
Submitted:
24 May 2024
Posted:
27 May 2024
You are already at the latest version
Abstract
Salinity, resulting from climate change and excessive mineral fertilization, burdens farmers and negatively impacts soil and water ecosystems in the Northeastern Nile Delta. Organic and biological approaches are crucial for addressing these issues. This study examined the effects of individual and combined inoculations with cyanobacteria, yeast, and Arbuscular Mycorrhizal Fungi (AMF), with or without K-humate, on enhancing salinity tolerance, increasing yield, and improving crop quality of wheat and faba bean. The aim was to reduce NPK mineral fertilizer application rates to 25% of recommended doses. An economic evaluation was also conducted. In a preliminary laboratory experiment, the interactive effects of these microbiomes on the production of plant antioxidant enzymes and soil enzymes were investigated under salinity stress. Results showed that co-inoculation, especially with K-humate, yielded superior outcomes compared to individual inoculations. The laboratory experiment findings were confirmed by a field trial conducted in saline-alkaline soil in the Northeastern Nile Delta region. The study found that the integrated inoculation of cyanobacteria, yeast, and mycorrhizae with K-humate, along with 25% mineral NPK fertilization enhanced microbial activity, including AMF colonization, spore density, and total bacteria, cyanobacteria, and yeast counts in the plant rhizosphere. It also increased soil enzyme activity, improving soil fertility compared to individual or mineral control treatments. This integration resulted in wheat yields of 2.77 tons/ha and faba bean yields of 4.75 tons/ha. Crop quality improved, with higher levels of nitrogen, potassium, phosphorus, and protein in wheat grains and faba bean seeds. Microbiomes inoculation increased potassium uptake over sodium, enhancing the potassium/sodium ratio and salt stress tolerance. This approach reduces reliance on costly mineral fertilizers by up to 75%, enabling economically viable cultivation in marginal lands. These sustainable practices optimize resource utilization, maximize economic efficiency, and preserve natural resources.
Keywords:
soil salinity
; cyanobacteria
; yeast
; mycorrhizae
; K-Humate
; faba bean
; wheat
; crop yield
; crop quality
; economic viability
1. Introduction
Global climate change and intensive agricultural practices have exacerbated the impact of abiotic stresses such as drought, salinity, UV rays, low and high temperatures that increase soil and water salinity especially in arid and semiarid areas and act as limiting factors that hinder plant growth and decrease agricultural productivity by 20% and 50% [1].These stressed conditions of soil salinization have grown up in Egypt to represent about 46% of the total Nile Delta zone due to the complex environmental risks from climate change and human-caused developments in addition to water scarcity problems [2,3]. Nowadays in Egypt, there is a tendency to expand the cultivated area of many crops in Northeastern Delta of Egypt, including wheat and faba bean in newly-reclaimed soil, although most of these soils are affected by salinity, which is a real global problem that requires urgent solutions [4]. El Husseiniya Plain in Sharkia Governorate in the Nile Delta region of Egypt of about 141.6 square kilometers area and irrigated from El-Salam Canal (drainage water and Nile river water 1:1) suffers from soil degradation caused by salinization (EC > 4 dSm-1 at 25 °C) and alkalinity that limits plant growth and development leading to yield losses [5]. Despite the continued expansion of wheat (Triticum aestivum L.) cultivations in Egypt, the increase in consumption is much higher than the rate of production as the average consumption was 16.6 million tons, while the production was estimated at 9.3 million tons, which led to an increase in imports that constitutes a huge burden on the Egyptian agricultural trade balance. The wheat gap is estimated at 7.3 million tons, and wheat and flour subsidies are estimated at 13.7 billion Egyptian pounds during the period 2012–2016 [6]. Royo and Abió [7] reported that about 397 million hectares under wheat cultivation are severely affected by salinity stress, crop yield starts to decline at a salinity stress level of 6–8 dS m−1 , which is imposing a serious threat to food security (WHO, 2019). Faba bean (Vicia faba L) is cultivated in the Mediterranean region as a rotational crop and is considered a popular legume food consumed worldwide and a major leguminous crop in Egypt; it is an important source of crude protein and essential amino acids for human and animal nutrition [8]. Vicia faba plants are proved to be moderately sensitive legumes to salinity, exhibiting a reduction in plant growth up to 50% under 6.7 dS m–1 salinity [9]. Since the yields of faba bean and wheat are negatively affected by abiotic stresses such as salinity and drought, developing effective, low-cost, and adaptive methods for managing and coping with abiotic stresses is a major challenge [10]. In addition to the negative effect of salinity stress on the plant growth and that inhibit crops productivity of these soils, it may also negatively affect its microbial diversity and decrease its fertility that lead to soil erosion and deterioration. However, some microbes that may tolerate high salinity conditions and still maintain natural activity, which may positively affect plant growth such as cyanobacteria, yeast and AMF, which get on/into the roots of plants and help in the absorption of water and nutrients from soil. Microorganisms could play an important role in adaptation strategies and increase of tolerance to abiotic stresses in agricultural plants. Plant-growth-promoting rhizobacteria (PGPR) mitigate most effectively the impact of abiotic stresses on plants through the production of exopolysaccharides and biofilm formation. AMF and dual symbiotic systems (endophytic rhizospheric bacteria and symbiotic fungi) also tend to mitigate the abiotic stress in plants [11]. Humic acids (HA) also convert elements to more beneficial forms and make them more effective, like PGBR. The beneficial effect of HA treatments on crops under salinity conditions can be attributed to its positive effect on the physical and chemical properties of soil, such as promoting microbial growth, increasing availability of nutrients and promoting enzyme activity, organic acid and amino acid content. Although, PGPRs and HA in different agricultural systems have already stimulated these processes reduced salinity damage in some crops under controlled experimental conditions, the field effect was insignificant and did not reflect productivity gains. This demonstrates the need for field tests to validate the potential of these biostimulants under real planting conditions [12].Thus, the main challenge of the Egyptian farmers is to grow up salt-tolerant plants under such saline and alkaline conditions and also to manage the ameliorating inputs to lessen further soil deterioration while sustaining soil productivity. Keeping this in mind, lab and field experiments were conducted to study the ability of cyanobacteria, yeast and AMF, individually or in combination, as inoculants to ameliorate salinity stress and enhance productivity of faba bean (Vicia faba L.) and Wheat (Triticum aestivum L.) in presence of humic acids and stimulatory dose (25%) of the recommended mineral fertilizers.
2. Material and Method
2.1. Preparation of Microbial Inoculation
Mixed cultures of cyanobacteria strains, yeast and Arbuscular Mycorrhizal Fungi (AMF), individually or in combination, were tested as inoculants to ameliorate salinity stress and enhance productivity of faba bean (Viciafaba L.), (cv-Nubaria 1) and Wheat (Triticum aestivum L.) (cv-Sakha 93) in presence of K-humateand stimulatory dose (25%) of the recommended mineral NPK fertilizer was tested.
2.1.1. Cyanobacteria (Cyano)
Four heterocystous (Nostoc muscorum isolate HSSASE1, Anabaena oryzae isolate HSSASE6, Anabaena sp. HSSASE11 and Wollea saccata isolate HSSASE7) and two non-heterocystous (Phormidium fragilis isolate HSSASE9and Spirulina platensis isolate HSSASE5) cyanobacteria strains were obtained from Microbiology Department, Soils, Water and Environment Res. Inst., Agric. Res., Center. The cyanobacteria strains were isolated from Egyptian soils and identified and deposited in GenBank under accession numbers (Table 1). All cyanobacterial strains were grown separately on BG11medium [13] except Spirulina platensis, which was grown on Zarrouk medium [14].
The cultures were incubated in a growth chamber with continuous shaking (150 rpm) under continuous illumination with Philips Florescent 40-W cool-white fluorescent lamps, at a relatively low light intensity (400-500 lux) and incubation at 27±2°C for 30 days to be used as inoculums for laboratory experiments. Some characterization parameters of cyanobacteria strains in the stationary phase such as pH values, algal dry weight [15] and optical density by spectrophotometer at 560 nm [16] are shown in Table 1.Equal portions on a volumetric basis from the suspensions of the cyanobacterial cultures, in the stationary phase, were mixed together for use in laboratory and field experiments. Seeds were soaked 12 hours before the laboratory germination experiment, while in the field experiment, 24 L/ha of cyanobacteria mixture was added in 3 equal doses after one month, 45 days and 60 days of sowing.
2.1.2. Yeast (Y)
Saccharomyces cerevisiae was kindly provided by microbiology Department Soils, Water and Environment Research Institute, Agriculture Research Center, Giza, Egypt. Saccharomyces cerevisiae was grown on glucose peptone and yeast extract (GPY) medium [17]. 100 ml of yeast culture containing 1×108cells.ml-1was used for single treatments, while, 50 ml was used just before inoculation for combined treatments.
2.1.3. Arbuscular Mycorrhizal Fungi (AMF)
Mixed spores of AMF (Glomus mosseas, Glomus intraradices and Glomus sp.) were originally extracted from the rhizosphere of maize plans grown in Sahl El-Hossynia Research Farm Station, El-Sharkia Governorate. The inoculum contains 38X102 spores/ Kg soil. AMF inoculated treatments received 100 g soil + 50 g infected maize roots + external hyphae. The inoculants were mixed with stickers (Arabic gum) and uniformly coated on the seeds and then air dried for 2 hours before sowing.
2.2. K-Humate Preparation (K-H)
Potassium humate (K-humate) was extracted from rice straw compost and characterized according to the standard method described by Sanchez – Monedero et al. [18]. Rice straw compost was mixed with water at a rate of 1:5 (w/v) and then treated with 0.5 N of KOH solution to extract the humic substances.
2.3. Lab and Field Trials
2.3.1. Preliminary Experiment
A one-month preliminary experiment was conducted in pots under laboratory conditions to study the antagonistic or synergistic effects of microbial treatments on rhizosphere enzyme activity (dehydrogenase and nitrogenase), plant oxidative enzyme activity (peroxidase and catalase), and the lengths of shoots and roots of faba bean (Vicia faba L. cv. Nubaria 1) and wheat (Triticum aestivum L. cv. Sakha 93). The aim was to assess the most promising inoculation for field trial application under salinity stress and mineral NPK deficiency. The seeds were sterilized with 5% Clorox, rinsed thoroughly, then presoaked in the biofertilizer treatments for 12 hours before being germinated in plastic pots (25 cm in diameter and 25 cm deep) filled with saline soil (18.57 dS.m-1) from the field experimental site. All treatments, except the control, were irrigated with saline water from El-Salam Canal for one month under laboratory conditions. The experimental design was as follows:
Based on the results of this preliminary experiment, the most promising interactions of seeds with different biofertilizer formulas, as well as their combination with K-humate under salinity stress will be selected for the field trial.
2.4. Field Experiment
2.4.1. Experimental location and Soil Analysis
The field experiment was conducted in one of the extension fields in the Al-Hussainiya area, Sharkia Governorate, Egypt (Figure 1), during the winter season of 2021/2022. The soil was clayey textured, highly alkaline, saline, and poor in organic matter. Soil texture characterization and chemical analyses (Table 2) were determined according to Page et al. [19]. Irrigation for this location was sourced from the El-Salam Canal (Table 3), which is a mix of Nile fresh water and agricultural drainage water (1:1).
This experiment aimed to investigate the individual and combined effects of cyanobacteria, yeast, AMF, and K-humate with 25% of the recommended mineral NPK fertilizers on the yield and yield components of faba bean and wheat under saline-alkaline soil conditions. Some soil biological activities were studied, including the total number of bacteria, cyanobacteria, and yeast, as well as soil enzymes (dehydrogenase and nitrogenase) after 75 days of cultivation. Additionally, AMF colonization was estimated after 45 and 75 days of cultivation, and spore numbers were assessed at harvest.
The field experimental design was as follows:
Measurements:
2.4.2. Soil Microbial Enzymes Analysis
2.4.3. Oxidative Enzymes Bioassay
2.4.4. Some Soil Biological Activity
Two soil enzymes activity (Dehydrogenase and Nitrogenase) were determined by the methods of Casidaet al. [24] and Dilworth [25], respectively. Concerning total count of bacteria, cyanobacteria and yeast were counted after 75th day from sowing. Total count of bacteriathe dilution plate method on nutrient agar [26] and total cyanobacterial counts were conducted by plating ten-fold serial soil suspension-dilutions in triplicate onto agarized BG11 medium [27] and Watanabe and Barraquie [28] or nitrogen fixing bacteria. AMF colonization (%) was estimated after 45, 75 days of sowing by the method described by Phillips and Hayman [29] while, spores number were determined at harvest stage according to Gerdemann and Nicolson [30].
2.4.5. Crop components and Some Chemical Analyses
At harvest, the collected wheat straw and grains samples were oven dried at 70 ºC to determine total N, P, K and Na contents in seeds were determined as described by Van Schouwenburg [31].Harvest index was calculated as a ratio of grain yield to total biological yield (straw yield + grain yield). Biological and grain yield were recorded at two central rows in each experimental unit. Subsequent sample was oven dried at 70oC for maximum 36 hours to estimate dry matter yield. 1000-grains were weighed on an electronic balance after drying for recording Seed Index.
2.4.6. Economic Evaluation of the Study
The data analysis was based on the averages of each treatment using partial budget analysis to obtain some economic and performance indicators [32] such as, profit /cost ratio, break-even yield and break-even price where is:
Profit = Revenue - Total cost; Profit /cost ratio = Profit/Total cost; Break-even yield = Total cost / Price; Break-even price = Total cost /Total yield.
2.4.7. Statistical Analysis
The obtained data were statistically analyzed using analysis of variance procedure according to Gomez and Gomez [32]. Means were compared using Gen-Stat software at 5% level of probability.
3. Results
3.1. Preliminary Experiment: Interactions of Plants, Microbiomes and K-Humate to Cope with Salinity Stress
Preliminary results showed that the biological treatments, with or without K-humate, significantly increased the activity of plant antioxidant enzymes (catalase and peroxidase) and soil enzymes (dehydrogenase and nitrogenase), except for the AMF treatment (T4), which failed to germinate. The root and shoot lengths of faba bean and wheat seedlings also exhibited different significant values compared to the control.
3.1.1. Faba Bean
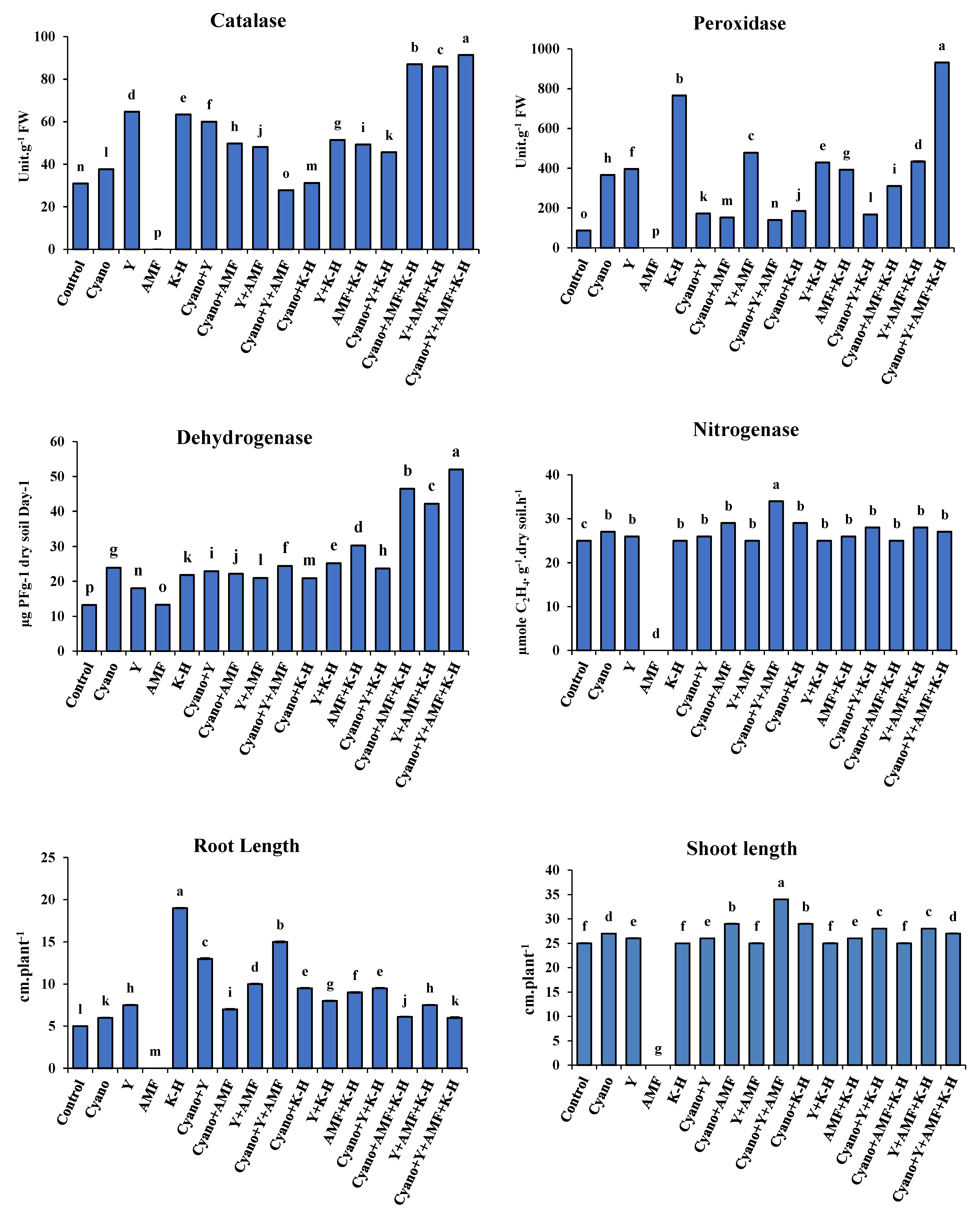
The results of Figure 2 and 3 illustrated that the combined treatment of cyano+Y+AMF+K-humate (T16) was the most effective treatment for improving the antioxidant enzyme activity (catalase and peroxidase), soil enzyme activity (dehydrogenase and nitrogenase), and seedling growth (root and shoot length) of faba bean plants under salinity stress. Specifically, the combined treatment of cyano+Y+AMF+K-humate (T16) resulted in the highest catalase and peroxidase enzyme activity in faba faba bean plants of 62.49 Unit.g-1 FW and 313.25 Unit.g-1 FW, respectively. The best significant increases in soil enzymes, dehydrogenase and nitrogenase, of 58.00 μg TPFg-1 dry soil Day-1 and 62.00 µg mol C2H4 g-1 soil day-1, respectively, were also achieved by the combined treatment of cyano+Y+AMF+K-humate (T16). Finally, the superior significant increases in root (20.67 cm) and shoot (61.00 cm) lengths were achieved by the combined treatment of Y+AMF+K-humate (T15) and Cyano+Y (T6), respectively.
3.1.2. Wheat
All treatments significantly increased soil enzymes (dehydrogenase and nitrogenase) and plant antioxidant enzyme activity (catalase and peroxidase), which was reflected in the lengths of roots and shoots of wheat seedlings. The combined treatment of cyano+Y+AMF+K-Humate (T16) resulted in the highest significant values of catalase (91.36 units g-1 FW), peroxidase (931.82 units g-1 FW), and dehydrogenase (52.0 g TPFg-1 dry soil Day-1) activities. While the combined treatment of cyano+Y+AMF (T9) showed the highest significant increases of nitrogenase activity (34.00 mole C2H4 g-1 soil day-1). K-Humate (T5) and cyano+Y+AMF (T9), respectively, showed the highest significant increases in the lengths of roots and shoots (Figure 2 and Table 5).
Figure 4.
Impact of microbiomes inoculation and k-humate on plant antioxidant enzymes, rhizosphere enzymes, shoot and root lengths of 30-day wheat seedlings.
Figure 4.
Impact of microbiomes inoculation and k-humate on plant antioxidant enzymes, rhizosphere enzymes, shoot and root lengths of 30-day wheat seedlings.
3.2. Field Experiment: Alleviating Effect of Salinity Stress via Microbiomes Inoculation and Organic Soil Amendment on Plants in Field Application
The preliminary laboratory experiment demonstrated the synergistic effect of cyanobacteria, yeast, and AMF, with or without K-humate. These results led to the development of different combinations of bio/organic fertilizers for field application on wheat and faba bean crops in the salt-stressed soils of the northeastern Egypt Delta that are irrigated with the El-Salam Canal (1:1 mixture of Nile River water and agricultural drainage water).
3.2.1. Soil Microbial Activity
The interactive effects of cyanobacteria, yeast, AMF, and K-humate, either inoculated alone or in combination, on soil microbial activity are shown in Table 6 and Table 7. The data in Table 6 shows that samples taken from the rhizosphere of wheat and faba bean at three stages (45 and 75 days after sowing and at harvest) showed higher significant differences (p ≤ 0.05) in the number of spores and total root colonization (%). AMF spores counts and root colonization (%) in the soil under study 45 and 75 days after sowing date showed significant variance between AMF inoculum alone (T4) versus being in combination with cyanobacteria, Saccharomyces cerevisiae, and K-humate (T8, T10, T11, and T12). The integrated application of the microbiomes consortium with K-humate T12 (Cyano+Y+AMF+K-Humate) had the highest inductive effects on AMF colonization and the spores number in the wheat and faba bean through the field experiment.
The results in Table 7 show that the total cyanobacterial count in the rhizosphere of wheat and faba bean was significantly (p ≤ 0.05) enhanced by the solitary, dual, and triple co-inoculation of microbiomes with or without K-humate under salinity stress, compared to the control (T1). This was true for all treatments except for the single treatment of K-humate in faba bean (T4). For individual inoculation, the cyanobacterial density was significantly (p ≤ 0.05) enhanced by the individual inoculation with cyanobacteria (36 and 30 cfu*10-3.g soil for wheat and faba bean, respectively) compared to the other individual treatments and the 100% NPK control (T1).
The dual co-treatments of K-humate with each of cyanobacteria, yeast and AMF significantly (p≤ 0.05) supported the cyanobacterial total count comparing to the individual microbiomes inoculation and 100% NPK control (T12). However, the superior significant (p≤ 0.05) total count of cyanobacteria in the rhizosphere of wheat and faba bean (78.00 and 61.00.f.u.*10-3.g-1 dry soil, respectively) was due to the co-treatment of K-humate with the consortium microbiomes (T12). Comparing to the control (T1), all the treatments significantly (p≤ 0.05) increased the total bacterial count in the rhizosphere of wheat and faba bean. The highest records of 200.00 and 307.00 cfu*10-3.g-1 dry soil were due to the co-treatment of k-humate with the consortium microbiomes (T12) for wheat and faba bean, respectively as shown in Table 6. Regarding to total count of yeast in the rhizosphere of wheat and faba bean (Table 7), it was significantly (p≤ 0.05) increased by all the individual microbiomes inoculation or the solo organic amendment of K-humate as well as their dual and triple inoculation (T12) that gave the highly significant (p≤ 0.05) results of 25.00 and 38.00 cfu.10-3.g-1 dry soil of the total count of yeast in the rhizosphere of wheat and faba bean, respectively. There was a significant variation in the response of soil enzymes (nitrogenase and dehydrogenase) to all treatments in both wheat and faba bean crops, as shown in Table 7. As for the effect of the individual treatments on the activity of N-ase enzyme, the AMF treatment failed to increase the N-ase enzyme, while the two treatments of cyanobacteria and yeast significantly (p≤ 0.05) increased N-ase in wheat rhizosphere. In contrary, only the AMF treatment significantly (p≤ 0.05) increased N-ase enzyme. However, the cyanobacteria and yeast treatments failed to increase the N-ase enzyme in faba bean rhizosphere compared to the mineral control treatment (T1). The highest significant (p≤ 0.05) increase of N-ase in wheat rhizosphere was achieved by Y+AMF+K-H (T11) and Cyano+Y+AMF+K-H (T12) while, the superior significant (p≤ 0.05) increases of N-ase in faba bean rhizosphere were achieved by these two treatments in addition to Cyano+AMF+K-H (T10). The activity of DHA-ase was increased by 166.16 and 99.71% via the individual treatment of cyanobacteria (T2) in the rhizosphere of wheat and faba bean, respectively. On the other hand, the integrated microbiomes consortium with K-humate (T12) resulted in the significant (p≤ 0.05) superior DHA-ase activity in the soil rhizosphere for both two crops.
3.2.2. Effects of Microbiomes and K-Humate on Wheat and Faba Bean Yield and Yield Components under Saline Soil Conditions
The solo, dual, and triple interacting effects of cyanobacteria, yeast, and AMF inoculation with/without K-humate significantly increased faba bean and wheat yield components as shown in Table 8 and Table 9, respectively. Among the individual effects of cyanobacteria, yeast, AMF, and K-humate on faba bean crop components (T2, T3, T4, and T5), cyanobacteria as a sole treatment had the most significant effect on the number of pods per plant, number of seeds per plant, weight of seeds per plant, weight of 100 seeds, biological yield per hectare, and harvest index, compared to the control (T1) with 100% NPK (Table 8).
The dual synergy of K-humate with each of cyanobacteria, yeast, and AMF (T6, T7 and T8) resulted in even greater significant increases in yield productivity, biological yield, and yield index compared to single inoculation of each microorganism alone (T2, T3, T4, and T5). However, the highest significant increase in these indicators was observed when K-humate was synergized with cyanobacteria (T6), compared to their synergy with other microorganisms.
Furthermore, it was found that the co-treatments of two microbes with K-humate (T9, T10 and T11) increased weight of 100 seeds (124.18%, 139.07% and 121.42%, respectively|), seed yield/ha (59.50, 95.04% and 82.64%, respectively) and straw yield/ha (45.52, 74.48% and 75.86%, respectively) compared to the full dose mineral NPK fertilizers (T1).
The significant superiority records of these indicators were achieved by the co-treatment of the microbiomes consortium and K-humate in terms of number of pods/plant (322.55%), number of seed/ plant (260.81%), seed yield/plant (692.45%), weight of 100 seeds (147.91%), Seed yield/ha (128.93%) and straw yield/ha (73.10%) compared to the full dose mineral NPK fertilizers (T1).
Table 9 shows the effect of inoculating wheat plants with microbiomes, with and without K-humate, on their yield components, compared to plants fertilized with the recommended full dose of mineral NPK fertilizers (T1). The results show that the highest significant number of spikes per plant (26) was recorded in plants inoculated with cyanobacteria alone (T2), followed by plants co-treated with K-humate and yeast (T7) and K-humate and AMF (T8), respectively. Cyanobacteria as a sole biofertilizer treatment (T2) also gave the highest significant number of grains per spike (60.17). The weight of 1000 grains was increased by 4.41% to 25.81% by inoculating wheat plants with microbiomes, with or without K-humate. The highest significant increase in the weight of 1000 grains (70.20 g) was recorded in plants inoculated with the consortium of microbiomes and K-humate (T12).
Moreover, inoculating wheat with microbiomes, with or without K-humate, increased straw and grain yields by 10.81% to 44.86% and 2.34% to 54.68%, respectively, compared to the control (T1). The combination of microbiomes with K-humate (T12) achieved the highest significant grain and straw yields of 4.75 and 6.43 ton.ha-1, respectively. The increase in yield and yield components also resulted in an increase in biological yield (ton/ha) by 8.62% to 48.88% compared to the control. The percentages of nitrogen (N), phosphorus (P), potassium (K), and sodium (Na) in wheat grains and faba bean seeds are indicators of crop quality. As shown in Figure 4 and Figure 5, the combination of exogenously added microbiomes (cyanobacteria, yeast, AMF, and K-humate) and the plant rhizosphere, along with the alleviating impact of salinity stress by K-humate, resulted in the highest NPK% while lowering Na% in faba bean seeds and wheat grains, respectively.
3.2.3. Influence of the Microbiomes and K-Humate on the Quality of Faba Bean Seeds and Wheat Grains
The percentages of nitrogen (N), phosphorus (P), potassium (K), and sodium (Na) in wheat grains and faba bean seeds are indicators of crop quality. As shown in Figure 4 and Figure 5, the combination of exogenously added microbiomes and the plant rhizosphere, along with the alleviating impact of salinity stress by K-humate, resulted in the highest NPK% while lowering Na% in faba bean seeds and wheat grains, respectively. The results also indicated that all of the individual and integrated microbiomes and K-humate with partial mineral fertilization (a quarter of the recommended dose of NPK mineral fertilizers for both faba bean and wheat crops) showed percentages of N, P and K significantly similar to that obtained by the control treatment (100% NPK). However, among all of the bio-organic amendments, T10 and T12 recorded the highest nitrogen percentages (4.45% and 4.55%, respectively), which reflected on the increase in protein percentage of faba bean seeds (Figure 4).
3.3. Economic Evaluation of the Study
Table 10 and Table 11 show the results of a feasibility study on using microbes and/or K-humate treatments with 25% of the recommended doses of NPK mineral fertilization for faba bean and wheat in soils under salinity stress in the northeastern Delta of Egypt. The study found that machinery, mineral fertilizers, and land rent (fixed costs) occupy the largest share of total production costs for the control treatments in faba bean and wheat, respectively. However, all of the treatments lowered these costs up to 25.50% and 11.53% of the total costs for faba bean and wheat, respectively, as a result of replacing 75% of NPK mineral fertilizers with the microbes and/or K-humate. The percentage of mineral fertilizer costs relative to total costs was lowered up to 7.32% and 2.96%, for faba bean and wheat, respectively, but this was not the main factor that increased the percentage of profits in response to these treatments, which was up to two times compared to the control. This indicated that the most influential factor in the profit is the yield quantity since all product sold at the same price. There are other important variables affecting the profit, such as the cost of the treatment production. The yield was calculated based on the average yield for each treatment, while the price was calculated based on the average market price per ton. All treatments raised the profit per hectare for wheat compared to the control, as well as in faba bean with the exception of AMF which was lower than the control as shown in Table 10 and Table 11.The most profitable treatments were T10, T11, and T12, which surpassed the control treatment in faba bean by 143.49, 37.33, and 164.95%, respectively, and in wheat by 90.47, 83.31, and 113.45%, respectively. Profit/cost ratio is the easiest method to determine the economic performance, the higher ratio is the better performance, If Profit/cost value is less than 1.00, then the production cost can be expected to higher than the returns. Data in Table 10 and Table 11 show that the lowest profit cost ratios of 1.6 and 0.85 were found by T1 (control) for faba bean and wheat, respectively while, the treatments from T2 to T12 recorded higher profit cost ratios than control (T1). Breakeven yield is the yield required to cover the cost of marketing, consequently, all treatments, with much higher yields, have achieved the higher profit. The price at which a crop must be sold in order to recoup its costs of production is known as the break-even price. Findings in Table 10 and Table 11 showed that the price needed to cover the production costs in the control treatment (T1) that received 100% mineral NPK fertilizers were 5.4 and 2.8 4 Egyptian pounds per kg for faba bean and wheat, respectively. Since the prices obtained by the microbiomes and K-humate treatments, either individually or co-treatments were higher than the control, consequently these treatments are the most profitable.
4. Discussion
4.1. Impact of Microbiomes and K-Humate on the Resistance of Faba Bean and Wheat Seedlings to Salt Stress
The results of this study revealed that faba bean and wheat seedlings' resilience to salinity stress was slightly enhanced by solitary applications of either microorganisms or K-Humate. Also, it was found that, compared to the single treatment or the control, the dual and triple combination microbial treatments significantly increased the resistance of plants to salinity stress. However, the co-treatments of K-Humate and microbes enhanced the microbial and plant resistance to salinity stress that led to the significant increases in the activities of plant antioxidant enzymes and soil enzymes, that was reflected in the successful growth of seedlings despite salinity stress with a significant increase in root and shoot lengths. Same findings were reported by Kthiri et al. [34], after conducting pots experiment for wheat germination, they found that inoculation with mixtures of microorganisms had a significant effect on increasing the biomass of stems and roots of wheat seedlings compared to untreated seedlings due to the enhancement of the antioxidative system (phenols and peroxidase enzymes) and reduce the harmful effect of NaCl on wheat seedlings and improve the resistance of seedlings to saline-alkali stress [35].
Recent studies have shown that using stress-tolerant microbiomes has emerged as a safer and sustainable means for mitigation of abiotic stresses resulting in improved crop productivity by enhancing the antioxidant potential, improving nutrient uptake, regulating the production of plant hormones, ACC deaminase, siderophore, exopolysaccharides and accumulating osmoprotectants and, thus, stimulating plant biomass and crop yield [36]. On the other hand, amendment of saline soil with humic acid significantly improved the variables affected by higher salinity as well as increased plant nitrate, nitrogen and phosphorus while, reduced electrical conductivity of soil, leakage of proline and electrolyte to plant, and enhanced plant root and dry weight of plants by allowing nutrients and water to be released to the plant as needed. Some studies reported that HA application mitigate the salinity effect in strawberry, maize and pepper seedlings in salty condition, thus it could be used as a growth regulator to regulate hormone level, improve plant growth and enhance stress tolerance. HA may stimulate shoot and root growth, and improve resistance to environmental stress in plant, but the physiological mechanism has not been well established [37, 38, 39].
4.2. The Integrated Effect of Microbiomes and K-Humate on Soil Microbial Activity under Salinity Stress
The combined application of cyanobacteria, yeast, AMF, and K-humate had a significant impact on the number of AMF spores and root colonization in wheat and faba bean plants under salt stress conditions. The differences were statistically significant, suggesting that the combined application of these microorganisms could be used as a biofertilizer to improve crop yields in the field, especially in salt-stressed soils. Mycorrhizal root infection rates as well as densities of spores in the rhizosphere of wheat and bean were outperformed by the consortium of microorganisms and K-humate (Table 6), which probably led to significant changes in the relative abundances of major groups of organic solutes, such as influencing carbohydrate formation and inducing the accumulation of specific osmoles such as proline). Additionally, mycorrhizal infection increases the phosphorus (P) uptake by the plant, mostly because of the ability of mycorrhizal fungi to absorb phosphate from the soil and transmit it to the roots of the host and increases the uptake of copper, zinc, and sulphate [40].
Sharma et al. [41] suggested that the impacts of AMF promoting the development of plant hormones are the cause of the resistance against biotic and abiotic stressors. Kumaret al. [42] compared the AMF-inoculated Jatropha curcas to non-inoculated plants under salinity exposure (1.7–8.5 dS m_1 NaCl for 60 d) and found increased soluble sugars and proline in the leaves of inoculated plants, allowing maintenance of leaf water status. Furthermore, inoculation of soybean (Glycine max) with Glomus etunicatum increased the root and shoot growth of plants grown with 50 and 100 mM NaCl, which was correlated with increased root by nutrient cycling and their interactions with other soil microorganisms [40].
The remarkable development in the biodensities of wheat and faba bean rhizosphere due to the co-treatment of consortium microbiomes and K-H in terms of the total number of cyanobacteria, bacteria and yeast, as well as soil enzymes (nitrogenase and dehydrogenase) as shown in the Table 7, could be attributed to the fact that microorganisms release phytohormones (indole acetic acid, gibberellins and cytocinine) and other PGPR that can induce defense programs such as systemic acquired resistance and induced systemic resistance, thus reducing phytotoxic microbial communities, increased root length, root surface area and number of root tips, leading to enhanced uptake of nutrients thereby improving plant health under stress conditions. It was also found that the combination of PGPR and Compost tea enhanced soil ion balance by decreasing Na+ in the root zone by creating IAA and exopolysaccharide, thus enhancing K+ absorption via the roots to the leaves in wheat plants [43 , 44].
Cyanobacteria are capable of solubilizing microbial nutrients and dissolving insoluble carbonatenodules through the secretion of oxalic acid. In this respect Han and Lee [44] showed that some cyanobacteria produce polysaccharide products, binding Na+ in the root zone and hence alleviating the salinity stress on plant and microbial growth and activities. Cyanobacterial exopolysaccharides improve the soil aggregation by lowering the pH and electrical conductivity and increasing the hydraulic conductivity of saline and alkali soil [46]. Exopolysaccharide production by the cyanobacteria increased when exposed to higher concentration of salt which seems to play a role in metal biosorption and the enrichment of saline soils with cyanobacteria improved the soil quality by decreasing pH, exchangeable sodium, Na/Ca, conserve organic C, organic N, and organic P as well as moisture and converts Na+ to Ca2+. These metabolites produced by the cyanobacteria affect the gene expression of the host plants and thereby bring about qualitative and quantitative changes in the phytochemical composition of the plants. Experiments carried out with live inoculum or with the extracts of cyanobacterial strains on several plant species, such as rice, wheat, maize, cotton etc., have demonstrated the synthesis of signalling metabolites [47,48,49].On the other hand, yeasts have been found in different soils and rhizosphere of various plants [50, 51]. Although the numbers of yeasts are low in comparison with other microorganisms, many investigators claimed that this group of organisms appear to play an important role in soil fertility and are capable of producing certain growth promoting substances such as hormones, amino acids, vitamins, proteins, organic acids, and soluble and volatile exudates [52, 53]. Nonetheless, despite the known ability of yeasts to produce organic acids, there have been very few reports on their ability to solubilise inorganic phosphate [54].
Interactions between AMF and other soil microorganisms may occur widely. Shifts in the presence or abundance of microbial species occurring in the rhizosphere of AMF plants and interaction between AM and other rhizosphere inhabitants can be detrimental to the AMF and certain rhizosphere microorganisms. Most research has focused on the effects of AMF on plant physiology by inoculating individual inocula or mixtures of only a few strains, and less attention has been paid to discerning the effects of complex inoculum mixtures. In nature, AMF colonize roots along with many other microbes, which are collectively called the “microbiome” Many other microbial organisms, such as fungal endophytes and bacteria, can also confer stress tolerance in plants [55].
Dastogeer et al. [56] drawn the attention to the consideration of microbe-microbe interactions could be an exciting area for future research as they found through reviewing 97 published peer reviews related to the effect of AMF on plant responses under salinity stress that the majority of these studies were conducted in controlled growth chamber or greenhouse conditions, thus we conducted lab prelaminary experiment followed by application in field experiment on two salinity sensitive crops (wheat and faba bean) to provide more data regarding the variability or stability of AMF-conferred salinity tolerance before advocating farmers for farm-level applications.
Furthermore, yeasts may enhance AMF development by supplying vitamin B12 to the rhizosphere, as AMF have been shown to be stimulated by this vitamin [57]. Thus, vitamin B12 produced by the soil yeasts might have resulted in better plant growth and yield in plants treated with both AMF and soil yeasts. The observations of Boby et al. [53] show the effect of inoculation with S. cerevisiae on non-AMF plants to be negligible, while it did increase the root colonization and spore count of AMF plants. This suggests that the yeasts specifically stimulate AMF development rather than the host plant, which upholds the observation made by Larsen and Jacobsen [58]. It is quite possible that Vitamin B12 production by soil yeasts could be the main reason for the stimulation of AMF development observed in this study and needs further investigation.
4.3. Effects of Microbiomes and K-Humate on Wheat and Faba Bean Yield in Saline Soil
Results in Table 8 and Table 9 showed that the integrated effect of microbiomes (cyanobacteria, yeast, and AMF) and K-humate with only 25% of the recommended NPK mineral fertilizers significantly improved faba bean and wheat yield components when compared to the individual K-humate or biofertilization as well as mineral fertilization 100% NPK. Our results are in accordance with Hamed et al. [59] who studied the effect of cyanobacteria and yeast inoculation, alone or both in combination, with different N-levels on wheat growth, grains quality and wheat yield components and they found that the integrated inoculation of cyanobacteria and yeast with 50% N, 75% N-fertilization enhanced wheat growth characteristic (Chl a,b), upgraded soil microbial community (nitrogenase activity and CO2 evolution), enhanced NPK-uptake Kg/fed and protein% in wheat grains. They also added that this combination recorded a wheat yield components (plant height, number of spikes/m2 , number of grains/spike, 1000 grains wt (gm) and grains yield (tonne/fed) insignificantly different from those obtained by the full recommended dose (100% N) in wheat cultivation.Same results were reported by Ghazal et al. l59] when combined cyanobacteria with R. radiobacter and different levels of nitrogen fertilization achieved a greater grain yield comparable to 100% N and induced the highest NPK-uptake in wheat plants either grains and straw. Generally, inoculation of N2-fxing cyanobacteria to cereals such as wheat and rice has been proven to induce N-availability in soil and consequently to plant leading to vigor seedlings with optimal yields at harvest stage [60, 61, 62]. Combined application of bio fertilizers caused considerable increase in plant height over all single treatments. Tillering enhanced significantly due to application of bio-fertilizers either alone or in combination. Greater wheat tillering was noticed when the crop received combined treatments than other single treatments. Similar trend of results was also observed in case of yield components of wheat such as spikes /m2 , grains/spike and 1000- grains weight when the crop received bio-fertilizers either alone or combined. Accordingly, the highest grain yield was recorded when the crop received combined bio-fertilizers [63]. This findings are supported our hypothesis that, in comparison to the control and the individual applications of the microbiomes inoculation or K-H soil amendment, the combined application of microbiomes consortium and K-Humate (T12) significantly improved the yield and quality of wheat and faba bean crops. Furthermore, the combined application markedly (p ≤ 0.05) boosted the antioxidant enzymes. Our research showed that the combination treatment could increase soil microbial activity and activate soil enzymes, primarily dehydrogenase and nitrogenase. In general, the combination treatment has demonstrated a good effect in terms of stimulating plant development and raising elements concentrations in wheat and faba bean under salt stress.Similarly, Alharbi et al. [12]concluded that the combined application of plant growth-promoting microbes (PGPMs; Bradyrhizobium japonicum (USDA 110) and Trichoderma harzianum) and potassium humate (K-humate) resulted in a substantial improvement in K+ levels in the leaves compared to Na+ levels, which has a direct positive impact on an enhancement in the antioxidants defense system (CAT, POX, SOD), which caused the decline of the oxidative stress indicators (H2O2 , MDA, and EL%) as well as proline content that led to a significant increase in root length, nodule weight, soybean relative water content (RWC), stomatal conductance, photosynthetic pigments, net photosynthetic rate, soluble protein, seed carbohydrate content as well as the number of pods. plant−1 and seed yield. Upon these findings they recommended to maximize the soybean growth and productivity under harsh growth conditions (e.g., water stress and soil salinity). Salt stress leads to ionic imbalance in plants due to excessive accumulation of Na+ and Cl–, which reduces the uptake of other mineral nutrients such as K+, Ca2+, and Mn2+. Excess Na+ accumulation leads to nutritional imbalance, membrane permeability and instability (resulting from Ca+ displacement by Na+), and an overproduction of reactive oxygen species (ROS), which cause oxidative damage on cellular macromolecules [64]. These biochemical alterations limit plant growth and performance, leading to crop yield reduction. To counteract salinity stress, plants induce a cascade of specific adaptative responses including K+ transport, phospholipid modifcations, activation of ROS-scavenging enzymes, and production of compatible solutes to compensate for the osmotic pressure of Na+ [64:67]. Such adequate natural stress responses play an important role in inducing plant tolerance on saline soils. Enhanced K+ uptake and reduced Na+/K+ ratio in the leaves of treated plants indicate microalgae-cyanobacteria extract formulations’s active role in reestablishing ion homeostasis. Nutrient uptake can be improved by enhanced root biomass, which subsequently increases the roots’ surface for nutrient absorption. These results indicate that microalgae-cyanobacteria extract formulations stimulated plant growth and tolerance responses through (i) enhanced antioxidant enzyme activities and (ii) improved root growth and nutrient uptake. Therefore, combined cyanobacteria-yeast-mycorhizae formulations could be another sustainable alternative to boost nutrient uptake, growth, and crop adaptability under normal and saline conditions [68].Nutrients accumulations in the grain were increased in plants treated with the biofertilizer consortium (Cyano+Y+AMF) plus K-humate with a quarter doses of chemical fertilizers, compared to those of complete mineral fertilization treatment. A clear benefit of biofertilizer application in the improvement of protein, N, P and K contents versus Na in the wheat and faba bean grains was observed in comparison with untreated control, especially under stress conditions. The most striking observation was the absence of significant differences between biofertilizer and chemical fertilizer treatments for most parameters. Moreover, the overall response to the biofertilizer consortium was accompanied by greater changes in the biological yield/ha and harvest index (%). In conclusion, the biofertilizer consortium + 25% mineral NPK fertilizers improved the yield and nutrient status of wheat grains and faba bean seeds to a similar extent as 100% chemical fertilizers, particularly under stress conditions, demonstrating the value of the integration of the used microbiomes consortium as a sustainable fertilization treatment.
4.4. Economic and Sustainability Evaluation of the Eco-Friendly Microbiomes Fertilizers
Soil salinization, one of the causes associated with the deterioration of strategic crops, such as wheat, maize, beans, etc.. low productivity is one of the environmental concerns felt most acutely by the poor in developing countries. In fact, according to our results in this applied study, we concluded that the total inputs cost of fertilizers used for the combined microbiomes and K-humate (T12), with only a quarter of the mineral dose of NPK fertilizers, were about 1135 and 760 L.E. for wheat and faba bean, respectively, which less than the traditional cropping system (100% NPK mineral fertilizers) by 1365 and 240 L.E, for wheat and faba bean, respectively. At the same time, the profitability of the microbiomes biofertilizers-based system surpassed the traditional cropping system by 113.45% and 164.95% for wheat and faba bean, respectively. Also our findings had proven economically and environmentally efficient, so we recommend further expansion of the production of these crops in such marginal saline soils, and also our findings pave the way for diffusion of this technology and its application in degraded lands in many other regions of Egypt through agricultural extension [69].On the other hand, market access for agricultural products remains the primary driver of production, due to the prominent role that the market plays in the development of agricultural value chains. The results of this study showed that wheat and faba bean production with the microbiomes biofertilizers-based system is globally profitable from an economic point of view. Bio-production of these strategic crops requires more investment comparing to the costly and environmentally polluted mineral fertilizers, keeping in mind that, fixed investments are the same for both production systems. At the level of variable costs, there is a big difference between the two cropping systems; however, most of these differences depend on the type of fertilization [70, 71].Thus, wheat and faba bean high production in alkaline-saline soils require vast inputs of microbiomes-based fertilizers as ecofriendly sustainable and economic input versus the mineral fertilizers cropping system. It is worthy to mention that the cost of biofertilizers produced from microbiomes in this study is only an experimental cost, which will be greatly reduced through the mass production of many tons if this technique is applied in the cultivation of wheat and faba bean. On the other hand, Ahmed et al. [72] concluded that wheat production contribute significantly to farm sustainability and contribute to alleviation of malnutrition. The actual production constraints restrict the sustainability of this important crop. Soni [73] conducted a study of the impact of improved wheat production technology, including high yielding varieties with cultural practices and comparing yield, input level and net returns. Farmers adopting advanced technology had 61.92%-76.07% higher yield as compared to the traditional system. Production costs are an important and fundamental issue in economic studies, because the production decisions depend largely on the level of production costs, as the volume of production is always linked to production costs, as the importance of studying the costs of production because it is a key factor in determining the net income [74].However, considering the values obtained from the cost-benefit ratios of the production systems (Table 10 and Table 11) we suggested that wheat and faba bean production with the microbiomes biofertilizers-based system is more profitable financially than the traditional system. Thus, this biotechnology represents a great challenge for the trend towards organic farming, opening up competitive national and international markets for young farmers and investors, as well as encouraging small holders who are open to innovation [75] and in principle, would be willing to adopt organic fertilizers if they understand the benefits of applying this technology well.
5. Conclusion
In the present study, effect of microbiomes inoculation and K-humate on plant antioxidant enzymes and rhizosphere enzymes, as well as stem and root lengths of wheat and faba bean seedlings 30 days after the initial laboratory experiment was determined to formulate the most promising combination under salinity stress for the field trial.
From the field trial we concluded that the integrated inoculation of cyanobacteria, yeast and mycorrhizae with K-humate along with 25% of mineral NPK fertilization of wheat and faba bean crops in saline-alkaline soils enhanced the microbial activity in terms of AMF-colonization, AMF-spores density, total counts of bacteria, cyanobacteria and yeast in plant rhizosphere, as well as increased soil enzymes activity (dehydrogenase and nitrogenase) which led to the improvement of soil fertility compared to the individual treatments or the mineral control. The integrated microbiomes fertilization with potassium humate proved highly efficient in increasing the productivity of wheat and faba bean crops and yield quality in terms of the proportions of nitrogen, potassium, phosphorus and protein in wheat grains and faba bean seeds. The study findings demonstrated that microbiomes inoculation significantly enhanced potassium uptake compared to sodium, resulting in an increased potassium/sodium ratio in plants. These results provide encouragement for farmers to expand the cultivation of wheat and faba beans by utilizing larger areas, particularly in marginal regions. The economic feasibility of this approach lies in its ability to optimize the utilization of available resources, leading to improved efficiency in wheat and faba bean crop production. Consequently, it also contributes to a reduction in the average cost of production. The need to develop and provide modern means and techniques that will raise the level of productivity and reduce costs to ensure the exploitation of productive resources optimization of economic efficiency. Focusing on the extension side in order to play its role in the transfer of information and the results of scientific research to farmers for adoption and raise their administrative capabilities and then raise the productive level in farm work, which is to achieve economic efficiency.
References
- Minhas, P.S.; Ramos, T.B.; Ben-Gal, A.; Pereira, L.S. 2020. Coping with salinity in irrigated agriculture: Crop evapotranspiration and water management issues. Agricultural Water Management 227: 105832.
- Negm, M. E., El- Kallawy, W. and Hefeina, A. (2019). Comparative study on rice germination and seedling growth under salinity and drought stresses. Env. Biodiv. Soil Security,, 3, 109-117. [CrossRef]
- Ouda, S. and Zohry A.E.H. (2020) Water Scarcity Leads to Food Insecurity. In: Deficit Irrigation. Springer, Cham,pp 1-13. [CrossRef]
- Abdellatif, K. F. ; E.A. El Absawy and Asmaa. M. Zakaria (2012). Drought stress tolerance of faba bean as studied by morphological traits and seed storage protein pattern. J. of Plant Studies, Vol. 1 (2): 47-54.
- Mohamed, A. I. (2013): Irrigation water quality evaluation In El-Salam canal project. International Journal of Engineering and Applied Sciences 3 (1): 21–28.
- Khalil, Y.M.M. , Abd El-Ghani, S.S. & Mansour, T.G.I. A standard analysis of Egyptian foreign trade structure for wheat. Bull Natl Res Cent 44, 20 (2020). [CrossRef]
- Royo, A., Abió, D. (2003). Salt tolerance in durum wheat cultivars. Spanish Journal of Agriculture Research, 1, 27–35. Phyton, 2021. [CrossRef]
- Vogelsang-O’dwyer, M. , Petersen, I. L., Joehnke, M. S., Sørensen, J. C., Bez, J., Detzel, A., et al. (2020). Comparison of faba bean protein ingredients produced using dry fractionation and isoelectric precipitation: techno-functional, nutritional and environmental performance. Foods 9:322. [CrossRef]
- Mass, E.V. , Hoffman, G.J., 1977. Crop salt tolerance: current assessment. American Society of Civil Engineers 103, 115–134.
- Al-Ashkar, I., Alderfasi, A., Romdhane, W., Seleiman, M. F., El-Said, R. A. et al. (2020). Morphological and genetic diversity within salt tolerance detection in eighteen wheat genotypes. Plants, 9, 287. [CrossRef]
- Inbaraj MP (2021) Plant-Microbe Interactions in Alleviating Abiotic Stress—A Mini Review. Front. Agron. 3:667903. [CrossRef]
- Alharbi, K.; Rashwan, E.; Hafez, E.; Omara, A.E.-D.; Mohamed, H.H.; Alshaal, T. Potassium Humate and Plant Growth-Promoting Microbes Jointly Mitigate Water Deficit Stress in Soybean Cultivated in Salt-Affected Soil. Plants 2022, 11, 3016. [CrossRef] [PubMed]
- Rippka, R., J. Deruelles, J. B. Waterburg, M. Herdman and R.Y. Stanier. 1979. Generic assignments, strain histories and properties of pure cultures of cyanobacteira. J. General Microbiol. 111:1-16.
- Zarrouk, C. 1966. Contribution á l’étuded’unecyanophycée. Influence de divers facteurs physiques et chimiquessur la croissance et la photosynthése de Spirulina maxima (Setch. et Gardner) Geitler. Ph. D. Thesis, University of Paris, France.
- Vonshak, A. 1986. Laboratory techniques for the cultivation of microalgae. In Handbook of microalgal mass culture. (Ed. A. Richmond).Boca Raton: CRC Press.
- Leduy, A. and N. Therien. 1977. An improved method for optical density measurement of semimcro blue-green alga Spirulina maxima. Biotechnol. Bioeng. 19:1219-1224.
- Difco Manual, 1985. Dehydrated culture media and reagents for microbiology. laboratories incorporated Detroit. Michigan, 48232 USS, p: 621.
- Sanchez-Monedero, M. A., Roid, A., Cegarra, J., Bernal,M.P. and Paredes, C. Effects of HCL-HF purification treatment on chemical composition and structure of humic acids. Eur. J. Soil Sci. 2002, 53, 375–381. [Google Scholar] [CrossRef]
- Page, A. L., R. H. Miller and D. R. Keeny. 1982. Methods of Soil Analysis. Part 2. Chemical and Microbiological properties. Second Edition. Madison, Wisconsin, USA.
- Glathe', H. and A. Thalmann (1970). Uber die microbielloactivitat and iherBeziehungenzuFruchtbrkeitsmerkmaleneinigerAcherbodenunterbesondererBerucksichtigung der dehydrogenase akativitat (TCC. Redukation).Zbl. Bakt. Abt. II, 124: 1-23.
- Dilworth MJ (1966) Acetylene reduction by nitrogen-fxing preparations from Clostridium pasteurianum. Biochimicaet Biophysica Acta 127:285– 294. [CrossRef]
- Allam, A.I. and Hollis, J.P. (1972) Sulphide Inhibition of Oxidases in Rice Roots. Phytopathology, 62, 634-639. [CrossRef]
- Góth, L. 1991. A simple method for determination of serum catalase activity and revision of reference range. Clinica Chimica Acta, 196, 143–151. [CrossRef]
- Casida, L.E., D. A. Klien and T. Suntoro. 1964. Soil dehydrogenase activity. Soil Sci. 98:371-376.
- Dilworth MJ (1966) Acetylene reduction by nitrogen-fxing preparations from Clostridium pasteurianum. Biochimicaet Biophysica Acta 127:285– 294. [CrossRef]
- APHA (1992) Standard Methods for the Examination of Water and Wastewater. 18th Edition, American Public Health Association (APHA), American Water Works Association (AWWA) and Water Pollution Control Federation (WPCF), Washington DC.
- Stanier, R. Y. , Kunisawa, R., Mandel, M., & Cohen-Bazire, G. (1971). Purification and properties of unicellular blue-green algae (order Chroococcales). Bacteriological reviews, 35(2), 171–205. [CrossRef]
- Watanabe, I. W. L. , and Barraque. 1979. Low levels of fixed N required for free-living organisms from rice roots. Nature 277:565-566.
- Phillips, J.M. and Hayman, D.A. (1970) Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Transactions of the British Mycological Society, 55, 158-161. [CrossRef]
- Gerdemann, J.W and Nicolson, T.H. (1963) Spores of Mycorrhizal Endogone Species Extracted from Soil by Wet Sieving and Decanting. Transactions of the British Mycological Society,46,235-244. [CrossRef]
- Van Schouwenburg,l. C. & Walinga I. (1967) The rapid determination of phosphorus in the presence of arsenic, silicon and germanium. Analytica. chim. Acta 37,269-271.
- Kay, J. A. (1986). Farm management, 2nd edition, Mc Graw-Hill, New York.
- Gomez MA, Gomez AA (1993) Statistical procedure for agricultural research (2nded). John Wile & Sons, New York, p 680.
- Kthiri, Z.; Ben Jabeur, M.; Machraoui, M.; Gargouri, S.; Hiba, K.; Hamada, W. Coating seeds with Trichoderma strains promotes plant growth and enhance the systemic resistance against Fusarium crown rot in durum wheat. Egypt. J. Biol. Pest Control 2020, 30, 139.
- Lau, SE. , Teo, W.F.A., Teoh, E.Y. et al. Microbiome engineering and plant biostimulants for sustainable crop improvement and mitigation of biotic and abiotic stresses. Discov Food 2, 9 (2022). [CrossRef]
- Phour M, Sindhu SS, Salama KHA, Morsy AA. 2022. Mitigating abiotic stress: microbiome engineering for improving agricultural production and environmental sustainability. Planta. 256(5):85. [CrossRef]
- Türkmen Ö, Şensoy S, Erdal İ. Effect of Potassium on Emergence and Seedling Growth of Cucumber Grown in Salty Conditions. Yuzuncu Yil University, J. Agric. Sci. 2000, 10, 113–117. [Google Scholar]
- Pinali N, Kaplan M. Investigation of effect on nutrient uptake of humic acid applications of different forms to strawberry plant. J. Plant Nutri. 2003, 26, 835–843. [Google Scholar] [CrossRef]
- Aydın, A., Kant, C., & Turan, M. Humic acid application alleviate salinity stress of bean (Phaseolus vulgaris L.) plants decreasing membrane leakage. African Journal of Agricultural Research 2012, 7, 1073–1086. [Google Scholar]
- Parial, R., Mohajan, S., Mt, H., Hashem, M., Das, M., & Islam, M. Isolation of arbuscular mycorrhizal Fungi and evaluation its effect on plant Growthover chemical fertilizers for better human health. SMU Medical Journal 2014, 1, 192–206. [Google Scholar]
- Sharma, J. , Zettler, L.W., Van Sambeek, J.W., Ellersieck, M.R., & Starbuck, C.J. (2003). Symbiotic Seed Germination and Mycorrhizae of Federally Threatened Platanthera praeclara (Orchidaceae).
- Kumar, A. , Sharma, S. & Mishra, S. Influence of Arbuscular Mycorrhizal (AM) Fungi and Salinity on Seedling Growth, Solute Accumulation, and Mycorrhizal Dependency of Jatropha curcas L.. J Plant Growth Regul 29, 297–306 (2010). [CrossRef]
- Shrivastava, P.; Kumar, R. Soil salinity: a serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi Journal of Biological Sciences 2015, 22, 123–131. [Google Scholar] [CrossRef]
- Ali, S.; Charles, T.C.; Glick, B.R. Amelioration of high salinity stress damage by plant growth-promoting bacterial endophytes that contain ACC deaminase. Plant Physiol. Biochem. 2014, 80, 160–167 [CrossRef]. [Google Scholar] [CrossRef] [PubMed]
- Han, H.S., & Lee, K.D. (2005). Plant Growth Promoting Rhizobacteria Effect on Antioxidant Status , Photosynthesis , Mineral Uptake and Growth of Lettuce under Soil Salinity.
- Anand Mohan, Baidyanath Kumar and Dina Nath. Cyanobacterial Consortium in the Improvement of Maize Crop. Int.J.Curr.Microbiol.App.Sci 2015, 4, 26. [Google Scholar]
- Ali, Laila K.M. and Mostafa, Soha S.M. (2009). Evaluation of potassium humate and Spirulina platensis as a bio-organic fertilizer for sesame plants grown under salinity stress. The 7th International Conference of Organic Agriculture, 13-15 December, Egypt. J. Res., 87(1):369-388.
- Mostafa, Soha S.M.; AbouElkhair, Amal W. and Shhata, Heba Sh. Effect of plant growth promoting substances from rhizo- and cyano-bacteria on sugar beet growth, yield and yield quality in saline soil. Intr. J. of Acad. Res. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Badawi, H. Mona; Mostafa, S. M. Soha; Abo El-Rakha, S. Eman; Mostafa, A. A. and Fayez, M. Cereal-PGPR interweave in salt-affected environments: towards plant persistence and growth promotion. International Journal of Scientific & Engineering Research, Volume 7, Issue 11, November-2016.
- Morais, P.B., M.B. Martins, A.N., Hagler, L.C. Mendonça-Hagler and L.B. Klaczko. 1995. Yeast succession in the Amazon fruit Parahancornia amapa as resource partitioning among Drosophila spp. Applied and Environmental Microbiology 1995, 61, 4251–4257. [Google Scholar]
- Ganter, P.F. 2006. Yeast and invertebrate associations. In: Biodiversity and Ecophysiology of Yeasts. The Yeast Handbook, (eds). C.A. Rosa and G. Peter (pp. 303-370). Heidelberg: Springer.
- Sampedro, I., E. Aranda, J. M. scervino, S. Fracchia, I. García-Romera, J. A. Ocampo and S. Godeas. Improvement by soil yeasts of arbuscular mycorrhizal symbiosis of soybean (Glycine max) colonized by Glomus mosseae. Mycorrhiza 2004, 14, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Boby, V.U., A.N. Balakrishna and D.J. Bagyaroj. Effect of combined inoculation of an AM fungus with soil yeasts on growth and nutrition of cowpea in sterilized soil. World Journal of Agriculture Sciences 2007, 3, 423–429. [Google Scholar]
- Hesham, A. and H.M. Mohamed. Molecular genetic identification of yeast strains isolation from Egyption soils for solubilization of inorganic phosphates and growth promotion of corn plants. J. Microbiolo. Biotechnol. 2011, 21, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, H.M. Effect of Arbuscular Mycorrhizal Fungus (Glomus Mosseae) and Soil Yeasts Interaction on Root Nodulation, N-Fixation and Growth of Faba Bean (Vichia faba). Malaysian Journal of Soil Science, 2015; 19, 157–168. [Google Scholar]
- Dastogeer KMG, Zahan MI, Tahjib-Ul-Arif M, Akter MA and Okazaki S (2020) Plant Salinity Tolerance Conferred by Arbuscular Mycorrhizal Fungi and Associated Mechanisms: A Meta-Analysis. Front. Plant Sci. 11:588550. [CrossRef]
- Singh, C.S., A. Kapoor and S.S. Wange. The enhancement of root colonization of legumes by vesicular-arbuscular mycorrhizal (VAM) fungi through the inoculation of the legume seed with commercial yeast (Saccharomyces cerevisiae). Plant Soil 1991, 131, 129–133. [Google Scholar] [CrossRef]
- Larsen, J. and I. Jacobsen. Interactions between a mycophagus Collembola, dry yeast and the external mycelium of an arbuscular mycorrhizal fungus. Mycorrhiza 1996, 6, 259–264. [Google Scholar] [CrossRef]
- Hamed, S.M. , El-Gaml, N.M. & Eissa, S.T. Integrated biofertilization using yeast with cyanobacteria on growth and productivity of wheat. Beni-Suef Univ J Basic Appl Sci 11, 112 (2022). [CrossRef]
- Ghazal FM, Moussa LAA, Fetyan NAH (2010) Cyanobacteria and Rhizobium radiobacter as possible biofertilizers in wheat production. J Agric Chemi Biotechnol 1(7):383–399. [CrossRef]
- Hussain A, Hasnain S (2011) Phytostimulation and biofertilization in wheat by cyanobacteria. J Ind Microbiol Biotechnol 38:85–92. [CrossRef]
- Chittapun S, Limbipichai S, Amnuaysin N, Boonkerd R, Charoensook M (2017) Efects of using cyanobacteria and fertilizer on growth and yield of rice, Pathum Thani I: a pot experiment. J Appl Phycol 30(1):79–85. [CrossRef]
- Singh, R. R. and K. Prasad, (2011). Effect of bio-fertilizers on growth and productivity of wheat (Triticum aestivum). J. of Farm Sci.,1(1): 1-8.
- Arif, Y. , Singh, P., Siddiqui, H., Bajguz, A., and Hayat, S. (2020). Salinity induced physiological and biochemical changes in plants: an omic approach towards salt stress tolerance. Plant Physiol. Biochem. 156, 64–77. [CrossRef]
- Acosta-Motos, J. R. , Ortuño, M. F., Bernal-Vicente, A., Diaz-Vivancos, P., Sanchez-Blanco, M. J., and Hernandez, J. A. (2017). Plant responses to salt stress: adaptive mechanisms. Agronomy 7:18. [CrossRef]
- Yang, Y. , Guo Y. Unraveling salt stress signaling in plants. J. Integr. Plant Biol. 2018;60:796–804. [CrossRef]
- Van Zelm, E. , Zhang Y., Testerink C. Salt Tolerance Mechanisms of Plants. Annu. Rev. Plant Biol. 2020;71:403–433. [CrossRef]
- Mutale-joan, C. , Rachidi, F., Mohamed, H. A., Mernissi, N. E., Aasfar, A., Barakate, M., et al. (2021). Microalgae-cyanobacteria–based biostimulant effect on salinity tolerance mechanisms, nutrient uptake, and tomato plant growth under salt stress. J. Appl. Phycol. 33, 3779–3795. [CrossRef]
- Aguégué, M. R. , Ahoyo Adjovi, N. R., Houédjofonon, E. M., Djinadou, K. A., Koda, D. A., Adoko, M. Y., et al. (2021a). Financial profitability of maize production with bio-fertilizer based on arbuscular mycorrhizal fungi native to Benin. J. Dev. Agric. Economics. 13 (1), 56–64. [CrossRef]
- Shiferaw B, Hellin J, Muricho G (2011). "Improving market access and agricultural productivity growth in Africa: what role for producer organizations and collective action institutions? Food Security 3(4):475-489. [CrossRef]
- Di-Marcantonio F, Morales-Opazo C, Barreiro-Hurlé J, Demeke M (2014). Determinants of food production in Sub Saharan Africa: The impact of policy, market access and governance. No. 727:2016- 50321. [CrossRef]
- Ahmed. E., J. Sulaiman and S. Mohd. Wheat Production and Economics. Am. J. Agric. Biol. Sci. 2011, 6, 332–338. [Google Scholar] [CrossRef]
- Soni, S. N. Economics of Technical Change of Wheat Production in Sagar District. M. P. Crop-Research-Hisar. 2000, 19, 452–456. [Google Scholar]
- Habib, W. Z. and I, Iskandar and A. Aziz. Economic efficiency of orange production in Syria. Damascus University Journal for Agricultural Sciences 2013, 29, 375–391. [Google Scholar]
- Maoba, S. (2016) Farmers’ Perception of Agricultural Extension Service Delivery in Germiston Region, Gauteng Province, South Africa. South African Journal of Agricultural Extension, 44, 167-173. [CrossRef]
Figure 1.
Field Experiment Site in the Northeast Nile Delta, Sharkia Governorate, Egypt: Landsat 8 Images (June 19, 2021).
Figure 1.
Field Experiment Site in the Northeast Nile Delta, Sharkia Governorate, Egypt: Landsat 8 Images (June 19, 2021).
Figure 2.
Effect of Microorganisms and K-humate on Faba Bean and Wheat Seedlings after 30 Days of Germination.
Figure 2.
Effect of Microorganisms and K-humate on Faba Bean and Wheat Seedlings after 30 Days of Germination.

Figure 3.
Impact of microbiomes inoculation and k-humate on plant antioxidant enzymes, rhizosphere enzymes, shoot and root lengths of 30-day faba bean seedlings.
Figure 3.
Impact of microbiomes inoculation and k-humate on plant antioxidant enzymes, rhizosphere enzymes, shoot and root lengths of 30-day faba bean seedlings.
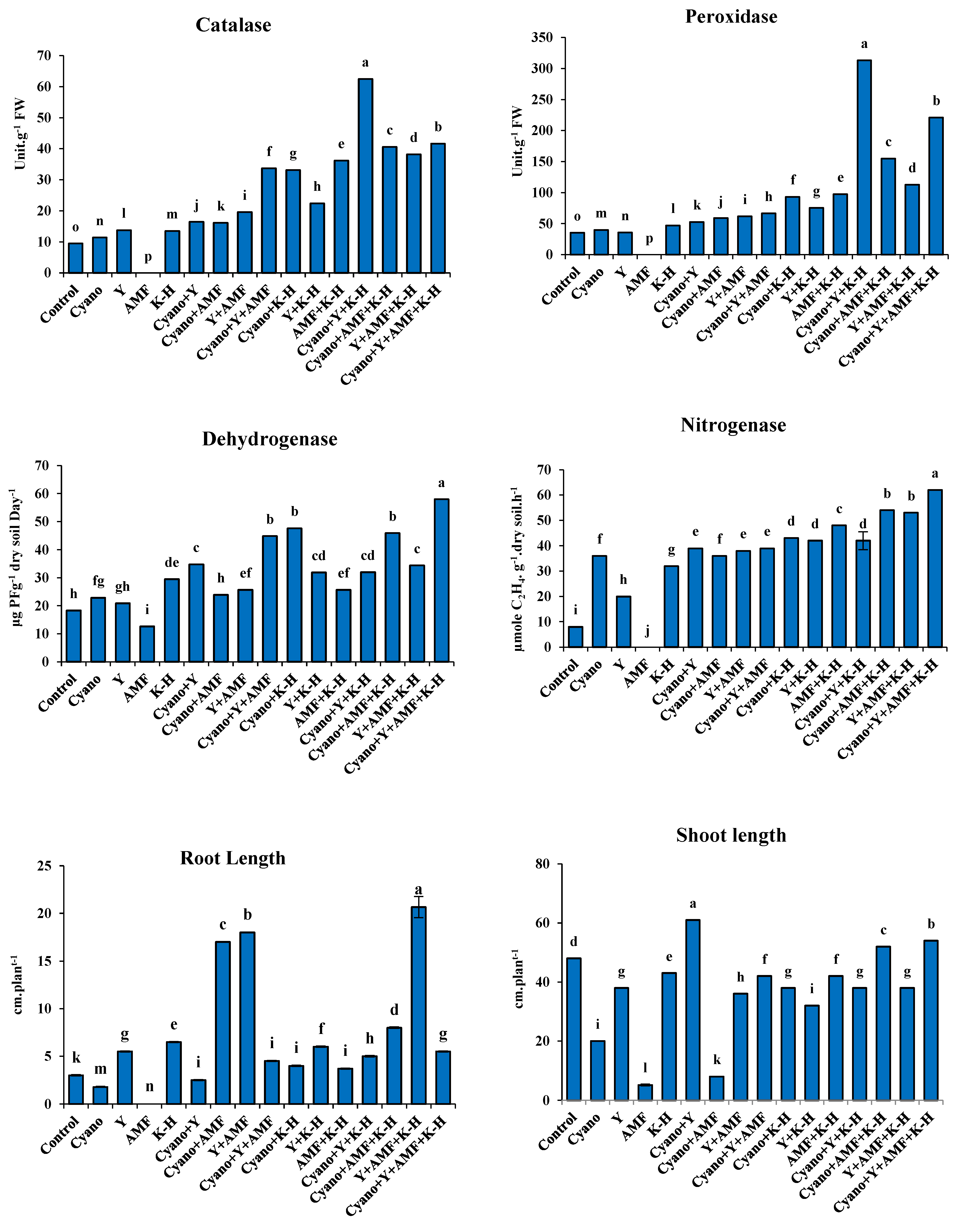
Figure 4.
Influence of treatments on K/Na ratio and percentages of protein, N, P, K, and Na in faba bean seeds.
Figure 4.
Influence of treatments on K/Na ratio and percentages of protein, N, P, K, and Na in faba bean seeds.
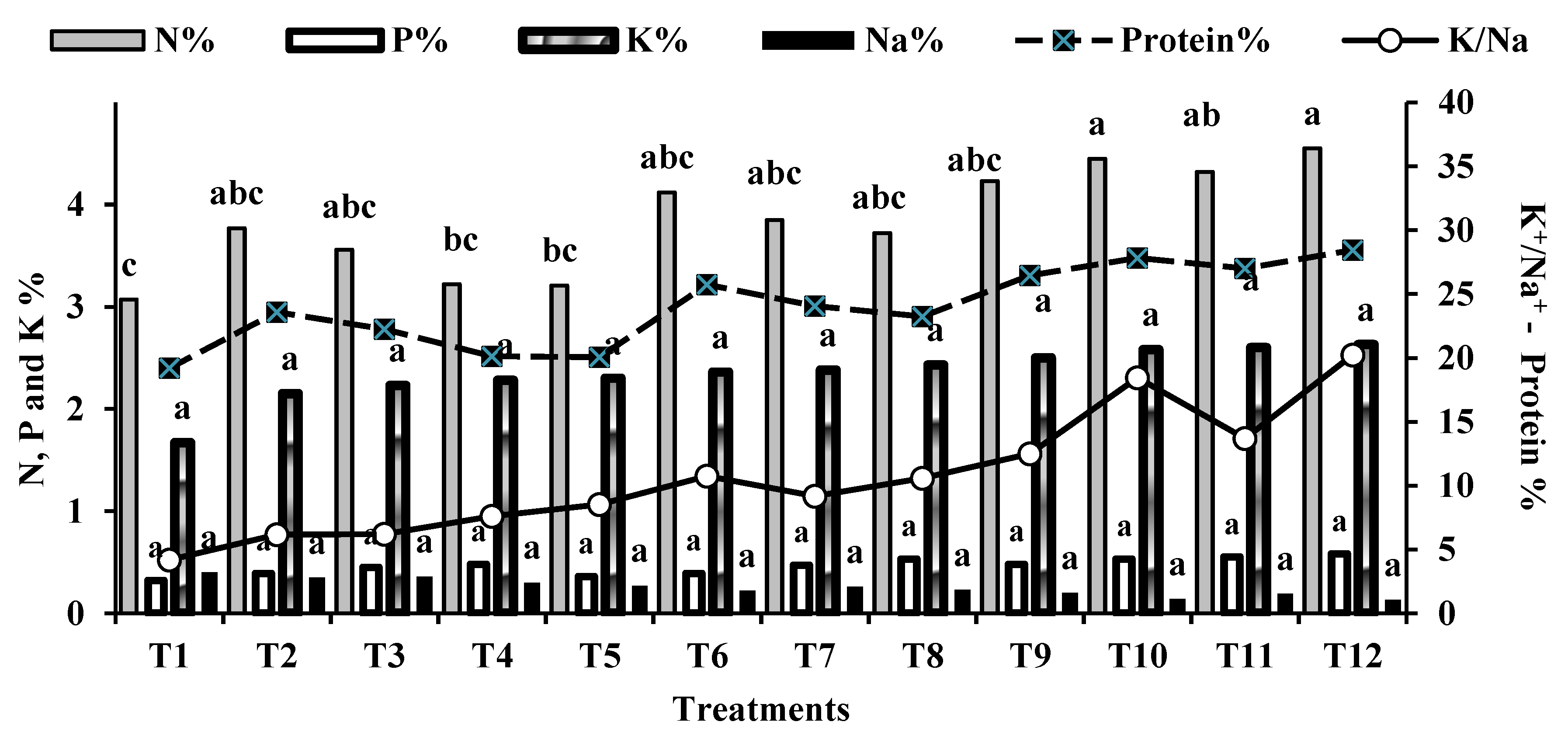
Figure 5.
Influence of treatments on K/Na ratio and percentages of Protein, N, P, K, and Na in wheat grains.
Figure 5.
Influence of treatments on K/Na ratio and percentages of Protein, N, P, K, and Na in wheat grains.
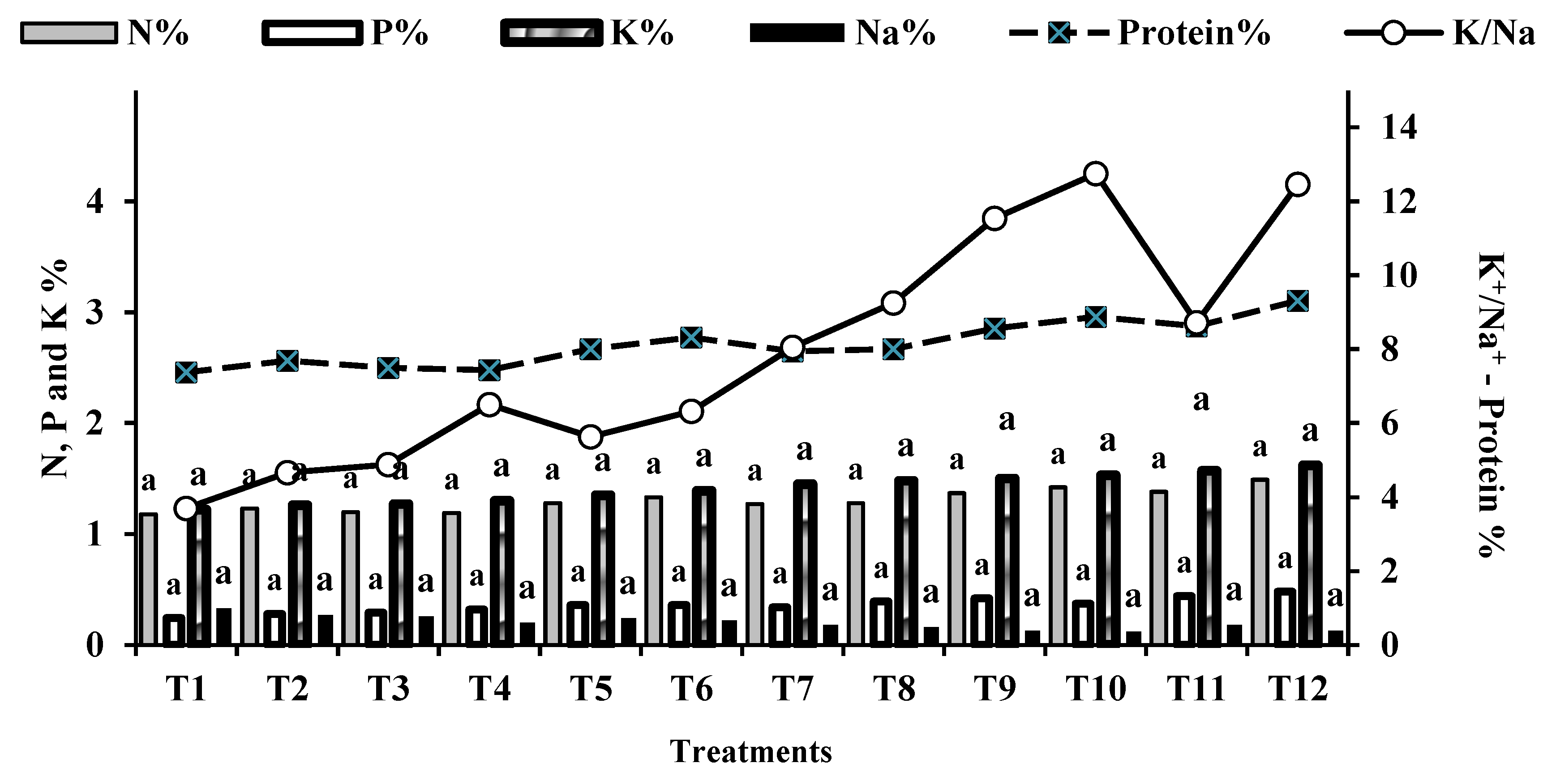

Table 1.
GenBank accession number and some characterization parameters of different cyanobacteria and green microalgae species under study.
Table 1.
GenBank accession number and some characterization parameters of different cyanobacteria and green microalgae species under study.
| Ser. | Microalgal strains | NCBI* Accession no. | Family | pH | Optical density at 560 nm | Total chlorophyll (mg.l-1) |
Dry weight (mg.l-1) |
|---|---|---|---|---|---|---|---|
| 1 | Nostoc muscorum isolate HSSASE1 | KT277784.1 | Nostocaceae | 8.11 | 1.19 | 5.26 | 760.96 |
| 2 | Spirulina platensis isolate HSSASE5 | KT277788.1 | Spirulinaceae | 10.16 | 2.77 | 11.63 | 1772.80 |
| 3 | Anabaena oryzae isolate HSSASE6 | KT277789.1 | Nostocaceae | 7.14 | 0.87 | 4.03 | 557.76 |
| 4 | Wollea saccata isolate HSSASE7 | KT277790.1 | Nostocaceae | 6.82 | 2.40 | 9.82 | 1532.80 |
| 5 | Phormidium fragilis isolate HSSASE9 | KT277792.1 | Phormidiaceae | 8.67 | 2.09 | 3.00 | 1334.40 |
| 6 | Anabaena sp. HSSASE11 | KT277794.1 | Nostocaceae | 8.05 | 1.67 | 7.56 | 1065.60 |
* National Center for Biotechnology Information (NCBI).
Table 2.
Soil Mechanical and Chemical Analyses.
| Coarse sand (%) | Fin sand (%) | Silt(%) | Clay (%) | Texture | O.M(%) | CaCO3 (%) | |||||||
| 3.14 | 8.29 | 28.76 | 59.81 | Clay | 0.48 | 5.19 | |||||||
| pH (1:2.5) | EC(dS.m-1) | Cations (meq/l) | Anions (meq/l) | ||||||||||
| Ca++ | Mg++ | Na+ | K+ | HCO-3 | Cl- | SO--4 | |||||||
| 8.25 | 18.57 | 12.46 | 21.73 | 150 | 0.76 | 8.25 | 132 | 44.70 | |||||
| Macronutrients (ppm) | Micronutrients (ppm) | ||||||||||||
| N | P | K | Fe | Mn | Zn | Cu | |||||||
| 37 | 5.67 | 189 | 1.37 | 3.25 | 0.73 | 0.048 | |||||||
Table 3.
Physical and Chemical Characterization of El Salam Canal.
| pH | EC (dS.m-1) | Cations (meq/l) | Anions (meq/l) | R.S.C. | SAR | ||||||||
| Ca++ | Mg++ | Na+ | K+ | HCO-3 | CO--3 | Cl- | SO--4 | 14.64 | 26.25 | ||||
| 9.00 | 4.94 | 2.12 | 2.29 | 39.00 | 2.40 | 12.45 | 6.60 | 16.27 | 10.49 | ||||
Table 6.
Influence of bio-inoculation and K-humate on AMF infection rate and spore densities in the rhizosphere of wheat and faba bean after 75-day field experiment.
Table 6.
Influence of bio-inoculation and K-humate on AMF infection rate and spore densities in the rhizosphere of wheat and faba bean after 75-day field experiment.
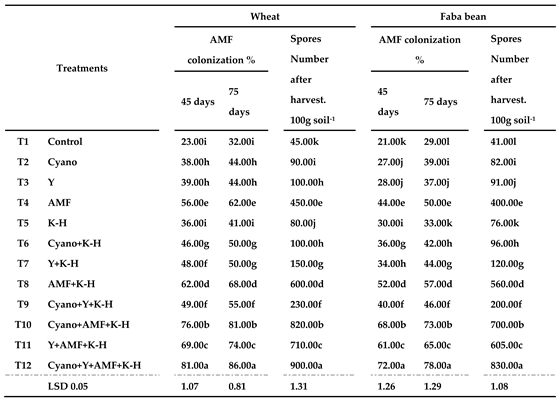 |
Table 7.
Soil biological activity as affected by cyanobacteria, yeast, AMF inoculation, and k-humate after 75 days of wheat and faba bean growth.
Table 7.
Soil biological activity as affected by cyanobacteria, yeast, AMF inoculation, and k-humate after 75 days of wheat and faba bean growth.
| Treatments | Wheat | Faba bean | ||||||||||
|
Cyano count c.f.u.* 10-3.g-1 dry soil |
Bacteria count c.f.u.* 10-6.g-1 dry soil |
Yeast count c.f.u.* 10-3.g-1 dry soil |
N-ase µmole C2H4. g-1 dry soil.h-1 |
DHA-ase μgTPF. g-1 dry soil.day-1 |
Cyano count c.f.u.* 10-3.g-1 dry soil |
Bacteria count c.f.u.* 10-6.g-1 dry soil |
Yeast count c.f.u.* 10-3.g-1 dry soil |
N-ase µmole C2H4. g-1 dry soil.h-1 |
DHA-ase μgTPF. g-1 dry soil.day-1 |
|||
| T1 | 10.00j | 21.50k | 1.00k | 0.85f | 5.29g | 15.02jk | 108.00j | 2.00l | 3.46gh | 4.67h | ||
| T2 | 36.00f | 61.50g | 3.00j | 1.60f | 14.08d | 30.00g | 250.25d | 4.00k | 1.13i | 9.33f | ||
| T3 | 17.00h | 60.00h | 7.00h | 2.70de | 3.71ghi | 24.00h | 245.25e | 5.00j | 2.84h | 7.46g | ||
| T4 | 12.00i | 26.00j | 6.00i | 0.70f | 2.87i | 14.00k | 147.33i | 7.00i | 4.8fg | 4.01h | ||
| T5 | 12.00i | 35.00i | 3.00j | 2.34de | 3.36hi | 32.00f | 108.50j | 10.00h | 8.41e | 8.95f | ||
| T6 | 38.00e | 95.50d | 9.00g | 5.89c | 11.45e | 36.00e | 207.00g | 11.00g | 12.76c | 10.85e | ||
| T7 | 23.00g | 79.00e | 12.00f | 5.52c | 7.19f | 16.00j | 250.00d | 17.00e | 17.46b | 5.01h | ||
| T8 | 18.00h | 66.00f | 13.00e | 3.12d | 4.84gh | 20.00i | 221.25f | 16.00f | 5.13f | 5.38h | ||
| T9 | 60.00c | 95.50d | 18.00c | 5.48c | 21.82b | 50.00b | 194.00h | 32.00c | 11.14d | 18.19b | ||
| T10 | 64.00b | 126.00c | 14.00d | 8.42b | 19.69c | 42.00c | 307.00b | 23.00d | 25.30a | 12.92d | ||
| T11 | 48.00d | 194.00b | 22.00b | 16.98a | 18.22c | 38.00d | 287.00c | 35.00b | 25.11a | 14.42c | ||
| T12 | 78.00a | 200.00a | 25.00a | 17.27a | 30.88a | 61.00a | 378.50a | 38.00a | 26.51a | 24.15a | ||
| LSD | 1.06 | 0.74 | 0.92 | 1.11 | 1.64 | 1.09 | 0.85 | 0.86 | 1.39 | 1.39 | ||
Table 8.
Impact of of microbiomes and K-humate on crop components and faba bean productivity.
| Treatments | Number of pods. plant-1 | Number of Seeds. plant-1 | Weight of seeds. plant-1 (g) | Weight of 100 seeds (g) | Seeds yield (ton.ha-1) | Straw yield (ton.ha-1) | Biological yield (ton.ha-1) |
Harvest index (%) | |
|---|---|---|---|---|---|---|---|---|---|
| T1 | Control | 6.83g | 16.10j | 5.83g | 31.56l | 1.21j | 1.45k | 2.66 | 0.45 |
| T2 | Cyano | 14.00d | 33.75g | 19.00c | 50.41h | 1.33g | 1.63h | 2.96 | 0.45 |
| T3 | Y | 11.00ef | 23.43i | 11.67f | 46.61j | 1.22i | 1.62i | 2.84 | 0.43 |
| T4 | AMF | 10.00f | 14.00k | 6.76g | 42.92k | 0.98k | 1.41l | 2.39 | 0.41 |
| T5 | K-H | 13.50de | 28.00h | 17.76e | 48.78i | 1.29h | 1.45j | 2.75 | 0.47 |
| T6 | Cyano+K-H | 19.73c | 55.24c | 33.50c | 61.90e | 1.42e | 1.66f | 3.08 | 0.46 |
| T7 | Y+K-H | 18.20c | 42.22f | 23.57d | 54.04g | 1.42e | 1.71e | 3.13 | 0.45 |
| T8 | AMF+K-H | 20.30c | 52.78e | 33.42c | 59.28f | 1.38f | 1.66g | 3.04 | 0.45 |
| T9 | Cyano+Y+K-H | 24.20b | 54.21d | 39.10b | 70.75c | 1.93d | 2.11d | 4.03 | 0.48 |
| T10 | Cyano+AMF+K-H | 25.74b | 57.14b | 43.87a | 75.45b | 2.36b | 2.53b | 4.89 | 0.48 |
| T11 | Y+AMF+K-H | 24.97b | 52.94e | 37.74b | 69.88d | 2.21c | 2.55a | 4.77 | 0.46 |
| T12 | Cyano+Y+AMF+K-H | 28.86a | 58.09a | 46.20a | 78.24a | 2.77a | 2.51k | 5.28 | 0.52 |
| LSD 0.05 | 2.96 | 0.65 | 3.82 | 0.59 | 0.57 | 0.72 |
Table 9.
Impact of of microbiomes and K-humate on crop components and wheat productivity.
| Treatments | Number of spike. plant-1 | Number of grains. spike-1 | Weight of 1000-grain (g) | Grain yield (ton.ha-1) | Straw yield (ton.ha-1) |
Biological yield (ton.ha-1) |
Harvest index (%) | ||
|---|---|---|---|---|---|---|---|---|---|
| T1 | Control | 18.00f | 35.57f | 55.80j | 3.07g | 4.44i | 7.51 | 0.41 | |
| T2 | Cyano | 26.00a | 60.17a | 62.30f | 3.53def | 5.28g | 8.81 | 0.40 | |
| T3 | Y | 19.00ef | 33.96g | 61.00g | 3.34efg | 4.92h | 8.26 | 0.40 | |
| T4 | AMF | 23.00b | 33.90g | 59.50h | 3.14fg | 5.02h | 8.16 | 0.39 | |
| T5 | K-H | 19.00ef | 44.89d | 60.98g | 3.43efg | 5.64ef | 9.07 | 0.38 | |
| T6 | Cyano+K-H | 22.00bc | 40.76e | 64.90e | 3.90bcd | 5.86de | 9.77 | 0.40 | |
| T7 | Y+K-H | 26.00a | 29.80h | 62.60f | 3.70cde | 5.59f | 9.31 | 0.40 | |
| T8 | AMF+K-H | 25.00a | 29.21h | 58.26i | 3.56def | 5.30g | 8.86 | 0.40 | |
| T9 | Cyano+Y+K-H | 14.00g | 34.81fg | 66.50c | 4.06bc | 6.07cd | 10.13 | 0.40 | |
| T10 | Cyano+AMF+K-H | 21.00cd | 44.20d | 68.10b | 4.10bc | 6.34ab | 10.44 | 0.39 | |
| T11 | Y+AMF+K-H | 20.00de | 48.23c | 65.80d | 4.10bc | 6.17bc | 10.27 | 0.40 | |
| T12 | Cyano+Y+AMF+K-H | 18.00f | 52.24b | 70.20a | 4.75a | 6.43a | 11.18 | 0.42 | |
| LSD 0.05 | 1.47 | 1.17 | 0.46 | 0.18 | 0.11 | ||||
Table 10.
Partial budget analysis of Faba bean.
| Items | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 |
| Seed yield (ton.ha-1) | 1.21 | 1.33 | 1.22 | 0.98 | 1.29 | 1.42 | 1.42 | 1.38 | 1.93 | 2.36 | 2.21 | 2.77 |
| Price ($.ton-1) | 640 | 640 | 640 | 640 | 640 | 640 | 640 | 640 | 640 | 640 | 640 | 640 |
| Straw yield (ton.ha-1) | 1.45 | 1.63 | 1.62 | 1.41 | 1.45 | 1.66 | 1.71 | 1.66 | 2.11 | 2.53 | 2.55 | 1.45 |
| Price ($.ton-1) | 80 | 80 | 80 | 80 | 80 | 80 | 80 | 80 | 80 | 80 | 80 | 80 |
| Revenue ($.ha-1) | 890.40 | 981.60 | 910.40 | 740.00 | 941.60 | 1041.60 | 1045.60 | 1016.00 | 1404.00 | 1712.80 | 1618.40 | 1888.80 |
| Seeds ($.ha-1) | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 | 46.54 |
| Labor ($.ha-1) | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 | 116.3 |
| Machine ($.ha-1) | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 | 193.85 |
| Menial Fertilizers ($.ha-1) | 193.85 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 | 48.46 |
| Cyanobacteria ($.ha-1) | 0 | 13.97 | 0 | 0 | 0 | 13.97 | 0 | 0 | 13.97 | 13.97 | 0 | 13.97 |
| Yeast ($.ha-1) | 0 | 0 | 6.98 | 0 | 0 | 0 | 6.98 | 0 | 6.98 | 0 | 6.98 | 6.98 |
| AMF($.ha-1) | 0 | 0 | 0 | 9.29 | 0 | 0 | 0 | 9.29 | 0 | 9.29 | 9.29 | 9.29 |
| K-humate ($.ha-1) | 0 | 0 | 0 | 0 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 |
| Pesticides ($.ha-1) | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 | 16.06 |
| Variable Cost ($.ha-1) | 566.60 | 435.18 | 428.19 | 430.50 | 430.50 | 444.47 | 437.48 | 439.79 | 451.45 | 453.76 | 446.77 | 460.74 |
| Fixed Cost ($.ha-1) | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 | 193.87 |
| Total Cost ($.ha-1) | 760.47 | 629.05 | 622.06 | 624.37 | 624.37 | 638.34 | 631.35 | 633.66 | 645.32 | 647.63 | 640.64 | 654.61 |
| Profit ($.ha-1) | 129.93 | 352.55 | 288.34 | 115.63 | 317.23 | 403.26 | 414.25 | 382.34 | 758.68 | 1065.17 | 977.76 | 1234.19 |
| Profit /cost ratio | 0.17 | 0.56 | 0.46 | 0.19 | 0.51 | 0.63 | 0.66 | 0.60 | 1.18 | 1.64 | 1.53 | 1.89 |
| Break-even yield | 1.06 | 0.87 | 0.86 | 0.87 | 0.87 | 0.89 | 0.88 | 0.88 | 0.90 | 0.90 | 0.89 | 0.91 |
| Break-even price | 285.89 | 212.52 | 219.04 | 261.24 | 227.87 | 207.25 | 201.71 | 208.44 | 159.73 | 132.44 | 134.59 | 155.12 |
Table 11.
Partial budget analysis of wheat.
| Items | T1 | T2 | T3 | T4 | T5 | T6 | T7 | T8 | T9 | T10 | T11 | T12 |
| Grain yield (ton.ha-1) | 3.07 | 3.53 | 3.34 | 3.14 | 3.43 | 3.91 | 3.72 | 3.55 | 4.06 | 4.10 | 4.10 | 4.75 |
| Price ($.ton-1) | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 | 240 |
| Straw yield (ton.ha-1) | 4.44 | 5.28 | 4.92 | 5.02 | 5.64 | 5.86 | 5.59 | 5.30 | 6.07 | 6.34 | 6.17 | 6.43 |
| Price ($.ton-1) | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 | 1100 |
| Revenue ($.ha-1) | 5621.00 | 6655.00 | 6214.00 | 6276.00 | 7027.00 | 7384.00 | 7041.80 | 6682.00 | 7651.00 | 7958.00 | 7771.00 | 8213.00 |
| Seeds ($.ha-1) | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 | 38.78 |
| Labor ($.ha-1) | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 |
| Machine ($.ha-1) | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 | 155.09 |
| Menial Fertilizers ($.ha-1) | 77.54 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 | 19.39 |
| Cyanobacteria ($.ha-1) | 0 | 13.97 | 0 | 0 | 0 | 13.97 | 0 | 0 | 13.97 | 13.97 | 0 | 13.97 |
| Yeast ($.ha-1) | 0 | 0 | 6.98 | 0 | 0 | 0 | 6.98 | 0 | 6.98 | 0 | 6.98 | 6.98 |
| AMF($.ha-1) | 0 | 0 | 0 | 9.29 | 0 | 0 | 0 | 9.29 | 0 | 9.29 | 9.29 | 9.29 |
| K-humate ($.ha-1) | 0 | 0 | 0 | 0 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 | 9.29 |
| Pesticides ($.ha-1) | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 | 13.56 |
| Variable Cost ($.ha-1) | 440.10 | 395.90 | 388.90 | 391.20 | 391.20 | 405.20 | 398.20 | 400.50 | 412.20 | 414.50 | 407.50 | 421.44 |
| Fixed Cost ($.ha-1) | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 | 232.63 |
| Total Cost ($.ha-1) | 672.69 | 628.51 | 621.52 | 623.83 | 623.83 | 637.80 | 630.81 | 633.12 | 644.78 | 647.09 | 640.10 | 654.07 |
| Profit ($.ha-1) | 4948.11 | 6026.69 | 5592.08 | 5651.77 | 6403.37 | 6746.60 | 6410.99 | 6048.88 | 7006.62 | 7310.91 | 7130.90 | 7558.93 |
| Profit /cost ratio | 7.36 | 9.59 | 9.00 | 9.06 | 10.26 | 10.58 | 10.16 | 9.55 | 10.87 | 11.30 | 11.14 | 11.56 |
| Break-even yield | 0.50 | 0.47 | 0.46 | 0.47 | 0.47 | 0.48 | 0.47 | 0.47 | 0.48 | 0.48 | 0.48 | 0.49 |
| Break-even price | 89.57 | 71.34 | 75.24 | 76.45 | 68.78 | 65.28 | 67.76 | 71.54 | 63.65 | 61.98 | 62.33 | 58.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



