
Submitted:
27 May 2024
Posted:
29 May 2024
You are already at the latest version
Abstract
Staphylococcus aureus and coagulase-negative staphylococci account for about 80% of infections associated with medical devices and are associated with increased virulence due to their ability to form biofilm. In this study, we aimed to construct a comprehensive reference map followed by significant pathway analysis in the proteome of S. aureus biofilm grown for 3-days as compared with 24 h planktonic using a high-resolution TMT based MS. We identified proteins associated with secondary metabolites, ABC transporters, biosynthesis of amino acids, response to stress, and amino sugar and nucleotide sugar metabolism were significantly upregulated in the 3-day biofilm. In contrast, proteins associated with virulence factors, microbial metabolism in diverse environments, translation, and energy metabolism were significantly downregulated. GO functional annotation indicated that more proteins are involved in metabolic processes, catalytic activity, and binding in biofilm, respectively. Among the significantly dysregulated proteins, hyaluronidase (hysA) in conjunction with chitinase may play a significant role in the elimination and/or prevention of biofilm development. This study advances the current S. aureus subproteomes, identified potential pathways significant to biofilm biology and helped to understand their potential role in S. aureus which may shed light on developing new therapeutic regimes including antibiofilm agents in the treatment of biofilm-infections related with implantable medical devices.
Keywords:
Staphylococcus aureus
; biofilms
; proteomics
; TMT-MS
; virulence factors
; biosynthetic processes
; stress responses.
1. Introduction
The Gram-positive opportunistic pathogen S. aureus represents a serious public health burden worldwide, particularly within healthcare settings where they are often associated with an increased virulence due to their ability to form biofilm. To establish infection bacteria initially have to attach to the tissue. S. aureus does this using Microbial Surface Components Recognising Adhesive Matrix Molecules abbreviated as MSCRAMMs, and Secreted Expanded-Repertoire Adhesive Molecules termed SERAMs. In addition, various types of enzymes also produced by S. aureus including exotoxins such as exfoliative toxins A and B (which increase host tissue invasion), lipases, proteases, thermonucleases, and hyaluronidases [1,2]. Planktonic cells (free-floating) generally cause acute infections by producing extracellular enzymes and secreted toxins [3]. S. aureus play significant role in chronic infections due to its biofilm development on host tissues or on implantable medical devices (e.g., prosthetic joints, catheters, breast implants, and pacemakers) [4,5,6,7,8,9] to withstand therapeutic intervention.
Biofilms are microbial communities embedded in a self-produced EPS matrix which can be found on any surface [10,11]. Although the exact composition of EPS differs between various bacterial species and environmental conditions, EPS consists mainly of polysaccharides, proteins, and extracellular DNA (eDNA) [11]. In general, biofilm development is characterised by three stages: initial attachment, biofilm maturation, and dispersal. Several studies highlighted primarily the elucidation of individual molecular variables essential in the growth of S. aureus biofilms. A very recent study by Graf et. al. (2019) mentioned some of the proteinaceous and non-proteinaceous factors responsible for various phases of biofilm formation and the synthesis and expression of these molecular factors are closely regulated by several biofilm regulators such as AgrA and RNAIII, Rot, SigB, SarA, IcaR, CodY, and others [12].
Attempts to comprehend the biochemical framework of biofilm formation and resilience have constantly demonstrated alterations in protein expression profile in S. aureus [13,14,15,16,17] compared with planktonic counterparts. However, numerous facets of complex structure and role of biofilms have yet to be elucidated.
In the present study, our goal was to construct a comprehensive proteomic reference map of S. aureus biofilm as compared with planktonic culture by employing TMT-based high-resolution MS. In addition, significant dysregulated marker proteins were identified and further characterised to better understand key proteins’ potential role(s) in S. aureus biofilm biology.
2. Materials and Methods
This research aimed to develop a quantitative proteomic analysis to delineate the differences between S. aureus cells transitioning between planktonic and biofilm lifestyles. To achieve this goal, we performed protein extraction, fractionation, reduction, alkylation, and in-solution digestion, generating samples for analysis using TMT-based MS. Each growth condition, planktonic and biofilm, was examined with three biological replicates.
2.1. Microorganism and Culture Conditions
Staphylococcus aureus reference strain (ATCC 25923) was cultured to stationary phase in 100% tryptic soy broth (TSB) for 24 h, maintaining constant agitation at 130 rpm and 37 °C. To produce 3-day biofilm, S. aureus was cultivated as previously described [18]. Briefly, Biofilm formation occurred on removable polycarbonate coupons within a Centers for Disease Control (CDC) biofilm reactor (BioSurface Technologies Corp, Bozeman, MT, USA) under batch conditions at 37 °C. Initially, 50% TSB was used for 48 h, after which the media was exchanged with 20% TSB every 48 h as needed to achieve a 3-day biofilm. Shear force was induced by baffle rotation at 130 rpm. Biofilm was cultivated and harvested across three independent experiments.
2.2. Protein Extraction and Fractionation
Protein extraction and fractionation were performed as described previously [18]. Concisely, planktonic bacteria were pooled from three separate growth of 24 h cultures of S. aureus and then mixed with a lysis buffer composed of 100 mM Triethylammonium bicarbonate (TEAB; Sigma-Aldrich, St. Louis, MO, USA) at pH 8.5, along with 1% (w/v) sodium deoxycholate (Sigma-Aldrich) at a ratio of 10 parts supernatant to 1 part lysis buffer. As for the S. aureus biofilm, growth of 3-day biofilm-coated coupons (n = 24) were washed to remove non-adherent cells, and then each coupon was placed individually in 2 mL of phosphate-buffered saline (PBS) and lysis buffer, followed by an overnight incubation with gentle shaking at 4 °C. The samples underwent probe sonication in an ice-cold environment (using Sonic Ruptor, Omni International, Kennesaw, Georgia, GA, USA) for 2 minutes at 50% power and 70% pulses. Following this, the samples were centrifuged at 12,000× g for 10 minutes, and the supernatant was then passed through a 10 kDa molecular weight cut-off (MWCO) ultra-membrane filter tube (Sigma-Aldrich) prior to another centrifugation step at 4000× g for 20 minutes. Protein samples underwent three washes with PBS to remove TSB and lysis buffer, and were subsequently concentrated using a 3 kDa MWCO filter tube (Sigma Aldrich).
The protein concentration was measured using the BCA protein assay (Thermo Fisher Scientific, Waltham, Massachusetts, USA) at a wavelength of 562 nm by following the manufacturer’s instructions, measuring absorbance.
2.3. Protein Reduction, Alkylation, and Digestion
Protein reduction, alkylation, and digestion were performed as described previously [18]. Concisely, a total of 40 µg protein was reduced (5 mM DTT, 15 min, RT), alkylated (10 mM IAA in darkness,30 min, RT) and then diluted with 100 mM TEAB pH 8.5. In-solution digestion step was carried out overnight at RT with Lys-C and trypsin at a ratio of 1:30 for 5.5 h at 37°C, respectively. Further steps of samples include: adjustment to 1% (v/v) TFA, removal of precipitated deoxycholate by centrifugation, then centrifuged at 14,100× g and desalted with 0.2% TFA washing by utilising SDB-RPS (3M-Empore) Stage Tips (Thermo Fisher Scientific). The elution of samples was carried out using 5% ammonium hydroxide in 80% acetone, followed by centrifugation at 1000× g for 5 minutes. Subsequently, the samples were vacuum dried and stored at -20°C until further processing.
2.4. TMT Labeling and High pH Fractionation
TMT labelling and high pH fractionation were performed as described previously [18]. Concisely, TMT (Thermo Fisher Scientific) reagents (0.8 mg) were dissolved in 85 µL of acetone, and 41 µL of this solution was added to the reconstituted samples (100 µL of 100 mM TEAB pH 8.5) and then incubated for 1 h at RT. Each TMT labeled sample mixed with 8 µL of hydroxylamine (5%), then incubated for 15 min at RT. A volume of 2 µL of each labelled sample was pooled, vacuum dried and then reconstituted in a 30 µL solution of FA(0.1%) (Merck, Kenilworth, NJ, USA). The mixture was centrifuged for 5 min at 14,000× g and subsequently analysed using a mass spectrometer.
Data searching was performed using Proteome Discoverer 1.3 (for detailed information, refer to the data processing section). Utilizing the normalization values derived from this search result, an equivalent number of peptides were taken from each sample, pooled, and then vacuum dried using miVac. The dried labelled sample was reconstituted in buffer A (5 mM ammonia, pH 10.5) and fractionated by high pH RP-HPLC (Agilent Technologies, Santa Clara, CA, USA). The dried labelled sample was reconstituted in buffer A. Following sample loading and a 10 min wash with 97% buffer A, the concentration of buffer B (5 mM ammonia solution with 90% acetone, pH 10.5) was ramped up from 3% to 30% over 55 min, then maintained at 70% for 10 min, and finally raised to 90% for 5 min, all at a flow rate of 300 µL/min. The eluent was collected at 2 min intervals initially, up to 16 min, and then at 1 min intervals for the rest of the gradient. The fractionated sample was divided into 19 fractions, dried using miVac, and subsequently resuspended in 55 µL of FA (0.1%) for MS analysis.
2.5. Nanoflow LC-ESI-MS/MS
The following steps were performed as previously described [18].
2.5.1. Nanoflow LC-ESI-MS/MS Using Orbitrap Elite
Data acquisition was performed using an Orbitrap Elite mass spectrometer (Thermo Fisher Scientific) coupled with a PicoView 550 Nanospray Source (New Objectives) and an Eksigent UPLC system (AB SCIEX) comprising an ekspert™ nanoLC 425 UPLC pump and ekspert™ nanoLC 400 autosampler (Thermo Fisher Scientific) was utilized for the experiment. Each fraction, totaling 20 µL, was loaded onto a self-packed trap column measuring 100 µm × 3.5 cm with Halo® 2.7 µm 160 Å ES-C18 (Advanced Materials Technology, Wilmington, DE, USA). The desalting process was conducted using a loading buffer [0.1% FA] at a flow rate of 4 µL/min for a duration of 10 min. The elution of peptides was achieved through linear gradients of mobile phase A (0.1% FA/5% DMSO) and mobile phase B (0.1% FA/5% DMSO). The gradient initiated with phase B (1-10%, 0.1 min), B (10–20%, 52 min), B (20–32%, 48 min) followed by (32–43%, 20 min) at a flow rate of 450 nL/min throughout the gradient. Before reaching the analytical column, the eluent from the trap underwent dilution with buffer A at a flow rate of 100 nL/min. Subsequently, the peptides underwent refocusing and separation on the analytical column maintained at 60 °C. Peptides were ionized by electrospray ionization, and data-dependent MS/MS acquisition was performed by utilising an Orbitrap Elite (Thermo Fisher Scientific) comprising 1 full MS1 (R = 120 K) scan acquisition from 380 to 1600 m/z, and 15 HCD type MS2 scans (R = 30 K).
2.5.2. Nanoflow LC-ESI-MS/MS Using Q Exactive
Data acquisition was performed using a Q Exactive (Thermo Fisher Scientific) Mass Spectrometer equipped with Nano spray Source and Easy nLC 1000 (Thermo Fisher Scientific). Each fraction, totaling 10 µL, was loaded onto a self-packed trap column measuring 100 µm × 3.5 cm with Halo® 2.7 µm 160 Å ES-C18 (Advanced Materials Technology, Wilmington, DE, USA). The desalting process was conducted using a loading buffer [0.1% FA] followed by peptides were eluted with the linear gradients of mobile phase A (0.1% FA) and buffer B [100%(v/v) Acetone, 0.1%(v/v) FA]. The gradient initiated with (1-30%, 110 min), B (30–85%, 2 min) followed by B (85%, 8 min) with a flow rate of 300 nL/min throughout the gradient. Peptides were ionized by electrospray ionization and data-dependent MS/MS acquisition was performed by utilising a Q-Exactive consisting of 1 full MS1 (R = 70 K) scan acquisition from 350 to 1850 m/z, and 10 HCD type MS2 scans (R = 70 K).
2.6. Database Search, Statistical Analysis, and Bioinformatics
Database search, statistical analysis, and bioinformatics were performed according to the recent publication [18]. Concisely, the raw data files were submitted to Proteome Discoverer (v 1.3, Thermo Fisher Scientific) and the processing of the data was conducted using Sequest and Mascot (Matrix Science, London, UK) against the S. aureus reference strain (ATCC 25923) sourced from Genbank CP009361 and CP009362. Protein identification utilised the following parameters: peptide mass tolerance set at 10 ppm; MS/MS tolerance at 0.1 Da; enzyme = trypsin, missed cleavage = 1; fixed modification, carbamidomethyl (C), TMT10-plex (K) and TMT10-plex (N-term); variable modification, oxidation (M), Deamination (N, Q) and Acetylation (N-Terminus). Quantification was carried out as per the peak intensities of reporter ions in the MS/MS spectra. Peptide identification utilised a threshold of less than 1% false discovery rate. Protein quantification was determined by the total intensity of the assigned peptides. Following the extraction of protein ratios through Proteome Discoverer, further processing and statistical analyses were conducted utilising the TMTPrePro R package. Afterwards, the BLAST search was executed using highly annotated strains S. aureus N315 and S. aureus COL. In the context of biofilm growth versus planktonic growth, proteins were deemed upregulated when the TMT ratio > 1.5, while those with a TMT ratio < 0.66 were considered downregulated with a significant p-value < 0.05. Proteins exhibiting significant differential expression (> 2-fold) were identified using VENNY (v.2.1) (http://bioinfogp.cnb.csic.es/tools/venny/, accessed on June 25, 2018) and subjected to further processing to obtain deeper functional insights. The functional pathways of the significantly identified proteins were analysed utilising the KEGG mapper (https://www.genome.jp/kegg/tool/map_pathway2.html, accessed on August 6, 2018). Subcellular localisation of the significantly identified proteins was assessed using PSORTb (v 3.0.2) (http://www.psort.org/psortb/index.html, accessed on 23 January 2018). The PPI network of the significantly differentially regulated proteins was examined using STRING-db (v 10.0) (http://string-db.org/, accessed on 25 June 2018). Identification of virulence factors was performed using the Virulence Factors of Pathogenic Bacteria Database (VFDB) (http://www.mgc.ac.cn/VFs/).
2.7. Validation of TMT Data with qPCR Results
qPCR was performed to validate TMT data as described previously [18]. To validate the expression variances between planktonic and biofilm states, we selected five genes: Hyaluronidase HysA, Chitinase SA0914, Glutamyl endopeptidase sspA, Pyruvate carboxylase pyc, and Succinate dehydrogenase sdhB as targets to analyse the levels of RNA expression. The 16S rRNA served as the internal control for data normalisation and compared the differential expression levels of the five genes between planktonic and 3-day biofilm.
RNA extraction from S. aureus planktonic and 3-day biofilm samples was carried out using a RNeasy Mini Kit (Qiagen, Hilden, Germany). To prevent degradation, RNAlater was utilised, and RNase-free DNase treatment was employed to eliminate genomic DNA. The RNA concentration was measured by absorbance at 260 nm, while its quality was evaluated based on the absorbance ratio (A260/A280). A total of 200 ng of RNA was used for cDNA synthesis, facilitated by the SuperScript™ IV VILO™ Master Mix (Thermo Fisher Scientific). The real-time (RT)-PCR primers utilized in this study (Table S1) were designed based on the genome sequences of S. aureus (ATCC 25923), accessible through Genbank accession numbers CP009361 and CP009362.
Quantitative real-time PCR (qPCR) was performed using an Applied Biosystems quantitative real-time PCR machine (ViiA™ 7 qPCR, ThermoFisher Scientific). Each experiment was conducted in duplicate using two biological replicates. The qPCR reaction mix, totaling 25 μL, comprised 12.5 μL of 2X PowerUp™ SYBR™ Green Master Mix (Thermo Fisher Scientific), 1 μL each of 10 μM reverse and forward primers (resulting in a final primer concentration of 400 nM), 8.5 μL of water, and 2 μL of 1:5 diluted cDNA. Each PCR run included a no template control (NTC) and a no reverse transcription control (no RT control). The NTC contained all PCR components except the cDNA template, replaced by nuclease free water. The no RT control involved the cDNA sample without the reverse transcriptase enzyme to check for contaminating genomic DNA in the RNA. The cycling conditions for RT-PCR commenced with an initial activation step at 95°C for 10 min to activate the polymerase. This was followed by 40 cycles comprising denaturation at 95°C for 15 s, annealing at either 50°C or 55°C for 40 s, and extension at 72°C for 30 s, or alternatively, annealing and extension at 60°C for 1 min.
Initially, the expressed copy number of each target gene was normalised to the copy number of 16S rRNA within the same growth condition. Subsequently, the levels of candidate gene expression in planktonic and 3-day biofilm were compared to investigate relative gene expression, employing a previously described method [19]. The ratios derived from the qPCR results were obtained by comparing them with the planktonic in 3-day biofilm.
3. Results
3.1. TMT Identification of Differentially Regulated Proteins in the Biofilm
In this study, a total of 1636 non-redundant proteins, each with at least one (1) unique peptide and less than 1% false discovery rate, were identified and quantified. Of these, 273 proteins were significantly (p <0.05) differentially regulated exclusive proteins (DREPs) greater than 2-fold change during biofilm growth in comparison to planktonic bacteria. These 273 DREPs could be regarded as potential variables responsible for difference in the proteome of biofilm compared with planktonic growth. Of these, 82 proteins were upregulated (Table S2) and 191 were downregulated in the biofilm (Table S3). Of the upregulated biofilm proteins 35 proteins (42.7%) were associated with recognised protein pathways. Similarly, 74 (38.7%) of the downregulated biofilm proteins were associated with recognised protein pathways. Among the DREPs, A total of 34 hypothetical proteins were identified and hypothetical protein KQ76_04110 had the highest differential expression being upregulated 5.09-fold.A molybdenum ABC transporter permease (4.68-fold upregulated) and transport of molybdenum via ABC transporter into the cells was identified as essential for bacterial growth [20]. Further, we identified DNA-directed RNA polymerase subunit omega (4.50-fold) which is involved in RNA polymerase, pyrimidine and purine metabolism, glycosyltransferase (4.32-fold), and branched-chain amino acid transporter II carrier protein (4.04-fold).
The most downregulated protein was delta-hemolysin (-34.02-fold) which is involved in quorum sensing. Delta-hemolysin is a small amphipathic membrane-damaging virulence factor protein and showed potential antimicrobial activity. Delta-hemolysin may act either by binding to the surface of the cell and aggregating to form transmembrane pores, thus destabilising the cytoplasmic membrane by affecting the membrane curvature, or function as a detergent by solubilising the membrane at higher concentration [21].
The protein PotD, involved in the ABC transporter pathway, was downregulated by 22.02-fold. PotD binds external polyamines like spermidine, crucial for creating a more compact protein structure and promoting cell growth. The reduced levels of this protein may hinder cell dispersal and promote biofilm formation.
PotD negatively regulates the spermidine-preferential uptake system transcription of the operon, thereby decrease in spermidine uptake activity leads to increased polyamines accumulation in cells [22,23]. In addition, extracellular proteins including alpha-hemolysin (-13.82-fold), phosphodiesterase (-12.33-fold), cysteine protease (-11.44-fold), and transglycosylase (-10.14-fold) were also downregulated. Several ribosomal proteins were also downregulated (Table S3). In addition, we identified 20 virulence factor proteins using VFDB among the DREPs (Table 1).
3.2. GO Analysis and Annotation of Differentially Regulated Proteins in the Biofilm
We performed GO functional annotation for all DREPs. PANTHER assessment showed the involvement of 12 distinct classes of proteins in the entire global repository of proteins (Figure 1). Nucleic acid-binding proteins (20.7%), hydrolases and transferases (17.2%), ligases (13.8%), transcription factors (8.6%) and oxidoreductases (6.9%) were the most prominent classes.
The molecular function categories by PANTHER revealed the seven most represented molecular functions (Figure 1). In which, maximum number of proteins were involved in catalytic activity (50.7%) followed by binding (31.9%), and structural molecule activity (5.8%). Upon assessing biological processes, we identified the seven most represented biological processes (Figure 1). Of these, metabolic processes are the most prevalent biological processes, representing 60.5% of the protein repository followed by cellular component organisation or biogenesis (19.7%), biological regulations (9.2%), and cellular processes (3.9%).
3.3. Significantly Dysregulated Proteins and Pathway Analysis in the Biofilm
We analysed the TMT results using KEGG pathways to establish pathways impacted by bacterial biofilm formation in S. aureus. We annotated 289 DREPs using the KEGG database, with all mapping onto 113 recognised pathways. Among these, 35 out of 82 significantly upregulated proteins were involved in recognised pathways. The 35 exclusively upregulated proteins were mainly involved in biosynthesis of secondary metabolites, biosynthesis of amino acids, microbial biosynthesis of antibiotics, metabolism in diverse environments, ABC transporters, alanine, aspartate and glutamate metabolism, amino sugar and nucleotide sugar metabolism, purine metabolism, ribosome, pyrimidine metabolism, arginine biosynthesis, tyrosine and tryptophan biosynthesis etc. (Figure 2). In addition, upregulated proteins were also found to be involved in energy metabolisms such as glycolysis and galactose, and synthesis of cell-wall components such as peptidoglycan biosynthesis.
On the other hand, 74 out of 191 significantly downregulated proteins were involved in recognised pathways. KEGG pathway analysis revealed that among the 74 exclusively downregulated proteins these were mainly involved in quorum sensing, citrate cycle (TCA cycle), carbon metabolism, pyruvate metabolism, aminoacyl-tRNA biosynthesis, cationic antimicrobial peptide (CAMP) resistance, methane metabolism, glycerophospholipid metabolism, and two-component system, etc. (Figure 2).
3.4. Protein-Protein Interaction (PPI) Analysis in the Biofilm
We established protein-protein interaction (PPI) networks using STRING software to assess the network of those proteins identified exclusively to be expressed differentially in the biofilm growth mode. In this PPI analysis, all predicted interactions tagged as “high-confidence” (≥0.7) and omitted nodes that are not connected in the network in STRING software. Among the 289 DREPs, 129 nodes (proteins) and 145 edges (interactions) formed the final network. (Figure S1).
PPI network revealed that one protein symbolises a majority of connections: 30S ribosomal protein S5 (rpsE). This protein was found to be upregulated in the biofilm growth mode and could connect with 39 other proteins (Figure S1). It plays an important role in translational accuracy. Among these 39 possible connections, most of them are involved in metabolic, catalytic activity, and binding (such as ion, nucleic acid, metal, and drug). The findings of this subnetwork are consistent with the top protein classes and biological processes identified through PANTHER analysis (Figure 1). Further relevant subnetworks consist of nodes associated with gene expression, translation, ATP biosynthesis, virulence factors, glucose metabolism, and stress-response (Figure S1).
3.5. Validation of TMT Data with Real-Time qPCR
The qPCR results ratios were derived by comparing them with the planktonic state in the 3-day biofilm. Individual normalised qPCR results are demonstrated in Table S4. The results of both upregulated and downregulated protein and gene expressions were measured in terms of fold change (FC) (Table S4). The qPCR results indicated that the ratios of the levels of gene expression were partially consistent with the data acquired from the TMT-based MS analysis (Table S4).
4. Discussion
Whilst the proteomics of S. aureus biofilm have previously been investigated [13,14,16,24], we have utilised highly powerful TMT-MS in this study. TMT-labelling combined with tandem MS can label and analyse up to 10 protein samples simultaneously in high-resolution in the low mass region [25,26]. This powerful proteomic strategy can be helpful for a deeper understanding of biological mechanisms as well as a screening of biomarkers by examining the variations in protein expression levels.
In our proteomics data, we identified several proteins associated with transporters, mostly ABC transporters (Table S2, Figure 2) as uniquely upregulated in the S. aureus biofilm state: molybdenum ABC transporter permease (4.68 fold), peptide ABC transporter substrate-binding protein (2.91 fold), spermidine/putrescine ABC transporter ATP-binding protein potA (2.39 fold), heme ABC transporter ATP-binding protein (2.32 fold), and glutamine ABC transporter ATP-binding protein (2.05 fold). Proteins were also exclusively downregulated in biofilm growth and included ABC transporter ATP-binding protein encoded by vga and iron ABC transporter substrate-binding protein encoded by SA0691. ATP-binding cassette transporters (ABC transporters) are members of a superfamily of proteins, that are transmembrane proteins which are linked with adenosine triphosphate (ATP) binding energy utilisation. They play substantial functions in molecular (macro and micro) uptake of nutrients, such as capsular polysaccharides, small molecule inhibitors, amino acids, lipids, and vitamins. To better understand of virulence and drug resistance, microbial ABC transporters are gaining attention as a potential target [27]. In previous studies, ABC transporters (such as ABC transporter lipoprotein, ABC transporter permease protein, ABC transporter periplasmic amino acid-binding protein, and ABC transporter ATP-binding protein) have been reported to be upregulated in biofilm formation in S. aureus [14,28,29,30] and in numerous other bacteria, including Cronobacter sp., Streptococcus uberis, Rhizobium leguminosarum, Pseudomonas fluorescens and Bacillus subtilis [31,32,33,34,35], but have also been reported to be downregulated (putative ABC transporter permease) in Listeria monocytogenes [36], and ABC transporter ATP-binding protein in S. aureus [37]. The specific role of the ABC transporters (i.e., up- or down-regulation) depends on the supplied substrates. The downregulation of ABC transporters may be due to the lower metabolic rate of the biofilm, reducing the need to transport ATP. A study by Brady et al. (2006) revealed that the upregulation of a membrane-bound ABC transporter protein in S. aureus biofilm growth and suggested that it may be an excellent vaccine candidate, as previous work reported it as immunogenic in S. aureus infections in humans [30,38]. The unique ABC transporter proteins, particularly those that are membrane-bound, identified in our study may play a crucial role in biofilm formation. These proteins could potentially serve as marker proteins, vaccine targets, and antimicrobial targets for biofilm-related infections.
Among the significant differentially regulated proteins in the biofilm extractomes, we identified most of the extracellular or cell-wall associated proteins to be primarily represented by virulence factors (Table 1). Proteins exclusively upregulated include fibrinogen-binding protein (SA1000), hypothetical protein KQ76_08475 (SA1452), hyaluronate lyase (hysA), and coagulase, while downregulated proteins (Table S3, Figure 2) include hemolysins (hld, SA1007, hlgCAB), proteases (sspABP, splCEF, SA1121, clpP), nucleases (nuc, rnhC, SA1526, cbf1, rnz), peptidases (lytM, SA0205, SA0620, sspA), lipases (lip1, lip 2), a chitinase (SA0914), a phenol soluble modulin (SACOL1186), fibronectin-binding protein (fnbA), and adhesin (sasF). Among the upregulated proteins, fibrinogen-binding protein is an MSCRAMM, vital for the attachment of S. aureus to human cells and thus for the spread of infections [39,40]. A recent in vitro study by Kot et al. (2018), demonstrated that the expression levels of fibrinogen-binding protein in weakly attaching strain of S. aureus was considerably smaller than in strongly attaching strain of S. aureus [41]. Studies by Resch et al. (2006), reported the upregulation of fibrinogen-binding protein in biofilm growth mode compared with planktonic which shows a similar trend with our study. In an in vivo rat model of central venous catheter infection using S. epidermidis, rat lacking fibrinogen-binding motif observed more robust biofilm on the catheter, indicating its significance in the in vivo biofilm development [42]. In addition, binding of S. aureus to fibrinogen-binding protein and coagulase demonstrates various evasive responses that protect bacteria against the immune system, and its binding is influenced by Rot and Agr mediated regulatory systems [16,42].
Hyaluronidase (hysA) an extracellular enzyme exclusively upregulated in biofilm state and play an important role in disseminating recognised biofilms by the degradation of hyaluronic acid (HA) (Figure 3). HA is an extracellular matrix component and revealed to enhance biofilm development in Gram-positive pathogens, including Streptococcus intermedius, and Streptococcus pneumoniae. A very recent in-depth study by Ibberson et al. (2016) demonstrated that S. aureus integrates HA into the biofilm matrix both in vivo (murine implant-associated infection model) and in vitro, and HysA acts as a spreading factor by dispersing the biofilm and disseminating to new locations of infection [43]. On the other hand, among the exclusively downregulated proteins, chitinase (SA0914) an exo-enzyme involved in quorum sensing that prevents the initial stage development of biofilms. Interestingly, HA is the structural constituent of N-acetyl-D-glucosamine (Figure 3) which can be hydrolysed by chitinase [44]. Therefore, we can speculate that hysA in conjunction with chitinase may play significant role in the elimination and/or prevention of biofilm development.
Further, the significant downregulation of virulence-related and cell wall proteins showed that the bacteria adapted to the diverse biofilm condition by reducing some less essential roles such as adhesion, invasion, and virulence. For example, agr quorum-sensing system regulates the expression of virulence genes and contributes to the dispersal and structuring of biofilms by regulating extracellular proteases (e.g., sspAB) and phenol-soluble modulin (PSMs) surfactant peptides [45,46]. Further, Staphylococcal accessory regulator (SarA) is a positive biofilm regulator through the downregulation of extracellular nuclease (nuc) and proteases [47]. Downregulation of these genes from our findings shows similarity with the findings of Resch et al. [28]. Studies have shown that S. aureus produces proteases which in most cases act as a virulence factor that may influence the chronicity of S. aureus infections [48]. In vivo the inflammatory response also contributes to tissue destruction by continually recruiting proinflammatory cells such as lymphocytes and macrophages, releasing proteases and inflammatory mediators [8]. Although proteases help dislodge biofilms, they also harm ordinary and curative tissues, whereas macrophages may form a fibrous capsule around the implants [49].
Further pathway analysis revealed that, the upregulation of glyceraldehyde-3-phosphate dehydrogenase encoded by gapA1 (2.34 fold), cystathionine gamma-synthase encoded by metB (2.33 fold), threonine synthase encoded by thrC (2.27 fold), argininosuccinate lyase encoded by argH (2.21 fold), acetolactate synthase encoded by alsS (2.21 fold), argininosuccinate synthase encoded by argG (2.15 fold), 3-phosphoshikimate 1-carboxyvinyltransferase encoded by aroA (2.09 fold), and histidinol-phosphate aminotransferase encoded by SACOL2701 (2.01 fold) involved in biosynthesis of amino acid (Table S2, Figure 2). Besides protein parts, amino acids function as signals for gene expression molecules and regulators. In the meantime, changes in the metabolism of amino acids contribute to the development of biofilms catheter infection, both in vitro and in vivo [50,51]. Studies by Ammons et al. (2014) reported that, in addition to the diverse role of amino acids in biofilm development, they also involved in substantial energy expenditure for adequate redox equilibrium maintenance, cell-wall synthesis components, and deposition of EPS matrix [52]. Notably, in our biofilm extractomes, we found exclusively upregulated proteins involved in amino sugar and nucleotide sugar metabolism (such as glmS, nanE, capG) which is linked with peptidoglycan biosynthesis (Figure 4). As we know, peptidoglycan is the major component of the bacterial cell-wall, and our study also observed significant accumulation of peptidoglycan biosynthesis associated protein (e.g., murA). Therefore, we can speculate that the proper utilisation of amino acids will stimulate cell-wall formation leading to EPS matrix deposition and enhance biofilm formation.
Many ribosomal subunit proteins such as 30S ribosomal protein S14 (rpsN), 50S ribosomal protein L27 (rpmA), 30S ribosomal protein S5 (rpsE), 50S ribosomal protein L10 (rplJ) were exclusively upregulated (Table S2, Figure 2) under biofilm growth state, while 50S ribosomal protein L17 (rplQ) and 50S ribosomal protein L20 (rplT) were downregulated (Table S3, Figure 2). Usually, ribosomal subunit proteins play a significant role in regulating the expression of whole proteins. 50S involves the activity that catalyses the formation of peptide bonds, protects premature polypeptide hydrolysis, and helps to fold proteins after synthesis, etc. Synthesising some peptides or proteins helps to promote resistance. For example, 50S ribosomal protein L27 (rpmA) plays a critical role in tRNA substrate stabilisation during the peptidyl transfer reaction as well as ribosome assembly and catalysis even with certain level of stress environment (e.g., deletion of some part) [53].
Among the significantly differentially regulated proteins, we identified several proteins related to different stress responses in the S. aureus biofilm extractomes: DNA-directed RNA polymerase subunit omega (RpoZ), dehydrogenases (e.g., bfmBAA, gap, ldhD), oxidoreductases (e.g., guaC, SACOL0959, SA0558, nfrA), reductases (e.g., SACOL1543, SA0759, SACOL1768, trxB), glutathione S-transferase, and heat shock protein GrpE (Table S3 and Table 1). The formation of a stress response is a significant characteristic of biofilm life cycle as it leads to changes in many gene expressions which increase antimicrobial resistance and is generally regulated by alternative RNA polymerase sigma factor B (SigB). Multiples studies have reported increased or decreased expression of stress response associated proteins in S. aureus biofilm [14,16,54] and other bacterial [55,56,57]. However, notably, we identified a unique DNA-directed RNA polymerase subunit omega (RpoZ) which is 4.52-fold upregulated in the biofilm. Even though very little is known about RpoZ, a very recent study reported its significant roles in stability, complex assembly, maintenance of transcriptional integrity, and cellular physiology in response to stress in S. aureus biofilm [54]. Another protein glyceraldehyde-3-phosphate dehydrogenase (Gap) was exclusively upregulated in biofilm growth mode and under oxidative stress environments, and showed a significant positive correlation between development, ATP level and Gap activity in planktonic S. aureus [58]. Pathway analysis revealed that the Gap, an enzyme involved in multiple pathways (such as biosynthesis of amino acids, carbon metabolism, microbial metabolism in diverse environments, glycolysis/gluconeogenesis etc), play an important role in the phosphorylation of glyceraldehyde-3 phosphate and contributes in phosphotransferase activity and repair apoptosis [59]. Gap is upregulated in biofilms developed by numerous bacterial species [60,61,62,63,64].
Metabolic activity and growth rate of the bacteria are affected by the changes in the gradient of oxygen and other nutrients within the biofilm. Many studies have demonstrated that cells within hypoxic conditions have decreased metabolic activity [65,66,67], this slow pace of development suggests tolerance as antimicrobials are most efficient against rapidly developing cells [68,69,70]. In addition, the deeper layers of cells are also located in biofilms with undergrowth-limiting conditions, with anaerobic or micro-aerobic conditions. Pyruvate fermentation could support these cells, allowing them to survive with little or no oxygen [71]. In our S. aureus biofilm extractomes, we observed significant upregulation of acetolactate synthase (alsS) which is responsible for the activation of butanediol pathway from pyruvate. Activation of this pathway will promote NADH oxidation and indicate that there is a tenuous redox balance during the development of biofilms [50]. Another study reported that alsS utilise pyruvate to produce acetoin which is essential for acid tolerance within biofilms [72].
Among the 273 DREPs, unique or exclusive proteins identified in S. aureus biofilm contain 34 of functionally unknown or very little-known hypothetical proteins (Tables S2 and S3) including a hypothetical protein namely hypothetical protein KQ76_08425 encoded by SA0772 with the highest upregulation (5.09 fold), suggests that the complex metabolic and regulatory reaction to biofilm is not yet fully elucidated. Even though the role of the hypothetical protein remains unknown, it is probable to play a part in the distinct physiological state of the biofilm. In particular, we can speculate for those exclusively upregulated in biofilm growth state. Although previous reports have suggested that certain proteins may be involved in altering biofilm structures [73,74,75], more studies are needed to determine their specific roles.to assess their role.
In this present study, we have constructed a comprehensive reference map of the proteome of S. aureus biofilm, observed a significant range of abundance variation in the biofilm, identified differentially expressed potential marker proteins, and elucidated potential role (s) of these exclusive proteins using this reference strain. In the future studies, identified significant marker proteins such as virulence factors, antibiofilm agents, will be further characterised and analysed using different platforms (e.g., targeted ELISAs, biochemical assays) to validate the proteomics results in numerous S. aureus strains.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: The network of DREPs of S. aureus biofilm was analysed by STRING software (v.10.0).; Table S1: Primers used in this study; Table S2: List of the significantly upregulated proteins in S. aureus biofilm in comparison to planktonic culture ; Table S3: List of the exclusively downregulated proteins in S. aureus biofilm in comparison to planktonic culture; Table S4: Relative correlation of selected qPCR gene expression data to TMT protein expression data in 3-day biofilm in comparison to planktonic culture.
Author Contributions
Conceptualization, M.A.R., K.V. and H.H.; methodology, M.A.R., K.V., H.H., A.A. and D.C.; formal analysis, M.A.R., K.V., H.H., D.C., and A.A.; investigation, M.A.R., K.V., H.H., and A.A.; data curation, M.A.R., K.V., H.H. and A.A.; writing—original draft preparation, M.A.R., K.V., H.H. and A.A.; writing—review and editing, M.A.R., K.V., H.H., and A.A.; supervision, K.V. and H.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding
Institutional Review Board Statement
Not applicable
Informed Consent Statement
Not applicable
Data Availability Statement
Data are available via ProteomeXchange [76] with identifier PXD033499.
Acknowledgments
We acknowledge the support of an International Macquarie University Research Excellence Scholarship from Macquarie University for M.A.R. We thank Maria Mempin for her kind advice and technique support for this study. We thank the technique support from the Australian Proteome Analysis Facility, Macquarie University. We also thank Dana Pascovici and the team for their great help in biostatistical and bioinformatics analysis at APAF.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Sandel, M.K.; McKillip, J.L. Virulence and recovery of Staphylococcus aureus relevant to the food industry using improvements on traditional approaches. Food Control 2004, 15, 5–10. [Google Scholar] [CrossRef]
- Bukowski, M.; Wladyka, B.; Dubin, G. Exfoliative Toxins of Staphylococcus aureus. Toxins 2010, 2, 1148–1165. [Google Scholar] [CrossRef]
- Gordon, R.J.; Lowy, F.D. Pathogenesis of methicillin-resistant Staphylococcus aureus infection. Clin. Infect. Dis. 2008, 46, S350–S359. [Google Scholar] [CrossRef] [PubMed]
- Parsek, M.R.; Singh, P.K. Bacterial Biofilms: An Emerging Link to Disease Pathogenesis. Annu. Rev. Microbiol. 2003, 57, 677–701. [Google Scholar] [CrossRef]
- Kiedrowski, M.R.; Horswill, A.R. New approaches for treating staphylococcal biofilm infections. Ann. New York Acad. Sci. 2011, 1241, 104–121. [Google Scholar] [CrossRef] [PubMed]
- Barrett, L.; Atkins, B. The clinical presentation of prosthetic joint infection. J. Antimicrob. Chemother. 2014, 69, i25–i27. [Google Scholar] [CrossRef] [PubMed]
- Maiti, P.; Chatterjee, S.; Kundu, A.; Dey, R. Biofilms on indwelling urologic devices: Microbes and antimicrobial management prospect. Ann. Med Heal. Sci. Res. 2014, 4, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Ajdic, D.; Zoghbi, Y.; Gerth, D.; Panthaki, Z.J.; Thaller, S. The Relationship of Bacterial Biofilms and Capsular Contracture in Breast Implants. Aesthetic Surg. J. 2016, 36, 297–309. [Google Scholar] [CrossRef]
- Pajkos, A.; Deva, A.K.; Vickery, K.; Cope, C.; Chang, L.; Cossart, Y.E. Detection of Subclinical Infection in Significant Breast Implant Capsules. Plast. Reconstr. Surg. 2003, 111, 1605–1611. [Google Scholar] [CrossRef]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: from the Natural environment to infectious diseases. Nat. Rev. Microbiol. 2004, 2, 95–108. [Google Scholar] [CrossRef]
- Flemming, H.-C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623. [Google Scholar] [CrossRef] [PubMed]
- Graf, A.C.; Leonard, A.; Schäuble, M.; Rieckmann, L.M.; Hoyer, J.; Maass, S.; Lalk, M.; Becher, D.; Pané-Farré, J.; Riedel, K. Virulence Factors Produced by Staphylococcus aureus Biofilms Have a Moonlighting Function Contributing to Biofilm Integrity. Mol. Cell. Proteom. 2019, 18, 1036–1053. [Google Scholar] [CrossRef]
- Resch, A.; Leicht, S.; Saric, M.; Pásztor, L.; Jakob, A.; Götz, F.; Nordheim, A. Comparative proteome analysis of Staphylococcus aureus biofilm and planktonic cells and correlation with transcriptome profiling. Proteomics 2006, 6, 1867–1877. [Google Scholar] [CrossRef] [PubMed]
- Islam, N.; Kim, Y.; Ross, J.M.; Marten, M.R. Proteomic analysis of Staphylococcus aureus biofilm cells grown under physiologically relevant fluid shear stress conditions. Proteome Sci. 2014, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Atshan, S.S.; Shamsudin, M.N.; Sekawi, Z.; Lung, L.T.T.; Barantalab, F.; Liew, Y.K.; Alreshidi, M.A.; Abduljaleel, S.A.; Hamat, R.A. Comparative proteomic analysis of extracellular proteins expressed by various clonal types of Staphylococcus aureus and during planktonic growth and biofilm development. Front. Microbiol. 2015, 6, 524. [Google Scholar] [CrossRef] [PubMed]
- Moche, M.; Schlüter, R.; Bernhardt, J.; Plate, K.; Riedel, K.; Hecker, M.; Becher, D. Time-Resolved Analysis of Cytosolic and Surface-Associated Proteins of Staphylococcus aureus HG001 under Planktonic and Biofilm Conditions. J. Proteome Res. 2015, 14, 3804–3822. [Google Scholar] [CrossRef] [PubMed]
- Ymele-Leki, P.; Ross, J.M. Erosion from Staphylococcus aureus Biofilms Grown under Physiologically Relevant Fluid Shear Forces Yields Bacterial Cells with Reduced Avidity to Collagen. Appl. Environ. Microbiol. 2007, 73, 1834–1841. [Google Scholar] [CrossRef]
- Rahman, A.; Amirkhani, A.; Chowdhury, D.; Mempin, M.; Molloy, M.P.; Deva, A.K.; Vickery, K.; Hu, H. Proteome of Staphylococcus aureus Biofilm Changes Significantly with Aging. Int. J. Mol. Sci. 2022, 23, 6415. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Cheng, G.; Karunakaran, R.; East, A.K.; Poole, P.S. Multiplicity of sulfate and molybdate transporters and their role in nitrogen fixation in Rhizobium leguminosarum bv. viciae Rlv3841. Mol. Plant-Microbe Interact. 2016, 29, 143–152. [Google Scholar] [CrossRef]
- Verdon, J.; Girardin, N.; Lacombe, C.; Berjeaud, J.-M.; Héchard, Y. δ-hemolysin, an update on a membrane-interacting peptide. Peptides 2009, 30, 817–823. [Google Scholar] [CrossRef] [PubMed]
- Shah, P.; Swiatlo, E. A multifaceted role for polyamines in bacterial pathogens. Mol. Microbiol. 2008, 68, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Antognoni, F.; Del Duca, S.; Kuraishi, A.; Kawabe, E.; Fukuchi-Shimogori, T.; Kashiwagi, K.; Igarashi, K. Transcriptional Inhibition of the Operon for the Spermidine Uptake System by the Substrate-binding Protein PotD. J. Biol. Chem. 1999, 274, 1942–1948. [Google Scholar] [CrossRef] [PubMed]
- Atshan, S.S.; Shamsudin, M.N.; Lung, T.; Than, L. Comparative characterisation of genotypically different clones of MRSA in the production of biofilms. BioMed Research International 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- da Silva, W.M., Bei, J., Amigo, N., Valacco, P. Quantification of Enterohemorrhagic Escherichia coli O157: H7 proteome using TMT-Based Analysis. bioRxiv 2018, 312652.
- Emery, S.J.; Mirzaei, M.; Vuong, D.; Pascovici, D.; Chick, J.M.; Lacey, E.; Haynes, P.A. Induction of virulence factors in Giardia duodenalis independent of host attachment. Sci. Rep. 2016, 6, 20765. [Google Scholar] [CrossRef] [PubMed]
- Theodoulou, F.L.; Kerr, I.D. ABC transporter research: going strong 40 years on. Biochem. Soc. Trans. 2015, 43, 1033–1040. [Google Scholar] [CrossRef]
- Resch, A.; Rosenstein, R.; Nerz, C.; Götz, F. Differential Gene Expression Profiling of Staphylococcus aureus Cultivated under Biofilm and Planktonic Conditions. Appl. Environ. Microbiol. 2005, 71, 2663–2676. [Google Scholar] [CrossRef]
- Islam, N.; Ross, J.M.; Marten, M.R. Proteome Analyses of Staphylococcus aureus Biofilm at Elevated Levels of NaCl. Clin. Microbiol. Open Access 2015, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; Leid, J.G.; Camper, A.K.; Costerton, J.W.; Shirtliff, M.E. Identification of Staphylococcus aureus Proteins Recognized by the Antibody-Mediated Immune Response to a Biofilm Infection. Infect. Immun. 2006, 74, 3415–3426. [Google Scholar] [CrossRef]
- Yang, J.; He, Y.; Jiang, J.; Chen, W.; Gao, Q.; Pan, L.; Shi, C. Comparative proteomic analysis by iTRAQ-2DLC-MS/MS provides insight into the key proteins involved in Cronobacter sp. biofilm formation. Food Control 2016, 63, 93–100. [Google Scholar] [CrossRef]
- Branda, S.S.; González-Pastor, J.E.; Dervyn, E.; Ehrlich, S.D.; Losick, R.; Kolter, R. Genes Involved in Formation of Structured Multicellular Communities by Bacillus subtilis. J. Bacteriol. 2004, 186, 3970–3979. [Google Scholar] [CrossRef] [PubMed]
- Crowley, R.C.; Leigh, J.A.; Ward, P.N.; Lappin-Scott, H.M.; Bowler, L.D. Differential Protein Expression in Streptococcus uberis under Planktonic and Biofilm Growth Conditions. Appl. Environ. Microbiol. 2011, 77, 382–384. [Google Scholar] [CrossRef] [PubMed]
- Hinsa, S.M.; Espinosa-Urgel, M.; Ramos, J.L.; O'Toole, G.A. Transition from reversible to irreversible attachment during biofilm formation by Pseudomonas fluorescens WCS365 requires an ABC transporter and a large secreted protein. Mol. Microbiol. 2003, 49, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Vanderlinde, E.M.; Harrison, J.J.; Muszyński, A.; Carlson, R.W.; Turner, R.J.; Yost, C.K. Identification of a novel ABC transporter required for desiccation tolerance, and biofilm formation inRhizobium leguminosarumbv. viciae3841. FEMS Microbiol. Ecol. 2010, 71, 327–340. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Long, F.; Chen, Y.; Knøchel, S.; She, Q.; Shi, X. A Putative ABC Transporter Is Involved in Negative Regulation of Biofilm Formation by Listeria monocytogenes. Appl. Environ. Microbiol. 2008, 74, 7675–7683. [Google Scholar] [CrossRef] [PubMed]
- Beenken, K.E.; Dunman, P.M.; McAleese, F.; Macapagal, D.; Murphy, E.; Projan, S.J.; Blevins, J.S.; Smeltzer, M.S. Global Gene Expression in Staphylococcus aureus Biofilms. J. Bacteriol. 2004, 186, 4665–4684. [Google Scholar] [CrossRef] [PubMed]
- Vytvytska, O.; Nagy, E.; Blüggel, M.; Meyer, H.E.; Kurzbauer, R.; Huber, L.A.; Klade, C.S. Identification of vaccine candidate antigens of Staphylococcus aureus by serological proteome analysis. Proteomics 2002, 2, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Corrigan, R.M.; Miajlovic, H.; Foster, T.J. Surface proteins that promote adherence of Staphylococcus aureus to human desquamated nasal epithelial cells. BMC Microbiol. 2009, 9, 22. [Google Scholar] [CrossRef] [PubMed]
- Hofbauer, B.; Vomacka, J.; Stahl, M.; Korotkov, V.S.; Jennings, M.C.; Wuest, W.M.; Sieber, S.A. Dual Inhibitor of Staphylococcus aureus Virulence and Biofilm Attenuates Expression of Major Toxins and Adhesins. Biochemistry 2018, 57, 1814–1820. [Google Scholar] [CrossRef]
- Kot, B.; Sytykiewicz, H.; Sprawka, I. Expression of the Biofilm-Associated Genes in Methicillin-Resistant Staphylococcus aureus in Biofilm and Planktonic Conditions. Int. J. Mol. Sci. 2018, 19, 3487. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The Staphylococcal Biofilm: Adhesins, Regulation, and Host Response. Microbiol. Spectr. 2016, 4. [Google Scholar] [CrossRef]
- Ibberson, C.B.; Parlet, C.P.; Kwiecinski, J.; Crosby, H.A.; Meyerholz, D.K.; Horswill, A.R. Hyaluronan Modulation Impacts Staphylococcus aureus Biofilm Infection. Infect. Immun. 2016, 84, 1917–1929. [Google Scholar] [CrossRef]
- Rahman, A.; Choi, Y.H.; Pradeep, G.C.; Yoo, J.C. An ammonium sulfate sensitive chitinase from Streptomyces sp. CS501. Arch. Pharmacal Res. 2014, 37, 1522–1529. [Google Scholar] [CrossRef] [PubMed]
- Boles, B.R.; Horswill, A.R. agr-Mediated Dispersal of Staphylococcus aureus Biofilms. PLOS Pathog. 2008, 4, e1000052. [Google Scholar] [CrossRef] [PubMed]
- Periasamy, S.; Joo, H.-S.; Duong, A.C.; Bach, T.-H.L.; Tan, V.Y.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. How Staphylococcus aureus biofilms develop their characteristic structure. Proc. Natl. Acad. Sci. 2012, 109, 1281–1286. [Google Scholar] [CrossRef]
- Tsang, L.H.; Cassat, J.E.; Shaw, L.N.; Beenken, K.E.; Smeltzer, M.S. Factors Contributing to the Biofilm-Deficient Phenotype of Staphylococcus aureus sarA Mutants. PLOS ONE 2008, 3, e3361. [Google Scholar] [CrossRef]
- Kantyka, T.; Plaza, K.; Koziel, J.; Florczyk, D.; Stennicke, H.R.; Thogersen, I.B.; Enghild, J.J.; Silverman, G.A.; Pak, S.C.; Potempa, J. Inhibition of Staphylococcus aureus cysteine proteases by human serpin potentially limits staphylococcal virulence. Biol. Chem. 2011, 392, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Bryers, J.D. Medical biofilms. Biotechnol. Bioeng. 2008, 100, 1–18. [Google Scholar] [CrossRef]
- Zhu, Y.; Weiss, E.C.; Otto, M.; Fey, P.D. Staphylococcus aureus biofilm metabolism and the influence of arginine on polysaccharide intercellular adhesin synthesis, biofilm formation, and pathogenesis. Infect. Immun. 2007, 75, 4219–4226. [Google Scholar] [CrossRef]
- Chen, X.; Wu, H.; Cao, Y.; Yao, X.; Zhao, L.; Wang, T.; Yang, Y.; Lv, D.; Chai, Y.; Cao, Y.; et al. Ion-pairing chromatography on a porous graphitic carbon column coupled with time-of-flight mass spectrometry for targeted and untargeted profiling of amino acid biomarkers involved in Candida albicans biofilm formation. Mol. Biosyst. 2013, 10, 74–85. [Google Scholar] [CrossRef]
- Ammons, M.C.B.; Tripet, B.P.; Carlson, R.P.; Kirker, K.R.; Gross, M.A.; Stanisich, J.J.; Copié, V. Quantitative NMR Metabolite Profiling of Methicillin-Resistant and Methicillin-Susceptible Staphylococcus aureus Discriminates between Biofilm and Planktonic Phenotypes. J. Proteome Res. 2014, 13, 2973–2985. [Google Scholar] [CrossRef] [PubMed]
- Wall, E.A.; Caufield, J.H.; Lyons, C.E.; Manning, K.A. Specific N-terminal cleavage of ribosomal protein L 27 in S taphylococcus aureus and related bacteria. Mol. Microbiol. 2015, 95, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Weiss, A.; Moore, B.D.; Tremblay, M.H.J.; Chaput, D.; Kremer, A.; Shaw, L.N. The ω Subunit Governs RNA Polymerase Stability and Transcriptional Specificity in Staphylococcus aureus. J. Bacteriol. 2017, 199, e00459-00416. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, H.; Qi, T.; Yan, X.; Wang, B.; Guan, J.; Li, Y. Comparative transcriptomics analyses of the different growth states of multidrug-resistant Acinetobacter baumannii. Biomed. Pharmacother. 2017, 85, 564–574. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.D.; Franklin, M.J.; Park, C.-H.; McFeters, G.A.; Stewart, P.S. Gene expression and protein levels of the stationary phase sigma factor, RpoS, in continuously-fed Pseudomonas aeruginosa biofilms. FEMS Microbiol. Lett. 2001, 199, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Yao, Z.; Sun, L.; Hu, W.; Cao, J.; Lin, W.; Lin, X. Proteomics Analysis Reveals a Potential Antibiotic Cocktail Therapy Strategy for Aeromonas hydrophila Infection in Biofilm. J. Proteome Res. 2016, 15, 1810–1820. [Google Scholar] [CrossRef] [PubMed]
- Weber, H.; Engelmann, S.; Becher, D.; Hecker, M. Oxidative stress triggers thiol oxidation in the glyceraldehyde-3-phosphate dehydrogenase of Staphylococcus aureus. Mol. Microbiol. 2004, 52, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, L.; Madureira, P.; Andrade, E.B.; Bouaboud, A.; Morello, E.; Ferreira, P.; Poyart, C.; Trieu-Cuot, P.; Dramsi, S. Group B Streptococcus GAPDH Is Released upon Cell Lysis, Associates with Bacterial Surface, and Induces Apoptosis in Murine Macrophages. PLOS ONE 2012, 7, e29963. [Google Scholar] [CrossRef] [PubMed]
- Foulston, L.; Elsholz, A.K.W.; DeFrancesco, A.S.; Losick, R. The Extracellular Matrix of Staphylococcus aureus Biofilms Comprises Cytoplasmic Proteins That Associate with the Cell Surface in Response to Decreasing pH. mBio 2014, 5, e01667-14. [Google Scholar] [CrossRef]
- Gil, C.; Solano, C.; Burgui, S.; Latasa, C.; García, B.; Toledo-Arana, A.; Lasa, I.; Valle, J. Biofilm Matrix Exoproteins Induce a Protective Immune Response against Staphylococcus aureus Biofilm Infection. Infect. Immun. 2014, 82, 1017–1029. [Google Scholar] [CrossRef]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa Displays Multiple Phenotypes during Development as a Biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef]
- Planchon, S.; Desvaux, M.; Chafsey, I.; Chambon, C.; Leroy, S.; Hébraud, M.; Talon, R. Comparative Subproteome Analyses of Planktonic and Sessile Staphylococcus xylosus C2a: New Insight in Cell Physiology of a Coagulase-Negative Staphylococcus in Biofilm. J. Proteome Res. 2009, 8, 1797–1809. [Google Scholar] [CrossRef]
- Wang, Y.; Yi, L.; Wu, Z.; Shao, J.; Liu, G.; Fan, H.; Zhang, W.; Lu, C. Comparative Proteomic Analysis of Streptococcus suis Biofilms and Planktonic Cells That Identified Biofilm Infection-Related Immunogenic Proteins. PLOS ONE 2012, 7, e33371. [Google Scholar] [CrossRef] [PubMed]
- Stewart, P.S.; Zhang, T.; Xu, R.; Pitts, B.; Walters, M.C.; Roe, F.; Kikhney, J.; Moter, A. Reaction–diffusion theory explains hypoxia and heterogeneous growth within microbial biofilms associated with chronic infections. npj Biofilms Microbiomes 2016, 2, 16012. [Google Scholar] [CrossRef]
- Walters, M.C.; Roe, F.; Bugnicourt, A.; Franklin, M.J.; Stewart, P.S. Contributions of antibiotic penetration, oxygen limitation, and low metabolic activity to tolerance of Pseudomonas aeruginosa biofilms to ciprofloxacin and tobramycin. Antimicrob. Agents Chemother. 2003, 47, 317–323. [Google Scholar] [CrossRef]
- Borriello, G.; Werner, E.; Roe, F.; Kim, A.M.; Ehrlich, G.D.; Stewart, P.S. Oxygen Limitation Contributes to Antibiotic Tolerance of Pseudomonas aeruginosa in Biofilms. Antimicrob. Agents Chemother. 2004, 48, 2659–2664. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Stewart, P.S. Growth limitation of Staphylococcus epidermidis in biofilms contributes to rifampin tolerance. Biofilms 2004, 1, 31–35. [Google Scholar] [CrossRef]
- Eng, R.H.; Padberg, F.T.; Smith, S.M.; Tan, E.N.; Cherubin, C.E. Bactericidal effects of antibiotics on slowly growing and nongrowing bacteria. Antimicrob. Agents Chemother. 1991, 35, 1824–1828. [Google Scholar] [CrossRef]
- Evans, D.J.; Allison, D.G.; Brown, M.R.W.; Gilbert, P. Susceptibility of Pseudomonas aeruginosa and Escherichia coli biofilms towards ciprofloxacin: effect of specific growth rate. J. Antimicrob. Chemother. 1991, 27, 177–184. [Google Scholar] [CrossRef]
- Eschbach, M.; Schreiber, K.; Trunk, K.; Buer, J.; Jahn, D.; Schobert, M. Long-Term Anaerobic Survival of the Opportunistic Pathogen Pseudomonas aeruginosa via Pyruvate Fermentation. J. Bacteriol. 2004, 186, 4596–4604. [Google Scholar] [CrossRef]
- Sambanthamoorthy, K.; Schwartz, A.; Nagarajan, V.; O Elasri, M. The Role of msa in Staphylococcus aureus Biofilm Formation. BMC Microbiol. 2008, 8, 221. [Google Scholar] [CrossRef] [PubMed]
- Klein, M.I.; Xiao, J.; Lu, B.; Delahunty, C.M.; Yates, J.R.; Koo, H. Streptococcus mutans Protein Synthesis during Mixed-Species Biofilm Development by High-Throughput Quantitative Proteomics. PLOS ONE 2012, 7, e45795. [Google Scholar] [CrossRef] [PubMed]
- Brady, R.A.; May, G.A.; Leid, J.G.; Prior, M.L. Resolution of Staphylococcus aureus Biofilm Infection Using Vaccination and Antibiotic Treatment. Infect. Immun. 2011, 79, 1797. [Google Scholar] [CrossRef] [PubMed]
- Chew, J.; Zilm, P.S.; Fuss, J.M.; Gully, N.J. A proteomic investigation of Fusobacterium nucleatum alkaline-induced biofilms. BMC Microbiol. 2012, 12, 189. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: a hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
Figure 1.
Classification of the DREPs of S. aureus biofilm using Gene Ontology based on their functional annotations. (A) GO Protein Classes; (B) GO Molecular Function; (C) GO Biological Processes.
Figure 1.
Classification of the DREPs of S. aureus biofilm using Gene Ontology based on their functional annotations. (A) GO Protein Classes; (B) GO Molecular Function; (C) GO Biological Processes.
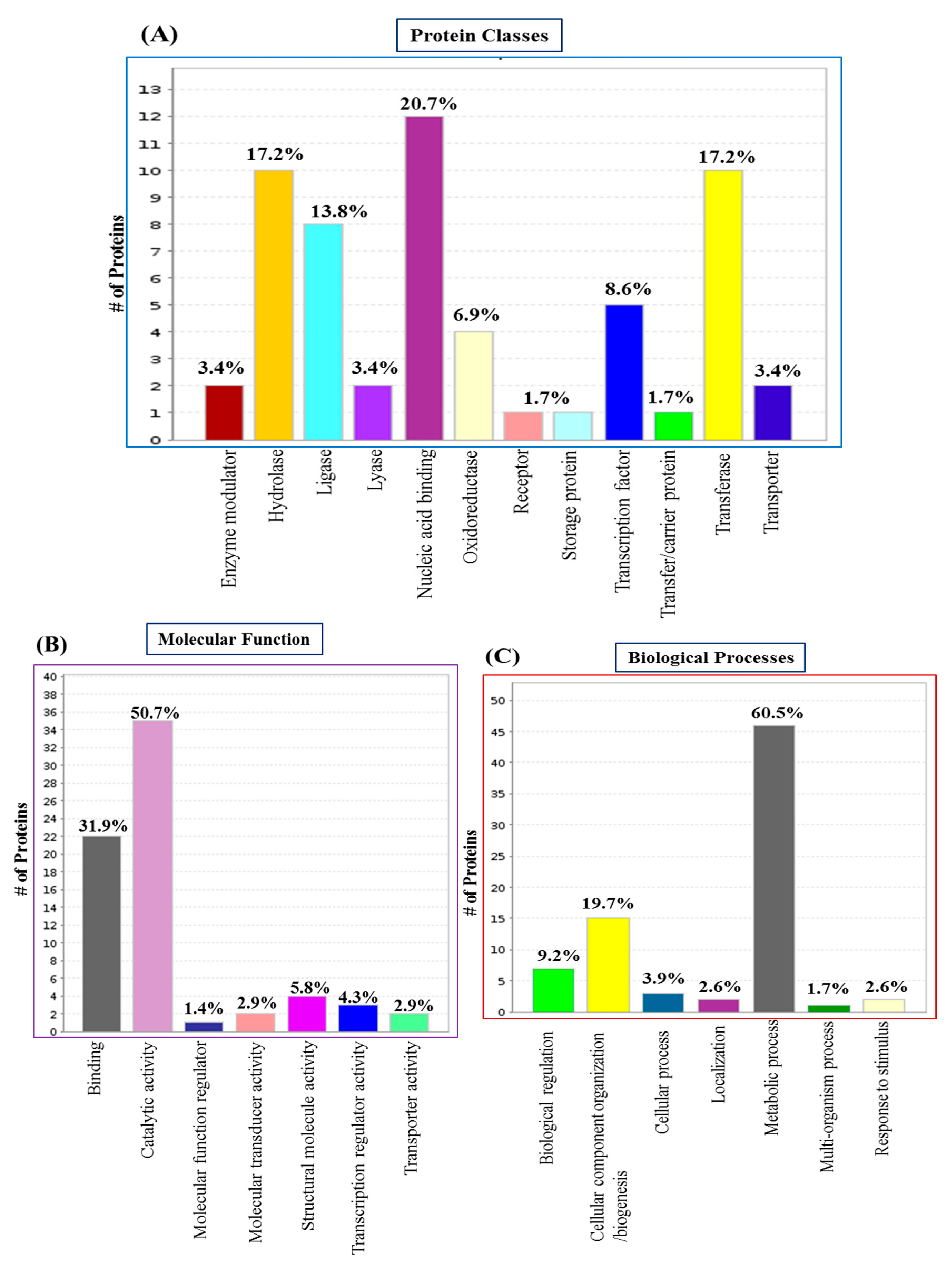
Figure 2.
The DREPs of S. aureus biofilm showed functional diversity using KEGG pathway analysis. (A) major pathways involved in exclusively upregulated proteins; (B) major pathways involved in exclusively downregulated proteins.
Figure 2.
The DREPs of S. aureus biofilm showed functional diversity using KEGG pathway analysis. (A) major pathways involved in exclusively upregulated proteins; (B) major pathways involved in exclusively downregulated proteins.
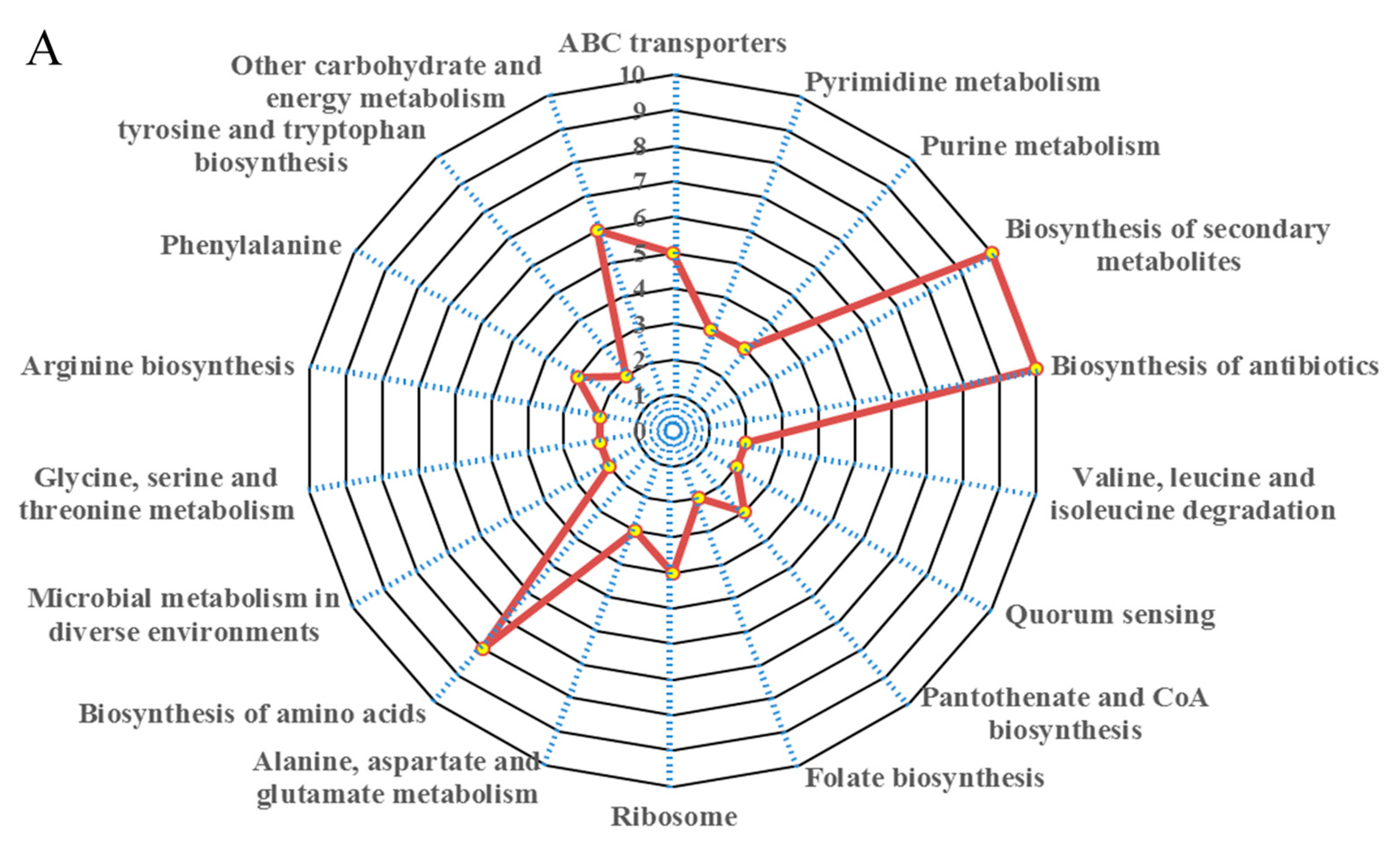

Figure 3.
This figure displays the structural component of hyaluronic acid (HA) and breaking point proteins (HysA and Chitinase). HA comprises of repeating disaccharide units of N-acetyl-D-glucosamine and D-glucuronic acid, and connected by repeating glycosidic bonds beta-1,4 and beta-1,3.
Figure 3.
This figure displays the structural component of hyaluronic acid (HA) and breaking point proteins (HysA and Chitinase). HA comprises of repeating disaccharide units of N-acetyl-D-glucosamine and D-glucuronic acid, and connected by repeating glycosidic bonds beta-1,4 and beta-1,3.
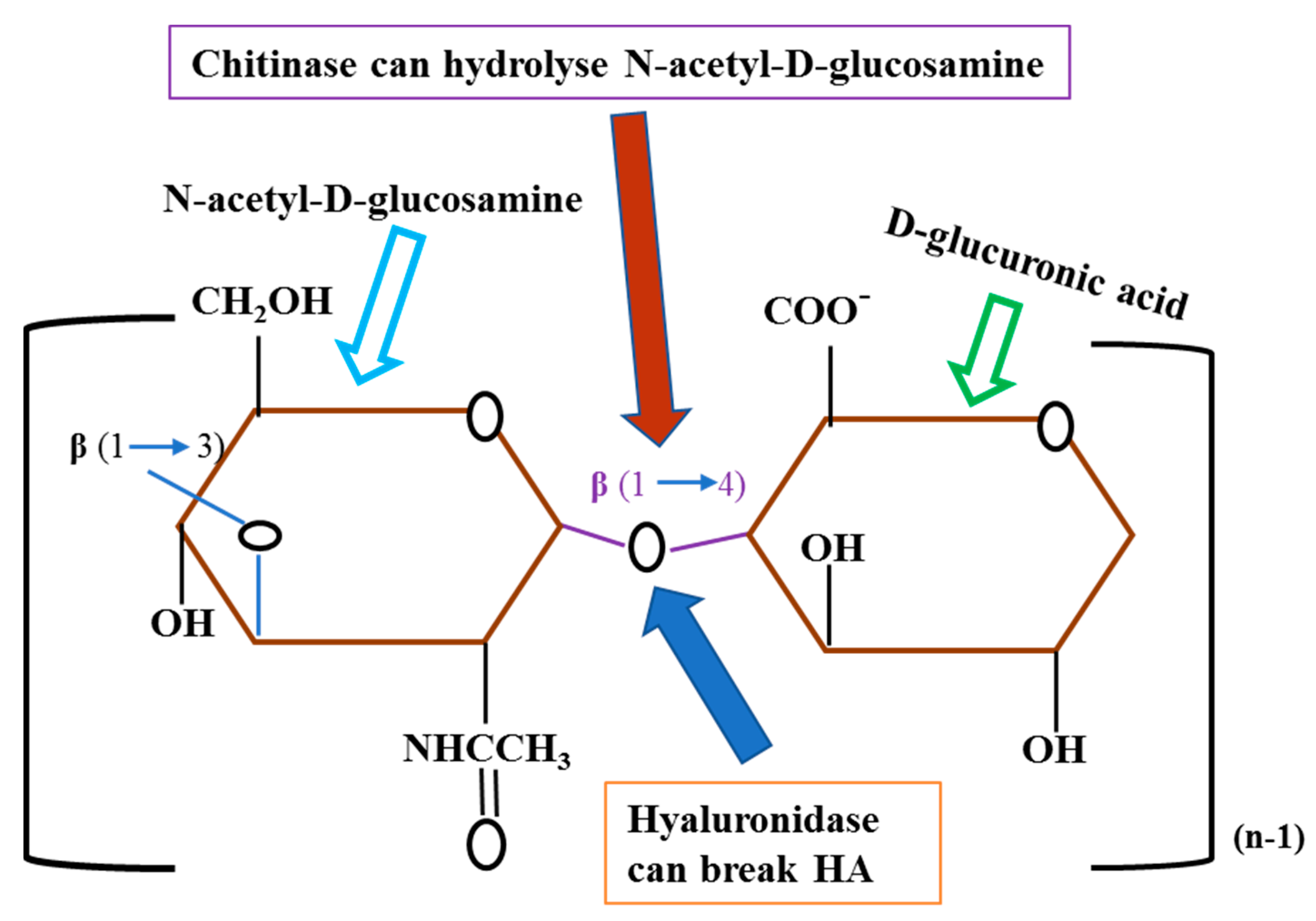
Figure 4.
Pathway demonstration showing exclusively upregulated proteins (red colour encoded genes) involved in amino sugar and nucleotide sugar metabolism and linked with peptidoglycan biosynthesis in 3-day biofilm.
Figure 4.
Pathway demonstration showing exclusively upregulated proteins (red colour encoded genes) involved in amino sugar and nucleotide sugar metabolism and linked with peptidoglycan biosynthesis in 3-day biofilm.
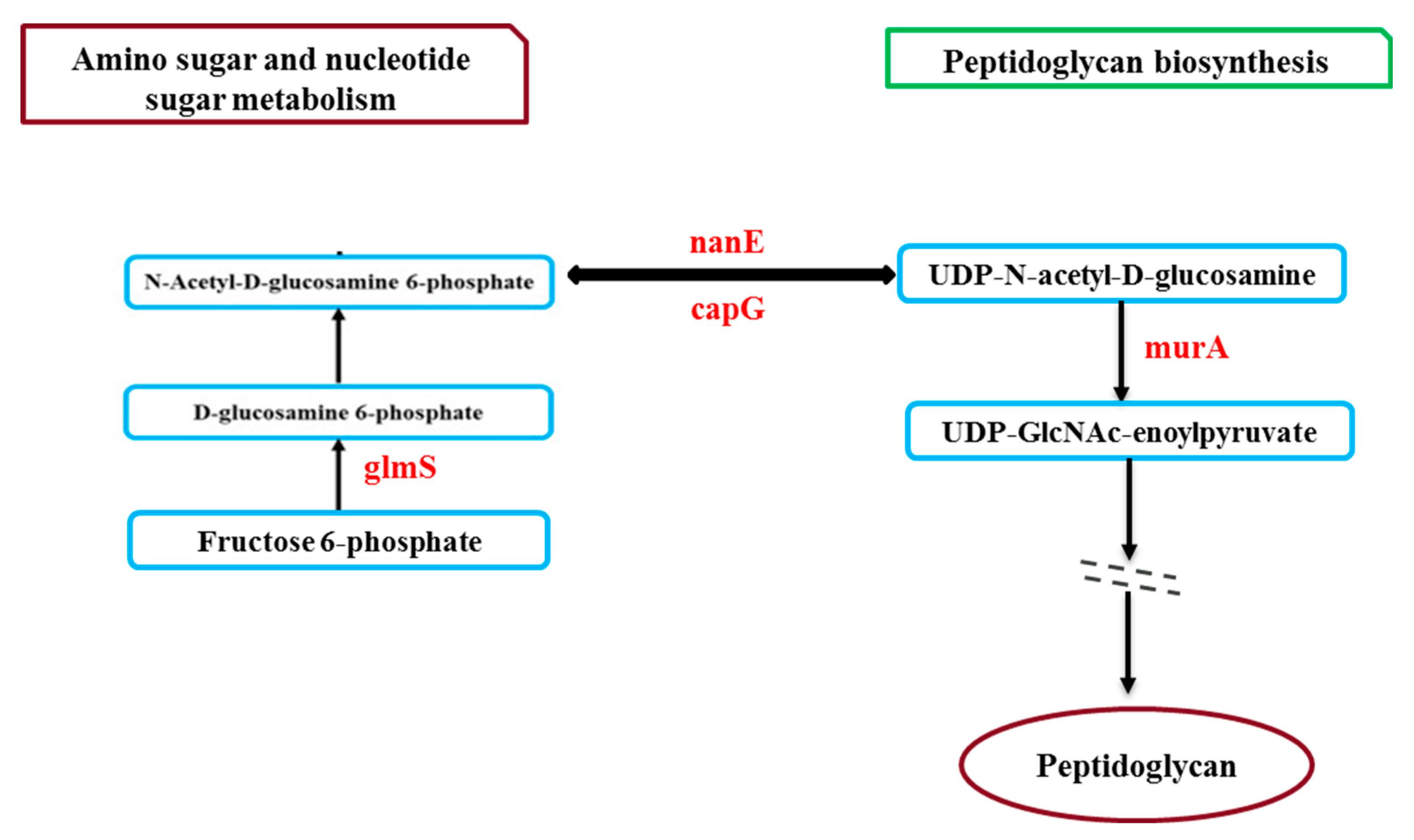

Table 1.
List of the exclusively differentially expressed virulence factor proteins identified by VFDB in S. aureus biofilm in comparison to planktonic culture (fold change >2, p <0.05). * CM denotes Cytoplasmic Membrane.
Table 1.
List of the exclusively differentially expressed virulence factor proteins identified by VFDB in S. aureus biofilm in comparison to planktonic culture (fold change >2, p <0.05). * CM denotes Cytoplasmic Membrane.
| Function | Accession ID | Uniprot ID | Virulence factors | Related genes | Fold change | Protein Pathway | Subcellular Localization |
|---|---|---|---|---|---|---|---|
| Adherence | AIO22275.1 | Q7A382 | Clumping factor B | clfB | -2.96 | S aureus infection | Cell wall |
| AIO22136.1 | Q7A3J7 | Fibronectin-binding protein A | fnbA SA2291 | -2.60 | Bacterial invasion of epithelial cells | Cell wall | |
| AIO19779.1 | A0A0H2X057 | Immunoglobulin G binding protein A | spa SACOL0095 | -4.71 | S aureus infection | Cell wall | |
| AIO20229.1 | Q5HIB2 | Serine-aspartate repeat-containing protein E | sdrE SACOL0610 | -3.82 | S aureus infection | Cell wall | |
| AIO20228.1 | Q7A780 | Serine-aspartate repeat-containing protein D | sdrD SA0520 | -4.49 | S aureus infection | Cell wall | |
| Toxins | AIO20763.1 | A0A0H3JMC2 | Alpha-Hemolysin | SA1007 | -13.83 | Extracellular | |
| AIO22369.1 | Q5HEI1 | Phospholipase C (EC 3.1.4.3) (Beta-hemolysin) (Beta-toxin) (Sphingomyelinase) (SMase) | hlb SACOL2003 | -12.33 | Quorum sensing, Inositol phosphate metabolism, Glycerophospholipid metabolism, Biosynthesis of secondary metabolites | Extracellular | |
| AIO21667.1 | P0A0M2 | Delta-hemolysin (Delta-lysin) (Delta-toxin) | hld SA1841.1 SAS065 | -34.02 | Quorum sensing | Extracellular | |
| AIO22060.1 | Q7A3S2 | Gamma-hemolysin component C | hlgC SA2208 | -3.79 | Staphylococcus aureus infection | Extracellular | |
| AIO20093.1 | A0A0H3JSX3 | Exotoxin 11 (superantigen-like protein) | set11 | -3.63 | Staphylococcus aureus infection | Extracellular | |
| Antiphagocytosis (Capsule) | AIO19823.1 | A0A0H3JKC9 | Capsular polysaccharide synthesis enzyme Cap5G | capG | 2.018 | Amino sugar and nucleotide sugar metabolism | Cytoplasmic |
| Exoenzyme | AIO21508.1 | Q5HEW4 | Serine protease SplE | splE | -4.18 | Quorum sensing | Extracellular |
| AIO21601.1 | P65826 | Cysteine proteinase A | scpA | -3.73 | Extracellular | ||
| AIO20644.1 | Q5HH36 | Cysteine proteinase B | sspB | -11.44 | Extracellular | ||
| AIO19987.1 | Q7A7P2 | lipase | geh | -4.12 | Extracellular | ||
| AIO21839.1 | A0A0H3JN21 | Hyaluronate lyase | hysA | 2.50 | Extracellular | ||
| AIO20645.1 | Q5HH35 | glutamyl endopeptidase | sspA | -6.52 | Quorum sensing | Extracellular | |
| OOC94232.1 | A0A0H2WZZ4 | aureolysin | aur | -3.22 | Staphylococcus aureus infection, Cationic antimicrobial peptide (CAMP) resistance | Extracellular | |
| AIO19888.1 | A0A0H3JNG8 | Staphylocoagulase | coa | 2.10 | Extracellular | ||
| Secretion system (Type VII secretion system) | AIO19949.1 | Q7A7S3 | type VII secretion protein EsaA | esaA SA0272 | -2.29 | CM |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



