
Submitted:
27 May 2024
Posted:
27 May 2024
You are already at the latest version
Abstract
Abiotic stress, especially drought stress poses a significant threat to the terrestrial plant growth, development, and productivity. Although mulberry have great genetic diversity and extensive stress tolerant traits in agroforestry systems, only a few reports offer preliminary insight into biochemical responses of mulberry leaves under drought conditions. In this study, we performed a comparative metabolomic and transcriptomic analysis on the “drooping mulberry” (Morus alba var. Pendula Dippel) under PEG-6000-simulated drought stress. Our research revealed that drought stress significantly enhanced flavonoid accumulation and up-regulated the expression of phenylpropanoid biosynthetic genes. Furthermore, the activities of superoxide dismutase (SOD), catalase (CAT) and malondialdehyde (MDA) content were elevated. In vitro enzyme assays and fermentation tests indicated the involvement of flavonol synthase/flavanone 3-hydroxylase (XM_010098126.2) and anthocyanidin 3-O-glucosyltransferase 5 (XM_010101521.2) in the biosynthesis of flavonol aglycones and glycosides, respectively. The recombinant MaF3GT5 protein was found to recognize kaempferol, quercetin, and UDP-glucose as substrates, but not 3-/7-O-glucosylated flavonols and UDP-rhamnose. MaF3GT5 is capable of forming 3-O- and 7-O-monoglucoside, but not di-O-glucosides, form kaempferol. This implies its role as a flavonol 3, 7-O-glucosyltransferase. The findings from this study provided insights into the biosynthesis of flavonoids and could have substantial implications for the future diversified utilization of mulberry.
Keywords:
drought stress
; mulberry
; flavonoids biosynthesis
; diversified utilization
1. Introduction
Terrestrial plant growth is strongly influenced by various abiotic stresses including drought, temperature, alkalinity, radiation, and salinity. Among these stresssors, drought is recognized as the most severe limiting factor affecting plant survival and yield[1]. Meanwhile, drought represents a persistent global ecological challenge, exacerbated by increasing temperatures[2]. Drought conditions typically lead to the accumulation of excess reactive oxygen species (ROS), reduced photosynthesis, and disruptions in plant metabolism[3,4]. In order to adapt to water scarcity, plants have developed various biological strategies, such as biochemical responses[5], antioxidant defense mechanisms[6], and the regulation of stomatal movement[7]. Numerous studies have revealed the differences in drought resistance among species and varieties, directly influencing factors such as total biomass, leaf area, and the root-to-shoot ratio[8,9,10]. Therefore, a comprehensive understanding of the mechanisms governing plant responses to drought stress is crucial for the effective screening and breeding of drought-resistant varieties.
The mulberry (Morus spp.) is an important tree crop in agroforestry, yielding fodder, fruit, fuel, and wood. It has been widely cultivated for thousands of years in China[11]. Mulberry leaves are used not only as forage for silkworms in sericulture, but also as herbal medicine and functional nutraceutical food for humans[12,13]. For a long time, growers have traditionally focused on the economic and pharmacological benefits due to the richness of mulberry leaves in various bioactive compounds like flavonoids, alkaloids, polyphenols, polysaccharides, and vitamins[14,15]. However, the growth and productivity of mulberry leaves are remarkably influenced by drought stress. Despite the genetic diversity of mulberry and its stress-tolerant traits in agroforestry systems, there is limited information available on biochemical responses of mulberry leaves to drought conditions[16]. The potential ecological benefits of mulberry leaves remain largely unexplored, promising diversified utilization of this valuable resource[17]. Further research in this area will contribute significantly to the broader understanding and application of mulberry’s ecological advantages.
Flavonoids are considered among the most critical constituents in mulberry leaves due to their diverse biochemical and pharmacological properties[18,19]. The biosynthesis of flavonoids is derived from phenylpropanoid metabolism and is regulated by a series of key genes[20]. In a study involving prolonged drought treatment in Achillea, it was observed that the expression of phenylalanine ammonia lyase (PAL) and flavone 3-hydroxylase (F3H) was significantly elevated at the onset of stress, later decreasing in the middle stage, whereas the expression of chalcone synthase (CHS), chalcone isomerase (CHI), and flavonoid 3′-hydroxylase (F3′H) increased during the middle stage. Toward the end of the stress period, the expression of all genes, except PAL, further increased, with the expression of these genes notably higher under stress conditions compared to non-stress conditions[21]. Recent studies have confirmed that flavonoids can enhance drought tolerance in maize (Zea mays)[22], sea buckthorn (Hippophae rhamnoides)[9], and wheat (Triticum aestivum)[23]. Mulberry leaves are rich in glycosylated flavonols such as astragalin, isoquercitrin, and rutin. Previous research has indicated a significant increase in rutin levels in drought-stressed Fava d’anta (Dimorphandra mollis) leaves, suggesting a potential role for rutin in protecting tissues against oxidative damage during drought periods[24]. Despite previous reports identifying 44 flavonoids in mulberry leaves and offering insights into rutin biosynthesis[13,25], little is known about their evaluation regarding drought stress tolerance. Given that comparative transcriptome analysis has revealed numerous key genes and pathways involved in stress response[26,27,28], a comprehensive widely-targeted metabolic profiling of mulberry leaves could provide further insights into the biochemical changes under drought condition[29,30].
The primary objective of this study was to elucidate drought-induced key genes or pathways in mulberry leaves and to explore the mechanisms governing flavonoid accumulation under drought stress conditions. To achieve this, we performed a comprehensive analysis of the metabolome and transcriptome, comparing drought-stressed “drooping mulberry” (Morus alba var. Pendula Dippel) leaves with untreated controls, as part of our investigation into mulberry drought biology. Our findings indicate significant alterations in phenylpropanoid metabolism and flavonoids content in response to drought stress. Furthermore, we conducted in vitro enzyme assays and fermentation tests to characterize flavonol synthase1 (MaFLS1) and flavonol 3, 7-O-glucosyltransferase (MaF3GT5). These enzymes play a pivotal role in the flavonols biosynthetic pathway responsible for synthesizing flavonol aglycones and glycosides[31]. These insights contribute to a better understanding of the biochemical mechanisms underlying mulberry drought tolerance, potentially facilitating the enhancement of flavonoid content and the medicinal utilization of mulberry resources.
2. Results
2.1. Measurement of Physiological Indices Related to Drought Damage
Under drought conditions, the accumulated free radicals and active oxygen is closely related to the activity of protective enzymes in plants, consequently leading to lipid peroxidation. In the present study, three key physiological indicators associated with drought stress were assessed: MDA content, SOD activity, and CAT activity. All measured parameters were compared to the control group (0 hour). The results revealed that CAT and SOD activities increased under drought stress compared to the control, showing a continuous increase from 0 hour to 72 hours, with the peak levels observed at 72 hours. Specifically, CAT activity rose from 1816.1 U/g at the control (0 hour) to 8089.9 U/g at 72 hours, while SOD activity increased from 34.2 U/g at the control to 181.2 U/g at 72 hours. Meanwhile, MDA concentration also exhibited an upward trend during drought stress. Unlike CAT and SOD, MDA levels increased steadily from the beginning of the experiment (28.5 nmol/g) up to 48 hours (51.3 nmol/g), followed by a slight decreased at 72 hours (43.91 nmol/g) (Figure 1).
2.2. Changes in Flavonoid Level under Drought Stress
The metabolic profiling analysis of mulberry leaves was conducted with three biological replicates under control (0 hour) and following 24, 48, and 72 hours of drought stress. Utilizing the mulberry metabolome database (MMHub, https://biodb.swu.edu.cn/mmdb/) established in a previous study[25], a total of 44 flavonoids present in mulberry and 2 authentic standards (kaempferol and quercetin) were assessed (Table S2). In this investigation, a total of 32 flavonoids were identified in M. Pendula leaves. O-glycosylated flavonols and malonylated flavonol glycosides were the predominant compounds. Moreover, some flavones such as apigenin 7-O-glucoside and luteolin 7-O-glucoside were also detected (Table S3).
The drought stress elicited varied responses in the individual compounds analyzed via UHPLC. Initially, at 0 hour (control), the malonylated, mono- and di-O-glycosylated flavonols, such as kaempferol 3-O-malonylglucoside, quercetin 3-O-glucoside, and rutin, exhibited the highest accumulation, while flavones and multiple- O-glycosylated flavonols were at notably low levels (Figure 2A). Subsequently, at 24 hours, all flavonoids showed increased levels compared to the control, with fold changes (log2FC) ranging from 0.31 to 9.26. Notably, quercetin O-rhamnosyl-O-hexosyl-O-hexoside-II experienced the most significant increase, while naringenin exhibited the least change. Moving to 48 hours, approximately half of compounds decreased in concentration compared to the levels observed at 24 hours. Particularly, quercetin O-hexosyl-O-hexosyl-O-hexoside displayed the most significant decrease with a log2FC of -3.87. Moreover, dertain flavonoids like apigenin maintained a continuous increase (log2FC = 3.70), while others showed no significant change. Almost all flavonoids reached their peak concentration at 72 hours. Quercetin O-rhamnosyl-O-hexosyl-O-hexoside-I and naringenin (0.12 and 0.17, respectively) showed no change compared to 48 hours, whereas kaempferol O-rhamnosyl-O-malonylhexoside exhibited the most significant increase with a log2FC of 4.15 (Figure 2B).
2.3. Identification of Flavonoid-Related Candidate Genes through Transcriptome Analysis
In order to identify the key genes related to the biosynthesis of flavonoids or involved in drought stress response in mulberry leaves, we performed an RNA sequencing (RNA-seq) analysis with three replications of two experimental groups: the control (C) and drought treated at 24 hours (D). In the present study, a total of 56,354,744 bases were assembled into 45,642 unigenes, displaying a mean length of 1,234 nucleotides (nt), an N50 length of 2,292 nt, an N50 count of 7,996, and a GC content of 40.44% (Table S4). The length distribution of all unigenes ranged from 200 to over 3,000 nt. Furthermore, 62.2% (28,401) of the unigenes were annotated through BLASTx searches (E-value threshold, 1 × 10−5) against the NR, Swiss-Prot, COG/KOG, and KEGG databases, resulting in 27,603, 18,371, 15,845, and 25,378 unigenes, respectively (Figure S1A).
We further performed the principal component analyses (PCA) on both the metabolome and transcriptome data. For the metabolome data, PC1 (47.7%) and PC2 (21.1%) collectively explained 68.8% of the variation in the metabolic profile (Figure S1B). Similarly, the PCA of transcriptome analysis indicated two distinct clusters along PC1, confirming the adequacy of our model (Figure S1C). Moreover, we identified 4,530 DEGs, with 2,495 genes showing up-regulation and the remainder displaying down-regulation (Figure S1D). Our KEGG pathway analysis revealed that the DEGs primarily participated in metabolism, genetic information processing, environmental information processing, cellular processes, and organismal systems, accounting for 60.8%, 30.2%, 4.6%, 1.6%, and 2.7% of the total, respectively (Figure S1E).
The drought-treated group exhibited a significantly higher relative abundance of flavonoids compared to the control group. Subsequently, an in silico filtration process[13,32] was employed to identify candidate genes related to flavonoid within the transcriptome libraries. In the present study, we identified 18 genes significantly changed in the flavonoid pathway (Table S3). Specifically, the expression levels of three PAL, two 4CL, two CHS, one F3H, two F3′H, one FLS, and four UGT genes displayed a 1.6 to 9.9 fold increased, while the expression of one CHS and two FLS genes decreased (Table S5). Besides, CHI and C4H did not exhibit significant changes in the transcriptome data. Given that flavonols are the most abundant flavonoids in mulberry leaves and certain key genes related to flavonol metabolism (e.g., MaF3GT (KP455729), MaF3G6″RT (KT324624), and MaF3H (EXC35356.1)) have been characterized in mulberry, we selected four candidate genes for further characterization —MaFLS1, MaUGT76A2, MaUGT90A1, and MaF3GT5— due to their high amino acid sequence identity (> 64%) with other sources (Figure S3). Furthermore, phylogenetic analysis suggested that MaUGT76A2 and MaUGT90A1 probably function as flavonoid rhamnosyltransferases, contributing to the biosynthesis of common O-rhamnosylated flavonols in mulberry leaves (e.g., kaempferol 3-O-rutinoside (K3R) and quercetin 3-O-rutinoside (Rutin)). Moreover, the distinct positioning of MaF3GT5 in the phylogenetic tree indicated functional differentiation of this candidate gene (Figure S2). Based on these results, we present a proposed flavonoid pathway and the key genes involved in drought response of mulberry leaves in Figure 3.
2.4. Expression Patterns of Phenylpropanoid Biosynthetic Genes under Drought Stress
We conducted a detailed analysis of the expression profiles of phenylpropanoid biosynthetic genes under prolonged drought conditions, focusing on 14 genes involved in the flavonoid pathway genes and 3 putative MaUGTs identified from the transcriptome data. Additionally, we examined a key enzyme in mulberry flavonol metabolism, flavonol 3-O-glucoside: 6″-O-rhamnosyltransferase (MaF3G6″RT), as previously described[13]. The results revealed that significant fluctuations in gene expression patterns in response to drought stress (Figure 4). Specifically, the expression levels of MaPAL1, MaPAL2, and MaPAL3 were markedly upregulated after 24 hours of drought treatment, followed by a significant decrease at 48 hours, and then stabilization until 72 hours. Consistent trends were observed for Ma4CL1, MaCHS1, and MaF3GT over the same time course. Notably, MaC4H, MaCHI, and MaF3G6″RhaT showed no significant changes throughout the experiment. On the other hand, Ma4CL2 and MaFLS1 exhibited a significant increase at 24 hours, followed by a gradual decline until 72 hours. MaCHS2 and MaF3H showed a similar pattern with an initial increase at 24 hours followed by a plateau until 72 hours. In contrast, MaF3′H1 displayed a consistent increase in expression levels during the entire experiment. Among the candidate MaUGTs, MaUGT90A1 initially increased, then decreased, and subsequently rose again, while MaUGT76A2 initially showed a slight increase, followed by a significant rise before returning to levels comparable to 24 hours (Figure 4). Noteworthy, all phenylpropanoid biosynthetic genes, except MaC4H2, MaCHI2, MaFLS2, MaF3G″6RT and MaUGT76A2, displayed significant changes in expression at 24 hours post-drought treatment, supporting the robustness of the RNA-seq data (Table S3).
2.5. Characterization of Recombinant MaFLS1 and MaF3GT5 Enzyems
The recombinant putative Morus alba L. flavonol synthase1 (MaFLS1) protein was successfully expressed in E. coil as a His tag-fused protein and then purified for enzymatic assays. MaFLS1 exhibited significant amino acid sequence homology with FLS proteins from various species such as Arabidopsis thaliana, Malus pumila, and Rosa multiflora (Figure S3A). It belongs to the typical 2-oxoglutarate-dependent dioxygenases (2-ODDs) family, which emphasizes its absolute dependence on iron (II), 2-oxoglutarate, and ascorbate for in vitro enzymic activity (Figure S3B). The recombinant MaFLS1 protein catalyzed the conversion of dihydroquercetin (7.35 min) and dihydrokaempferol (8.31 min) into distinct products (N1 and N2, respectively) (Figure 5). In the UHPLC-MS/MS analysis, N1 and N2 displayed distinct peaks and molecular ions at a mass-to-charge ratio of 303.0501 [M+H]+ and 287.0553 [M+H]+, respectively, consistent with the retention time and mass calculation (│m/z error│ ≤ 10 ppm) of quercetin (C15H10O7, 303.0499 [M+H]+, 9.45 min) and kaempferol (C15H10O6, 287.0550 [M+H]+, 10.62 min) (Figure 5A). Furthermore, the mass spectra of N1 and N2 exhibited identical fragmentation patterns to those of the authentic standards of kaempferol (153.0185, 121.0288, and 68.9981) and quercetin (153.0186, 137.0236, 257.0450, and 229.0496) (Figure 5B). In addition, the fermentation liquor of MaFLS1 changed in color from milky white to yellow after 48 hours of fermentation at 25°C (Figure 5A). It is worth noting that the detected fermented products were consistent with the result of the enzymatic assay described above. These observations indicated that MaFLS1 encodes a flavonol synthase that recognizes dihydroquercetin and dihydrokaempferol as potential substrates in mulberry.
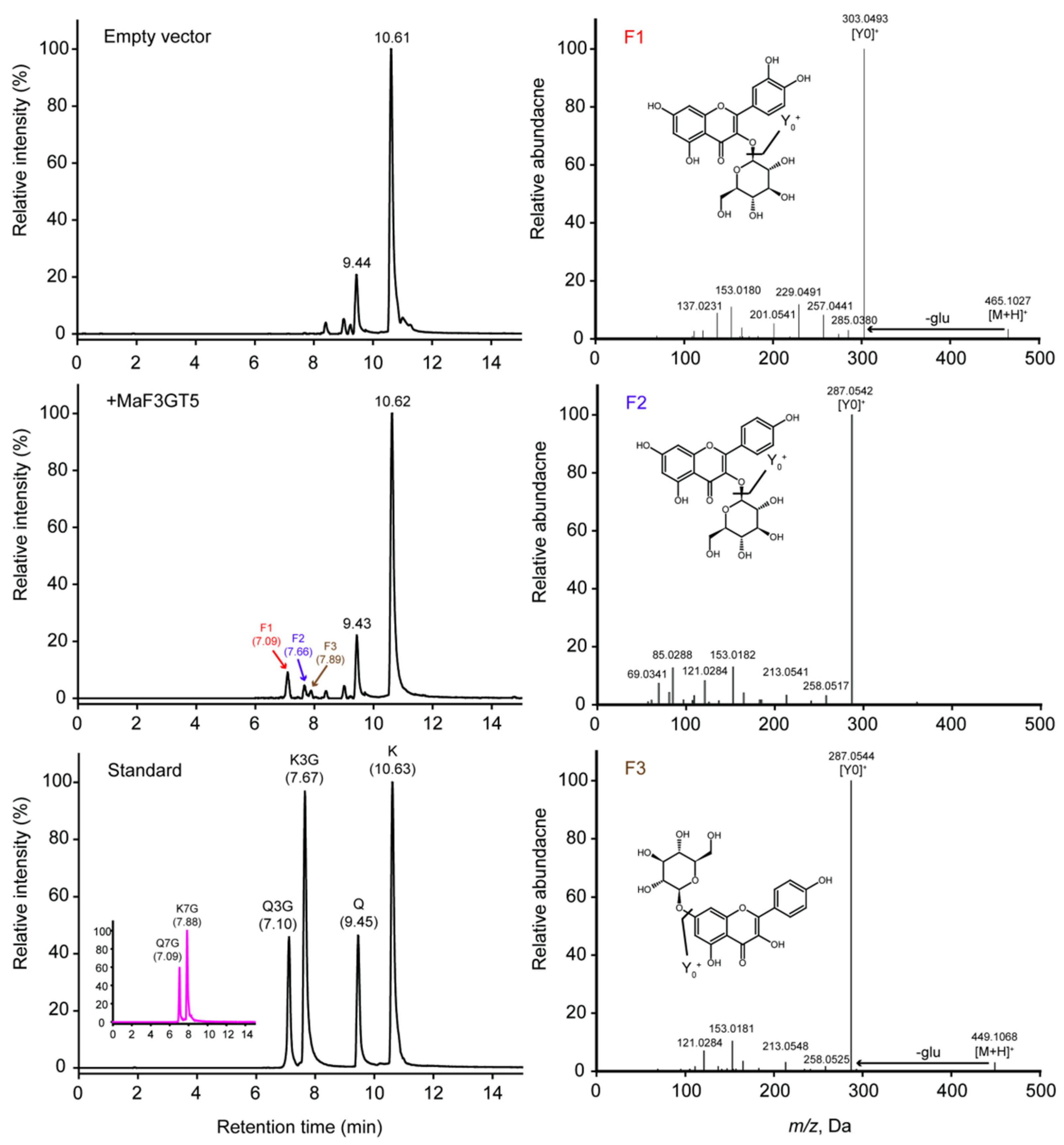
Three recombinant Morus alba L. flavonoid related UGTs were also obtained using the same method as MaFLS1. Two sugar donors (UDP-glucose and UDP-rhamnose) and six typical mulberry leaf flavonol aglycones or glycosides were tested as potential substrates, including kaempferol, quercetin, kaempferol 3-O-glucoside, kaempferol 7-O-glucoside, quercetin 3-O-glucoside, and quercetin-7-O-glucoside. Among these, only the recombinant MaF3GT5 protein catalyzed the conversion of kaempferol and quercetin into three products (F1, F2, and F3) (Figure 6). F2 and F3 had distinct peaks and molecular ions with a mass-to-charge ratio of 449.1070 [M+H]+ and 449.1068 [M+H]+, respectively, consistent with the retention time and mass calculation (│m/z error│ ≤ 10 ppm) of kaempferol 3-O-glucoside (C21H20O11, 7.66 min) and kaempferol 7-O-glucoside (C21H20O11, 7.89 min), as shown in Figure 6A. The MS2 spectra of F2 and F3 also displayed identical fragmentation profiles to those of the authentic standards of kaempferol 3-O-glucoside and kaempferol 7-O-glucoside, in which Y0+ ions were observed at m/z 287.0542 and 287.0544 due to the loss of the glucose moiety from the O-glycosyl group of the phenolic hydroxyl (-162) (Figure 6B). Remarkably, although F1 had a single peak and molecular ions at a mass-to-charge ratio of 465.1027 [M+H]+, consistent with the retention time, mass calculation, and MS2 spectra of quercetin 7-O-glucoside (C21H20O12, 7.09 min), 3-O- and 7-O-glycosylated quercetin presented the same elution profile (RTΔ = 0.01 min) under our condition (Figure 6A). The recombinant MaF3GT5 protein was found to recognize flavonol aglycones and UDP-glucose but not flavonol glycosides or UDP-rhamnose. These observations indicated that MaF3GT5 functions as a flavonol 3, 7-O-glucosyltransferase.
3. Discussion
Abiotic stress, particularly drought, can trigger a burst of ROS and affect the metabolic processes of plants, leading to an increased accumulation of H2O2 and other hydroxyl radicals, which seriously threaten the plant growth, development, and production. SOD and CAT are key enzymes within the protective enzymes system and plays a pivotal role in scavenging ROS[33]. Besides, it is likely that SOD and CAT also synergistically interact with protective secondary metabolites to uphold an optimal redox balance and individual fitness in plants[5]. Our study observed a continuous increase in the activities of SOD and CAT during PEG-6000 treatment, thus indicating the reliability of simulating drought stress. Drought stress invariably leads to lipid peroxidation, resulting in the accumulation of MDA. The MDA content serves as an indicator of stress severity. Although the MDA accumulation presented a certain degree of decrease at 72 hours, the content of MDA was significantly higher than 0 hour (no stress condition) in our research.
Flavonoids, widely distributed secondary metabolites, play important roles in plant growth and are considered as a major metabolite response to abiotic stress[34]. The 32 flavonoids detected in this study (Tables S2 and S3) can be categorized as one flavanone, five flavones, and 26 flavonols, which are generally consistent with the known composition of flavonoids in M. Pendula leaf reported before[13]. The antioxidant and free-radical-scavenging ability of flavonoids is attributed to their hydroxyl groups, double bounds and their predisposition to glycosylation, acylation, and methylation[35,36,37]. Yang et al.[38] reported that drought stress triggers biosynthesis of flavonoids in Bupleurum chinense leaves. Jan et al.[39] also observed that over-accumulation of non-enzymatic antioxidant flavonoids enhanced drought and UV radiation stress tolerance in rice. Recently, a basic leucine zipper (bZIP) transcription factor, FlbZIP12 from Fagopyrum leptopodum, was shown to enhance drought tolerance by modulating flavonoid biosynthesis[40]. In our study, compared to the untreated counterparts (0 hour), PEG-6000 treatment significantly increased the accumulation of flavonoids, potentially reducing ROS levels and enhancing mulberry leaf adaptability to drought stress. Our transcriptome analysis further revealed significant differences in the expression of key genes involved in flavonoid biosynthesis pre- and post-drought treatment, suggesting that flavonoids play a role in the drought tolerance of mulberry. Flavonoids have attracted more attention due to their diverse pharmacological effects. Controlled drought stress appears capable of enhancing the abundance of flavonoids, which could hold considerable medicinal value and broaden the utilization of mulberry leaves.
Through metabolome and transcriptome analysis performed in both drought-stressed and unstressed mulberry, a more comprehensive understanding of the response of the phenylpropanoids pathway to drought was obtained. The drought stress activated the biosynthesis of phenylpropanoids in mulberry leaves. However, despite naringenin being considered an intermediate and rapidly catalyzed[41] as the common precursor of the large number of downstream flavones and flavonols, its content remained at a relatively low and almost unchanged level (│log2FC│ < 0.35) throughout. Similarly, the detection of flavonols presented a comparable situation. While mulberry leaves has abundant O-glycosylated flavonols and malonylated flavonol glycosides, flavonol aglycones such as kaempferol and quercetin were never found in either previous study or the present work, implying their immediate conversion to downstream products and suggesting highly efficient in vivo synthesis of terminally modified flavonols[42,43]. Our findings indicate that the accumulation pattern of flavonoids under drought showed a strong correlation with the expression of key genes in the phenylpropanoids pathway, notably MaPAL1, MaPAL2, MAPAL3, MaCHS1, MaF3H, MaF3GT and MaUGT90A1. A similar trend was also reported in Achillea pachycephala.[21] However, not all gene expression coincides with the accumulation trend of flavonoids, such as MaC4H, MaF3′H1, MaF3G6″RT, and MaUGT76A2. A possible explanation for this observation might be a homeostatic regulation of phenylpropanoid pathway[44]. Furthermore, the metabolism of flavonoids is closely intertwined with metabolic signaling pathways that are activated in response to drought stress. A notable example is abscisic acid (ABA), a phytohormone associated with drought tolerance in plants. Gao et al.[9] have confirmed that the mutual regulation of ABA and flavonoids signaling contributed to the variation in drought resistance among different sea buckthorn subspecies. While the ABA content was not measured in this study, it is speculated that an increased ABA content would be evident based on findings from Liu’s research in mulberry[29].
The integration of multi-omics profiling and analysis has provided new insights on the biosynthesis studies of flavonoids and other bioactive compounds such as 1-deoxynojirimycin (DNJ) in mulberry[13,45]. This study not only investigated the biochemical response of flavonoids under drought but also endeavored to identify potential key genes involved in flavonoid biosynthesis. The enzyme FLS catalyzes the conversion of dihydroflavonol to flavonol, serving as the initial key enzyme in the flavonol pathway. MaFLS1 protein characterized in this study catalyzed the conversion of dihydroflavonols to flavonols, potentially facilitating the enrichment of flavonols intermediator such as kaempferol and quercetin in mulberry. While a negative correlation was observed between the expression levels of MaFLS2 and MaFLS3 and flavoniods accumulation, the specific functions of these enzymes remained unexplored. Flavonols have various bioactivities beneficial to human health, however, their practical application is hindered by the high costs and extraction challenges. Compared to chemical synthesis and plant extraction, the idea of synthesizing flavonoids through fermentation using mulberry genes holds promise for producing secondary metabolites under controlled conditions[42]. This approach may potentially reduce production costs, avoid side effects, and mitigate environmental pollution problems[46].
Flavonoid related UGTs in plants usually shows strong substrate specificity[47]. In our study, the MaF3GT5 protein catalyzed the conversion of flavonol aglycones (e.g., kaempferol and quercetin) to flavonol 3-O-glucoside and flavonol 7-O-glucoside, but it did not utilize 3-O- and 7-O-glucosylated flavonols or UDP-rhamnose as potential substrates. Similarly, FaGT7 protein from strawberry (Fragaria x ananassa) primarily catalyzed the glycosylation at the 3-OH or 4′-OH position of kaempferol and quercetin, but it did not directly mediate the synthesis of flavonol di-O-glucoside[48]. A novel anthocyanidin 3-O-glucoside-2″-O-glucosyltransferase (In3GGT) was also reported in Japanese morning glory (Ipomoea nil), which recognized anthocyanidin 3-O-glucoside as substrates but not anthocyanidin 3, 5-di-O-glucoside[49]. Moreover, a genetic study of AtUGT89C1 and AtUGT79B6 in Arabidopsis observed that flavonol 7-O-rhamnosylation occurs after 3-O-glycosylation and glycosylation at 5-0H or 7-OH may occur after full modification at 3-OH[50,51], suggesting that several, as-yet uncharacterized MaUGTs involved in flavonol 3, 7-di-O-glucoside biosynthesis are encoded in the mulberry genome. Unlike the characterization of MaFLS1, fermentation test was not performed in the functional study of MaF3GT5 protein duo to its lower efficiency of flavonoid production (Figure 6). To our knowledge, MaF3GT5 is the first UDP-glucosyltransferase identified in mulberry that forms 3-O- and 7-O-monoglucoside but not di-O-glucosides from flavonols such as kaempferol. Besides, flavonoid biosynthetic and modification enzymes, such as flavonoid 3-O-glucoside-7-O-glucosyltransferase (3GGT) in mulberry, remains to be elucidated in further research.
4. Material and Methods
4.1. Plant Materials and Drought Treatment
The seeds of M. Pendula were initially stratified in bottles with water at 4°C for 48 hours to break dormancy. After vernalization, the seeds were disinfected with 0.1% HgCl2 for 10 min, followed by three washes with sterilized distilled water. Subsequently, the treated seeds were cultured in a modified Hoagland solution within a growth chamber under constant conditions (16 h light/8 h dark photoperiod, 70% relative humidity, and a culture temperature of 25°C). When the seedlings reached 45 days, they were transferred to a Hoagland solution medium supplemented with 30% PEG-6000 to simulate drought stress. Leaf samples were then collected at 0, 24, 48, and 72 hours of continuous treatment and immediately stored at −80°C. Throughout the sampling period, regular water supply was maintained to ensure a consistent concentration of PEG-6000. To assess the impact of drought stress on the mulberry plants, the concentration of superoxide dismutase (SOD), malondialdehyde (MDA), and catalase (CAT) were analyzed after treatment. The content of SOD, MDA, and CAT were measured using the Grace reagent kit (Suzhou Grace Biotechnolgy Co., Ltd., Jiangsu, China).
4.2. Sample Extraction for Metabolomic Analysis
The frozen mulberry leaves were pulverized into powder using a mixer mill (MM400, Retsch, Haan, Germany) with a zirconia bead for 1.5 min at 30 Hz. To extract flavonoids, 100 mg of the powder was weighed and then subjected to extraction with 0.5 ml 70% (v/v) aqueous methanol for 8 h (or overnight) at 4°C. After centrifuged at 11,000 g for 10 min, 0.4 ml of the supernatant was filtered through a 0.22 μm filter (SCAA-104, ANPEL, Shanghai, China) and kept in 1.5 ml chromatographic sample bottles before LC-MS/MS analysis.
4.3. UHPLC-MS/MS Analysis Conditions
The sample extracts were analyzed using an LC-ESI-MS/MS system (UHPLC, Thermo Scientific™ Dionex™ UltiMate™ 3000; MS, Q Exactive hybrid quadrupole-orbitrap mass spectrometer; Thermo Fisher Scientific, Waltham, MA, USA). The UHPLC conditions were as follows: column, Aquity UPLC-BEH-C18 (1.7 μm particle size, length 2.1×150 mm); solvent system, mobile phase A: ultrapure water (0.04% acetic acid), mobile phase B: acetonitrile (0.04% acetic acid); gradient program, 95:5 VA/VB at 0 min, 5:95 VA/VB at 20.0 min, 5:95 VA/VB at 22.0 min, 95:5 VA/VB at 22.1 min, 95:5 VA/VB at 26.0 min; flow rate, 0.25 ml/min; column temperature, 40°C; injection volume, 5μl. The analytical MS/MS conditions were as follows: ESI source operation parameters: sheath gas, 35 arbitrary units; auxiliary gas, 10 arbitrary units; sweep gas, 0 arbitrary units; spray voltage, 3.5KV; capillary temperature, 350°C; and S-lens RF level, 50. The full MS parameters were as follows: MS scan range, 100-1,000 m/z; resolution, 70,000; microscans, 1; automatic gain control (AGC) target, 1e6; Max IT, 200 ms. The data-dependent MS2 (dd-MS2) quantification method parameters were as follows: resolution, 35,000; microscans, 1; AGC target, 2e4; Max IT, 100 ms; loop count, 5; topN, 5; isolation window, 1.0 m/z; (N)CE: 15, 30, 60; apex-trigger, 2-6s. Instrument tuning and mass calibration were performed with PierceTM LTQ Velos ESI positive ion calibration solution (Pierce, Rockford, IL, USA).
4.4. Transcriptome Sequencing, Assembly, and Analysis
Libraries were constructed from mulberry leaf mRNA and subjected to sequencing using Illumina HiSeqTM 2500 platform by Gene Denovo Biotechnology Co. (Guangzhou, China). High-quality clean reads were obtained after removing sequences containing adapters, sequences with all A bases, sequences with more than 10% of unknown nucleotides (N), and sequences with over 50% of low-quality bases (Q-value ≤ 20). The reads, devoid of RNA (rRNA), from each sample were aligned to the reference genome using TopHat2. Transcript reconstruction was performed using the Cufflinks software. Gene expression levels were normalized using the FPKM (fragments per kilobase of transcript per million mapped reads) method. Significantly differentially expressed genes (DEGs) were selected based on a fold change ≥ 2 and a false discovery rate (FDR) < 0.05. The DEGs were further subjected to GO functions and KEGG pathways enrichment analyses, with thresholds of P ≤ 0.01 and FDR ≤ 0.05 for both analyses. Unigenes were annotated by conducting BLASTx searches against the NCBI non-redundant protein (Nr) database, the Swiss-Prot protein database, the Kyoto Encyclopedia of Genes and Genomes (KEGG) database, and the COG/KOG databases, utilizing an E-value of 1e-5. Protein functional annotations were obtained according to the best alignment results.
4.5. Quantitative Real-Time PCR
Quantitative real-time PCR (qRT-PCR) was conducted using the Step One real-time PCR system (Applied Biosystems, Foster City, CA, USA). Gene-specific primers were designed by Primer Premier 5 software and listed in Table S1. Each qRT-PCR reaction was performed in a 20 μl mixture consisting of 1 μl of cDNA as template, 10 μl 2 × QuantiNova SYBR Green PCR Master Mix, 2 μl Rox dye (QiagenGmBH, Hilden, Germany), 0.6 μl of each primer, and DEPC-treated water. The amplification program consisted of an initial denaturation at 95°C for 15 min, followed by 41 cycles of denaturation at 95°C for 20 s, annealing at 55°C for 40 s, and extension at 72°C for 30 s. The 2−ΔΔCt method was used to calculate the relative expression levels of the genes. All reported data are presented as means ± SD (n= 3).
4.6. Production of Recombinant Protein, In Vitro Enzyme Assay, and Fermentation Test
The full-length cDNA of MaFLS1 and MaF3GT5 were cloned into the pcold-TF expression vector and expressed as recombinant proteins in Escherichia coli strain BL21 (DE3). The bacterial culture was incubated at 37°C until reaching an optical density at 600 nm of 0.6, followed by induction with a final concentration of 0.1 mM isopropyl-β-D-thiogalactopyranoside (IPTG) and further incubation at 25°C for 8 h.
For the in vitro enzyme assays, cells were collected by centrifugation, and the fusion protein was purified using high-affinity Ni-NTA resin (L00250, GenScript, Nanjing, China) according to the manufacturer’s instructions. In the MaFLS1 reaction, 30 µl of purified MaFLS1 protein was dissolved in 100 mM phosphate buffer (pH 6.8) containing 100 µM dihydroflavonols substrate, 10 mM ascorbic acid, 10 mM 2-oxoglutarate, and 0.25 mM ferrous sulfate (FeSO4). In the MaF3GT5 reaction, 30 µl purified MaF3GT5 protein was dissolved in phosphate buffer saline (pH = 7.3) containing 100 µM flavonoid substrate, 100 µM UDP-glucose, and 100 µM UDP-rhamnose. Both reactions were incubated at 28°C for 30 min with continuous agitation. The fermentation process mirrored that previously utilized for characterizing MaCHS[42]. Cells were harvested by centrifugation, resuspended in M9 medium containing 0.1 mM IPTG, 0.1 mg/ml ampicillin, and 1 mM dihydroflavonols, and incubated with shaking at 25°C for 48 h. Flavonoids in the resulting solution and the final fermentation product were extracted with 70% (v/v) aqueous methanol, filtered through a 0.22 μm filter, and subjected to LC-MS/MS analysis.
4.7. Chemicals and Standards
All solvents and reagents used in this study were of analytical grade. Methanol, acetonitrile, and acetic acid (HPLC grade) were purchased from Thermo-Fisher Scientific (Shanghai, China). Authentic standards including kaempferol, quercetin, 3-O- and 7-O-glycosylated flavonols, kaempferol 3-O-rutinoside (K3R), and rutin were purchased from Chroma Biotechnology Co., Ltd. (Chengdu, China). Dihydrokaempferol and dihydroquercetin were purchased from DeSiTe Biological Technology Co., Ltd. (Chengdu, China). Other standards were purchased from Sigma-Aldrich (Shanghai, China). UDP-glucose and UDP-rhamnose were purchased from EnzymeWorks Inc. (Jiangsu, China).
4.8. Statistical Analyses
The metabolome and RNA-seq data were analyzed using R software, incorporating heatmap analysis, principal component analysis (PCA), and multiple testing. Figures were generated with SigmaPlot 12.0 (Systat, CA, USA) and Adobe Illustrator CS6 (Adobe, CA, USA). The results are presented as means ± standard deviation (SD). The calculated p-value was calibrated using false discovery rate (FDR) correction.
5. Conclusion
In this study, we conducted a comprehensive investigation of the biochemical response of flavonoids in mulberry leaves under drought conditions through multi-omics profiling and analysis. Our findings have enhanced our understanding of the phenylpropanoid pathway in mulberry. Specifically, we observed that drought stress led to a significant increase in the activity of protective enzymes, upregulated the expression of genes involved in the phenylpropanoid pathway, and stimulated the biosynthesis of flavonoids. Additionally, we identified and characterized one MaFLS and three flavonoid-related UPD-glycosyltransferases by analyzing transcriptome data. The recombinant MaFLS1 and MaF3GT5 protein were found to catalyze the synthesis of flavonol aglycones and flavonol O-monoglucoside, respectively. These results represent a significant advancement in our knowledge of flavonol biosynthesis in mulberry and provide valuable insights into the biosynthesis of other bioactive compounds.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Figure S1: Metabolic and transcriptional analysis of M. Pendula leaves under drought stress. Figure S2: Non-rooted molecular phylogenetic tree of flavonoid-related UDP-glycosyltransferase. Figure S3: Bioinformatic analysis of MaFLS1. Table S1: List of primer sequences used in this study. Table S2: Checklist of flavonoids measured in this study and recommendations for LC-MS/MS. Table is on a separate sheet. Table S3: Metabolic profiling of flavonoids in M. Pendula leaves under drought stress. Table is on a separate sheet. Table S4: Statistical results of RNA-seq assembly data. Table S5: Details of significantly changed genes involved in flavonoid biosynthesis in this study.
Authors’ Contributions
NH designed the experiments and revised the manuscript; GC performed the drought treatment and RNA-Seq data analysis; DL and DB performed the metabolic profiling analysis; PY and FC performed the fermentation test and in vitro characterization; JY prepared the plant materials; BM provided the R analysis; ZY performed the expression analysis; DL and GC wrote the manuscript. All authors have read and approved the final manuscript.
Funding
This work was funded by the National Key Research and Development Program of China (No.2022YFD1201602), the Innovation Research 2035 Pilot Plan of Southwest University (SWU-XDZD22008), the Fundamental Research Funds for the Central Universities (swu120028), and the Chongqing Research Program of Basic Research and Frontier Technology (No. cstc2021yszx-jcyj0004).
Notes
The authors declare no competing financial interest.
References
- Bhaskarla, V.; Zinta, G.; Ford, R.; Jain, M.; Varshney, R.K.; Mantri, N. Comparative Root Transcriptomics Provide Insights into Drought Adaptation Strategies in Chickpea (Cicer arietinum L.). Int. J. Mol. Sci. 2020, 21, 1781, . [CrossRef]
- Zhang, Q.; Han, L.; Jia, J.; Song, L.; Wang, J.S. Management of drought risk under global warming. Theor. Appl. Climatol. 2016, 125, 187–196.
- Yu, F.; Wan, W.; Lv, M.-J.; Zhang, J.-L.; Meng, L.-S. Molecular Mechanism Underlying the Effect of the Intraspecific Alternation of Seed Size on Plant Drought Tolerance. J. Agric. Food Chem. 2020, 68, 703–711, . [CrossRef]
- Wang, J.; Zhang, L.; Cao, Y.; Qi, C.; Li, S.; Liu, L.; Wang, G.; Mao, A.; Ren, S.; Guo, Y.-D. CsATAF1 Positively Regulates Drought Stress Tolerance by an ABA-Dependent Pathway and by Promoting ROS Scavenging in Cucumber. Plant Cell Physiol. 2018, 59, 930–945, . [CrossRef]
- Cao, P.; Yang, J.; Xia, L.; Zhang, Z.; Wu, Z.; Hao, Y.; Liu, P.; Wang, C.; Li, C.; Yang, J.; et al. Two gene clusters and their positive regulator SlMYB13 that have undergone domestication-associated negative selection control phenolamide accumulation and drought tolerance in tomato. Mol. Plant 2024, 17, 579–597, . [CrossRef]
- Scharwies, J. D.; Dinneny, J. R., Water transport, perception, and response in plants. J Plant Res 2019, 132, (3), 311-324.
- Li, S.; Li, X.; Wei, Z.; Liu, F. ABA-mediated modulation of elevated CO2 on stomatal response to drought. Curr. Opin. Plant Biol. 2020, 56, 174–180, . [CrossRef]
- Guha, A.; Rasineni, G.K.; Reddy, A.R. DROUGHT TOLERANCE IN MULBERRY (MORUS SPP.): A PHYSIOLOGICAL APPROACH WITH INSIGHTS INTO GROWTH DYNAMICS AND LEAF YIELD PRODUCTION. Exp. Agric. 2010, 46, 471–488, . [CrossRef]
- Gao, G.; Lv, Z.; Zhang, G.; Li, J.; Zhang, J.; He, C. An ABA–flavonoid relationship contributes to the differences in drought resistance between different sea buckthorn subspecies. Tree Physiol. 2021, 41, 744–755, . [CrossRef]
- Corso, M.; Vannozzi, A.; Maza, E.; Vitulo, N.; Meggio, F.; Pitacco, A.; Telatin, A.; D’angelo, M.; Feltrin, E.; Negri, A.S.; et al. Comprehensive transcript profiling of two grapevine rootstock genotypes contrasting in drought susceptibility links the phenylpropanoid pathway to enhanced tolerance. J. Exp. Bot. 2015, 66, 5739–5752, . [CrossRef]
- He, N.; Zhang, C.; Qi, X.; Zhao, S.; Tao, Y.; Yang, G.; Lee, T.-H.; Wang, X.; Cai, Q.; Li, D.; et al. Draft genome sequence of the mulberry tree Morus notabilis. Nat. Commun. 2013, 4, 2445. [CrossRef]
- Srivastava, S.; Kapoor, R.; Thathola, A.; Srivastava, R. P., Mulberry (Morus alba) leaves as human food: a new dimension of sericulture. Int. J. Food Sci. Nutr. 2003, 54, (6), 411-416.
- Li, D.; Chen, G.; Ma, B.; Zhong, C.; He, N. Metabolic Profiling and Transcriptome Analysis of Mulberry Leaves Provide Insights into Flavonoid Biosynthesis. J. Agric. Food Chem. 2020, 68, 1494–1504, . [CrossRef]
- Hunyadi, A.; Liktor-Busa, E.; Márki, .; Martins, A.; Jedlinszki, N.; Hsieh, T.J.; Báthori, M.; Hohmann, J.; Zupkó, I. Metabolic Effects of Mulberry Leaves: Exploring Potential Benefits in Type 2 Diabetes and Hyperuricemia. Evidence-Based Complement. Altern. Med. 2013, 2013, 1–10, . [CrossRef]
- Wen, P.; Hu, T.-G.; Linhardt, R.J.; Liao, S.-T.; Wu, H.; Zou, Y.-X. Mulberry: A review of bioactive compounds and advanced processing technology. Trends Food Sci. Technol. 2019, 83, 138–158, . [CrossRef]
- Chaitanya, K.V.; Rasineni, G.K.; Reddy, A.R. Biochemical responses to drought stress in mulberry (Morus alba L.): evaluation of proline, glycine betaine and abscisic acid accumulation in five cultivars. Acta Physiol. Plant. 2009, 31, 437–443, . [CrossRef]
- Li, H.; Li, D.; Yang, Z.; Zeng, Q.; Luo, Y.; He, N. Flavones Produced by Mulberry Flavone Synthase Type I Constitute a Defense Line against the Ultraviolet-B Stress. Plants 2020, 9, 215, . [CrossRef]
- Katsube, T.; Imawaka, N.; Kawano, Y.; Yamazaki, Y.; Shiwaku, K.; Yamane, Y. Antioxidant flavonol glycosides in mulberry (Morus alba L.) leaves isolated based on LDL antioxidant activity. Food Chem. 2006, 97, 25–31, . [CrossRef]
- Chen, A.Y.; Chen, Y.C. A review of the dietary flavonoid, kaempferol on human health and cancer chemoprevention. Food Chem. 2013, 138, 2099–2107, doi:10.1016/j.foodchem.2012.11.139.
- Zhao, S.; Park, C.H.; Li, X.; Kim, Y.B.; Yang, J.; Sung, G.B.; Park, N.I.; Kim, S.; Park, S.U. Accumulation of Rutin and Betulinic Acid and Expression of Phenylpropanoid and Triterpenoid Biosynthetic Genes in Mulberry (Morus alba L.). J. Agric. Food Chem. 2015, 63, 8622–8630, . [CrossRef]
- Gharibi, S.; Tabatabaei, B.E.S.; Saeidi, G.; Talebi, M.; Matkowski, A. The effect of drought stress on polyphenolic compounds and expression of flavonoid biosynthesis related genes in Achillea pachycephala Rech.f. Phytochemistry 2019, 162, 90–98, . [CrossRef]
- Li, B.; Fan, R.; Sun, G.; Sun, T.; Fan, Y.; Bai, S.; Guo, S.; Huang, S.; Liu, J.; Zhang, H.; et al. Flavonoids improve drought tolerance of maize seedlings by regulating the homeostasis of reactive oxygen species. Plant Soil 2021, 461, 389–405, . [CrossRef]
- Niu, Y.; Li, J.; Sun, F.; Song, T.; Han, B.; Liu, Z.; Su, P. Comparative transcriptome analysis reveals the key genes and pathways involved in drought stress response of two wheat (Triticum aestivum L) varieties. Genomics 2023, 115, 110688, . [CrossRef]
- Lucci, N.; Mazzafera, P. Distribution of rutin in fava d'anta (Dimorphandra mollis) seedlings under stress. J. Plant Interactions 2009, 4, 203–208, . [CrossRef]
- Li, D.; Ma, B.; Xu, X.; Chen, G.; Li, T.; He, N. MMHub, a database for the mulberry metabolome. Database 2020, 2020, . [CrossRef]
- Liu, F. R.; Xie, L. F.; Yao, Z. Y.; Zhou, Y. L.; Zhou, W. F.; Wang, J. H.; Sun, Y. Y.; Gong, C. M., Caragana korshinskii phenylalanine ammonialyase is up-regulated in the phenylpropanoid biosynthesis pathway in response to drought stress. Biotechnol Biotec Eq 2019, 33, (1), 842-854.
- Li, M.; Li, H.; Sun, A.; Wang, L.; Ren, C.; Liu, J.; Gao, X. Transcriptome analysis reveals key drought-stress-responsive genes in soybean. Front. Genet. 2022, 13, 1060529, . [CrossRef]
- Cao, X.; Hu, Y.; Song, J.; Feng, H.; Wang, J.; Chen, L.; Wang, L.; Diao, X.; Wan, Y.; Liu, S.; et al. Transcriptome Sequencing and Metabolome Analysis Reveals the Molecular Mechanism of Drought Stress in Millet. Int. J. Mol. Sci. 2022, 23, 10792, . [CrossRef]
- Liu, H.; Sun, H.; Bao, L.; Han, S.; Hui, T.; Zhang, R.; Zhang, M.; Su, C.; Qian, Y.; Jiao, F. Secondary Metabolism and Hormone Response Reveal the Molecular Mechanism of Triploid Mulberry (Morus Alba L.) Trees Against Drought. Front. Plant Sci. 2021, 12, . [CrossRef]
- Yang, J.; Chen, R.; Wang, C.; Li, C.; Ye, W.; Zhang, Z.; Wang, S. A widely targeted metabolite modificomics strategy for modified metabolites identification in tomato. J. Integr. Plant Biol. 2024, . [CrossRef]
- Prescott, A.G.; Stamford, N.P.; Wheeler, G.; Firmin, J.L. In vitro properties of a recombinant flavonol synthase from Arabidopsis thaliana. Phytochemistry 2002, 60, 589–593, . [CrossRef]
- Polturak, G.; Heinig, U.; Grossman, N.; Battat, M.; Leshkowitz, D.; Malitsky, S.; Rogachev, I.; Aharoni, A. Transcriptome and Metabolic Profiling Provides Insights into Betalain Biosynthesis and Evolution in Mirabilis jalapa. Mol. Plant 2018, 11, 189–204, . [CrossRef]
- Guo, Y.; Li, D.; Liu, L.; Sun, H.; Zhu, L.; Zhang, K.; Zhao, H.; Zhang, Y.; Li, A.; Bai, Z.; et al. Seed Priming With Melatonin Promotes Seed Germination and Seedling Growth of Triticale hexaploide L. Under PEG-6000 Induced Drought Stress. Front. Plant Sci. 2022, 13, 932912, . [CrossRef]
- Sharma, A.; Shahzad, B.; Rehman, A.; Bhardwaj, R.; Landi, M.; Zheng, B. Response of Phenylpropanoid Pathway and the Role of Polyphenols in Plants under Abiotic Stress. Molecules 2019, 24, 2452, doi:10.3390/molecules24132452.
- Kim, G. N.; Jang, H. D., Flavonol Content in the Water Extract of the Mulberry (Morus alba L.) Leaf and Their Antioxidant Capacities. J Food Sci 2011, 76, (6), C869-C873.
- Nouraei, S.; Rahimmalek, M.; Saeidi, G., Variation in polyphenolic composition, antioxidants and physiological characteristics of globe artichoke (Cynara cardunculus var. scolymus Hayek L.) as affected by drought stress. Sci Hortic-Amsterdam 2018, 233, 378-385.
- Ju, W.-T.; Kwon, O.-C.; Kim, H.-B.; Sung, G.-B.; Kim, H.-W.; Kim, Y.-S. Qualitative and quantitative analysis of flavonoids from 12 species of Korean mulberry leaves. J. Food Sci. Technol. 2018, 55, 1789–1796, . [CrossRef]
- Yang, L.-L.; Yang, L.; Yang, X.; Zhang, T.; Lan, Y.-M.; Zhao, Y.; Han, M.; Yang, L.-M. Drought stress induces biosynthesis of flavonoids in leaves and saikosaponins in roots of Bupleurum chinense DC. Phytochemistry 2020, 177, 112434, doi:10.1016/j.phytochem.2020.112434.
- Jan, R.; Khan, M.-A.; Asaf, S.; Lubna; Waqas, M.; Park, J.-R.; Asif, S.; Kim, N.; Lee, I.-J.; Kim, K.-M. Drought and UV Radiation Stress Tolerance in Rice Is Improved by Overaccumulation of Non-Enzymatic Antioxidant Flavonoids. Antioxidants 2022, 11, 917, . [CrossRef]
- Wang, A.; Liu, Y.; Li, Q.; Li, X.; Zhang, X.; Kong, J.; Liu, Z.; Yang, Y.; Wang, J. FlbZIP12 gene enhances drought tolerance via modulating flavonoid biosynthesis in Fagopyrum leptopodum. Front. Plant Sci. 2023, 14, 1279468, . [CrossRef]
- Wang, Y.; Chen, S.; Yu, O. Metabolic engineering of flavonoids in plants and microorganisms. Appl. Microbiol. Biotechnol. 2011, 91, 949–956, . [CrossRef]
- Wang, C.; Zhi, S.; Liu, C.; Xu, F.; Zhao, A.; Wang, X.; Tang, X.; Li, Z.; Huang, P.; Yu, M. Isolation and characterization of a novel chalcone synthase gene family from mulberry. Plant Physiol. Biochem. 2017, 115, 107–118, . [CrossRef]
- Ju, W.-T.; Kwon, O.-C.; Kim, H.-B.; Sung, G.-B.; Kim, H.-W.; Kim, Y.-S. Qualitative and quantitative analysis of flavonoids from 12 species of Korean mulberry leaves. J. Food Sci. Technol. 2018, 55, 1789–1796, . [CrossRef]
- Shi, J.; Yan, X.; Sun, T.; Shen, Y.; Shi, Q.; Wang, W.; Bao, M.; Luo, H.; Nian, F.; Ning, G. Homeostatic regulation of flavonoid and lignin biosynthesis in phenylpropanoid pathway of transgenic tobacco. Gene 2022, 809, 146017, . [CrossRef]
- Yang, Z.; Luo, Y.; Xia, X.; He, J.; Zhang, J.; Zeng, Q.; Li, D.; Ma, B.; Zhang, S.; Zhai, C.; et al. Dehydrogenase MnGutB1 catalyzes 1-deoxynojirimycin biosynthesis in mulberry. Plant Physiol. 2023, 192, 1307–1320, . [CrossRef]
- Mavel, S.; Dikic, B.; Palakas, S.; Emond, P.; Greguric, I.; de Gracia, A. G.; Mattner, F.; Garrigos, M.; Guilloteau, D.; Katsifis, A., Synthesis and biological evaluation of a series of flavone derivatives as potential radioligands for imaging the multidrug resistance-associated protein 1 (ABCC1/MRP1). Bioorgan Med Chem 2006, 14, (5), 1599-1607.
- Guo, H.; Cao, P.; Wang, C.; Lai, J.; Deng, Y.; Li, C.; Hao, Y.; Wu, Z.; Chen, R.; Qiang, Q.; et al. Population analysis reveals the roles of DNA methylation in tomato domestication and metabolic diversity. Sci. China Life Sci. 2023, 66, 1888–1902, . [CrossRef]
- Griesser, M.; Vitzthum, F.; Fink, B.; Bellido, M. L.; Raasch, C.; Munoz-Blanco, J.; Schwab, W., Multi-substrate flavonol O-glucosyltransferases from strawberry (Fragaria x ananassa) achene and receptacle. J Exp Bot 2008, 59, (10), 2611-2625.
- Morita, Y.; Hoshino, A.; Kikuchi, Y.; Okuhara, H.; Ono, E.; Tanaka, Y.; Fukui, Y.; Saito, N.; Nitasaka, E.; Noguchi, H.; Iida, S., Japanese morning glory dusky mutants displaying reddish-brown or purplish-gray flowers are deficient in a novel glycosylation enzyme for anthocyanin biosynthesis, UDP-glucose:anthocyanidin 3-O-glucoside-2″-O-glucosyltransferase, due to 4-bp insertions in the gene. Plant J. 2005, 42, (3), 353-363.
- Yonekura-Sakakibara, K.; Nakabayashi, R.; Sugawara, S.; Tohge, T.; Ito, T.; Koyanagi, M.; Kitajima, M.; Takayama, H.; Saito, K., A flavonoid 3-O-glucoside: 2”-O-glucosyltransferase responsible for terminal modification of pollen-specific flavonols in Arabidopsis thaliana. Plant J. 2014, 79, (5), 769-782.
- Yonekura-Sakakibara, K.; Tohge, T.; Niida, R.; Saito, K. Identification of a Flavonol 7-O-Rhamnosyltransferase Gene Determining Flavonoid Pattern in Arabidopsis by Transcriptome Coexpression Analysis and Reverse Genetics. J. Biol. Chem. 2007, 282, 14932–14941, . [CrossRef]
Figure 1.
Variation in catalase (CAT), superoxide dismutase (SOD), and malondialdehyde (MDA) levels at different hours under drought stress in Morus alba var. Pendula Dippel leaves. Data represent the mean of three replicates with standard deviation (± SD).
Figure 1.
Variation in catalase (CAT), superoxide dismutase (SOD), and malondialdehyde (MDA) levels at different hours under drought stress in Morus alba var. Pendula Dippel leaves. Data represent the mean of three replicates with standard deviation (± SD).
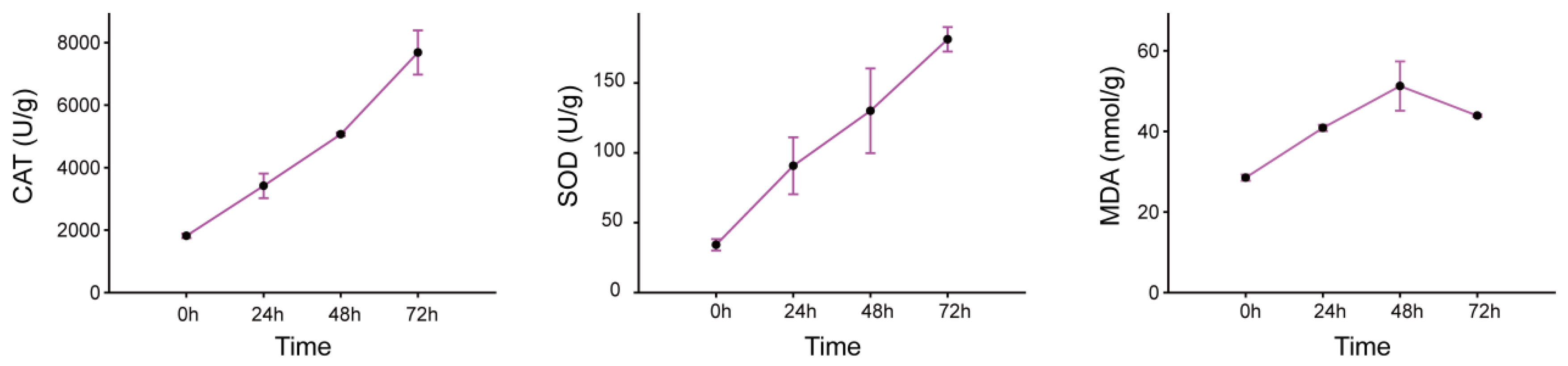
Figure 2.
Metabolic profiling and flavonoid variation in M. Pendula leaves under drought stress. (A) LC/MS profiling of flavonoids at 0, 24, 48, and 72 h in mulberry leaves under drought treatment, each with three biological replicates. The heat-map shows values displayed on log2 of relative peak area. (B) Fold changes of flavonoids at 24, 48, and 72 h. Increased flavonoids are indicated in green color, while decreased flavonoids are indicated in blue.
Figure 2.
Metabolic profiling and flavonoid variation in M. Pendula leaves under drought stress. (A) LC/MS profiling of flavonoids at 0, 24, 48, and 72 h in mulberry leaves under drought treatment, each with three biological replicates. The heat-map shows values displayed on log2 of relative peak area. (B) Fold changes of flavonoids at 24, 48, and 72 h. Increased flavonoids are indicated in green color, while decreased flavonoids are indicated in blue.
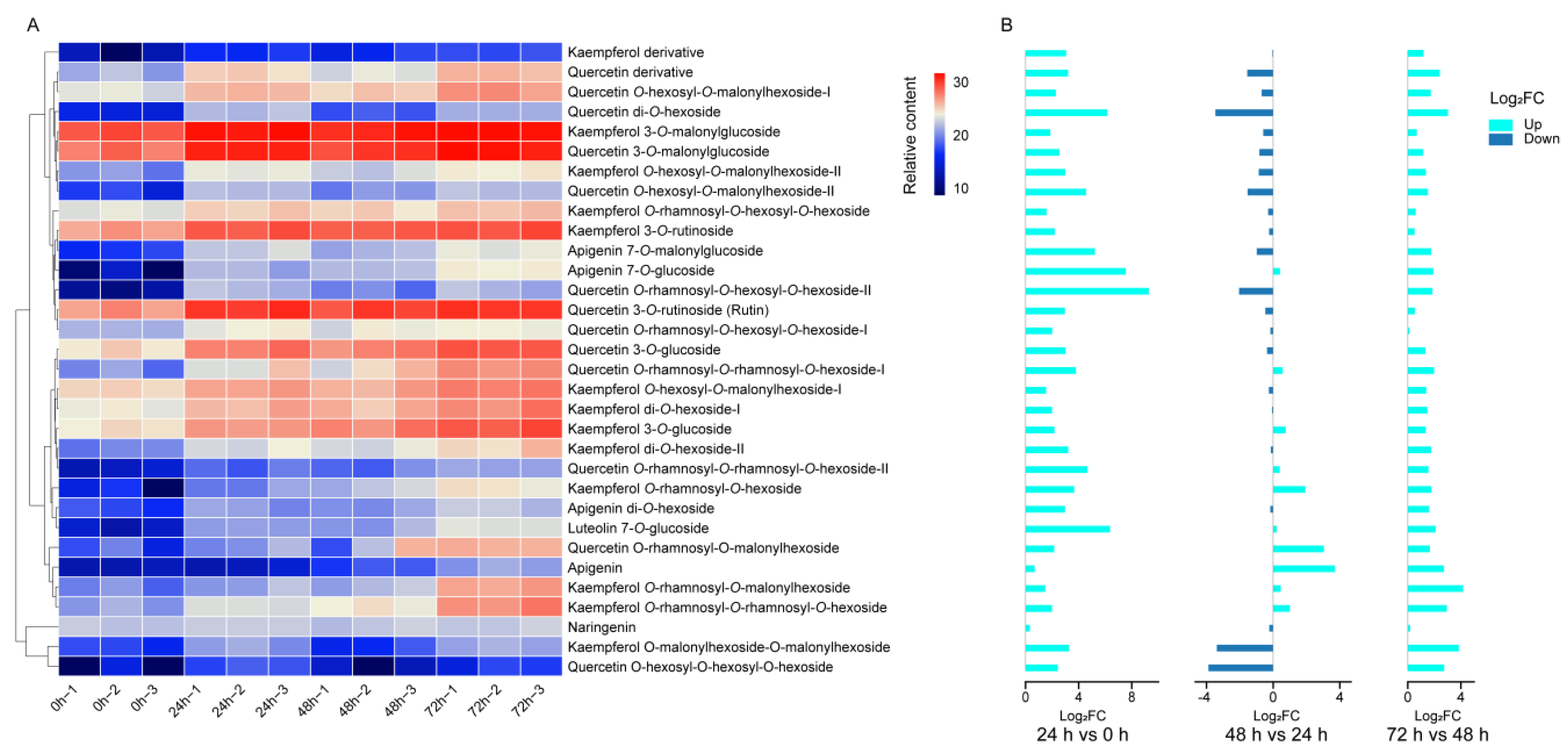
Figure 3.
The proposed flavonoid pathway and heat-map of key genes involved in the drought response of mulberry leaves. Colored blocks indicated different relative expression levels (from blue to red). C1 to C3 represents three replicates of the control, while D1 to D3 represents drought-treated group. Abbreviations: PAL, phenylalanine ammonia lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate-CoA ligase; CHS, chalcone synthase; CHI, chalcone isomerase; FNS, flavone synthase; F3H, flavanone 3-hydroxylase; F3′H, flavanone 3′-hydroxylase; FLS, flavonol synthase; F3GT, flavonoid 3-O-glycosyltransferase; F3G6”RT, flavonol 3-O-glucoside: 6″-O-rhamnosyltransferase; K3G, kaempferol 3-O-glucoside; Q3G, quercetin 3-O-glucoside; K3R, kaempferol 3-O-rutinoside.
Figure 3.
The proposed flavonoid pathway and heat-map of key genes involved in the drought response of mulberry leaves. Colored blocks indicated different relative expression levels (from blue to red). C1 to C3 represents three replicates of the control, while D1 to D3 represents drought-treated group. Abbreviations: PAL, phenylalanine ammonia lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate-CoA ligase; CHS, chalcone synthase; CHI, chalcone isomerase; FNS, flavone synthase; F3H, flavanone 3-hydroxylase; F3′H, flavanone 3′-hydroxylase; FLS, flavonol synthase; F3GT, flavonoid 3-O-glycosyltransferase; F3G6”RT, flavonol 3-O-glucoside: 6″-O-rhamnosyltransferase; K3G, kaempferol 3-O-glucoside; Q3G, quercetin 3-O-glucoside; K3R, kaempferol 3-O-rutinoside.
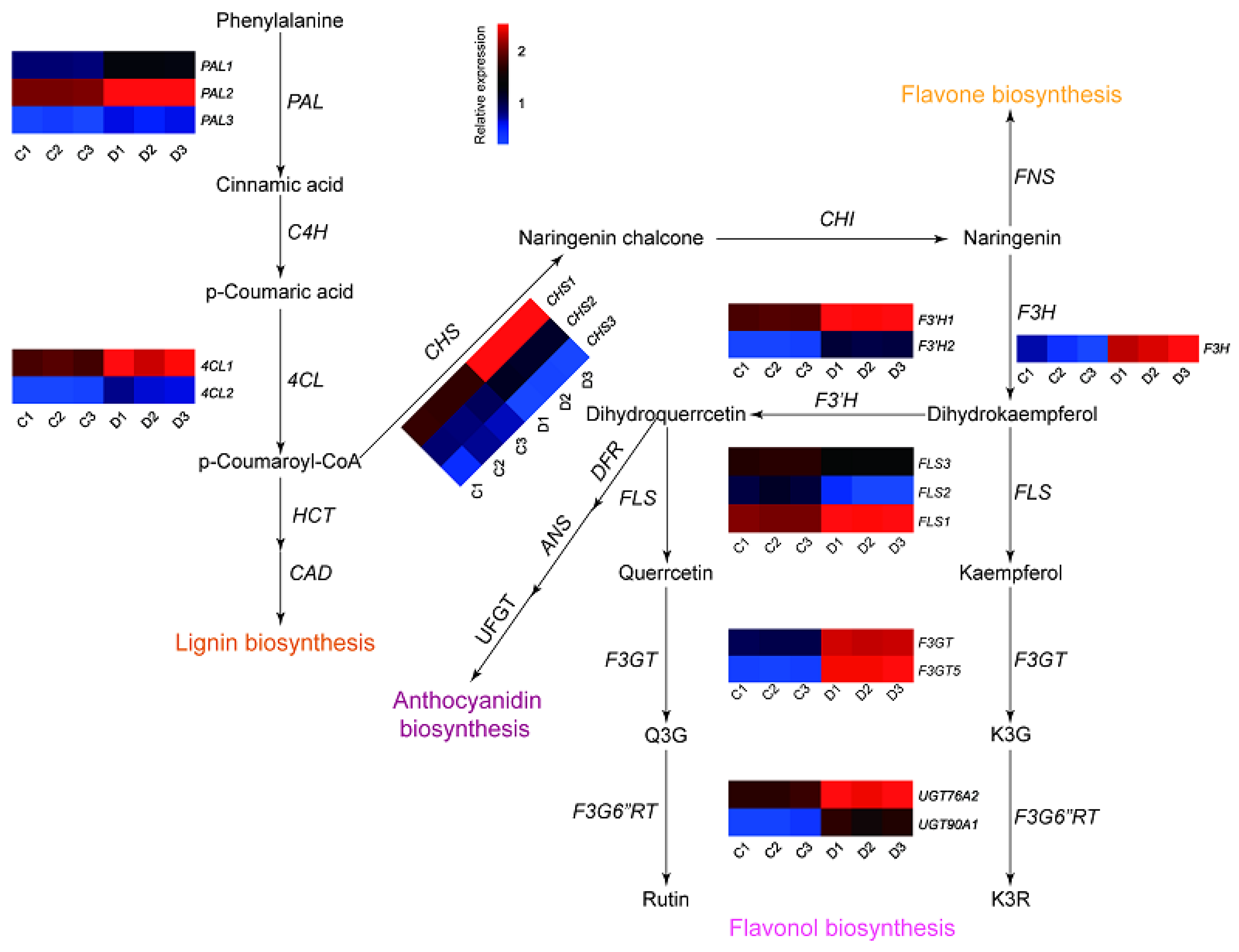
Figure 4.
Transcription levels of phenylpropanoid biosynthetic genes and candidate genes under PEG-6000 induced drought stress. Data represent the mean of three replicates with standard deviation (±SD).
Figure 4.
Transcription levels of phenylpropanoid biosynthetic genes and candidate genes under PEG-6000 induced drought stress. Data represent the mean of three replicates with standard deviation (±SD).
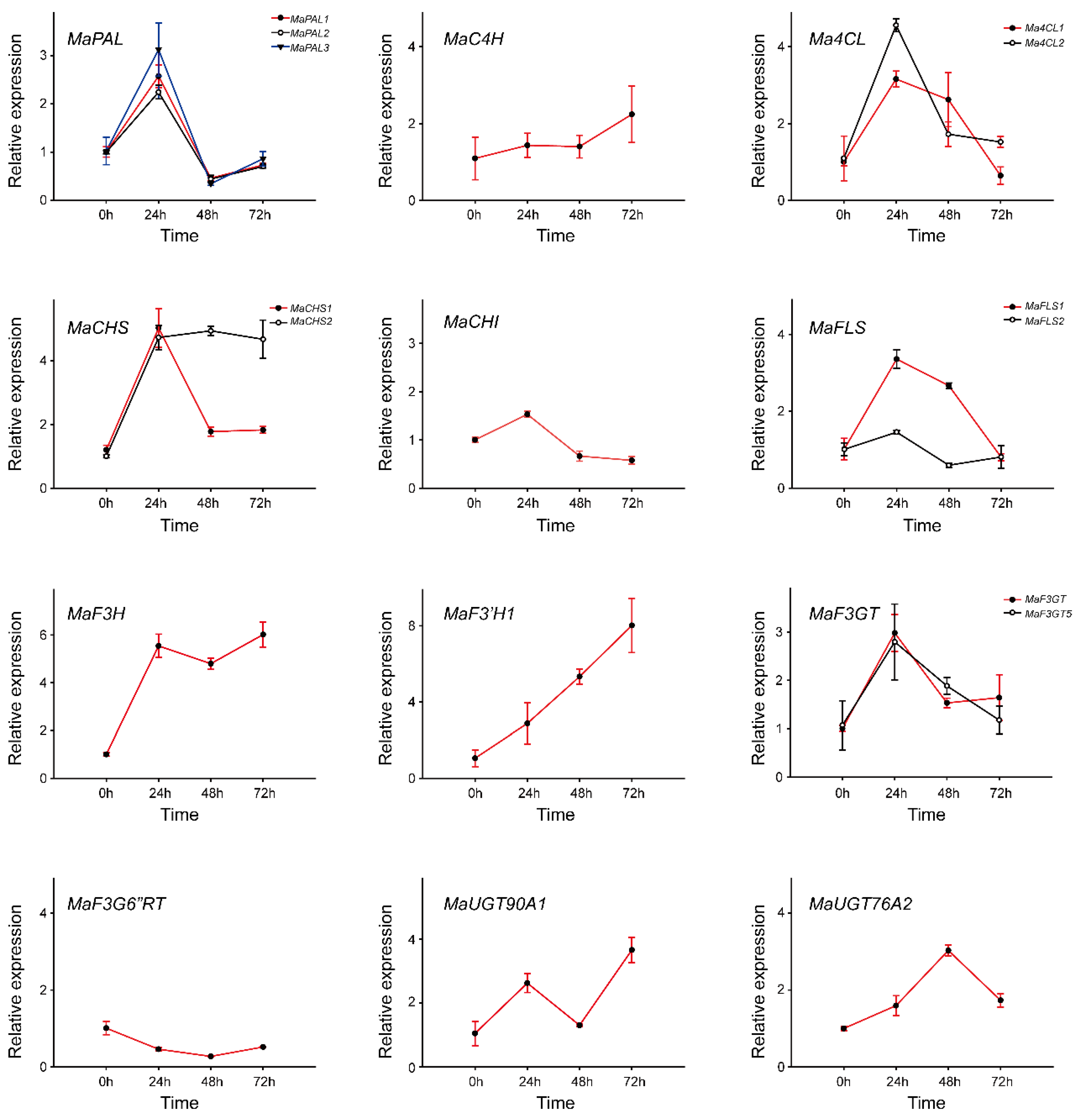
Figure 5.
UHPLC-MS/MS analyses of the reaction of recombinant MaFLS1 protein and fermented products. (A) Elution profile of the reaction products of His tag protein (empty vector), MaFLS1 protein (+MaFLS1), and authentic standards. The chromatograms of fermentation extracts present a similar pattern to the in vitro enzyme assay. (B) Extracted fragment mass chromatograms of reaction products (N1 and N2). Abbreviations: DHK, dihydrokaempferol; DHQ, dihydroquercetin; K, kaempferol; Q, quercetin.
Figure 5.
UHPLC-MS/MS analyses of the reaction of recombinant MaFLS1 protein and fermented products. (A) Elution profile of the reaction products of His tag protein (empty vector), MaFLS1 protein (+MaFLS1), and authentic standards. The chromatograms of fermentation extracts present a similar pattern to the in vitro enzyme assay. (B) Extracted fragment mass chromatograms of reaction products (N1 and N2). Abbreviations: DHK, dihydrokaempferol; DHQ, dihydroquercetin; K, kaempferol; Q, quercetin.
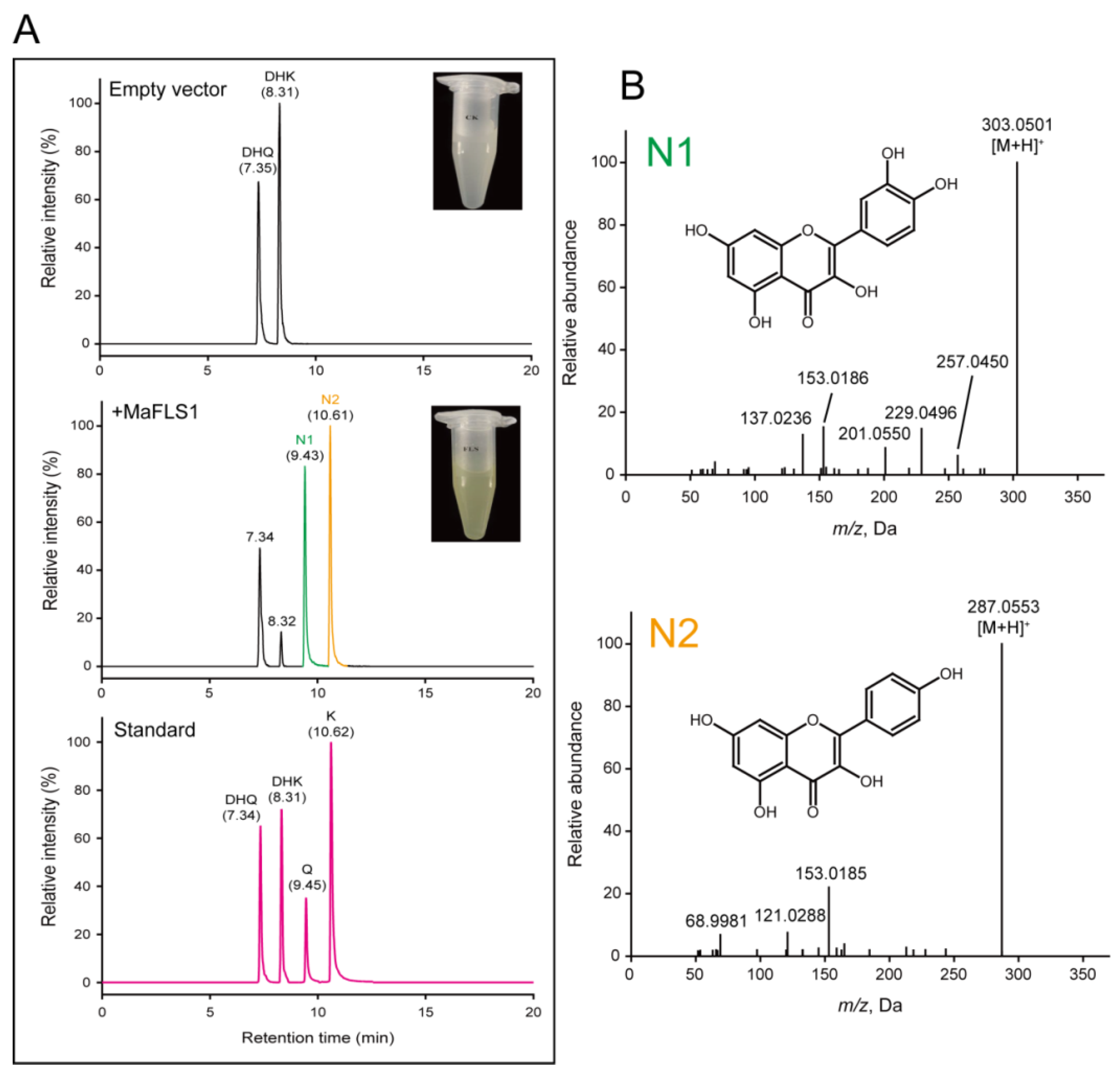
Figure 6.
HPLC-MS/MS analyses of the reaction of recombinant MaF3GT5 protein. The elution profile of the reaction products involving His-tag protein (empty vector), MaF3GT5 protein (+MaF3GT5), and chromatograms of standards (K, Q, K3G, Q3G, K7G, and Q7G) are shown on the left. Extracted fragment mass chromatograms of reaction products (F1, F2, and F3) are shown on the right. Abbreviations: K, kaempferol; Q, quercetin; K3G, kaempferol 3-O-glucoside; Q3G, quercetin 3-O-glucoside; K7G, kaempferol 7-O-glucoside; Q7G, quercetin 7-O-glucoside; -glu, natural loss of glucoside.
Figure 6.
HPLC-MS/MS analyses of the reaction of recombinant MaF3GT5 protein. The elution profile of the reaction products involving His-tag protein (empty vector), MaF3GT5 protein (+MaF3GT5), and chromatograms of standards (K, Q, K3G, Q3G, K7G, and Q7G) are shown on the left. Extracted fragment mass chromatograms of reaction products (F1, F2, and F3) are shown on the right. Abbreviations: K, kaempferol; Q, quercetin; K3G, kaempferol 3-O-glucoside; Q3G, quercetin 3-O-glucoside; K7G, kaempferol 7-O-glucoside; Q7G, quercetin 7-O-glucoside; -glu, natural loss of glucoside.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



