Submitted:
17 May 2024
Posted:
23 May 2024
You are already at the latest version
Abstract
Introduction: Transforming growth factor beta receptor 2 (Tgfbr2) signaling regulates odontoblast differentiation, dentin mineralization, and the paracrine signaling that guides sensory innervation in developing mouse molars. We hypothesized that Tgfbr2 also regulates the neuro-pulpal responses to injury to activate axon sprouting and tertiary dentin secretion. Methods: We per-formed a timed deletion of Tgfbr2 using tetracycline-responsive Osterix-Cre;Tgfbr2f/f (Tgfbr2^cko) mice, which allowed them to develop normally. At 3 months, we drilled a shallow hole on the mesial side of the first mandibular molar (M1) in both control and Tgfbr2^cko mice. Hemi-mandibles were collected 4, 8, 21, and 56 days post-injury (dpi). In situ hybridization (ISH) for Sp7 was performed to confirm Osterix-Cre transcription. Microcomputed tomography (micro-CT) imaging and histology were used to examine the tertiary dentin, and immunofluorescence was used to visualize CGRP+ axons following the injury. Results: Tgfbr2 deletion was inferred by Sp7 ISH. Micro-CT and histology indicated a lower dentin volume in 21 dpi Tgfbr2^cko M1s compared to WT M1s, but the volume was comparable by 56 dpi. The duration of axon sprouting was longer in injured Tgfbr2^cko M1s compared to WT M1s. Conclusions: These results suggest that sensory afferents may support dentin repair in Tgfbr2-deficient odontoblasts.
Keywords:
transforming growth factor beta
; calcitonin gene-related peptide
; tertiary dentin
; neuropeptide
; dental pulp cells
; pulp biology
1. Introduction
Odontoblasts secrete dentin, a mineralized layer of the tooth beneath the enamel, to encase and protect the soft pulp tissue inside of the tooth organ. Dentinogenesis occurs throughout the lifetime of an animal, with primary dentin laid down during tooth development, secondary dentin slowly secreted throughout adulthood, and tertiary dentin secreted in response to injury or infection [1,2]. The secretion and mineralization of dentin involves the TGF superfamily, predominantly mediated through TGFβ receptors I and II, which are expressed in both odontoblasts and dental pulp cells [3,4,5,6,7]. We previously reported that Tgfbr2 promotes the proliferation and differentiation of mineralizing cells required for bones and teeth to grow and mineralize [3,7,8,9]. If Tgfbr2 is deleted in mature odontoblasts, the cells lose their polarity, resulting in ectopic matrix formation, osteodentin formation, and even pulpal obliteration [5,10]. This prompted us to question whether the response to dentin injury also requires TGFβ signaling and whether the absence of Tgfbr2 would result in aberrant tertiary dentin formation. Additionally, sensory afferents sprout and provide peptidergic signals following injury of the dentin-pulp complex, and this mediates the healing process [11,12,13,14,15]. Several studies reported that CGRP moderates the neuroinflammatory response to dentin injury [11,14,16], yet the roles of CGRP in pulp repair, particularly in the absence of pulp exposure, are not yet fully understood. Since Tgfbr2 signaling in the dental pulp cells promotes axon sprouting during tooth development [3,17], we hypothesized that this could be recapitulated during dentin repair. In the present study, we evaluated whether Tgfbr2 deletion in the odontoblasts and dental pulp cells during a shallow dentin injury would impact CGRP+ axon sprouting and tertiary dentin secretion.
2. Results
2.1.1. Tgfbr2cko model characterization
To confirm that dentin injury did not affect eating behaviors or body weight, which could affect tooth healing, mice were weighed pre- and post-surgery. We found no significant differences in weights between WT and Tgfbr2cko male and females (Figure 1E).
Mice were raised from in utero until 2 weeks prior to surgery with doxycycline-infused chow. To infer the activity of Cre after transitioning to standard chow, we performed ISH for Sp7 (Osterix) on injured and control teeth. ISH revealed active transcription of Sp7, indicating that there was Cre expression and therefore conditional deletion Tgfbr2 in the Tgfbr2cko mice (Figure 1F-G).
2.1.2. Tertiary dentin deposition is delayed in Tgfbr2cko M1s
We did not find evidence of tertiary dentin at 4 and 8 dpi irrespective of mouse genotype (Figure 2).
While we found tertiary dentin in injured WT molars at 21 dpi, we did not find any in Tgfbr2cko molars (Figure 3 B,E and H, K). The WT molars did not demonstrate increases in tertiary dentin beyond the 21 dpi time point (Figure 3 B,C, and H,I). Intriguingly, we found tertiary dentin in the Tgfbr2cko mice at 56 dpi similar to what we saw in the WT mice (Figure 3 C, F and I, L). Histological staining depicted polarized odontoblasts in both uninjured and injured molars in both genotypes, with tubular tertiary dentin. Masson’s trichrome generally stained the outer, or more external, regions of the dentin in Biebrich scarlet and the inner regions in aniline blue at the cemento-enamel junction. The regions of tertiary dentin were also stained in Biebrich scarlet.
We also performed micro-CT to quantify changes in dentin between the genotypes within the injury area. We did not find any evidence of tertiary dentin until 21 dpi. At 21 dpi, we found differences in dentin volume within the ROI between the WT and Tgfbr2cko mice (Figure 4 B, E, G; p<0.001). Intriguingly, we found that the tertiary dentin volume levels in the Tgfbr2cko mice at 56 dpi were equivalent to what was present in the WT at 21 and 56 dpi (Figure 4 B, C, E, F, G; p<0.05). We did not find any changes in dentin density (Appendix A, Figure A1) nor differences between sexes (data not shown).
2.1.3. CGRP+ axon sprouting is elevated in Tgfbr2cko M1s
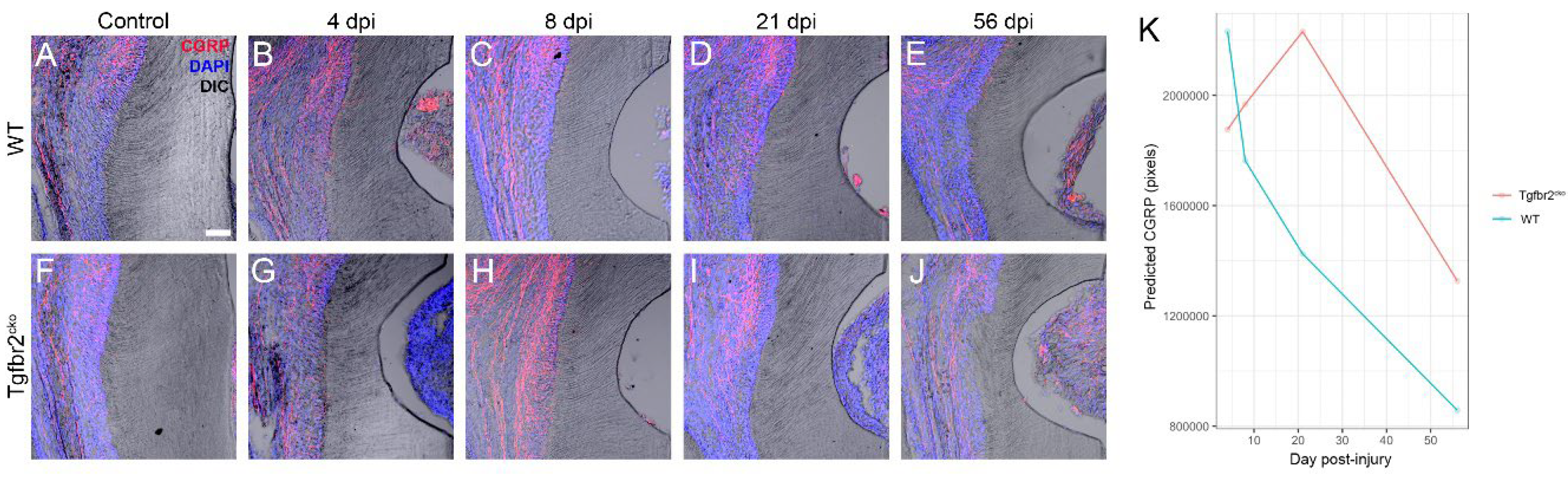
We imaged and quantified the pixel density of CGRP+ axons in uninjured and 4, 8, 21, and 56 dpi M1s from WT and Tgfbr2cko mice to determine the extent of peptidergic axon sprouting in response to injury (Figure 5A-J, Figure A2). The analysis was based on imaging CGRP with confocal microscopy and converting the z-stacks into a maximum projection for 20 μm sections in the area of the injury. Similar areas were chosen for control samples. The zstacks were autothresholded and pixel density of CGRP was then calculated with Image J for control areas for 4, 8, and 21 time points for both genotypes. We then utilized generalized estimating equations (GEE) [18] (Figure 5K, Table A1) to analyze the data while accounting for the repeated measures. Model prediction results of CGRP are presented based on the genotype, injury status, and time point, and a detailed statistical analysis is included in Appendix A, Table A1. We found that the predicted CGRP at 4 dpi was not statistically different between WT and Tgfbr2cko M1s (Figure 5B, G). However, the change in CGRP from 4 to 21 dpi differed between the two genotypes for injured mice (p=0.004), with the predicted model indicating a slower decline in CGRP in the injured Tgfbr2cko mice, as shown by the red line in Figure 5K. By 56 dpi, the CGRP seems to stabilize in both injured genotypes. No significant differences were observed in CGRP+ sprouting by sex across all time points and genotypes (data not shown). Our results suggest that CGRP sprouting in response to injury remains elevated longer in Tgfbr2cko compared to WT molars and predicts a different healing trajectory.
3. Discussion
We previously reported that Tgfbr2 in dental pulp fibroblasts and odontoblasts regulates the postnatal neuro-pulpal development of mouse molars [3,7,9]. A shallow dentin injury without pulp exposure has been shown to initiate the expression of neurotrophic factors, such as nerve growth factors (NGF), by odontoblasts [5,6,7]. This triggers a neuroinflammatory response in which sensory afferents sprout and secrete CGRP [8,9] to activate odontogenic activities for dentin repair [11,14]. In this report, we sought to determine whether the signaling downstream of Tgfbr2 in the developing dental pulp mesenchyme also regulates reactionary dentinogenesis and odontoblast secretion of neurotrophic signals that promote axon sprouting. Since conditional deletion of Tgfbr2 in the Osteocalcin-Cre model results in pulpal obliteration [5], and deletion in the Osterix-Cre model results in late postnatal death [3,7,8,9], we utilized the tetracycline-responsive element in the Osterix-Cre mouse model to isolate conditional deletion of Tgfbr2 to the time period immediately before the dentin injury through the studies on the injury response. Our ISH of Sp7 indicated that there was Tgfb2 deletion in the dental pulp, confirming the validity of our model. Unexpectedly, we found that while tertiary dentin secretion was delayed in the Tgfbr2cko molars, the levels of tertiary dentin were eventually equivalent to those demonstrated in WT control mice. In addition, the axon sprouting in Tgfbr2cko mice was equivalent to that in the WT mice at 4 dpi but remained elevated for a longer period of time following injury than in the WT mice. Even though recent research has demonstrated CGRP signaling differs between male and female models of pain, we did not find a difference in CGRP+ axon sprouting between the sexes [19], indicating sex did not play a primary role in reactionary dentinogenesis in our models. Together, our results suggest that the sensory afferents may have secreted CGRP for a longer period in the Tgfbr2cko mice to assist with dentin repair.
Research showing that CGRP+ sensory afferents sprout during dentin injury was first published more than three decades ago [13], but conflicting data have recently emerged regarding the involvement of peptidergic signaling from the sensory neurons in reactionary and/or reparative dentinogenesis. One group reported that CGRP has minimal effects on the expression of differentiation markers in dental pulp stem cells (DPSCs) in vitro unless paired with Sonic Hedgehog (Shh) [14]. However, they also reported that CGRP alone promotes DPSC mineralization [20]. One group found that applying CGRP promoted pulpal healing in ferrets, with increased levels of tertiary dentin and osteodentin [16]. A different group reported that CGRP inhibits mineralization [21]. It should be noted that these reports experimented directly on DPSCs, while we and others have shown an intimate, bi-directional communication between the DPSCs and trigeminal neurons [11,15,22,23,24,25,26,27,28,29,30]. Interestingly, bone research has also found that sensory afferents assist bone healing via CGRP signaling. For instance, bone healing around orthopedic implants can be attenuated with denervation of sensory nerves or knockdown of the CGRP receptors genes, Calcrl or Ramp1. Conversely, upregulating the receptors with adenoviruses to overexpress them (AdV-Calcrl) enhanced osteogenesis [31]. Another group observed increased CGRP around 3 days after a femoral fracture in mice, similar to the CGRP spike found in molars [13]. However, the CGRP receptors were not upregulated until 1-2 weeks post injury. This indicated that the increased expression of CGRP and its receptors during bone healing were not aligned [32]. A follow-up study performed an ACL reconstruction where they injected hydrogel microparticles loaded with adenoviruses to silence or over-express the CGRP receptors (adv-shCalcrl or adv-Calcrl) in the bone tunnels. Their results showed striking reductions/increases in mineralization markers and bone volume following the silencing/overexpression of the CGRP receptors, similar to the findings of the orthodontic implant study [31]. This demonstrated that increasing CGRP receptors at the earlier timepoints when CGRP levels peak dramatically improves bone healing [33]. We hypothesize that the prolonged peak of CGRP in our Tgfbr2cko mice was present when the receptor expression levels would be higher, which allowed for additional mineralization signals in the absence of Tgfbr2. Future studies investigating CGRP receptors directly in conjunction with CGRP signaling in the pulp will be important to confirm this hypothesis.
We previously demonstrated that several members of the semaphorin family (SEMAs) in the dental pulp regulate neurite outgrowth in developing molars [3]. It is possible that changes in the odontoblast expression levels of these chemoattractants in our Tgfbr2cko mice could be directly altering, i.e. prolonging, the neurite outgrowth during the injury response. When recombinant SEMAs were applied to mineralized tissues, it led to a range of results, from aggravating existing periapical lesions [34] to promoting reparative dentin formation in pulp capping experiments [35] and bone healing in fracture calluses and calvarial defects [36,37]. In addition, SEMA-regulated bone repair has been shown to be driven by the sensory nerves, rather than osteoblasts [38,39], indicating that there is complex neuronal-mesenchymal crosstalk during repair. Mouse models of different semaphorin deletions in mineralizing and innervating populations of the dental pulp should be evaluated to fully address the roles these chemoattractants play during reactionary and/or reparative dentinogenesis and the associated sensory afferent signals.
It is common practice to etch dentin during endodontic procedures, but etching and some pulp capping materials can induce cellular damage and hypersensitivity [40,41,42,43,44,45,46]. In mouse dentin injuries, etching can cause severe damage and hyperactive responses that do not replicate the scenarios one would encounter in a clinical setting [11]. In our experiment, we used a low-speed drill to prevent high heat during the procedure and did not etch the dentin afterward. This protected the nearby axon terminals and underlying odontoblast layer, as evidenced in our confocal images of the afferents and histological images of the odontoblast layers. In addition, axonal coverage can drastically vary between ROIs, and a thin histological section could misrepresent the true nature of the wound response. Therefore, we based our quantifications on confocal imaging of the CGRP+ axon sprouting of two consecutive, 20-µm-thick sections surrounding the injured area. Since our micro-CT analyses indicated that the injury spans approximately 90 µm deep, our analysis represents the sprouting that covered almost half of the injured area. Our surprising results indicate that more comprehensive monitoring of the areas of interest and analyzing longer healing timelines are important to advance our knowledge of how the pulp tissue responds to injury. We also suggest that etching and the use of toxic pulp capping materials, such as calcium hydroxide, be avoided whenever possible in dental procedures because these may impede repair processes.
Previous reports suggested that inhibiting TGFβ receptor signaling disrupts the odontoblast layer and dentin secretion, leading to osteodentin [5,10] rather than tubular dentin. Here, we show that tubular dentin secretion is possible from TGFβ-deficient cells. Interestingly, two reports showed that less porous dentin stains scarlet [47,48], which we saw in the outer regions of dentin and tertiary dentin that were not evident in the H&E images. By focusing on the levels of red and blue in tooth healing, it would be possible to better ascertain the dentin integrity and porosity and perform more subtle investigations of the dentin-pulp complex, such as in pulp regeneration with implanted bone marrow stem cells or aged teeth. Our initial results from 21 dpi would have been misleading had we not additionally collected samples at 56 dpi based on previous reports indicating that tertiary dentin is sometimes not seen until then in C57/B6 mice [49]. These histological and micro-CT analyses again strongly support longer term studies in regenerative endodontics to fully assess whether healing is prevented or simply delayed. This is particularly relevant to our study and to dental clinicians given the research showing that TGFβ signaling plays a large role in regulating the dentin-pulp complex [3,5,6,9,50] and that disrupted TGFβ signaling results in oral-dental anomalies [51,52] and delayed oral wound healing [53]. It is important to note that the dentin proteome between the Tgfbr2cko and WT mice should be nearly identical due to the Cre recombinase suppression until the experimental timeline. The dentin proteome has been shown to promote neurite outgrowth [54,55] and to stimulate odontoblast differentiation and/or tertiary dentin secretion [56,57]. These growth factors in dentin can be released from the dentin into the pulp during endodontic treatments [58,59,60,61,62,63] and are being studied for their potential in regenerative endodontics. It is possible that the dentin similarities between our Tgfbr2cko and WT mice may be masking a differential response that would occur if the proteomes were different. Since we did not apply acid or EDTA to release dentin proteins, future studies will be required to confirm this. However, in order to develop better treatments for patients with skeletal defects due to disrupted/deficient TGFβ signaling, such as patients with Loeys-Dietz Syndrome, Marfan Syndrome [52,64,65] or diabetes [66,67,68], future investigations should focus on developing models where the fibroblasts and the dentin are both modified.
4. Materials and Methods
4.1. Mice
All mouse experiments were approved by the OSU Institutional Animal Care and Use Committee. The Tgfbr2cko mice were described previously [3,7,8]. To inhibit Osx-Cre activity, Tgfbr2cko breeding cages were maintained on a doxycycline-enhanced diet (7012, 1 g/kg, TD.08826, Teklad). Tgfbr2cko mice were moved to standard chow two weeks before creating a shallow dentin injury (Figure 1A). Wild type (WT) C57BL/6J mice were maintained on standard chow.
4.2. Dentin Injury and Mandible Harvest
Three-month-old mice were anesthetized with isoflurane, and with the aid of an endodontic microscope (Enova Illumination, Minneapolis, MN), the cemento-enamel junction (CEJ) of the mesial mandibular first molar (M1; Figure 1B-D) was exposed. A shallow dentin injury was then created with a low-speed handpiece fitted with a #1/16 carbide round bur (Komet, H1.314.003). At 4, 8, 21, and 56 days post-injury (dpi), the mice were injected with a ketamine/xylazine cocktail (1 g/10 g body weight; 25 mg/ml ketamine, 2.5 mg/ml xylazine, Covetrus North America), weighed, and transcardially perfused with cold PBS followed by 4% paraformaldehyde (PFA) to collect mandibles. For mice collected on 4-21 dpi, the right mandible was injured and the left mandible served as a contralateral control. For those sacrificed at 56 dpi, the right and left mandibles were both injured for separate experiments. Dissected mandibles were post-fixed in 4% PFA for 2 hours at room temperature.
4.3. In Situ Hybridization (ISH) and Histological Staining
Paraffin-embedded tissues were sectioned coronally at 7 µm in preparation for ISH or histological staining. Active Sp7 transcription was identified by ISH, which was performed using the RNAscope RED kit (ACD, 322373) and an Sp7 probe (ACD, 403401) per the manufacturer’s instructions. The presence of Sp7 transcripts confirmed the Tgfbr2 deletion in Osterix-Cre+ mice. Hematoxylin and eosin (H&E) and Masson’s Trichrome stains were applied according to standard protocols.
4.4. Micro-CT Scanning and Analysis
Fixed hemi-mandibles were stored and scanned in 70% ethanol with a µCT 50 (Scanco Medical, Bassersdorf, Switzerland) at 70 kVp, 76 µA, with a 0.5 mm Al filter, 900-ms integration and 6 µm voxel dimension. DICOM files were calibrated to a standard curve calculated from five known densities of hydroxyapatite (mg/cm3 HA). The mandibles were oriented with the mid-sagittal plane of the tooth. The first mandibular molar’s dentin was segmented at 500-600 mg/cm3 HA and enamel was segmented at 1600 mg/cm3 HA. This segmentation map was applied for n=5-6 sample size. The control mandibles were analyzed as described, and their respective segmentation maps were applied. Reconstructed images were analyzed using Analyze 14.0 (AnalyzeDirect, Overland Park, KS), as described previously [69,70]. A region of interest (ROI) was created to encompass the largest region of tertiary dentin found in any of the samples (90 µm total) and this was applied to all injured samples. Control images are shown for reference.
4.5. Immunofluorescence, Confocal Imaging, and Image Analysis
Hemi-mandibles were decalcified in 10% EDTA and prepared for cryo-embedding as described previously [3]. Twenty-micron sections were permeabilized using 0.5% Tween-20 blocking solution, followed by application of the primary antibody (anti-calcitonin gene-related peptide [CGRP]; Immunostar, 24112, 1:2000), secondary antibody (goat anti-rabbit IgG, Invitrogen, 1:500), and DAPI. Negative control experiments without primary antibodies did not yield significant staining (data not shown).
Confocal microscopy was used to image CGRP fluorescence in Z-stacks (20 µm) in the area of injury and similar areas for uninjured (control) molars. The results were converted to maximum projection images for analysis using ImageJ (National Institutes of Health, Bethesda, Maryland). CGRP confocal images were then converted to 8-bit, auto-thresholded with the mean option, and the injury area (or similar area in control molars) was isolated for pixel quantification. When possible, two serial sections per animal (40 µm total) were used for quantification and statistical analyses.
4.6. Statistical Analyses
A two-way mixed effects analysis of variance (ANOVA) with Šídák’s multiple comparisons was used to assess weight changes after injury, and differences in dentin deposition in micro-CT were evaluated with a two-way mixed effects ANOVA with Tukey’s multiple comparisons using GraphPad Prism 10.2.2 (GraphPad Software, www.graphpad.com). Significance was set at 5% for both ANOVAs.
Generalized estimating equations (GEE) [18] were used to analyze CGRP sprouting over all time points for both injured and control mice, controlling for genotype (Tgfbr2cko or WT) and accounting for repeated measurements resulting from evaluating two sections from each sample. Since only injured M1s were investigated at 56 dpi, injured M1s were considered the reference group for modeling (See Supplemental Table 1). Significance from estimated effects was tested using a Wald test with a 5% significance level.
5. Conclusions
The aim of this study was to determine if the neuro-pulpal crosstalk regulated by Tgfbr2 in the dental pulp fibroblasts was recapitulated during dentin regeneration, i.e. reactionary dentinogenesis. To this end, we performed a timed deletion of Tgfbr2 in the dental pulp mesenchyme immediately prior to a shallow dentin injury and quantified the levels of tertiary dentin and CGRP+ axon sprouting. Micro-CT analysis, H&E images, and Masson’s trichrome images all demonstrate that tertiary dentin secretion was delayed in the Tgfbr2cko mice, but reached equivalence to the WT controls by two months. CGRP+ axon sprouting levels were similar at 4 days but were elevated for longer duration in the Tgfbr2cko mice. This was in striking conflict with the developmental studies showing Tgfbr2 deletion resulted in severely hypomineralized and hypoinnervated dentition and instead suggests a compensation by the peripheral nervous system to assist healing. Given the current debates about the role(s) of CGRP in bone, tooth, and overall health, we believe subsequent studies should address the CGRP ligand-receptor complex during injury of the dentin-pulp complex. These investigations are likely to provide information that will lead to better long-term outcomes for dental restorations and regenerative endodontics.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org.
Author Contributions
The following statements should be used “Conceptualization, MS, FS, SBP.; methodology, MS, FF, AH, KK, DK, IK, NM, FS, SBP.; software, MS, KK, NM, FS, SBP.; validation, MS, FS, SBP.; formal analysis, FS, SBP.; investigation, MS, SBP.; resources, SBP.; data curation, MS, SBP.; writing—original draft preparation, MS, SBP.; writing—review and editing, MS, FS, SBP.; visualization, MS, KK, NM, FS, SBP.; supervision, FS, SBP.; project administration, SBP.; funding acquisition, SBP. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by NIH/NIDCR K99/R00 DE027706 and 3R00DE027706-05W1.
Acknowledgments
The authors thank Dr. Susan Travers for help in our experimental techniques and many discussions about the data. We especially thank Dr. Margie Byers for her assistance in optimizing the surgery and sample processing, as well as many theoretical discussions. BioRender was used to create Figure 1A.
Conflicts of Interest
The authors declare no conflicts of interest.
Appendix A
Figure A1.
Micro-CT analysis of dentin density. No differences in dentin density were found at 21 or 56 dpi between WT and Tgfbr2cko M1s.
Figure A1.
Micro-CT analysis of dentin density. No differences in dentin density were found at 21 or 56 dpi between WT and Tgfbr2cko M1s.
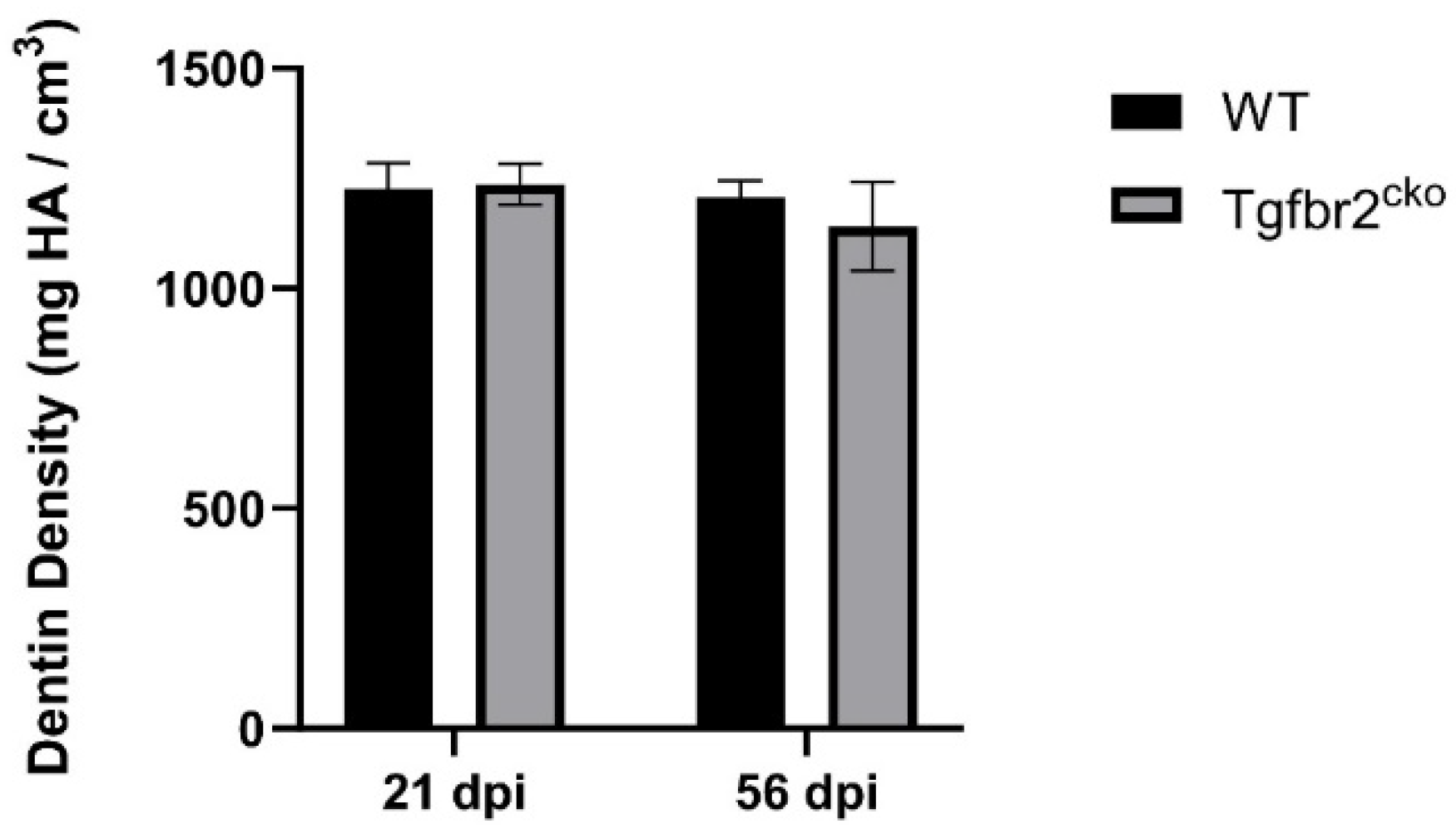
Figure A2.
Isolated CGRP+ axon sprouting in response to injury. Timeline of isolated CGRP+ axon outgrowth (white) in uninjured, and 4, 8, 21, and 56 dpi WT (A-E) and Tgfbr2cko (F-J) M1s, demonstrating divergent outgrowth patterns from 4-21 dpi between genotypes. Dotted yellow lines indicate areas of dentin injury. Scale bar in (A) = 50 μm.
Figure A2.
Isolated CGRP+ axon sprouting in response to injury. Timeline of isolated CGRP+ axon outgrowth (white) in uninjured, and 4, 8, 21, and 56 dpi WT (A-E) and Tgfbr2cko (F-J) M1s, demonstrating divergent outgrowth patterns from 4-21 dpi between genotypes. Dotted yellow lines indicate areas of dentin injury. Scale bar in (A) = 50 μm.
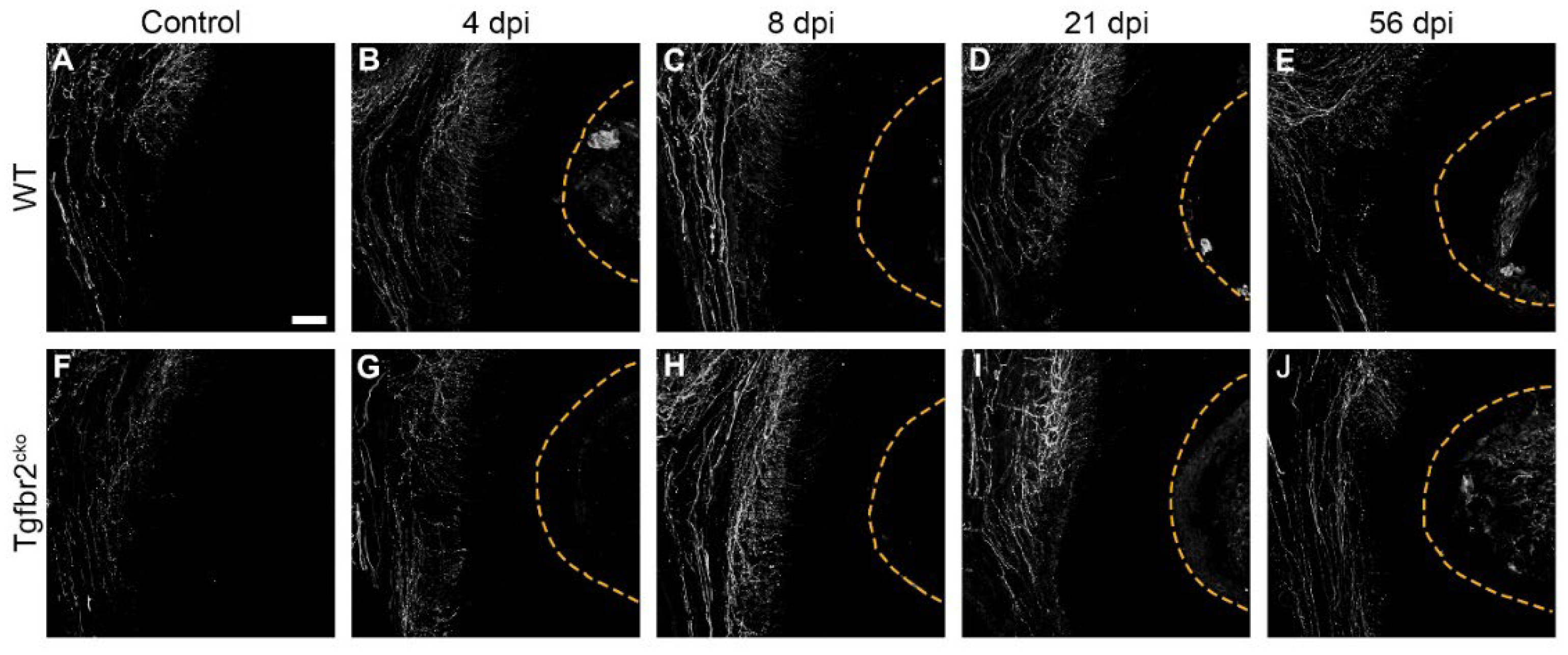

Table A1.
Results from the fitted GEE, accounting for repeated measures from evaluating two sections from each sample.
Table A1.
Results from the fitted GEE, accounting for repeated measures from evaluating two sections from each sample.
| term | estimate | std.error | statistic | p.value |
|---|---|---|---|---|
| Intercept | 1875690 | 199702 | 88.218 | <0.0001 |
| WT | 355742 | 272975 | 1.698 | 0.1925 |
| Ctl | -301958 | 302287 | 0.998 | 0.3178 |
| 8dpi | 92072 | 308701 | 0.089 | 0.7655 |
| 21dpi | 355390 | 335635 | 1.121 | 0.2897 |
| 56dpi | -548734 | 463553 | 1.401 | 0.2365 |
| WT:Ctl | -719330 | 412341 | 3.043 | 0.0811 |
| WT:8dpi | -560670 | 413896 | 1.835 | 0.1755 |
| WT:21dpi | -1161959 | 407350 | 8.137 | 0.0043 |
| Ctl:8dpi | -93531 | 414278 | 0.051 | 0.8214 |
| Ctl:21dpi | -18368 | 448516 | 0.002 | 0.9673 |
| WT:56dpi | -824827 | 521689 | 2.5 | 0.1139 |
| WT:Ctl:8dpi | 1354414 | 581367 | 5.428 | 0.0198 |
| WT:Ctl:21dpi | 921794 | 576407 | 2.557 | 0.1098 |
| Reference group: Tgfbr2cko, Day 4, injured | ||||
Generalized Estimating Equations (GEE) is a method for fitting (generalized) linear regression models to clustered data, which yields from having more than one measurement per mouse. By using the Huber-White method to estimate standard errors, valid inferential results are expected even when the working correlation is misspecified. In particular, we considered an identity link, normal variance, and working independence. The injured group was used as reference (as opposed to the control) since data from 56 dpi was only available for injured mice. However, results are equivalent since only two groups (injured and control) were considered. Overall, A:B denotes the interaction term between factor A and factor B, and A:B:C denotes the 3-way interaction between factors A, B, and C. The statistically significant findings are described below:
- WT:21dpi: The change from 4 dpi CGRP to 21 dpi CGRP in Wild Type mice is significantly different than the change from 4 dpi CGRP to 21 dpi CGRP in Tgfbr2cko mice. In other words, the effect of changing dpi from 4 to 21 is significantly different between genotypes, with a p-value of 0.00434.
- The difference in the change from Day 4 dpi CGRP to Day 8 dpi CGRP between Wild Type mice and Tgfbr2cko mice in control teeth is significantly different than the difference in the change from Day 4 dpi CGRP to Day 8 dpi CGRP between Wild Type mice and Tgfbr2cko mice in injured teeth, with a p-value of 0.0198.
Taken together, this indicates that due to the long time period in which the axon sprouting is equivalent at the beginning and at the end, we found many similarities between the CGRP levels. However, we found that the timeline from 4 dpi to 21 dpi was different between the genotypes, indicating a differential healing response via CGRP.
References
- Linde, A.; Goldberg, M. Dentinogenesis. Critical Reviews in Oral Biology & Medicine 1993, 4, 679.
- Goldberg, M.; Kulkarni, A.B.; Young, M.; Boskey, A. Dentin: Structure, Composition and Mineralization. Front Biosci (Elite Ed) 2011, 3, 711–735.
- Stanwick, M.; Barkley, C.; Serra, R.; Kruggel, A.; Webb, A.; Zhao, Y.; Pietrzak, M.; Ashman, C.; Staats, A.; Shahid, S.; et al. Tgfbr2 in Dental Pulp Cells Guides Neurite Outgrowth in Developing Teeth. Frontiers in Cell and Developmental Biology 2022, 0, 215. [CrossRef]
- Sloan, A.J.; Couble, M.L.; Bleicher, F.; Magloire, H.; Smith, A.J.; Farges, J.C. Expression of TGF-Beta Receptors I and II in the Human Dental Pulp by in Situ Hybridization. Adv Dent Res 2001, 15, 63–67. [CrossRef]
- Ahn, Y.H.; Kim, T.H.; Choi, H.; Bae, C.H.; Yang, Y.M.; Baek, J.A.; Lee, J.C.; Cho, E.S. Disruption of Tgfbr2 in Odontoblasts Leads to Aberrant Pulp Calcification. Journal of dental research 2015, 94, 828–835. [CrossRef]
- Niwa, T.; Yamakoshi, Y.; Yamazaki, H.; Karakida, T.; Chiba, R.; Hu, J.C.C.; Nagano, T.; Yamamoto, R.; Simmer, J.P.; Margolis, H.C.; et al. The Dynamics of TGF-β in Dental Pulp, Odontoblasts and Dentin. Scientific Reports 2018, 8. [CrossRef]
- Wang, Y.; Cox, M.K.; Coricor, G.; MacDougall, M.; Serra, R. Inactivation of Tgfbr2 in Osterix-Cre Expressing Dental Mesenchyme Disrupts Molar Root Formation. Developmental biology 2013, 382, 27–37. [CrossRef]
- Peters, S.B.; Wang, Y.; Serra, R. Tgfbr2 Is Required in Osterix Expressing Cells for Postnatal Skeletal Development. Bone 2017, 97, 54–64. [CrossRef]
- Corps, K.; Stanwick, M.; Rectenwald, J.; Kruggel, A.; Peters, S.B. Skeletal Deformities in Osterix-Cre;Tgfbr2f/f Mice May Cause Postnatal Death. Genes 2021, 12. [CrossRef]
- Neves, V.C.M.; Sharpe, P.T. Regulation of Reactionary Dentine Formation. Journal of Dental Research 2018, 97, 416–422. [CrossRef]
- Zhan, C.; Huang, M.; Chen, J.; Lu, Y.; Yang, X.; Hou, J. Sensory Nerves, but Not Sympathetic Nerves, Promote Reparative Dentine Formation after Dentine Injury via CGRP-Mediated Angiogenesis: An in Vivo Study. Int Endod J 2024, 57, 37–49. [CrossRef]
- Sarram, S.; Lee, K.F.; Byers, M.R. Dental Innervation and CGRP in Adult P75-Deficient Mice. The Journal of comparative neurology 1997, 385, 297–308.
- Taylor, P.E.; Byers, M.R.; Redd, P.E. Sprouting of CGRP Nerve Fibers in Response to Dentin Injury in Rat Molars. Brain research 1988, 461, 371–376. [CrossRef]
- Moore, E.R.; Michot, B.; Erdogan, O.; Ba, A.; Gibbs, J.L.; Yang, Y. CGRP and Shh Mediate the Dental Pulp Cell Response to Neuron Stimulation. Journal of dental research 2022, 101. [CrossRef]
- de Almeida, J.F.A.; Chen, P.; Henry, M.A.; Diogenes, A. Stem Cells of the Apical Papilla Regulate Trigeminal Neurite Outgrowth and Targeting through a BDNF-Dependent Mechanism. Tissue engineering. Part A 2014, 20, 3089–3100. [CrossRef]
- Kline, L.W.; Yu, D.C. Effects of Calcitonin, Calcitonin Gene-Related Peptide, Human Recombinant Bone Morphogenetic Protein-2, and Parathyroid Hormone-Related Protein on Endodontically Treated Ferret Canines. Journal of Endodontics 2009, 35, 866–869. [CrossRef]
- Barkley, C.; Serra, R.; Peters, S.B. A Co-Culture Method to Study Neurite Outgrowth in Response to Dental Pulp Paracrine Signals. Journal of visualized experiments : JoVE 2020. [CrossRef]
- Hardin, J.W.; Hilbe, J.M. Generalized Estimating Equations; 2nd ed.; Chapman and Hall/CRC: New York, 2013; ISBN 978-0-429-11103-7.
- Paige, C.; Plasencia-Fernandez, I.; Kume, M.; Papalampropoulou-Tsiridou, M.; Lorenzo, L.E.; David, E.T.; He, L.; Mejia, G.L.; Driskill, C.; Ferrini, F.; et al. A Female-Specific Role for Calcitonin Gene-Related Peptide (CGRP) in Rodent Pain Models. Journal of Neuroscience 2022, 42, 1930–1944. [CrossRef]
- B, M.; SM, C.; JL, G. Effects of Calcitonin Gene-Related Peptide on Dental Pulp Stem Cell Viability, Proliferation, and Differentiation. Journal of endodontics 2020, 46, 950–956. [CrossRef]
- Saito, N.; Kimura, M.; Ouchi, T.; Ichinohe, T.; Shibukawa, Y. Gαs-Coupled CGRP Receptor Signaling Axis from the Trigeminal Ganglion Neuron to Odontoblast Negatively Regulates Dentin Mineralization. Biomolecules 2022, 12, 1747. [CrossRef]
- Sultan, N.; Amin, L.E.; Zaher, A.R.; Grawish, M.E.; Scheven, B.A. Neurotrophic Effects of Dental Pulp Stem Cells on Trigeminal Neuronal Cells. Scientific Reports 2020 10:1 2020, 10, 1–13. [CrossRef]
- Pagella, P.; Miran, S.; Neto, E.; Martin, I.; Lamghari, M.; Mitsiadis, T.A. Human Dental Pulp Stem Cells Exhibit Enhanced Properties in Comparison to Human Bone Marrow Stem Cells on Neurites Outgrowth. The FASEB Journal 2020, 34, 5499–5511. [CrossRef]
- Zhan, C.; Huang, M.; Zeng, J.; Chen, T.; Lu, Y.; Chen, J.; Li, X.; Yin, L.; Yang, X.; Hou, J. Irritation of Dental Sensory Nerves Promotes the Occurrence of Pulp Calcification. Journal of Endodontics 2023. [CrossRef]
- Byers, M.R.; Swift, M.L.; Wheeler, E.F. Reactions of Sensory Nerves to Dental Restorative Procedures. Proc Finn Dent Soc 1992, 88 Suppl 1, 73–82.
- Woodnutt, D.A.; Wager-Miller, J.; O’Neill, P.C.; Bothwell, M.; Byers, M.R. Neurotrophin Receptors and Nerve Growth Factor Are Differentially Expressed in Adjacent Nonneuronal Cells of Normal and Injured Tooth Pulp. Cell and Tissue Research 2000, 299, 225–236. [CrossRef]
- Byers, M.R.; Wheeler, E.F.; Bothwell, M. Altered Expression of NGF and P75 NGF-Receptor by Fibroblasts of Injured Teeth Precedes Sensory Nerve Sprouting. Growth factors (Chur, Switzerland) 1992, 6, 41–52.
- Diogenes, A. Trigeminal Sensory Neurons and Pulp Regeneration. Journal of Endodontics 2020, 46, S71–S80. [CrossRef]
- Bowles, W.R.; Burke, R.; Sabino, M.; Harding-Rose, C.; Lunos, S.; Hargreaves, K.M. Sex Differences in Neuropeptide Content and Release from Rat Dental Pulp. J Endod 2011, 37, 1098–1101. [CrossRef]
- Buck, S.; Reese, K.; Hargreaves, K.M. Pulpal Exposure Alters Neuropeptide Levels in Inflamed Dental Pulp and Trigeminal Ganglia: Evaluation of Axonal Transport. J Endod 1999, 25, 718–721. [CrossRef]
- Zhang, Y.; Xu, J.; Ruan, Y.C.; Yu, M.K.; O’Laughlin, M.; Wise, H.; Chen, D.; Tian, L.; Shi, D.; Wang, J.; et al. Implant-Derived Magnesium Induces Local Neuronal Production of CGRP to Improve Bone-Fracture Healing in Rats. Nat Med 2016, 22, 1160–1169. [CrossRef]
- Appelt, J.; Baranowsky, A.; Jahn, D.; Yorgan, T.; Köhli, P.; Otto, E.; Farahani, S.K.; Graef, F.; Fuchs, M.; Herrera, A.; et al. The Neuropeptide Calcitonin Gene-Related Peptide Alpha Is Essential for Bone Healing. EBioMedicine 2020, 59. [CrossRef]
- Zhao, X.; Wu, G.; Zhang, J.; Yu, Z.; Wang, J. Activation of CGRP Receptor–Mediated Signaling Promotes Tendon-Bone Healing. Science Advances 2024, 10, eadg7380. [CrossRef]
- Wang, L.; Song, Y.; Yi, X.; Wu, C.; Guo, Q.; Zhou, X.; Song, D.; Zhang, L.; Huang, D. Semaphorin 7A Accelerates the Inflammatory Osteolysis of Periapical Lesions. Journal of Endodontics 2022, 48, 641-649.e2. [CrossRef]
- Yoshida, S.; Wada, N.; Hasegawa, D.; Miyaji, H.; Mitarai, H.; Tomokiyo, A.; Hamano, S.; Maeda, H. Semaphorin 3A Induces Odontoblastic Phenotype in Dental Pulp Stem Cells. Journal of Dental Research 2016, 95, 1282–1290. [CrossRef]
- Li, Y.; Yang, L.; He, S.; Hu, J. The Effect of Semaphorin 3A on Fracture Healing in Osteoporotic Rats. Orthop Sci 2015, 20, 1114–1121. [CrossRef]
- Kenan, S.; Onur, Ö.D.; Solakoğlu, S.; Kotil, T.; Ramazanoğlu, M.; Çelik, H.H.; Ocak, M.; Uzuner, B.; Fıratlı, E. Investigation of the Effects of Semaphorin 3A on New Bone Formation in a Rat Calvarial Defect Model. Journal of Cranio-Maxillofacial Surgery 2019, 47, 473–483. [CrossRef]
- Fukuda, T.; Takeda, S.; Xu, R.; Ochi, H.; Sunamura, S.; Sato, T.; Shibata, S.; Yoshida, Y.; Gu, Z.; Kimura, A.; et al. Sema3A Regulates Bone-Mass Accrual through Sensory Innervations. Nature 2013, 497, 490–493. [CrossRef]
- Mei, H.; Li, Z.; Lv, Q.; Li, X.; Wu, Y.; Feng, Q.; Jiang, Z.; Zhou, Y.; Zheng, Y.; Gao, Z.; et al. Sema3A Secreted by Sensory Nerve Induces Bone Formation under Mechanical Loads. Int J Oral Sci 2024, 16, 1–11. [CrossRef]
- Berkowitz, G.S.; Spielman, H.; Matthews, A.G.; Vena, D.; Craig, R.G.; Curro, F.A.; Thompson, V.P. Postoperative Hypersensitivity and Its Relationship to Preparation Variables in Class I Resin-Based Composite Restorations: Findings from the Practitioners Engaged in Applied Research and Learning (PEARL) Network. Part 1. Compend Contin Educ Dent 2013, 34, e44–e52.
- Younus, M.Z.; Ahmed, M.A.; Syed, A.U.Y.; Baloch, J.M.; Ali, M.; Sheikh, A. Comparison between Effectiveness of Dentine Desensitizer and One Bottle Self-Etch Adhesive on Dentine Hypersensitivity. Technol Health Care 2021, 29, 1153–1159. [CrossRef]
- Dammaschke, T. The History of Direct Pulp Capping. J Hist Dent 2008, 56, 9–23.
- Dammaschke, T.; Nowicka, A.; Lipski, M.; Ricucci, D. Histological Evaluation of Hard Tissue Formation after Direct Pulp Capping with a Fast-Setting Mineral Trioxide Aggregate (RetroMTA) in Humans. Clin Oral Investig 2019, 23, 4289–4299. [CrossRef]
- Kenneth M. Hargreaves / Harold E. Goodis / Franklin R. Tay (Editor) | Seltzer and Bender’s Dental Pulp Available online: https://www.quintessence-publishing.com/gbr/en/product/seltzer-and-benders-dental-pulp (accessed on 25 April 2024).
- Cox, C.F.; Sübay, R.K.; Ostro, E.; Suzuki, S.; Suzuki, S.H. Tunnel Defects in Dentin Bridges: Their Formation Following Direct Pulp Capping. Oper Dent 1996, 21, 4–11.
- Giraud, T.; Jeanneau, C.; Rombouts, C.; Bakhtiar, H.; Laurent, P.; About, I. Pulp Capping Materials Modulate the Balance between Inflammation and Regeneration. Dent Mater 2019, 35, 24–35. [CrossRef]
- Rabea, A.A. Assessment of Bone Marrow-Derived Mesenchymal Stem Cells Capacity for Odontogenic Differentiation and Dentin Regeneration in Methimazole-Treated Albino Rats (Light Microscopic Study). The Saudi Dental Journal 2022, 34, 27–35. [CrossRef]
- Rabea, A.A. Histological, Histochemical and Immunohistochemical Evaluation of the Role of Bone Marrow-Derived Mesenchymal Stem Cells on the Structure of Periodontal Tissues in Carbimazole-Treated Albino Rats. Archives of Oral Biology 2020, 119, 104887. [CrossRef]
- Frozoni, M.; Balic, A.; Sagomonyants, K.; Zaia, A.A.; Line, S.R.P.; Mina, M. A Feasibility Study for the Analysis of Reparative Dentinogenesis in pOBCol3.6GFPtpz Transgenic Mice. Int Endod J 2012, 45, 907–914. [CrossRef]
- Oka, S.; Oka, K.; Xu, X.; Sasaki, T.; Bringas, P.; Chai, Y. Cell Autonomous Requirement for TGF-Beta Signaling during Odontoblast Differentiation and Dentin Matrix Formation. Mechanisms of development 2007, 124, 409–415. [CrossRef]
- Yapijakis, C.; Davaria, S.; Gintoni, I.; Chrousos, G.P. The Impact of Genetic Variability of TGF-Beta Signaling Biomarkers in Major Craniofacial Syndromes. Adv Exp Med Biol 2023, 1423, 187–191. [CrossRef]
- Jani, P.; Nguyen, Q.C.; Almpani, K.; Keyvanfar, C.; Mishra, R.; Liberton, D.; Orzechowski, P.; Frischmeyer-Guerrerio, P.A.; Duverger, O.; Lee, J.S. Severity of Oro-Dental Anomalies in Loeys-Dietz Syndrome Segregates by Gene Mutation. Journal of Medical Genetics 2020, 57, 699–707. [CrossRef]
- Yamano, S.; Kuo, W.P.; Sukotjo, C. Downregulated Gene Expression of TGF-Βs in Diabetic Oral Wound Healing. Journal of cranio-maxillo-facial surgery : Official publication of the European Association for Cranio-Maxillo-Facial Surgery 2013, 41, e42-8. [CrossRef]
- Widbiller, M.; Austah, O.; Lindner, S.R.; Sun, J.; Diogenes, A. Neurotrophic Proteins in Dentin and Their Effect on Trigeminal Sensory Neurons. Journal of Endodontics2 2019, 45, 729–765. [CrossRef]
- Austah, O.; Widbiller, M.; Tomson, P.L.; Diogenes, A. Expression of Neurotrophic Factors in Human Dentin and Their Regulation of Trigeminal Neurite Outgrowth. Journal of Endodontics 2018, 45, 414–419. [CrossRef]
- Smith, A.J.; Cassidy, N.; Perry, H.; Bègue-Kirn, C.; Ruch, J.V.; Lesot, H. Reactionary Dentinogenesis. Int J Dev Biol 1995, 39, 273–280.
- Ivica, A.; Deari, S.; Patcas, R.; Weber, F.E.; Zehnder, M. Transforming Growth Factor Beta 1 Distribution and Content in the Root Dentin of Young Mature and Immature Human Premolars. Journal of Endodontics 2020, 46, 641–647. [CrossRef]
- Widbiller, M.; Eidt, A.; Lindner, S.R.; Hiller, K.-A.; Schweikl, H.; Buchalla, W.; Galler, K.M. Dentine Matrix Proteins: Isolation and Effects on Human Pulp Cells. International Endodontic Journal 2018, 51, e278–e290. [CrossRef]
- Galler, K.M.; Widbiller, M.; Buchalla, W.; Eidt, A.; Hiller, K.-A.; Hoffer, P.C.; Schmalz, G. EDTA Conditioning of Dentine Promotes Adhesion, Migration and Differentiation of Dental Pulp Stem Cells. International Endodontic Journal 2016, 49, 581–590. [CrossRef]
- Mohammadi, Z.; Shalavi, S.; Jafarzadeh, H. Ethylenediaminetetraacetic Acid in Endodontics. Eur J Dent 2013, 7, S135–S142. [CrossRef]
- Dos Reis-Prado, A.H.; Abreu, L.G.; Fagundes, R.R.; Oliveira, S. de C.; Bottino, M.C.; Ribeiro-Sobrinho, A.P.; Benetti, F. Influence of Ethylenediaminetetraacetic Acid on Regenerative Endodontics: A Systematic Review. Int Endod J 2022, 55, 579–612. [CrossRef]
- Graham, L.; Cooper, P.R.; Cassidy, N.; Nor, J.E.; Sloan, A.J.; Smith, A.J. The Effect of Calcium Hydroxide on Solubilisation of Bio-Active Dentine Matrix Components. Biomaterials 2006, 27, 2865–2873. [CrossRef]
- Ferracane, J.L.; Cooper, P.R.; Smith, A.J. Dentin Matrix Component Solubilization by Solutions at pH Relevant to Self-Etching Dental Adhesives. The journal of adhesive dentistry 2013, 15, 407–40712. [CrossRef]
- Singh, K.K.; Rommel, K.; Mishra, A.; Karck, M.; Haverich, A.; Schmidtke, J.; Arslan-Kirchner, M. TGFBR1 and TGFBR2 Mutations in Patients with Features of Marfan Syndrome and Loeys-Dietz Syndrome. Human Mutation 2006, 27, 770–777. [CrossRef]
- Loeys, B.L.; Chen, J.; Neptune, E.R.; Judge, D.P.; Podowski, M.; Holm, T.; Meyers, J.; Leitch, C.C.; Katsanis, N.; Sharifi, N.; et al. A Syndrome of Altered Cardiovascular, Craniofacial, Neurocognitive and Skeletal Development Caused by Mutations in TGFBR1 or TGFBR2. Nature Genetics 2005, 37, 275–281. [CrossRef]
- Jiao, H.; Xiao, E.; Graves, D.T. Diabetes and Its Effect on Bone and Fracture Healing. Curr Osteoporos Rep 2015, 13, 327–335. [CrossRef]
- Xu, M.T.; Sun, S.; Zhang, L.; Xu, F.; Du, S.L.; Zhang, X.D.; Wang, D.W. Diabetes Mellitus Affects the Biomechanical Function of the Callus and the Expression of TGF-Beta1 and BMP2 in an Early Stage of Fracture Healing. Braz J Med Biol Res 2015, 49, e4736. [CrossRef]
- Deng, Z.; Fan, T.; Xiao, C.; Tian, H.; Zheng, Y.; Li, C.; He, J. TGF-β Signaling in Health, Disease, and Therapeutics. Sig Transduct Target Ther 2024, 9, 1–40. [CrossRef]
- Nagasaki, A.; Nagasaki, K.; Kear, B.D.; Tadesse, W.D.; Thumbigere-Math, V.; Millán, J.L.; Foster, B.L.; Somerman, M.J. Delivery of Alkaline Phosphatase Promotes Periodontal Regeneration in Mice. J Dent Res 2021, 100, 993–1001. [CrossRef]
- Ao, M.; Chavez, M.B.; Chu, E.Y.; Hemstreet, K.C.; Yin, Y.; Yadav, M.C.; Millán, J.L.; Fisher, L.W.; Goldberg, H.A.; Somerman, M.J.; et al. Overlapping Functions of Bone Sialoprotein and Pyrophosphate Regulators in Directing Cementogenesis. Bone 2017, 105, 134–147. [CrossRef]
Figure 1.
Experimental timeline for dentin injury and model validation. (A) Timeline of dentin injury activities, beginning with doxycycline removal for Tgfbr2cko mice 14 days prior to dentin injury and the collection of tissue on specific days post-injury (dpi). (B-D) Creation of the dentin injury. Pre-injury (B), injury in process with a #1/16 carbide round bur (C), and post-injury (D), with an inset showing a higher magnification image of the injury (red arrows). (E) Percent weight change post-dentin injury in male and female WT and Tgfbr2cko mice at 4 dpi and 56 dpi. There was no significant difference in weight pre- to post-dentin injury in either genotype, sex, or dpi. (F-G) Sp7 (Osterix) in situ hybridization of 4 dpi Tgfbr2cko control (F) and injured (G) mice to confirm Osterix-Cre expression (N=4). Osterix was being actively transcribed in both control and injured Tgfbr2cko sections (black arrows). Insets show lower magnification of sections, with dotted lines outlining the main images of (F) and (G). Dentin injury is marked with the yellow * in the inset of (G). Od = odontoblasts, DP = dental pulp, D = dentin. Scale bars = 10 μm (F) and 100 μm (F inset).
Figure 1.
Experimental timeline for dentin injury and model validation. (A) Timeline of dentin injury activities, beginning with doxycycline removal for Tgfbr2cko mice 14 days prior to dentin injury and the collection of tissue on specific days post-injury (dpi). (B-D) Creation of the dentin injury. Pre-injury (B), injury in process with a #1/16 carbide round bur (C), and post-injury (D), with an inset showing a higher magnification image of the injury (red arrows). (E) Percent weight change post-dentin injury in male and female WT and Tgfbr2cko mice at 4 dpi and 56 dpi. There was no significant difference in weight pre- to post-dentin injury in either genotype, sex, or dpi. (F-G) Sp7 (Osterix) in situ hybridization of 4 dpi Tgfbr2cko control (F) and injured (G) mice to confirm Osterix-Cre expression (N=4). Osterix was being actively transcribed in both control and injured Tgfbr2cko sections (black arrows). Insets show lower magnification of sections, with dotted lines outlining the main images of (F) and (G). Dentin injury is marked with the yellow * in the inset of (G). Od = odontoblasts, DP = dental pulp, D = dentin. Scale bars = 10 μm (F) and 100 μm (F inset).
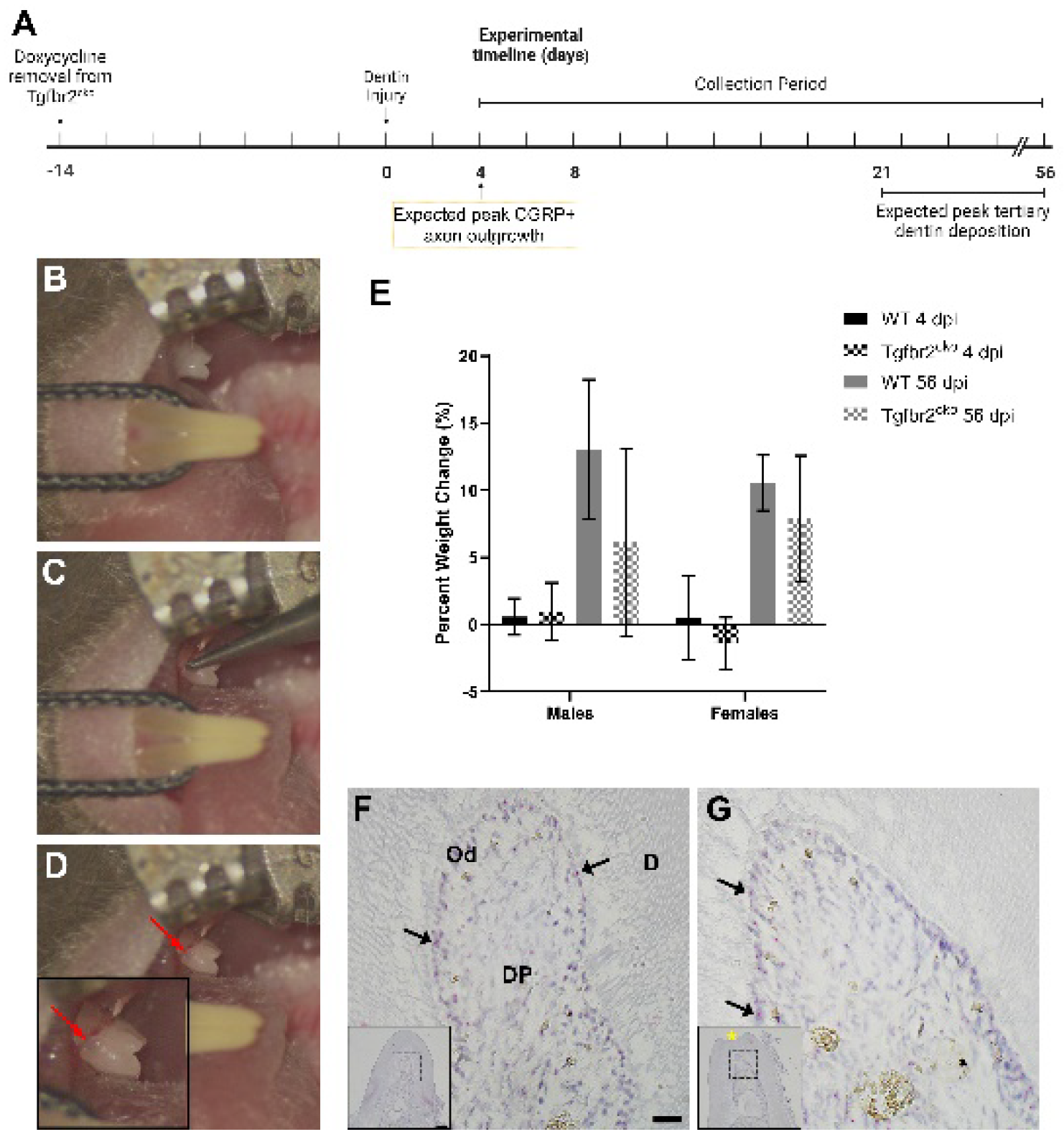
Figure 2.
Histological analyses of 4 and 8 dpi M1s. No tertiary dentin was present at 4 or 8 dpi in WT (A-C, G-I) or Tgfbr2cko (D-F, J-L) M1s as depicted by H&E and Masson’s trichrome staining. The yellow asterisks indicate the area drilled dentin injury. Scalebars in (A, G) = 50 µm.
Figure 2.
Histological analyses of 4 and 8 dpi M1s. No tertiary dentin was present at 4 or 8 dpi in WT (A-C, G-I) or Tgfbr2cko (D-F, J-L) M1s as depicted by H&E and Masson’s trichrome staining. The yellow asterisks indicate the area drilled dentin injury. Scalebars in (A, G) = 50 µm.
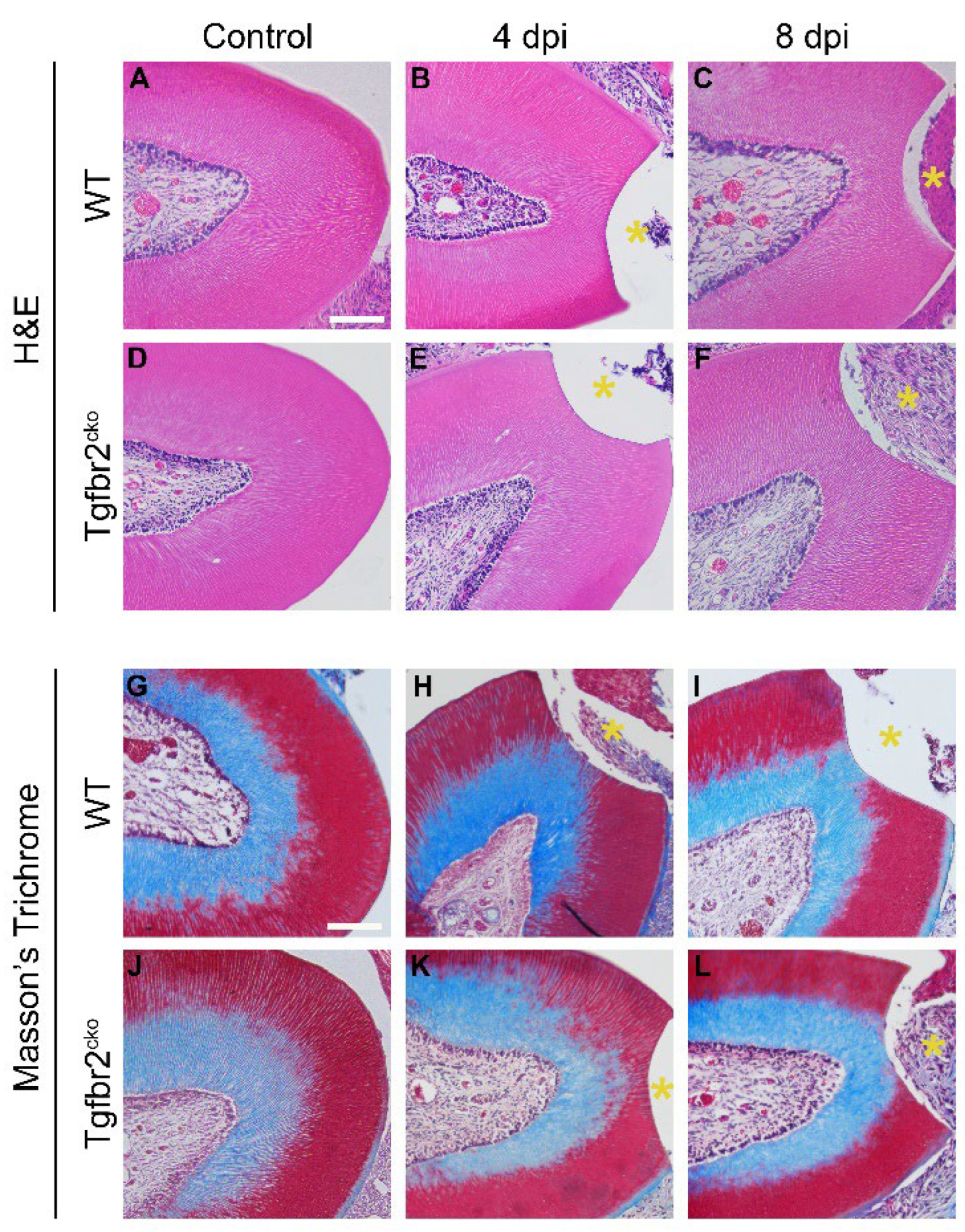
Figure 3.
Histology of tertiary dentin formation. (A-F) H&E and (G-L) Masson’s trichrome stained coronal sections (7 µm thick) of control, 21 dpi, and 56 dpi M1s. Tertiary dentin formation was not demonstrated at 21 dpi in the Tgfbr2cko mice (E, K) compared to WT mice (B, H). Comparable tertiary dentin was formed by 56 dpi in both the Tgfbr2cko (F, L) and WT (C, I) mice. The yellow asterisks indicate the area drilled dentin injury. White dotted lines demarcate the tertiary dentin border. Scale bar (shown in A) = 10 µm.
Figure 3.
Histology of tertiary dentin formation. (A-F) H&E and (G-L) Masson’s trichrome stained coronal sections (7 µm thick) of control, 21 dpi, and 56 dpi M1s. Tertiary dentin formation was not demonstrated at 21 dpi in the Tgfbr2cko mice (E, K) compared to WT mice (B, H). Comparable tertiary dentin was formed by 56 dpi in both the Tgfbr2cko (F, L) and WT (C, I) mice. The yellow asterisks indicate the area drilled dentin injury. White dotted lines demarcate the tertiary dentin border. Scale bar (shown in A) = 10 µm.
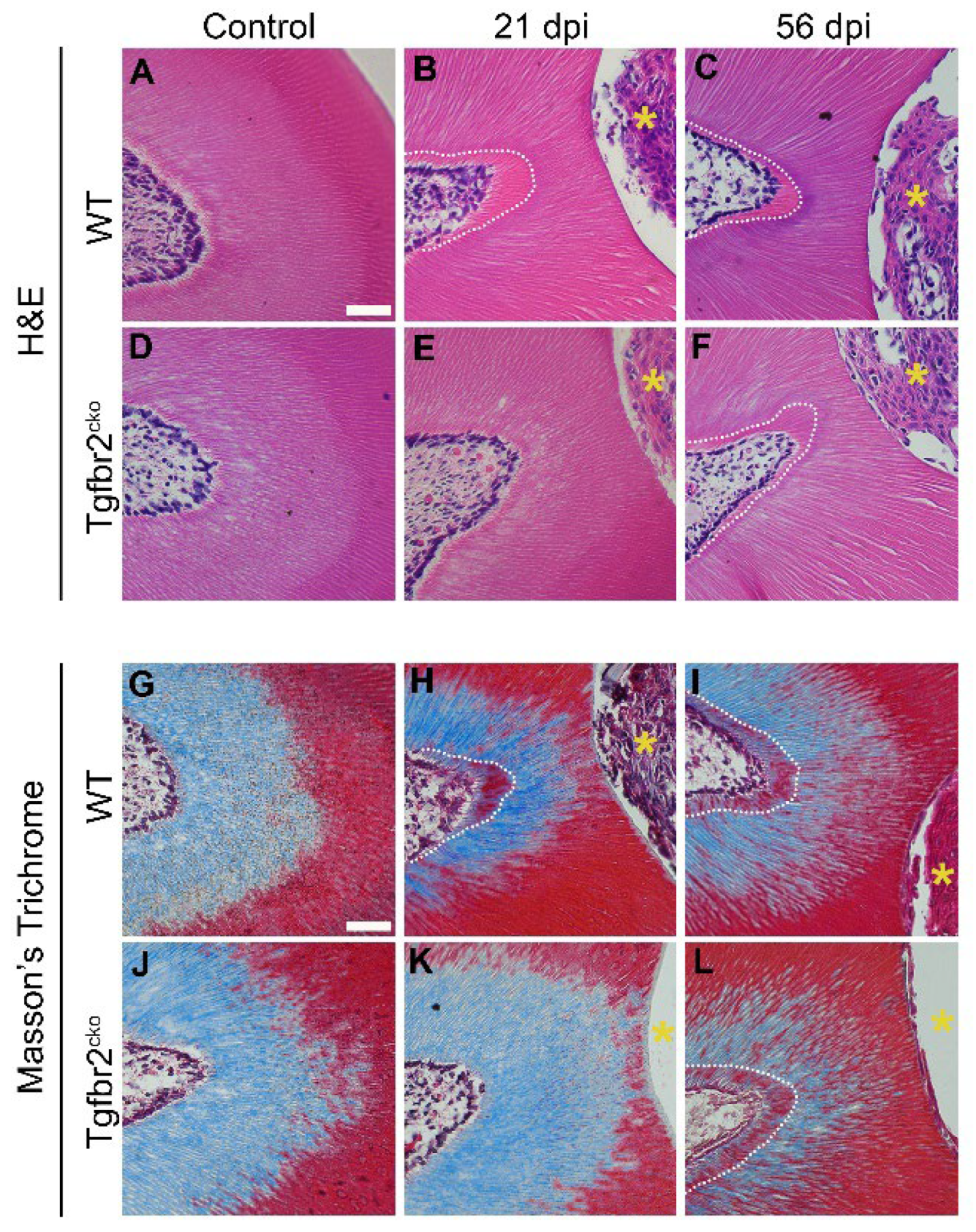
Figure 4.
Micro-CT analysis of tertiary dentin formation. (A-G) Micro-CT analysis (6 µm resolution) of control, 21 dpi, and 56 dpi M1s from all groups. Yellow dotted lines indicate inset images in (A-F) that represent the ROIs used for quantifications. There was a significantly lower dentin volume in Tgfbr2cko mice at 21 dpi compared to WT mice (B, E G), but at 56 dpi, the tertiary dentin volume was comparable between genotypes (C, F, G). Scale bars = 1 mm. In (G), * indicates p<0.05 and *** indicates p<0.001 by mixed effects 2-way ANOVA.
Figure 4.
Micro-CT analysis of tertiary dentin formation. (A-G) Micro-CT analysis (6 µm resolution) of control, 21 dpi, and 56 dpi M1s from all groups. Yellow dotted lines indicate inset images in (A-F) that represent the ROIs used for quantifications. There was a significantly lower dentin volume in Tgfbr2cko mice at 21 dpi compared to WT mice (B, E G), but at 56 dpi, the tertiary dentin volume was comparable between genotypes (C, F, G). Scale bars = 1 mm. In (G), * indicates p<0.05 and *** indicates p<0.001 by mixed effects 2-way ANOVA.
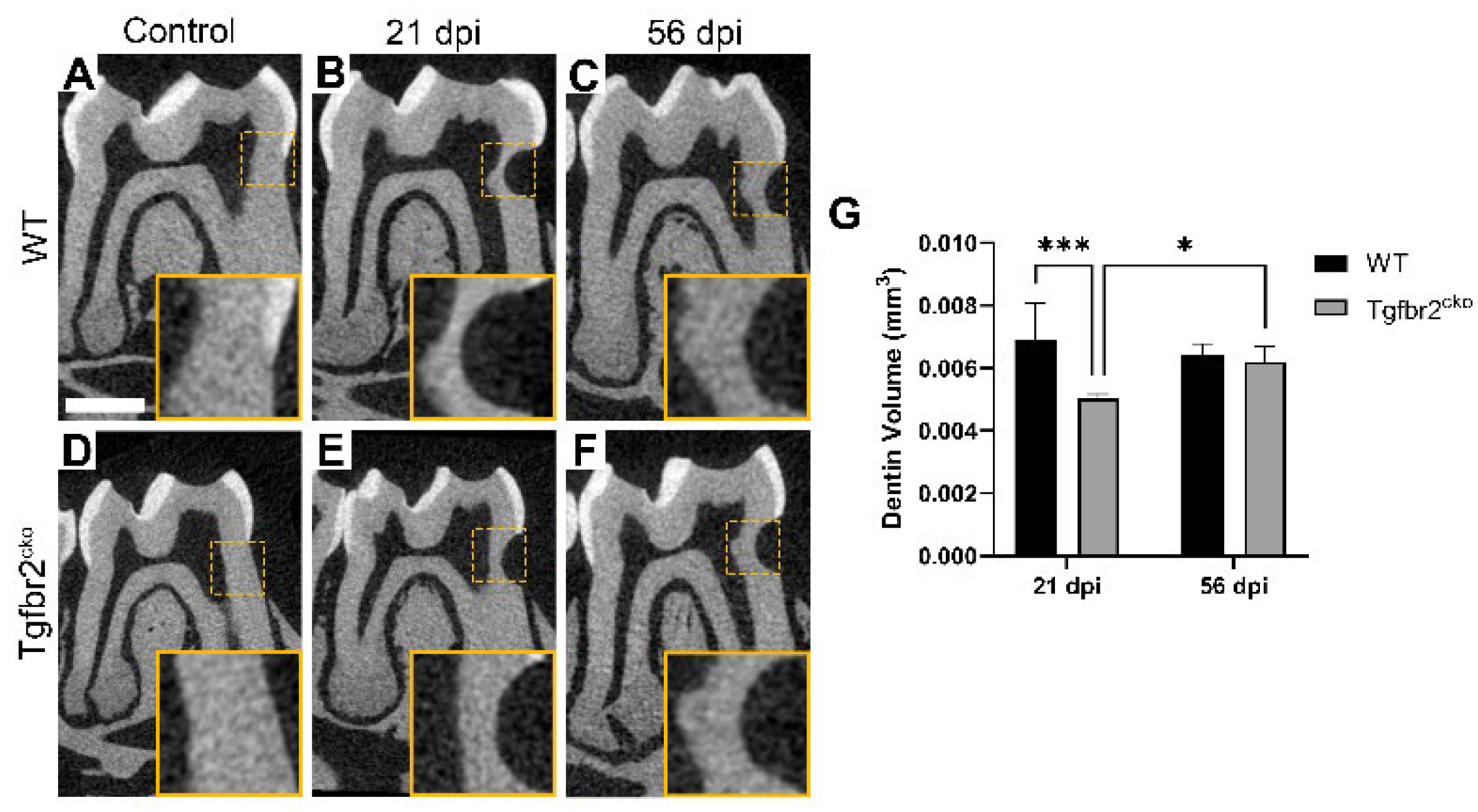
Figure 5.
CGRP+ axon sprouting in response to dentin injury. (A-J) Representative maximum projections of confocal images (20 μm thick) showing the CGRP+ (red) axon outgrowth in control mice and WT and Tgfbr2cko M1s collected at 4-56 dpi (N = 4-8/group). The injury is visible as a half-moon on the right of each frame (B-E, G-J) via differential interference contrast (DIC) imaging. In some images, gingiva is apparent in the injured area. Tgfbr2cko CGRP+ axon sprouting increased significantly between 4 and 21 dpi compared to that in the WT mice (B, D, G, I). At 56 dpi, there were no significant differences in sprouting between the genotypes (E, J). Scalebar in (A) = 50 μm. (K) Prediction of CGRP pixels in response to injury for each genotype fit from a generalized estimating equation model.
Figure 5.
CGRP+ axon sprouting in response to dentin injury. (A-J) Representative maximum projections of confocal images (20 μm thick) showing the CGRP+ (red) axon outgrowth in control mice and WT and Tgfbr2cko M1s collected at 4-56 dpi (N = 4-8/group). The injury is visible as a half-moon on the right of each frame (B-E, G-J) via differential interference contrast (DIC) imaging. In some images, gingiva is apparent in the injured area. Tgfbr2cko CGRP+ axon sprouting increased significantly between 4 and 21 dpi compared to that in the WT mice (B, D, G, I). At 56 dpi, there were no significant differences in sprouting between the genotypes (E, J). Scalebar in (A) = 50 μm. (K) Prediction of CGRP pixels in response to injury for each genotype fit from a generalized estimating equation model.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



