
Submitted:
27 May 2024
Posted:
28 May 2024
You are already at the latest version
Abstract
Macrophages as a source and target of GDF-15
Keywords:
cytokine
; growth factor
; receptor
; inflammation
; healing
; fibrosis
1. Introduction
Macrophages are ubiquitous in almost all human tissues and profoundly influence the healing and remodeling of damaged tissue. At the core of their impressive versatility lies their ability to rapidly polarize in response to stimuli. For example, during healing processes, macrophages remove local debris via phagocytosis and secrete various factors that promote angiogenesis and scar formation through auto- and paracrine mechanisms [1,2]. This dynamic adaptability to their environment has made macrophages essential to the body's natural tissue maintenance mechanisms [3].
During the inflammatory phase of injury, there is an increase in macrophage infiltration, accompanied by an enhanced production of proinflammatory cytokines. This, in turn, induces the recruitment and proliferation of macrophage progenitor cells. A phenotypic transition to M2 favors the resolution of inflammation through the secretion of IL-10, transforming growth factor β (TGF-β), and vascular endothelial growth factor (VEGF), which supports fibroblast proliferation and promotes angiogenesis. The macrophage infiltration then decreases until wound resolution progresses [4,5].
The ontogeny of macrophages may influence their function during healing processes. Tissue-resident macrophages can originate from resident cells that locally proliferate, from a spleen monocyte reservoir, or from blood peripheral circulating bone marrow-derived monocytes (BMDM) [6]. The initial proinflammatory response seems to be supported by spleen-derived monocytes that differentiate into M1. In contrast, the resolution-like M2 phenotype appears to be derived from resident macrophages and circulating monocytes [6].
On the opposite end of the healing spectrum lies fibrosis, a process resulting from excessive extracellular matrix (ECM) accumulation and defective remodeling [7]. Macrophages contribute to fibrogenesis by recruiting and activating fibroblasts, secreting TGF-β1, and modulating the tissue microenvironment. They also contribute to the resolution of fibrosis by secreting matrix metalloproteinases (MMPs) that degrade the ECM, such as MMP2 and MMP13, and by participating in the clearance of senescent cells [8].
The balance between reparative processes and fibrotic responses is regulated by the interplay of local inflammatory cues. For instance, members of the TGF-β superfamily exert dualistic effects, depending on their content, concentration, spatial and temporal production and clearance, as well as the activation status of target cells [9]. Our review focuses on growth differentiation factor 15 (GDF-15), a member of the transforming growth factor-beta (TGF-β) superfamily, which can be both produced and cleared by macrophages. GDF-15 production can be induced by inflammatory mediators, yet the majority of its effects are anti-inflammatory and pro-fibrotic. We have summarized the available data on the role of GDF-15 in pathologies in which monocytes and macrophages play a major role.
2. GDF-15 Is a Member of the TGF-β Superfamily
GDF-15, also known as macrophage inhibitory cytokine-1 (MIC-1), placental transforming growth factor-β (PTGF-β), prostate-derived factor (PDF), and NSAID-activated gene-1 (NAG-1), is a multifunctional cytokine and is a distant member of the glial cell-derived neurotrophic factor (GDNF) family and the TGF-β superfamily [10,11,12]. Numerous TGF-β family members are known to be produced by macrophages and to target macrophages, including TGF-β1, GDFN, and bone morphogenetic protein 2 (BMP-2) [13,14,15]. GDF-15 was discovered in the late 1990s when it was identified as an autocrine cytokine capable of reducing the proinflammatory macrophage activation after lipopolysaccharide (LPS) stimulation in the human myelomonocytic cell line U93737, In parallel, Lawton et al. discovered its expression in the placenta during early and late gestation [16]. Moreover, Paralkar et al. found elevated expression of this protein in the prostate and named it PDF [17]. A few years later, Baek et al., observed the upregulation of GDF-15 in HCT-116 colon cancer cells upon treatment with non-steroidal anti-inflammatory drugs (NSAIDs) [18].
GDF15 gene is found throughout the animal kingdom and is well-conserved in vertebrates, particularly in mammals [19,20]. It is located on chromosome 19p12-13.1 and consists of two exons (309 bp and 891 bp) separated by a 2,9 kb intron [16]. Analogous to other members of the TGF-β family, GDF-15 has a dimeric disulfide-bonded configuration and is synthesized as a proprotein [10]. Its general structure includes a propeptide followed by an RXXR furine-like site and the mature protein. GDF-15 conserves the seven cysteine domains typical of the TGF-β family, sharing a 20% amino acid identity that gives rise to its cysteine knot crystal motif (Figure 1a and c) [12]. In addition to its proximity to the TGF-β family, GDF-15 resembles the structure of the GDNF family ligands, sharing 16% amino acid identity (Figure 1b and c) [10]. GDF-15 immature form consists of 308 amino acids, including a 9-amino acid signal peptide, a 167-amino acid propeptide, and a 112-amino acid mature protein. As part of its post-translational modifications, the immature form undergoes proteolytic cleavage, leaving the mature (13 kD) and propeptide (30 kD) forms, which are later cleaved at the RXXR furine-like site [12]. This site is recognized and cleaved by proprotein convertase subtilisin/kexin type (PCSK)-3, -5, and -6 or by MMPs, facilitating GDF-15 maturation [21,22,23]. After dimerization, the mature form, consisting of 224 amino acids (25 kD), and the propeptide are secreted. In contrast to the general structure of the TGF-β family, the propeptide of GDF-15 lacks cysteine residues [12]. Interestingly, the propeptide appears to function autonomously [24]. Latent stromal stores of this immature form have been found in various tissues and pathologies, such as in prostate cancer. These stores serve as a reservoir for GDF-15 [25]. Moreover, the secretion of the propeptide is faster than that of the mature GDF-15 [26].
2.1 Producing Cells and Regulators of GDF-15 Expression
Under physiological conditions, GDF-15 is produced at high levels by trophoblasts in the terminal villi of the placenta, with levels reaching up to 54,000 pg/mL [28,29]. Other human tissues produce lower levels of GDF-15, including stomach, skeletal muscle, subcutaneous adipose tissue, prostate epithelium, bladder, kidney, and lung [30,31,32,33,34,35,36,37]. Furthermore, GDF-15 has been found in most human fluids, including blood, amniotic fluid, bronchoalveolar fluid, and cerebrospinal fluid [28,34] (Figure 2).
A diverse array of cell types, such as fibroblasts, adipocytes, macrophages, epithelial and endothelial cells contribute to the production of GDF-15 (Table 1). However, most of them express GDF-15 under stress conditions, such as exposure to LPS, bleomycin and oligomycin [38,39,40]. GDF-15 production is specialized in some organs, such as in the stomach, where it is produced by chief cells, located at the bottom of the gastric glands [37]. Among the blood cells, the myeloid population exhibits higher expression levels of GDF-15 compared to the lymphoid population. Following clodronate-induced depletion of myeloid cells in mice, GDF-15 mRNA levels significantly dropped, highlighting a major myeloid contribution to systemic GDF-15 levels [41]. During the hematopoietic differentiation of CD34+ progenitors, erythroid precursors secrete high levels of GDF-15. In contrast, the myeloid cell lineage exhibited minimal levels of GDF-15 during maturation [42]. During erythroblast maturation, GDF-15 levels are found up to 74000 pg/mL [43]. Megakaryocytes were also shown to be key cells expressing GDF-15 in the bone marrow. In primary myelofibrosis, megakaryocytes are the most GDF-15 expressing cells in the bone marrow microenvironment [44].
Several transcriptional factors have been identified that induce GDF-15 expression (Table 2). For example, upon exposure to C-reactive protein (CRP), p53 binds to the GDF-15 promoter and induces GDF-15 transcription in human aortic endothelial cells [53]. This highlights an association between the two biomarkers, GDF-15 and CRP, which are co-elevated in acute inflammatory conditions. Also induces GDF-15 expression following damage induced in enterocytes. In this context, GDF-15 functions as a pro-apoptotic factor and triggers the expression of activating transcription factor 3 (ATF3), a pro-survival protein induced during cellular stress [56]. The interaction between ATF3 and GDF-15 also been reported in human colorectal cancer cells [59].
Transcription factor EB (TFEB), a regulator of energy expenditure and an inducer of autophagy, binds to the GDF-15 promoter and increases GDF-15 expression following exposure to lysosomal stressors in adipose tissue macrophages. This mechanism mediates a reduction in metabolic inflammation during high-fat-induced obesity in mice [41]. In adipocytes, signal transducer and activator of transcription 6 (STAT6) triggers the expression of GDF-15 when exposed to IL-13 treatment. This process seems to enhance glucose tolerance [54]. In addition, under mitochondrial stress, C/EBP homologous protein (CHOP) promotes GDF-15 expression in murine myoblasts and hepatocytes [55]. Another known transcription factor is early growth response 1 (EGR1), which forms a positive feedback loop with GDF-15 and acts as a promoter for EGR1 [57]. Proliferator-activated receptor γ (PPARγ) and retinoid X receptor α (RXRα) are regulatory elements of the GDF15 locus in monocyte-derived macrophages that are active during muscle regeneration [60]. Nuclear factor erythroid 2-related factor 2 (NRF2), a factor involved in the transcription of antioxidant proteins, also promotes GDF-15 expression in BMDMs treated with NSAIDs [61]. Table 2 highlights the diverse range of GDF-15 inducers.
2.2. Known Receptors and Target Cells of GDF-15
In 2017, GDNF family receptor α-like (GFRAL), a member of the GDNF receptor α family, was identified as a receptor for GDF-15. This receptor is highly expressed in the area postrema and nucleus of the solitary tract of the hindbrain in mice, non-human primates, and humans [65,66,67]. Using an unbiased ligand-receptor coupling approach, Mullican et al. and Yang et al. found that GDF-15 binds exclusively to GFRAL, excluding high-affinity binding to other receptors, including those of the TGF-β receptor family [66,67]. Other potential ligands for GFRAL, such as TGF-β and GDNF-similar ligands (GFLs), were also ruled out, highlighting an exclusive partnership between GDF-15 and GFRAL [66]. Together with Emerson et al, these three research groups dissected the mechanism of GDF-15-GFRAL binding and its effect on appetite regulation [67]. Moreover, the receptor tyrosine-protein kinase (RET) was found to be a co-receptor necessary for the metabolic effects of GDF-15 through GFRAL [65,67]. Upon its binding, GDF-15 promotes the physical interaction between GFRAL and RET, and mediates the activation of RET phosphorylation and the intracellular phosphorylation cascade of extracellular-signal-regulated kinase (ERK), protein kinase B (Akt), and phospholipase C [65,66].
Like GFRAL, RET expression is particularly high in the area postrema and nucleus of the solitary tract in the hindbrain of both mice and non-human primates. Beyond this, GFRAL is also expressed in the human spleen, thymus, testis, and adipose tissue, as well as in isolated adipocytes, but not in macrophages [32,40]. RET is expressed in most mouse tissues except liver, kidney, and adrenal glands [66]. GFRAL/RET expression was also shown in osteoblasts in a murine model of prostate cancer bone metastasis [58]. In the immune cells, GFRAL is expressed by regulatory T cells (Treg) treated with recombinant human GDF-15 (rGDF-15) [68]. These findings highlight that GFRAL expression extends beyond the boundaries of the central nervous system, possibly increasing in response to pathological stimuli, and varies among different cell types.
To date, other receptors have been investigated as possible mediators of GDF-15 peripheral actions. For instance, rGDF-15 was shown to increase the phosphorylation of downstream components of the TGF-βI receptor cascade, namely Smad2/3, in fibroblasts and in human colorectal carcinoma cells, which was associated with metastasis induction [40,69]. In human acute monocytic leukemia (THP-1) cells and neutrophils, rGDF-15 binds to the TGF-βI receptor and effectively blocks the cytokine-induced phosphorylation cascade, promoting anti-inflammatory activity [70]. However, concerns have been raised about the validity of these results due to reported TGF-β contamination of rGDF-15 [71]. Other alternative mechanisms could explain the peripheral effects of GDF-15, such as the existence of a soluble GFRAL that would form a complex with GDF-15 and facilitate its recognition and internalization in further tissues [72]. Another consideration is that GDF-15 may bind to other receptors depending on the cell type and underlying pathological condition. Therefore, the involvement of TGF-β receptors or other receptors in the response of cells to GDF-15 is still controversial.
Using His-tag mediated pull-down assays and mass spectrometry, Wang et al. identified CD48 as a receptor for GDF-15 in Jurkat T cells. The interaction between GDF-15 and CD48 leads to inhibition of CD48 cascade pathways, resulting in downregulation of STIP1 Homology And U-Box Containing Protein 1 (STUB1) and subsequent accumulation of Forkhead-Box-Protein P3 (FOXP3), thereby facilitating Treg activation. This binding was found to be exclusive to GDF-15 and not to TGF-β [73].
GDF-15 is also recognized as a common endocytic ligand for both stabilin-1 and 2 [74]. Stabilin-1 (STAB1, FEEL-1, CLEVER-1, KIAA0246) is a multifunctional scavenger receptor that mediates the endocytic and phagocytic internalization of various unwanted self-ligands, thereby contributing to tissue homeostasis [75]. Stabilin-1 expression is observed in sinusoidal endothelial cells within the spleen, liver, and lymphatic vessels. It is also present in resident macrophages, excluding Kupffer cells, and in tumor-associated macrophages (TAMs). Macrophages in inflamed or healing cardiac tissue also show expression of stabilin-1 [76,77,78,79,80,81]. Macrophages express stabilin-1 in response to IL-4 and dexamethasone, thus stabilin-1 is considered a marker of M2 polarization [82]. Stabilin-1 expression in TAMs supports tumor growth in animal models and correlates with poor prognosis for patients with various cancers [79,83,84]. Some ligands known to interact with stabilin-1 extracellular domain include modified low-density lipoproteins (LDL), such as oxidized LDL (oxLDL) and acetylated LDL (acLDL), and advanced glycation end products (AGEs), secreted protein acidic and rich in cysteine (SPARC), placental lactogen and epidermal growth factor (EGF) [81,85,86,87]. As an endocytic receptor, stabilin-1 engages the clathrin-dependent pathway to facilitate the intracellular trafficking of internalized ligands. This process involves sorting within endosomes, leading to either lysosomal degradation or storage in secretory vesicles [76,85,88]. Using GGA adaptors, stabilin-1 can also sort intracellular ligands from the biosynthetic pathways into the lysosomal secretory route in macrophages [87].
We identified GDF-15 as a directly interacting protein of stabilin-1 through yeast two-hybrid screening. This interaction was confirmed by affinity chromatography and endocytosis assays. Furthermore, we observed that impaired clearance of GDF-15 in STAB-1-/-STAB-2-/- mice contributed to severe glomerular fibrosis and mild perisinusoidal hepatic fibrosis [74]. However, the fate of GDF-15 after internalization by these receptors remains unclear.
In vitro studies have investigated the target cells of rGDF-15 primarily by assessing its impact on the transcriptome through RT-PCR. Table 3 summarizes the major effects of rGDF-15. Takenochi et al. showed that rGDF-15 supplementation increases fibroblast activation measured by increased secretion of alpha-smooth muscle actin (α-SMA). This effect was not mediated by the GFRAL/RET activation pathway [40]. Other myeloid cells strongly affected by rGDF-15 treatment are erythrocytes. rGDF-15 supplementation leads to decreased erythroid colony formation and decreased transcription of erythroid differentiation factors [49]. In vitro rGDF-15 supplementation has also been linked to increased angiogenesis and increased VEGFA expression in glioblastoma cells [89].
3. GDF-15 in Health and Pathology
GDF-15 basal plasmatic levels range from 337 to 1060 pg/mL. These levels may increase during certain physiological changes, such as muscle contraction and exercise [32,48], and under the pathological conditions reviewed in the following sections. Circulating GDF-15 increases with age and is unaffected by gender [32,39,90]. Notably, GDF-15 is significantly elevated at birth and during the first 4 months of life, reaching around three times the average adult concentration [91]. Its levels also progressively rise during pregnancy, correlating with gestational week and peaking in the third trimester [28].
Under physiological conditions, GDF-15 expression positively correlates with higher maturation states in the erythroid lineage. Furthermore, supplementing erythroid cells with GDF-15 results in reduced metabolic activity and proliferation [49]. Moon et al. found that rGDF-15 enhances the regulatory effect of Treg cells on activated T cells. Moreover, rs7226, a single nucleotide polymorphism associated with increased production of GDF-15 in humans, correlates with an increased count of lymphocytes and monocytes and decreased concentrations of innate immune cells and granulocytes [68].
In general, GDF-15 is overexpressed in cellular stress states and its function seems to be protective. Several pathological conditions show elevated plasma GDF-15 levels, including metabolic, cardiovascular, hematological diseases and cancer, reaching concentrations up to one hundred times the physiological value (see Table 4) [90].
The most studied role of GDF-15 is its anorexigenic effect. GDF-15 mediates weight loss by reducing energy intake, an effect that is a direct consequence of GDF-15-GFRAL binding [72]. Its circulating levels have been consistently found to be elevated in obesity [102,103]. Pena-Leon et al. demonstrated in a mouse model that plasma levels of GDF-15 decrease during fasting and are restored after refeeding. This effect was associated with GDF-15 production by chief cells of the gastric mucosa. This group also showed that high GDF-15 in obesity is associated with increased post-transcriptional processing of the GDF-15 precursor, pro-GDF-15, which is highly accumulated in the gastric mucosa. Moreover, this post-transcriptional processing seems to be impaired in the fasting state [37].
GDF-15 supplementation improves the metabolic profile in high-fat diet-induced obese mice, showing a significant reduction in body weight, food intake, and glycemia. These effects may be due to taste aversion rather than increased gastric motility [102,103]. This anorexic effect is effectively abolished by blocking with a monoclonal antibody directed against GDF-15 and by GDF-15 or GFRAL knockout (KO) [66,102]. Recent studies have suggested that GDF-15 effect on body weight may be also associated with increased energy expenditure and thermogenesis [104]. Recently, Feng Lu et al. found that GDF-15/GFRAL binding is an important determinant of the efficacy of ketogenic diet-induced weight loss. They found that GDF-15 hepatic production significantly increases after introduction of ketogenic diet in obese pigs and mice, which was an effect of increased PPARγ, a known transcription factor of GDF-15. Additionally, they found that the ketogenic diet-induced weight loss was mainly an effect of decreased energy intake, which corroborates previous studies highlighting the anorexigenic role of GDF-15 [105]. A recent review summarizes the relationship between GDF-15 and body weight [106].
High plasmatic GDF-15 levels are also found in association with impaired glucose tolerance, insulin resistance, diabetes, and diabetes-related complications, but its role in glucose homeostasis is unclear. Other references provide a detailed review of this matter [107,108]. GDF-15 is not only elevated during pregnancy but has also been showed diagnostic relevance in preeclampsia and gestational diabetes mellitus [109,110].
3.1. GDF-15/Macrophage Interaction in Physiological and Pathological Conditions
In murine and human BMDM, as well as in THP-1 and RAW264.7 cells, GDF-15 expression is increased under stimulation of pro- and anti-inflammatory mediators, including IL-4, IL-1β, tumor necrosis factor α (TNF-α), IL-2 and macrophage colony-stimulating factor (M-CSF) [45,51]. At the same time, GDF-15 has been widely associated with M2 differentiation and with the inhibition of M1 polarization [40,51,54]. Pence et al. examined the relationship between human serum GDF-15 levels and different parameters of monocyte immunosenescence. They observed significantly elevated levels of GDF-15 in the elderly population compared to younger individuals. Also, circulating GDF-15 levels displayed a negative correlation with maximal monocyte respiratory capacity [114]. The particular interplay between GDF-15 and macrophages in various pathological contexts will be reviewed in the following sections.
3.1.1. Sepsis and Inflammation
Serum GDF-15 levels are elevated in sepsis, regardless of the pathogen involved, and have been correlated with prognosis, severity, and survival, as well as, in SARS-CoV-2 in adults and children [51,115,116,117,118,119,120]. Assessment of the diagnostic value of GDF-15 highlighted its role as a biomarker of sepsis severity, including septic shock. GDF-15 showed a positive correlation with procalcitonin, IL-6, and IL-10 [51]. GDF-15 KO mice were protected against cecal ligation- and puncture-induced sepsis, showing less severe symptoms, lower markers of inflammation, and lower bacterial load [117]. Several other studies have proposed GDF-15 as a protective factor in sepsis [30,121,122]. High GDF-15 levels during bacterial inflammation stimulate beta-adrenergic sympathetic outflow and hepatic triglyceride production, mediate cardiac protection and improve thermal homeostasis [123]. Supporting this mechanism, Kim et al found increased catecholamine synthesis-related gene expression and increased thermogenesis in mice overexpressing GDF-15 [41]. Recently, Wang et al. proved that GDF-15-treated mice show increased skeletal muscle noradrenaline and oxygen consumption, which is associated with increased maintenance of energy expenditure [124].
In vitro stimulation with LPS induces a dose- and time-dependent increase in both GDF-15 expression and secretion in THP-1 and RAW264.7 macrophages [51]. Similarly, in vivo LPS injection increases plasmatic GDF-15 concentrations in mice, rats, and humans (in humans, LPS was administered at a dose of 1 ng/kg) [30,125]. Pretreatment with rGDF-15 resulted in a dose-dependent decrease in the expression of proinflammatory cytokines such as TNF-α, IL-6, monocyte chemoattractant protein-1 (MCP-1), and IL-10. GDF-15 also appears to improve the phagocytic and bactericidal function of macrophages in the THP-1 and RAW264.7 cell lines [51]. Similar experiments by Govaere et al. showed decreased TNF-α and CCL2 secretion after treatment with rGDF-15 in THP-1 cells challenged with LPS [111]. Possible molecular pathways responsible for the decreased secretion of proinflammatory cytokines under GDF-15 exposure have been considered. Zhang et al. found that rGDF-15 promoted phosphatidylinositol 3-kinase (PI3K)/Akt phosphorylation in macrophages under LPS-induced inflammation. This effect was reduced by treatment with the PI3K/Akt inhibitor [119]. Other research groups proposed the decrease in phosphorylation of JAK1/STAT3 and nuclear translocation of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) after rGDF-15 treatment as cascade pathways [121]. The inhibitory effects of GDF-15 on NF-κB have also been shown by others [63,126]. However, no specific receptor has been proposed to mediate this effect.
3.1.2. Fibrosis
GDF-15 has been linked to fibrotic diseases. Recent mouse model experiments related to fibrosis are summarized in Table 5. Govaere et al. and Chung et al. found that GDF-15 expression in hepatic tissue positively correlated with fibrosis progression in non-alcoholic fatty liver disease and carbon tetrachloride- (CCl4) induced liver injury, respectively [111,127]. In contrast, Li et al. observed decreased GDF-15 expression levels in mouse and human liver fibrotic tissue compared to healthy liver tissue during fibrosis progression [128]. These research groups showed that GDF-15 deficient mice display increased liver fibrosis, which can be mitigated with rGDF-15 treatment. Chung et al. also demonstrated that mitochondrial damage, caused by oligomycin and rotenone, induces GDF-15 expression. In a co-culture model involving hepatocytes and Kupffer cells, they observed that the absence of GDF-15 production by hepatocytes resulted in increased expression of proinflammatory cytokines following LPS stimulation by Kupffer cells. GDF-15 deficiency altered immune infiltration in hepatic tissue, increasing the number of CD4+, CD8+ T cells and neutrophils while leaving the monocyte population unaffected. This effect was reversed through the supplementation with rGDF-15 [127]. These findings highlight the paracrine effects of GDF-15 in fibrosis progression.
GDF-15 deficient mice display an increased macrophage infiltration in hepatic tissue and a phenotype shift from Ly6Clow to Ly6Chi macrophages. Li et al. highlighted an inhibitory effect of GDF-15 on NF-κB signaling by showing an increased activation of the pathway in the GDF-15 KO mice. The effects of GDF-15 on fibrosis progression may result from decreased M1 polarization and reduced proinflammatory cytokine expression in the liver. However, the observed effects were not explained due to the binding of GDF-15 to a peripheral receptor or a macrophage receptor. Additionally, Li et al. tested the systemic effect of parenteral GDF-15 preprogrammed macrophages in mice with CCl4-induced liver fibrosis. The study revealed a decrease in the severity of hepatic fibrosis [128].
Kim et al. proposed a mechanism linking liver fibrosis to increased levels of liver and portal catecholamines along with GDF-15 [130]. This association was explained by the increased ethanol-induced oxidative stress in the mitochondria, with catecholamines facilitating increased levels of CYP2E1, which correlated with increased GDF-15 levels. In hepatic cells deficient in GDF-15, there was a significant decrease in Kupffer cell apoptosis and in the expression of apoptotic genes. This research group identified beta-1 adrenergic receptor 1/2 (ADRB1/2) as the receptor responsible for inducing apoptosis in Kupffer cells when exposed to ethanol-induced damage. ADRB1/2 is a catecholamine receptor, which is stimulated by GDF-15. These findings shed light on the mechanism by which GDF-15 functions as a stress-induced cytokine, promoting apoptosis of inflammatory Kupffer cells. These cells play a key role in the fibrotic changes of alcohol-induced liver injury, thereby mitigating further hepatic damage [130].
GDF-15 also associates with lung fibrosis. GDF-15-deficient neonatal mice, which were briefly exposed to hyperoxia, presented decreased survival rates along with impaired alveolarization and perturbed macrophage activation in lung tissue [129]. In a bleomycin-induced lung fibrosis mouse model, GDF-15 expression and protein levels are increased in lung tissue, bronchoalveolar fluid, and plasma of mice with pulmonary fibrosis. Within the lung tissue, the highest GDF-15 positivity was found in epithelial cells and macrophages [40].
3.1.3. Regenerative Processes
Titanium is a widely used implant material in fields such as orthopedics, cardiology, and dentistry [132]. Our group has shown that macrophages exposed to titanium nanoparticles (TiNPs) increased the expression and secretion of GDF-15 [133]. Siddiqui et al. investigated the role of GDF-15 in prostate cancer bone metastasis. They found that prostate cancer cells highly express and secrete GDF-15, which further induces the expression of osteoclastogenesis-related genes in osteoclasts and the expression of MCP-1/CCL2, which is involved in macrophage recruitment to osteoblasts in mice [58]. This suggests that GDF-15 may activate osteoclastogenesis through a paracrine mechanism and further recruit macrophages in the bone. Furthermore, this group found the presence of GFRAL/RET in osteoblasts and showed that GFRAL silencing decreases the osteoclastogenesis and macrophage recruitment markers expression, induced by GDF-15 [58]. This proposes a novel GDF-15/GFRAL/RET functional interaction. We also found that TiNPs decrease the expression of stabilin-1, the clearance receptor of GDF-15, in macrophages, and decrease their endocytic function [133]. This mechanism could further increase the local levels of GDF-15 surrounding titanium implant microenvironment and contribute to inadequate implant osseointegration and aseptic loosening.
3.1.4. Cancer
GDF-15 has been recognized as a potential diagnostic and prognostic biomarker for several gastrointestinal tumors, including pancreatic, colorectal, esophageal, hepatocellular, and gastric cancers [107]. In addition, other cancers such as glioblastoma, breast, lung, cervical, ovarian, endometrial, lung, prostate, renal, urothelial, thyroid, and melanoma have also shown elevated levels of the cytokine [89,100,108,134].
GDF-15 appears to have a dualistic function in the process of carcinogenesis. It has an inhibitory effect on tumor growth in the early stages and subsequently facilitates progression and metastasis in the advanced stages [135].
Overall, GDF-15 has been reported to exert pleiotropic effects on carcinogenesis, as shown in Figure 3. Concerning primary tumor growth, rGDF-15 has been shown to promote the proliferation of esophageal carcinoma cells [136]. In contrast, in bladder cancer, GDF-15 has been shown to decrease cell proliferation and invasion [35]. Likewise, patients with renal cell carcinoma (RCC) and increased GDF-15 protein levels in tumor tissue show a better outcome. Yang et al. proposed GDF-15 as a possible regulator of ferroptosis in RCC [137]. However, the specific pathway surrounding this hypothesis needs to be investigated. In addition, GDF-15 promotes angiogenesis by activating the hypoxia-inducible factor-1α (HIF-1α)/VEGF signaling pathway in colon, gastric, and breast cancer cells [100,138,139]. This is supported by the observation that rGDF-15 promotes p53 degradation, increased HIF-1α accumulation, and vessel formation in human umbilical vein endothelial cells [140].
GDF-15 also plays a role in the tumor microenvironment. Recently, Chitinase-3-like protein 1 (YKL-40) produced by macrophages was shown to promote GDF-15 expression in tumor cells. The interaction of YKL-40 and GDF-15 leads to tumor invasion and suppression of the immune response of CD8+ T lymphocytes through the induction of PD-L1 in gallbladder tumor cells [141]. This has also been demonstrated in glioblastoma cells [142]. In support of this observation, patients with low plasmatic levels of GDF-15 show better response rates to anti-programmed cell death protein-1 (PD-1)/PD-L1 inhibitors in advanced non-small cell lung cancer and in melanoma [143,144].
Recently, Haake et al. reported that GDF-15 produced by melanoma cells inhibits lymphocyte adhesion to endothelium and migration through inhibition of the lymphocyte function-associated antigen (LFA-1)/intercellular adhesion molecule 1 (ICAM-1) axis in T lymphocytes. This inhibition results in reduced lymphocyte infiltration at the tumor site when GDF-15 tissue levels are elevated. In addition, combined anti-GDF-15 and anti-PD-1 therapy results in increased T lymphocyte infiltration in mouse models of pancreatic cancer [144]. The use of GDF-15 as a marker for patients who would benefit from such therapy may be a valuable option in such cancers, where anti-PD-1/PD-L1 inhibitors are used as first-line therapy in intermediate- and poor-risk metastatic tumors.
Other authors linked GDF-15 to chemotherapy resistance, as in the case of Zheng et al. who showed that TAMs derived from a mouse xenograft model of colorectal cancer secrete high levels of GDF-15 and contribute to the reduced chemosensitivity in colorectal cancer cells by increasing fatty acid oxidation metabolism [145]. A similar mechanism has been suggested by Yu et al. in gastric cancer [146].
Bonaterra et al. showed that the presence of GDF-15 in prostate cancer tissue was associated with macrophage infiltration, and the presence of GDF-15+ macrophages was associated with high-grade malignancy [147]. This was also highlighted by Sadasivan et al. who found a higher risk of biochemical recurrence in patients whose prostate cancer biopsy was enriched for M2 macrophages and characterized by elevated GDF-15 expression [148].
Lv et al. studied the effect of GDF-15 enriched conditioned medium from M1-polarized THP-1 macrophages on SCC25, a tongue squamous cell carcinoma cell line, and showed that phosphorylation of ErbB2 and its signaling proteins ERK and AKT was increased. This effect was reduced by knocking out GDF-15 in SCC25 cells [149]. This activation pattern has been widely implicated in tumor progression in several cancer types, particularly breast cancer [141,150]. These observations again highlight a tumorigenic effect of GDF-15. However, a direct correspondence between the presence of GDF-15 in the tumor microenvironment and squamous cell carcinoma progression has not been established [149]. Ratman et al. highlighted the interaction between NF-κB and GDF-15 in the tumor microenvironment by showing that constitutive activation of NF-κB in pancreatic cancer cells leads to secretion of GDF-15, a known NF-κB inhibitor, and induces a decreased cytotoxic capacity in TAMs [63].
Elevated GDF-15 has been strongly associated with metastasis in prostate, esophageal, hepatocellular, colorectal, pancreatic, gastric, and endometrial cancers [151]. Esophageal, breast, and colon cancer models show that GDF-15 correlates with the loss of E-cadherin and that the inhibition of GDF-15 expression decreases cell migration and invasion ability [70,152,153]. In contrast, in A549 lung cancer cells, overexpression of GDF-15 reduces cell growth and migration and decreases the spread of lung and bone metastases [154]. The exact effects and mechanisms explaining GDF-15 behavior in cancer are still controversial and often paradoxical. Regarding the interplay between GDF-15 and macrophages in metastasis, Ding et al. showed that GDF-15 secreted by macrophages contributes to an invasive phenotype in colon cancer cells, an effect that was reversed by GDF-15 neutralizing antibodies. The proposed mechanism was increased phosphorylation of c-Fos via Erk1/2 activation by GDF-15, which induced the expression of epithelial-mesenchymal transition in colon cancer cells [155].
3.1.5. Metabolic and Cardiovascular Disorders
Jung et al. investigated the effect of reduced mitochondrial oxidative function on insulin resistance in mice. They found that adipose tissue resident macrophages express low levels of GDF-15 and show a shift toward M1 polarization. Further experiments showed that treatment with rGDF-15 decreased the expression of IL-6, nitric oxide synthase 2, and TNF-α, and promoted M2 polarization by increasing the production of Arg1 and resistin-like alpha [45]. Campderrós et al. found that GDF-15 produced by murine brown adipocytes suppressed the expression of TNF-α, CCL2, and IL-6 in M1-polarized RAW264.7. In contrast, M2 gene expression was not affected by GDF-15 [156].
Besides its anorexigenic effect and its effect on glucose metabolism, GDF-15 has also been proposed as a biomarker for increased mortality risk and recurrent myocardial infarction (MI) after acute coronary syndrome. Similarly, GDF-15 serves as a biomarker in heart failure, a common complication in patients with coronary heart disease and in atrial fibrillation [101,157,158,159]. In fact, GDF-15 is positively correlated with cardiovascular mortality and all-cause mortality [160]. Mice deficient in GDF-15 have higher mortality after induced MI. They also display an increased recruitment of polymorphonuclear leukocytes, monocytes, and macrophages in the myocardial tissue as compared to controls. Additionally, this recruitment is reduced upon treatment with rGDF-15, which decreases leukocyte adhesion, arrest, and transmigration on the endothelium [161]. Taken together, GDF-15 elevated levels after MI may exert a protective function by reducing immune cell recruitment and, thereby, MI complications, such as cardiac remodeling and heart failure.
In the context of allogeneic transplantation of cardiac progenitor cells (CPCs) as a therapy for myocardial infarction, downregulation of GDF-15 in CPCs resulted decreased activation of Tregs and M2 macrophages, preventing an adequate engraftment into the injured myocardium [131]. In contrast, GDF-15 secreted by CPCs inhibited NF-κB activation and promoted a shift toward M2 polarization and Tregs activation, ultimately associated with a cardioprotective outcome. In this series of experiments, the effects of GDF-15 as part of the secretome of CPCs were shown to be mediated by Tregs, as CPCs injected in absence of Tregs failed to promote the protective effects [131].
GDF-15 has emerged as a significant player in mechanisms involved in atherosclerosis and macrophage function. For example, treatment of THP-1 with rGDF-15 is associated with lipid accumulation, whereas GDF-15 knockdown resulted in reduced lipid burden. In addition, rGDF-15 increased the levels of autophagy-related proteins, suggesting a possible role for GDF-15 in autophagosome formation in foam cells [162]. Heduschke et al. supported these findings by showing that siGDF-15 decreased the autophagic activity in THP-1, an effect that was reversed by rGDF-15 supplementation in THP-1 cells [163]. The observation that GDF-15 decreases the release of proinflammatory cytokines has been reported to be associated with the decreased expression of TLR4 in macrophages under oxLDL treatment [164]. This suggests a potential immunomodulatory role of GDF-15 in the progression of atherosclerosis by reducing the proinflammatory plaque surroundings.
4. Conclusions
The reviewed studies emphasize the significant interplay of GDF-15 in the regulation of immune responses and paracrine effects involving the macrophage system. Although the interplay between GDF-15 and the macrophage system has been identified since the discovery of GDF-15, there is still a knowledge gap regarding the extent to which this interaction impacts human pathology as well as in the understanding of the dynamic mechanism of GDF-15 production and responses of the target cells. In order to understand the sequence of events in GDF-15 controlled process, further experimentation is needed and mechanisms of target cell responses to GDF-15 have to be deciphered. One notable gap is the lack of identification of macrophage receptors for GDF-15 that can clarify its immunomodulatory effects.
5. Future Directions
Due to its crucial roles in multiple biological processes, GDF-15 has been identified as a potential therapeutic target for various diseases. Currently, there is an ongoing recruitment for Phase I/II clinical trials investigating the impact of neutralizing GDF-15. Visugromab is an anti-GDF-15 monoclonal antibody currently being evaluated in a Phase II clinical trial as a combination therapy with the checkpoint inhibitor anti-PD-1/PD-L1 for treating advanced solid tumors (NCT04725474). AV-380 and Ponsegromab, also anti-GDF-15, are under evaluation as potential therapies for cancer-induced cachexia in non-small cell lung, pancreatic, and colorectal cancer patients, as well as, in metastatic colorectal cancer through Phase I and II studies (NCT05865535)(NCT05546476). Additionally, a current application for monoclonal anti-GDF-15 is in the treatment of heart failure, with Ponsegromab undergoing recruitment in a Phase II study (NCT05492500). Finally, a multicenter Phase II study on Visugromab is recruiting participants to assess the effects of combination therapy with Nivolumab on muscle-invasive bladder carcinoma (NCT06059547). The Phase I/IIa clinical trial of AZD8853 in patients with metastatic solid tumors was terminated prematurely due to an overall evaluation of its risk-benefit profile (NCT05397171).
In overall, targeting GDF-15 is a very appealing therapeutic field for those pathologies where GDF-15 activity has demonstrated a clear impact, such as fibrosis-associated diseases. Conversely, animal studies on GDF-15 modulation in sepsis and cancer display significant discrepancies. Consequently, a comprehensive investigation of tissue-specific and context-dependent effects is necessary to clarify these inconsistencies.
Further research should focus on deciphering the complex molecular mechanisms governing the actions of GDF-15, exploring its potential as a therapeutic target, and elucidating the contextual factors that impact its expression in diverse conditions. In addition, exploring the role of cell-specific receptors in mediating the effects of GDF-15 presents an exciting opportunity for further investigation. A thorough comprehension of the role of GDF-15 in fibrosis may facilitate the development of innovative therapeutic interventions and improve the clinical management of fibrotic diseases, as well as, prevent the possible adverse effects of the GDF-15-based therapies.
As reviewed here, the new findings on GDF-15 in the macrophage system reveal a significant potential for displaying immunomodulatory properties in contexts involving remodeling, such as MI and fibrosis. For example, macrophages expressing GDF-15 have been suggested as a novel macrophage type with distinct transcriptomics, which mediates remodeling in sterile muscle injury, specifically by enhancing myoblast proliferation and decreasing inflammatory infiltration [60]. Figure 4 summarizes the reported cellular effects of GDF-15 in the macrophage system as well as its systemic interactions.
Author Contributions
L.SB writing—original draft preparation, J.K. and H.K.; writing—review and editing.
Funding
This research was funded by DRK-Blutspendedienst Baden-Württemberg - Hessen gGmbH.
Conflicts of Interest
The authors declare no conflict of interest
Abbreviation list
| α-SMA | alpha Smooth Muscle Actin |
| acLDL | acetylated LDL |
| ADRB1/2 | beta-1 adrenergic receptor 1/2 |
| AGEs | advanced glycation end products |
| Akt | protein kinase B |
| Arg-1 | Arginase 1 |
| ATF | activating transcription factor |
| bp | base pairs |
| BM | bone marrow |
| BMDM | blood peripheral circulating bone marrow-derived monocytes |
| BMP | bone morphogenetic protein |
| CCL | chemokine (C-C motif) ligand |
| CCl4 | carbon tetrachloride |
| CD | cluster of differentiation |
| CHOP | C/EBP homologous protein |
| CFSE | carboxyfluorescein succinimidyl ester |
| CHO | chinese hamster ovary |
| CPC | cardiac progenitor cells |
| CRIF1 | cytokine response 6 (CR6)-interacting factor 1 |
| CRP | c reactive protein |
| DDC | 3,5-Diethoxycarbonyl-1,4-Dihydrocollidine |
| ECM | extracellular matrix |
| EGF | epidermal growth factor |
| EGR1 | early growth response 1 |
| ELISA | enzyme-linked immunosorbent assay |
| ERK | extracellular-signal-regulated kinase |
| FOXP3 | Forkhead-Box-Protein P3 |
| GDF | growth differentiation factor |
| GDNF | glial cell-derived neurotrophic factor |
| GFRAL | GDNF family receptor α-like |
| HBMVEC | human brain microvascular endothelial cells |
| HIF | hypoxia-inducible factor |
| ICAM-1 | intercellular adhesion molecule 1 |
| IL | interleukin |
| i.m. | intramuscular |
| i.v | intravenous |
| KLF | kruppel-like factor |
| LDL | low-density lipoprotein |
| LFA-1 | lymphocyte function-associated antigen |
| LPS | lipopolysaccharide |
| M-CSF | macrophage colony-stimulating factor |
| MCP-1 | monocyte chemoattractant protein-1 |
| MI | myocardial infarction |
| MIC-1 | macrophage inhibitory cytokine-1 |
| MMP | matrix metalloproteinase |
| mRNA | messenger RNA |
| NAG-1 | NSAID-activated gene-1 |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| NOS2 | nitric oxide synthase 2 |
| NRF2 | nuclear factor erythroid 2-related factor 2 |
| NSAIDs | non-steroidal anti-inflammatory drugs |
| oxLDL | oxidized LDL |
| PBMC | peripheral blood mononuclear cell |
| PD-1 | programmed cell death protein-1 |
| prostate-derived factor | |
| PI3K | phosphatidylinositol 3-kinase |
| PPAR | proliferator-activated receptor |
| PCSK | proprotein convertase subtilisin/kexin |
| PTGF-β | placental transforming growth factor-β |
| RCC | renal cell carcinoma |
| RET | receptor tyrosine-protein kinase |
| RNA | ribonucleic acid |
| RT-PCR | reverse transcription real-time polymerase chain reaction |
| RXRα | retinoid X receptor α |
| SAT | subcutaneous adipose tissue |
| SPARC | secreted protein acidic and rich in cysteine |
| STAT | signal transducer and activator of transcription |
| STUB1 | STIP1 Homology And U-Box Containing Protein 1 |
| TAMs | tumor-associated macrophages |
| TGF | transforming growth factor |
| TFEB | transcription factor EB |
| THP-1 | human acute monocytic leukemia cell line |
| TiNPs | titanium nanoparticles |
| TNF | tumor necrosis factor |
| VEGF | vascular endothelial growth factor |
| YAP | yes-associated protein |
| YKL-40 | chitinase-3-like protein 1 (CHI3L1) |
References
- Xue, J.; et al. Transcriptome-based network analysis reveals a spectrum model of human macrophage activation. Immunity 2014, 40, 274–88. [Google Scholar] [CrossRef] [PubMed]
- Shapouri-Moghaddam, A.; et al. Macrophage plasticity, polarization, and function in health and disease. J Cell Physiol 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Mosser, D.M., K. Hamidzadeh, and R. Goncalves, Macrophages and the maintenance of homeostasis. Cell Mol Immunol 2021, 18, 579–587. [CrossRef]
- Bryan, B.A.; et al. Coordinated vascular endothelial growth factor expression and signaling during skeletal myogenic differentiation. Mol Biol Cell 2008, 19, 994–1006. [Google Scholar] [CrossRef]
- Oishi, Y. and I. Manabe, Macrophages in inflammation, repair and regeneration. Int Immunol 2018, 30, 511–528. [CrossRef]
- Milich, L.M., C.B. Ryan, and J.K. Lee, The origin, fate, and contribution of macrophages to spinal cord injury pathology. Acta Neuropathol 2019, 137, 785–797. [CrossRef] [PubMed]
- de Castro Bras, L.E. and N.G. Frangogiannis, Extracellular matrix-derived peptides in tissue remodeling and fibrosis. Matrix Biol 2020, 91-92, 176–187. [CrossRef]
- Wynn, T.A., A. Chawla, and J.W. Pollard, Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–55. [CrossRef]
- Tian, M. and W.P. Schiemann, The TGF-beta paradox in human cancer: an update. Future Oncol 2009, 5, 259–71. [CrossRef] [PubMed]
- Bootcov, M.R.; et al. MIC-1, a novel macrophage inhibitory cytokine, is a divergent member of the TGF-beta superfamily. Proc Natl Acad Sci U S A 1997, 94, 11514–9. [Google Scholar] [CrossRef]
- Hsu, J.Y.; et al. Non-homeostatic body weight regulation through a brainstem-restricted receptor for GDF15. Nature 2017, 550, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Strelau, J.; et al. GDF-15/MIC-1 a novel member of the TGF-beta superfamily. J Neural Transm Suppl 2000, 273–276. [Google Scholar]
- Wynn, T.A. and L. Barron, Macrophages: master regulators of inflammation and fibrosis. Semin Liver Dis 2010, 30, 245–57. [CrossRef] [PubMed]
- Hashimoto, M.; et al. Stimulation of production of glial cell line-derived neurotrophic factor and nitric oxide by lipopolysaccharide with different dose-responsiveness in cultured rat macrophages. Biomed Res 2005, 26, 223–9. [Google Scholar] [CrossRef] [PubMed]
- Dube, P.R., L. Birnbaumer, and G. Vazquez, Evidence for constitutive bone morphogenetic protein-2 secretion by M1 macrophages: Constitutive auto/paracrine osteogenic signaling by BMP-2 in M1 macrophages. Biochem Biophys Res Commun 2017, 491, 154–158. [CrossRef] [PubMed]
- Lawton, L.N.; et al. Identification of a novel member of the TGF-beta superfamily highly expressed in human placenta. Gene 1997, 203, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Paralkar, V.M.; et al. Cloning and characterization of a novel member of the transforming growth factor-beta/bone morphogenetic protein family. J Biol Chem 1998, 273, 13760–7. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.J.; et al. Cyclooxygenase inhibitors regulate the expression of a TGF-beta superfamily member that has proapoptotic and antitumorigenic activities. Mol Pharmacol 2001, 59, 901–8. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, P.; et al. Conserved function of zebrafish (Danio rerio) Gdf15 as a sepsis tolerance mediator. Dev Comp Immunol 2020, 109, 103698. [Google Scholar] [CrossRef]
- Schreiber, F.; et al. TreeFam v9: a new website, more species and orthology-on-the-fly. Nucleic Acids Res 2014, 42, D922–D925. [Google Scholar] [CrossRef]
- Li, J.J.; et al. Growth Differentiation Factor 15 Maturation Requires Proteolytic Cleavage by PCSK3, -5, and -6. Mol Cell Biol, 2018; 38. [Google Scholar] [CrossRef]
- Li, S.; et al. Maturation of growth differentiation factor 15 in human placental trophoblast cells depends on the interaction with Matrix Metalloproteinase-26. J Clin Endocrinol Metab 2014, 99, E2277–E2287. [Google Scholar] [CrossRef]
- Wischhusen, J., I. Melero, and W.H. Fridman, Growth/Differentiation Factor-15 (GDF-15): From Biomarker to Novel Targetable Immune Checkpoint. Front Immunol 2020, 11, 951. [CrossRef]
- Fairlie, W.D.; et al. The propeptide of the transforming growth factor-beta superfamily member, macrophage inhibitory cytokine-1 (MIC-1), is a multifunctional domain that can facilitate protein folding and secretion. J Biol Chem 2001, 276, 16911–8. [Google Scholar] [CrossRef] [PubMed]
- Bauskin, A.R.; et al. The propeptide mediates formation of stromal stores of PROMIC-1: role in determining prostate cancer outcome. Cancer Res 2005, 65, 2330–6. [Google Scholar] [CrossRef] [PubMed]
- Bauskin, A.R.; et al. The TGF-beta superfamily cytokine MIC-1/GDF15: secretory mechanisms facilitate creation of latent stromal stores. J Interferon Cytokine Res 2010, 30, 389–97. [Google Scholar] [CrossRef]
- Blum, M.; et al. The InterPro protein families and domains database: 20 years on. Nucleic Acids Res 2021, 49, D344–D354. [Google Scholar] [CrossRef] [PubMed]
- Moore, A.G.; et al. The transforming growth factor-ss superfamily cytokine macrophage inhibitory cytokine-1 is present in high concentrations in the serum of pregnant women. J Clin Endocrinol Metab 2000, 85, 4781–8. [Google Scholar] [CrossRef]
- Zeng, Y.T.; et al. GDF15 deficiency hinders human trophoblast invasion to mediate pregnancy loss through downregulating Smad1/5 phosphorylation. iScience 2023, 26, 107902. [Google Scholar] [CrossRef]
- Abulizi, P.; et al. Growth Differentiation Factor-15 Deficiency Augments Inflammatory Response and Exacerbates Septic Heart and Renal Injury Induced by Lipopolysaccharide. Sci Rep 2017, 7, 1037. [Google Scholar] [CrossRef]
- Cheung, P.K.; et al. Protein profiling of microdissected prostate tissue links growth differentiation factor 15 to prostate carcinogenesis. Cancer Res 2004, 64, 5929–33. [Google Scholar] [CrossRef]
- Laurens, C.; et al. Growth and differentiation factor 15 is secreted by skeletal muscle during exercise and promotes lipolysis in humans. JCI Insight 2020, 5. [Google Scholar] [CrossRef] [PubMed]
- Perez-Gomez, M.V.; et al. Urinary Growth Differentiation Factor-15 (GDF15) levels as a biomarker of adverse outcomes and biopsy findings in chronic kidney disease. J Nephrol 2021, 34, 1819–1832. [Google Scholar] [CrossRef] [PubMed]
- Shnaper, S.; et al. Elevated levels of MIC-1/GDF15 in the cerebrospinal fluid of patients are associated with glioblastoma and worse outcome. Int J Cancer 2009, 125, 2624–30. [Google Scholar] [CrossRef] [PubMed]
- Tsui, K.H.; et al. Growth differentiation factor-15: a p53- and demethylation-upregulating gene represses cell proliferation, invasion, and tumorigenesis in bladder carcinoma cells. Sci Rep 2015, 5, 12870. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q., D. Jiang, and H.W. Chu, Cigarette smoke induces growth differentiation factor 15 production in human lung epithelial cells: implication in mucin over-expression. Innate Immun 2012, 18, 617–26. [CrossRef] [PubMed]
- Pena-Leon, V.; et al. Gastric GDF15 levels are regulated by age, sex, and nutritional status in rodents and humans. J Endocrinol Invest 2023. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; et al. Integrated stress response regulates GDF15 secretion from adipocytes, preferentially suppresses appetite for a high-fat diet and improves obesity. iScience 2021, 24, 103448. [Google Scholar] [CrossRef]
- Sramkova, V.; et al. Expression of lipogenic markers is decreased in subcutaneous adipose tissue and adipocytes of older women and is negatively linked to GDF15 expression. J Physiol Biochem 2019, 75, 253–262. [Google Scholar] [CrossRef]
- Takenouchi, Y.; et al. Growth differentiation factor 15 facilitates lung fibrosis by activating macrophages and fibroblasts. Exp Cell Res 2020, 391, 112010. [Google Scholar] [CrossRef]
- Kim, J.; et al. TFEB-GDF15 axis protects against obesity and insulin resistance as a lysosomal stress response. Nat Metab 2021, 3, 410–427. [Google Scholar] [CrossRef]
- Ramirez, J.M.; et al. Growth differentiation factor 15 production is necessary for normal erythroid differentiation and is increased in refractory anaemia with ring-sideroblasts. Br J Haematol 2009, 144, 251–62. [Google Scholar] [CrossRef] [PubMed]
- Tanno, T.; et al. High levels of GDF15 in thalassemia suppress expression of the iron regulatory protein hepcidin. Nat Med 2007, 13, 1096–101. [Google Scholar] [CrossRef] [PubMed]
- Uchiyama, T.; et al. The role of growth differentiation factor 15 in the pathogenesis of primary myelofibrosis. Cancer Med 2015, 4, 1558–72. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.B.; et al. Reduced oxidative capacity in macrophages results in systemic insulin resistance. Nat Commun 2018, 9, 1551. [Google Scholar] [CrossRef] [PubMed]
- Torres-Castro, I.; et al. Human monocytes and macrophages undergo M1-type inflammatory polarization in response to high levels of glucose. Immunol Lett 2016, 176, 81–9. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, S.; et al. Growth differentiation factor 15, a marker of lung involvement in systemic sclerosis, is involved in fibrosis development but is not indispensable for fibrosis development. Arthritis Rheumatol 2014, 66, 418–27. [Google Scholar] [CrossRef]
- Frank, D.; et al. Gene expression pattern in biomechanically stretched cardiomyocytes: evidence for a stretch-specific gene program. Hypertension 2008, 51, 309–18. [Google Scholar] [CrossRef] [PubMed]
- Ranjbaran, R.; et al. GDF-15 negatively regulates excess erythropoiesis and its overexpression is involved in erythroid hyperplasia. Exp Cell Res 2020, 397, 112346. [Google Scholar] [CrossRef] [PubMed]
- Day, E.A.; et al. Metformin-induced increases in GDF15 are important for suppressing appetite and promoting weight loss. Nat Metab 2019, 1, 1202–1208. [Google Scholar] [CrossRef]
- Li, H.; et al. The Clinical Value of GDF15 and Its Prospective Mechanism in Sepsis. Front Immunol 2021, 12, 710977. [Google Scholar] [CrossRef]
- Ha, G.; et al. GDF15 secreted by senescent endothelial cells improves vascular progenitor cell functions. PLoS One 2019, 14, e0216602. [Google Scholar] [CrossRef]
- Kim, Y., N. Noren Hooten, and M.K. Evans, CRP Stimulates GDF15 Expression in Endothelial Cells through p53. Mediators Inflamm 2018, 2018, 8278039. [CrossRef]
- Lee, S.E.; et al. Growth Differentiation Factor 15 Mediates Systemic Glucose Regulatory Action of T-Helper Type 2 Cytokines. Diabetes 2017, 66, 2774–2788. [Google Scholar] [CrossRef]
- Chung, H.K.; et al. Growth differentiation factor 15 is a myomitokine governing systemic energy homeostasis. J Cell Biol 2017, 216, 149–165. [Google Scholar] [CrossRef]
- Zhao, C.; et al. C5a induces A549 cell proliferation of non-small cell lung cancer via GDF15 gene activation mediated by GCN5-dependent KLF5 acetylation. Oncogene 2018, 37, 4821–4837. [Google Scholar] [CrossRef]
- Jin, Y.; et al. Transcriptional Regulation of GDF15 by EGR1 Promotes Head and Neck Cancer Progression through a Positive Feedback Loop. Int J Mol Sci 2021, 22. [Google Scholar] [CrossRef]
- Siddiqui, J.A.; et al. GDF15 promotes prostate cancer bone metastasis and colonization through osteoblastic CCL2 and RANKL activation. Bone Res 2022, 10, 6. [Google Scholar] [CrossRef]
- Lee, S.H., C. Krisanapun, and S.J. Baek, NSAID-activated gene-1 as a molecular target for capsaicin-induced apoptosis through a novel molecular mechanism involving GSK3beta, C/EBPbeta and ATF3. Carcinogenesis 2010, 31, 719–28. [CrossRef]
- Patsalos, A.; et al. A growth factor-expressing macrophage subpopulation orchestrates regenerative inflammation via GDF-15. J Exp Med 2022, 219. [Google Scholar] [CrossRef]
- Eisenstein, A.; et al. Activation of the transcription factor NRF2 mediates the anti-inflammatory properties of a subset of over-the-counter and prescription NSAIDs. Immunity 2022, 55, 1082–1095. [Google Scholar] [CrossRef]
- L'Homme, L.; et al. Saturated Fatty Acids Promote GDF15 Expression in Human Macrophages through the PERK/eIF2/CHOP Signaling Pathway. Nutrients 2020, 12. [Google Scholar] [CrossRef]
- Ratnam, N.M.; et al. NF-kappaB regulates GDF-15 to suppress macrophage surveillance during early tumor development. J Clin Invest 2017, 127, 3796–3809. [Google Scholar] [CrossRef]
- Wang, T.; et al. YAP promotes breast cancer metastasis by repressing growth differentiation factor-15. Biochim Biophys Acta Mol Basis Dis 2018, 1864 5 Pt A, 1744–1753. [Google Scholar] [CrossRef]
- Emmerson, P.J.; et al. The metabolic effects of GDF15 are mediated by the orphan receptor GFRAL. Nat Med 2017, 23, 1215–1219. [Google Scholar] [CrossRef]
- Yang, L.; et al. GFRAL is the receptor for GDF15 and is required for the anti-obesity effects of the ligand. Nat Med 2017, 23, 1158–1166. [Google Scholar] [CrossRef]
- Mullican, S.E.; et al. GFRAL is the receptor for GDF15 and the ligand promotes weight loss in mice and nonhuman primates. Nat Med 2017, 23, 1150–1157. [Google Scholar] [CrossRef]
- Moon, J.S.; et al. Growth differentiation factor 15 protects against the aging-mediated systemic inflammatory response in humans and mice. Aging Cell 2020, 19, e13195. [Google Scholar] [CrossRef]
- Li, C.; et al. GDF15 promotes EMT and metastasis in colorectal cancer. Oncotarget 2016, 7, 860–72. [Google Scholar] [CrossRef]
- Artz, A., S. Butz, and D. Vestweber, GDF-15 inhibits integrin activation and mouse neutrophil recruitment through the ALK-5/TGF-betaRII heterodimer. Blood 2016, 128, 529–41. [CrossRef]
- Olsen, O.E.; et al. TGF-beta contamination of purified recombinant GDF15. PLoS One 2017, 12, e0187349. [Google Scholar] [CrossRef]
- Tsai, V.W.W.; et al. The MIC-1/GDF15-GFRAL Pathway in Energy Homeostasis: Implications for Obesity, Cachexia, and Other Associated Diseases. Cell Metab 2018, 28, 353–368. [Google Scholar] [CrossRef]
- Wang, Z.; et al. GDF15 induces immunosuppression via CD48 on regulatory T cells in hepatocellular carcinoma. J Immunother Cancer 2021, 9. [Google Scholar] [CrossRef]
- Schledzewski, K.; et al. Deficiency of liver sinusoidal scavenger receptors stabilin-1 and -2 in mice causes glomerulofibrotic nephropathy via impaired hepatic clearance of noxious blood factors. J Clin Invest 2011, 121, 703–14. [Google Scholar] [CrossRef]
- Larionova, I.; et al. New Angiogenic Regulators Produced by TAMs: Perspective for Targeting Tumor Angiogenesis. Cancers (Basel) 2021, 13. [Google Scholar] [CrossRef]
- Kzhyshkowska, J. , Multifunctional receptor stabilin-1 in homeostasis and disease. ScientificWorldJournal 2010, 10, 2039–53. [Google Scholar] [CrossRef]
- Carai, P.; et al. Stabilin-1 mediates beneficial monocyte recruitment and tolerogenic macrophage programming during CVB3-induced viral myocarditis. J Mol Cell Cardiol 2022, 165, 31–39. [Google Scholar] [CrossRef]
- Ryabov, V.; et al. Cardiac CD68+ and stabilin-1+ macrophages in wound healing following myocardial infarction: From experiment to clinic. Immunobiology 2018, 223, 413–421. [Google Scholar] [CrossRef]
- Larionova, I.; et al. Tumor-Associated Macrophages in Human Breast, Colorectal, Lung, Ovarian and Prostate Cancers. Front Oncol 2020, 10, 566511. [Google Scholar] [CrossRef]
- Holte, C.; et al. Highly oxidized albumin is cleared by liver sinusoidal endothelial cells via the receptors stabilin-1 and -2. Sci Rep 2023, 13, 19121. [Google Scholar] [CrossRef]
- Larionova, I.; et al. Tumor-associated macrophages respond to chemotherapy by detrimental transcriptional reprogramming and suppressing stabilin-1 mediated clearance of EGF. Front Immunol 2023, 14, 1000497. [Google Scholar] [CrossRef]
- Kzhyshkowska, J., A. Gratchev, and S. Goerdt, Stabilin-1, a homeostatic scavenger receptor with multiple functions. J Cell Mol Med 2006, 10, 635–49. [CrossRef]
- Riabov, V.; et al. Stabilin-1 is expressed in human breast cancer and supports tumor growth in mammary adenocarcinoma mouse model. Oncotarget 2016, 7, 31097–110. [Google Scholar] [CrossRef]
- Riabov, V.; et al. Role of tumor associated macrophages in tumor angiogenesis and lymphangiogenesis. Front Physiol 2014, 5, 75. [Google Scholar] [CrossRef]
- Kzhyshkowska, J.; et al. Alternatively activated macrophages regulate extracellular levels of the hormone placental lactogen via receptor-mediated uptake and transcytosis. J Immunol 2008, 180, 3028–37. [Google Scholar] [CrossRef]
- Tamura, Y.; et al. FEEL-1 and FEEL-2 are endocytic receptors for advanced glycation end products. J Biol Chem 2003, 278, 12613–7. [Google Scholar] [CrossRef]
- Zhang, J.; et al. A novel GGA-binding site is required for intracellular sorting mediated by stabilin-1. Mol Cell Biol 2009, 29, 6097–105. [Google Scholar] [CrossRef]
- Kzhyshkowska, J.; et al. Phosphatidylinositide 3-kinase activity is required for stabilin-1-mediated endosomal transport of acLDL. Immunobiology 2005, 210, 161–173. [Google Scholar] [CrossRef]
- Park, H.; et al. Ionizing Radiation-Induced GDF15 Promotes Angiogenesis in Human Glioblastoma Models by Promoting VEGFA Expression Through p-MAPK1/SP1 Signaling. Front Oncol 2022, 12, 801230. [Google Scholar] [CrossRef]
- Ho, J.E.; et al. Clinical and genetic correlates of growth differentiation factor 15 in the community. Clin Chem 2012, 58, 1582–91. [Google Scholar] [CrossRef] [PubMed]
- Diaz, M.; et al. Circulating growth-and-differentiation factor-15 in early life: relation to prenatal and postnatal growth and adiposity measurements. Pediatr Res 2020, 87, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; et al. Measurement of serum levels of macrophage inhibitory cytokine 1 combined with prostate-specific antigen improves prostate cancer diagnosis. Clin Cancer Res 2006, 12, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Koopmann, J.; et al. Serum markers in patients with resectable pancreatic adenocarcinoma: macrophage inhibitory cytokine 1 versus CA19-9. Clin Cancer Res 2006, 12, 442–6. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.A.; et al. MIC-1 serum level and genotype: associations with progress and prognosis of colorectal carcinoma. Clin Cancer Res 2003, 9, 2642–50. [Google Scholar] [PubMed]
- Notz, Q.; et al. Pro- and Anti-Inflammatory Responses in Severe COVID-19-Induced Acute Respiratory Distress Syndrome-An Observational Pilot Study. Front Immunol 2020, 11, 581338. [Google Scholar] [CrossRef] [PubMed]
- Staff, A.C.; et al. Elevated plasma growth differentiation factor-15 correlates with lymph node metastases and poor survival in endometrial cancer. Clin Cancer Res 2011, 17, 4825–33. [Google Scholar] [CrossRef] [PubMed]
- Kempf, T.; et al. Circulating concentrations of growth-differentiation factor 15 in apparently healthy elderly individuals and patients with chronic heart failure as assessed by a new immunoradiometric sandwich assay. Clin Chem 2007, 53, 284–91. [Google Scholar] [CrossRef] [PubMed]
- Yatsuga, S.; et al. Growth differentiation factor 15 as a useful biomarker for mitochondrial disorders. Ann Neurol 2015, 78, 814–23. [Google Scholar] [CrossRef]
- Temel Yuksel, I.; et al. Maternal levels of growth differentiation factor-15 in patients with preeclampsia. Hypertens Pregnancy 2018, 37, 192–196. [Google Scholar] [CrossRef]
- Traeger, L.; et al. Serum Hepcidin and GDF-15 levels as prognostic markers in urothelial carcinoma of the upper urinary tract and renal cell carcinoma. BMC Cancer 2019, 19, 74. [Google Scholar] [CrossRef]
- Hagstrom, E.; et al. Growth Differentiation Factor 15 Predicts All-Cause Morbidity and Mortality in Stable Coronary Heart Disease. Clin Chem 2017, 63, 325–333. [Google Scholar] [CrossRef]
- Xiong, Y.; et al. Long-acting MIC-1/GDF15 molecules to treat obesity: Evidence from mice to monkeys. Sci Transl Med 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Breen, D.M.; et al. Growth differentiation factor 15 neutralization does not impact anorexia or survival in lipopolysaccharide-induced inflammation. iScience 2021, 24, 102554. [Google Scholar] [CrossRef]
- Jena, J.; et al. GDF15 is required for cold-induced thermogenesis and contributes to improved systemic metabolic health following loss of OPA1 in brown adipocytes. Elife 2023, 12. [Google Scholar] [CrossRef]
- Lu, J.F.; et al. GDF15 is a major determinant of ketogenic diet-induced weight loss. Cell Metab 2023, 35, 2165–2182. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.C. and D.Y. Xu, Research progress on the role and mechanism of GDF15 in body weight regulation. Obes Facts 2023. [Google Scholar] [CrossRef] [PubMed]
- Breit, S.N., D.A. Brown, and V.W. Tsai, The GDF15-GFRAL Pathway in Health and Metabolic Disease: Friend or Foe? Annu Rev Physiol 2021, 83, 127–151. [CrossRef]
- Wang, D.; et al. GDF15: emerging biology and therapeutic applications for obesity and cardiometabolic disease. Nat Rev Endocrinol 2021, 17, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.C.; et al. Association between growth differentiation factor 15 levels and gestational diabetes mellitus: A combined analysis. Front Endocrinol (Lausanne) 2023, 14, 1084896. [Google Scholar] [CrossRef] [PubMed]
- Wang, L. and Q. Yang, Circulating Growth Differentiation Factor 15 and Preeclampsia: A Meta-Analysis. Horm Metab Res 2023, 55, 114–123. [CrossRef]
- Govaere, O.; et al. Transcriptomic profiling across the nonalcoholic fatty liver disease spectrum reveals gene signatures for steatohepatitis and fibrosis. Sci Transl Med 2020, 12. [Google Scholar] [CrossRef]
- Li, X.; et al. GDF15 Ameliorates Liver Fibrosis by Metabolic Reprogramming of Macrophages to Acquire Anti-Inflammatory Properties. Cell Mol Gastroenterol Hepatol 2023. [Google Scholar] [CrossRef] [PubMed]
- Riabov, V.; et al. High erythroferrone expression in CD71(+) erythroid progenitors predicts superior survival in myelodysplastic syndromes. Br J Haematol 2021, 192, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Pence, B.D., J.R. Yarbro, and R.S. Emmons, Growth differentiation factor-15 is associated with age-related monocyte dysfunction. Aging Med (Milton) 2021, 4, 47–52. [CrossRef] [PubMed]
- Ebihara, T.; et al. Cytokine Elevation in Severe COVID-19 From Longitudinal Proteomics Analysis: Comparison With Sepsis. Front Immunol 2021, 12, 798338. [Google Scholar] [CrossRef] [PubMed]
- Buendgens, L.; et al. Growth Differentiation Factor-15 Is a Predictor of Mortality in Critically Ill Patients with Sepsis. Dis Markers 2017, 2017, 5271203. [Google Scholar] [CrossRef]
- Santos, I.; et al. CXCL5-mediated recruitment of neutrophils into the peritoneal cavity of Gdf15-deficient mice protects against abdominal sepsis. Proc Natl Acad Sci U S A 2020, 117, 12281–12287. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; et al. [Study on the value of mitochondrial associated protein fumarylacetoacetate domain containing protein 1 and growth differentiation factor-15 in the diagnosis of sepsis: test results from the patients of a multicenter study]. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2021, 33, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X. and S. Dong, Protective effect of growth differentiation factor 15 in sepsis by regulating macrophage polarization and its mechanism. Bioengineered 2022, 13, 9687–9707. [CrossRef]
- Ishak, S.R.; et al. Serum growth differentiation factor-15 (GDF-15) is a biomarker of cardiac manifestations in children with COVID-19. Eur J Med Res 2023, 28, 527. [Google Scholar] [CrossRef]
- Li, M.; et al. GDF-15 prevents LPS and D-galactosamine-induced in fl ammation and acute liver injury in mice. Int J Mol Med 2018, 42, 1756–1764. [Google Scholar] [CrossRef]
- Luan, H.H.; et al. GDF15 Is an Inflammation-Induced Central Mediator of Tissue Tolerance. Cell 2019, 178, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Luan, H.H.; et al. GDF15 Is an Inflammation-Induced Central Mediator of Tissue Tolerance. Cell 2019, 178, 1231–1244. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; et al. GDF15 promotes weight loss by enhancing energy expenditure in muscle. Nature 2023, 619, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Patel, A.R.; et al. LPS induces rapid increase in GDF15 levels in mice, rats, and humans but is not required for anorexia in mice. Am J Physiol Gastrointest Liver Physiol 2022, 322, G247–G255. [Google Scholar] [CrossRef] [PubMed]
- Vanhara, P.; et al. Growth/differentiation factor-15 inhibits differentiation into osteoclasts--a novel factor involved in control of osteoclast differentiation. Differentiation 2009, 78, 213–22. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.K.; et al. GDF15 deficiency exacerbates chronic alcohol- and carbon tetrachloride-induced liver injury. Sci Rep 2017, 7, 17238. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; et al. GDF15 Ameliorates Liver Fibrosis by Metabolic Reprogramming of Macrophages to Acquire Anti-Inflammatory Properties. Cell Mol Gastroenterol Hepatol 2023, 16, 711–734. [Google Scholar] [CrossRef]
- Al-Mudares, F.; et al. Loss of growth differentiation factor 15 exacerbates lung injury in neonatal mice. Am J Physiol Lung Cell Mol Physiol 2023, 325, L314–L326. [Google Scholar] [CrossRef]
- Kim, H.H.; et al. Catecholamine induces Kupffer cell apoptosis via growth differentiation factor 15 in alcohol-associated liver disease. Exp Mol Med 2023, 55, 158–170. [Google Scholar] [CrossRef]
- Mishra, R.; et al. Transplanted allogeneic cardiac progenitor cells secrete GDF-15 and stimulate an active immune remodeling process in the ischemic myocardium. J Transl Med 2022, 20, 323. [Google Scholar] [CrossRef]
- Saini, M.; et al. Implant biomaterials: A comprehensive review. World J Clin Cases 2015, 3, 52–7. [Google Scholar] [CrossRef] [PubMed]
- Silva-Bermudez, L.S.; et al. Titanium Nanoparticles Enhance Production and Suppress Stabilin-1-Mediated Clearance of GDF-15 in Human Primary Macrophages. Front Immunol 2021, 12, 760577. [Google Scholar] [CrossRef] [PubMed]
- Muniyan, S.; et al. Macrophage inhibitory cytokine-1 in cancer: Beyond the cellular phenotype. Cancer Lett 2022, 536, 215664. [Google Scholar] [CrossRef] [PubMed]
- Spanopoulou, A. and V. Gkretsi, Growth differentiation factor 15 (GDF15) in cancer cell metastasis: from the cells to the patients. Clin Exp Metastasis 2020, 37, 451–464. [CrossRef] [PubMed]
- Urakawa, N.; et al. GDF15 derived from both tumor-associated macrophages and esophageal squamous cell carcinomas contributes to tumor progression via Akt and Erk pathways. Lab Invest 2015, 95, 491–503. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; et al. Downregulation of GDF15 suppresses ferroptosis and predicts unfavorable prognosis in clear cell renal cell carcinoma. Cell Div 2023, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Mielcarska, S.; et al. GDF-15 Level Correlates with CMKLR1 and VEGF-A in Tumor-free Margin in Colorectal Cancer. Curr Med Sci 2021, 41, 522–528. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.K.; et al. Macrophage inhibitory cytokine-1 activates AKT and ERK-1/2 via the transactivation of ErbB2 in human breast and gastric cancer cells. Carcinogenesis 2008, 29, 704–12. [Google Scholar] [CrossRef] [PubMed]
- Song, H., D. Yin, and Z. Liu, GDF-15 promotes angiogenesis through modulating p53/HIF-1alpha signaling pathway in hypoxic human umbilical vein endothelial cells. Mol Biol Rep 2012, 39, 4017–22. [CrossRef]
- Wang, Z.; et al. YKL-40 derived from infiltrating macrophages cooperates with GDF15 to establish an immune suppressive microenvironment in gallbladder cancer. Cancer Lett 2023, 563, 216184. [Google Scholar] [CrossRef]
- Peng, H.; et al. Growth and differentiation factor 15 regulates PD-L1 expression in glioblastoma. Cancer Manag Res 2019, 11, 2653–2661. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.; et al. Plasma GDF15 levels associated with circulating immune cells predict the efficacy of PD-1/PD-L1 inhibitor treatment and prognosis in patients with advanced non-small cell lung cancer. J Cancer Res Clin Oncol 2023, 149, 159–171. [Google Scholar] [CrossRef]
- Haake, M.; et al. Tumor-derived GDF-15 blocks LFA-1 dependent T cell recruitment and suppresses responses to anti-PD-1 treatment. Nat Commun 2023, 14, 4253. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; et al. HIF1alpha promotes tumor chemoresistance via recruiting GDF15-producing TAMs in colorectal cancer. Exp Cell Res 2021, 398, 112394. [Google Scholar] [CrossRef]
- Yu, S.; et al. Activated HIF1alpha of tumor cells promotes chemoresistance development via recruiting GDF15-producing tumor-associated macrophages in gastric cancer. Cancer Immunol Immunother 2020, 69, 1973–1987. [Google Scholar] [CrossRef] [PubMed]
- Bonaterra, G.A.; et al. Increased Density of Growth Differentiation Factor-15+ Immunoreactive M1/M2 Macrophages in Prostate Cancer of Different Gleason Scores Compared with Benign Prostate Hyperplasia. Cancers (Basel) 2022, 14. [Google Scholar] [CrossRef]
- Sadasivan, S.M.; et al. The interplay of growth differentiation factor 15 (GDF15) expression and M2 macrophages during prostate carcinogenesis. Carcinogenesis 2020, 41, 1074–1082. [Google Scholar] [CrossRef]
- Lv, C.; et al. M1 Macrophages Enhance Survival and Invasion of Oral Squamous Cell Carcinoma by Inducing GDF15-Mediated ErbB2 Phosphorylation. ACS Omega 2022, 7, 11405–11414. [Google Scholar] [CrossRef]
- Li, S.; et al. GDF15 promotes the proliferation of cervical cancer cells by phosphorylating AKT1 and Erk1/2 through the receptor ErbB2. J Exp Clin Cancer Res 2018, 37, 80. [Google Scholar] [CrossRef]
- Joo, M.; et al. GDF15 Promotes Cell Growth, Migration, and Invasion in Gastric Cancer by Inducing STAT3 Activation. Int J Mol Sci 2023, 24. [Google Scholar] [CrossRef]
- Peake, B.F.; et al. Growth differentiation factor 15 mediates epithelial mesenchymal transition and invasion of breast cancers through IGF-1R-FoxM1 signaling. Oncotarget 2017, 8, 94393–94406. [Google Scholar] [CrossRef] [PubMed]
- Dong, G.; et al. SCAP Mediated GDF15-Induced Invasion and EMT of Esophageal Cancer. Front Oncol 2020, 10, 564785. [Google Scholar] [CrossRef]
- Duan, L.; et al. The role of GDF15 in bone metastasis of lung adenocarcinoma cells. Oncol Rep 2019, 41, 2379–2388. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; et al. c-Fos separation from Lamin A/C by GDF15 promotes colon cancer invasion and metastasis in inflammatory microenvironment. J Cell Physiol 2020, 235, 4407–4421. [Google Scholar] [CrossRef]
- Campderros, L.; et al. Brown Adipocytes Secrete GDF15 in Response to Thermogenic Activation. Obesity (Silver Spring) 2019, 27, 1606–1616. [Google Scholar] [CrossRef]
- Wang, Y.; et al. Growth-differentiation factor-15 predicts adverse cardiac events in patients with acute coronary syndrome: A meta-analysis. Am J Emerg Med 2019, 37, 1346–1352. [Google Scholar] [CrossRef]
- Ferreira, J.P.; et al. Growth differentiation factor-15 and the effect of empagliflozin in heart failure: Findings from the EMPEROR program. Eur J Heart Fail 2023. [Google Scholar] [CrossRef]
- Molek-Dziadosz, P.; et al. Elevated growth differentiation factor 15 is associated with echocardiographic markers of right heart overload in atrial fibrillation. Pol Arch Intern Med 2023, 133. [Google Scholar] [CrossRef]
- Xie, S., L. Lu, and L. Liu, Growth differentiation factor-15 and the risk of cardiovascular diseases and all-cause mortality: A meta-analysis of prospective studies. Clin Cardiol 2019, 42, 513–523. [CrossRef] [PubMed]
- Kempf, T.; et al. GDF-15 is an inhibitor of leukocyte integrin activation required for survival after myocardial infarction in mice. Nat Med 2011, 17, 581–8. [Google Scholar] [CrossRef]
- Ackermann, K.; et al. Growth differentiation factor-15 regulates oxLDL-induced lipid homeostasis and autophagy in human macrophages. Atherosclerosis 2019, 281, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Heduschke, A.; et al. GDF-15 Deficiency Reduces Autophagic Activity in Human Macrophages In Vitro and Decreases p62-Accumulation in Atherosclerotic Lesions in Mice. Cells 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; et al. GDF-15 Suppresses Atherosclerosis by Inhibiting oxLDL-Induced Lipid Accumulation and Inflammation in Macrophages. Evid Based Complement Alternat Med 2021, 2021, 6497568. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
GDF-15 crystal protein structure and phylogenetic associations. (a) Crystal structure of the GDF-15 protein. Obtained from InterPro: https://www.ebi.ac.uk/interpro/structure/PDB/5vt2/#table [27]; (b) Phylogenetic tree illustrating relationships among GDF-15, TGF-β, and GDNF superfamily members. Generated using TaxOnTree (bioinfo.icb.ufmg.br/taxontree/#x); (c) Protein sequence alignment of GDF-15 compared to TGF-β and GDNF superfamily members. Sequence alignment was performed using Protein BLAST (blast.ncbi.nlm.nih.gov). cov: percentage of coverage, pid: percentage of identity.
Figure 1.
GDF-15 crystal protein structure and phylogenetic associations. (a) Crystal structure of the GDF-15 protein. Obtained from InterPro: https://www.ebi.ac.uk/interpro/structure/PDB/5vt2/#table [27]; (b) Phylogenetic tree illustrating relationships among GDF-15, TGF-β, and GDNF superfamily members. Generated using TaxOnTree (bioinfo.icb.ufmg.br/taxontree/#x); (c) Protein sequence alignment of GDF-15 compared to TGF-β and GDNF superfamily members. Sequence alignment was performed using Protein BLAST (blast.ncbi.nlm.nih.gov). cov: percentage of coverage, pid: percentage of identity.
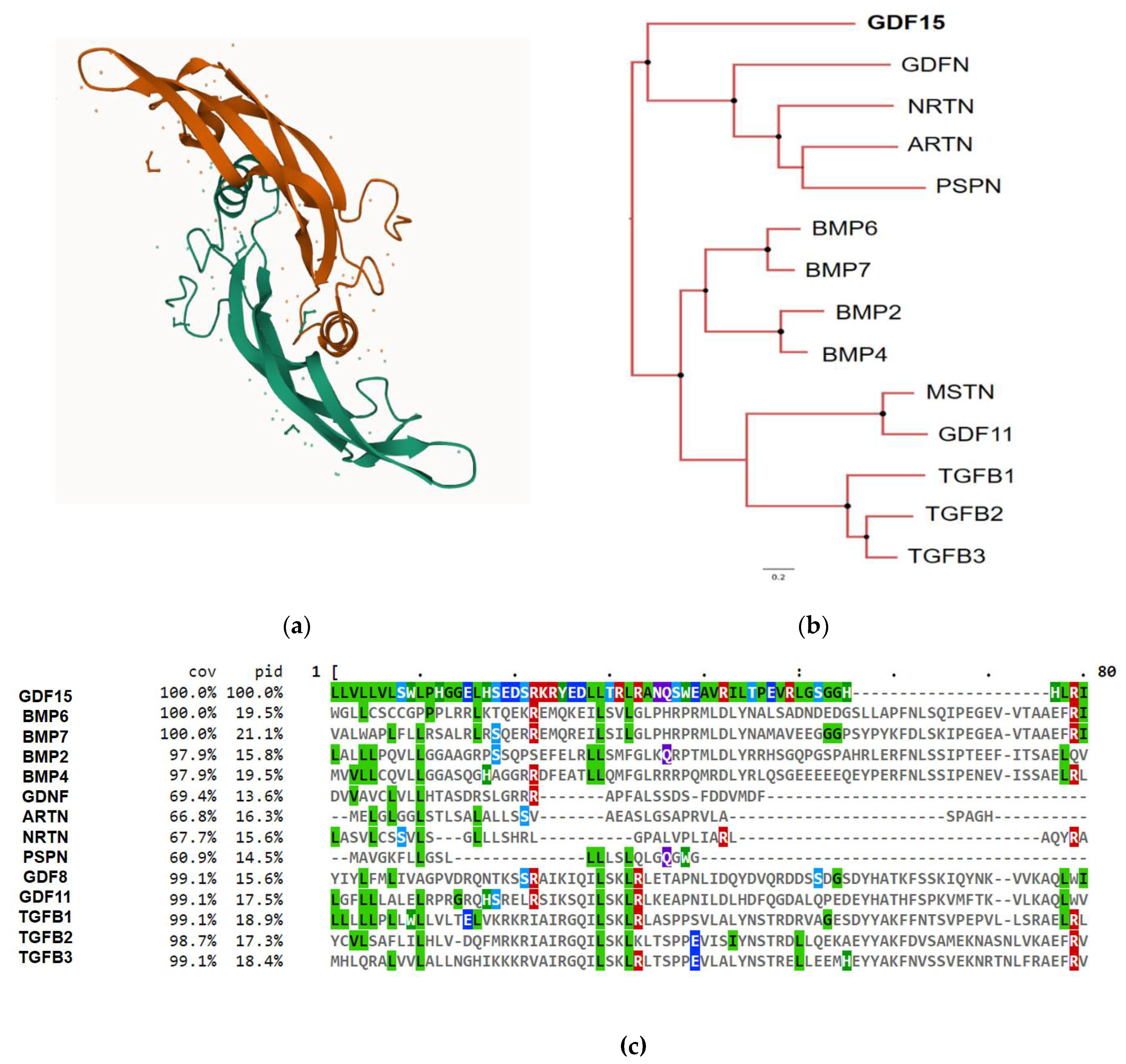
Figure 2.
GDF-15 RNA and protein expression in the human body. GDF-15 expression is elevated in various tissues, including kidney, bladder, brain, and female and male tissues. RNA expression levels are represented using normalized transcript expression values (nTPM). Adapted from: Human Protein Atlas https://www.proteinatlas.org/ENSG00000130513-GDF15/tissue.
Figure 2.
GDF-15 RNA and protein expression in the human body. GDF-15 expression is elevated in various tissues, including kidney, bladder, brain, and female and male tissues. RNA expression levels are represented using normalized transcript expression values (nTPM). Adapted from: Human Protein Atlas https://www.proteinatlas.org/ENSG00000130513-GDF15/tissue.
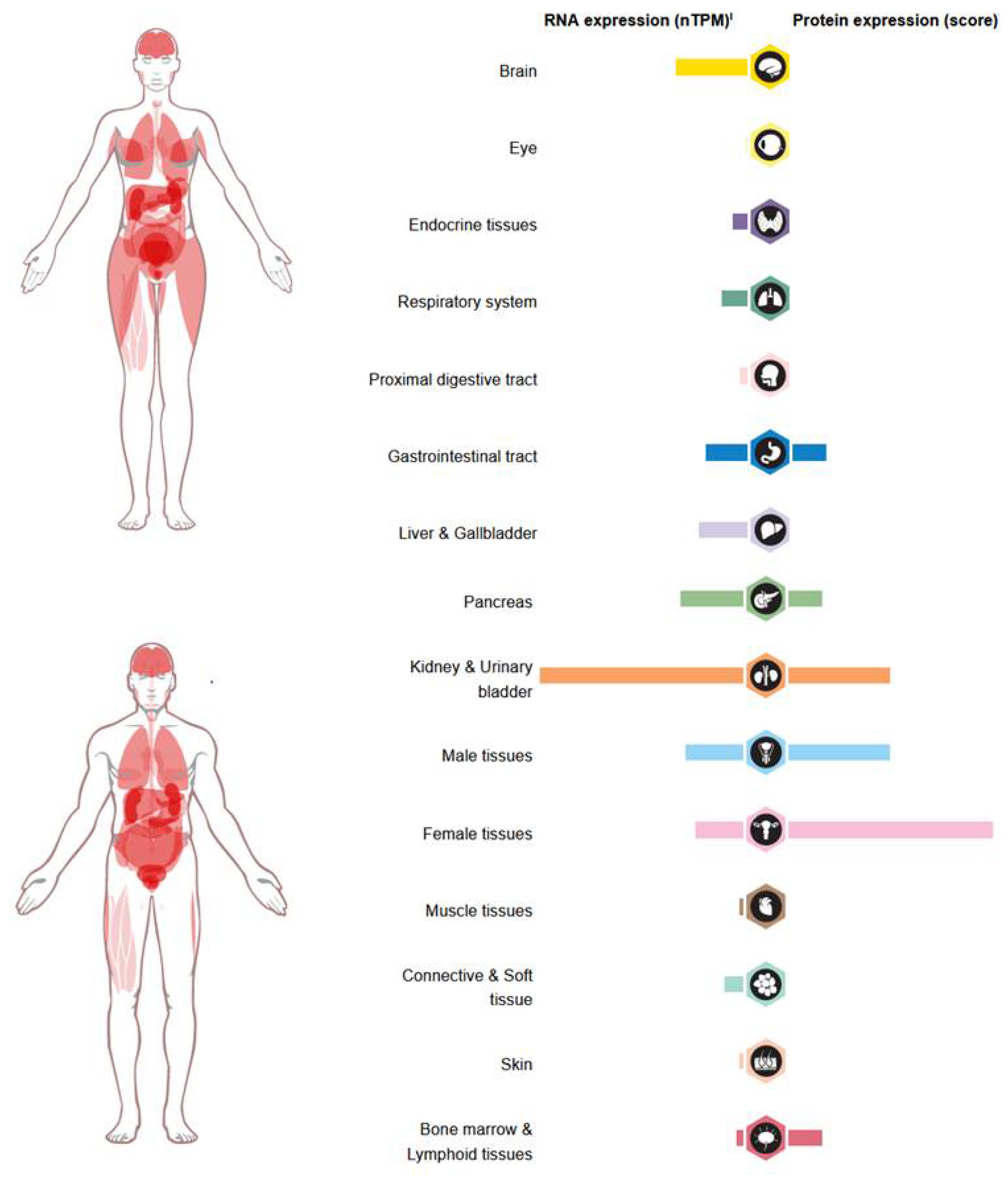
Figure 3.
Effects of GDF-15 in carcinogenesis. Image created by biorender.com
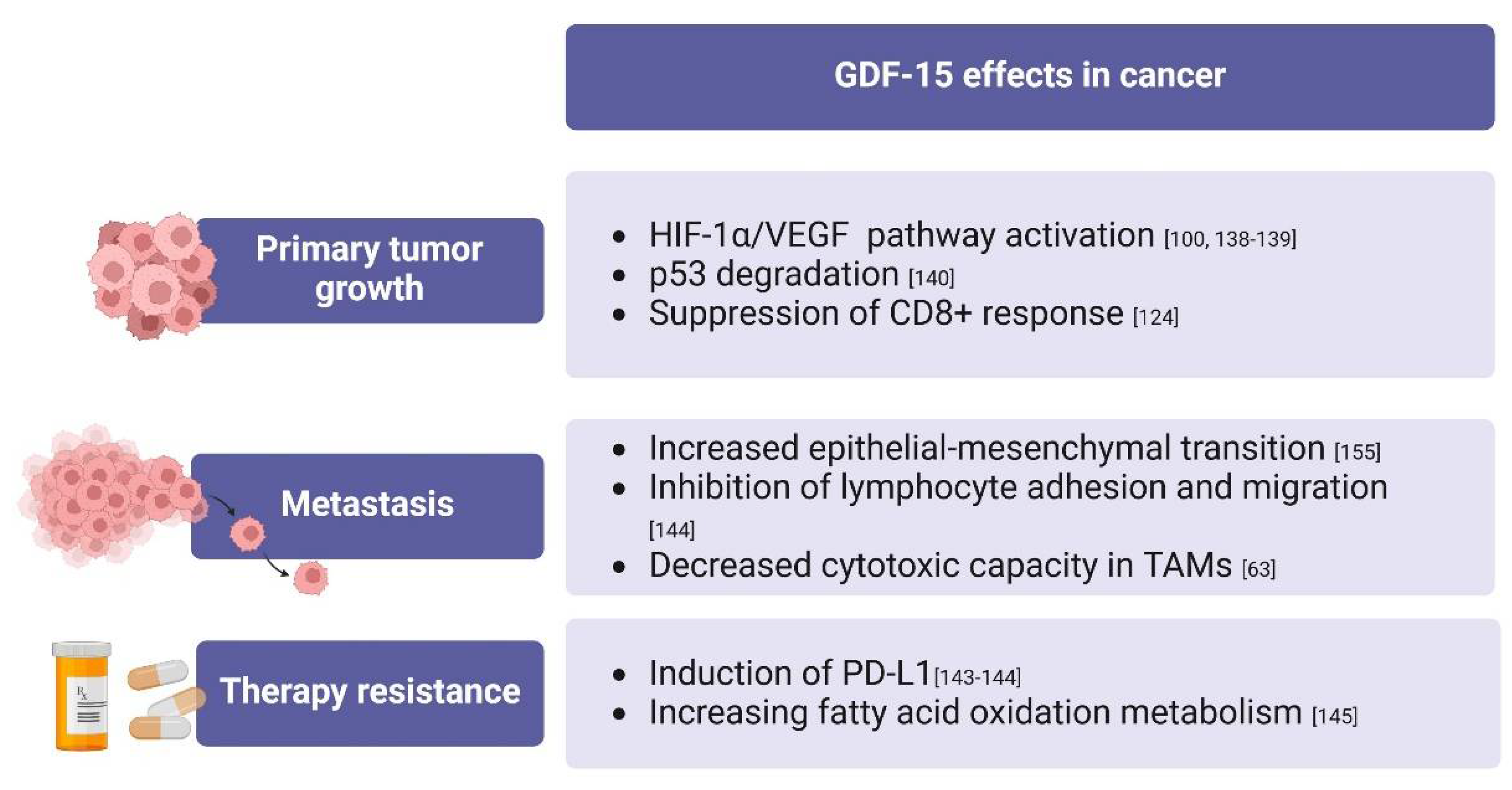
Figure 4.
Overview of the effects of GDF-15 in the macrophage system. Image created by biorender.com
Figure 4.
Overview of the effects of GDF-15 in the macrophage system. Image created by biorender.com
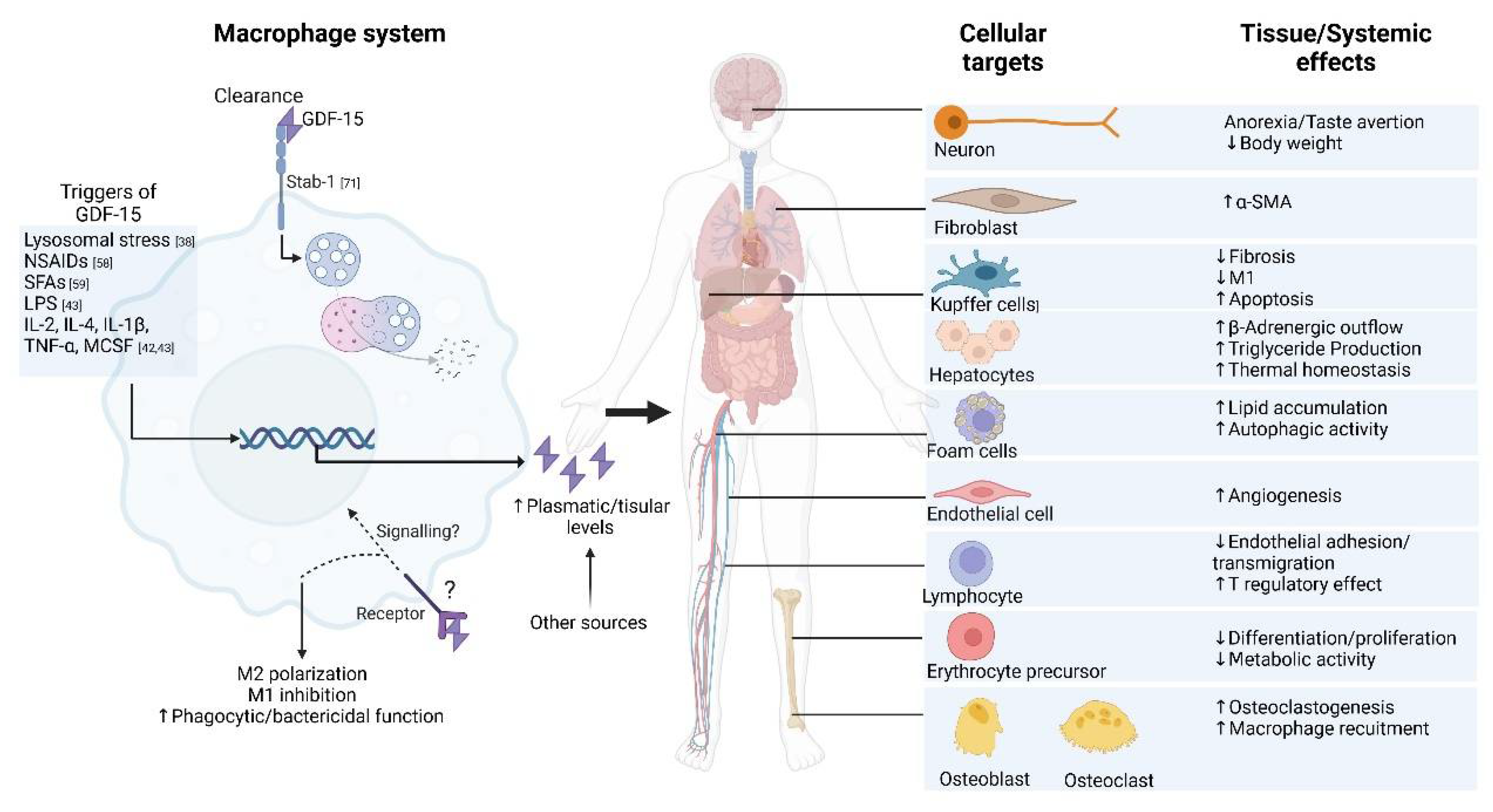

Table 1.
GDF-15 producing cells.
| Primary cells | Species | Conditions | Measurement | Reference | |
|---|---|---|---|---|---|
| Adipose tissue macrophages | Human | Obesity | RT-PCR | [41] | |
| Rosiglitazone-treated | Microarray | [45] | |||
| Human nasal epithelial cell | Human | LPS treatment | RT-PCR | [46] | |
| Lung fibroblasts | Mouse | Bleomycin treatment | RT-PCR | [47] | |
| Cardiomyocytes | Rat | Stretch conditions | RT-PCR | [48] | |
| Chief cells | Rat/Human | Obesity | Immunohistochemistry | [37] | |
| Erythroid cells | Human | Maturation | RT-PCR | [49] | |
| Megakaryocytes | Human | Primary myelofibrosis | Immunohistochemistry | [44] | |
| Primary erythroblasts | Human | 7 and 14d | RT-PCR, ELISA | [43] | |
| Hepatocytes | Human | Metformin treatment | RT-PCR | [50] | |
| Cell lines | |||||
| Macrophages-like cells | |||||
| THP-1 | Human | LPS treatment | RT-PCR | [51] | |
| RAW264.7 | Mouse | ||||
| Endothelial cells | |||||
| Endothelial colony-forming cells generated from blood | Human | 7d | RT-PCR, ELISA | [52] | |
| Human aortic endothelial cells | Human | hrCRP | RT-PCR, ELISA | [53] | |
| Connective and soft tissue cells | |||||
| Adipocytes | Human | SAT-differentiated | RT-PCR, ELISA | [38] | |
| Human | Oligomycin treatment | ELISA | |||
| Mouse | rIL-4 and rIL-13 | RT-PCR, ELISA | [54] | ||
| Embryonic adipocyte-like cell line (3T3-L1) | Mouse | Tunicamycin treatment | RT-PCR | [39] | |
| Myoblasts (C2C12) | Mouse | . | RT-PCR, Western blot | [55] | |
| Trophoblastic cells (BeWo) | Human | 24h | ELISA | [28] | |
| Cancer cells | |||||
| Alveolar basal epithelial cells (A549) | Human | Bleomycin treatment, C5a treatment | RT-PCR | [40,56] | |
| Colorectal cancer cells (HCT-116) | Human | NSAID, Indomethacin | Northern/ Western blot | [18] | |
| Hypopharyngeal carcinoma cells (FaDu cells) | Human | Head and neck cancer | Western blot | [57] | |
| Prostate cancer cells (LNCaP-C81 cell line) |
Human | Prostate cancer | Western blot | [58] | |
CRP: C-reactive protein; hr: human recombinant; GDF-15: growth differentiation factor 15; ELISA: enzyme-linked immunosorbent assay; LPS: lipopolysaccharide; NSAID: non-steroidal anti-inflammatory drug; RT-PCR: reverse transcription real-time polymerase chain reaction; SAT: Subcutaneous adipose tissue; THP-1: human acute monocytic leukemia cell line.
Table 2.
Transcription factors regulating GDF-15 expression.
| Transcription factors | Expressed in macrophages | Cells | Conditions | Reference |
|---|---|---|---|---|
| ATF4 | + | Human nasal epithelial cells | LPS treatment | [46] |
| Murine hepatocytes | Metformin treatment | [50] | ||
| Murine embryonic adipocyte-like cell line (3T3-L1) | Tunicamycin treatment | [39] | ||
| CHOP | PMA-differentiated THP-1, PBMCs | SFAs treatment | [62] | |
| + | Murine C2C12 myoblasts | CRIF1 deficiency | [55] | |
| Murine hepatocytes | Metformin treatment | [50] | ||
| EGR1 | + | Hypopharyngeal carcinoma cell line (FaDu), human epithelial carcinoma cell line (KB) | . | [57] |
| KLF5 | + | Adenocarcinoma alveolar basal epithelial cells (A549) | C5a treatment | [56] |
| NF-κB | + | Immortalized mouse embryonic Fibroblasts | . | [63] |
| NRF2 | + | Murine and human BMDM | NSAIDs | [61] |
| p53 | + | Human aortic endothelial cells | CRP supplementation | [53] |
| Sp1, Sp3 | + | Human colorectal cancer cells (HCT-116) | . | [18] |
| STAT6 | + | Murine adipocytes | IL-13 treatment | [54] |
| TFEB | + | Human and murine adipose tissue macrophages | Obesity | [41] |
| YAP* | - | Breast cancer cells and cytotrophoblast | . | [64] |
*YAP has been shown to act as a negative regulator of GDF-15 expression. ATF: activating transcription factor; BMDM: blood peripheral circulating bone marrow-derived monocytes; CHOP: C/EBP homologous protein; CRIF1: Cytokine response 6 (CR6)-interacting factor 1; CRP: C-reactive protein; EGR1: early growth response 1; KLF: Kruppel-like factor; NF-κB: nuclear factor kappa-light-chain-enhancer of activated B cells; NRF2: Nuclear factor erythroid 2-related factor 2; NSAID: non-steroidal anti-inflammatory drug; PBMC: peripheral blood mononuclear cells; PMA: phorbol 12-myristate 13-acetate; SFA: saturated fatty acids; STAT6: signal transducer and activator of transcription 6; TFEB: transcription factor EB; THP-1: human acute monocytic leukemia cell line; YAP: Yes-associated protein.
Table 3.
Effects of rGDF-15.
| Effects | Evidence | Method | Target cells | Conditions | GDF-15 source | Concentration (ng/mL) | Reference |
|---|---|---|---|---|---|---|---|
| Increased M2 and decreased M1 polarization | Increased Arg-1 expression | RT-PCR, Flow cytometry | THP-1 and RAW264.7 | rGDF15 for 48h | CHO; E. coli | 100 | [40] |
| Decreased IL-6, TNF-α, MCP-1, and IL-10 secretion, decreased CD80, increased CD163 mRNA levels | |||||||
| Fibroblast activation | Increased α-SMA protein expression | Western blot | WI-38 | Preincubation for 48h; rGDF15 for 72h | CHO; E. coli | 0-100 | [40] |
| Increased Smad2/3 phosphorylation through TGF-β I receptor | Western blot | WI-38 | Preincubation for 48h; rGDF15 for 20 min-1h | CHO; E. coli | 100 | [40] | |
| Reduced metabolic activity in erythroid cells | Decreased optical density with increasing GDF-15 concentration | RT-PCR, Flow cytometry, MTT cell metabolic activity assay | K562 | Treatment with β-thalassemia serum containing 48 ng/mL of GDF-15 or GDF-15 for 24h, 48h and 72h | CHO | 2-50 | [49] |
| Reduced proliferation in erythroid cells | Decreased optical density with increasing GDF-15 concentration in MTT assay | CFSE proliferation assay | K562 | GDF-15 for 5d | CHO | 10-50 | [49] |
| Increased angiogenesis | Increased VEGFA expression | Tube formation assay, RT-PCR | U373, HBMVEC | 3 d in culture | CHO | 100 | [89] |
Arg-1: Arginase-1; CFSE: carboxyfluorescein succinimidyl ester; CHO: chinese hamster ovary; GDF-15: growth differentiation factor 15; HBMVEC: human brain microvascular endothelial cells; K562: lymphoblast; MCP-1: Monocyte Chemoattractant Protein-1; RT-PCR: reverse transcription real-time polymerase chain reaction; TGF-β: transforming growth factor-beta; THP-1: human acute monocytic leukemia cell line; TNF-α: tumor necrosis factor α; U373: glioblastoma cells; VEGF: vascular endothelial growth factor; WI-38: fibroblasts from lung tissue.
Table 4.
GDF-15 serum levels in pathology.
| Condition | Concentration (pg/mL) | Reference |
|---|---|---|
| α-thalassemia syndrome | 5900 ± 1200 | [43] |
| Benign prostate hyperplasia | 731 ± 500 | [92] |
| β-thalassemia major | 66000 ± 9600 | [43] |
| Chronic pancreatitis | 2368 ± 2431 | [93] |
| Colorectal carcinoma | 783 ± 491 | [94] |
| SARS-CoV-2 in ICU | 12400 | [95] |
| Endometrial cancer | 1077 | [96] |
| Exercise | 200 -1000 | [32] |
| Heart failure | 2705 | [97] |
| Hereditary hemochromatosis | 720 ± 50 | [43] |
| Mitochondrial myopathy | 2711 ± 2459 | [98] |
| Ovarian cancer | 96,1 - 1876 | [98] |
| Pancreatic cancer | 5388 ± 3720 | [93] |
| Preeclampsia | 421 ± 187 | [99] |
| Pregnancy | 6300-15300 | [28] |
| Prostate cancer | 860 ± 850 | [92] |
| Renal cell carcinoma | 1100 ± 150 | [100] |
| Sepsis | 4900 | [51] |
| Sickle cell anemia | 880 ± 160 | [43] |
| Smoking | 1835 | [47] |
| Stable coronary heart disease | 915 - 1827 | [101] |
| Systemic sclerosis | 1367 | [47] |
GDF-15: growth differentiation factor 15; ICU: intensive care unit
Table 1.
GDF-15 effects in fibrosis mice models.
| Setting | Intervention | Finding | Reference |
|---|---|---|---|
| CCl4-induced liver fibrosis | CRISPR-Cas GDF-15 KO | Histopathology (H&E and Sirius red staining): increased fibrosis. Immunohistochemistry: increased macrophage F4/80 and neutrophil MPO infiltration, upregulated Ly6Chi Serum: increased hepatic enzymes |
[128] |
| DDC-induced liver fibrosis | CRISPR-Cas GDF-15 KO | Histopathology (H&E and Sirius red staining): increased fibrosis and collagen deposition | |
| CCl4-induced liver fibrosis | AAV8 gene vector GDF-15 overexpression | Histopathology (H&E and Sirius red staining): reduced liver injury and fibrosis. Serum: reduced hepatic enzyme levels in blood. mRNA levels (RT-PCR): reduced IL-1β, TNF-α, and NOS2. Increased YM1, Arg1 and CD206 |
|
| DDC-induced liver fibrosis | CRISPR-Cas GDF-15 KO | Histopathology (H&E and Sirius red staining): reduced liver injury and fibrosis | |
| CCl4-induced liver fibrosis | Tail vein infusion with GDF15-preprogrammed macrophages 24 h | Histopathology (H&E and Sirius red staining): reduced liver injury and fibrosis.Serum: reduced hepatic enzyme levels | |
| Hyperoxia 95% after birth | GDF15-/- mice | Higher mortality and lower body weight. Immunofluorescence for von Willebrand factor: impaired alveolarization and lung vascular development, lower macrophage F4/80 infiltration |
[129] |
| Ethanol-induced liver disease | Genetic ablation of hepatocyte-derivedGDF-15 | Annexin V apoptosis assay: decrease in Kupffer cell apoptosis in liver perivenous region | [130] |
| Ethanol-induced liver disease | GDF15 KO | Histopathology (H&E and Oil Red O staining): increased hepatic fat accumulation. Serum: increased hepatic enzymes and triglyceride. Increased TNF-α and IL-6 |
[127] |
| CCl4-induced liver fibrosis | GDF15 KO | Histopathology (H&E and Oil Red O staining): increased hepatic fat accumulation, Serum: increased hepatic enzymes and triglyceride. Increased TNF-α and IL-6 |
|
| CCl4-induced liver fibrosis GDF15 KO | rGDF-15 0,5 mg/kg i.v. | Histopathology (H&E and Oil Red O staining): reduced collagen accumulation. Western blot: inhibition of NF-κB, JNK, and p38 signaling pathways |
|
| Coronary artery ligation-induced myocardial infarction | Allogenic cardiac progenitor cells transplant with GDF-15 KD | Flow cytometry of cell suspension: Decrease in M2 phenotype and Treg activation | [131] |
| Sterile muscle injury with cardiotoxin injection | rGDF-15 i.m. | Flow cytometry: decrease CD45+ muscle infiltration. Increased MCHII expression by monocyte-derived macrophages, anti-inflammatory phenotype | [60] |
DDC: 3,5-Diethoxycarbonyl-1,4-Dihydrocollidine; CCl4: carbon tetrachloride; GDF-15: growth differentiation factor 15; H&E: Hematoxylin-eosin, i.v.: intravenous; i.m.: intramuscular; KO: knock-out; KD: knock-down; NF-κB: nuclear factor kappa-light-chain enhancer of activated B cells; NOS2: nitric oxide synthase 2; TNF-α: tumor necrosis factor α.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



