
Submitted:
23 May 2024
Posted:
23 May 2024
You are already at the latest version
Abstract
Objective: Daratumumab (Dara) is the first-in-class human-specific anti-CD38 mAb approved for treating multiple myeloma (MM). However, DARA-induced depletion of CD38high natural killer (NK) cells results in crippled antibody-dependent cellular cytotoxicity (ADCC), which limits its clinical detection and prediction for Dara efficacy. Therefore, we aimed to explore new NK subsets that could predict Dara efficacy and the underlying mechanism in Dara development.
Methods: Multicolour spectral flow cytometry monitored changes in the immune cells of 13 paired RRMM patients. We used machine learning-driven clustering, FlowSOM (Flow Self-Organizing Maps), and dimensional reduction with tSNE (t-distributed Stochastic Neighbour Embedding) to identify unique cell populations pre- and posttreatment. Then, in vitro, fratricide and proliferation assays were performed to explore the function of new NK subsets against MM under Dara treatment.
Results: The NK cells were divided into 25 subsets, among which we found that the percentages of KIR-NKP46+ NK cells decreased in paired RRMM patients compared to healthy controls but increased significantly post-Dara treatment. The fold change in KIR-NKP46+ after one dose of Dara treatment can predict whether the best response could be achieved within one month (ROC=0.86, p=0.011). Further in vitro experiments showed that KIR-NKP46+ NK cells are superior to KIR+ NK cells, cooperatively acting with daratumumab to sustainably eradicate MM cells. Meanwhile, apoptosis was significantly decreased, and the proliferation ability was relatively increased post-Dara treatment.
Conclusion: In conclusion, the fold change in KIR-NKP46+ NK cell subsets after one dose of Dara treatment can predict the outcome of the curative effect, indicating a putative role of KIR-NKP46+ subsets against MM cells. The mechanism attributed to a relative expansion of the subset due to decreased apoptosis and increased proliferation ability.
Keywords:
Natural killer cells
; daratumumab
; multiple myeloma
; KIR-NKP46+
Xun-Hong Cao, Xue-Lin Dou, Lei Wen, Xiao-Jun Huang, Jin Lu * and Xiang-Yu Zhao *
Introduction
Multiple myeloma (MM) is a malignant clonal plasma cell dyscrasia [1]. The prognosis varies greatly, and almost all patients face disease relapse. CD38 monoclonal antibody is the first monoclonal antibody drug approved for the treatment of MM and has shown efficacy in the treatment of relapsed and refractory MM [2,3,4]. Dara induces MM cell death through several mechanisms, including complement-dependent cytotoxicity (CDC), antibody-dependent cellular cytotoxicity (ADCC), antibody-dependent cellular phagocytosis (ADCP), apoptosis induced by cross-linking of CD38 on the target cells, and immunomodulatory effects via elimination of CD38+ immunosuppressive cells [5,6]. However, a simple and practical biomarker that can accurately predict the treatment response of MM patients undergoing Dara therapy is limited.
Natural killer (NK) cells are an important part of the innate immune system and play an important role in anti-infection and antitumour immunity. The cytotoxic activity of NK cells, particularly against MM cells, has been confirmed in many studies [7,8]. Nonetheless, significant changes in the quantity and quality of NK cells have been described in MM patients. Therefore, many studies have focused on enhancing the functional ability of NK cells, relieving the inhibitory effect of NK cells by MM niches, or the adoptive transfer of NK cells to increase their ability against MM cells. However, due to the highly expressed level of CD38 in NK cells, treatment with dara leads to rapid depletion of CD38high NK cells in patients, which results in crippled ADCC and consequently increases the incidence of infection [9]. Whether the subtype of NK cells before and after Dara treatment can predict the clinical response is currently unclear.
Previous studies have shown that surviving expanded CD38-knockout NK cells exhibit stronger cytotoxicity against MM cell lines and primary MM cells than CD38WT NK cells during Dara treatment, and adoptive infusion of CD38KO NK cells expanded in vitro could control MM.1S progression in mice [10,11]. Despite this, the prognostic role of CD38-negative NK cells in Dara treatment has not yet been determined. High-throughput spectral flow cytometry is a powerful tool to broadly assess surface markers on immune cells. Dimensional reduction and visualization with tSNE (t-distributed stochastic neighbour embedding) combined with machine learning-driven clustering with methods such as FlowSOM (Flow self-organizing maps) allow for discrimination of distinct immune cell clusters in an unbiased way. Therefore, we aimed to gain more insight into NK cell immunity in daratumumab-treated RRMM patients via spectral flow cytometry and found that KIR-NKP46+ NK subsets were resistant to Dara-induced apoptosis and correlated with clinical efficacy after Dara-treatment. Further functional analysis showed that KIR-NKP46+ NK subsets maintained strong proliferation ability and anti-MM effects during Dara treatment, indicating that higher KIR-NKP46+ NK cells post-Dara treatment may be a good indicator of a clinical response.
Materials and Methods
Patients
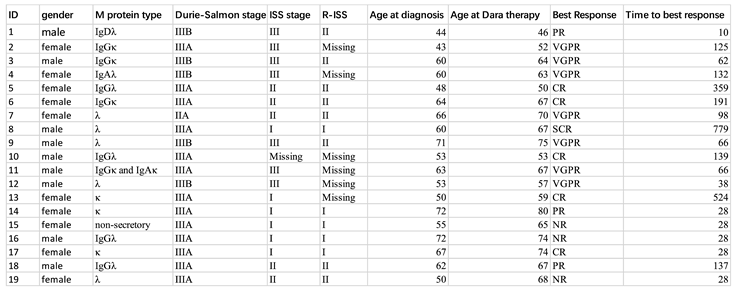
Paired peripheral blood samples from a total of 19 RRMM patients at Peking University of People’s Hospital before and after daratumumab treatment were collected. The clinical characteristics and outcomes of those patients are listed in Table 1. Among these 19 patients, 13 were monitored using a 5-laser 25-color panel (Table S1) and the other 6 patients were monitored using a 3-laser 9-color panel (Table S2). Written informed consent was obtained from all patients. Peripheral blood mononuclear cells (PBMCs) from those 13 patients were isolated using Ficoll density gradient centrifugation and cryo-stored before the experiment and the other 6 cases were newly enrolled patients.
Cell Lines
MM cell lines (U266 and LP-1) were kindly obtained from Professor G.R. Ruan’s laboratory and K562 cells that were maintained in RPMI-1640 medium supplemented with 10% heat-inactivated foetal bovine serum (FBS).
Multiparametric Spectral Cytometry Immunophenotypic Studies
Frozen PBMCs from 13-paried patients before and after one-dose daratumumab treatment as well as from healthy donors were rapidly thawed in a 37 °C water bath and then rested in prewarmed growth cultured medium overnight. The spectral cytometry panel including 25 antibody-fluorophore conjugates was listed in Table S1. All antibodies were titrated to find their best concentration and the results of similarity index matrix (SIM) which measures how similar two spectra are to each other as well as stain index were shown in Table S3(A,B). Flow cytometry was performed with a 5-Laser Cytek Aurora flow cytometer for those 13-paired patients using 25-color panel and new enrolled 6 patients were test using 3-Laser Cytek Aurora (Cytek® Biosciences Inc., Fremont, California, United States) via 9-color panel.
Cytotoxicity and Proliferation Assay
A flow-based cytotoxicity assay was performed as previously reported [12]. Briefly, PBMCs from healthy donors or RRMM patients were cocultured with U-266 and LP-1 cell lines at 5:1 effector(2*105)/target(4*104) ratios in 96-well u-shaped plate at 37 °C with or without daratumumab (MCE, 10 µg/ml) for 4 h, and the cytotoxicity of NK cells was measured via surface CD107a, followed by intracellular IFN-γ and TNF--α staining in another smaller panel as CD3,CD56,KIR,NKP46, IFN-γ and TNF--α.
To evaluate the fratricide of NK cell subsets, purified NK cells from PBMCs of RRMM patients were sorted first by negative selection (NK cell isolation Kit, Miltenyi, USA) and then further sorted into KIR+ and KIR- subpopulations via flow cytometry (ARIA SORP, BD, USA) via CD3-V500, CD56-APC and KIR-FITC. Purified NK subpopulations were cocultured with or without targets cells (U266, LP-1 and K562 cells, the K562 cells were treated as negative control to exclude education effect in NK cells) in the treatment of Dara at a 5:1 effector (1*105)/target (2*104) ratios for 5 hours. Before coculture, the targets were firstly labelled with CFSE(5μM). The cell death was monitored by flow cytometric analysis with an anti-Annexin V antibody and 7-ADD. In addition, KIR- NK cell subsets were blocked with or without NKP46 (5 µg/ml, BD).
For the cell proliferation assay, PBMCs from healthy donors or RRMM patients were labelled with Cell Trace CFSE Cell Proliferation Dye (Invitrogen) and placed in culture medium supplemented with 200 U IL-2 with or without daratumumab (10 ng/ml, MCE) for 7 days, followed by surface staining to test the proliferation of NK cells.
Data Analysis
Data analysis was performed on the unmixed FCS files using FlowJo software (FlowJo, LLC). The measurements and spectral unmixing were performed using Spectroflo® Software (Cytek Biosciences Inc). To check that the unmixing of the 25-color panel was accurate, data was cleaned up (singlets, live, scatter gate) and NxN plot permutations were screened as shown in Figure S1B.
For clustering analysis, data were manually compensated, and live single cells were gated based on forwards scatter, and side scatter (Figure S1A). To remove anomalous events, FlowAI was applied based on the flow rate and dynamic range of fluorochrome parameters. Next, NK cells were selected by gating for low side scatter CD45+ events and further separated into CD3-CD56+ subsets. For NK cells, only healthy control samples were down-sampled to equal numbers, while all NK cells from patients post daratumumab treatment were included due to their low numbers. Clustering of the NK cells was based on panel from Table S1.
Statistical Analysis
Visualizations and statistical analyses were performed in either GraphPad Prism v8.4.3 (GraphPad Software, California, United States) or R. Continuous outcome measures were compared between two groups using multiple unpaired t tests. The Holm‒Sidak correction was applied to correct for multiple testing. Statistical significance was set as an adjusted p value of <0.05.
Results
Cohort Characteristics
In the current cohort, 19 paired RRMM patients treated with daratumumab and 13 among them were included in the higher dimensional analysis. The clinical characteristics, including age, sex, and response to daratumumab, are summarized in Table 1.
For spectral flow cytometry, frozen PBMCs from 13 paired RRMM patients and 7 healthy donors were stained with the 25 monoclonal antibodies listed in Table S1.
An unsupervised cell clustering view of the immune cell landscape before and after daratumumab treatment in RRMM patients
To investigate immunological changes in pre- and post-daratumumab treated RRMM patients, we evaluated healthy donors and paired analyses of PBMCs from RRMM patients by spectral cytometry using a broad immunophenotype panel and NK cell-focused panel.
As an overview of our approach, we first applied unsupervised cell clustering using FlowSOM (Flow Self-Organizing Maps) and dimensionality reduction by tSNE to the cross-sectional spectral cytometry from 13 paired RRMM patients before and post Dara treatment and 7 healthy donors to identify distinct populations in an unbiased way.
For an initial, high-level view of the circulating immune cell populations, we first performed tSNE clustering of immunophenotype panel spectral cytometry data from the cross-sectional cohorts (Figure 1A). tSNE allowed clear visualization of distinct immune cell clusters, including major CD3+ T-cell populations, CD3-CD56+ NK cells, CD3-CD19+ B cells, and CD3+CD56+ NKT cells. Among these high-level immune cell subsets, the proportion of CD3-CD56+ NK and CD19+ B cells among nuclear cells was significantly higher in RRMM patients regardless of whether they were treated with Dara than in healthy controls, while CD3+ T-cell populations were lower in RRMM patients before and after Dara treatment (Figure 1B). The absolute number of NK cells were also significantly decrease post Dara treatment (Figure1C, p=0.0016)
Increased Activated KIR-NKP46+ NK Cells Post Daratumumab Treatment
Next, we investigated differences in CD3-CD56+ NK cell populations in healthy donors and paired RRMM patients. The tSNE diagram shows
the distribution of NK subsets among the healthy donors and diversely paired with RRMM patients, suggesting a great impact of tumour and Dara treatment on NK cell subsets (Figure 2A). To identify cell populations that differ between controls and paired patients, we clustered NK cells based on the 20-marker NK cell-focused panel using FlowSOM. We then compared the abundances of the clusters between healthy donors and paired RRMM patients and identified metacluster 9 as significantly decreased in paired RRMM patients but increased posttreatment compared to pre-treatment (Figure 2B). Heatmap expression analysis revealed that metacluster 9 contained cells with higher expression of NKP46 and lower expression of KIR, and there was a total of 25 pops among the three groups, while the pop6 subset contained cells with high expression of Tigit, MICA, and CD155. The pop9 subset showed higher expression of NKG2A. (Figure 2B,C) Manual gating proved that the percentages of KIR-NKP46+ (%of NK cells) were significantly decreased before and post Dara treatment in RRMM patients compared with the healthy controls but increased significantly post-Dara treatment (Figure 2D). However, the absolute number of KIR-NKP46+ NK cells were showed no significant statistical difference pre and post Dara treatment (Figure S2B).
To explore the properties of KIR-NKP46+ NK subsets in more detail, we further evaluated the expression of activating and inhibitory receptors compared with KIR+, KIR-, and NKP46+ NK subsets among healthy donors and paired RRMM patients by manual gating. The results showed that the KIR-NKP46+ NK subsets in RRMM patients without Dara treatment tended to be more activated than those in the healthy controls, as the percentages of CD226, CD122, and CD38 on the NK subsets were higher than those in the healthy controls (Figure 2E). Thus, suggesting an activation state of RRMM patients without Dara treatment, which is in accordance with a report from Souza et al. [13]. We further performed a detailed analysis of receptor expression between the KIR-NKP46+ NK subsets and other subsets before and after Dara treatment. The results showed that the percentages of CD122, NKP30, and CD38 on KIR-NKP46+ NK subsets were higher than those on KIR+NKP46+ and KIR- NK subsets whether pre- or post-Dara treatment, suggesting a more activated state of KIR-NKP46+ subpopulations (Figure 2F). While the inhibitory receptors Tigit showed no significant difference among NK subsets in paired RRMM patients except for the higher percentages of NKG2A expression in KIR-NKP46+ cells in pre- and posttreatment patients (Figure S2A), activating receptors such as CD226(DNAM-1), NKG2D, and CD16 showed no significant differences among the groups (data not shown).
KIR-NKP46+ Subpopulations Are Resistant to Dara-Induced NK Cell Fratricide and Exert Sustainable Anti-MM Effects
To clarify the functional status of KIR-NKP46+ NK subsets post-Dara stimulation, we first evaluated the cytotoxicity of NK subsets when cocultured with U-266 and LP-1 cell lines. Interestingly, we found that the percentages of CD107a and TNF-α in NK subsets in RRMM patients after Dara stimulation in vitro significantly decreased compared with those in healthy donors when cocultured with LP-1 and U266 cells (Figure 3A, Figure S3A), suggesting impaired NK cell function in MM patients.
Further analysis into the function of KIR-NKP46+ NK cells showed no significant difference in CD107a expression in RRMM patients among KIR+, KIR- and KIR-NKP46+ subpopulations when cocultured with U266 cells with or without Dara stimulation, while higher CD107a expression was found in KIR- NK subsets than in KIR+ NK and KIR-NKP46+ NK subpopulations when cocultured with LP-1 cells with or without Dara treatment in RRMM patients (Figure 3B, p= 0.0260, p=0.0109), and KIR-NKP46+ NK subsets tended to have higher expression of CD107a than KIR+ subsets when cocultured with LP-1 cells, although there was no significant difference. Meanwhile, KIR-NKP46+ NK cells from the bone marrow of RRMM patients also showed an increased tendency to kill MM cell lines under Dara stimulation compared to KIR+ NK subsets when cocultured with LP-1 cells and U266 cells (Figure 3B).
Previous studies have shown that Dara treatment leads to fratricide of NK cells, which is the main reason for the decreased absolute number of NK cells after Dara treatment in vivo [10]. However, our study found that the percentages of KIR-NKP46+ NK subsets increased significantly post-Dara treatment, while the absolute number of KIR-NKP46+ NK cells showed no statistical difference under the significant decreased number of NK cells post-Dara treatment, suggesting a relative expansion of this subset in NK cells. We therefore evaluated the proliferation and fratricide of KIR-NKP46+ NK cells under Dara stimulation in vitro. The results showed that Dara treatment significantly decreased the proliferation of NK cells in healthy donors but decreased the proliferation of NK cells in RRMM patients (p=0.0440 and p=0.1081, respectively, Figure S3B). However, further subpopulation analysis showed that the proliferation ability of the KIR-NKP46+ NK cell subset was significantly higher than that of other subpopulations (KIR-NKP46-,KIR+NKP46-,KIR+NKP46+) under Dara stimulation (Figure 3D, Figure S3C, p=0.0493), resulting in a relatively higher proportion of KIR-NKP46+ NK cells after Dara treatment. Furthermore, we evaluated the fratricide of KIR+ NK cells and KIR- NK cells with or without NKP46 blockade. Remarkably, Dara-induced early and late apoptosis was more frequently observed in KIR+ NK cells (p=0.0489 and p= 0.0417, respectively, Figure 3E, Figure S3D) than in KIR- subsets without NKP46 blockade, and NKP46 blockade tended to impair the ability of KIR-NKP46+ NK cells to resist late apoptosis compared to KIR-NKP46- NK cells, although no significant difference was shown.
Considering that KIR receptors are important receptors involved in the education process of NK cells and are related to effector function [14], which determines that the fratricide effect of NK cells may be affected differently when against tumour cells. Therefore, we purified KIR+ and KIR- were co-incubated with LP-1 and U266. K562 is a chronic myelogenous leukaemia (CML)cell line expressing no CD38 as a negative control. The result showed that KIR+ NK cells had a higher percentage of 7ADD+AnnexinV+(% of NK cells) when cocultured with LP-1 and there is a higher tendency when cocultured with U266 under the stimulation of Dara. While no significant difference was seen when cocultured with K562 cells, further indicating that the fratricide of NK cells depends on the ADCC effect (Figure S3E). Interestingly enough, we found that KIR- and KIR+ NK cells showed comparable percentage of 7ADD+Annexinv+ cells(%of target cells) whether cocultured with U266 or LP-1 cells under the stimulation of Dara treatment, while significantly higher apoptosis rates of K562 cells were found in Dara stimulated KIR-NK cells compared with KIR+ NK cells(Figure S3F). Together, these results indicate that compared to KIR+ NK cells or KIR-NKP46- NK cells, the KIR-NKP46+ subset is superior at acting cooperatively with Dara to sustainably eradicate MM cells due to the decreased apoptosis and relatively increased proliferation ability.
Discussion
By broad cellular immunophenotyping profiling, we identified multiple changed immune cell populations in RRMM patients pre- and post-Dara treatment. We found that NK cells were significantly higher in RRMM patients, whether treated with Dara or not, than in healthy donors. In addition, NK cells in RRMM patients correlated with an activated phenotype. However, the percentages of CD107a and TNFα expression by NK cells against MM cell lines were significantly lower compared with the healthy controls, indicating an inversion of the quality and quantity of NK cells in the MM tumour environment, which is in accordance with Souza et al.’s report [13]. Interestingly, we found that the secretion of CD107a and TNFα in NK cells of RRMM patients significantly decreased compared with that in NK cells of healthy donors after Dara treatment in vitro, but it had no effect on the secretion of IFN-γ. A previous study showed that IFN-γ production and degranulation are differentially regulated in response to stimulation in murine NK cells [15], indicating that Dara mainly plays an anti-MM role by enhancing the NK cell degranulation pathway.
Further analysis of NK cells (CD3-CD56+) using tSNE showed diverse distributions of NK cell subsets among the healthy donors and paired RRMM patients, indicating a huge impact of the MM tumour environment on the phenotype of NK cells [16], and Dara treatment may function greatly on the quantity of NK depletion. Flowsome clustering and manual gating analysis found that KIR-NKP46+ significantly increased post Dara treatment, and a higher frequency of KIR-NKP46+ subsets post Dara treatment correlated with a better response to Dara treatment. Although in vitro studies found that the addition of a clinical anti-KIR antibody (IPH2101) increased NK cell cytotoxicity against HLA-C-positive acute myeloid leukaemia and lymphoma cells [17], less clear clinical responses were found in relapsed/refractory MM patients in a phase I clinical study [18] and smouldering MM patients in a phase II trial [19]. However, IPH2101 combined with an immunomodulatory augmented NK cell function and resulted in an objective response [20,21], indicating that the combination effect of anti-KIR and Dara treatment is involved in the anti-MM effect, which suggests the importance of the KIR- NK subset against MM in the setting of Dara treatment.
In addition, a previous report showed that the activating receptor NKP46 in NK cells plays an important role against MM cells in vitro; therefore, we speculated that the anti-MM effect of the KIR-NKP46+ subpopulation is stronger under Dara stimulation. We found that the percentages of CD107a in KIR-NKP46+ NK cells were comparable to those in KIR+ and KIR- NK cells, suggesting that the overall anti-MM ability of NK cells was enhanced after Dara treatment. Although Dara can significantly enhance the anti-MM effect of NK cells, it can also cause NK cell suicide and therefore reduce the number of NK cells. Previous studies have found that CD38-negative NK cells can resist NK cell fratricide and therefore continue to play a sustained anti-MM role. We also found that KIR-NKP46+ NK cells had less fratricide compared with other subsets and sustained a higher proliferation ability among the NK cells, which can explain its predictive effect on Dara efficacy. Interestingly, we found higher expression of CD38 in KIR-NKP46+ NK subsets with less fratricide. This may be due to the impaired expression of CD16 in the KIR-NKP46+ NK subset, as fratricide of NK cells induced by Dara treatment is mainly caused by the ADCC effect [10]. Therefore, the mechanism of KIR-NKP46+ NK cell resistance to NK cell fratricide needs to be determined in the future.
Compared to KIR-NK cells, KIR+ NK cells are considered licensed and therefore exhibit lower levels of robust activation. However, the functional differences between KIR- and KIR+ NK cells, resulting from educational variances, are also influenced by the cytokine environment. For instance, the presence of IL-12, IL-15, and IL-18 has been shown to decrease KIR downregulation, thereby enhancing the killing of tumour cells by NK cells [22]. In our study, we have demonstrated that sorted healthy KIR- NK cells exhibit comparable anti-multiple myeloma (MM) effects to KIR+ NK cells when stimulated with Dara treatment. This phenomenon may be attributed to the higher level of fratricide (NK cells killing each other) observed in KIR+ NK cells. Furthermore, the fratricide level is significantly higher in KIR+ NK cells compared to KIR- NK cells, suggesting that a relatively higher proportion of KIR- NK cells may survive in the body and exert a sustained anti-MM effect.
Our study has several limitations. The relatively small cohort of RRMM patients treated with Dara limits the ability to identify correlated immune features and precludes exact evaluation of clinical correlates of the cellular features identified. A larger cohort will be required in subsequent studies to determine the immune features detected here. In addition, our study focuses only on immune changes in peripheral blood samples and does not contain bone marrow samples. As there are few MM cells in peripheral blood, the determination of NK-related receptor ligands cannot be performed. Nonetheless, the substantial alterations demonstrated in circulating immune cells from RRMM patients’ response to Dara treatment support the idea that clinically relevant signals may be detectable in NK cells from PB samples.
In conclusion, this study highlighted that the expansion of KIR-NKP46+ NK subsets in RRMM patients post Dara treatment correlated with a better clinical response to Dara treatment, which may be attributed to the decreased apoptosis and relatively increased proliferation ability under Dara treatment compared with other subsets. In the future, the KIR-NKP46+ subsets might be superior at acting cooperatively with daratumumab to sustainably eradicate MM cells.
Author Contributions
J.L., and X.Z. designed the study. X.C. and X.D. performed data analysis. X.C. performed the experiments. X.H., X.Z.,X.D., and J.L. drafted the manuscript. X.D and L.W. collected the clinical samples. All authors read and approved the final version.
Funding
This study was supported by grants from the National Natural Science Foundation of China (Grant No. 81670166, 81870140, 82070184 and 81621001), Peking University People’s Hospital Research and Development Funds (No. RDL2021-01), Beijing Life Oasis Public Service Center (No. CARTFR-01), and CSH Young Scholars and 3SBio Pharmaceutical joint research project (No. KYC2201001).
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Acknowledgment
Thanks to Cyteck for the technical support.
Declaration of interests
The authors declare no competing financial interests.
References
- Kumar, S.K.; Rajkumar, V.; Kyle, R.A.; et al. Multiple myeloma. Nat Rev Dis Primers. 2017, 3, 17046. [Google Scholar] [CrossRef]
- Usmani, S.Z.; Weiss, B.M.; Plesner, T.; Bahlis, N.J.; Belch, A.; Lonial, S.; Lokhorst, H.M.; Voorhees, P.M.; Richardson, P.G.; Chari, A.; et al. Clinical efficacy of daratumumab monotherapy in patients with heavily pretreated relapsed or refractory multiple myeloma. Blood 2016, 128, 37–44. [Google Scholar] [CrossRef]
- Bhatnagar, V.; Gormley, N.J.; Luo, L.; et al. FDA approval summary: Daratumumab for treat- ment of multiple myeloma after one prior therapy. Oncologist. 2017, 22, 1347–1353. [Google Scholar] [CrossRef]
- Dimopoulos, M.A.; Oriol, A.; Nahi, H.; et al. ; POLLUX Investigators. Daratumumab, lenali- domide, and dexamethasone for multiple myeloma. N Engl J Med. 2016, 375, 1319–1331. [Google Scholar]
- Van De Donk, N.W.C.J.; Janmaat, M.L.; Mutis, T.; Lammerts Van Bueren, J.J.; Ahmadi, T.; Sasser, A.K.; Lokhorst, H.M.; Parren, P.W.H.I. Monoclonal antibodies targeting CD 38 in hematological malignancies and beyond. Immunol. Rev. 2016, 270, 95–112. [Google Scholar] [CrossRef]
- Krejcik, J.; Casneuf, T.; Nijhof, I.S.; et al. Daratumumab depletes CD38 immune regulatory cells, promotes T-cell expansion, and skews T-cell repertoire in multiple mye- loma. Blood. 2016, 128, 384–394. [Google Scholar] [CrossRef]
- Bigley A B, Spade S, Agha N H. ; et al.FcεRIγ-negative NK cells persist in vivo and enhance efficacy of therapeutic monoclonal antibodies in multiple myeloma. Blood advances 2021, 5, 3021–3031.
- Sivori, S.; Pende, D.; Bottino, C.; Marcenaro, E.; Pessino, A.; Biassoni, R.; Moretta, L.; Moretta, A. NKp46 is the major triggering receptor involved in the natural cytotoxicity of fresh or cultured human NK cells. Correlation between surface density of NKp46 and natural cytotoxicity against autologous, allogeneic or xenogeneic target cells. Eur. J. Immunol. 1999, 29, 1656–1666. [Google Scholar] [CrossRef]
- Nahi, H.; Chrobok, M.; Gran, C.; Lund, J.; Gruber, A.; Gahrton, G.; Ljungman, P.; Wagner, A.K.; Alici, E. Infectious complications and NK cell depletion following daratumumab treatment of Multiple Myeloma. PLOS ONE 2019, 14, e0211927. [Google Scholar] [CrossRef]
- Kararoudi, M.N.; Nagai, Y.; Elmas, E.; Pereira, M.d.S.F.; Ali, S.A.; Imus, P.H.; Wethington, D.; Borrello, I.M.; Lee, D.A.; Ghiaur, G. CD38 deletion of human primary NK cells eliminates daratumumab-induced fratricide and boosts their effector activity. Blood The Journal of the American Society of Hematology 2020, 136, 2416–2427. [Google Scholar] [CrossRef]
- Clara, J.A.; Levy, E.R.; Reger, R.; Barisic, S.; Chen, L.; Cherkasova, E.; Chakraborty, M.; Allan, D.S.J.; Childs, R. High-affinity CD16 integration into a CRISPR/Cas9-edited CD38 locus augments CD38-directed antitumor activity of primary human natural killer cells. J. Immunother. Cancer 2022, 10, e003804. [Google Scholar] [CrossRef]
- Kim, J.; Phan, M.-T.T.; Kweon, S.; Yu, H.; Park, J.; Kim, K.-H.; Hwang, I.; Han, S.; Kwon, M.-J.; Cho, D. A Flow Cytometry-Based Whole Blood Natural Killer Cell Cytotoxicity Assay Using Overnight Cytokine Activation. Front. Immunol. 2020, 11, 1851. [Google Scholar] [CrossRef]
- D’souza, C.; Keam, S.P.; Yeang, H.X.A.; Neeson, M.; Richardson, K.; Hsu, A.K.; Canfield, R.; Bezman, N.; Robbins, M.; Quach, H.; et al. Myeloma natural killer cells are exhausted and have impaired regulation of activation. Haematologica 2021, 106, 2522–2526. [Google Scholar] [CrossRef]
- Anfossi, N.; André, P.; Guia, S.; Falk, C.S.; Roetynck, S.; Stewart, C.A.; Breso, V.; Frassati, C.; Reviron, D.; Middleton, D.; et al. Human NK Cell Education by Inhibitory Receptors for MHC Class I. Immunity 2006, 25, 331–342. [Google Scholar] [CrossRef]
- Vahlne, G.; Becker, S.; Brodin, P.; Johansson, M.H. IFN-γ Production and Degranulation are Differentially Regulated in Response to Stimulation in Murine Natural Killer Cells. Scand. J. Immunol. 2007, 67, 1–11. [Google Scholar] [CrossRef]
- Pazina, T.; MacFarlane, I.V. ; A. W.; Bernabei, L.; et al.Alterations of NK cell phenotype in the disease course of multiple myeloma. Cancers 2021, 13, 226. [Google Scholar]
- Romagne, F.; Andre, P.; Spee, P.; Zahn, S.; Anfossi, N.; Gauthier, L.; et al. Preclinical characterization of 1-7F9, a novel human anti-KIR receptor therapeutic antibody that augments natural killer-mediated killing of tumor cells. Blood 2009, 114, 2667–77. [Google Scholar] [CrossRef]
- Benson, D.M.; Hofmeister, C.C.; Padmanabhan, S.; Suvannasankha, A.; Jagannath, S.; Abonour, R.; et al. A phase 1 trial of the anti-KIR antibody IPH2101 in patients with relapsed / refractory multiple myeloma. Blood 2012, 120, 4324–33. [Google Scholar] [CrossRef]
- Korde, N.; Carlsten, M.; Lee, M.-J.; Minter, A.; Tan, E.; Kwok, M.; Manasanch, E.; Bhutani, M.; Tageja, N.; Roschewski, M.; et al. A phase II trial of pan-KIR2D blockade with IPH2101 in smoldering multiple myeloma. Haematologica 2014, 99, e81–e83. [Google Scholar] [CrossRef]
- Benson, D.M.; Cohen, A.D.; Jagannath, S.; Munshi, N.C.; Spitzer, G.; Hofmeister, C.C.; Efebera, Y.A.; Andre, P.; Zerbib, R.; Caligiuri, M.A. A Phase I Trial of the Anti-KIR Antibody IPH2101 and Lenalidomide in Patients with Relapsed/Refractory Multiple Myeloma. Clin. Cancer Res. 2015, 21, 4055–4061. [Google Scholar] [CrossRef]
- Kohrt, H.E.; Thielens, A.; Marabelle, A.; Sagiv-Barfi, I.; Sola, C.; Chanuc, F.; Fuseri, N.; Bonnafous, C.; Czerwinski, D.; Rajapaksa, A.; et al. Anti-KIR antibody enhancement of anti-lymphoma activity of natural killer cells as monotherapy and in combination with anti-CD20 antibodies. Blood 2014, 123, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Ewen, E.M.; Pahl, J.H.; Miller, M.; et al. KIR downregulation by IL-12/15/18 unleashes human NK cells from KIR/HLA-I inhibition and enhances killing of tumor cells. European journal of immunology 2018, 48, 355–365. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
(A):Tsne distribution of CD45+ positive cells among the healthy donors, paired RRMM patients before and post dara treatment. (B): The percentages of lymophocytes among the healthy donors, paired patients before and post dara treatment.(From left to right),(left):the percentages of NK cells in CD45+ cells;(middle): the percentages of T cells in CD45+ cells;(right):the percentages of B cells in CD45+ cells.
Figure 1.
(A):Tsne distribution of CD45+ positive cells among the healthy donors, paired RRMM patients before and post dara treatment. (B): The percentages of lymophocytes among the healthy donors, paired patients before and post dara treatment.(From left to right),(left):the percentages of NK cells in CD45+ cells;(middle): the percentages of T cells in CD45+ cells;(right):the percentages of B cells in CD45+ cells.
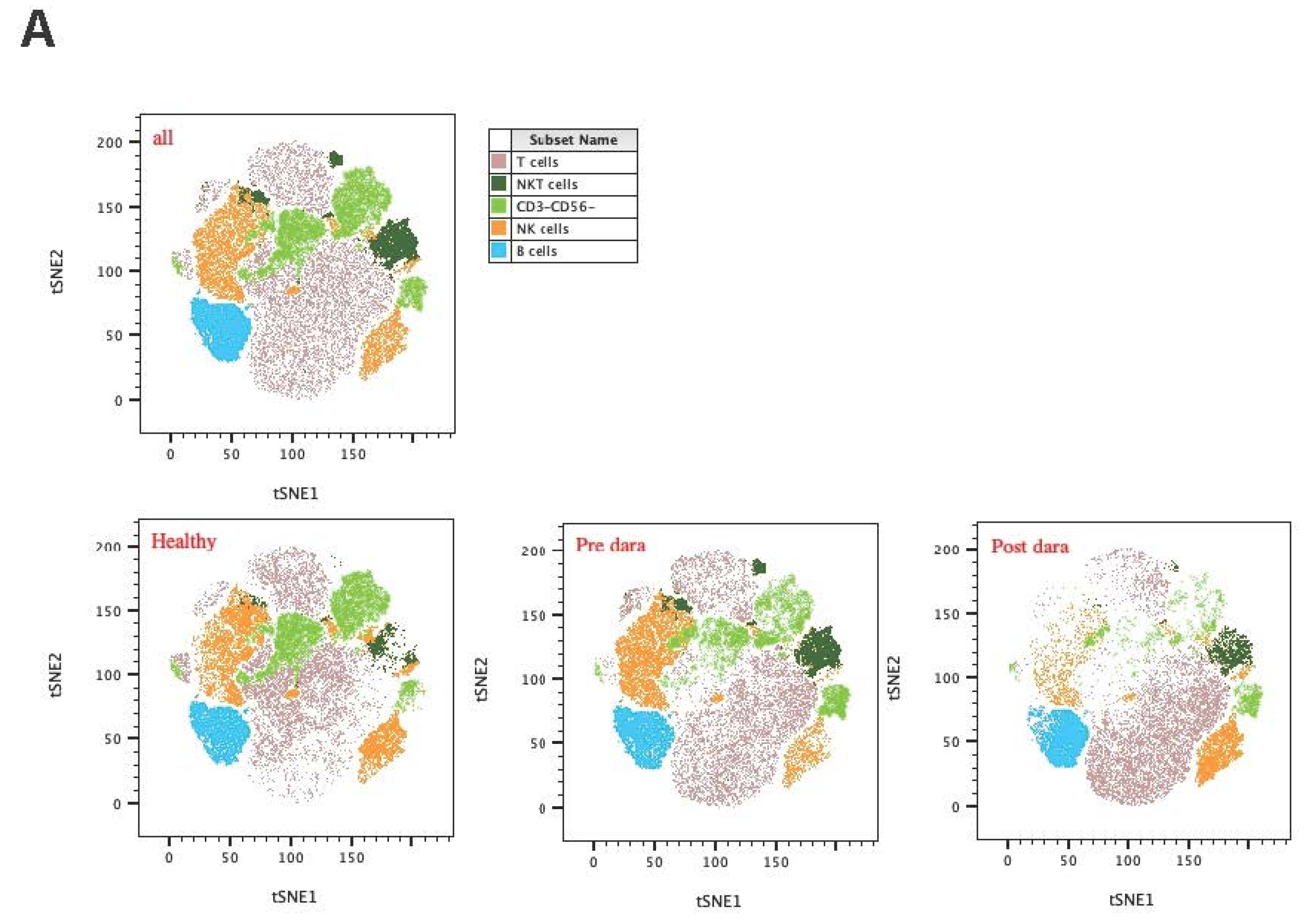
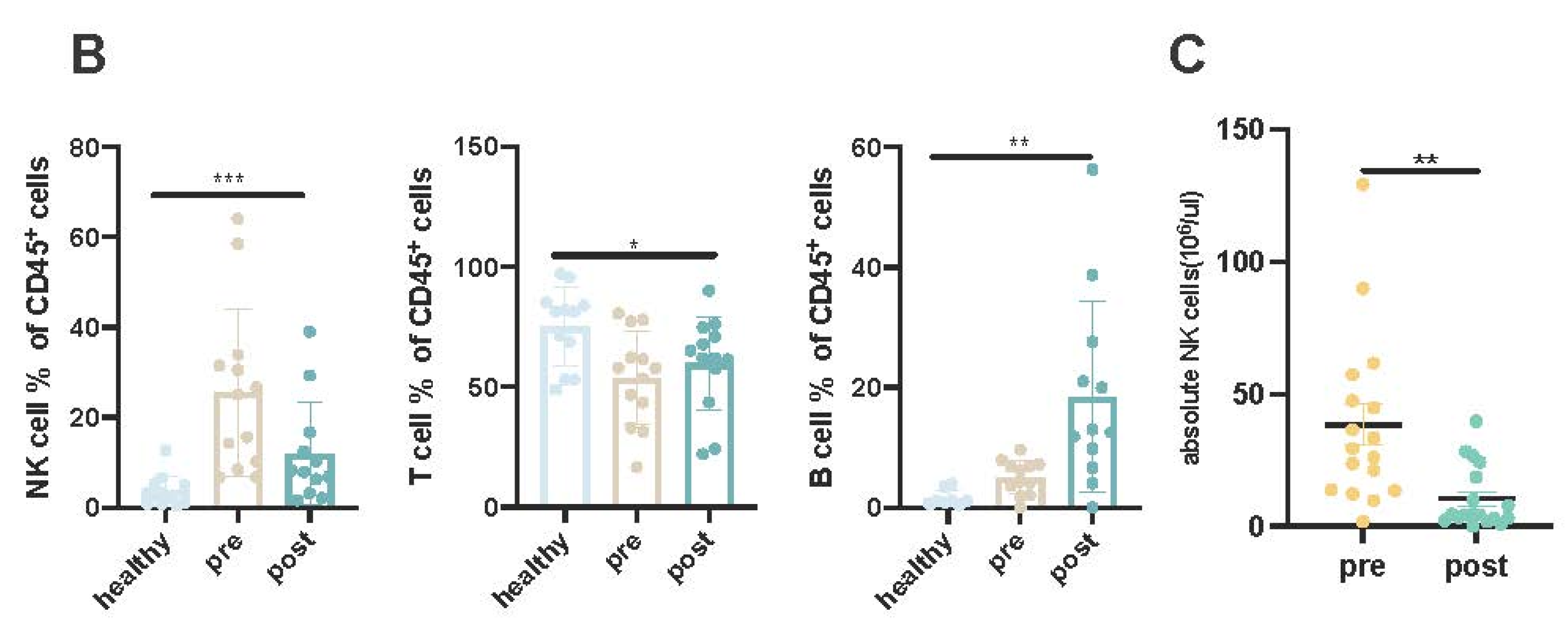
Figure 2.
(A): tSNE distribution of NK cells among the healthy donors and paired RRMM patients. (B):Metacluster of NK cell pops among the groups. (C):Heatmap expression of metaclusters. (D):The percentages of KIR-NKP46+ of NK cells among the healthy donors and paired RRMM patients. (E): Activation receptor expression in NK cells.(left): the percentages of CD226(MFI) expression in NK cells among the group.(middle):the percentages of CD122(MFI) expression in NK cells among the group.(right):the percentages of CD38(MFI) expression in NK cells among the group. (F):Activation receptor expression in NK subpopulations. the percentages of CD122(MFI) expression in NK cells among the group.(middle):the percentages of NKP30(MFI) expression in NK cells among the group.(right):the percentages of CD38(MFI) expression in NK cells among the group.
Figure 2.
(A): tSNE distribution of NK cells among the healthy donors and paired RRMM patients. (B):Metacluster of NK cell pops among the groups. (C):Heatmap expression of metaclusters. (D):The percentages of KIR-NKP46+ of NK cells among the healthy donors and paired RRMM patients. (E): Activation receptor expression in NK cells.(left): the percentages of CD226(MFI) expression in NK cells among the group.(middle):the percentages of CD122(MFI) expression in NK cells among the group.(right):the percentages of CD38(MFI) expression in NK cells among the group. (F):Activation receptor expression in NK subpopulations. the percentages of CD122(MFI) expression in NK cells among the group.(middle):the percentages of NKP30(MFI) expression in NK cells among the group.(right):the percentages of CD38(MFI) expression in NK cells among the group.
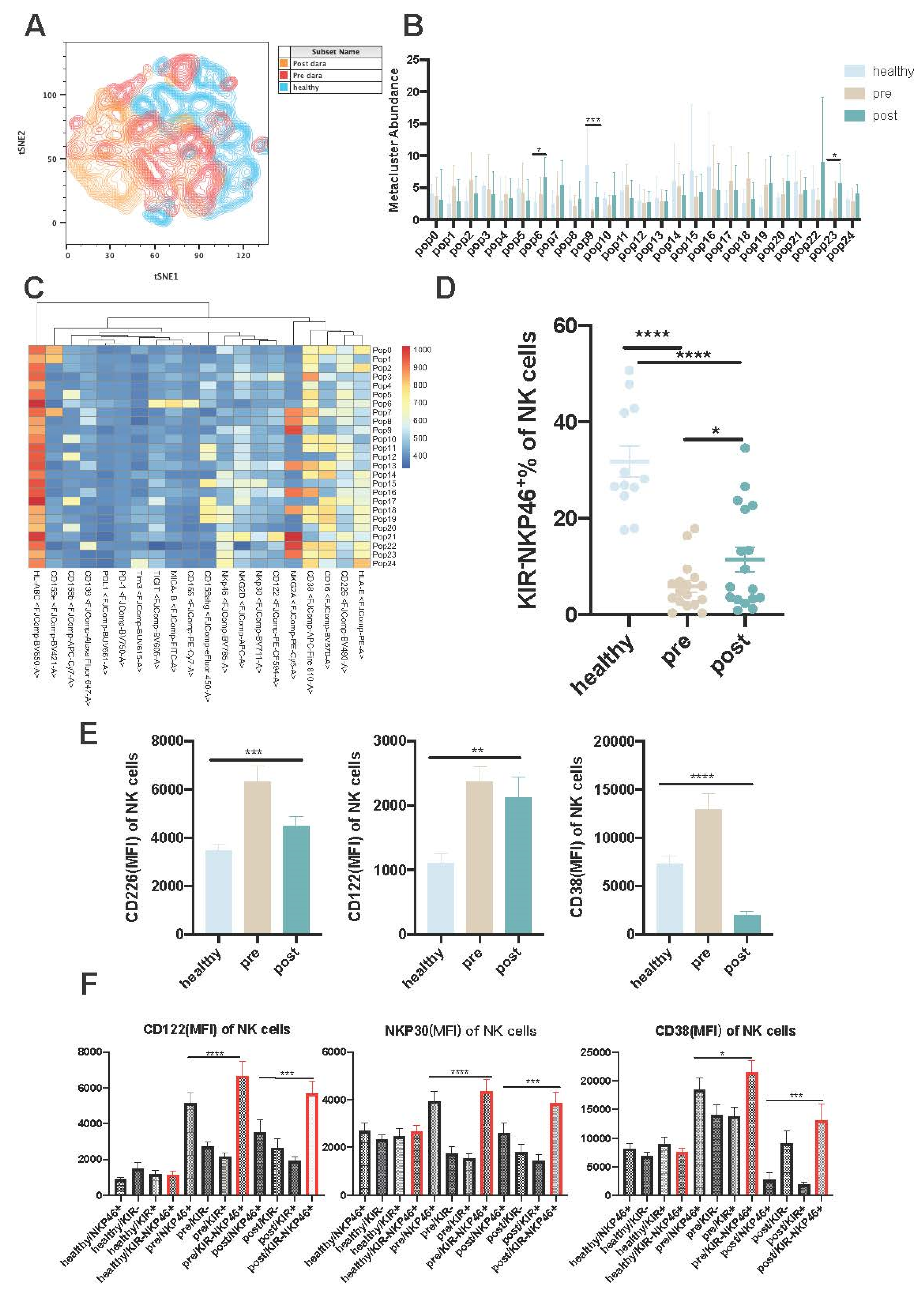
Figure 3.
(A): The percentages of CD107a of NK subsets from PB in healthy donors and RRMM patients with or without dara treatment when cocultured with U266 or LP-1 cells. (B): The percentages of CD107a of NK subsets from BM in RRMM patients with or without dara treatment when cocultured with U266 or LP-1 cells. (C):The percentages of CD107a and TNF-α in PB NK cells between the healthy donors and RRMM patients after dara stimulation in vitro. (D): The proliferation ability of NK subsets with or without dara treatment.(n=5). (E):The early and late apoptosis level among NK subsets with or without dara treatment(n=3). (E):The percentages of 7ADD+Annexinv+ in KIR+ and KIR- NK cells with or without Dara treatment(n=6). (F):The percentages of 7ADD+Annexinv+ in target cells when cocultured with KIR+ or KIR-NK cells with or without Dara treatment(n=6).
Figure 3.
(A): The percentages of CD107a of NK subsets from PB in healthy donors and RRMM patients with or without dara treatment when cocultured with U266 or LP-1 cells. (B): The percentages of CD107a of NK subsets from BM in RRMM patients with or without dara treatment when cocultured with U266 or LP-1 cells. (C):The percentages of CD107a and TNF-α in PB NK cells between the healthy donors and RRMM patients after dara stimulation in vitro. (D): The proliferation ability of NK subsets with or without dara treatment.(n=5). (E):The early and late apoptosis level among NK subsets with or without dara treatment(n=3). (E):The percentages of 7ADD+Annexinv+ in KIR+ and KIR- NK cells with or without Dara treatment(n=6). (F):The percentages of 7ADD+Annexinv+ in target cells when cocultured with KIR+ or KIR-NK cells with or without Dara treatment(n=6).
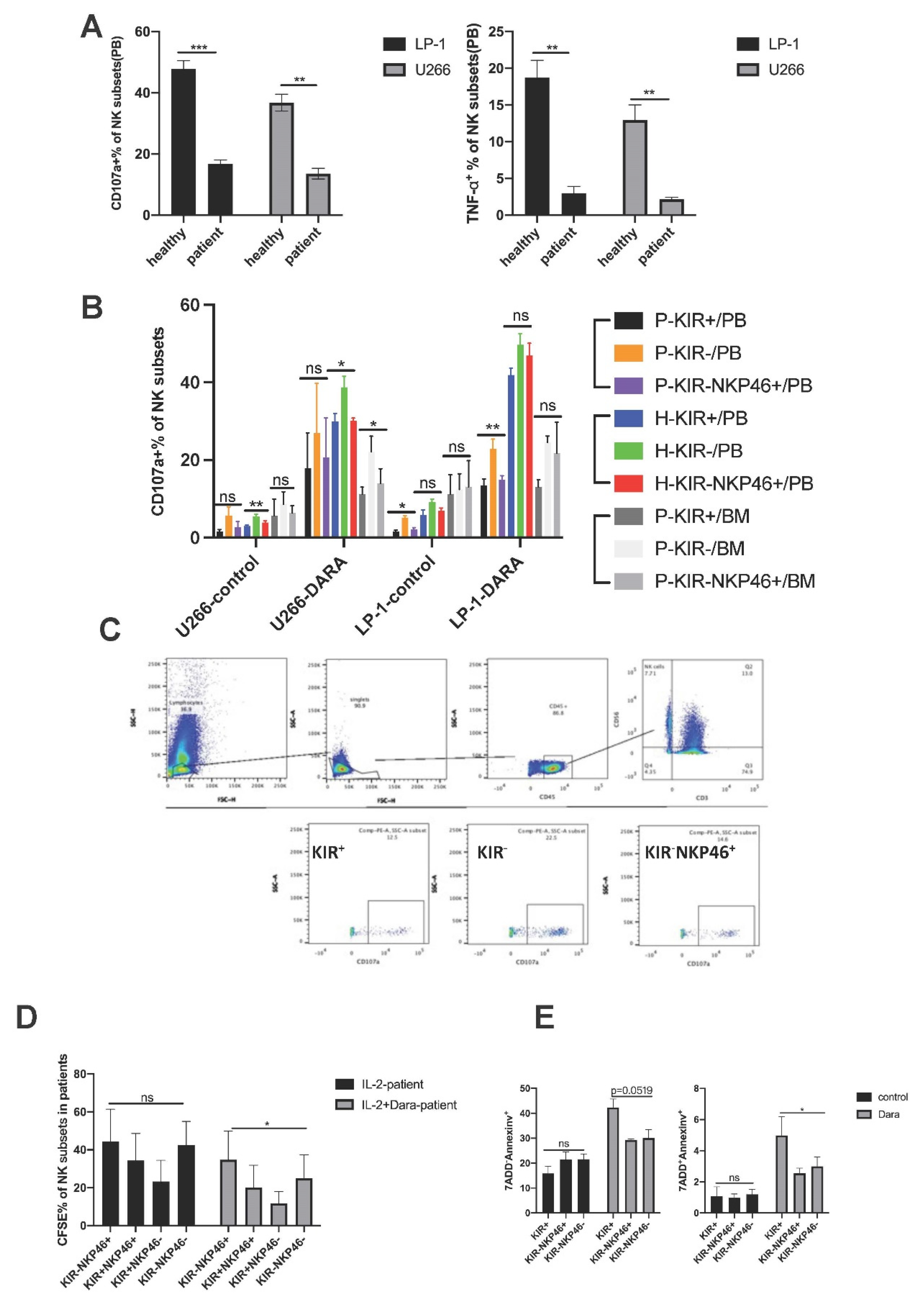
Figure 4.
(A): ROC curve of KIR-NKP46+ NK subsets from post dara-treatment RRMM patients.
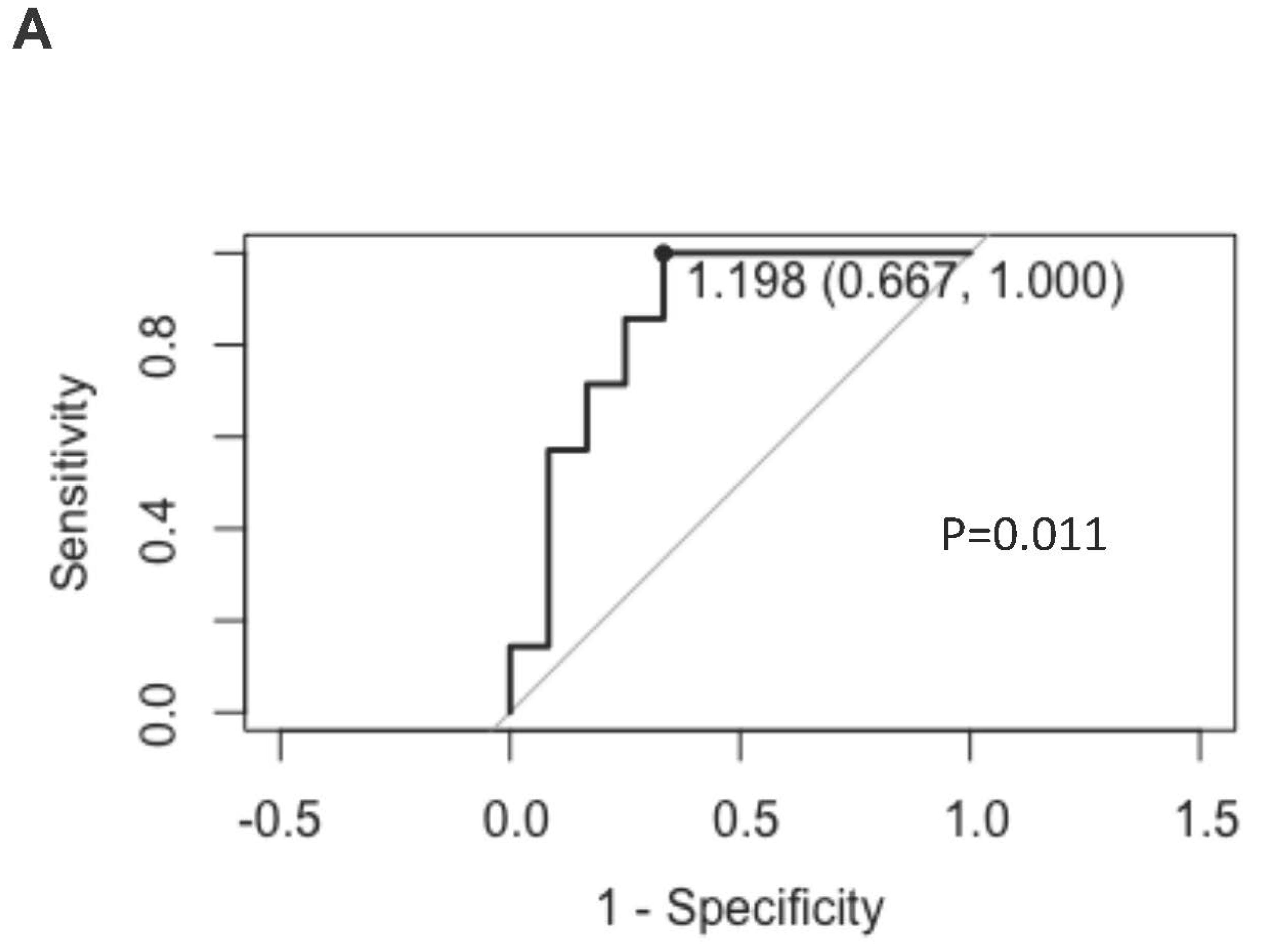

Table 1.
The clinical characteristics of paired RRMM patients.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



