
Submitted:
24 May 2024
Posted:
24 May 2024
You are already at the latest version
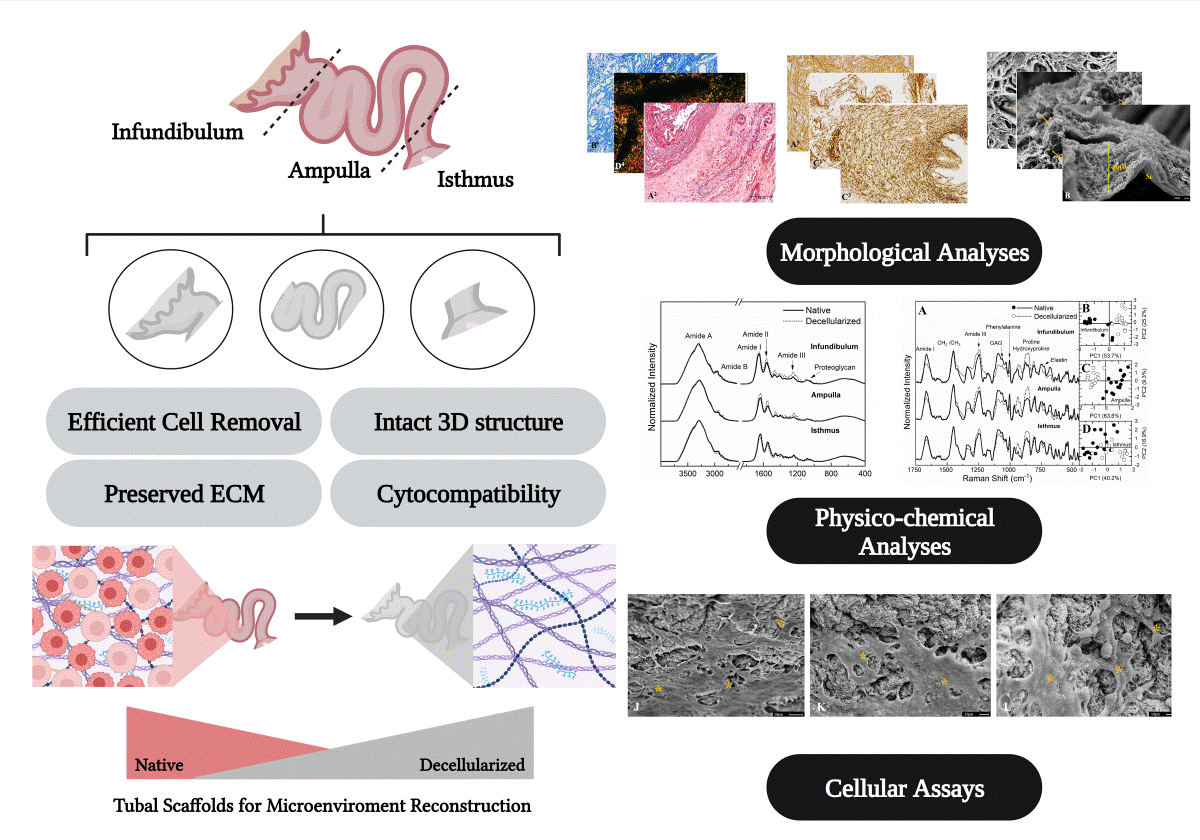
Abstract
Uterine tube extracellular matrix is a key component that regulates tubal tissue physiology, and it has a region-specific structural distribution, which are directly associated to their functions. Considering this, application of biological matrices in culture systems is an interesting strategy to develop biomimetic tubal microenvironments and enhance their complexity. However, there are no established protocols to produce tubal biological matrices that consider the organ morphophysiology for such applications. Therefore, this study aimed to establish region-specific protocols to obtain decellularized scaffolds derived from porcine infundibulum, ampulla, and isthmus to provide suitable sources of biomaterials for tissue-engineering approaches. Porcine uterine tubes were decellularized in solutions of 0.1% SDS and 0.5% Triton X-100. Decellularization efficiency was evaluated by DAPI staining and DNA quantification. We analyzed ECM composition and structure by optical and scanning electronic microscopy, FTIR and Raman spectroscopy. DNA and DAPI assays validated the decellularization, presenting significative reduction of cellular content. Structural and spectroscopy analyses revealed that produced scaffolds remained well structured and with ECM composition preserved. YS and HEK293 cells were used to attest cytocompatibility, allowing cell survival and successful interaction with the scaffolds. These results suggest that such matrices are applicable for future biotechnological approaches in the reproductive field.
Keywords:
uterine tube
; extracellular matrix
; reproduction
; decellularization
; biomaterial
1. Introduction
The female reproductive tract microenvironment plays a direct and essential influence on embryonic morphogenesis, from the oocyte release in the ovarian follicle to the conceptus implantation in the endometrium, modulating cellular processes for the formation of a new individual [1]. Although the endometrium is considered the main tissue for the proper establishment of maternal-embryo interaction, promoting communication between the maternal blood system and the placenta, the uterine tube is actually the first microenvironment that the newly fertilized embryo interacts with [2]. During this process, the tubal epithelium inhibits polyspermy, and assists on spermatic capacitation and flagellar movement, through ciliary motility and the secretion of a fluid enriched of nutrients and signaling molecules [3]. On the other hand, the tubal microenvironment specifically in the ampulla region acts on the genome activation of embryonic cells and influences on the quality of inner mass cells, and such interaction contributes to embryonic survival [4].
Despite the importance of uterine tubes in natural conception in mammals, this structure became non-essential for the in vitro fertilization (IVF) techniques process, once the embryos can be produced in vitro and transferred to the maternal endometrium after a short period in culture, even for human samples[5,6]. Nevertheless, even with the advances in in vitro assisted reproduction techniques, live birth rates after embryo transfer are lower than expected, leading to a high percentage of gestational loss and an increase in the expenses of IVF [7]. According to the Society for Reproductive Technology (SART), the successful birth rate in women under 35 who have undergone IVF in the United States is only 55.6% [8] and the percentage decreases in more advanced ages, as 38-40 years old women [8]. In other species which IVF is practiced because of economic interest, as bovines, the success rate of embryos transferred after IVF is only 38%, which is much lower compared to the natural mating process [9].
In this context, studies have focused on elucidating the molecular mechanisms presented in the female reproductive tract and their influence on embryo development and in the implantational process [10,11]. Tubal microenvironment has gained more protagonism in pregnancy success, once the interaction between the embryo and the tubal epithelium establish the basis for embryonic development [12]. Indeed, the molecular communication that ciliated epithelial cells play on germ cells and in the newly formed embryo is able to modulate phenotypic characteristics and induce important modifications in cellular processes, contributing to fertilization and, consequently, implantation success [13].
The extracellular matrix (ECM) of the tubes provides not only structural support to the cells, but also is able to self-modulate due to cyclical hormonal stimulation in the female reproductive tract [14,15]. The tubal ECM molecular signaling is able to stimulate cell proliferation in the mucosa through mechanotransduction, as well as their differentiation in the several specialized cell types along the tubal epithelium and stroma [16]. Mechanic forces that the tubal wall plays on the embryo, as shear stress and compression forces, also modulate cell fate and contribute to embryonic development and survival [17,18]. This complex microenvironment demonstrates that a better understanding of its cellular and molecular constituents may provide important data for the development of new approaches for early infertility diagnosis, as well as the improvement of in vitro reproduction techniques to increase well-succeeding pregnancy rates [19,20].
The study of early development events is very difficult in humans, once the management of human samples in the pre-implantational phase is limited for sample collection due to be a very short and dynamic phase , and also there are several technical and ethical parameters involved in the studies with of human embryos [21]. Thus, it has been proposed the production of in vitro models that recapitulate the tubal morphophisiology to study the interaction between the embryo and the components that structure the tubal wall under controlled parameters [21,22]. A number of artificial 2D and 3D methods were developed to mimic the ciliated secretory epithelium, as transwell systems and tubal organoid; however, none of them were able to recapitulate the uterine tube histoarchitecture [21,23]. Cell-cell and cell-ECM contact is essential to structure a functionally appropriated biomimetic model that allow co-culture with embryos or embryonic organoids [23].
Although these models aim to rebuild portions of the tubal wall, cell-cell and cell-environment interactions are essential to mimic in vivo conditions [24,25]. To develop more complex models, it was proposed to use natural and synthetic biomaterials that may provide a functional three-dimensional structure that allows cellular adhesion and maintenance to reconstitute the tissue microarchitecture [14,15]. Several natural biomaterials as collagen, alginate, Matrigel®, and synthetic ones as polyethylene glycol (PEG) and polycaprolactone have a number of mechanical, physical-chemical and biological properties for application in biomimetic tissues reconstruction; however none of them is able to mimic efficiently and precisely the ECM properties [26]. Tubal ECM bioactive peptides present specific characteristics and properties that are singular to the uterine tube microenvironment [16], thus, obtaining biological matrices by decellularization is an innovative and accessible approach that allows the preservation of ECM composition and three-dimensionality [27,28]. Recent studies demonstrated that biological matrices are able to support cells from several biological sources and species, revealing their xenogeneic compatibility [29,30]. The application of biological scaffolds derived from decellularized tubal matrices has great biotechnological potential, not only for tubal microenvironment reconstruction, but also as a substrate to stimulate oocyte maturation and in vitro embryo development from different species [27,28].
Therefore, this study aimed to establish region-specific decellularization protocols to produce biological acellular extracellular matrices from porcine infundibulum, ampulla and isthmus to access the preservation of main ECM components for the development of high-quality porcine tubal matrices. Structural and physico-chemical analyses were performed to evaluate the efficiency of each presented protocol as well as the cytocompatibility of the scaffolds was also verified.
2. Materials and Methods
2.1. Samples Acquisition
Uterine tubes (porcine) (n = 10 per group) from pre-pubertal sows at approximately 6 months of age were obtained from the slaughterhouse of the Faculty of Animal Science and Food Engineering of the University of São Paulo, Pirassununga campus, Brazil. Immediately after the collection, the samples were stored on ice and transported to the laboratory. The methodology is summarized in Figure 1.
2.2. Uterine Tubes Segments Decellularization
The uterine tubes were dissected and divided into infundibulum, ampulla and isthmus. Each fragment had 1cm size, which were submitted to the decellularization process. The decellularization process took place in three steps varying according to the segment: for infundibulum and ampulla (1) initial washing - 2h of dH2O and 2h of sodium phosphate buffer (PBS) at 100rpm of agitation; (2) decellularization - sodium dodecyl phosphate (SDS) 0.1% for 10h and Triton X-100 0.5% for 2h. For the isthmus, no differences in step 1 and step 2 had longer exposure to SDS 0.1%, for 24h. Samples were stored in PBS 1x at 4 °C for further analysis.
2.3. 4,6-Diamidino-2-Fenilindole (DAPI) Staining
DAPI staining was used to verify the presence of nuclei in the samples after the decellularization. Native and decellularized samples were frozen in Tissue Plus O.C.T. (Fisher Health Care, Houston, TX, USA), then microsectioned using a cryostat (CM1860 model, Leica Biosystems, Baden-Wurttemberg, Germany). The slides were stained with DAPI solution (1:10,000) at room temperature without light for 10 min, and then washed with PBS 1x for analysis using fluorescent microscopy (Nikon ECLIPSE 80I, CADI FMVZ-USP, Tokyo, Japan).
2.4. Genomic DNA Quantification
For genomic DNA quantification, the BioGene® K204-4 (BioGene, Recife, Brazil) was used to extract the DNA from native and decellularized samples according to the manufacturer’s specifications. The fragments were digested at 56 °C by the action of Proteinase K. The fragments were filtered and analyzed by spectrophotometry at 260 nm (Nanodrop, Thermo Scientific, Waltham, MA, USA).
2.5. Histological Analysis
Histological evaluation was conducted to assess the decellularization efficiency and to verify the morphological integrity of ECM components. Native and decellularized tubal fragments were fixed in 4% buffered paraformaldehyde (PFA) for 48 h, dehydrated using sequential concentrations of alcohol (70, 80, 90, and 100%), diaphanized in xylol, and embedded in paraffin. Microsections of 5µm (n° RM2265; Leica) were stained with Hematoxylin & Eosin (HE) to verify nuclei presence and the general tissue morphology; Masson’s trichrome for collagen content evaluation; Picrosirius Red to evaluate the pattern of collagen distribution of different thicknesses; Alcian blue (pH = 2.5) to evaluate GAGs general content and Weigert’s fuchsin-resorcin to detect elastic fibers presence. Slides were photographed and analyzed using a light microscope (Nikon ECLIPSE 80I, CADI FMVZ-USP). To perform measurements, the relative occupied area of these ECM elements were quantified in five semiseries slices per animal, totaling 10 fields at the magnification of 20 times with the software ImageJ version 8.0.
2.6. Immunohistochemistry Analysis
At first, samples microsections were rehydrated in citrate buffer in the microwave for antigen retrieval for 1 minute. Then, the endogenous peroxidase blockage was performed with 3% hydrogen peroxide in distilled water for 40 min in the dark. After that, for the non-specific protein interaction blockage, 2% bovine serum albumin (BSA) in PBS was used. The primary antibodies chosen were anti-collagen I (#PA5-29569, 1:250, Invitrogen, Carlsbad, CA, USA), anti-collagen III (#PA1-28870, 1:250; Invitrogen), anti-fibronectin (#Ab2413, 1:100, Abcam, Cambridge, UK), anti-laminin subunit α2 (#PA1-16730, 1:200; Invitrogen), anti-elastin (#Ab9519, 1:100, Abcam), and the secondary antibodies were IgG anti-mouse/anti-rabbit (#K800; Dako, CA, United States). The incubation took place overnight at 4 °C. The reaction was detected by Dako Advance HRP (#K6068; Dako) followed by DAB (#k3468; Dako) revelation, according to the manufacturer’s instructions. Slides were photographed and analyzed using a light microscope (Nikon ECLIPSE 80I, CADI FMVZ-USP). To perform a measurement, the relative occupied area of these main ECM components was quantified in five semiseries slices per animal, totaling 10 fields at the magnification of 20 times with the software ImageJ version 8.0.
2.7. Scanning Electronic Microscopy (SEM)
Samples were fixed in Karnovsky solution (2.5% glutaraldehyde and 4% paraformaldehyde in a buffered 0.1 M sodium cacodylate) for 48 h and dehydrated in alcohol concentrations for 5 min each. Then, the fragments were dried in a supercritical point device (LEICA EM CPD 300®) and coated with gold (EMITECH K550®, Quorum Technologies, United Kingdom). Lastly, the samples were photographed under a scanning electron microscope (LEO 435 VP®, Oberkochen, Germany).
2.8. Fourier Transform Infrared Spectroscopy (FTIR) Analysis
To evaluate the ECM molecular composition of native and decellularized samples and verify if the main proteins remained preserved, the Fourier transform infrared spectroscopy (FTIR) was applied. The assay was conducted in a Bruker Vertex 70v FTIR spectrometer (Bruker Optik GmbH, Ettilingen, Germany) with an attenuated total reflectance (ATR) accessory located at the Department of Physics from the State University of Maringá, Paraná, Brazil. The spectrum of each uterine tube segment (n=5), between 4000 and 400 cm−1, is an average of three measurements with 128 scans and 4cm−1 of spectral resolution. All measurements were performed at room temperature, and the spectra were vector normalized using OPUS software 8.5.
2.9. Raman Spectroscopy Analysis
To complement the FTIR analysis, Raman spectroscopy was also applied to evaluate physical-chemical alterations in native and decellularized tissues. Raman spectra were obtained using a Senterra Confocal Raman microscope (Bruker Optik GmbH, Ettilingen, Germany) equipped with an objective lens (20× magnification) focusing the excitation laser (785 nm, 100 mW) on the sample. The spectrum of each tubal segment, from 1750 to 400 cm−1, is an average of three measurements with 3 scans, 9-15 cm−1 of spectral resolution, and 20 s of detector integration time. All measurements were performed at room temperature and the spectra were vector normalized using OPUS software 8.5.
2.10. Scaffolds Sterilization
At the end of decellularization, the scaffolds were washed with dH2O for 48h to remove the detergents. For sterilization, inside the laminar flow hood, the scaffolds were fragmented and immersed in progressive and regressive alcohol concentrations (70%, 80%, 90%, 100%, 90%, 80%, and 70%) for 5 min in each one. After that, the segments were washed with PBS 1x with 2% antibiotics (Penicillin-Streptomycin 10.000 μg/mL, LGC Biotecnologia, Cotia, Brazil). Then, they were also exposed to UV light for 5 min and stored for further analysis. To evaluate the scaffolds sterility, they were immersed in DMEM medium (Sigma, St. Louis, MO, USA) without antibiotics and incubated for 72h in the cell culture incubator.
2.11. Cytotoxicity Assay
To evaluate cell viability in the biomaterials, the resazurin assay was conducted. At first, canine Yolk sac-derived (YS) cells, which were isolated, characterized and donated by Professor Ana Claudia Oliveira Carreira from University of São Paulo [31], and HEK293 cells were culture in DMEM medium supplemented with 10% fetal bovine serum (FBS, Gibco, UK) and antibiotics, and incubated at 37°C under 5% CO2 condition. After 18h, the cells were tripsinized and a density of 2x103 cells/mL were seeded on tubal scaffolds in a 24 well plate. Then, 1mL of resazurin solution (0,14mg/mL in PBS; Thermo Fisher, Waltham, MA, USA) were added to each well. The seeded scaffolds were incubated for 1, 3, 7 and 10 days. In each day, 200μL were collected and stored in a 96 well plate in the refrigerator. At the end of the experiment, the collected samples were read in a spectrophotometer (µQuant – Bio-Tek Instruments, INC) at an optical density of 570nm. The same culture conditions were applied for cells cultured without the scaffolds, which were the control groups.
2.12. Cytocompatibility Assay
To verify the cell adhesion on the scaffolds, 1x104 YS and HEK293 cells were seeded on infundibulum, ampulla and isthmus fragments immersed in supplemented D-MEM medium for 7 days at the same culture conditions. Then, the seeded scaffolds were fixed for SEM analysis.
2.13. Statistical Analysis
For quantitative data, Shapiro-Wilk normality test was conducted to confirm data normal distribution. For DNA content data, Mann-Whitney was applied. For morphoquantitative analyses, multiple t-student tests were applied. Data were showed as mean ± SD (standard deviation). For FTIR and Raman experiments, a PCA analysis was conducted, and a one-way ANOVA was performed to compare the means of quantitative data. Statistical significance was considered for p < 0.05. A Tukey post-hoc test was applied to compare native and decellularized samples. The data were analyzed with GraphPad Prism 8.0 (GraphPad Software, Inc., San Diego, CA, USA).
3. Results
3.1. Tubal Segments Decellularization: Protocol Standardization
Considering that each tubal segment has a distinct structural composition, varying in thickness, more than one decellularization protocol was applied to allow the efficient cellular removal and maintenance of the structural scaffold, as well as the ECM composition of each segment. Several analyses were performed for each segment separately to verify their properties as a biological scaffold. Macroscopically, it was possible to observe a whitening in the infundibulum, ampulla and isthmus compared to the native tissue with little alteration in the macroscopic structure, respectively (Figure 2A, Figure 2C and Figure 2D). At first, it was proposed a unique protocol to decellularize the uterine tube as a whole; however, preliminary histological analyses showed that the exposure period to decellularize the infundibulum and the ampulla was insufficient to remove isthmus cellular content, which led to apply a longer exposure in the decellularizing solution. By DAPI staining analysis, it was verified that, when compared to the native tissue, the 10h SDS-based protocol for the infundibulum (Figure 2E and Figure 2F) and for ampulla (Figure 2G and Figure 2H) was able to promote cellular removal efficiently. Regarding the isthmus, the same analysis showed that the 24h SDS-based protocol was more efficient to decellularization (Figure 2I and Figure 2J), since it presents a higher muscle layer thickness (data not shown). Corroborating these findings, the total genomic DNA analysis showed a reduction in the DNA content of 97.6% in the infundibulum, 97.8% in the ampulla and 94.7% in the isthmus compared to the native tissue, which demonstrated the decellularization efficiency (Figure 2K).
3.2. Morphological and Ultrastructural Characterization
The ECM components preservation after the decellularization process is vital in the development of biological scaffolds, once these elements structure the tissue microenvironment [32]. For this purpose, histological (Figure 3); immunohistochemical (Figure 4) and ultrastructural (Figure 5) analysis of the three decellularized tubal segments were conducted to perform a morphological and three-dimensional characterization of the generated biomaterials. Through Hematoxylin and Eosin (H&E) staining, it was possible to observe the tissue general structure, as well as the presence or absence of nuclei. In the native infundibulum (INF), there were evident epithelial invaginations toward to the lumen, as well as several blood vessels, glandular structures and a thin muscle layer (Figure 3A). In the decellularized infundibulum, it is possible to visualize nuclei absence, maintenance of some invaginations and rounded structures, corresponding to blood vessels and glands (Figure 3A1). Regarding the ampulla, there is a reduced number of glands, a more prominent lumen with fewer invaginations and a more intense presence of connective tissue (Figure 3A2). The decellularized ampulla (dAMP) also presented nuclei absence, maintained the vascular and glandular structures preserved and it is possible to distinguish the regions with prominent connective tissue (Figure 3A3). In the isthmus (IST), the uterine tube muscular portion, there is a more reduced quantity of glands with a more stained connective portion, demonstrating a more dense tissue (Figure 3A4). In the decellularized isthmus (dIST), it is also noteworthy the nuclei absence and a more structured and dense extracellular matrix (Figure 3A5).
Total collagen content was evaluated using Masson’s trichrome staining, which highlighted the collagen fibers in dark blue (Figure 3B-3B5). Morphologically, decellularized samples did not show any extracellular matrix degradation or abrupt disarrangement, being more evident in the dIST region, due to its highly vascularized characteristic (Figure 3B5). In dINF and dAMP, the morphology of ECM presents minor alterations, which it was expected due to the higher presence of loose connective tissue in these samples. The quantitative analysis of collagen fibers after the decellularization did not reveal any statistically significant difference between the native tissue and the decellularized scaffolds, demonstrating that there was no collagen degradation (Figure 3G). The means of the relative occupied area are summarized in Table S1, Supplementary Information.
Additionally, Picrosirius Red staining was used to highlight the collagen content distribution (Figure 3C-3C5); under polarized light, thick collagen fibers present red and yellow color while thin collagen fibers are green (Figure 3D-3D5). Morphologically, there is a higher proportion of thin collagen fibers compared to the thick ones in the INF; in the AMP, this proportion is more balanced although the thin fibers are still more abundant , while in the IST it is opposite, since the percentage of thick fibers is higher than thin fibers. This morphological pattern remains similar in the decellularized samples of the three segments; indeed, quantitative data corroborated the morphological findings, demonstrating no statistical difference for both thick and thin collagen fibers distribution between native and decellularized samples (Figure 3H and 3I).
The main non-fibrillary molecular group presented in the ECM is general glycosaminoglycan (GAGs) content, thus we evaluated by the Alcian Blue staining. Alcian Blue highlights the GAG content in light blue in a diffuse pattern through the tissue (Figure 3E-3E5); morphologically, GAGS had a similar distribution pattern in the three tubal segments both in native and decellularized samples. It is noteworthy that the decellularization process caused a mild degradation in the GAG content, since those molecules are more sensitive to the effects of detergents. However, the quantitative analysis did not evidence any statistic difference between the groups, which demonstrated that even with a small loss; the majority of GAG content remained in the scaffolds after the decellularization process (Figure 3J).
The presence of elastic fibers was detected by Weigert’s fuchsin-resorcin staining, which stains the elastic components in intense dark purple (Figure 3F-3F5). In tubal samples, elastic fibers were more concentrated in blood vessels, mainly in elastic arteries and veins, with some fibers diffused through the non-modeled connective tissue. In the absence of cells, the elastic fibers became more evident, mainly in dAMP (Figure 3F3) and dIST (Figure 3F5). Quantitative analysis of the relative occupied area demonstrated an increase of elastic fibers in dAMP and dIST samples compared to the native tissues (Figure 3K). This increase may be related to the absence of cells on the samples, which allowed the fibers to be more evident in the staining, however, there was no relevant statistic difference between INF and dINF samples.
After a histological evaluation of tubal ECM, immunohistochemical analyses were performed to detect and quantify the main matrix proteins (Figure 4A-4E5). Type I and III collagens, which are main tubal structural components, presented a similar expression pattern between native and decellularized samples, with similar values in quantification (Figure 4F and Figure 4G). Elastin, the main elastic fibers component, presented an increase in dAMP and dIST compared to native tissues, but no statistic difference was observed between INF and dINF samples (Figure 4H), as previously observed in histological staining. Regarding the adhesive glycoproteins, such as fibronectin and laminin, there was no statistical difference between native and decellularized groups (Figure 4I and Figure 4J). These data confirmed the histological analyses and demonstrated that the produced scaffolds maintained important structural proteins for tissue microarchitecture preservation, as well as adhesive glycoproteins important for cell-ECM interaction. The means of the relative occupied area are summarized in Table S2, Supplementary Information.
To evaluate the three-dimensionality of the generated scaffolds, an ultrastructural analysis by scanning electronic microscopy was performed for native and decellularized tubal segments (Figure 5). It is noteworthy that the decellularization maintained tubal wall stratification of dINF-derived scaffolds, with organized fibers and absence of cells (Figure 5B). In the native infundibulum, there is a dense ciliated secretory epithelium with several epithelial invaginations (Figure 5A and Figure 5C), while in the decellularized biomaterial, the mucosa portion is completely acellular. At the highest magnification, it was observed in the dINF collagen bundles are organized in a stratified and interspersed manner, with no signs of degradation (Figure 5D). In addition, in the native tissue, there is an intense presence of fimbriae and mucus accumulations (Figure 5E e Figure 5G), which are absent after the decellularization. It is also observed in dINF the maintenance of thick and thin collagen fibers refinement, which remained complexly arranged, demonstrating that delicate structures were preserved after the decellularization process (Figure 5F e Figure 5H).
As expected, dAMP-derived scaffolds preserved the three tubal wall layers maintenance along with the absence of cells (Figure 5J). In native ampulla, there is the presence of epithelial invagination and mucus (Figure 5I, 5K and 5M), which is absent in the acellular scaffolds. At higher magnification, dense and structured collagen bundles can be observed, demonstrating that the tissue microstructure remained intact in dAMP (Figure 5N). In addition, ECM layers superimposed in a stratified pattern and remained well structured and more distinct in the absence of cells. Similarly to the infundibulum, the fimbria presented in the ampulla epithelium and mucus deposits were abundant in native tissue (Figure 5O), and absent in dAMP. Thick and thin collagen fibers were also preserved in acellular scaffolds, with a greater proportion of thick fibers, corroborating the histological findings (Figure 5P).
The dIST-derived scaffolds presented a more compact and fibrous structure, with the decellularized mucosa and muscular layers more prominent than in the other segments (Figure 5R). In addition, the tissue stratification is well preserved compared to the native tissue. While the native stroma is denser due to the cell anchorage in the ECM components (Figure 5S), in the decellularized material the collagen layers and bundles are more visible, which allowed the observation of a preserved structure (Figure 5T). In native IST, there was an intense presence of blood vessels surrounded by a dense non-shaped connective tissue (Figure 5Q and Figure 5U). In the dIST, due to the absence of cells, it is possible to observe round structural scaffolds for the blood vessels (Figure 5V). At higher magnification, in the IST, the collagen bundles are more interspaced due to the cell anchoring (Figure 5W). In the dIST, the collagen bundles are thicker and more agglomerated, which makes the scaffold aspect more fibrous (Figure 5X).
3.3. ECM Physic-Chemical Composition Characterization
The spectroscopy emerged as an valuable tool to evaluate biological samples in the bioengineering field, vastly contributing to biomaterials characterization [33]. Vibrational spectroscopy techniques are relatively simple, reproducible, non-damaging and able to provide valuable information at a molecular level. These techniques also allow the investigation of functional groups, types of chemical bonds and molecular conformation, leading to direct information about biochemical composition [34,35]. In this study, Fourier Transform Infrared (FTIR-ATR) and Raman spectroscopy were applied to evaluate the physic-chemical alterations in native and decellularized tubal segments.
FTIR spectra (Figure 6A) were previously characterized by [33]. Among the bands, the most noticeable are associated with the amides, which are directly correlated to the collagen spectrum. Amide I can be related to protein secondary structure. Amide II is associated with the proteins hydration, but also can indicate collagen self-assembly [36,37]. The ratio between the amide III spectrum and the deformation of CH2 at 1450 cm-1 can be related to the collagen triple helical structure [37]. At last, the band at 1080 cm-1 is associated with the proteoglycan content, which are composed by sulfated and non-sulfated glycosaminoglycans (GAGs) covalently bonded to core proteins [37,38]. Based on the spectra results, to evaluate the level of preservation of ECM molecules content and provide quantitative data after the decellularization process, an integration of band areas of Amides I and II, proteoglycans and the ratio between amide III and 1450 cm-1 was performed (Figure 6B–E). There was a statistically significant increase of 22.67% of Amide I in dAMP compared to the native tissue (Figure 6B). On the other hand, in dAMP and dIST, there was an increase of 26.28% and 24.30% respectively in the collagen triple helical structure-related band (Figure 6E). No statistically significant alterations for Amide II and proteoglycans content were detected after the decellularization compared to native (Figure 6C,D).
For Raman spectroscopy (Figure 7A), the Amide I band has its center at 1665 cm-1, while amide III is at 1247 cm-1 [38,39,40,41,42,43]. GAG molecules are characteristic at 1063 cm-1 and the vibratory mode associated with the phenylalanine aromatic ring is at 1003 cm-1 [40,41,43,44,45,46]. The region between 856 and 875 cm-1 can be associated directly to proline and hydroxyproline, which are the major aminoacids that form the collagen fibers [39,42,43,44,45,46]. Finally, elastin is characterized at 725 cm-1 [39,44].
PCA statistical method was applied to evaluate the spectral differentiation of each region independently. Figure 7B, Figure 7C and Figure 7D present the score graphic for native and decellularized infundibulum, ampulla and isthmus, respectively. Each PC1 score graphic has a variation of 53.7%, 63.6% and 40.2% between the regions’ datasets. This result indicates that there is a sample separation tendency, which means that this differentiation does not indicate a full separation between the spectra, and may present similar band contributions between both samples despite the existence of a known spectral difference between native and decellularized samples.
Similar to FTIR-ATR, Raman spectra had their areas integrated to evaluate the spectral contribution of each functional group. The quantitative data are exposed in Figure 7E to Figure 7J. Amide I, elastin and GAG did not present any statistically significant difference between native and decellularized samples for the three regions. Regarding the Amide III spectrum, only in the ampulla region there was an increase of 19.41% in the decellularized compared to the native group. In the proline/hydroxyproline spectrum, there was an increase of 41.13% in the dAMP and 20.51% in the dIST compared to the native samples. Finally, the Raman spectroscopy detected a decrease in the phenylalanine spectra in all regions. Together, the data demonstrate no evident statistical differences, even presenting a slight divergent tendency compared to the FTIR-ATR, predominantly for Amide I, demonstrating a corroboration of the data provided by both techniques. Therefore, spectroscopically, it is possible to assume that the major ECM components of the decellularized components were preserved. These data corroborated the structural and ultrastructural findings regarding the maintenance of scaffolds ECM integrity after the removal of cells.
3.4. Cytocompatibility Evaluation
The ability to allow cell culture is one of the most important properties for a biomaterial, once the interaction between the scaffold and cells is the main characteristic for the proper microenvironment establishment [47]. To evaluate the scaffolds cytocompatibility, two assays were performed: the resazurin assay, to gather cell viability, and a scaffold-cell adhesion assay. We also performed a viability assay using immortalized HEK293 epithelial cells derived from human embryonic kidney, and canine Yolk sac-derived (YS) cells, which are endothelial precursors [31]. These cells were chosen to demonstrate the non-cytotoxic potential of the generated scaffolds and to evaluate the scaffolds capacity to support non-tubal and non-porcine cells. Through SEM analysis, it was possible to observe that after 7 days of culture on the scaffolds both cell types were able to interact with the biological matrices, allowing cell anchorage (Figure 8A-8R). In all three scaffolds, YS cells were able to adhere to the ECM fibers, presenting a fibroblastoid shape. There were cell-cell interactions in specific areas, but they did not form a layer on the scaffolds. On the other hand, HEK cells not only adhered to the scaffolds, but also formed layers as observed in native tissue. This demonstrated that the matrices allowed the epithelial cells to organize and anchor on the ECM, accordingly to the expectations since the adhesive glycoproteins of the scaffold remained preserved (Figure 8A-8R).
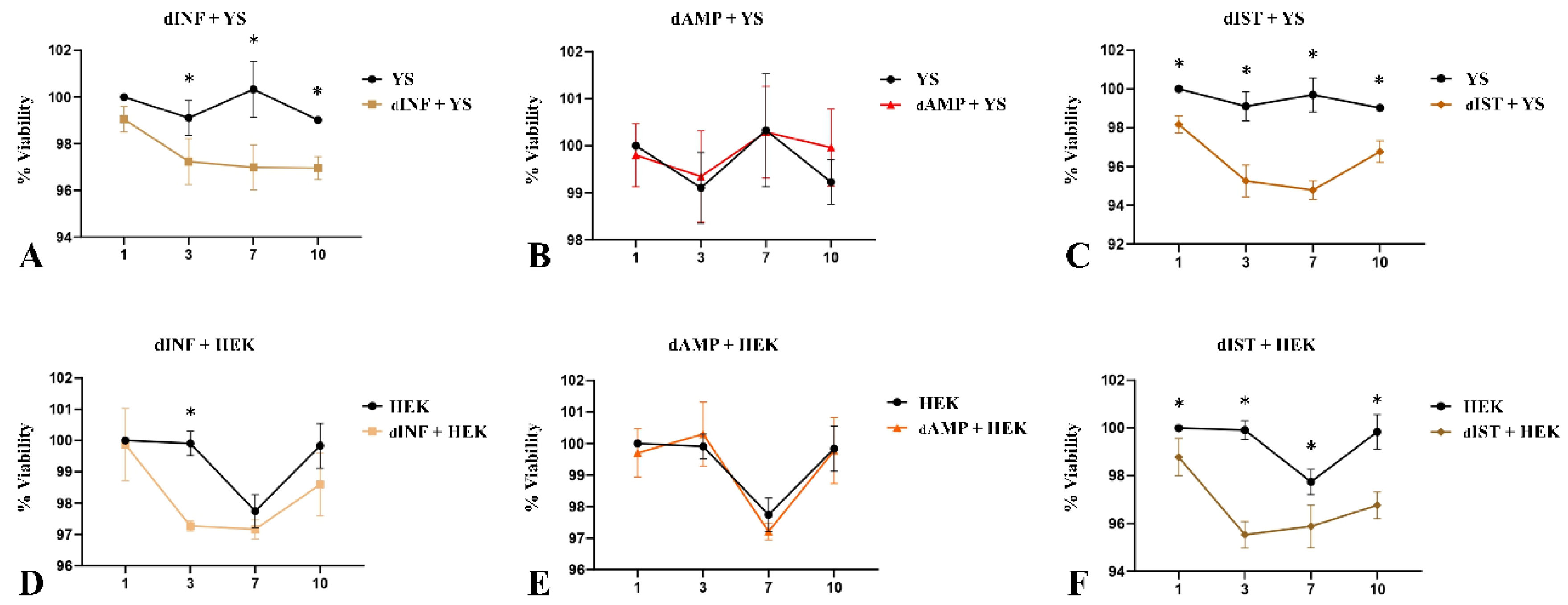
For the cytotoxicity assay, these cells were cultured on scaffolds for 10 days in the presence of resazurin, a purple composite that is reduced by mitochondria becoming a fluorescent pink composite (resofurin) used to check cell viability [48]. The results showed that in all three scaffolds the cell viability was above 95% after1, 3, 7 and 10 days, which verified the scaffolds non-cytotoxicity (Figure 9). In terms of performance, dAMP-derived scaffolds presented the best results, with no statistical difference in any of analyzed periods for both cell types (Figure 9B and Figure 9E). Concerning dINF-derived scaffolds, HEK cells presented a better performance on the scaffolds compared to YS cells (Figure 9A and Figure 9D). A similar pattern was observed for dIST-derived scaffolds (Figure 9C and Figure 9F). In general, the scaffolds allowed satisfactory adherence and survival of epithelial cells from different origins, supporting their broad ability to be used as a platform for in vitro cell culture and tissue reconstruction.
4. Discussion
Tubal microarchitecture is responsible for supporting an appropriated microenvironment for the beginning of embryonic development, providing structure and molecular signaling for both gametes and the newly generated embryo to be nourished by the specialized cells of the ciliated secretory epithelium and ECM[13,49]. The structural and compositional configuration of each tubal segment ECM is related to its function in the establishment of maternal-embryonic communication [50,51]. The proportion of ECM components, as well as the stiffness or malleability of each tubal tissue, is directly correlated to its function. The infundibulum has a more flexible structure that allows a more dynamic movement of the fimbriae [52,53]. On the other hand, the isthmus has a more rigid and less flexible structure, being the region that connects the uterine tube to the uterus [53]. The ampulla has an intermediate structural configuration compared to the other two, allowing a greater interaction between the embryo and the tubal tissue [54]. Due to these particularities, the development of decellularization protocols is delicate, being necessary the standardization of each step to preserve the ECM without compromising the efficiency of cells removal processes.
These matrices may have a great biotechnological potential, since the ECM is able to modulate important intracellular processes through the interaction with transmembrane receptors as integrins, as well as influence on cytoskeleton reorganization during cell migration [55]. Once they may provide molecular substrate for in vitro models production and supplementary condition for embryo culture, the establishment of proper scaffolds and matrices are essential for biotechnology of reproduction [14,15,33]. Despite these indicatives, only few studies previously used decellularized tubal matrices for experimental assays until the day [28,56]. Francés-Herrero et al. [28] used decellularized uterine tubes from hormonally stimulated rabbits to develop a hydrogel that could be applied as a supplement for in vitro embryo culture. Despite of their pioneering and success in stablishing acellular tubal matrices, the study didn’t considered the anatomic distinction of tubal segments, decellularizing the tube as whole, and consequently facing some challenges [28]. This methodology presented some loss of major tubal ECM proteins, which is demonstrated by GAG general content degradation and the decrease in the identification of some proteoglycan and glycoproteins by proteomic analysis. In comparison, our data revealed that there was a preservation of the main ECM components for the three tubal segments once they were decellularized separately accordingly to tissue density. The degradation of rabbit tubal ECM reflected directly on the hydrogel bioactivity, once the loss of some bioactive peptides originally present in tubal microenvironment could compromise the mimicking of in vivo conditions [57,58]. Although the limitations, the use of acellular tube-derived hydrogels was able to induce changes in the metabolic profile of embryos, which revealed that the ECM components interacted with the embryo in its early stages, allowing an adequate in vitro development with a satisfactory metabolic performance [28].
In different study conducted by Venkata et al. [56], tubal and uterine organoids derived from human fetal and adult cells were produced and transplanted in tubal and uterine three-dimensional scaffolds, to develop an in vitro model that allows the study of the molecular mechanisms related with Muller ducts development anomalies. The results demonstrated differences in proteomic composition of the organoids and the fetal ones were able to develop properly with Wnt ligands supplementation in the medium [56]. This study innovates on the development of tubal organoids harvested on biological matrices, providing more complexity to the system; however, the protocol used to obtain the tubal acellular matrices differs from our results.
Once the tissues were exposed to a high concentration of SDS detergent for ten days, the ECM molecular composition were very likely affected and [59,60,61]due to the lack of an acute analysis of scaffolds structural and compositional characterization, it is inviable to compare this model to the one we presented in this study.
Besides the methodological parameters for the generation of preserved acellular scaffolds, the biological material source is an important variable to be considered in the development of technologies for both experimental assays and large scale applications [62,63]. Pre-pubertal porcine uterine tubes were chosen in this study since pig slaughter occurs with high frequency in commercial slaughterhouses, and reproductive tissues are discarded or underutilized, which reinforces the relevance of recycling these biological materials for biotechnological purposes [64,65]. Another interesting factor to be considered is the homogeneity of the samples used to generate the biomaterials, due to the animals reproductive cycle consistency [66,67]. Indeed, hormonal stimulation is able to induce structural and functional alteration on reproductive tissues, which reflects on ECM organization and composition, and thus, in the production of biomaterial [68,69,70]. Consequently, the translational potential of porcine tissue-derived biomaterials is considerable for in vitro reproduction technologies for both humans and other species of economic interest [33].
The maintenance of matrix scaffolds three-dimensional structure is another key factor for the development of more trustworthy in vitro models that mimic a suitable microenvironment for pre-implanted embryos [71,72]. It is noteworthy that 3D culture systems provide adequate conditions for cell culture compared to the traditional 2D ones, allowing a greater cell-cell and cell-environment interaction, which impacts in the cellular performance, gene expression and protein synthesis [73]. Besides the three-dimensionality of tubal microenvironment, , the biomechanical forces that the conceptus is exposed are important for embryo culture, acting directly on cell viability and embryo survival [14,15]. Physiologically inside the uterine tubes and in direct contact with the tubal epithelium, the trophectoderm cells of the conceptus are exposed to mechanical action such as shear stresses and compression, which are able to modulate important intracellular events [74,75,76], and their excessive application may induce cell gene expression alterations in the embryo [74,77]. This aspect of biomaterials production is an interesting challenge for the field.
To compose a complex system, the 3D structure must be cytocompatible to provide the required conditions for the several cell types to repopulate the scaffold and allow a tissue reorganization similar to those found in vivo [78,79]. Although there are several cytocompatible biomaterials, many of them lack adhesive molecules that allow a suitable cell anchorage and a proper interaction between cells and the environment, which is crucial for signaling and structure maintenance [80,81]. Our results demonstrate that the decellularized ampulla-derived matrix (dAMP) presented the best performance for cell culture amongst the obtained biomaterials from the three tubal regions. This result might be related to dAMP intermediate structural pattern, which allowed a more appropriate interaction between cells and the ECM. This finding is interesting for reproducibility due to the availability of material, since the ampulla is the most extensive tubal region, besides being the area which the early embryo interacts for longer periods [82,83,84]. The matrices ability to support non-tubal native cells and from other species is also very important for biotechnological purposes, presenting significant translational characteristics [33]. Altogether, this data proposed decellularization protocols that are efficient to promote a balance between cell removal and ECM preservation for the three uterine tube regions. The produced matrices also presented great cytocompatibility, allowing both cell populations viability in vitro.
5. Conclusions
This study was the first to present a structural and compositional characterization of decellularized scaffolds derived from the three porcine tubal regions. The generated scaffolds remained three-dimensionally structured and with the major ECM components preserved. In addition, it was verified their cytocompatibility, once allowed cell survival and provided the conditions for cell adhesion and integration with the scaffolds. These findings demonstrated that matrices might be used as a valuable tool for future applications as supplement for specific embryo culture mediums or even substrate for 3D bioprintable hydrogels production. Such biotechnologies can be used for regenerative therapies in the female reproductive tract or for tubal organoids production to perform drugs screening test or even to study unknown molecular mechanisms related to infertility in the pre-implantational stages in humans and high-interest commercial species. Given the biological and molecular role of extracellular matrix in the distinct uterine tube portions, these decellularized region-specific matrices may be suitable bioactive compounds for a great number of applications in cell and embryo, which provide unique molecular stimulation, enhancing the system complexity parameters to those found in vivo.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1: Quantitative data of ECM general components of native and decellularized tubal tissues. The data are expressed by the mean ± standard deviation (SD); Table S2: Quantitative data of ECM main components of native and decellularized tubal tissues. The data are expressed by the mean ± standard deviation (SD).
Author Contributions
Conceptualization, G.H.DR.A., R.S.d.S. and A.C.O.C.; methodology, G.H.DR.A., R.S.d.S., M.S.G., V.H.d.S.G., H.d.S. and A.C.O.C.; validation, G.H.D.R.A., L.A.F., M.S.G., A.C.O.C.; formal analysis, G.H.D.R.A., R.S.d.S., M.S.G., V.H.d.S.G., H.d.S., R.P.I., L.A.F., I.C.F., T.N.G.N., C.S.A-F.; investigation, G.H.D.R.A., M.S.G., F.S., M.L.B., L.H., J.d.C.R., F.V.M., A.J.P.F.; resources, G.H.D.R.A., M.L.B., L.H., A.C.O.C.; data curation, G.H.D.R.A.; writing—original draft preparation, G.H.D.R.A., R.P.I., J.d.C.R., L.H.; writing—review and editing, F.S., M.L.B., L.H., J.d.C.R., F.V.M., C.S.A-F., A.J.P.F., A.C.O.C.; visualization, F.S., M.L.B., L.H., J.d.C.R., F.V.M., A.J.P.F., A.C.OC.; supervision, A.C.O.C.; project administration, G.H.D.R.A and A.C.O.C.; funding acquisition, A.C.O.C. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by Coordination for the Improvement of Higher Education Personnel (CAPES), grant n° 726 88887.488264/2020-00. Gustavo Henrique Doná Rodrigues Almeida has received research support from CAPES.
Institutional Review Board Statement
This research was conducted in acquiescence with the Ethics Committee on the Use of Animals regulations of School of Veterinary Medicine and Animal Science of University of São Paulo (CEUAx protocol no. 2250270223).
Data Availability Statement
All data of this study are available in the article. Additional raw data sets generated during the study are too large to be shared publicly but are available upon request from the corresponding author.
Acknowledgments
We thank Professor Maria Angelica Miglino for the provided infrastructure to perform the study. We thank the Advanced Center for Imaging Diagnosis (CADI-FMVZ/USP), Dr. Rose Eli Graci Rici for the support in the SEM images and technicians Ronaldo Agostinho and Diogo Palermo in optical microscopy analyses.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Klohonatz, K.M.; Nulton, L.C.; Hess, A.M.; Bouma, G.J.; Bruemmer, J.E. The role of embryo contact and focal adhesions during maternal recognition of pregnancy. PLoS One 2019, 14. [Google Scholar] [CrossRef] [PubMed]
- Kölle, S.; Hughes, B.; Steele, H. Early embryo-maternal communication in the oviduct: A review. Mol. Reprod. Dev. 2020, 87, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Harris, E.A.; Stephens, K.K.; Winuthayanon, W. Extracellular Vesicles and the Oviduct Function. Int. J. Mol. Sci. 2020, 21, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Csöbönyeiová, M.; Varga, I.; Lapides, L.; Pavlíková, L.; Feitscherová, C.; Klein, M. From a Passive Conduit to Highly Dynamic Organ. What are the Roles of Uterine Tube Epithelium in Reproduction? Physiol. Res. 2022, 71, S11–S20. [Google Scholar] [CrossRef] [PubMed]
- Kleijkers, S.H.M.; Eijssen, L.M.T.; Coonen, E.; Derhaag, J.G.; Mantikou, E.; Jonker, M.J.; Mastenbroek, S.; Repping, S.; Evers, J.L.H.; Dumoulin, J.C.M.; et al. Differences in gene expression profiles between human preimplantation embryos cultured in two different IVF culture media. Hum. Reprod. 2015, 30, 2303–2311. [Google Scholar] [CrossRef] [PubMed]
- Krisher, R.L.; Schlenker, T. Culture of Human Preimplantation Embryos in a Clinical ART Setting. Methods Mol. Biol. 2019, 2006, 355–371. [Google Scholar] [CrossRef] [PubMed]
- Shaulov, T.; Sierra, S.; Sylvestre, C. Recurrent implantation failure in IVF: A Canadian Fertility and Andrology Society Clinical Practice Guideline. Reprod. Biomed. Online 2020, 41, 819–833. [Google Scholar] [CrossRef] [PubMed]
- Romanski, P.A.; Gelvin, B.; Bortoletto, P.; Rosenwaks, Z.; Kang, H.J. Live-Birth Outcomes Among Women With Infertility and Anti-Müllerian Hormone Levels of 0.3 ng/mL or Lower. Obstet. Gynecol. 2022, 140, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Demetrio, D.G.B.; Benedetti, E.; Demetrio, C.G.B.; Fonseca, J.; Oliveira, M.; Magalhaes, A.; dos Santos, R.M. How can we improve embryo production and pregnancy outcomes of Holstein embryos produced in vitro? (12 years of practical results at a California dairy farm). Anim. Reprod. 2020, 17. [Google Scholar] [CrossRef]
- Ranjbaran, A.; Latifi, Z.; Nejabati, H.R.; Abroon, S.; Mihanfar, A.; Sadigh, A.R.; Fattahi, A.; Nouri, M.; Raffel, N. Exosome-based intercellular communication in female reproductive microenvironments. J. Cell. Physiol. 2019, 234, 19212–19222. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Lin, H.; Kong, S.; Wang, S.; Wang, H.; Armant, D.R. Physiological and molecular determinants of embryo implantation. Mol. Aspects Med. 2013, 34, 939–980. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, L.; Guo, N.; Liu, C.; Wang, M.; Zhu, L.; Li, F.; Jin, L.; Sui, C. Oviductal extracellular vesicles from women with endometriosis impair embryo development. Front. Endocrinol. (Lausanne). 2023, 14, 1171778. [Google Scholar] [CrossRef] [PubMed]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef]
- Almeida, G.H.D.R.; Iglesia, R.P.; Rinaldi, J. de C.; Murai, M.K.; Calomeno, C.V.A.Q.; da Silva Junior, L.N.; Horvath-Pereira, B. de O.; Pinho, L.B.M.; Miglino, M.A.; Carreira, A.C.O. Current Trends on Bioengineering Approaches for Ovarian Microenvironment Reconstruction. Tissue Eng. Part B. Rev. 2022. [CrossRef] [PubMed]
- Almeida, G.H.D.; Iglesia, R.P.; Araujo, M.S.; Carreira, A.C.O.; Santos, E.X. Dos; Calomeno, C.V.A.Q.; Miglino, M.A. Uterine Tissue Engineering: Where We Stand and the Challenges Ahead. Tissue Eng. Part B. Rev. 2022, 28, 861–890. [Google Scholar] [CrossRef]
- Godoy-Guzmán, C.; Nuñez, C.; Orihuela, P.; Campos, A.; Carriel, V. Distribution of extracellular matrix molecules in human uterine tubes during the menstrual cycle: a histological and immunohistochemical analysis. J. Anat. 2018, 233, 73. [Google Scholar] [CrossRef]
- Elad, D.; Jaffa, A.J.; Grisaru, D. Biomechanics of early life in the female reproductive tract. Physiology 2020, 35, 134–143. [Google Scholar] [CrossRef]
- Matsuzaki, S. Mechanobiology of the female reproductive system. Reprod. Med. Biol. 2021, 20, 371. [Google Scholar] [CrossRef]
- Choucair, F.; Avella, M. Basic, translational and clinical studies in reproductive medicine and clinical reproductive sciences. J. Transl. Med. 2023, 21. [Google Scholar] [CrossRef]
- Jiao, S.Y.; Yang, Y.H.; Chen, S.R. Molecular genetics of infertility: loss-of-function mutations in humans and corresponding knockout/mutated mice. Hum. Reprod. Update 2021, 27, 154–189. [Google Scholar] [CrossRef]
- Rugg-Gunn, P.J.; Moris, N.; Tam, P.P.L. Technical challenges of studying early human development. Development 2023, 150. [Google Scholar] [CrossRef] [PubMed]
- Ferraz, M.A.M.M.; Henning, H.H.W.; Stout, T.A.E.; Vos, P.L.A.M.; Gadella, B.M. Designing 3-Dimensional In Vitro Oviduct Culture Systems to Study Mammalian Fertilization and Embryo Production. [CrossRef]
- Alzamil, L.; Nikolakopoulou, K.; Turco, M.Y. Organoid systems to study the human female reproductive tract and pregnancy. Cell Death Differ. 2021, 28, 35. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D cell cultures – a comparison of different types of cancer cell cultures. Arch. Med. Sci. 2018, 14, 910. [Google Scholar] [CrossRef] [PubMed]
- Nishida-Aoki, N.; Gujral, T.S. Emerging approaches to study cell–cell interactions in tumor microenvironment. Oncotarget 2019, 10, 785. [Google Scholar] [CrossRef]
- Cramer, M.C.; Badylak, S.F. Extracellular Matrix-Based Biomaterials and Their Influence Upon Cell Behavior. Ann. Biomed. Eng. 2020, 48, 2132. [Google Scholar] [CrossRef] [PubMed]
- Francés-Herrero, E.; Rodríguez-Eguren, A.; Gómez-álvarez, M.; Miguel-Gómez, L. de; Ferrero, H.; Cervelló, I. Future Challenges and Opportunities of Extracellular Matrix Hydrogels in Female Reproductive Medicine. Int. J. Mol. Sci. 2022, 23, 3765. [Google Scholar] [CrossRef] [PubMed]
- Francés-Herrero, E.; De Miguel-Gómez, L.; López-Martínez, S.; Campo, H.; Garcia-Dominguez, X.; Diretto, G.; Faus, A.; Vicente, J.S.; Marco-Jiménez, F.; Cervelló, I. Development of Decellularized Oviductal Hydrogels as a Support for Rabbit Embryo Culture. Reprod. Sci. 2021, 28, 1644–1658. [Google Scholar] [CrossRef]
- Zhang, Q.; Hu, Y.; Long, X.; Hu, L.; Wu, Y.; Wu, J.; Shi, X.; Xie, R.; Bi, Y.; Yu, F.; et al. Preparation and Application of Decellularized ECM-Based Biological Scaffolds for Articular Cartilage Repair: A Review. Front. Bioeng. Biotechnol. 2022, 10, 908082. [Google Scholar] [CrossRef]
- McInnes, A.D.; Moser, M.A.J.; Chen, X. Preparation and Use of Decellularized Extracellular Matrix for Tissue Engineering. J. Funct. Biomater. 2022, 13. [Google Scholar] [CrossRef]
- Fratini, P.; Carreira, A.C.O.; Alcântara, D.; de Oliveira e Silva, F.M.; Rodrigues, M.N.; Miglino, M.A. Endothelial differentiation of canine yolk sac cells transduced with VEGF. Res. Vet. Sci. 2016, 104, 71–76. [Google Scholar] [CrossRef]
- Liu, C.; Pei, M.; Li, Q.; Zhang, Y. Decellularized extracellular matrix mediates tissue construction and regeneration. Front. Med. 2022, 16, 56–82. [Google Scholar] [CrossRef] [PubMed]
- Almeida, G.H.D.R.; da Silva-Júnior, L.N.; Gibin, M.S.; dos Santos, H.; de Oliveira Horvath-Pereira, B.; Pinho, L.B.M.; Baesso, M.L.; Sato, F.; Hernandes, L.; Long, C.R.; et al. Perfusion and Ultrasonication Produce a Decellularized Porcine Whole-Ovary Scaffold with a Preserved Microarchitecture. Cells 2023, 12. [Google Scholar] [CrossRef] [PubMed]
- Movasaghi, Z.; Rehman, S.; Rehman, I.U. Raman Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2007, 42, 493–541. [Google Scholar] [CrossRef]
- Movasaghi, Z.; Rehman, S.; ur Rehman, I. Fourier Transform Infrared (FTIR) Spectroscopy of Biological Tissues. Appl. Spectrosc. Rev. 2008, 43, 134–179. [Google Scholar] [CrossRef]
- Sohutskay, D.O.; Buno, K.P.; Tholpady, S.S.; Nier, S.J.; Voytik-Harbin, S.L. Design and biofabrication of dermal regeneration scaffolds: role of oligomeric collagen fibril density and architecture. Regen. Med. 2020, 15, 1295. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, L.; Lin, S.; Liang, Q.; Shi, Z.; Xu, J.; Ma, H. Isolation and characterization of collagen from the muscle of Amur sturgeon (Acipenser schrenckii). Biotechnol. Bioprocess Eng. 2014, 19, 935–941. [Google Scholar] [CrossRef]
- Nara, S.; Chameettachal, S.; Midha, S.; Murab, S.; Ghosh, S. Preservation of biomacromolecular composition and ultrastructure of a decellularized cornea using a perfusion bioreactor. RSC Adv. 2015, 6, 2225–2240. [Google Scholar] [CrossRef]
- Arezoo, N.; Mohammad, H.; Malihezaman, M. Tissue engineering of mouse uterus using menstrual blood stem cells (MenSCs) and decellularized uterine scaffold. Stem Cell Res. Ther. 2021, 12, 475. [Google Scholar] [CrossRef] [PubMed]
- Bergholt, M.S.; Serio, A.; Albro, M.B. Raman Spectroscopy: Guiding Light for the Extracellular Matrix. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef]
- Nguyen, T.T.; Happillon, T.; Feru, J.; Brassart-Passco, S.; Angiboust, J.F.; Manfait, M.; Piot, O. Raman comparison of skin dermis of different ages: focus on spectral markers of collagen hydration. J. Raman Spectrosc. 2013, 44, 1230–1237. [Google Scholar] [CrossRef]
- Zhang, Q.; Andrew Chan, K.L.; Zhang, G.; Gillece, T.; Senak, L.; Moore, D.J.; Mendelsohn, R.; Flach, C.R. Raman microspectroscopic and dynamic vapor sorption characterization of hydration in collagen and dermal tissue. Biopolymers 2011, 95, 607–615. [Google Scholar] [CrossRef] [PubMed]
- Timchenko, E. V.; Timchenko, P.E.; Lichtenberg, A.; Assmann, A.; Aubin, H.; Akhyari, P.; Volova, L.T.; Pershutkina, S. V. Assessment of decellularization of heart bioimplants using a Raman spectroscopy method. J. Biomed. Opt. 2017, 22, 091511. [Google Scholar] [CrossRef] [PubMed]
- Villaret, A.; Ipinazar, C.; Satar, T.; Gravier, E.; Mias, C.; Questel, E.; Schmitt, A.M.; Samouillan, V.; Nadal, F.; Josse, G. Raman characterization of human skin aging. Skin Res. Technol. 2019, 25, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Naeem, E.M.; Sajad, D.; Talaei-Khozani, T.; Khajeh, S.; Azarpira, N.; Alaei, S.; Tanideh, N.; Reza, T.M.; Razban, V. Decellularized liver transplant could be recellularized in rat partial hepatectomy model. J. Biomed. Mater. Res. A 2019, 107, 2576–2588. [Google Scholar] [CrossRef] [PubMed]
- Emami, A.; Talaei-Khozani, T.; Vojdani, Z.; Zarei fard, N. Comparative assessment of the efficiency of various decellularization agents for bone tissue engineering. J. Biomed. Mater. Res. B. Appl. Biomater. 2021, 109, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; Li, F.; Zhao, X.; Ma, Y.; Li, Y.; Lin, M.; Jin, G.; Lu, T.J.; Genin, G.M.; Xu, F. Functional and Biomimetic Materials for Engineering of theThree-Dimensional Cell Microenvironment. Chem. Rev. 2017, 117, 12764. [Google Scholar] [CrossRef] [PubMed]
- Vieira-da-Silva, B.; Castanho, M.A.R.B. Resazurin Reduction-Based Assays Revisited: Guidelines for Accurate Reporting of Relative Differences on Metabolic Status. Molecules 2023, 28. [Google Scholar] [CrossRef] [PubMed]
- Kölle, S.; Hughes, B.; Steele, H. Early embryo-maternal communication in the oviduct: A review. Mol. Reprod. Dev. 2020, 87, 650–662. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Dai, M.; Gong, G.; Chen, M.; Cao, C.; Wang, T.; Hou, Z.; Shi, Y.; Guo, J.; Zhang, Y.; et al. The role of extracellular matrix on unfavorable maternal–fetal interface: focusing on the function of collagen in human fertility. J. Leather Sci. Eng. 2022 41 2022, 4, 1–17. [Google Scholar] [CrossRef]
- Idelevich, A.; Vilella, F. Mother and Embryo Cross-Communication. Genes (Basel). 2020, 11. [Google Scholar] [CrossRef]
- Slayden, O.D.; Luo, F.; Bishop, C. V. Physiological Action of Progesterone in the Nonhuman Primate Oviduct. Cells 2022, Vol. 11, Page 1534 2022, 11, 1534. [Google Scholar] [CrossRef]
- Matsuzaki, S. Mechanobiology of the female reproductive system. Reprod. Med. Biol. 2021, 20, 371. [Google Scholar] [CrossRef] [PubMed]
- Fujii, D.T.; Yohannes, E.; Por, E.D.; Gillette, L.; Beesley, R.D.; Heitmann, R.J.; Chow, G.E.; Burney, R.O. The proteome of human Fallopian tube lavages during the phase of embryo transit reveals candidate proteins for the optimization of preimplantation embryo culture. Hum. Reprod. 2021, 36, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Martino, F.; Perestrelo, A.R.; Vinarský, V.; Pagliari, S.; Forte, G. Cellular mechanotransduction: From tension to function. Front. Physiol. 2018, 9, 378185. [Google Scholar] [CrossRef]
- Venkata, V.D.; Fairuz, M.; Jamaluddin, B.; Goad, J.; Drury, H.R.; Tadros, M.A.; Lim, R.; Karakoti, A.; O’sullivan, R.; Ius, Y.; et al. Development and characterization of human fetal female reproductive tract organoids to understand M € ullerian duct anomalies. 2022. [Google Scholar] [CrossRef]
- Nicolas, J.; Magli, S.; Rabbachin, L.; Sampaolesi, S.; Nicotra, F.; Russo, L. 3D Extracellular Matrix Mimics: Fundamental Concepts and Role of Materials Chemistry to Influence Stem Cell Fate. Biomacromolecules 2020, 21, 1968–1994. [Google Scholar] [CrossRef] [PubMed]
- Urciuolo, F.; Imparato, G.; Netti, P.A. In vitro strategies for mimicking dynamic cell–ECM reciprocity in 3D culture models. Front. Bioeng. Biotechnol. 2023, 11, 1197075. [Google Scholar] [CrossRef]
- Moffat, D.; Ye, K.; Jin, S. Decellularization for the retention of tissue niches. J. Tissue Eng. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Kasturi, M.; Vasanthan, K.S. Effect of decellularization using sodium dodecyl sulfate on glycosaminoglycans content in the liver. https://doi.org/10.2217/rme-2023-0050 2023, 18, 527–530. [Google Scholar] [CrossRef]
- White, L.J.; Taylor, A.J.; Faulk, D.M.; Keane, T.J.; Saldin, L.T.; Reing, J.E.; Swinehart, I.T.; Turner, N.J.; Ratner, B.D.; Badylak, S.F. The impact of detergents on the tissue decellularization process: a ToF-SIMS study. Acta Biomater. 2017, 50, 207. [Google Scholar] [CrossRef]
- Ahsan, T.; Nerem, R.M. Bioengineered tissues: the science, the technology, and the industry. Orthod. Craniofac. Res. 2005, 8, 134–140. [Google Scholar] [CrossRef] [PubMed]
- de Jongh, D.; Massey, E.K.; Cronin, A.J.; Schermer, M.H.N.; Bunnik, E.M. Early-Phase Clinical Trials of Bio-Artificial Organ Technology: A Systematic Review of Ethical Issues. Transpl. Int. 2022, 35. [Google Scholar] [CrossRef] [PubMed]
- Khan, R.L.; Khraibi, A.A.; Dumée, L.F.; Corridon, P.R. From waste to wealth: Repurposing slaughterhouse waste for xenotransplantation. Front. Bioeng. Biotechnol. 2023, 11, 1091554. [Google Scholar] [CrossRef]
- Mozhiarasi, V.; Natarajan, T.S. Slaughterhouse and poultry wastes: management practices, feedstocks for renewable energy production, and recovery of value added products. Biomass Convers. Biorefinery 2022, 1, 1. [Google Scholar] [CrossRef]
- Diaz, P.S.; Solar, P.A.; Juica, N.E.; Orihuela, P.A.; Cardenas, H.; Christodoulides, M.; Vargas, R.; Velasquez, L.A. Differential expression of extracellular matrix components in the Fallopian tubes throughout the menstrual cycle. Reprod. Biol. Endocrinol. 2012, 10, 1–8. [Google Scholar] [CrossRef]
- Varga, I.; Csöbönyeiová, M.; Visnyaiová, K.; Záhumenský, J.; Pavlíková, L.; Feitscherová, C.; Klein, M. Functional Morphology of the Human Uterine Tubes in the 21st Century: Anatomical Novelties and Their Possible Clinical Applications. Physiol. Res. 2022, 71, S151–S159. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, B.B.; Pope, B.D.; Peters, M.M.; Ris-Stalpers, C.; Parker, K.K. The role of extracellular matrix in normal and pathological pregnancy: Future applications of microphysiological systems in reproductive medicine. Exp. Biol. Med. 2020, 245, 1163. [Google Scholar] [CrossRef] [PubMed]
- Jorge, S.; Chang, S.; Barzilai, J.J.; Leppert, P.; Segars, J.H. Mechanical Signaling in Reproductive Tissues: Mechanisms and Importance. Reprod. Sci. 2014, 21, 1093. [Google Scholar] [CrossRef] [PubMed]
- Ouni, E.; Bouzin, C.; Dolmans, M.M.; Marbaix, E.; Pyr Dit Ruys, S.; Vertommen, D.; Amorim, C.A. Spatiotemporal changes in mechanical matrisome components of the human ovary from prepuberty to menopause. Hum. Reprod. 2020, 35, 1391–1410. [Google Scholar] [CrossRef]
- Karvas, R.M.; Zemke, J.E.; Ali, S.S.; Upton, E.; Sane, E.; Fischer, L.A.; Dong, C.; Park, K. mi; Wang, F.; Park, K.; et al. 3D-cultured blastoids model human embryogenesis from pre-implantation to early gastrulation stages. Cell Stem Cell 2023, 30, 1148–1165. [Google Scholar] [CrossRef]
- Tu, Z.; Bi, Y.; Zhu, X.; Liu, W.; Hu, J.; Wu, L.; Mao, T.; Zhou, J.; Wang, H.; Wang, H.; et al. Modeling human pregastrulation development by 3D culture of blastoids generated from primed-to-naïve transitioning intermediates. Protein Cell 2023, 14, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Fontoura, J.C.; Viezzer, C.; dos Santos, F.G.; Ligabue, R.A.; Weinlich, R.; Puga, R.D.; Antonow, D.; Severino, P.; Bonorino, C. Comparison of 2D and 3D cell culture models for cell growth, gene expression and drug resistance. Mater. Sci. Eng. C. Mater. Biol. Appl. 2020, 107. [Google Scholar] [CrossRef] [PubMed]
- Hawkins, J.; Miao, X.; Cui, W.; Sun, Y. Biophysical optimization of preimplantation embryo culture: what mechanics can offer ART. [CrossRef]
- Besenfelder, U.; Havlicek, V. The interaction between the environment and embryo development in assisted reproduction. Anim. Reprod. 2023, 20. [Google Scholar] [CrossRef] [PubMed]
- Jafarbeglou, F.; Nazari, M.A.; Keikha, F.; Amanpour, S.; Azadi, M. Visco-hyperelastic characterization of the mechanical properties of human fallopian tube tissue using atomic force microscopy. Materialia 2021, 16, 101074. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, F.; Zhong, W.; Puscheck, E.; Shen, H.; Rappolee, D.A. Shear stress induces preimplantation embryo death that is delayed by the zona pellucida and associated with stress-activated protein kinase-mediated apoptosis. Biol. Reprod. 2006, 75, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Adil, A.; Xu, M.; Haykal, S. Recellularization of Bioengineered Scaffolds for Vascular Composite Allotransplantation. Front. Surg. 2022, 9. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, E.; Saleh, T.; Xu, M. Recellularization of Native Tissue Derived Acellular Scaffolds with Mesenchymal Stem Cells. Cells 2021, Vol. 10, Page 1787 2021, 10, 1787. [Google Scholar] [CrossRef] [PubMed]
- Joyce, K.; Fabra, G.T.; Bozkurt, Y.; Pandit, A. Bioactive potential of natural biomaterials: identification, retention and assessment of biological properties. Signal Transduct. Target. Ther. 2021, 6. [Google Scholar] [CrossRef] [PubMed]
- Khalili, A.A.; Ahmad, M.R. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int. J. Mol. Sci. 2015, 16, 18149. [Google Scholar] [CrossRef]
- Sowamber, R.; Nelson, O.; Dodds, L.; Decastro, V.; Paudel, I.; Milea, A.; Considine, M.; Cope, L.; Pinto, A.; Schlumbrecht, M.; et al. Integrative Transcriptome Analyses of the Human Fallopian Tube: Fimbria and Ampulla—Site of Origin of Serous Carcinoma of the Ovary. Cancers (Basel). 2020, 12, 1090. [Google Scholar] [CrossRef]
- Cruz Walma, D.A.; Yamada, K.M. The extracellular matrix in development. Development 2020, 147. [Google Scholar] [CrossRef] [PubMed]
- Saint-Dizier, M.; Schoen, J.; Chen, S.; Banliat, C.; Mermillod, P. Composing the Early Embryonic Microenvironment: Physiology and Regulation of Oviductal Secretions. Int. J. Mol. Sci. 2020, 21. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Representative scheme demonstrating the methodology used in the study. Created with BioRender.com.
Figure 1.
Representative scheme demonstrating the methodology used in the study. Created with BioRender.com.
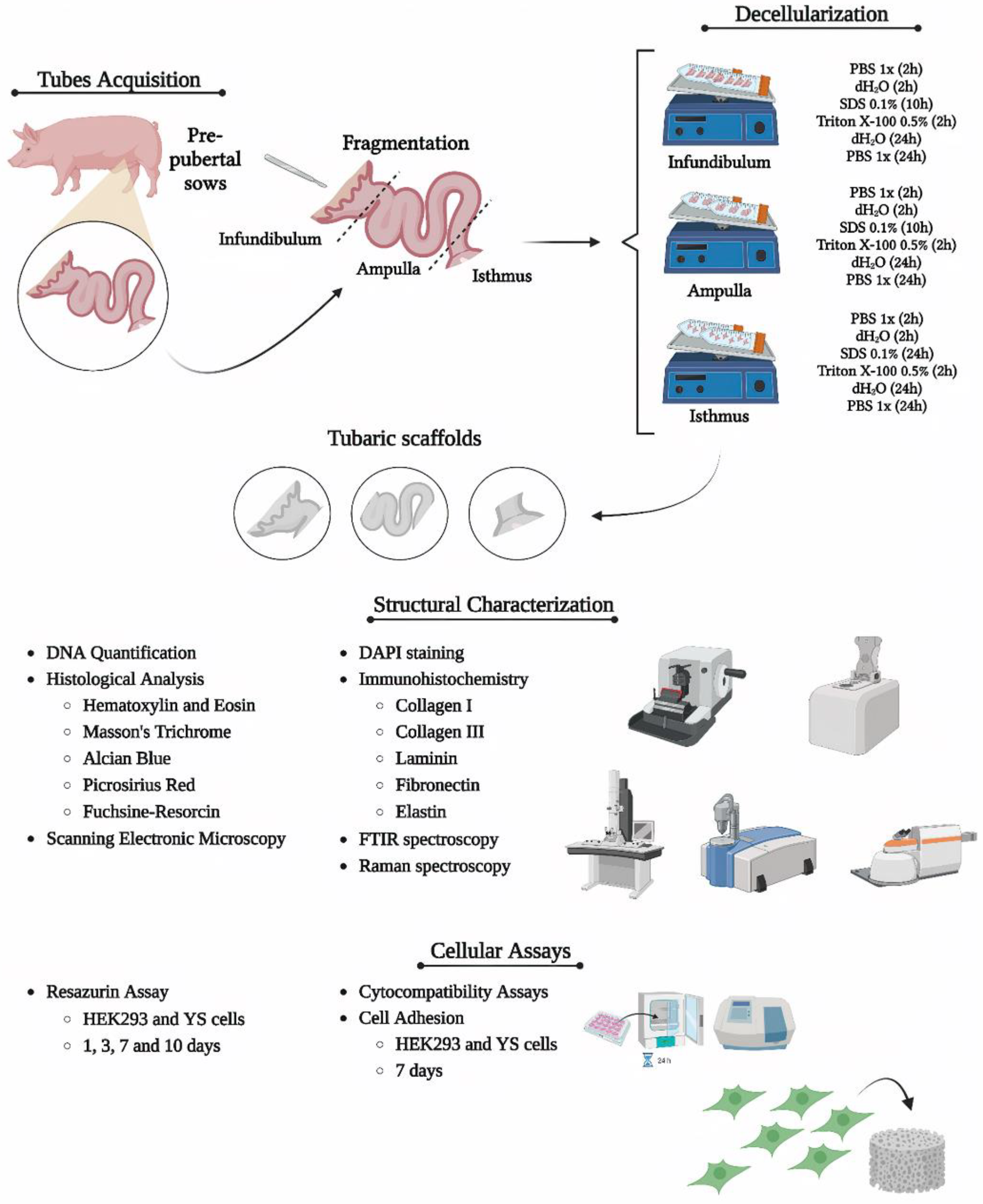
Figure 2.
Decellularization of porcine fallopian tube segments. Anatomic division of porcine fallopian tube, highlighting the three segments: infundibulum, ampulla and isthmus (A). Comparison of native infundibulum (INF) and decellularized infundibulum scaffolds (dINF) (B). Comparison of native ampulla (AMP) and decellularized ampulla scaffolds (dAMP) (C). Comparison of native isthmus (IST) and decellularized isthmus scaffolds (dIST) (D). DAPI staining of native tubal segments (E, G, I) and decellularized scaffolds (f, h, j). DNA quantification of tubal tissues before and after the decellularization process (K). * p < 0.05 compared to the native tissue.
Figure 2.
Decellularization of porcine fallopian tube segments. Anatomic division of porcine fallopian tube, highlighting the three segments: infundibulum, ampulla and isthmus (A). Comparison of native infundibulum (INF) and decellularized infundibulum scaffolds (dINF) (B). Comparison of native ampulla (AMP) and decellularized ampulla scaffolds (dAMP) (C). Comparison of native isthmus (IST) and decellularized isthmus scaffolds (dIST) (D). DAPI staining of native tubal segments (E, G, I) and decellularized scaffolds (f, h, j). DNA quantification of tubal tissues before and after the decellularization process (K). * p < 0.05 compared to the native tissue.
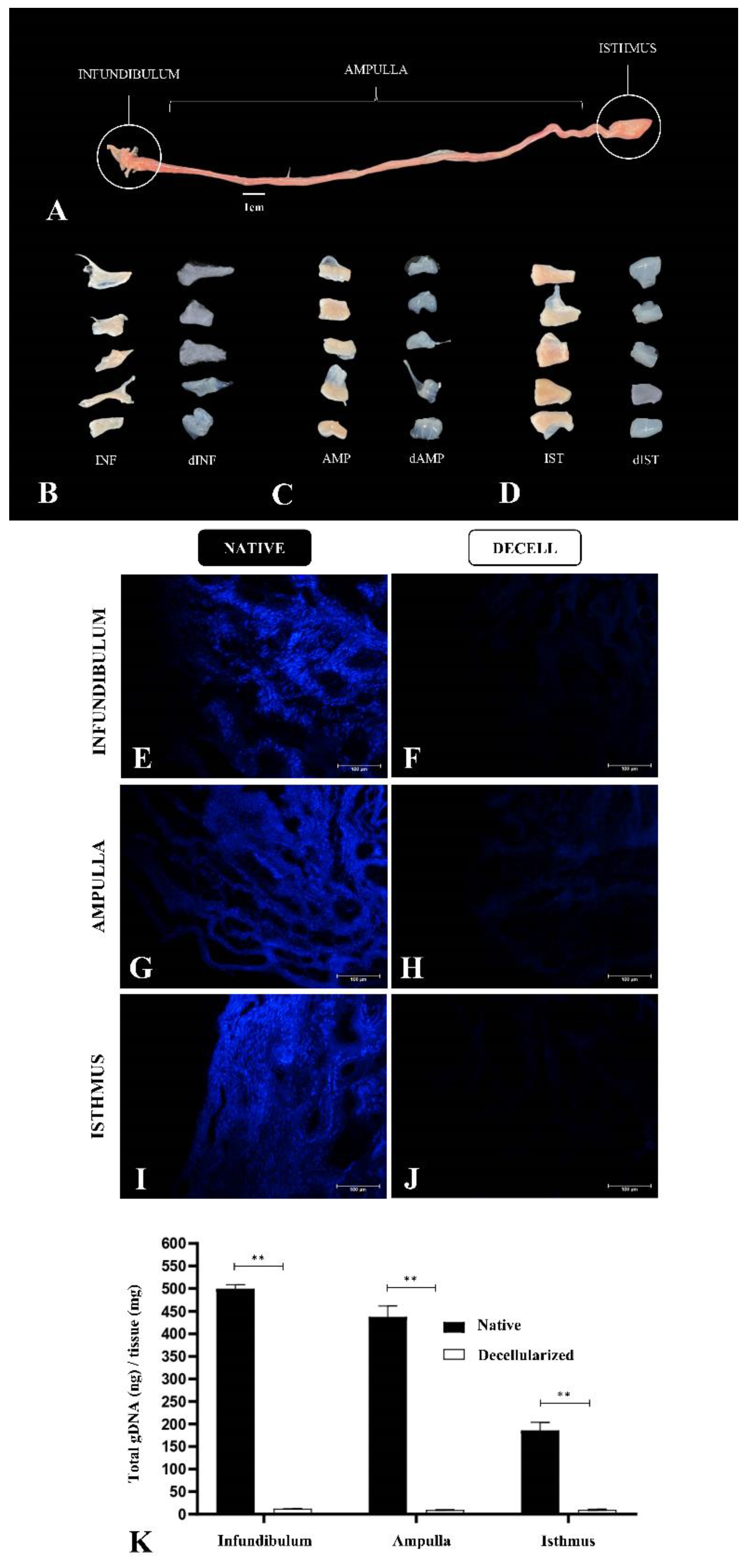
Figure 3.
Histological analysis of tubal ECM from native and decellularized infundibulum, ampulla and isthmus. Hematoxylin and Eosin staining indicates the tissue general structure, presence or absence of cells in native and decellularized samples (A-A5). Masson’s Trichrome staining highlights total collagen content in blue in native and decellularized samples (B-B5). Picrosirius Red staining also indicates collagen content (C-C5). Polarized Picrosirius Red staining distinguishes mature collagen (red and yellow) and immature collagen (green) (D-D5). Alcian Blue staining evidences GAG content in native and decellularized samples (E-E5). Weigert’s fuchsin-resorcin staining highlights elastic fibers content in native and decellularized samples (F-F5). Quantitative analysis of the relative occupied area of general ECM components in native and decellularized porcine infundibulum, ampulla and isthmus. Comparison of total collagen content (G), thick collagen fibers distribution (H), thin collagen fibers distribution (I), total glycosaminoglycan content (J) and elastic fibers content (K) between native and decellularized tubal samples. ** Statistically significant with p < 0.05 when compared with native group.
Figure 3.
Histological analysis of tubal ECM from native and decellularized infundibulum, ampulla and isthmus. Hematoxylin and Eosin staining indicates the tissue general structure, presence or absence of cells in native and decellularized samples (A-A5). Masson’s Trichrome staining highlights total collagen content in blue in native and decellularized samples (B-B5). Picrosirius Red staining also indicates collagen content (C-C5). Polarized Picrosirius Red staining distinguishes mature collagen (red and yellow) and immature collagen (green) (D-D5). Alcian Blue staining evidences GAG content in native and decellularized samples (E-E5). Weigert’s fuchsin-resorcin staining highlights elastic fibers content in native and decellularized samples (F-F5). Quantitative analysis of the relative occupied area of general ECM components in native and decellularized porcine infundibulum, ampulla and isthmus. Comparison of total collagen content (G), thick collagen fibers distribution (H), thin collagen fibers distribution (I), total glycosaminoglycan content (J) and elastic fibers content (K) between native and decellularized tubal samples. ** Statistically significant with p < 0.05 when compared with native group.
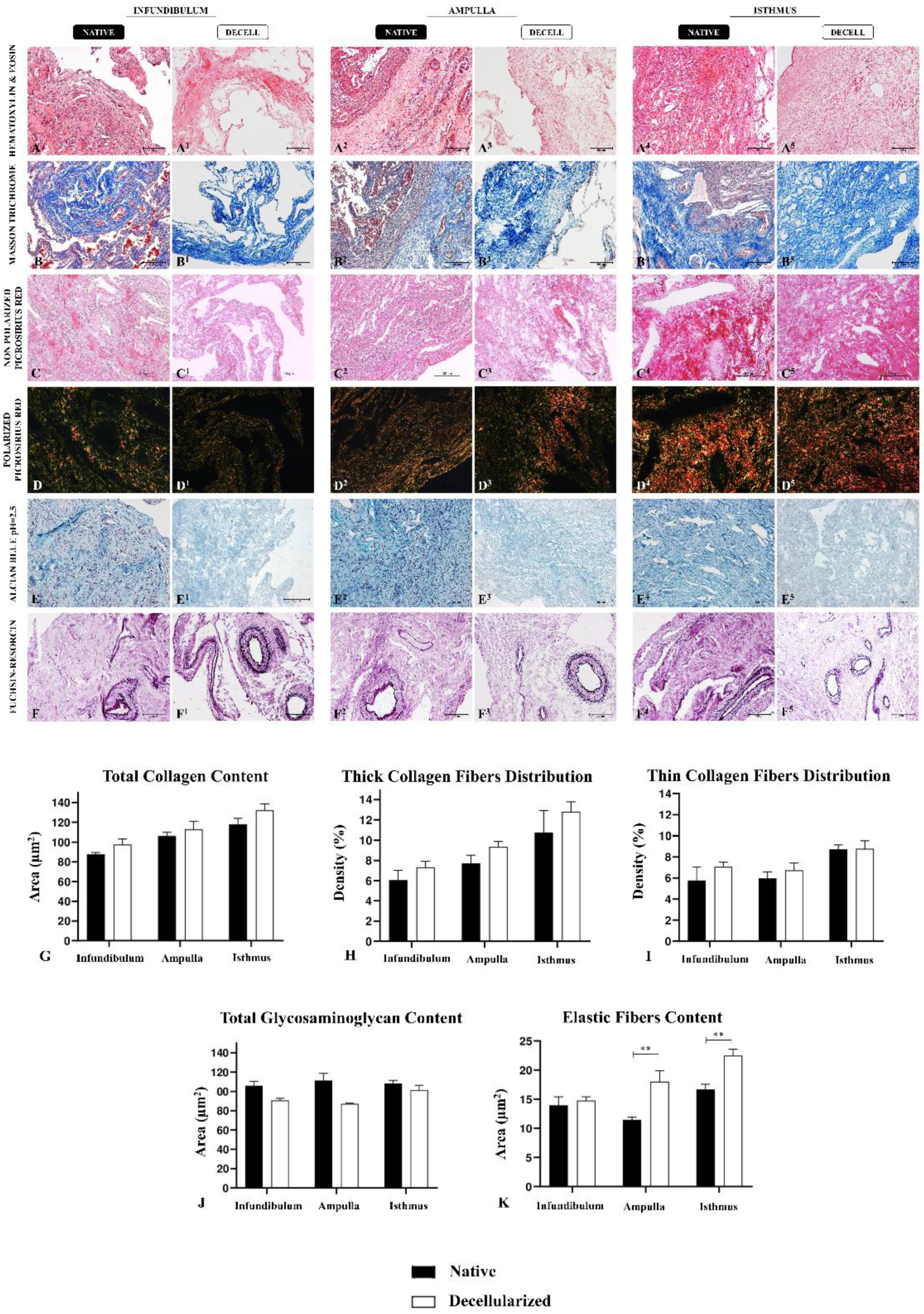
Figure 4.
Immunohistochemical analysis of the main ECM components of native and decellularized infundibulum, ampulla and isthmus. Immunolocalization of type I collagen (A-A5), type III collagen (B-B5), elastin (C-C5), fibronectin (D-D5) and laminin (E-E5) in native and decellularized samples. Quantitative analysis of the relative occupied area of main ECM components in native and decellularized porcine infundibulum, ampulla and isthmus. Comparison of type I collagen content (F), type III collagen content (G), elastin content (H), fibronectin content (I) and laminin content (J) between native and decellularized tubal samples. ** Statistically significant with p < 0.05 when compared with native group.
Figure 4.
Immunohistochemical analysis of the main ECM components of native and decellularized infundibulum, ampulla and isthmus. Immunolocalization of type I collagen (A-A5), type III collagen (B-B5), elastin (C-C5), fibronectin (D-D5) and laminin (E-E5) in native and decellularized samples. Quantitative analysis of the relative occupied area of main ECM components in native and decellularized porcine infundibulum, ampulla and isthmus. Comparison of type I collagen content (F), type III collagen content (G), elastin content (H), fibronectin content (I) and laminin content (J) between native and decellularized tubal samples. ** Statistically significant with p < 0.05 when compared with native group.
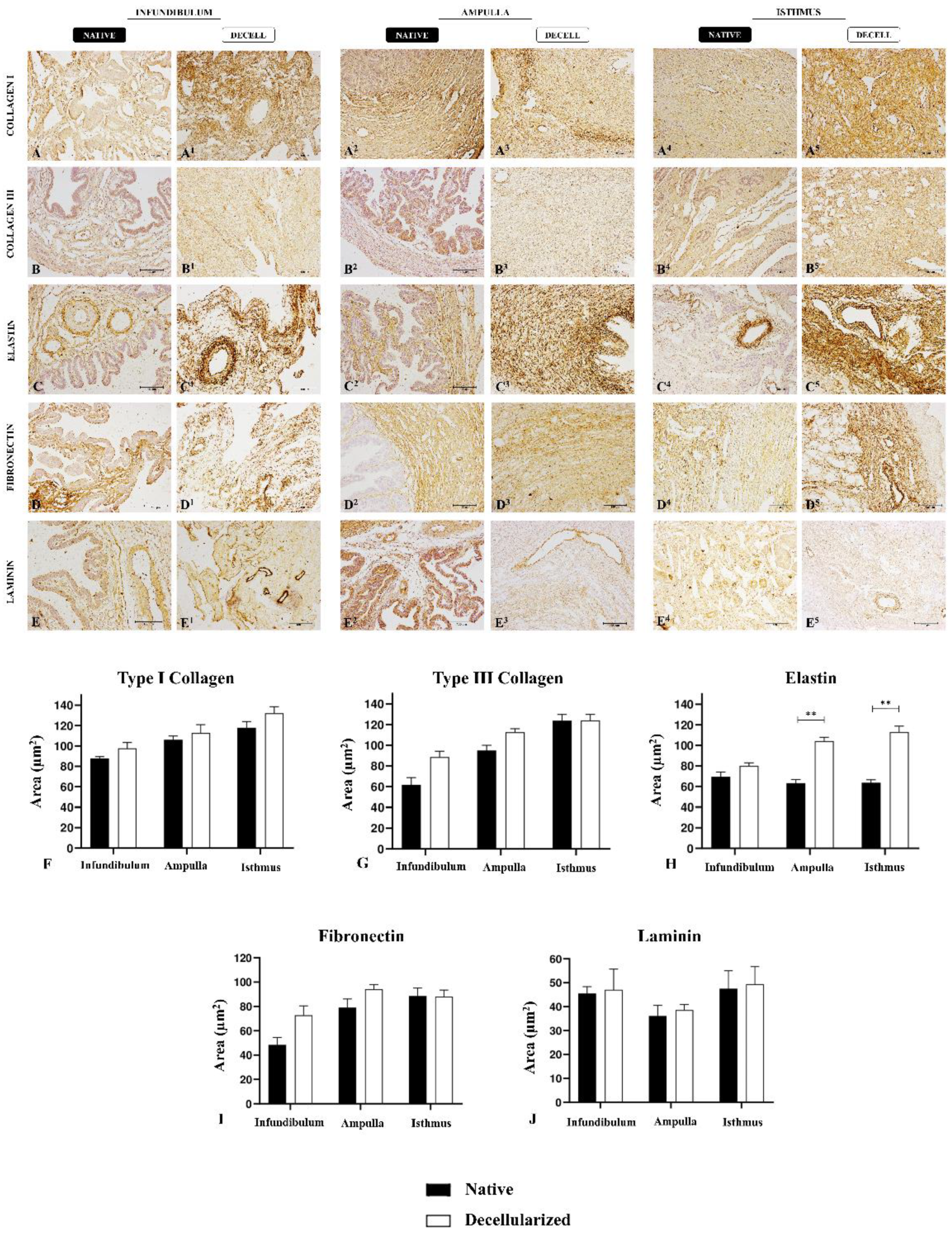
Figure 5.
Scanning electronic micrographs of native and decellularized infundibulum (A-H), ampulla (I-P) and isthmus (Q-X). Ep (epithelium), dutW (decellularized uterine tubal wall), Muc (mucosa), Mus (muscularis), Se (serous), EI (epithelial invaginations), CE (ciliated epithelium), CB (collagen bundles), infC (cilia of the infundibulum), CA (collagen arrangements), inv (invagination), CL (collagen layer), ampC (cilia of the ampulla), bv (blood vessels), Str (stroma). Full yellow arrows indicate thick fibers. Dotted arrows indicate thin fibers. Yellow asterisks indicate cells among ECM fibers. Yellow “x” indicates presence of mucus.
Figure 5.
Scanning electronic micrographs of native and decellularized infundibulum (A-H), ampulla (I-P) and isthmus (Q-X). Ep (epithelium), dutW (decellularized uterine tubal wall), Muc (mucosa), Mus (muscularis), Se (serous), EI (epithelial invaginations), CE (ciliated epithelium), CB (collagen bundles), infC (cilia of the infundibulum), CA (collagen arrangements), inv (invagination), CL (collagen layer), ampC (cilia of the ampulla), bv (blood vessels), Str (stroma). Full yellow arrows indicate thick fibers. Dotted arrows indicate thin fibers. Yellow asterisks indicate cells among ECM fibers. Yellow “x” indicates presence of mucus.
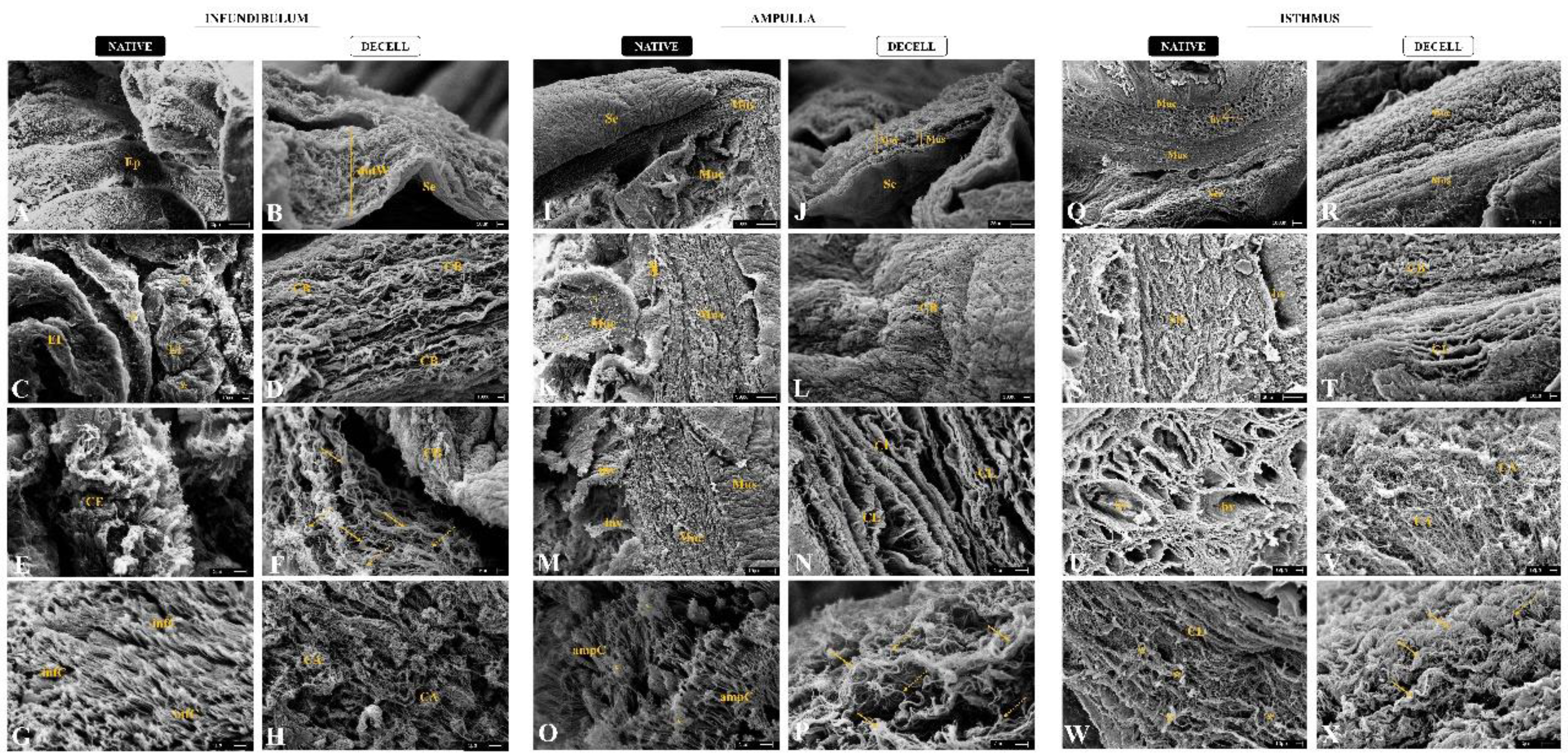
Figure 6.
FTIR-ATR spectra from the regions of native and decellularized infundibulum, ampulla and isthmus (A). Integrated band areas of Amide I (B), Amide II (C), proteoglycan content (D), and band area ratio of Amide III: 1450 cm-1 (triple helical structure of collagens) (E) in native and decellularized porcine infundibulum, ampulla and isthmus. ** Statistically significant with p < 0.05 when compared with native group.
Figure 6.
FTIR-ATR spectra from the regions of native and decellularized infundibulum, ampulla and isthmus (A). Integrated band areas of Amide I (B), Amide II (C), proteoglycan content (D), and band area ratio of Amide III: 1450 cm-1 (triple helical structure of collagens) (E) in native and decellularized porcine infundibulum, ampulla and isthmus. ** Statistically significant with p < 0.05 when compared with native group.
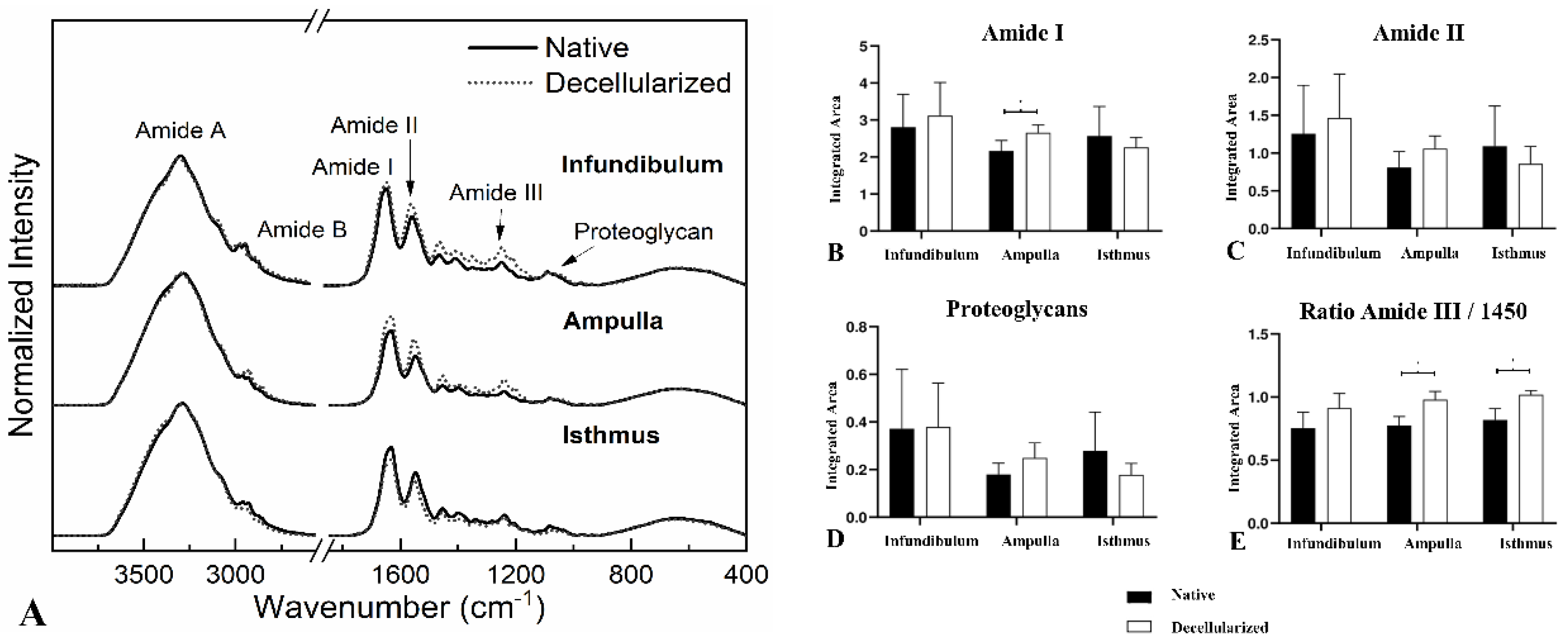
Figure 7.
Average Raman spectroscopy spectrum of the regions for native and decellularized samples from the porcine fallopian tube (A). Principal component analysis (PCA) score graph obtained for the infundibulum region (B). PCA core graph for the ampulla region (C). PCA core graph for the isthmus region (D). Integrated band areas of Amide I (E), Amide III (F), elastin (G), GAG content (H), phenylalanine (I) and proline/hydroxyproline (J) in native and decellularized porcine infundibulum, ampulla and isthmus. ** Statistically significant with p < 0.05 when compared with native group.
Figure 7.
Average Raman spectroscopy spectrum of the regions for native and decellularized samples from the porcine fallopian tube (A). Principal component analysis (PCA) score graph obtained for the infundibulum region (B). PCA core graph for the ampulla region (C). PCA core graph for the isthmus region (D). Integrated band areas of Amide I (E), Amide III (F), elastin (G), GAG content (H), phenylalanine (I) and proline/hydroxyproline (J) in native and decellularized porcine infundibulum, ampulla and isthmus. ** Statistically significant with p < 0.05 when compared with native group.
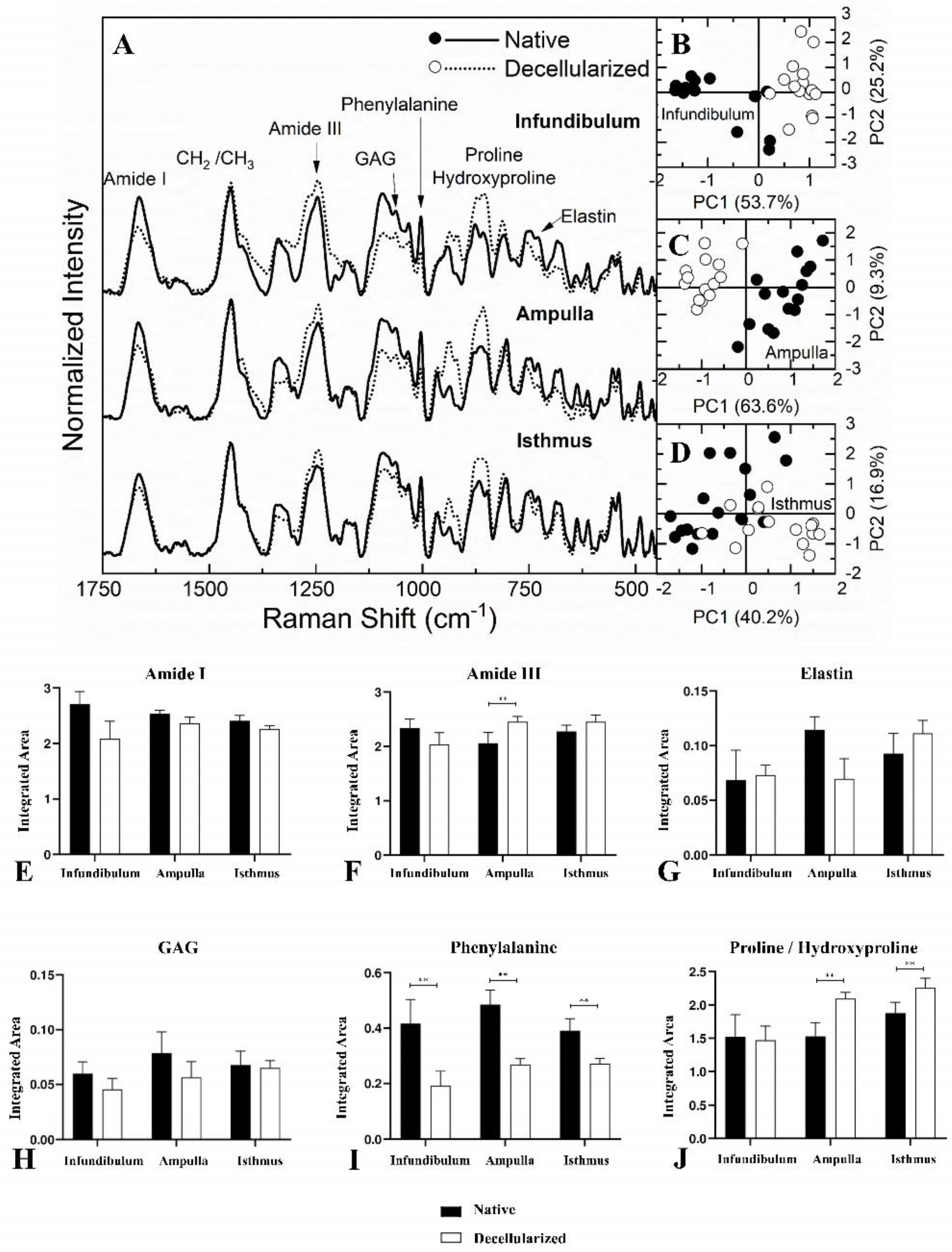
Figure 8.
Scanning electronic micrographs of the adhesion assay performed with YS and HEK293 cells seeded on decellularized infundibulum, ampulla and isthmus scaffolds after 7 days (A-R). Yellow asterisks indicate the cells anchored on the scaffolds ECM fibers.
Figure 8.
Scanning electronic micrographs of the adhesion assay performed with YS and HEK293 cells seeded on decellularized infundibulum, ampulla and isthmus scaffolds after 7 days (A-R). Yellow asterisks indicate the cells anchored on the scaffolds ECM fibers.
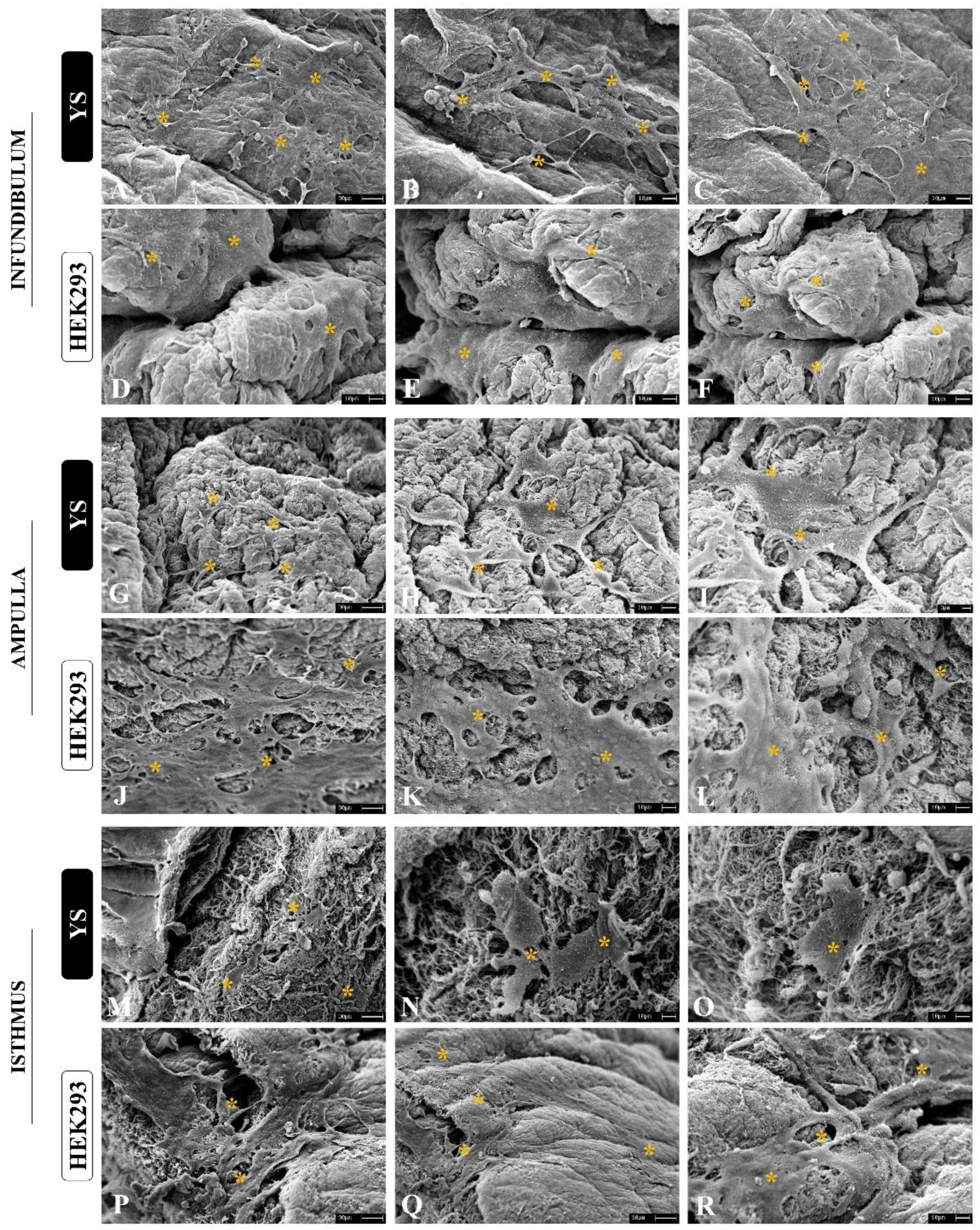
Figure 9.
Cell viability assessment of YS and HEK293 cells cultured on decellularized infundibulum (dINF), ampulla (dAMP) and isthmus (dIST) after 1, 3, 7 and 10 days. Trend graphs of YS cells (A,B,C) and HEK293 cells (D,E,F). The obtained absorbance data were converted and expressed in viability percentage. *Statistically significant for p < 0.01 when compared to the control group.
Figure 9.
Cell viability assessment of YS and HEK293 cells cultured on decellularized infundibulum (dINF), ampulla (dAMP) and isthmus (dIST) after 1, 3, 7 and 10 days. Trend graphs of YS cells (A,B,C) and HEK293 cells (D,E,F). The obtained absorbance data were converted and expressed in viability percentage. *Statistically significant for p < 0.01 when compared to the control group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



