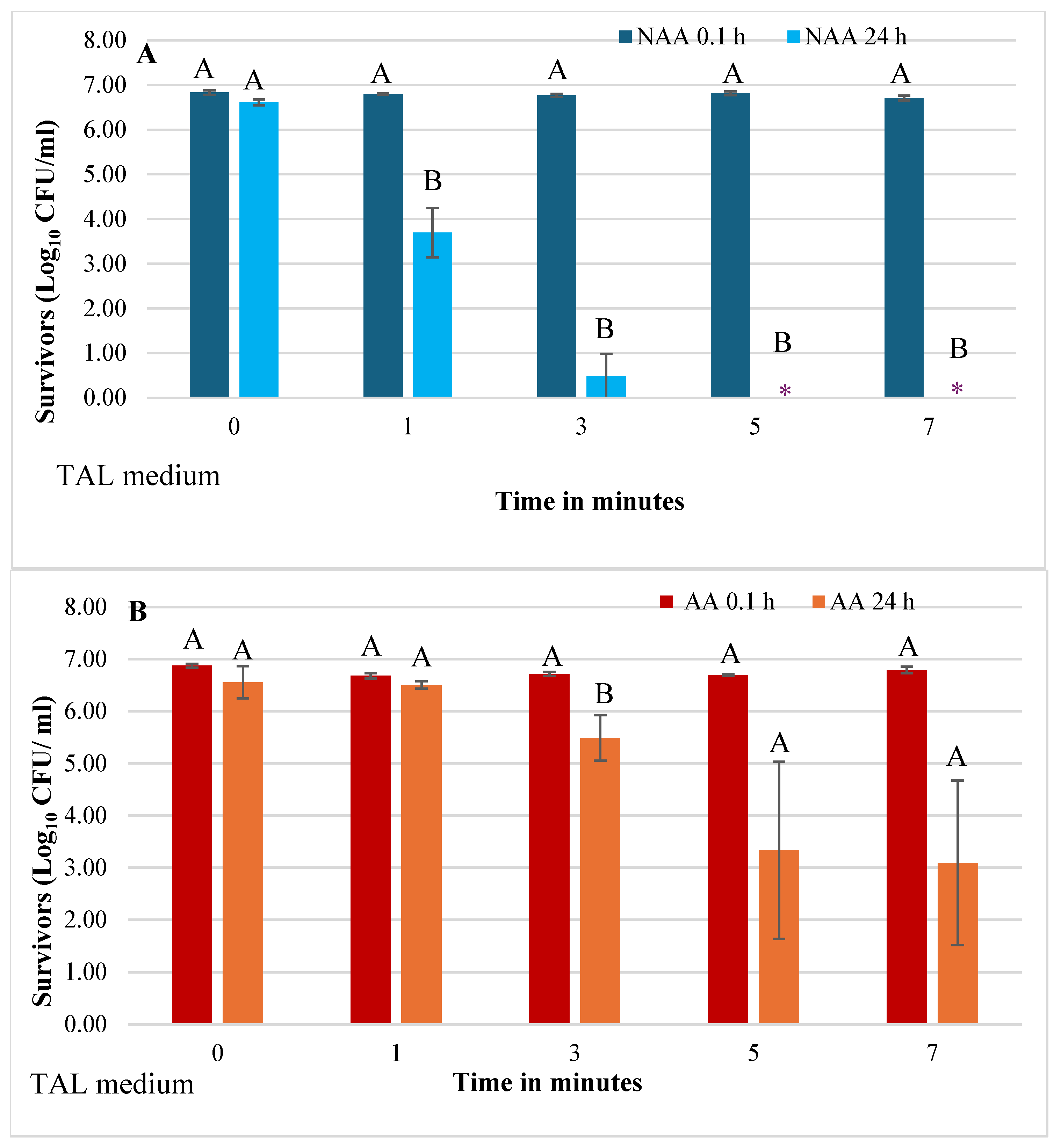
Figure 4.
Effect of physiological state (NAA and AA) on survival of Escherichia coli O157:H7 in raw pineapple juice held at 4 ᵒC for 24 h after HVACP (70 kV) treatment. Survivors were recovered on sorbitol MacConkey (SMAC) agar (A) and TAL medium (B). For each treatment time, different first letters (A or B) over the bars indicate a significant difference in viable counts of NAA and AA cells (P<0.05).
Figure 4.
Effect of physiological state (NAA and AA) on survival of Escherichia coli O157:H7 in raw pineapple juice held at 4 ᵒC for 24 h after HVACP (70 kV) treatment. Survivors were recovered on sorbitol MacConkey (SMAC) agar (A) and TAL medium (B). For each treatment time, different first letters (A or B) over the bars indicate a significant difference in viable counts of NAA and AA cells (P<0.05).

Table 1.
*D-values (min) of non-acid-adapted (NAA) and acid adapted (AA) Escherichia coli O157:H7 following exposure to HVACP treatment of pineapple juice.
Table 1.
*D-values (min) of non-acid-adapted (NAA) and acid adapted (AA) Escherichia coli O157:H7 following exposure to HVACP treatment of pineapple juice.
| Physiological state | SMAC agar | TAL Medium |
| NAA | 0.57 ± 0.12 Ax | 0.57 ± 0.06 Ax |
| AA | 3.03 ± 2.87 Ax | 3.43 ± 3.18 Ax |
*Each D-value (in minutes) represents the mean (± standard deviation) of four replications of the experiment. D-values were derived from the slopes of survivor curves based on bacterial colony counts (log10 CFU/mL) on SMAC agar and TAL medium. Survivors were recovered from juice (4 °C) at 24 h after HVACP treatment.
Table 2.
Sub-lethal injury (based on linear regression parameters*) in survivors of non-acid-adapted (NAA) and acid-adapted (AA) Escherichia coli O157:H7 in pineapple juice treated with HVACP (70kv) and recovered at 0.1 h and 24 h after HVACP treatment.
Table 2.
Sub-lethal injury (based on linear regression parameters*) in survivors of non-acid-adapted (NAA) and acid-adapted (AA) Escherichia coli O157:H7 in pineapple juice treated with HVACP (70kv) and recovered at 0.1 h and 24 h after HVACP treatment.
| Physiological state | Time | Slope | y-intercept | R2 |
| NAA | 0.1 h | 0.06±0.55 | 0.056±0.05 | 0.154±0.097 |
| NAA | 24 h | 0.983±0.01 | 0.093±0.03a | 0.997±0.002 |
| AA | 0.1 h | 0.73±0.51 | 0.063±0.05 | 0.417±0.33 |
| AA | 24 h | 1.01±0.12 | 0.28±0.02a | 0.942±0.07 |
* Values are averages ± standard deviations from three replications of the experiment; a Intercept significantly different from 0 (P < 0.05).
Table 3.
The pH and degrees Brix of pineapple juice at 0.1h and 24h after treatment with HVACP (70 kV).
Table 3.
The pH and degrees Brix of pineapple juice at 0.1h and 24h after treatment with HVACP (70 kV).
| pH | °Brix | |||
|---|---|---|---|---|
| Time (min) | pH; 0.1h | pH; 24h | ᵒBrix; 0.1h | ᵒBrix; 24h |
| 0 | 3.36±0.03 A,x | 3.36±0.03 A,x | 14.3±0.00 A,x | 14.5±0.00 A,y |
| 3 | 3.35±0.04 A,x | 3.35±0.02 A,x | 14.4±0.00 B,x | 14.5±0.00 A,y |
| 7 | 3.33±0.01 A,x | 3.34±0.01 A,x | 14.4±0.00 B,x | 14.5±0.00 A,y |
* Values are averages ± standard deviations from four replications of the experiment. Different uppercase letters (A, B) within a column indicate significant differences in pH or ᵒBrix (p<0.05). Different lowercase letters (x, y) within a row indicate significant differences (p<0.05) for pH or Brix taken at 0.1 h or 24 h at specific treatment times (0, 3, or 7 min).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



