
Submitted:
27 May 2024
Posted:
27 May 2024
You are already at the latest version
Abstract
The yeast Saccharomyces cerevisiae ensures successful fermentation in winemaking, although the persistent use of commercial strains lead the loss of aroma complexity of wines. Hence, the research of indigenous S. cerevisiae with proper oenological features and well adapted to specific wine-growing area become of great interest for winemakers. Here, 206 S. cerevisiae strains were isolated from two wineries during two-year sampling campaign and bio-typed with the aim to evaluate the occurrence and persistence of S. cerevisiae wild population linked to each winery. Both wineries belonging to the same Verdicchio DOC wine area (Castelli di Jesi), and never used commercial yeasts during fermentation. Results showed a specific and varied population of S. cerevisiae in each winery without cross-contamination each other and with commercial starter strains. Moreover, inside each winery it was observed a persistent of some dominant biotypes over the time, indicating a sort of “winery-effect”. The evaluation of S. cerevisiae populations for the oenological characters showed a proper and well distinct aromatic imprinting on the resulted wines confirming the “winery effect”.
Keywords:
native Saccharomyces cerevisiae
; spontaneous fermentation
; resident yeasts
; winery-effect
1. Introduction
Saccharomyces cerevisiae represents the yeast mainly responsible for alcoholic fermentation in wine. The use of S. cerevisiae commercial active dry wine yeasts drives a vigorous fermentation process linked to a rapid start of fermentation and a complete consumption of fermentable sugars [1], ensuring uniform and constant quality of the wine over the time. On the other hand, pure fermentation could lead the loss of aromatic variability, complexity, and sensorial varietal aroma of wines [2]. For this reason, a great interest of winemakers is addressed to use of indigenous S. cerevisiae yeasts, that are well adapted to climatic conditions and grape varieties surviving during winemaking practices of a specific wine-growing area [3] with a possible enhancement of the typicity of wines. In this regard, spontaneous fermentation, that involves indigenous non-Saccharomyces yeasts, is conducted by several indigenous S. cerevisiae strains. It has been widely demonstrated that, although different indigenous S. cerevisiae strains are involved in the process, only few strains represent the dominant population of the fermentation process [4,5,6,7]. This advantage could be related to the highest competitivity of the dominant yeasts to survive under oenological conditions such as increasing ethanol concentrations, low pH values, temperature variations and cell-to-cell interactions with other yeasts [8,9]. Another feature of indigenous yeasts is the so called “winery effect” meaning that some indigenous S. cerevisiae strains may persist in the same winery for several consecutive years characterising the wines of the territory [6,7,10]. Probably this is due to the better adaptation of these resident yeasts to a specific winery’s winemaking managements, including climate and agricultural practices. They play a key role in determining organoleptic properties on the wines and to be representative to a single winery [6]. Aspects that reinforce the ascertained close relationship between the provenience of indigenous S. cerevisiae strains and the aroma profile of the resulting wine [11]. On the contrary, Santamaría and co-workers [3] found that there are no representative strains from the winery or the area in the long term (3-4 consecutive years) investigating on the biodiversity of native S. cerevisiae in 11 wineries.
In this work, the occurrence and persistence of S. cerevisiae populations of two wineries situated within the Verdicchio DOC winemaking area (distant c.a. 20 Km) was investigated. Both wineries never used commercial wine strains using spontaneous fermentations of a pre-inoculum (“pie de cuve”). The isolated S. cerevisiae strains, were bio typed and oenologically characterized to evaluate the relation among origin, biotype, and oenological characters.
2. Materials and Methods
2.1. S. cerevisiae Strains Origin
A total of 206 S. cerevisiae strains, belonging to the microbial collection of the Dipartimento di Scienze della Vita e dell’Ambiente (DiSVA), Università Politecnica delle Marche (Italy), were previously isolated from two different wineries, located 20 km away from each other. The first cellar (winery 1) was in the town of Cupramontana (coordinates: 43.45939, 13.12515), the second cellar (winery 2) was located in Montecarotto (coordinates: 43.532023, 13.075781), both in winemaking area of Verdicchio DOC Castelli di Jesi Marche region, Italy. Ninety-nine S. cerevisiae strains coming from winery 1, while 107 strains coming from winery 2. These strains were collected during two-year sampling campaign. The wineries did not use commercial starter strains.
2.2. Genotyping Characterization of S. cerevisiae Strains
All S. cerevisiae strains were subjected to genotyping using primer pairs delta12 (5’-TCAACAATGGAATCCCAAC-3’)/delta21 (5’-CATCTTAACACCGTATATGA-3’) to analyse interdelta sequence, as described by Legras and Karst [12]. The genomic DNA of yeasts was extracted at 95 °C for 10 min using a Biorad Thermal Cycler [13] and used to carried out the amplification as follow: 3 min at 95 °C, then 25 s at 94 °C, 30 s at 45 °C, and 90 s at 72 °C for 9 cycles and 25 s at 94 °C, 30 s at 50 °C, and 90 s at 72 °C for 21 cycles. It was performed a final extension at 72 °C for 10 min. Amplicons obtained were separated by electrophoresis on 1.5 % (w/v) of agarose gel submitted to 66 V for 1.5 h in 0.5 × TBE buffer, to compare between them the electrophoretic profile of yeasts and define the different biotypes. Four commercial starter strains, commonly used in Verdicchio wine production, were used to exclude any commercial contamination: three strains of S. cerevisiae, Lalvin EC1118, Lalvin ICV OKAY® (Lallemand Inc., Toulouse, France) and VIN13 (Anchor Wine Yeast, Cape Town, South Africa), and a strain of Saccharomyces bayanus N96 (Anchor Wine Yeast, Cape Town, South Africa).
2.3. Analyses of Biotypes for H2S Production and Killer Activity
Biotypes were tested for their ability to produce H2S (hydrogen sulphide), an undesirable compound in wine (confers rotten egg aromatic note), following the procedure reported by Agarbati et al. [14]: pure culture of each biotype was spread on BiGGY agar medium (Oxoid Ltd., Cheshire, England) and incubated at 25 °C for 48 h. In this medium, the colonies H2S-negative appear white, while appear brown/black those H2S-positive. Biotypes were tested also for their killer activity, as described by Comitini et al. [15]: a sensitive S. cerevisiae strain, DiSVA 42, were seeded into Malt agar medium (Biolife, Monza, Italia) buffered to pH 4.2 with 0.1 M citric acid/dibasic sodium phosphate, at a final concentration of 106 CFU/mL Pure cultures of biotypes were spotted on Malt agar surface and after 48 h of incubation at 25 °C, strains were designated as killer when around the spot there was a clear zone of growth inhibition of the sensitive yeast.
2.4. Fermentation Trials
Biotypes of both wineries were oenologically characterized carrying out microfermentation using Verdicchio must, whose characteristics as follow: initial sugar content 236 g/L, pH 3.22, total acidity 4.42 g/L, malic acid 2.70 g/L, total SO2 35 mg/L, and yeast assimilable nitrogen (YAN) content 89 mgN/L. The YAN was adjusted to 250 mgN/L by the addition of diammonium phosphate and yeast derivative (Genesis Lift® Oenofrance, Bordeaux, France), then, the must was treated overnight at 4 °C with 0.2 mL/L of dimethyl dicarbonate to suppress the wild yeasts. It was confirmed through viable cell counts using WL nutrient agar (Wallerstain Laboratories, Oxoid, Hampshire, UK). Biotypes were precultured in modified YPD (0.5% w/v yeast extract, 2% w/v glucose, and 0.1% w/v peptone) for 24 h at 25 °C, washed twice with sterile water and used to inoculate (c.a. 1 × 106 cells/mL) flasks containing 70 mL of Verdicchio must. The flasks were locked with Pasteur hydraulic valve, incubated at 22 °C under static conditions and daily monitored by weight loss, due to the CO2 evolution, until the end of the fermentation (constant weight for almost two consecutive days). The trials were carried out in duplicate and S. cerevisiae Lalvin ICV OKAY® and S. bayanus N96, were used as controls.
2.5. Main Analytical Characters and Volatile Compounds of Microfermentation Trials
The final wines were analysed for the main analytical characters. The fermentation rate was calculated as g CO2/day over the 3rd day of fermentation, volatile acidity and ethanol content were determined following the methods reported by the International Organisation of Vine and Wine [16,17]. Acetaldehyde, ethyl acetate, n-propanol, isobutanol, amyl- and isoamyl alcohols were determined by direct injection of the sample in a gas chromatograph (GC-2014, Shimadzu, Kyoto, Japan) following the procedure of Canonico et al. [18]. The volatile compounds were determined using the solid phase microextraction (HS-SPME) approach: 5 mL of sample was placed into a vial with 1 g of NaCl and 1.6 mg/L of 3-octanol as the internal standard. The sample was stirred for 10 min at 25 °C and placed at 40 °C for 30 min inserting the fiber Divinylbenzene/Carboxen/Polydimethylsiloxane (DVB/CAR/PDMS) into the vial headspace. Then, the fibre was inserted into a Shimadzu gas chromatograph GC injector, in split–splitless modes, using a glass capillary column of 0.25 µm Supelcowax 10, length 60 m, internal diameter 0.32 mm, to allow the desorption of the compounds [19].
2.6. Data Analyses
The experimental data regarding fermentation rate, volatile acidity and ethanol were subjected to analysis of variance (ANOVA) through Duncan’s tests, considering an associated p-value < 0.05 (software STATISTICA 7, Statsoft inc., Tulsa, OK, USA). The mean values of by-products of fermentation and volatile compounds were used to carry out Principal Component Analysis (PCA) using JMP 11® statistical software (Statistical discovery from SAS, New York, NY, USA).
3. Results
3.1. Genotyping, Occurrence, H2S Production and Killer Activity of S. cerevisiae Strains
Interdelta sequence analyses of the 206 S. cerevisiae strains revealed a total of 19 different biotypes, as showed in Figure 1. Seven different biotypes had been found in winery 1, and 12 different biotypes in winery 2, highlighting more biodiversity in the lastly winery. Biotypes from the two wineries were different and both biotypes groups were different from the more diffused commercial strains used as reference.
Regarding the occurrence of each biotype within the winery, in both there are two biotypes that represent the dominant population, biotype I and II in the winery 1 and biotype I and IV in winery 2 (Table 1). In winery 1, the two biotypes represented more than 90% of the entire population of S. cerevisiae strains, the other biotypes occurrence was ≤ 2%. Moreover, both dominant biotypes and the biotype IV had been found in both two-year sampling campaign, as “resident yeasts”. In winery 2 it was observed more biotype’s occurrence variability. The dominant biotypes represented c.a. 40% of the S. cerevisiae population, followed by the biotype III (c.a. 17 %) and biotype II (15%). The remaining 28% of the population was grouped in the other biotypes. Similarly to winery 1, in winery 2 was also found a resident biotype belonging to the dominant yeasts, the biotype IV.
The results (Table 1) show low or absent production of this undesired compound (rotten egg aroma) for all biotypes of both wineries. All of them showed H2S production with a score ≤ 2.0. The evaluation of the presence of killer phenomenon showed that a wide diffusion among the biotypes. Indeed, all biotypes of both wineries showed killer activity against the sensitive S. cerevisiae yeast DiSVA 42, with the only exception of the biotype II belonging to winery 2 (Table 1).
3.2. Oenologically characterization
3.2.1. Fermentation rate, volatile acidity and ethanol production of biotypes
Results of main oenological characters of the biotypes of both wineries are reported in Table 2. No significant differences were detected regarding fermentation rate, volatile acidity and ethanol production comparing the wineries between them and with the commercial starters. Nonetheless, greater variability in the fermentation rate was observed, ranging from 0.9 (Okay ®) to 1.4 (winery 1) gCO2/day.
3.2.2. Main by-products of fermentation
Main by-products of fermentation, responsible for the aromatic profiles of the wines, are reported in Table 3. Overall, a variable production of these compounds is observed among the samples. Nevertheless, S. cerevisiae biotypes of winery 1 and winery 2 differ from both commercial strains for the higher production of β-phenyl ethanol and phenylethyl acetate able to confer rose aroma, and ethyl hexanoate responsible for refreshing fruity aroma. On the contrary, they showed lower production of amyl alcohol, isoamyl alcohol (solvent aroma) and diethyl succinate (raspberry and apple aroma) than commercial strains.
Results of main by-products of fermentation were also elaborated through principal component analysis (PCA) to evaluate the clear distinction between wineries and commercial starters (Figure 2) on aroma compounds production. The elaboration explained a total variability of 100% (PCA 1 73.3%; PCA2 26.7%). On the bases of the compounds evaluated the wineries had well separated each other and from the commercial strains. Winery 1 was distributed in the lower right quadrant and mainly characterized by ethyl octanoate and diethyl succinate production. Winery 2 was in the upper right quadrant and mainly characterized by terpens and isoamyl acetate while commercial strains were placed in upper left quadrant.
4. Discussion
The concept of “terroir”, that refers to wines with distinctive features linked to the characteristics of the geographical area of production, is defined by several characteristics such as the cultivar of the fermented grapes, geographical factors (climate, soil geology and pedology) and the agronomic approach used. Biotic factors are also involved, such as grape’s microbial communities. However, the occurrence and effects of regional/winery specific microbiota in defining wine characteristics are more controversial issues [20,21].
Several studies using high throughput sequencing technologies described microbial communities of grapes and related wines and winery, revealing a link between grape must and soil microbial communities, as well as the geography of the territory [22,23,24]. Other studies showed that there are no specific strains representative of a winery or a winemaking area [3,25,26]. In addition, other factors could be involved in the “microbial terroir” concept, as the role of non-Saccharomyces yeasts at strains level. Indeed, two vineyards can be sharing a very similar yeast consortium at the genus or species level but may be very different at strain level since strong strain’s differences may produce very different wines [20]. Based on these observations, the concept of “microbial terroir” emerged, although its meaning seems more complex, involving many factors, related to both the plant and the physical environment, then terroir must be considered by a pluri-disciplinary approach.
Another aspect regards the concept of “winery effect”. Rosini [27] showed that in a new winery after the initially addition of selected S. cerevisiae, it is able to impose itself on the spontaneous fermentation carried out the next year. In another work [28], the selectivity of the winery environment (“winery effect”) was detected between S. cerevisiae population isolated from modern, working winery, and abandoned one (since 1914). S. cerevisiae yeasts population from modern winery showed higher values for characters typically subjected to selective pressure (fermentation power, fermentation rate, SO2 resistance).
In the present work it was evaluated S. cerevisiae population of two wineries located in the same winemaking area (Verdicchio DOC Castelli di Jesi) and with the same methods of conducting fermentation (never used commercial strains, use of spontaneous fermentations with pre-inoculum). It was found a distinct and stable S. cerevisiae yeast population in both wineries. Some specific genotypes were dominant in each winery and in different years, confirming the results reported by other authors [6,10,29]. These results support the so called “winery effect” or a microbial terroir at a smaller scale was also suggested by other works [6,29,30,31] where some predominant S. cerevisiae strains persisted in different fermentations in the same winery from one year to another and they seemed to be representative of a single winery rather than of an oenological area. Moreover, in agreement with Stefanini et al [32], the results of the PCA analysis of oenological characters of S. cerevisiae population showed distinctive aromatic profiles in each winery population suggesting the existence of a very winery-specific “microbial-terroir” contributing significantly to the final product. To have a more comprehensive picture of the influence of specific yeast populations at the winery or at regional level, further investigations on the microbiota at strain level will be necessary. In this regard, the oenological involvement of non-Saccharomyces could also be useful for clarify the picture of specific biotic component on the specific wine and ultimately on “terroir”.
Non-Saccharomyces starters are increasingly used in combination with conventional starters. This consideration leads to a growing interest in the social life of wine yeasts, to better understand the different species of wine yeasts and their ecological context [33,34]. In the short term, the development of multispecies starter cultures and the study of interactions between wine microorganisms are also likely to become key drivers of wine innovation.
Author Contributions
Investigation, data curation and writing—original draft preparation, A.A.; validation and writing—review and editing, F.C.; conceptualization, methodology, writing—review and editing, validation and supervision, M.C.; validation and writing—review and editing, L.C. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Longo, E.; Cansado, J.; Agrelo, D.; Villa, T.G. Effect of Climatic Conditions on Yeast Diversity in Grape Musts from Northwest Spain. Am J Enol Vitic. 1991, 42, 141–144. [Google Scholar] [CrossRef]
- Capece, A.; Pietrafesa, R.; Siesto, G.; Romaniello, R.; Condelli, N.; Romano, P. Selected Indigenous Saccharomyces Cerevisiae Strains as Profitable Strategy to Preserve Typical Traits of Primitivo Wine. Fermentation 2019, 5, 87. [Google Scholar] [CrossRef]
- Santamaría, P.; López, R.; del Patrocinio Garijo, M.; Escribano, R.; González-Arenzana, L.; López-Alfaro, I., Rosa Gutiérrez, A. Biodiversity of Saccharomyces cerevisiae yeasts in spontaneous alcoholic fermentations: typical cellar or zone strains?. In Morata, A.; Loira, I. Advances in Grape and Wine Biotechnology; 2019 1-15.
- Capece, A.; Romaniello, R.; Siesto, G.; Romano, P. Diversity of Saccharomyces Cerevisiae Yeasts Associated to Spontaneously Fermenting Grapes from an Italian “Heroic Vine-Growing Area”. Food Microbiology 2012, 31, 159–166. [Google Scholar] [CrossRef]
- Pérez-Torrado, R.; Rantsiou, K.; Perrone, B.; Navarro-Tapia, E.; Querol, A.; Cocolin, L. Ecological Interactions among Saccharomyces cerevisiae Strains: Insight into the Dominance Phenomenon. Sci Rep 2017, 7, 43603. [Google Scholar] [CrossRef]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The Biodiversity of Saccharomyces cerevisiae in Spontaneous Wine Fermentation: The Occurrence and Persistence of Winery-Strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef]
- Agarbati, A.; Canonico, L.; Comitini, F.; Ciani, M. Ecological Distribution and Oenological Characterization of Native Saccharomyces cerevisiae in an Organic Winery. Fermentation 2022, 8, 224. [Google Scholar] [CrossRef]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in Alcoholic Fermentation Processes: Role of Physiological Fitness and Microbial Interactions. Appl Microbiol Biotechnol 2016, 100, 2035–2046. [Google Scholar] [CrossRef]
- Ganucci, D.; Guerrini, S.; Mangani, S.; Vincenzini, M.; Granchi, L. Quantifying the Effects of Ethanol and Temperature on the Fitness Advantage of Predominant Saccharomyces cerevisiae Strains Occurring in Spontaneous Wine Fermentations. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Börlin, M.; Venet, P.; Claisse, O.; Salin, F.; Legras, J.-L.; Masneuf-Pomarede, I. Cellar-Associated Saccharomyces Cerevisiae Population Structure Revealed High-Level Diversity and Perennial Persistence at Sauternes Wine Estates. Applied and Environmental Microbiology 2016, 82, 2909–2918. [Google Scholar] [CrossRef]
- Lappa, I.K.; Kachrimanidou, V.; Pateraki, C.; Koulougliotis, D.; Eriotou, E.; Kopsahelis, N. Indigenous Yeasts: Emerging Trends and Challenges in Winemaking. Current Opinion in Food Science 2020, 32, 133–143. [Google Scholar] [CrossRef]
- Legras, J.-L.; Karst, F. Optimisation of Interdelta Analysis for Saccharomyces cerevisiae Strain Characterisation. FEMS Microbiology Letters 2003, 221, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Ciani, M.; Comitini, F. The Impact of Fungicide Treatments on Yeast Biota of Verdicchio and Montepulciano Grape Varieties. PLOS ONE 2019, 14, e0217385. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Comitini, F.; Ciani, M. Reduction of Sulfur Compounds through Genetic Improvement of Native Saccharomyces cerevisiae Useful for Organic and Sulfite-Free Wine. Foods 2020, 9, 658. [Google Scholar] [CrossRef]
- Comitini, F.; Agarbati, A.; Canonico, L.; Galli, E.; Ciani, M. Purification and Characterization of WA18, a New Mycocin Produced by Wickerhamomyces anomalus Active in Wine Against Brettanomyces bruxellensis Spoilage Yeasts. Microorganisms 2021, 9, 56. [Google Scholar] [CrossRef] [PubMed]
- OIV-Compendium of International Methods of Wine and Must Analysis MA-AS313-02. Available online: https://www.oiv.int/standards/annex-a-methods-of-analysis-of-wines-and-musts/section-3-chemical-analysis/section-3-1-organic-compounds/section-3-1-3-acids/volatile-acidity-%28type-i%29 (accessed on 15 May 2024).
- OIV-MA-AS313-24. Available online: https://www.oiv.int/standards/annex-a-methods-of-analysis-of-wines-and-musts/section-3-chemical-analysis/section-3-1-organic-compounds/section-3-1-3-acids/determination-of-total-ethanol-in-wine-by-high-performance-liquid-chromatography-%28type-iv%29 (accessed on 15 May 2024).
- Canonico, L.; Comitini, F.; Ciani, M. Influence of Vintage and Selected Starter on Torulaspora delbrueckii/Saccharomyces cerevisiae Sequential Fermentation. Eur Food Res Technol 2015, 241, 827–833. [Google Scholar] [CrossRef]
- Canonico, L.; Ciani, E.; Galli, E.; Comitini, F.; Ciani, M. Evolution of Aromatic Profile of Torulaspora delbrueckii Mixed Fermentation at Microbrewery Plant. Fermentation 2020, 6, 7. [Google Scholar] [CrossRef]
- Alexandre, H. Wine Yeast Terroir: Separating the Wheat from the Chaff—for an Open Debate. Microorganisms 2020, 8, 787. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, I.S. Tasting the Terroir of Wine Yeast Innovation. FEMS yeast research 2020, 20, foz084. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Thorngate, J.H.; Richardson, P.M.; Mills, D.A. Microbial Biogeography of Wine Grapes Is Conditioned by Cultivar, Vintage, and Climate. Proceedings of the National Academy of Sciences 2014, 111, E139–E148. [Google Scholar] [CrossRef]
- Knight, S.; Klaere, S.; Fedrizzi, B.; Goddard, M.R. Regional Microbial Signatures Positively Correlate with Differential Wine Phenotypes: Evidence for a Microbial Aspect to Terroir. Sci Rep 2015, 5, 14233. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Collins, T.S.; Masarweh, C.; Allen, G.; Heymann, H.; Ebeler, S.E.; Mills, D.A. Associations among Wine Grape Microbiome, Metabolome, and Fermentation Behavior Suggest Microbial Contribution to Regional Wine Characteristics. mBio 2016, 7. [Google Scholar] [CrossRef]
- Vigentini, I.; De Lorenzis, G.; Fabrizio, V.; Valdetara, F.; Faccincani, M.; Panont, C.A.; Picozzi, C.; Imazio, S.; Failla, O.; Foschino, R. The Vintage Effect Overcomes the Terroir Effect: A Three Year Survey on the Wine Yeast Biodiversity in Franciacorta and Oltrepò Pavese, Two Northern Italian Vine-Growing Areas. Microbiology 2015, 161, 362–373. [Google Scholar] [CrossRef]
- Torija, M.J.; Rozès, N.; Poblet, M.; Guillamón, J.M.; Mas, A. Yeast Population Dynamics in Spontaneous Fermentations: Comparison between Two Different Wine-Producing Areas over a Period of Three Years. Antonie Van Leeuwenhoek 2001, 79, 345–352. [Google Scholar] [CrossRef]
- Rosini, G. Assessment of Dominance of Added Yeast in Wine Fermentation and Origin of Saccharomyces cerevisiae in Wine-Making. The Journal of General and Applied Microbiology 1984, 30, 249–256. [Google Scholar] [CrossRef]
- Cocolin, L.; Pepe, V.; Comitini, F.; Comi, G.; Ciani, M. Enological and Genetic Traits of Saccharomyces Cerevisiae Isolated from Former and Modern Wineries. FEMS Yeast Research 2004, 5, 237–245. [Google Scholar] [CrossRef]
- Abdo, H.; Catacchio, C.R.; Ventura, M.; D’Addabbo, P.; Calabrese, F.M.; Laurent, J.; David-Vaizant, V.; Alexandre, H.; Guilloux-Bénatier, M.; Rousseaux, S. Colonization of Wild Saccharomyces Cerevisiae Strains in a New Winery. Beverages 2020, 6, 9. [Google Scholar] [CrossRef]
- Vezinhet, F.; Hallet, J.-N.; Valade, M.; Poulard, A. Ecological Survey of Wine Yeast Strains by Molecular Methods of Identification. Am J Enol Vitic. 1992, 43, 83–86. [Google Scholar] [CrossRef]
- Gutiérrez, A.R.; Santamaría, P.; Epifanio, S.; Garijo, P.; Ópez, R.L. Ecology of Spontaneous Fermentation in One Winery during 5 Consecutive Years. Letters in Applied Microbiology 1999, 29, 411–415. [Google Scholar] [CrossRef]
- Stefanini, I.; Albanese, D.; Cavazza, A.; Franciosi, E.; De Filippo, C.; Donati, C.; Cavalieri, D. Dynamic Changes in Microbiota and Mycobiota during Spontaneous ‘Vino Santo Trentino’ Fermentation. Microbial Biotechnology 2016, 9, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Jouhten, P.; Ponomarova, O.; Gonzalez, R.; Patil, K.R. Saccharomyces Cerevisiae Metabolism in Ecological Context. FEMS Yeast Research 2016, 16, fow080. [Google Scholar] [CrossRef] [PubMed]
- Conacher, C.; Luyt, N.; Naidoo-Blassoples, R.; Rossouw, D.; Setati, M.; Bauer, F. The Ecology of Wine Fermentation: A Model for the Study of Complex Microbial Ecosystems. Appl Microbiol Biotechnol 2021, 105, 3027–3043. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Different interdelta profiles of biotypes of both wineries. Each biotype is represented as Roman numeral, white colored for winery 1 and orange colored for winery 2. All biotype’s profiles were compared with those of commercial strains OKAY®, N96, EC1118 and VIN13.
Figure 1.
Different interdelta profiles of biotypes of both wineries. Each biotype is represented as Roman numeral, white colored for winery 1 and orange colored for winery 2. All biotype’s profiles were compared with those of commercial strains OKAY®, N96, EC1118 and VIN13.
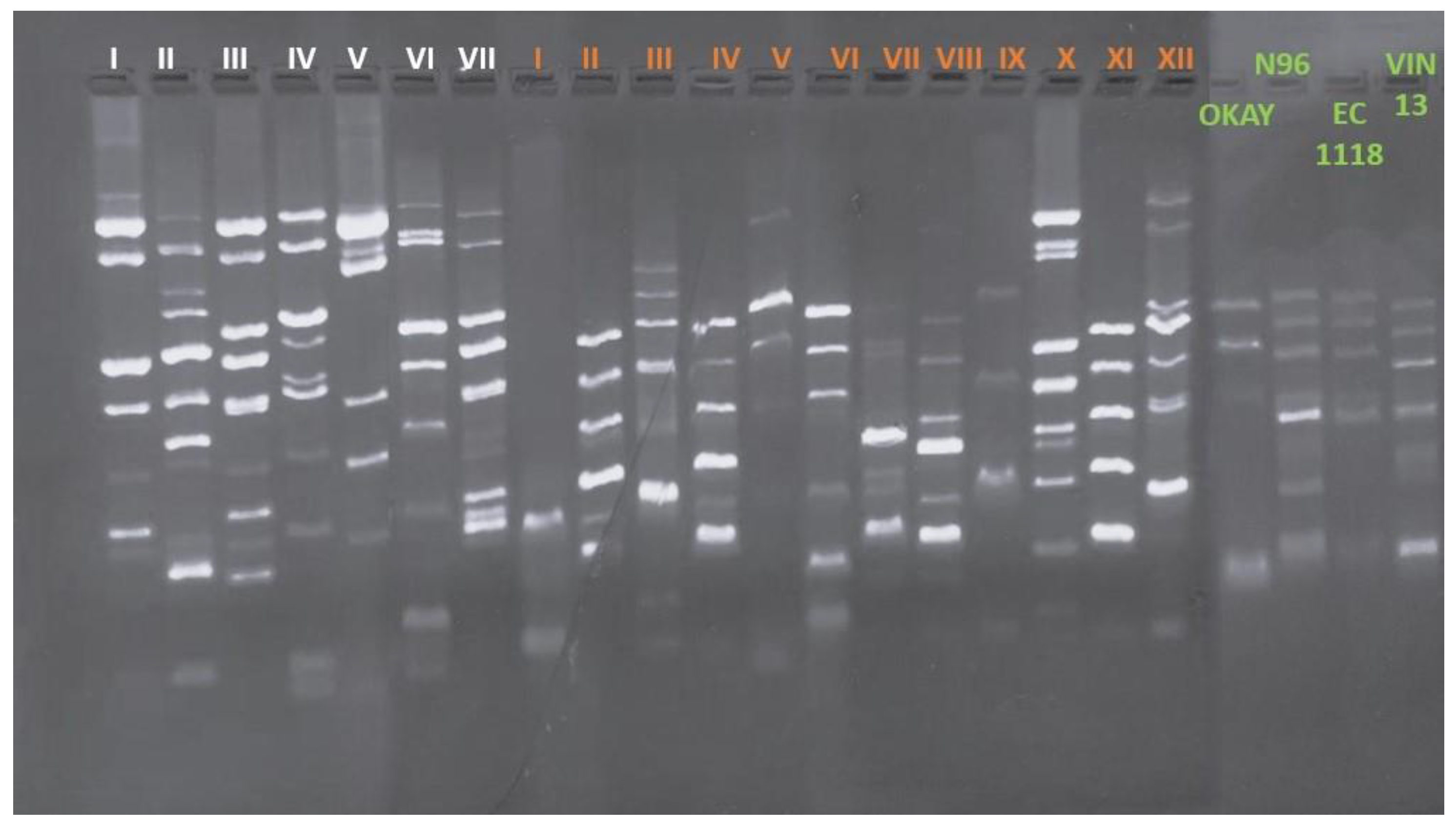
Figure 2.
Principal Component Analysis (PCA) based on the data for the main by-products of fermentation of wines obtained by all biotypes belonging to the two wineries, compared with commercial starter strains.
Figure 2.
Principal Component Analysis (PCA) based on the data for the main by-products of fermentation of wines obtained by all biotypes belonging to the two wineries, compared with commercial starter strains.
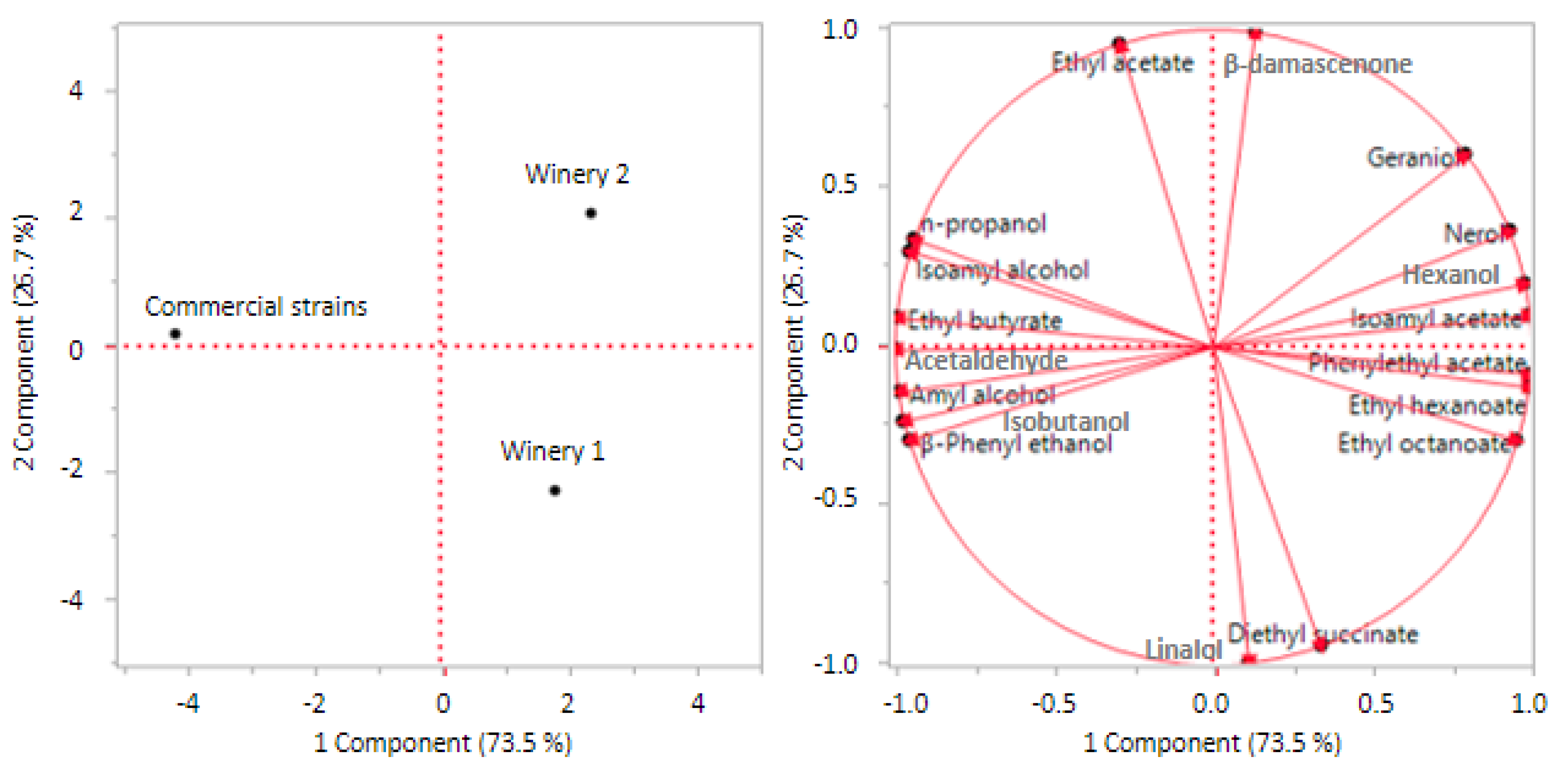

Table 1.
Biotypes occurring in wineries, their frequency and characterization in H2S production and killer activity.
Table 1.
Biotypes occurring in wineries, their frequency and characterization in H2S production and killer activity.
| Winery | Biotype | Frequency (%) | H2S production | Killer activity |
|---|---|---|---|---|
| 1 | I* | 49.5 | 2.0 | +/− |
| II* | 43.4 | 0.0 | + | |
| III | 1.0 | 1.5 | + | |
| IV* | 2.0 | 0.0 | + | |
| V | 1.0 | 0.5 | + | |
| VI | 1.0 | 0.5 | +/− | |
| VII | 2.0 | 0.8 | + | |
| 2 | I | 23.4 | 0.5 | + |
| II | 15.0 | 0.0 | − | |
| III | 16.8 | 0.0 | + | |
| IV* | 19.6 | 1.0 | + | |
| V | 2.8 | 0.5 | +/− | |
| VI | 7.5 | 0.5 | + | |
| VII | 1.9 | 1.5 | +/− | |
| VIII | 1.9 | 0.8 | +/− | |
| IX | 3.7 | 0.0 | +/− | |
| X | 0.9 | 1.0 | + | |
| XI | 5.6 | 1.0 | + | |
| XII | 0.9 | 1.0 | +/− |
*Biotypes found in both sampling years within the same winery.
Table 2.
Fermentation rate of biotypes and volatile acidity and ethanol content of resulted wines. Biotypes of the same winery were analysed as unique population reported as winery 1 and winery 2 and compared with both commercial starter strains, OKAY® and N96. Data are reported as mean value ± standard deviation and Duncan’s test (p-value < 0.05) reported as different superscript letters (a,b,c) within each column represent significant differences.
Table 2.
Fermentation rate of biotypes and volatile acidity and ethanol content of resulted wines. Biotypes of the same winery were analysed as unique population reported as winery 1 and winery 2 and compared with both commercial starter strains, OKAY® and N96. Data are reported as mean value ± standard deviation and Duncan’s test (p-value < 0.05) reported as different superscript letters (a,b,c) within each column represent significant differences.
| Strains population | Fermentation rate (gCO2/day)* | Volatile acidity (g/L) | Ethanol (% v/v) |
|---|---|---|---|
| Winery 1 | 1.4±0.2a | 0.4±0.1a | 12.8±0.3a |
| Winery 2 | 1.2±0.1ab | 0.5±0.1a | 12.4±0.3a |
| OKAY® | 0.9±0.0c | 0.4±0.0a | 12.4±0.0a |
| N96 | 1.1±0.0bc | 0.5±0.0a | 12.4±0.1a |
*Fermentation rate calculated at the 3rd day of fermentation.
Table 3.
Main by-products of fermentation of wines obtained by biotype’s populations of winery 1 and winery 2 compared with those obtained by commercial starter strains OKAY® and N96. Data are reported as mean value ± standard deviation and Duncan’s test (p-value < 0.05) reported as different superscript letters (a,b,c) within each row represent significant differences.
Table 3.
Main by-products of fermentation of wines obtained by biotype’s populations of winery 1 and winery 2 compared with those obtained by commercial starter strains OKAY® and N96. Data are reported as mean value ± standard deviation and Duncan’s test (p-value < 0.05) reported as different superscript letters (a,b,c) within each row represent significant differences.
| Fermentation by-products | Winery 1 | Winery 2 | OKAY® | N96 |
|---|---|---|---|---|
| Alcohols | ||||
| Hexanol (mg/L) | 11.84±0.99a | 12.66±1.77a | 10.38±0.61ab | 9.13±0.74b |
| β-Phenyl ethanol (mg/L) | 7.48±1.12a | 7.39±1.11a | 7.96±0.03b | 7.32±0.00b |
| n-propanol (mg/L) | 12.60±8.26b | 16.88±2.96b | 42.70±0.09a | 13.51±0.42b |
| Amyl alcohol (mg/L) | 7.28±4.44b | 4.85±2.39b | 15.42±0.39a | 15.55±0.38a |
| Isoamyl alcohol (mg/L) | 36.96±10.7b | 41.60±12.07b | 52.01±0.17ab | 60.78±0.37a |
| Isobutanol (mg/L) | 5.22±3.94a | 8.24±1.78a | 5.29±0.03a | 7.89±0.04a |
| Carbonyl compounds | ||||
| Acetaldehyde (mg/L) | 15.59±9.85b | 12.21±2.06b | 15.05±0.47b | 80.97±0.23a |
| Esters | ||||
| Isoamyl acetate (mg/L) | 0.80±0.39ab | 0.90±0.20a | 0.39±0.00b | 0.35±0.01b |
| Phenyl ethyl acetate (mg/L) | 0.47±0.16a | 0.47±0.09a | 0.21±0.01b | 0.38±0.02ab |
| Ethyl hexanoate (mg/L) | 0.65±0.19a | 0.63±0.22a | 0.39±0.00ab | 0.16±0.01b |
| Ethyl butyrate (mg/L) | 0.03±0.03a | 0.03±0.05a | 0.06±0.00a | 0.05±0.01a |
| Ethyl octanoate (µg/L) | 3.75±1.04a | 3.60±1.13a | 3.45±0.34a | 2.78±0.05a |
| Diethyl succinate (mg/L) | 0.04±0.01b | 0.03±0.01b | 0.01±0.00c | 0.06±0.00a |
| Ethyl acetate (mg/L) | 10.16±3.34b | 11.61±4.61ab | 17.62±0.20a | 5.13±0.03b |
| Terpenes | ||||
| Linalol (µg/L) | 17.85±8.96a | 7.12±5.04a | 8.89±0.53a | 12.51±1.07a |
| Nerol (µg/L) | 3.41±1.48ab | 4.56±1.50a | 1.95±0.50b | 1.88±0.39b |
| Geraniol (µg/L) | 4.71±1.58a | 5.69±1.53a | 4.02±0.57a | 4.49±0.47a |
| Enones | ||||
| β-Damascenone (µg/L) | 4.56±2.05a | 4.15±1.41a | 5.89±0.34a | 4.98±0.28a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



