
Submitted:
21 May 2024
Posted:
23 May 2024
You are already at the latest version
Abstract
(Background) Canine mammary tumors (CMT) have emerged as an important model for understanding pathophysiological aspects of human disease. Liquid biopsy (LB), which relies on blood-borne biomarkers and offers minimal invasiveness, holds promise for reflecting the disease status of patients. Small extracellular vesicles (SEVs) and their protein cargo have recently gained attention as potential tools for disease screening and monitoring. (Objectives) This study aimed to isolate SEVs from canine patients and analyze their proteomic profile to assess their diagnostic and prognostic potential. (Methods) Plasma samples were collected from female dogs grouped into CMT (malignant and benign), healthy controls, relapse, and remission groups. SEVs were isolated and characterized using ultracentrifugation (UC), nanoparticle tracking analysis (NTA) and transmission electron microscopy (TEM). Proteomic analysis of circulating SEVs was conducted using Liquid Chromatography–Mass Spectrometry (LC–MS). (Results) While no significant differences were observed in the concentration and size of exosomes among the studied groups, proteomic profiling revealed important variations. Mass spectrometry identified exclusive proteins that could serve as potential biomarkers for mammary cancer. These included Inter-alpha-trypsin inhibitor heavy chain (ITIH2 and ITI4), phosphopyruvate hydratase or alpha enolase (ENO1), eukaryotic translation elongation factor 2 (eEF2), actin (ACTB), transthyretin (TTR), beta-2-glycoprotein 1 (APOH) and gelsolin (GSN) found in female dogs with malignant tumors. Additionally, Vitamin D-binding protein (VDBP), also known as group-specific component (GC), was identified as a protein present during remission. (Conclusion) The results underscore the potential of proteins found in SEVs as valuable biomarkers in CMT. Despite the lack of differences in vesicle concentration and size between the groups, the analysis of protein content revealed promising markers with potential applications in CMT diagnosis and monitoring. These findings suggest a novel approach in the development of more precise and effective diagnostic tools for this challenging clinical condition.
Keywords:
canine
; biomarker
; proteomics
; small extracellular vesicles
; liquid biopsy
; mammary carcinoma
; diagnosis
1. Introduction
Canine mammary tumors (CMT) have garnered significant attention in veterinary medicine, not only due to their high prevalence among female dogs but also because of their intriguing similarities with human breast cancer. As the most common tumor in intact female dogs, CMT provides a unique opportunity to bridge the gap between veterinary and human medicine, offering insights into shared mechanisms and potential breakthroughs. In addition to anatomical and histological similarities, CMT and human breast cancer share genetic and molecular features, making the former an invaluable comparative model for understanding the complexities of both diseases. Studying the canine counterpart can provide essential understanding into disease progression, treatment strategies, and the development of potential biomarkers that are applicable to both species [1,2].
In both veterinary and human medicine, the identification of reliable biomarkers plays a crucial role in early detection, accurate diagnosis, and effective prognosis of disease. Biomarkers serve as indicative tools for the presence or progression of diseases [3,4], making their exploration in CMT not only beneficial for our beloved canine companions but also significant for advancing our understanding of human breast cancer.
Tissue biopsy limitations are increasingly recognized in precision medicine. In contrast, liquid biopsy (LB) offers minimal invasiveness, easy sample acquisition, and dynamic analysis, enabling the early detection of malignant tumors [5]. LB relies on unique biomarkers in circulation, which reflect the patient’s disease status [6].
One promising avenue in biomarker discovery involves investigating the cargo of small extracellular vesicles (SEVs). These small membrane-bound structures released by cells carry a payload of proteins, nucleic acids, and other bioactive molecules [6,7]. Exploring the extracellular vesicle cargo in CMT may unveil specific proteins usable as biomarkers, revolutionizing diagnosis and prognosis [8,9,10], as well as distinct molecular pathways deregulated in the disease that could be further investigated as a therapeutic strategy. By embracing the convergence of veterinary and human medicine, we propose a scientific endeavor that not only benefits our companion animals but also advances cancer research with far-reaching implications for both species. In this study, we performed an exploratory analysis and evaluated, using mass spectrometry, the proteomic profile of SEVs in dogs bearing mammary tumors, either benign or malignant, before and after mastectomy, under relapse or remission.
2. Materials and Methods
Experimental Groups
Forty adult female dogs were distributed into the following groups – (G1) 8 female dogs with malignant mammary neoplasia (Simple Carcinomas); (G2) 8 female dogs with benign mammary neoplasia (Carcinoma in mixed tumor); (G3) 8 female dogs with tumor relapse after mastectomy (metastasis); and (G4) 8 female dogs in clinical remission (over a year following mastectomy of malignant tumors); and (G5) 8 healthy female controls. For G1 and G2 groups, blood was collected at the time of diagnosis of mammary tumor (G1.1 and G2.1) and at the time of surgical suture removal (G1.2 and G2.2), approximately 10 days after mastectomy. G3, G4, and G5 groups had only one collection. A summary of our methodological approach is shown in Figure 1. The average age of the female dogs in all groups was 8.5 years, with mixed breed dogs being the most prevalent among all the breeds.
The inclusion criteria for the neoplastic group were adult dogs with a clinical and histological diagnosis of CMT, with or without metastasis. The control group consisted of healthy adult spayed patients without comorbidities or treatments. Histopathological grading was performed using the Goldschmidt classification (2011) [11], which is based on Nottingham’s classification.
Liquid Biopsy
Three to five milliliters of blood were collected into tubes with EDTA anticoagulant and immediately centrifuged twice at 2,500g at 23˚C for 15 min to obtain platelet-poor plasma. Subsequently, the plasma underwent to three consecutive centrifugations at 4˚C, first at 300g for 10 min to remove cells, then at 2,500g for 10 min to remove cellular debris, and finally at 16,500g for 30 min to remove small extracellular vesicles (SEVs) larger than 150 nm. After five centrifugation cycles, the plasma was aliquoted and stored at -80˚C for subsequent isolation and characterization of the SEVs.
Tissue samples (tumor fragment) and histopathology
The tumor fragment obtained during surgical excision was fixed in 10% formaldehyde for 24 h and then transferred to 70% alcohol for subsequent histopathological diagnosis. Histopathological analysis of the canine patients was conducted at the PatAnimal Laboratory in São José do Rio Preto, SP. The samples were processed following the standard protocol, which included sectioning in a microtome and staining with hematoxylin and eosin (HE). The slides were examined by two independent veterinary pathologists using a common Nikon E20 optical microscope.
Exosomes Isolation
Exosome isolation was performed using ultracentrifugation (UC) (Optima XE-90 Ultracentrifuge; rotor 70 Ti; Beckman Coulter) at the Laboratory of Molecular Morphophysiology and Development (LMMD/ZMV), University of São Paulo, Pirassununga/SP/Brazil. To obtain exosome-enriched content, the pellets were filtered through a 0.22 μm pore filter and ultracentrifuged twice at 119,700 g for 70 minutes at 4 ºC. Following the second ultracentrifugation, the pellet was diluted in phosphate-saline (PBS) and stored at -80 ºC.
Nanoparticle Tracking Analysis (NTA)
The pellets containing SEVs were characterized for size, morphology, and quantity. Initially, they were diluted in 50 µL of magnesium-calcium-free PBS at a dilution factor of 1:500 in PBS. A laser beam was passed through a sampling chamber containing this liquid suspension. The suspended particles scattered the light, allowing visualization under a microscope at 20x magnification. A video camera (sCMOS in Camera Level 15 at a temperature of 37 ºC) captured 5 images of 30 seconds of these particles in Brownian motion. Using the Nano-Sight software (NS300; NTA 3.1 Build 3.1.45; Malvern), the particles were individually tracked, and the hydrodynamic diameter of the particles was calculated using the Stokes-Einstein equation.
Transmission Electron Microscopy (TEM)
Following ultracentrifugation, the pellet was diluted in a fixative solution (0.1M sodium cacodylate, 2.5% Glutaraldehyde, 4% paraformaldehyde, pH between 7.2 and 7.4) for 2 hours at room temperature (RT). Then, 2 mL of ultrapure water were added followed by centrifugation at 119,700 g for 70 minutes at 4 ºC to remove the fixative solution. The pellet was diluted in 20 uL of ultrapure water and stored at 4ºC. The contents were placed on a copper grid coated with pioloform for 5 min, and the excess was removed with moist filter paper. The grid was then inserted into a drop of 2% aqueous uranyl acetate for 3 min, removing the excess again with moist filter paper. Transmission electron microscopy (FEI 200kV, model Tecnai 20, emitter LAB6) was performed at the Multiuser Electron Microscopy Laboratory, USP/Ribeirão Preto/Brazil. Images were captured at 140,000 X magnification.
Proteomics analysis
Mass spectrometry analysis was conducted using the nanoElute nanoflow chromatographic system from Bruker Daltonics, Bremen, Germany, coupled online to a hybrid trapped ion mobility spectrometry-quadrupole time-of-flight mass spectrometer-timsTof Pro mass spectrometer, also from Bruker Daltonics at the Max Feffer Genetics of Plants Laboratory, Departament of Genetics, USP/Piracicaba/SP/Brazil. An aliquot (1µL) of the sample, equivalent to 200 ng of digested peptides, was injected into an Aurora 2 C18 trap column (1.6µm, 250mm x 75µm), from ionOpticks, Australia. A typical reverse-phase gradient (Solvent A: 0.1% AF, 99.9% H2O MilliQ; Solvent B: 0.1% AF, 99.9% CH3CN) was established in a liquid chromatography nanoflow system and separated at a flow rate of 400 nL.min. 1. The column temperature was maintained at 50°C. The chromatographic run was 120 min (2% to 15% of Solvent B for 60 min; raised to 25% at 90 min; raised to 37% at 100 min; raised to 95% at 110 min and finally 95% for 10 min for column washing). The column was coupled online with a timsTOF-Pro with a CaptiveSpray ion source, both from Bruker Daltonik GmbH. The temperature of the capillary ion transfer line was set at 180°C. Ion accumulation for 123ms and mobility separation were obtained with an input potential ramp from -160V to -20V within 123s. During acquisition, to enable the parallel accumulation-serial fragmentation (PASEF) method, precursor m/z and mobility information were first derived from a Tims-MS full scan experiment, with an m/z range of 100-1700. Monocharged precursors were excluded based on their position in the m/z-ion plane of mobility, and precursors that reached the target value of 20,000 a.u. were dynamically excluded for 0.4 min. The operational mode of TIMS-ToF, MS, and PASEF were controlled and synchronized using OtofControl 5.1 instrumental control software from Bruker Daltonik.
For data processing, the raw files were analyzed using MaxQuant software, version 2.4.0.0. Perseus analysis software, version 2.0.9.0 (Tyanova and Cox, 2018), was used to rigorously filter the proteomic results (90%), followed by a refinement using a script in the R programming language (https://www.R-project.org/). Processed peak intensities were then further submitted to Metaboanalyst 5.0 [12] to generate heatmaps after Pareto scaling, presenting only the 25 most significant proteins. Confirmed proteins retrieved from the Uniprot database for Canis lupus familiaris (CANF) were displayed using their gene names, while predicted, inferred, uncertain or Ig-like proteins were indicated by their primary accession. Exclusive and shared proteins were visualized using Venn diagrams (https://bioinformatics.psb.ugent.be/webtools/Venn/). Gene Ontology Enrichment Analysis [13] was performed for the unique features found in each group, with outputs having a false discovery rate (FDR) < 0.05 considered enriched, and -log (p value) reported. To assess protein interaction or network formation, the protein list was submitted to the STRING database [14]. Finally, unique proteins were cross-referenced in the Uniprot database, and squares were inserted in the figures to show only the confirmed, predicted, or inferred proteins while uncertain or Ig-like proteins were not shown.
Statistical Analysis
Statistical calculations were performed for SEVs using and GraphPad Prism software (v.10.2.3). All quantitative data were expressed either as the median with interquartile ranges or as means with standard errors. Quantitative significance was performed by One-way ANOVA for comparison between groups followed by Tukey’s post-hoc test. Comparison between two paired groups was determined by Student's t test. p <0.05 was considered statistically significant.
3. Results
3.1. Extracellular Vesicles Isolated from Canine Plasma Exhibited Characteristics Typical of Small Extracellular Vesicles (SEVs), with Consistent Concentration and Size across Different Groups
The average concentration and size of SEVs for each canine group are presented in Figure 2. While there were not statistically differences in SEVs concentration and size among most groups, the recurrence and remission groups showed distinct patterns. This suggests parameters may not offer significant diagnostic or prognostic value independently. It highlights the importance of considering a more comprehensive approach or exploring other SEV characteristics, such as cargo content, for a more thorough assessment. SEVs were isolated from serum using ultracentrifugation following a standard procedure described in previous studies and characterized according to the guidelines of the International Society for Extracellular Vesicles (ISEV) [15] . Validation was conducted using NTA (Figure 2A–H) and TEM (Figure 2I), revealing a typical “donut-like” appearance by TEM. Our data collectively indicate that the SEVs isolated from canine plasma exhibit characteristics typical of exosomes.
3.2. SEVs` Proteomic Profile from Dogs with Malignant Tumors Exhibit a Larger Number of Unique Proteins, Most of Them Associated with Immune and Wound Healing Processes
We initially investigated whether the proteomic profile of SEVs differed among healthy (G5), benign (G2.1), and malignant (G1.1) condition. The heatmap (Figure 3A) revealed distinct protein clusters specific to each group, with SEVs from dogs bearing malignant tumors containing a higher number of exclusive proteins or proteins with increased intensity. Indeed, this group showed the highest number of total (85) and unique (31) proteins (Figure 3B), while the benign group had 49 total and 1 unique protein, and the healthy group showed 56 total and 2 unique proteins. The squares in Figure 3B represent only the exclusive proteins. These findings suggest that the cargo composition between the pathological status is sharply distinct, potentially serving as biomarkers for disease staging. Gene Ontology enrichment analysis was performed using all output from data processing, revealing 14 significantly enriched biological processes in the malignant group, most of which were related to immune- or wound healing (Figure 3C). The unique proteins identified appeared to cluster and form a network with interactions, as depicted in Figure 3D.
3.3. SEV`s Cargo from Dogs with Malignant Tumor Decreases after Mastectomy, and This Trend Persists Even after Surgery in Cases of Remission or Relapse
Having identified a specific cargo in SEVs, including unique proteins, we interrogated whether mastectomy would alter the proteomic profile in benign or malignant conditions. For this purpose, we performed a paired analysis before and after surgery, as summarized in Figure 4. The heat map showed distinct clusters for benign (Figure 4A) and malignant (Figure 4B) conditions, while unique proteins identified before and after surgery were detailed in Figure 4C,D. In the benign group, 9 proteins were exclusive in SEVs cargo before surgery and 10 after. Conversely, in the malignant group, 39 proteins were found in SEVs collected before surgery compared to 3 after, indicating that mastectomy remarkably decreased the number of proteins in SEVs, particularly in the malignant group. Gene Ontology analysis retrieved significantly enriched biological processes only in the malignant group, including pathways involved in cholesterol and lipid transport, in addition to immune-related processes (Figure 4E).
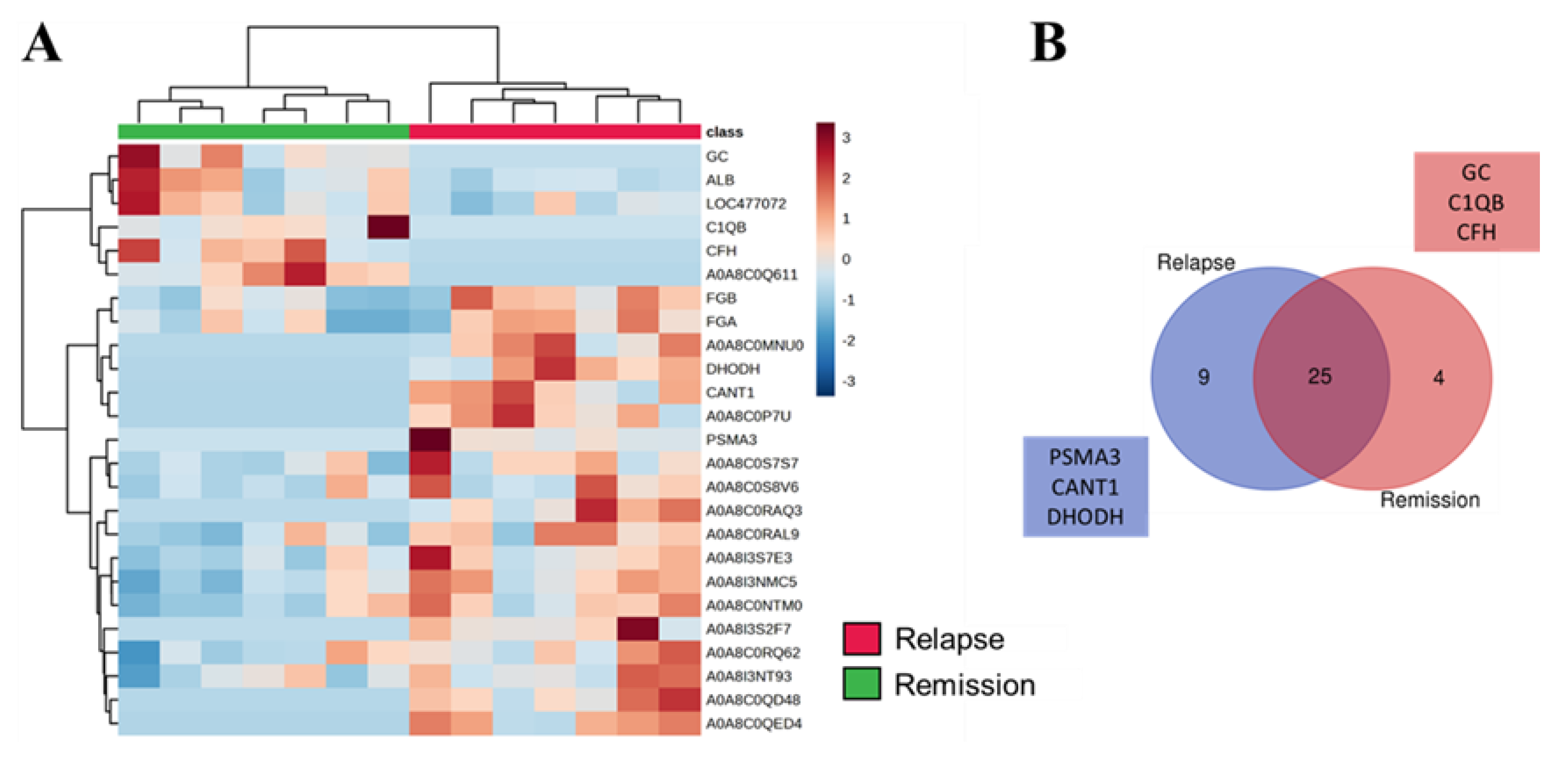
Next, we examined whether proteins found in malignant condition would change upon remission or relapse (Figure 5). We observed that SEVs from dogs with malignant tumors had increased peak intensity for most proteins, while these proteins were decreased or absent in the relapse or remission groups, as shown in the heatmap (Figure 5A). This was further confirmed by the increase in the number of exclusive proteins (51) found in the malignant group (Figure 5B) compared to the relapse (3) and remission (1) groups. Once again, most of the enriched biological processes were related to lipid and cholesterol metabolism, innate and adaptative immune pathways, wound healing, coagulation, and response to stress (Figure 5C). The majority of identified proteins formed a network of interactions (Figure 5D). The difference between the relapse and remission groups is described in Figure 6. Both groups showed clustered proteins, but most of them were not validated or were primarily Ig-like proteins (Figure 6A), indicating a possible regulation of the immune system. Both groups exhibited exclusive proteins (9 for relapse and 4 for remission), but no significant enrichment output was obtained from the gene ontology analysis.
4. Discussion
Our findings demonstrate a diverse array of proteins present in each investigated condition. As depicted in Figure 3, the heatmap reveals distinct patterns of protein expression between healthy dogs and those with benign and malignant tumors, suggesting significant alterations in the proteomic content of SEVs in different pathological states. Additionally, the Venn diagram highlights exclusive proteins found in each group, providing information on potential molecular markers associated with each condition. Gene ontology enrichment analysis reveals specific biological pathways and molecular functions that may be affected in malignant tumors, indicating potential therapeutic targets. Similarly, Figure 4, Figure 5 and Figure 6 provide valuable information into changes in the proteomic profile of SEVs at different stages of the disease and treatment response. These results suggest that SEVs may play a significant role in the progression of canine mammary cancer and could serve as useful biomarkers for monitoring treatment efficacy and predicting patient prognosis.
EVs have emerged as pivotal mediators of intercellular communication and are increasingly recognized as potential source of biomarkers for liquid biopsy in various cancer types [16]. They exhibit selectivity in tumor cell signaling and are continuously secreted into body fluids from the early stages of the disease [16,17]. Aguilera-Rojas and colleagues (2018) [17] isolated SEVs from blood serum of dogs and reported a nanoparticle concentration (xE10/mL) ranging from 107.4 +/− 6.8 to 403.2 +/− 25.8 in non-cancer patients, and 225.6 +/− 10.4 to 500.4 +/− 76.4 in cancer patients, respectively. However, no significant difference was observed between non-cancerous and cancerous samples, despite notable variation within individual samples [18]. Similarly, our results demonstrated that the SEVs concentration did not exhibit statistically significant difference in the canine cohorts, either before or after surgical excision of the primary tumor. In fact, the concentration before mastectomy was higher than after mastectomy, considering individual values. However, SEVs concentration in the remission, relapse, and control groups was even higher than that obtained for the mammary tumor groups. Like Aguilera-Rojas and colleagues (2018) [17], we also noted considerable variation within individual samples, despite the lack of statistical significance. The constant influx of SEVs, the dynamic release-uptake dynamics of different cells, and the lack of precise characterization of SEVs' origin make it challenging to determine whether the quantity of SEVs in cancer patients differs significantly from that in healthy control patients.
On the contrary, scientific evidence suggests that the cargo of SEVs, comprising proteins, nucleic acids, and lipids, may be more relevant than merely their quantity or size in discerning between cancer and non-cancer patients. Their cargo could impact the signaling of the host's immune defenses, facilitating the spread of the neoplasm by priming distant environments for metastasis [19,20]. This aspect is particularly crucial for identifying biomarkers and gaining a better understanding of cancer pathophysiology. Consequently, the number of studies focusing on the proteomics of SEVs in patients diagnosed with mammary tumor has increased, with the potential for early diagnosis and longitudinal prognostic evaluation [9].
In this study we observed that the protein characteristics of SEV’s cargo can be more discriminatory than their mere presence or absence. The relative abundance of proteins among the various groups did not exhibit significant differences, rendering it an insignificant factor within the study's context. However, the pertinent observation lies in the consistent detection of distinct proteins across all groups, despite certain groups sharing common proteins, presumably linked to normal cellular constituents of the species. This discernment has enabled the identification of potential biomarkers for diagnostic and prognostic purposes, and conceivably for guiding treatment modalities.
The proteins identified in blood samples from female dogs, particularly in the malignant mammary tumor group, encompass various roles in cancer biology. One of these very abundant proteins, inter-alpha-trypsin inhibitor heavy chain (ITIH2) contributes to matrix stability and may serve as a hyaluronan transporter or binding protein [21,22]. These proteins can be positively or negatively regulated, but there is already evidence that all members of the ITIH family play a significant role in cell malignant processes and tumor growth [22]. Kopylov and colleagues (2020) [22] studied the proteomics of plasma samples from colorectal cancer patients at different stages and healthy patients. They observed significant changes in the levels of inter-alpha-trypsin inhibitor heavy chains (such as ITIH2) due to their implication in tumor growth and the malignancy process [22]. The literature describes that ITIH2 may present significant downregulation in solid tumors [23], contrary to the results obtained in the study by Kopylov and colleagues (2020) [22] and in our study, which observed a greater abundance of the ITIH2 protein in the malignant carcinoma group. According to Hamm (2008) [20] ITIH2 expression was clearly detectable in normal breast epithelium, in hyperplastic gland epithelium, and in ductal carcinoma in-situ. However, in 44% of invasive breast carcinomas, ITIH2 expression was strongly reduced or completely lost, whereas 56% of invasive carcinomas maintained moderate to strong ITIH2 expression. Furthermore, strong expression of ITIH2 was highly significantly associated with the presence of estrogen receptor. Our results may reflect differences between canine and human carcinomas, and further studies are needed to prove or refute these findings. Also, van den Broek and colleagues (2010) [23] observed significantly increased serum concentrations in the breast cancer group for ITIH4, significantly decreasing after surgery. The authors suggested a potential for ITIH4 in post-surgical breast cancer monitoring [23]. Accordingly, van Winden and colleagues (2010) [23] and Opstal-van Winden and colleagues (2011) [24] discovered that isoform 1 of the heavy chain of inter-alpha trypsin inhibitor H4 (ITIH4) presented elevated values in pre-diagnosis breast cancer and were already altered up to three years before cancer detection. Corroborating the previous studies, the study by Yang and colleagues (2016) [25] aimed to identify new serum peptide biomarkers for women with breast cancer (BC) through the analysis of the serum proteomic profile. Three peptide biomarkers were identified, including ITIH4, and the authors concluded that this protein has diagnostic and prognostic potential for breast cancer.
Another high abundant protein found mostly in the malign carcinoma group was phosphopyruvate hydratase or alpha enolase (ENO1), that plays a crucial role in cancer development by promoting cell proliferation, invasion and metastasis. Its overexpression has been linked to chemoresistance and poor prognosis [26]. Song and colleagues (2015) [27] analyzed the involvement of ENO1 in tumor progression and prognosis of human glioma. They concluded that overexpression of ENO1 was associated with glioma progression, while reducing ENO1 expression led to suppression of cell growth, migration and invasion progression by inactivating the PI3K/Akt pathway in glioma cells. In cancer metabolic reprogramming, ENO1 stimulates cancer cells to create energy largely through the breakdown of glucose in a non-oxidative way, rather than typical oxidative phosphorylation, which is known as the Warburg effect [28,29]. Indeed, mammary canine tumors undergo metabolic alterations [30] that favors tumor growth. Chu and colleagues (2011) [31] examined ENO1 expression by immunohistochemical staining and evaluated its importance in canine mammary carcinoma. ENO1 overexpression significantly correlated with shorter survival but was not associated with ER positivity in canine mammary carcinoma [31]. The study of Zamani-Ahmadmahmudi and colleagues (2014) [32] used serological proteomic analysis to detect autoantigens that provoke a humoral response in dogs with mammary tumors. They found four autoantigens, including ENO1, with significantly greater immunoreactivity in tumor samples than in normal samples, which were identified as biomarker candidates [32].
Besides ITIH2, ITIH4 and ENO1, we also found eukaryotic translation elongation factor 2 (eEF2) as abundant protein. It’s a member of the GTP-binding translation elongation factor family that is essential for protein synthesis. In our study, this protein was abundantly found in the SEVs proteomic of the malignant carcinoma group. Eukaryotic translation elongation factor 2 (eEF2) and its kinase eEF2K regulates many cellular processes, such as protein synthesis, cellular differentiation and malignant transformation [33]. Scientific evidence shows that eEF2K regulates the cell cycle, autophagy, apoptosis, angiogenesis, invasion and metastasis in various types of cancer. Expression of eEF2K promotes cancer cell survival, and the level of this protein is increased in many cancer cells to adapt them to microenvironmental conditions, including hypoxia, nutrient depletion, and acidosis [34]. The study by Wang and colleagues (2019) [35] investigated the role of autophagy and its regulator, eukaryotic elongation factor 2 kinase (eEF2K), in determining the biological nature of triple-negative breast cancers (TNBCs). eEF2K and autophagy play key roles in maintaining aggressive tumor behavior and chemoresistance in TNBC, and eEF2K silencing may be a new strategy for the treatment of TNBC.
Actin B (ACTB) was also found as abundant and exclusive protein in this study. Beta-actin (ACTB) is a cytoskeletal structural protein widely distributed in all eukaryotic cells and plays critical roles in cell migration and cell division. ACTB is generally upregulated in most tumor cells and tissues, being associated with the invasiveness and metastasis of cancers [36]. Fang et al. (2019) [37] evaluated the role of serum actin-4 as a biomarker for the diagnosis of BC, as well as the association between its levels and clinicopathological characteristics. They reported that serum ACTN4 level was upregulated in BC patients compared to healthy controls [37]. Furthermore, high ACTN4 expression was significantly associated with clinical stage, tumor grade, and lymph node status [37,38]. Numerous clinical studies showed that changes in ACTN4 gene expression are correlated with aggressiveness, invasion, and metastasis in certain tumors [39]. Wang and colleagues (2017) [40] sought to determine the role and regulation of ACTN4 expression in human breast cancer metastasis under ellagic acid (EA)-based therapy. EA inhibited breast cancer growth and metastasis by directly targeting ACTN4 in vitro and in vivo. ACTN4 knockdown resulted in blocking malignant cell proliferation, colony formation and improving metastasis potency [40]. Increased ACTN4 expression was directly associated with advanced cancer stage, increased incidence of metastases and short survival period global [40].
Chung and colleagues (2014) [41] used mass spectrometry to identify proteins differentially expressed in sera from women with breast cancer and healthy volunteers. From 51 protein peaks that were significantly up- or down-regulated, they obtained five protein peaks that showed positive association with large tumor size and lymph node involvement and could distinguish women with breast cancer with high sensitivity and specificity. Interestingly, transthyretin and beta2-glycoprotein were between those proteins. In agreement, our study also identified an abundance and exclusive peak for both proteins. However, previously, the study by Nasim et al. (2012) [42] identified several acute phase proteins as potential biomarkers for breast carcinoma but reported that transthyretin was significantly downregulated in the sera of BC patients [42]. Recently, Sharma et al. (2023) [43] studied the action of organochlorine pesticides (OCP) and their relationship with the spread of breast cancer in Indian women, through the proteomic analysis of plasma from breast cancer patients. They found 17 dysregulated proteins, but transthyretin (TTR) was three times higher than in healthy controls, and docking and molecular dynamics studies revealed a competitive affinity between the pesticide endosulfan II and the thyroxine-binding site of TTR that could result in endocrine dysregulation and lead to breast cancer. The authors pointed to a putative role of TTR in OCP-mediated CM [43].
Apolipoprotein H (APOH) or beta2-glycoprotein I (β2-GPI) is a plasma glycoprotein that has been implicated in a variety of physiological functions. Apolipoproteins (APOs) bind to lipids to form lipoproteins. By functioning as lipid carriers, apolipoproteins act as ligands for cell membrane receptors, enzyme cofactors and structural components of lipoproteins [44,45]. Several studies demonstrate the interaction of APOs with classical tumorigenesis pathways. Furthermore, dysregulation of APOs may indicate the occurrence and progression of cancer, thus serving as potential biomarkers for cancer patients [46]. Besides Chung and colleagues [41], Lee and colleagues (2023) [47] also investigated the role of β2-GPI in tumor cells from mammary cancer patients and its correlation with tumor prognosis. β2-GPI expression was predominantly observed in the cells from breast cancer patients and significantly correlated with tumor stage and lymph node metastasis of breast cancer. High expression of β2-GPI was significantly correlated with better overall survival (OS) and disease-free survival (DFS). Their results corroborated Lin and colleagues previously results [48].
Cytoskeletal rearrangement occurs in several cellular processes and involves a broad spectrum of proteins such as gelsolin superfamily proteins [49]. Gelsolin (GSN) is a multifunctional actin-binding protein that is greatly decreased in many transformed cell lines in tumor tissues, including breast cancers. The study by Mielnicki et al. (1999) [50] demonstrated epigenetic modification leading to downregulation of gelsolin expression in human breast cancer. Baig et al. (2013) [49] investigated the germline mutations and expressional profile of gelsolin in human breast cancer tissues. Different types of mutations were observed in gelsolin coding regions and the transcript level was significantly lower in breast tumor tissues compared to control samples as well as in metastatic patients compared to disease-free patients at final follow-up. Downregulation of gelsolin was also described by Winston and colleagues (2001) [51] in invasive mammary carcinomas. Stock et al. (2015) [52] analyzed gelsolin mRNA levels and concluded that high gelsolin levels are associated with a better prognosis in ER(+) HER2(-) breast cancer and a reduction in tumor cell migration. However, gelsolin was also found as exclusive protein in this study, instead of downregulated. This finding was intriguing but considering the differences between canine and human species and the scarcity of studies in dogs, we hypothesize that the abundance of gelsolin protein in samples from dogs with malignant mammary tumors may be related to a better prognosis in these individuals, and not related to its function as a diagnostic biomarker. Therefore, it is likely that gelsolin may play a dual role in cancer genesis. For example, Rao and colleagues (2002) [53] reported decreased gelsolin expression in the early stages of malignant transformation in urothelial carcinomas. However, there was an increase in gelsolin expression in the transition from non-invasive to invasive urothelial carcinomas, indicating a biphasic expression pattern. Therefore, the authors concluded that increased gelsolin expression could indicate the conversion of a superficial tumor into an invasive tumor. Corroborating our finding, Van den Abbeele and colleagues (2007) [54] demonstrated that downregulation of gelsolin in several cancer cell types significantly reduces the invasive and motile properties of the cells, as well as cell aggregation, pointing to a role for gelsolin as tumor activator. Also, Chen et al. (2015) [53] investigated the role of gelsolin (GSN) in epithelial-mesenchymal transition (EMT) in breast cancer cells in response to transforming growth factor beta 1 (TGF-β1). The results showed that TGF-β1 induced the expression of GSN and EMT and increased cell migration and invasion [55]. GSN overexpression affected cell proliferation, cell cycle and migration, and modulated vimentin expression [55]. The authors concluded that TGF-β1 promoted the demethylation of the GSN gene promoter in these cells, inducing epigenetic modifications that contributed to the progression of EMT in breast cancer cells [55]. In the study by Zhang et al. (2020) [56] the role of gelsolin (GSN) in hepatocellular carcinoma (HCC) was investigated. GSN was found to be overexpressed in HCC tissues and correlated with advanced tumor grade and poor prognosis, while GSN knockdown inhibited tumor cell migration and invasion and GSN overexpression had the opposite effect. The authors suggested that GSN promotes HCC progression by regulating epithelial-mesenchymal transition (EMT).
Interestingly, Vitamin D-binding protein (VDBP) or group-specific component (GC) was identified as an abundant protein in the remission group. We didn’t find literature about the relationship between cancer and VDBP, but Tagliabue and colleagues (2015) [57] performed a meta-analysis of vitamin D binding protein and cancer risk. They included 28 independent studies relating to the following tumors: basal cell carcinoma, bladder, breast, colon-rectum, endometrium, liver, esophagus, stomach, melanoma, pancreas, prostate and kidney. The authors concluded they could not assert that DBP would be a marker of cancer remission, but the results suggest a trend towards a decreased risk of cancer in individuals with higher levels of DBP, which could indicate a potential protective role of DBP in cancer.
5. Conclusions
Based on our main findings, the concentration and size of SEVs did not consistently vary in the canine mammary tumor (CMT) groups, before and after surgical treatment, as well as in the relapse, remission, and control groups. Therefore, SEVs did not prove to be reliable biomarkers and could not be strictly correlated with CMC diagnosis. However, in this descriptive study, the identification of candidate proteins for biomarkers underscores their pivotal role in defining diagnostic and prognostic strategies for CMT. This achievement highlights the potential to enhance patient care and advocate for the well-being of our canine companions. The cargo of SEVs shows promise in predicting disease progression, motivating us to pursue further investigations for a comprehensive understanding. Importantly, there is limited literature on these proteins in dogs, and this study is the first to introduce them to the scientific community. This underscores the need for further investigations to comprehensively understand their potential in predicting disease progression and improving patient care for CMT.
Author Contributions
AAN and DAPCZ have made substantial contributions to conception and design. AAN and LMMM have been involved in acquisition of biological material. AAN, GS, SFS, CAS, JCS have been involved in isolation and characterization of extracellular vesicles. AAN, GHT, FFS and CAL have been involved in the proteomic analysis. AAN, GHT, and DAPCZ have been involved in interpretation of data. AAN, GHT, LGAC, SB and DAPCZ have been involved in drafting the manuscript or revising it critically for important intellectual content. DAPCZ have given final approval of the version to be published. Each author has participated sufficiently in the work to take public responsibility for appropriate portions of the content and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work were appropriately investigated and resolved. .
Funding
This research was funded by FAPESP, grant number 20/12970-8.
Institutional Review Board Statement
The animal study protocol was approved by the Ethics Committee on the Use of Animals (CEUA), being registered under the protocol number 001-004391/2019. .
Data Availability Statement
Data are available upon request to interested researchers.
Acknowledgments
We would like to thank FAPESP for their support of this project (Proc. No. 20/12970-8), the veterinary staff of UFMT/Sinop Veterinary Hospital, Núcleo Veterinário, Clinivet and “Governador Laudo Natel” Veterinary Hospital, for their assistance in collecting canine samples for this research. We also thank Max Feffer Genetics of Plants Laboratory, Departament of Genetics - Escola Superior de Agricultura Luiz de Queiroz, University of São Paulo, Piracicaba, Brazil, for the support in proteomic research, under the guidance of Prof. Dr. Carlos Alberto Labate and his technician, Prof. Dr. Thaís Regiani Cataldi. Their expertise and assistance have greatly contributed to our work. .
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- L. Zeng, W. Li, C.-S. Chen, Breast cancer animal models and applications., Zool. Res. 41 (2020) 477–494. [CrossRef]
- J.Y. Kwon, N. Moskwa, W. Kang, T.M. Fan, C. Lee, Canine as a comparative and translational model for human mammary tumor., J. Breast Cancer. 26 (2023) 1–13. [CrossRef]
- L. Neves Rebello Alves, D. Dummer Meira, L. Poppe Merigueti, M. Correia Casotti, D. do Prado Ventorim, J. Ferreira Figueiredo Almeida, V. Pereira de Sousa, M. Cindra Sant’Ana, R. Gonçalves Coutinho da Cruz, L. Santos Louro, G. Mendonça Santana, T. Erik Santos Louro, R. Evangelista Salazar, D. Ribeiro Campos da Silva, A. Stefani Siqueira Zetum, R. Silva Dos Reis Trabach, F. Imbroisi Valle Errera, F. de Paula, E. de Vargas Wolfgramm Dos Santos, E. Fagundes de Carvalho, I. Drumond Louro, Biomarkers in Breast Cancer: An Old Story with a New End., Genes. 14 (2023). [CrossRef]
- S. Afzal, M. Hassan, S. Ullah, H. Abbas, F. Tawakkal, M.A. Khan, Breast cancer; discovery of novel diagnostic biomarkers, drug resistance, and therapeutic implications., Front. Mol. Biosci. 9 (2022) 783450. [CrossRef]
- T.K.Y. Tay, P.H. Tan, Liquid biopsy in breast cancer: A focused review., Arch. Pathol. Lab. Med. 145 (2021) 678–686. [CrossRef]
- D. Yu, Y. Li, M. Wang, J. Gu, W. Xu, H. Cai, X. Fang, X. Zhang, Exosomes as a new frontier of cancer liquid biopsy., Mol. Cancer. 21 (2022) 56. [CrossRef]
- E. Diomaiuto, V. Principe, A. De Luca, F. Laperuta, C. Alterisio, A. Di Loria, Exosomes in Dogs and Cats: An Innovative Approach to Neoplastic and Non-Neoplastic Diseases., Pharmaceuticals (Basel). 14 (2021). [CrossRef]
- R.J. Simpson, S.S. Jensen, J.W.E. Lim, Proteomic profiling of exosomes: current perspectives., Proteomics. 8 (2008) 4083–4099. [CrossRef]
- A.A. Novais, G.H. Tamarindo, L.G. de A. Chuffa, D.A.P. de C. Zuccari, Decoding hidden messengers: proteomic profiling of exosomes in mammary cancer research., Biomedicines. 11 (2023). [CrossRef]
- S. Rontogianni, E. Synadaki, B. Li, M.C. Liefaard, E.H. Lips, J. Wesseling, W. Wu, M. Altelaar, Proteomic profiling of extracellular vesicles allows for human breast cancer subtyping., Commun. Biol. 2 (2019) 325. [CrossRef]
- M. Goldschmidt, L. Peña, R. Rasotto, V. Zappulli, Classification and grading of canine mammary tumors., Vet. Pathol. 48 (2011) 117–131. [CrossRef]
- J. Xia, N. Psychogios, N. Young, D.S. Wishart, MetaboAnalyst: a web server for metabolomic data analysis and interpretation., Nucleic Acids Res. 37 (2009) W652-60. [CrossRef]
- P.D. Thomas, D. Ebert, A. Muruganujan, T. Mushayahama, L.-P. Albou, H. Mi, PANTHER: Making genome-scale phylogenetics accessible to all., Protein Sci. 31 (2022) 8–22. [CrossRef]
- D. Szklarczyk, R. Kirsch, M. Koutrouli, K. Nastou, F. Mehryary, R. Hachilif, A.L. Gable, T. Fang, N.T. Doncheva, S. Pyysalo, P. Bork, L.J. Jensen, C. von Mering, The STRING database in 2023: protein-protein association networks and functional enrichment analyses for any sequenced genome of interest., Nucleic Acids Res. 51 (2023) D638–D646. [CrossRef]
- C. Théry, K.W. C. Théry, K.W. Witwer, E. Aikawa, M.J. Alcaraz, J.D. Anderson, R. Andriantsitohaina, A. Antoniou, T. Arab, F. Archer, G.K. Atkin-Smith, D.C. Ayre, J.-M. Bach, D. Bachurski, H. Baharvand, L. Balaj, S. Baldacchino, N.N. Bauer, A.A. Baxter, M. Bebawy, C. Beckham, A. Bedina Zavec, A. Benmoussa, A.C. Berardi, P. Bergese, E. Bielska, C. Blenkiron, S. Bobis-Wozowicz, E. Boilard, W. Boireau, A. Bongiovanni, F.E. Borràs, S. Bosch, C.M. Boulanger, X. Breakefield, A.M. Breglio, M.Á. Brennan, D.R. Brigstock, A. Brisson, M.L. Broekman, J.F. Bromberg, P. Bryl-Górecka, S. Buch, A.H. Buck, D. Burger, S. Busatto, D. Buschmann, B. Bussolati, E.I. Buzás, J.B. Byrd, G. Camussi, D.R. Carter, S. Caruso, L.W. Chamley, Y.-T. Chang, C. Chen, S. Chen, L. Cheng, A.R. Chin, A. Clayton, S.P. Clerici, A. Cocks, E. Cocucci, R.J. Coffey, A. Cordeiro-da-Silva, Y. Couch, F.A. Coumans, B. Coyle, R. Crescitelli, M.F. Criado, C. D’Souza-Schorey, S. Das, A. Datta Chaudhuri, P. de Candia, E.F. De Santana, O. De Wever, H.A. Del Portillo, T. Demaret, S. Deville, A. Devitt, B. Dhondt, D. Di Vizio, L.C. Dieterich, V. Dolo, A.P. Dominguez Rubio, M. Dominici, M.R. Dourado, T.A. Driedonks, F.V. Duarte, H.M. Duncan, R.M. Eichenberger, K. Ekström, S. El Andaloussi, C. Elie-Caille, U. Erdbrügger, J.M. Falcón-Pérez, F. Fatima, J.E. Fish, M. Flores-Bellver, et al., Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines., J. Extracell. Vesicles. 7 (2018) 1535750. [CrossRef]
- M. Szajnik, M. Derbis, M. Lach, P. Patalas, M. Michalak, H. Drzewiecka, D. Szpurek, A. Nowakowski, M. Spaczynski, W. Baranowski, T.L. Whiteside, Exosomes in Plasma of Patients with Ovarian Carcinoma: Potential Biomarkers of Tumor Progression and Response to Therapy., Gynecol Obstet (Sunnyvale). Suppl 4 (2013) 3. [CrossRef]
- M. Aguilera-Rojas, B. Badewien-Rentzsch, J. Plendl, B. Kohn, R. Einspanier, Exploration of serum- and cell culture-derived exosomes from dogs., BMC Vet. Res. 14 (2018) 179. [CrossRef]
- J. Maia, S. Caja, M.C. Strano Moraes, N. Couto, B. Costa-Silva, Exosome-Based Cell-Cell Communication in the Tumor Microenvironment., Front. Cell Dev. Biol. 6 (2018) 18. [CrossRef]
- M. Żmigrodzka, O. Witkowska-Piłaszewicz, A. Rzepecka, A. Cywińska, D. Jagielski, A. Winnicka, Extracellular Vesicles in the Blood of Dogs with Cancer-A Preliminary Study., Animals (Basel). 9 (2019). [CrossRef]
- A. Hamm, J. Veeck, N. Bektas, P.J. Wild, A. Hartmann, U. Heindrichs, G. Kristiansen, T. Werbowetski-Ogilvie, R. Del Maestro, R. Knuechel, E. Dahl, Frequent expression loss of Inter-alpha-trypsin inhibitor heavy chain (ITIH) genes in multiple human solid tumors: a systematic expression analysis., BMC Cancer. 8 (2008) 25. [CrossRef]
- X. Jiang, X.-Y. Bai, B. Li, Y. Li, K. Xia, M. Wang, S. Li, H. Wu, Plasma Inter-Alpha-Trypsin Inhibitor Heavy Chains H3 and H4 Serve as Novel Diagnostic Biomarkers in Human Colorectal Cancer., Dis. Markers. 2019 (2019) 5069614. [CrossRef]
- A.T. Kopylov, A.A. Stepanov, K.A. Malsagova, D. Soni, N.E. Kushlinsky, D.V. Enikeev, N.V. Potoldykova, A.V. Lisitsa, A.L. Kaysheva, Revelation of proteomic indicators for colorectal cancer in initial stages of development., Molecules. 25 (2020). [CrossRef]
- I.van den Broek, R.W. Sparidans, A.W.J. van Winden, M.-C.W. Gast, E.J. van Dulken, J.H.M. Schellens, J.H. Beijnen, The absolute quantification of eight inter-α-trypsin inhibitor heavy chain 4 (ITIH4)-derived peptides in serum from breast cancer patients., Proteomics Clin. Appl. 4 (2010) 931–939. [CrossRef]
- A.W.J. Opstal-van Winden, E.J.M. Krop, M.H. Kåredal, M.-C.W. Gast, C.H. Lindh, M.C. Jeppsson, B.A.G. Jönsson, D.E. Grobbee, P.H.M. Peeters, J.H. Beijnen, C.H. van Gils, R.C.H. Vermeulen, Searching for early breast cancer biomarkers by serum protein profiling of pre-diagnostic serum; a nested case-control study., BMC Cancer. 11 (2011) 381. [CrossRef]
- J. Yang, X. Xiong, S. Liu, J. Zhu, M. Luo, L. Liu, L. Zhao, Y. Qin, T. Song, C. Huang, Identification of novel serum peptides biomarkers for female breast cancer patients in Western China., Proteomics. 16 (2016) 925–934. [CrossRef]
- A.T. Hoang, B. Vizio, L. Chiusa, A. Cimino, D. Solerio, N.H. Do, S. Pileci, M. Camandona, G. Bellone, Impact of tissue enolase 1 protein overexpression in esophageal cancer progression., Int. J. Med. Sci. 18 (2021) 1406–1414. [CrossRef]
- Y. Song, Q. Luo, H. Long, Z. Hu, T. Que, X. Zhang, Z. Li, G. Wang, L. Yi, Z. Liu, W. Fang, S. Qi, Correction: Alpha-enolase as a potential cancer prognostic marker promotes cell growth, migration, and invasion in glioma., Mol. Cancer. 13 (2015) 235. [CrossRef]
- Y. Zhang, Q. Li, Z. Huang, B. Li, E.C. Nice, C. Huang, L. Wei, B. Zou, Targeting glucose metabolism enzymes in cancer treatment: current and emerging strategies., Cancers (Basel). 14 (2022). [CrossRef]
- G. Qiao, A. Wu, X. Chen, Y. Tian, X. Lin, Enolase 1, a moonlighting protein, as a potential target for cancer treatment., Int. J. Biol. Sci. 17 (2021) 3981–3992. [CrossRef]
- G.H. Tamarindo, A.A. Novais, L.G.A. Chuffa, D.A.P.C. Zuccari, Metabolic alterations in canine mammary tumors., Animals (Basel). 13 (2023). [CrossRef]
- P.-Y. Chu, N.C. Hsu, A.T. Liao, N.-Y. Shih, M.-F. Hou, C.-H. Liu, Overexpression of α-enolase correlates with poor survival in canine mammary carcinoma., BMC Vet. Res. 7 (2011) 62. [CrossRef]
- M. Zamani-Ahmadmahmudi, S.M. Nassiri, R. Rahbarghazi, Serological proteome analysis of dogs with breast cancer unveils common serum biomarkers with human counterparts., Electrophoresis. 35 (2014) 901–910. [CrossRef]
- H. Zhu, X. Yang, J. Liu, L. Zhou, C. Zhang, L. Xu, Q. Qin, L. Zhan, J. Lu, H. Cheng, X. Sun, Eukaryotic elongation factor 2 kinase confers tolerance to stress conditions in cancer cells., Cell Stress Chaperones. 20 (2015) 217–220. [CrossRef]
- B. Zhang, J. Zou, Q. Zhang, Z. Wang, N. Wang, S. He, Y. Zhao, C.B. Naman, Progress in the development of eukaryotic elongation factor 2 kinase (eef2k) natural product and synthetic small molecule inhibitors for cancer chemotherapy., Int. J. Mol. Sci. 22 (2021). [CrossRef]
- R.-X. Wang, X.-E. Xu, L. Huang, S. Chen, Z.-M. Shao, eEF2 kinase mediated autophagy as a potential therapeutic target for paclitaxel-resistant triple-negative breast cancer., Ann. Transl. Med. 7 (2019) 783. [CrossRef]
- C. Guo, S. Liu, J. Wang, M.-Z. Sun, F.T. Greenaway, ACTB in cancer., Clin. Chim. Acta. 417 (2013) 39–44. [CrossRef]
- C. Fang, J.-J. Li, T. Deng, B.-H. Li, P.-L. Geng, X.-T. Zeng, Actinin-4 as a Diagnostic Biomarker in Serum of Breast Cancer Patients., Med. Sci. Monit. 25 (2019) 3298–3302. [CrossRef]
- Lenčo, A. Fučíková, J. Dresler, L. Čápková, R. Hrstka, R. Nenutil, P. Bouchal, Targeted proteomics driven verification of biomarker candidates associated with breast cancer aggressiveness., Biochim. Biophys. Acta Proteins Proteom. 1865 (2017) 488–498. [CrossRef]
- D. Tentler, E. Lomert, K. Novitskaya, N.A. Barlev, Role of ACTN4 in tumorigenesis, metastasis, and EMT., Cells. 8 (2019). [CrossRef]
- N. Wang, Q. Wang, H. Tang, F. Zhang, Y. Zheng, S. Wang, J. Zhang, Z. Wang, X. Xie, Direct inhibition of ACTN4 by ellagic acid limits breast cancer metastasis via regulation of β-catenin stabilization in cancer stem cells., J. Exp. Clin. Cancer Res. 36 (2017) 172. [CrossRef]
- L. Chung, K. Moore, L. Phillips, F.M. Boyle, D.J. Marsh, R.C. Baxter, Novel serum protein biomarker panel revealed by mass spectrometry and its prognostic value in breast cancer., Breast Cancer Res. 16 (2014) R63. [CrossRef]
- F.-U.-H. Nasim, S. Ejaz, M. Ashraf, A.R. Asif, M. Oellerich, G. Ahmad, G.A. Malik, Attiq-Ur-Rehman, Potential biomarkers in the sera of breast cancer patients from bahawalpur, pakistan., Biomark. Cancer. 4 (2012) 19–34. [CrossRef]
- S. Sharma, L. Malhotra, P. Mukherjee, N. Kaur, T. Krishanlata, C.V. Srikanth, V. Mishra, B.D. Banerjee, A.S. Ethayathulla, R.S. Sharma, Putative interactions between transthyretin and endosulfan II and its relevance in breast cancer., Int. J. Biol. Macromol. 235 (2023) 123670. [CrossRef]
- L. Ren, J. Yi, W. Li, X. Zheng, J. Liu, J. Wang, G. Du, Apolipoproteins and cancer., Cancer Med. 8 (2019) 7032–7043. [CrossRef]
- J.J.P. Kastelein, W.A. van der Steeg, I. Holme, M. Gaffney, N.B. Cater, P. Barter, P. Deedwania, A.G. Olsson, S.M. Boekholdt, D.A. Demicco, M. Szarek, J.C. LaRosa, T.R. Pedersen, S.M. Grundy, TNT Study Group, IDEAL Study Group, Lipids, apolipoproteins, and their ratios in relation to cardiovascular events with statin treatment., Circulation. 117 (2008) 3002–3009. [CrossRef]
- Y. He, J. Chen, Y. Ma, H. Chen, Apolipoproteins: New players in cancers., Front. Pharmacol. 13 (2022) 1051280. [CrossRef]
- Y.-C. Lee, C.-T. Tang, J.-Y. Kan, C.-P. Chiang, H.-T. Li, J.-N. Sung, W.-C. Chiu, Correlation of Beta2-Glycoprotein I With Tumor Prognosis in Breast Cancer Patients., Anticancer Res. 43 (2023) 3455–3462. [CrossRef]
- X. Lin, S. Hong, J. Huang, Y. Chen, Y. Chen, Z. Wu, Plasma apolipoprotein A1 levels at diagnosis are independent prognostic factors in invasive ductal breast cancer., Discov. Med. 23 (2017) 247–258.
- R.M. Baig, I. Mahjabeen, M. Sabir, N. Masood, K. Ali, F.A. Malik, M.A. Kayani, Mutational spectrum of Gelsolin and its down regulation is associated with breast cancer., Dis. Markers. 34 (2013) 71–80. [CrossRef]
- L.M. Mielnicki, A.M. Ying, K.L. Head, H.L. Asch, B.B. Asch, Epigenetic regulation of gelsolin expression in human breast cancer cells., Exp. Cell Res. 249 (1999) 161–176. [CrossRef]
- J.S. Winston, H.L. Asch, P.J. Zhang, S.B. Edge, A. Hyland, B.B. Asch, Downregulation of gelsolin correlates with the progression to breast carcinoma., Breast Cancer Res. Treat. 65 (2001) 11–21. [CrossRef]
- A.-M. Stock, F. Klee, K. Edlund, M. Grinberg, S. Hammad, R. Marchan, C. Cadenas, B. Niggemann, K.S. Zänker, J. Rahnenführer, M. Schmidt, J.G. Hengstler, F. Entschladen, Gelsolin Is Associated with Longer Metastasis-free Survival and Reduced Cell Migration in Estrogen Receptor-positive Breast Cancer., Anticancer Res. 35 (2015) 5277–5285.
- J. Rao, D. Seligson, H. Visapaa, S. Horvath, M. Eeva, K. Michel, A. Pantuck, A. Belldegrun, A. Palotie, Tissue microarray analysis of cytoskeletal actin-associated biomarkers gelsolin and E-cadherin in urothelial carcinoma., Cancer. 95 (2002) 1247–1257. [CrossRef]
- A. Van den Abbeele, V. De Corte, K. Van Impe, E. Bruyneel, C. Boucherie, M. Bracke, J. Vandekerckhove, J. Gettemans, Downregulation of gelsolin family proteins counteracts cancer cell invasion in vitro., Cancer Lett. 255 (2007) 57–70. [CrossRef]
- Z.-Y. Chen, P.-W. Wang, D.-B. Shieh, K.-Y. Chiu, Y.-M. Liou, Involvement of gelsolin in TGF-beta 1 induced epithelial to mesenchymal transition in breast cancer cells., J. Biomed. Sci. 22 (2015) 90. [CrossRef]
- Y. Zhang, X. Luo, J. Lin, S. Fu, P. Feng, H. Su, X. He, X. Liang, K. Liu, W. Deng, Gelsolin Promotes Cancer Progression by Regulating Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma and Correlates with a Poor Prognosis., J. Oncol. 2020 (2020) 1980368. [CrossRef]
- E. Tagliabue, S. Raimondi, S. Gandini, Meta-analysis of vitamin D-binding protein and cancer risk., Cancer Epidemiol. Biomarkers Prev. 24 (2015) 1758–1765. [CrossRef]
Figure 1.
A) Summary of the methodological approach. Dogs bearing mammary tumors were recruited for the study, and samples were collected for isolation of SEVs, their characterization, and assessment of their proteomic profile by mass spectrometry. B) Dogs recruited were assigned to five groups: G1 – simple carcinomas (malignant); G2 – mixed tumor carcinoma (benign); G3 – Relapse of disease; G4 – Remission of disease; G5 – Healthy controls. Dogs in the groups G1 and G2 had samples collected at two timepoints: G1.1 and G2.1 – before any surgical intervention; G1.2 and G2.2 – after mastectomy. The remission and relapse groups were independent, meaning that they do not have paired comparisons.
Figure 1.
A) Summary of the methodological approach. Dogs bearing mammary tumors were recruited for the study, and samples were collected for isolation of SEVs, their characterization, and assessment of their proteomic profile by mass spectrometry. B) Dogs recruited were assigned to five groups: G1 – simple carcinomas (malignant); G2 – mixed tumor carcinoma (benign); G3 – Relapse of disease; G4 – Remission of disease; G5 – Healthy controls. Dogs in the groups G1 and G2 had samples collected at two timepoints: G1.1 and G2.1 – before any surgical intervention; G1.2 and G2.2 – after mastectomy. The remission and relapse groups were independent, meaning that they do not have paired comparisons.
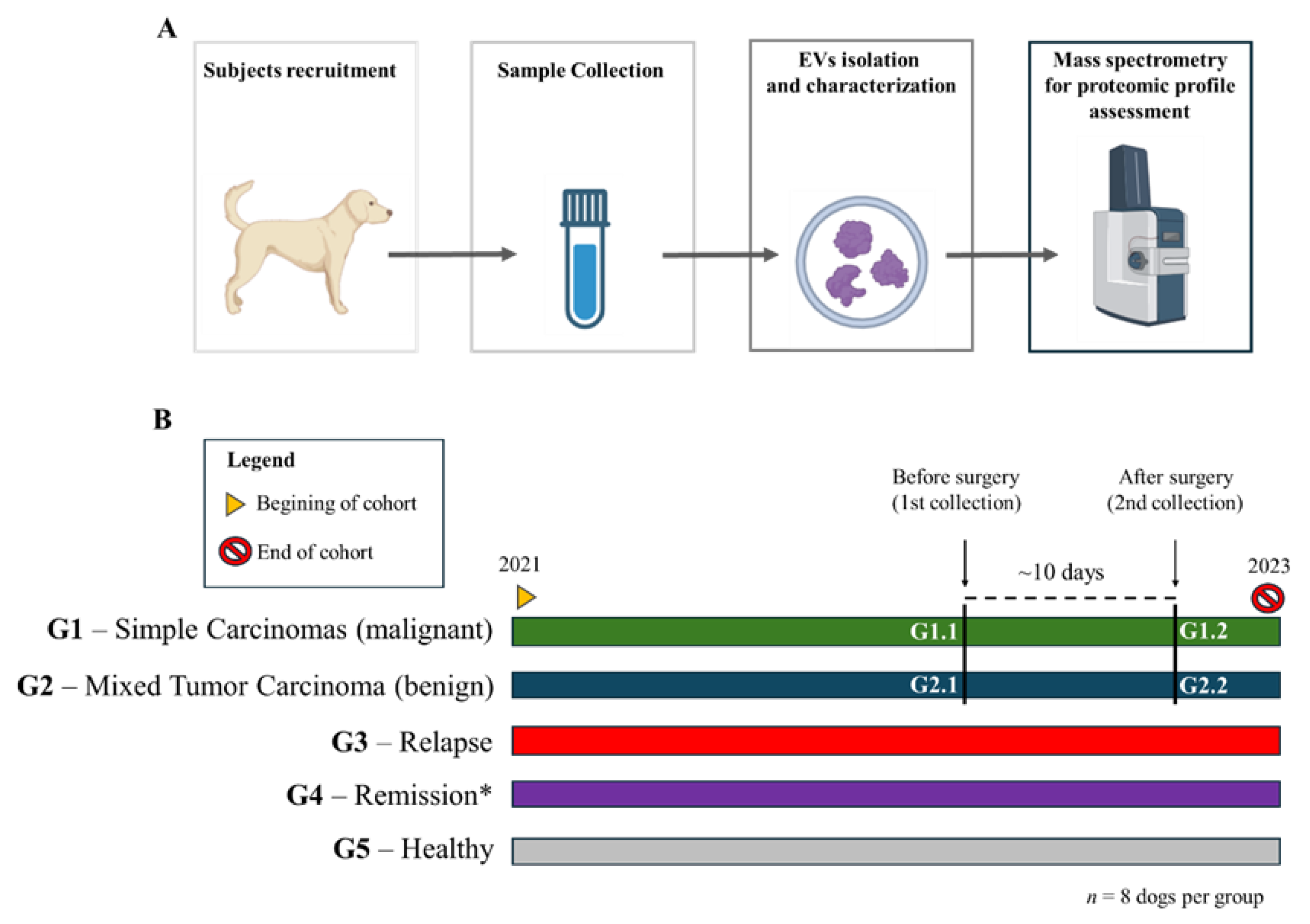
Figure 2.
Characterization of SEVs. Concentration of SEVs isolated from plasma samples of dogs with mammary tumors A) all groups without surgical intervention. Before and after mastectomy in B) Benign and C) Malignant tumors. D) Representative Nanoparticle Tracking Analysis (NTA) profile showing SEVs concentration, with values displayed as particles/mL (n=56). Size of SEVs in the respective groups: E) All groups without surgical intervention, before and after mastectomy in F) Benign and G) Malignant tumors. H) Representative NTA profile displaying size distribution, with values shown as arbitrary units (n=56). I) Transmission Electron Micrography (TEM) of SEVs isolated from plasma, magnified at 140,000 X. White arrows indicated SEVs in the grids. Legend: * - statistically significant; SEVs – small extracellular vesicles.
Figure 2.
Characterization of SEVs. Concentration of SEVs isolated from plasma samples of dogs with mammary tumors A) all groups without surgical intervention. Before and after mastectomy in B) Benign and C) Malignant tumors. D) Representative Nanoparticle Tracking Analysis (NTA) profile showing SEVs concentration, with values displayed as particles/mL (n=56). Size of SEVs in the respective groups: E) All groups without surgical intervention, before and after mastectomy in F) Benign and G) Malignant tumors. H) Representative NTA profile displaying size distribution, with values shown as arbitrary units (n=56). I) Transmission Electron Micrography (TEM) of SEVs isolated from plasma, magnified at 140,000 X. White arrows indicated SEVs in the grids. Legend: * - statistically significant; SEVs – small extracellular vesicles.
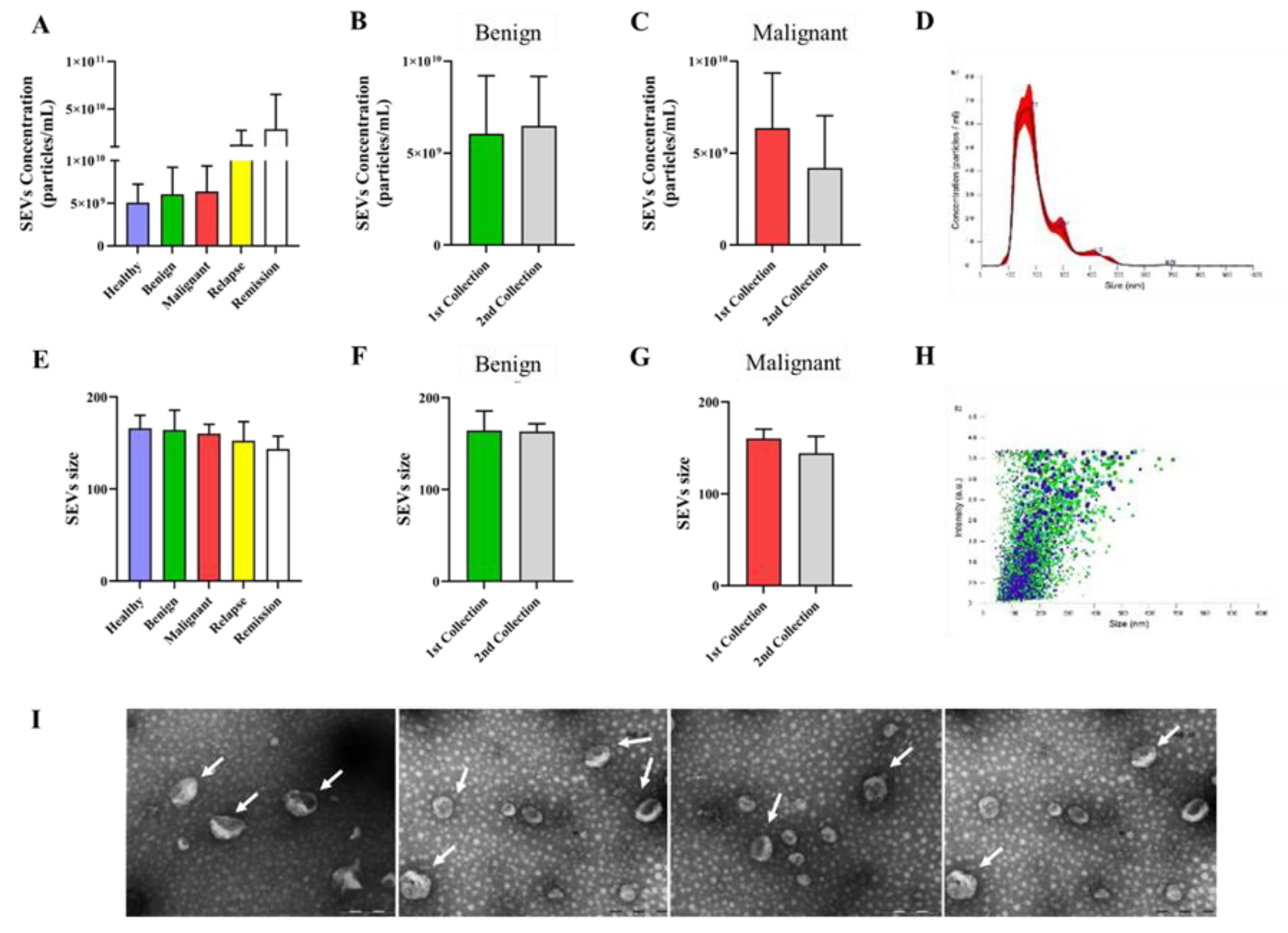
Figure 3.
Proteomic profile of SEVs isolated from dogs with benign or malignant tumors compared to healthy controls. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified in benign, malignant, and healthy subjects. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. C) Gene Ontology Enrichment analysis for the unique proteins identified in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log(pvalue). D) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database.
Figure 3.
Proteomic profile of SEVs isolated from dogs with benign or malignant tumors compared to healthy controls. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified in benign, malignant, and healthy subjects. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. C) Gene Ontology Enrichment analysis for the unique proteins identified in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log(pvalue). D) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database.
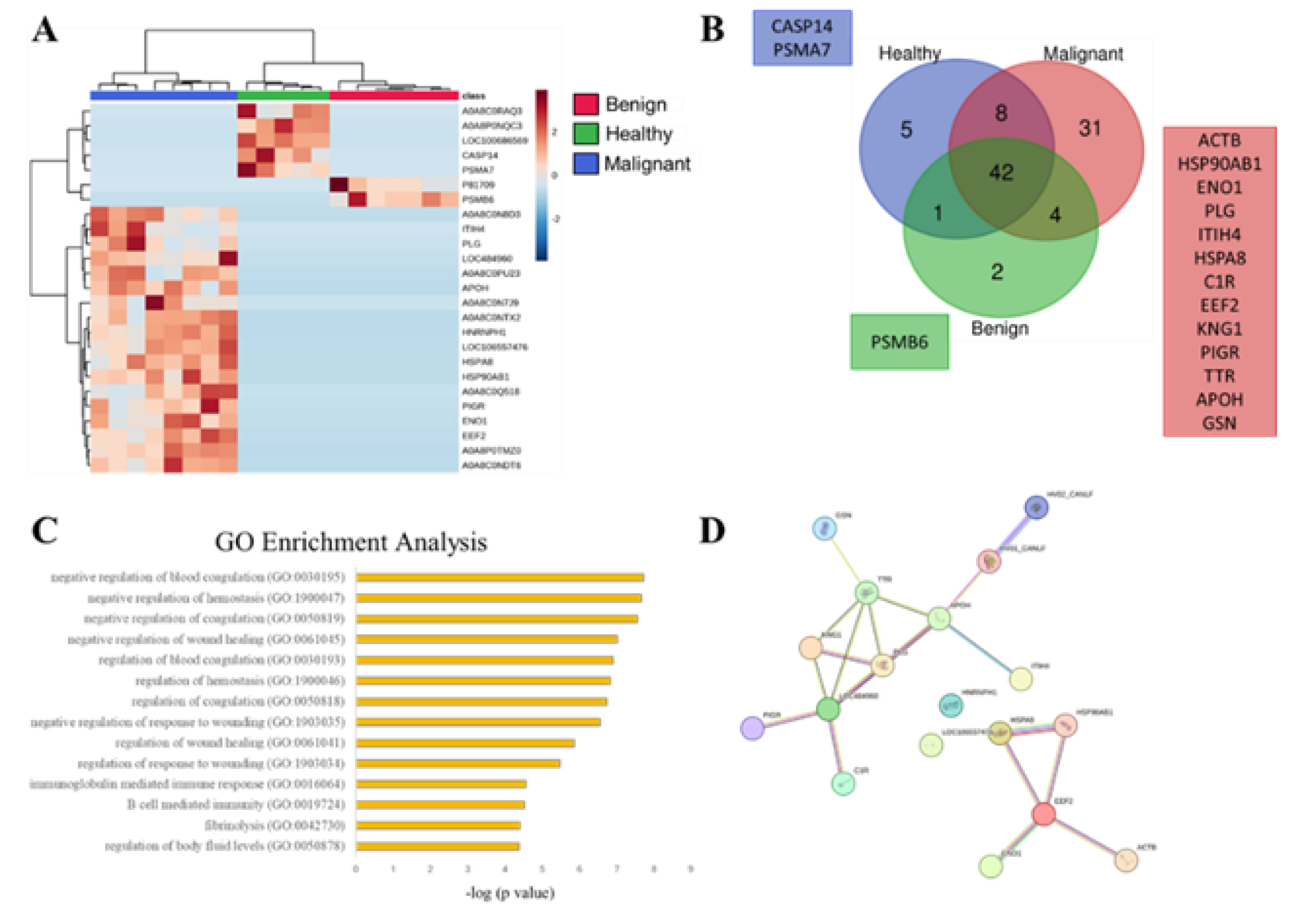
Figure 4.
Proteomic profile of SEVs isolated from dogs with benign or malignant mammary tumors before and after surgery. A) Heatmap displaying the top 25 different proteins found in benign condition. B) Heatmap displaying the top 25 different proteins found in malignant condition. C) Venn diagram illustrating the shared and unique proteins identified in benign tumors. D) Venn diagram illustrating the shared and unique proteins identified in malignant tumors before and after mastectomy. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. E) Gene Ontology Enrichment analysis for the unique proteins found in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log (p value). F) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database.
Figure 4.
Proteomic profile of SEVs isolated from dogs with benign or malignant mammary tumors before and after surgery. A) Heatmap displaying the top 25 different proteins found in benign condition. B) Heatmap displaying the top 25 different proteins found in malignant condition. C) Venn diagram illustrating the shared and unique proteins identified in benign tumors. D) Venn diagram illustrating the shared and unique proteins identified in malignant tumors before and after mastectomy. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. E) Gene Ontology Enrichment analysis for the unique proteins found in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log (p value). F) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database.
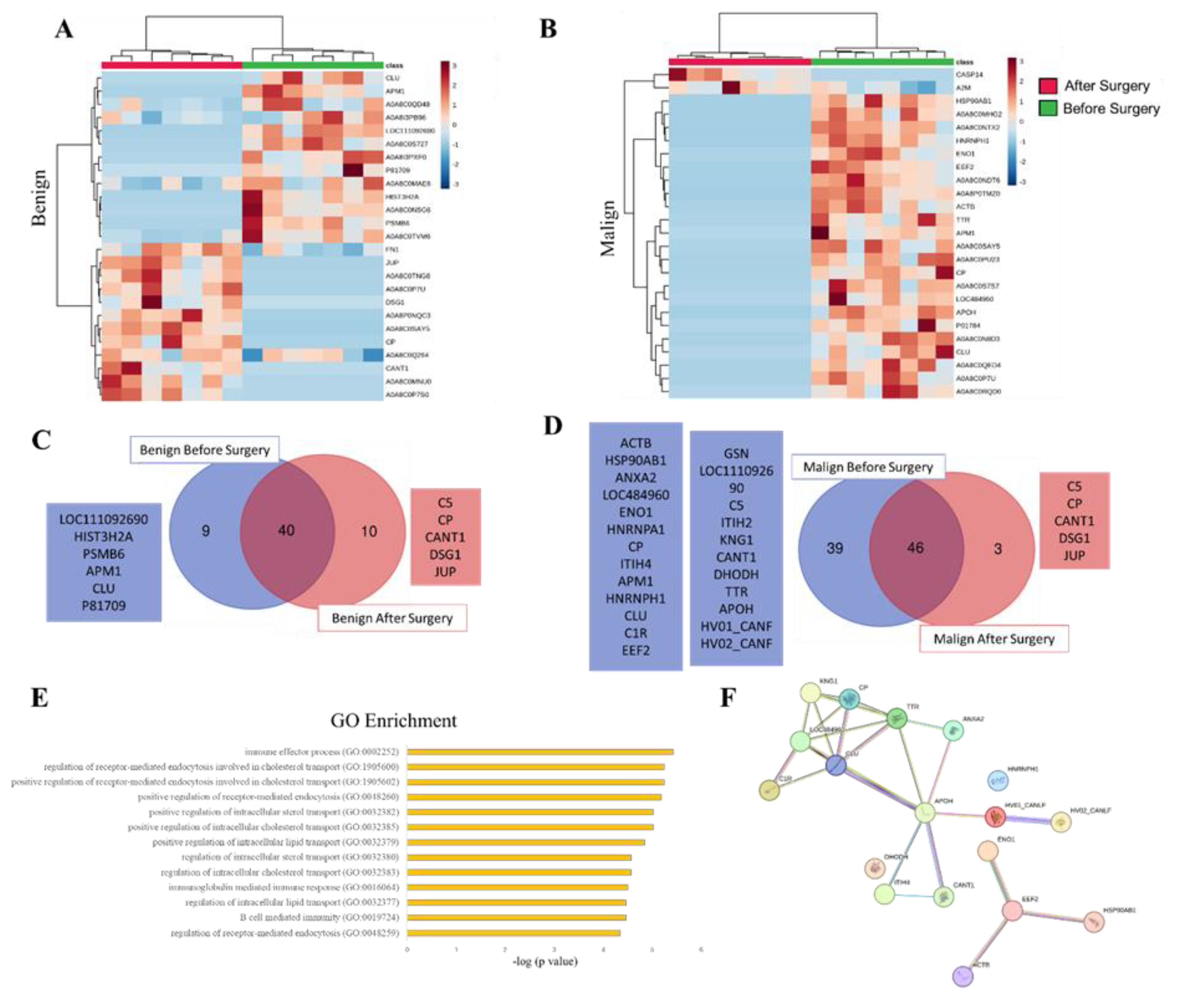
Figure 5.
Proteomic profile of SEVs isolated from dogs with malignant mammary tumors compared to SEVs from remission and relapse condition. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. C) Gene Ontology Enrichment analysis for the unique proteins found in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log (p value). D) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database. Neither remission nor relapse condition showed set enrichment. .
Figure 5.
Proteomic profile of SEVs isolated from dogs with malignant mammary tumors compared to SEVs from remission and relapse condition. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. C) Gene Ontology Enrichment analysis for the unique proteins found in the malignant group. Only enrichments with p < 0.05 and FDR < 0.05 are reported, displayed as -log (p value). D) Molecular interactions potentially involved in the unique proteins exclusively found in the malignant condition, retrieved from STRING database. Neither remission nor relapse condition showed set enrichment. .
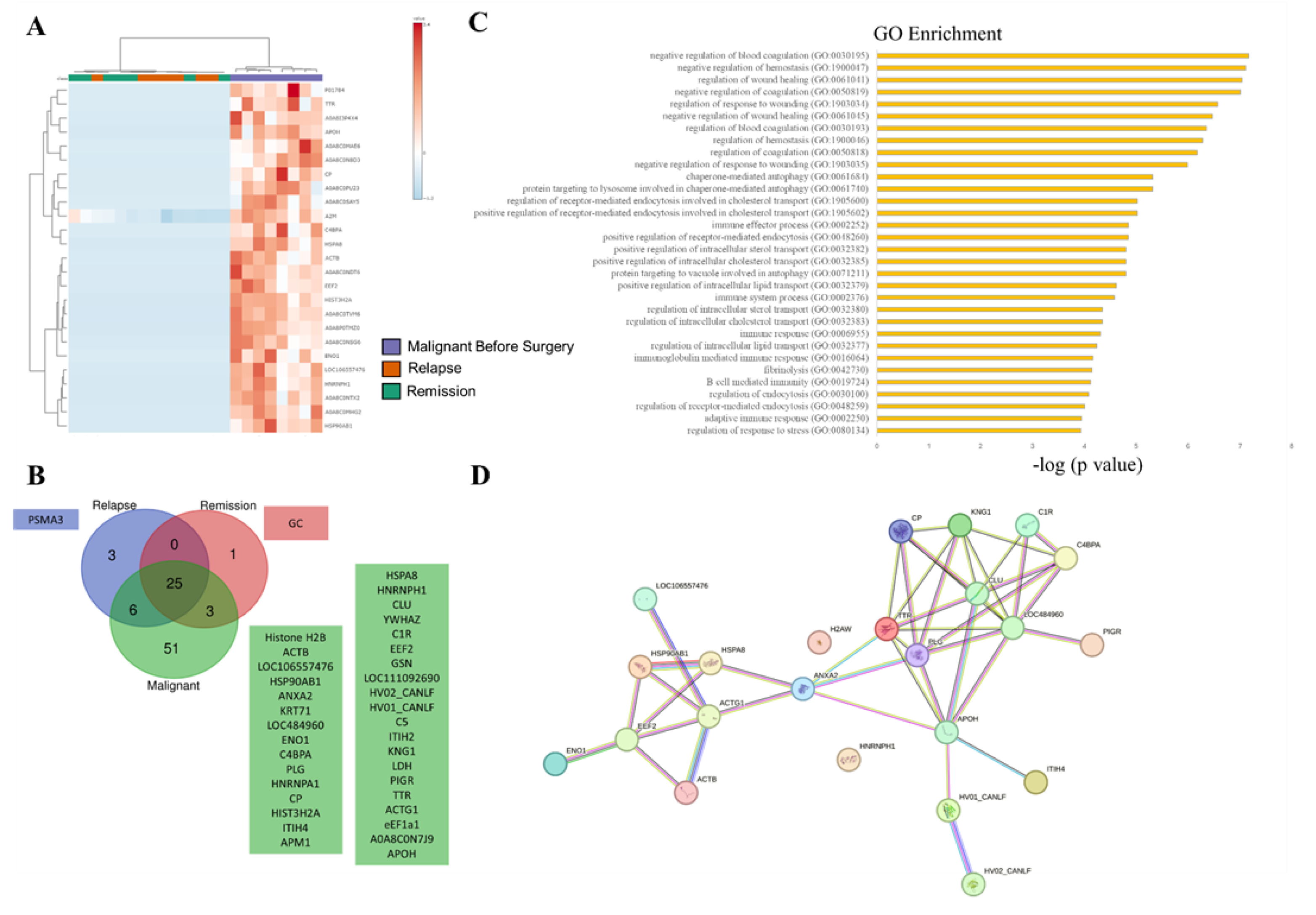
Figure 6.
Comparison of the SEVs proteomic profile isolated from dogs that underwent remission or relapse. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. Neither remission nor relapse showed set enrichment. .
Figure 6.
Comparison of the SEVs proteomic profile isolated from dogs that underwent remission or relapse. A) Heatmap displaying the top 25 different proteins found in each condition. B) Venn diagram illustrating the shared and unique proteins identified. Ig-like or uncertain proteins retrieved from Uniprot were excluded. Squares denote the list of exclusive proteins. Neither remission nor relapse showed set enrichment. .
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



