
Submitted:
27 May 2024
Posted:
28 May 2024
You are already at the latest version
Abstract
The maturation process of Arabica coffee fruits is influenced by plant endogenous and external factors. The maturation stage of the fruits affects the bean's chemical composition, which is related to coffee beverage quality. During fruit maturation, the colour of the peel changes from green to red (cherry), which indicates the harvest time and supposedly that the fruits have high quality. However, many tones of red may define fruit maturity. This work aimed to study the dynamics of soluble sugar accumulation related to fruit redness in coffee fruit tissues. We classified the ripe fruits into six ripeness classes based on the intensity of the red colour of the epicarp using a colourimeter. Total soluble sugar, sucrose and starch were analyzed in the coat (exocarp+mesocarp), coat juice (obtained by squeezing the coat), and beans. We showed that endosperm sugar dynamics do not follow changes in the coat, indicating a separate control of sugar accumulation, mainly related to sucrose, an essential component of coffee quality. Our data show that there is no transfer of sucrose and reducing sugars from the red coat to the bean.
Keywords:
coffee quality
; sucrose
; Coffea arabica
1. Introduction
The maturation process of Arabica coffee fruits (Coffea arabica L.) is influenced by factors related to the plant genetic background, edaphoclimatic conditions and their interaction [1]. Coffee variety, age and plant physiology strongly influence bean quality. For instance, different varieties present different cycle lengths, ranging from early to late maturation. The position of the fruits in the plant canopy also affects maturation, as the source-sink balance is different among fruits, and they experience light incidence and temperature differently according to their position [2]. Climate, altitude, and their regulatory role over the mean air temperature affect the maturation rate and the plant demand for water, which also relates to quality [3].
Immature and overripe beans are directly related to lower coffee quality as their physicochemical properties and chemical composition are significantly different from the cherry fruits [4,5]. Their presence in a coffee lot is a defect in the international coffee evaluation system. Immature coffee beans are lighter, more acidic and have lower sucrose, oil, and protein concentrations than mature beans [4]. Regarding the overripe fruits, they are exposed to microorganisms attack, which favours the development of undesirable products from the microbial metabolism [5]. Therefore, maximizing the collection of ripe fruits and minimizing immature and overripe fruits is a key factor for coffee beverage quality [1,4].
The chemical composition of coffee beans is related to beverage aroma, flavour, sourness, and sweetness. These sensorial parameters result from a complex process of post-harvest and roasting reactions [6]. Of the several compounds in coffee beans, sucrose is a precursor of coffee aroma and flavour [7,8].
C. arabica fruits have a bimodal curve growth [9]. In the first growth phase, the endosperm is completely formed, and the fruit is at full size. In the second phase occurs the maturation. Contrary to proteins, alkaloids caffeine and trigonelline, and most of the chlorogenic acids found in the endosperm, which reach the highest concentration about the first fruit growth phase [10], sucrose continues to accumulate until the end of maturation [9,11]. Sucrose accumulates in the endosperm also in a bimodal curve [9,12], reaching about 8% in the endosperm of mature fruits [13,14]. Such an increase during maturation is probably supported by sucrose imported from the leaves, and it is uncertain how much epicarp photosynthesis contributes to such sucrose increase [9]. During bean roasting, sucrose reacts with amino acids, peptides, and proteins in the Maillard reaction, forming new compounds. The Maillard reaction is responsible for developing coffee organoleptic attributes (Kerler et al., 2010; Mazzafera et al., 2019). Thus, bean sucrose concentration of great importance for final coffee quality, and it is supposed that its highest content is found in mature fruits, which, according to the coffee farmer, is defined by the fruit colour.
During maturation, the fruit metabolism changes markedly, and the colour changes in the fruit peel (epicarp), from green to red or yellow according to the cultivar. Coffee maturation can be divided into five classes based on the colour of the fruit peel: green (immature fruits), yellowish (semimature fruits), cherry (ripe fruits), dark red or dark yellow (overripe fruits), and black (dry fruits) [15,16]. In red fruit varieties, cherry fruits should be entirely red when they are physiologically mature and supposedly at the highest quality point. Therefore, colour is intimately linked to coffee quality, indicating the maturation stage and proper harvest timing for farmers.
Here, we investigated the dynamics of soluble sugar accumulation related to fruit redness in coffee fruit tissues. We classified the ripe fruits into six ripeness classes based on the intensity of the red colour of the pericarp using a colourimeter. Total soluble sugar, sucrose and starch were analyzed in the pericarp, pericarp juice, and beans. We showed that changes in the coat (exocarp + mesocarp) are not followed by endosperm, indicating a separate control of sugar accumulation, mainly related to sucrose, an essential component of coffee quality.
2. Results
The fruits were separated in six classes of red, according the colourimeter measurements (Figure 1). The ripeness stages were divided arbitrarily into six groups, based on the upper and lower boundary of each class from light to dark red: Stage 1 (a* = +23.0 to +21.0), Stage 2 (a* = +20.9 to +19.0), Stage 3 (a* = +18.9 to +17.0), Stage 4 (a* = +16.9 to +15.0), Stage 5 (a* = +14.9 to +12.0) and Stage 6 (a* = +11.9 to +8.0).
The anthocyanin on the coffee coat increased up to the fifth stage, implying an augment in the red colour of the fruits (Figure 2).
The total soluble sugars (TSS) increased in the coat, juice and beans as the maturation occurred (Figure 3). TSS in the coat and juice presented a linear tendency of increase, while in the beans, this was less evident because of the considerable variation among the samples. Mean TSS concentration was higher in the coat, followed by juice, and the beans had the lowest values.
3. Discussion
The division we adopted to separate distinct classes of red was efficient (Figure 1b). The darkening of the red colour in the fruit coat from stage one to six results from the increase in anthocyanin (Figure 2). There were visual similarities between the nearer ripeness stages (i.e., 1 and 2 or 2 and 3), but the differences between every other stage were distinguishable (Figure 1b). This implies that if redness is related to the ideal stage for harvesting, there is a variation that, in principle, might be used to separate high-quality beans, which maximizes coffee cup quality.
Previous work also used a colourimeter to separate beans at different developmental stages [21]. Unfortunately, among fruits at four developmental stages, only two were red, and all metabolomic and proteomic analyses were carried out with whole fruits and not separated tissues.
Coffee fruits increase in volume and dry mass during maturation, which happens in the second growth phase of the bimodal growth curve of coffee beans. There is an increase in the growth rate of the pericarp. Meanwhile, the beans cease growth and reach their maximum size and dry mass in the first growth phase [9,22]. Sucrose concentrations increase all over the fruit growth, contrary to other compounds like caffeine, trigonelline, proteins, and chlorogenic acids that reach their maximum concentration at the first growth phase [10,23], also known as the storage phase [24]. The transcriptomic analysis also showed that most genes related to seed reserves had been expressed in the transition of green (with formed endosperm) to yellow (beginning of maturation) fruits, with 2058 differentially expressed transcripts (down and upregulated). The comparison between the red (mature) and yellow stages showed only 130 differentially expressed genes [25].
Such an increase was investigated by feeding coffee fruits or leaves in branches bearing fruits with 14CO2. 14C-sucrose, the product of the fruit and leaves photosynthesis, was high in the coat of fruits where 14CO2 was supplied to leaves [9]. But, about 30-40% of the total radioactivity found in the fruit was in the endosperm and perisperm. Thus, sucrose from leaves was imported into any part of the fruit. When the fruits were incubated with 14CO2, most radioactivity was found in the perisperm, suggesting this is a passage tissue. Indeed, the perisperm is linked to the pericarp by a sort of bridge formed by a few cells [9]. However, the endosperm is not connected to any other fruit tissue, suggesting that molecules transported from the leaf or the coat photosynthesis to the endosperm follow an apoplastic pathway [9].
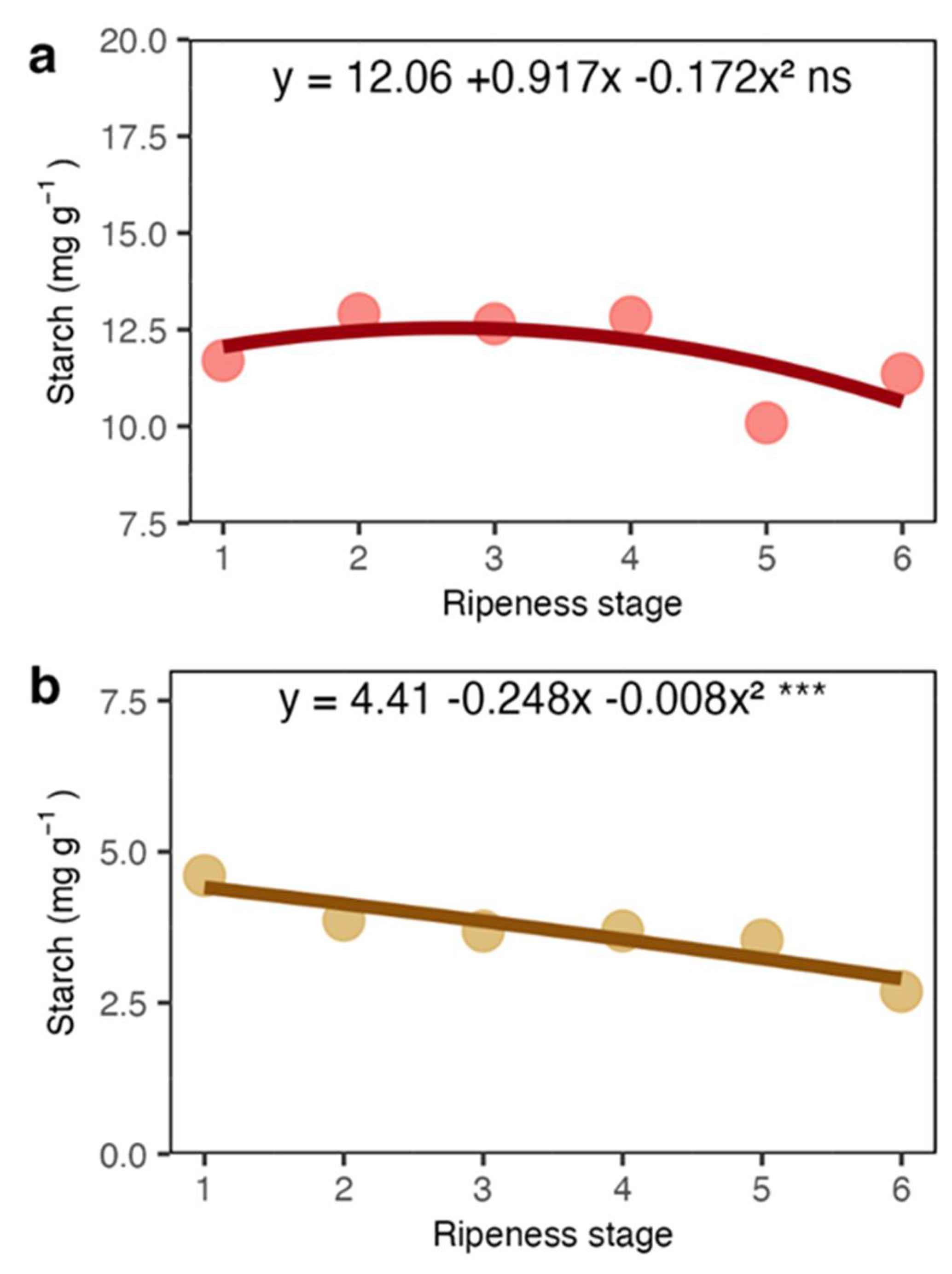
Our results showed that total soluble sugars and sucrose showed a sharp increase in the coat and juice (the liquid obtained from squeezed coats). The increase in sucrose may be related to the high respiration occurring in fruits during maturation [26] and sucrose is probably imported from leaves [9]. However, most of the total sugars were represented by reducing sugars (Figures 3a and 5a), which may have appeared from the pectin degradation in mesocarp. The sugars detected in the juice were certainly a rich fraction of these sugars. The coffee pericarp is rich in pectin [27], and at maturation, there is an increase in enzyme activity related to pectin degradation, such as pectinase and pectin lyase [28,29]. Increases in the reducing sugars arabinose and galactose concentration, among other sugars in the fruit accompany this process. Because of the low amount of starch in the coat and endosperm (Figures 6a and 6b), its contribution seems very limited.
It is well known that while sucrose increases in the endosperm, reducing sugars like fructose and glucose are reduced [9,13,30]. Here, both sucrose (Figure 4c) and reducing sugars (Figure 5c) in the endosperm were stable. Thus, although a marked increase of soluble sugars in the coat, which includes sucrose and reducing sugars, they seem not transferred to the endosperm. We suggest that the endocarp, which is mainly composed of cellulose (40-49%), hemicellulose (25-32%) and lignin (33-35%) and relatively impermeable [31], may work as a barrier for the transport of sugars from the coat to the endosperm. Due to its lignocellulosic composition, the endocarp (parchment in dry seeds) has a low hygroscopic nature, offering water transfer resistance of parchment [32].
Although sucrose increase was not so pronounced in the endosperm in the six red classes we separated here, it would be worth testing the beverage quality to verify if such a discrete increase may affect quality.
Identifying the optimal ripeness stage is fundamental to defining harvest timing and increasing coffee quality. However, depending on the size of the coffee farm, harvesting may become a logistic problem if maturation occurs homogeneously on the whole farm. Thus, variety selection aiming at different maturity groups to scale the harvest into a more controlled process would optimize the process to avoid green fruits. Moreover, remote sensing and colour sorting machines could help identify the coffee crop's ideal ripeness stage and sort the fruits by colour after harvest, producing more uniform and high-quality coffee lots. Ultimately, this would result in better products offered to the market and increase the profitability and sustainability of coffee crops.
4. Materials and Methods
4.1. Experimental Design and Fruit Harvest
The samples were collected from coffee trees ("Red Obatã" variety) in the 2017/2018 season at the experimental area of "Luiz de Queiroz" College of Agriculture – Esalq/Usp at Piracicaba-SP. Fruits were manually harvested from five six-year-old coffee trees. The branches collected were in the mid-third of the plants. In the lab, the fruits were visually classified into green (unripe fruits), yellowish (moderately ripe), cherry (completely red fruits) and overripe fruits. As indicated by Figure 1, most of the fruits were at the red stage, and only these fruits were used for analysis further.
4.2. Fruit Classification by Colour
Two hundred red fruits were selected randomly from each plant for colour classification. The colour was measured by placing each fruit on a Minolta CR300 colourimeter laterally, which measures colour using the CIELAB pattern (International Commission on Illumination). In this system, colour is classified by the L*, a* and b* coordinates in a three-dimensional space, where the first axis, L*, is the lightness of the colour and varies from 0 (black) to 100 (white), while the other two axes, a* and b*, indicate the relative coordinate of the colour among red, green, blue, and yellow. The a* coordinate is the position between red (positive values) and green (negative values), and the b* coordinate is the position between blue (positive values) and yellow (negative values). All three parameters were obtained, but only the a* parameter (coordinate red/green) was used to classify the fruits into six ripeness stages based on the red colour.
4.3. Sample Preparation and Analysis
Each fruit's coat (epicarp and mesocarp) was manually removed and then squeezed into tubes maintained on ice to extract the juice from the mesocarp. We estimated the volume of the juice in a 10 ml cylinder, quickly centrifuged at 4oC, and then immediately froze the supernatant at -20oC. The coat was also frozen. The beans were rubbed in a cotton tissue to remove the adherent mucilage, and the hard parchment (endocarp) was manually removed and discarded. The beans were frozen, too. The frozen coat and beans were lyophilized and finally grounded in a mortar with a pestle.
We determined total soluble sugars, sucrose, reducing sugars (subtracting sucrose from total soluble sugars) and starch content in all samples, except for the juice where starch was not determined. Coat and beans were extracted with 70% ethanol (100 mg / 1 ml of) at 80°C for 10 min. The samples were centrifuged at 10,000 xg for 10 min, the supernatant was collected, and the precipitate was re-extracted five more times in 70% ethanol. The six extractions were combined. The remaining residue was used for starch determination, using 1 ml of 30% perchloric acid [17]. The samples were left resting overnight on the bench, and the supernatant was recovered by centrifugation. The juice was used for analysis without any previous extraction. Total soluble sugar was determined using the phenol sulfuric method, and sucrose was used as standard [18]. Sucrose was determined in the extracts after the hydrolysis of the reducing sugars with a 10% KOH incubation at 100oC, as proposed by Van Handel (1968). The reducing sugar concentration was obtained by the difference between total soluble sugar and sucrose determinations. Anthocyanin was also estimated in the coat [20]. Briefly, extraction was done by adding 1 ml of a solution containing 6 M HCl, water and methanol (7:13:70, v/v/v) to 100 mg grounded samples, followed by incubation at 4°C for 24h in the dark. Next, the samples were centrifuged at 14,000 rpm for 10 min, and the supernatant was collected and analyzed in a spectrophotometer at two wavelengths (530 and 653 nm). The anthocyanin was calculated by the formula C=A530-(0.24*A653), where C is the anthocyanin concentration in absorbance units; A530 is the absorbance at 530 nm; and A653 is the absorbance at 653 nm.
4.4. Data Analysis
All data analysis was done using R software (version 4.2.2), and all plots were made using the ggplot2 package. We used linear mixed models (package lme4), setting the coffee ripeness stage as the fixed effect and the plants (replicate) as random factors for all variables. Analysis of variance was done using the car package.
Author Contributions
Conceptualization, P.M.; methodology, J.L.C.B., G.A. and M.S.S.; formal analysis, J.L.C.B., G.A. and M.S.S.; resources, P.M.; writing, J.L.C.B. and P.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are available under request to the corresponding author.
Acknowledgments
The authors thank Fundação de Amparo a Pesquisa do Estado de São Paulo (FAPESP) for MSc fellowships to J.L.C.B. and M.S.S. and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for a research fellowship to PM.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Leroy, T.; Ribeyre, F.; Bertrand, B.; Charmetant, P.; Dufour, M.; Montagnon, C.; Marraccini, P.; Pot, D. Genetics of Coffee Quality. Braz J Plant Physiol 2006, 18, 229–242. [Google Scholar] [CrossRef]
- Vaast, P.; Bertrand, B.; Perriot, J.J.; Guyot, B.; Génard, M. Fruit Thinning and Shade Improve Bean Characteristics and Beverage Quality of Coffee (Coffea Arabica L.) under Optimal Conditions. J Sci Food Agric 2006, 86, 197–204. [Google Scholar] [CrossRef]
- Ribeiro, D.E.; Borem, F.M.; Cirillo, M.A.; Vilela, M.B.P.; Ferraz, V.P.; Alves, H.M.R.; Taveira, J.H. da S. Interaction of Genotype, Environment and Processing in the Chemical Composition Expression and Sensorial Quality of Arabica Coffee. Afr J Agric Res 2016, 11, 2412–2422. [Google Scholar] [CrossRef]
- Mazzafera, P. Chemical Composition of Defective Coffee Beans. Food Chem 1999, 64, 547–554. [Google Scholar] [CrossRef]
- Osorio Pérez, V.; Matallana Pérez, L.G.; Fernandez-Alduenda, M.R.; Alvarez Barreto, C.I.; Gallego Agudelo, C.P.; Montoya Restrepo, E.C. Chemical Composition and Sensory Quality of Coffee Fruits at Different Stages of Maturity. Agronomy 2023, 13, 341. [Google Scholar] [CrossRef]
- Borém, F.M.; Figueiredo, L.P.; Ribeiro, F.C.; Taveira, J.H.S.; Giomo, G.S.; Salva, T.J.G. The Relationship between Organic Acids, Sucrose and the Quality of Specialty Coffees. Afr J Agric Res 2016, 11, 709–717. [Google Scholar] [CrossRef]
- Kerler, J.; Winkel, C.; Davidek, T.; Blank, I. Basic Chemistry and Process Conditions for Reaction FLavours with Particular Focus on Maillard-Type Reactions. In Food Flavour Technology; Taylor, A.J., Linforth, R.S.T., Eds.; Blackwell, 2010; pp. 51–88. ISBN 9781405185431. [Google Scholar]
- Mazzafera, P.; Schimpl, F.; Kiyota, E. Proteins of Coffee Beans: Recent. In Coffee: production, quality and chemistry; Farah, A., Ed.; The Royal Society of Chemistry: London, 2019; pp. 431–444. [Google Scholar]
- Geromel, C.; Ferreira, L.P.; Guerreiro, S.M.C.; Cavalari, A.A.; Pot, D.; Pereira, L.F.P.; Leroy, T.; Vieira, L.G.E.; Mazzafera, P.; Marraccini, P. Biochemical and Genomic Analysis of Sucrose Metabolism during Coffee (Coffea Arabica) Fruit Development. J Exp Bot 2006, 57. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N.; Kazi, T. The Influence of Coffee Bean Maturity on the Content of Chlorogenic Acids, Caffeine and Trigonelline. Food Chem 1987, 26, 59–69. [Google Scholar] [CrossRef]
- Geromel, C.; Ferreira, L.P.; Davrieux, F.; Guyot, B.; Ribeyre, F.; Brígida dos Santos Scholz, M.; Protasio Pereira, L.F.; Vaast, P.; Pot, D.; Leroy, T.; et al. Effects of Shade on the Development and Sugar Metabolism of Coffee (Coffea Arabica L.) Fruits. Plant Physiology and Biochemistry 2008, 46. [Google Scholar] [CrossRef]
- Cannell, M.G.R. Physiology of the Coffee Crop. <italic>Coffee: Botany, Biochemistry and Production of Beans and Beverage</italic> 1985, 108–134.
- Trugo, L.C. Carbohydrates. In Coffee: VOlume 1 - Chemistry; Clarke, R.J., Macrae, R., Eds.; Springer Netherlands: Dordrecht, 1985; ISBN 978-94-010-8693-6. [Google Scholar]
- Rogers, W.J.; Michaux, S.; Bastin, M.; Bucheli, P. Changes to the Content of Sugars, Sugar Alcohols, Myo-Inositol, Carboxylic Acids and Inorganic Anions in Developing Grains from Different Varieties of Robusta (Coffea Canephora) and Arabica (C. Arabica) Coffees. Plant Science 1999, 149, 115–123. [Google Scholar] [CrossRef]
- Morais, H.; Caramori, P.H.; Koguishi, M.S.; Ribeiro, A.M. de A. Escala Fenológica Da Fase Reprodutiva de Coffea Arabica. Bragantia 2008, 67, 257–260. [Google Scholar] [CrossRef]
- Pezzopane, J.R.M.; Pedro Júnior, M.J.; Thomaziello, R.A.; Camargo, M.B.P. de Coffee Phenological Stages Evaluation Scale. Bragantia 2003, 62, 499–505. [Google Scholar] [CrossRef]
- Cocking, E.C.; Yemm, E.W. Estimation of Amino Acids by Ninhydrin. Biochem J 1954, 58, xii. [Google Scholar]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal Chem 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Van Handel, E. Direct Microdetermination of Sucrose. Anal Biochem 1968, 22, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Hughes, N.M.; Neufeld, H.S.; Burkey, K.O. Functional Role of Anthocyanins in High-light Winter Leaves of the Evergreen Herb Galax Urceolata. New Phytologist 2005, 168, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Zhou, B.; Zheng, T.; Zhao, C.; Shen, X.; Wang, X.; Qiu, M.; Fan, J. Integrating Metabolomics and Proteomics Technologies Provides Insights into the Flavor Precursor Changes at Different Maturity Stages of Arabica Coffee Cherries. Foods 2023, 12, 1432. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.G.R. Physiology of the Coffee Crop. Coffee 1985, 108–134. [Google Scholar] [CrossRef]
- Rogers, W.J.; Bézard, G.; Deshayes, A.; Meyer, I.; Pétiard, V.; Marraccini, P. Biochemical and Molecular Characterization and Expression of the 11S-Type Storage Protein from Coffea Arabica Endosperm. Plant Physiology and Biochemistry 1999, 37, 261–272. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, C.; Zhang, Y.; Zeng, W.; Cesarino, I. Coffee Cell Walls—Composition, Influence on Cup Quality and Opportunities for Coffee Improvements. Food Quality and Safety 2021, 5. [Google Scholar] [CrossRef]
- Cheng, B.; Furtado, A.; Henry, R.J. The Coffee Bean Transcriptome Explains the Accumulation of the Major Bean Components through Ripening. Sci Rep 2018, 8, 11414. [Google Scholar] [CrossRef] [PubMed]
- Cannell, M.G.R. Changes in the Respiration and Growth Rates of Developing Fruits of Coffea Arabica L. Journal of Horticultural Science 1971, 46, 263–272. [Google Scholar] [CrossRef]
- Avallone, S.; Guiraud, J.P.; Guyot, B.; Olguin, E.; Brillouet, J.M. Polysaccharide Constituents of Coffee-Bean Mucilage. J Food Sci 2000, 65, 1308–1311. [Google Scholar] [CrossRef]
- Cheng, B.; Furtado, A.; Henry, R.J. The Coffee Bean Transcriptome Explains the Accumulation of the Major Bean Components through Ripening. Sci Rep 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Cação, S.M.B.; Leite, T.F.; Budzinski, I.G.F.; dos Santos, T.B.; Scholz, M.B.S.; Carpentieri-Pipolo, V.; Domingues, D.S.; Vieira, L.G.E.; Pereira, L.F.P. Gene Expression and Enzymatic Activity of Pectin Methylesterase during Fruit Development and Ripening in Coffea Arabica L. Genetics and Molecular Research 2012, 11, 3186–3197. [Google Scholar] [CrossRef] [PubMed]
- Redgwell, R.; Fischer, M. Coffee Carbohydrates. Brazilian Journal of Plant Physiology 2006, 18, 165–174. [Google Scholar] [CrossRef]
- Munõz, M.; Roa, M.; Correa, R. Endocarp Analysis of a Traditional Variety of Colombian Coffee. Coffee Sci 2019, 14, 206. [Google Scholar] [CrossRef]
- Ramírez-Martínez, A.; Salgado-Cervantes, M.A.; Rodríguez-Jimenes, G.C.; García-Alvarado, M.A.; Cherblanc, F.; Bénet, J.C. Water Transport in Parchment and Endosperm of Coffee Bean. J Food Eng 2013, 114, 375–383. [Google Scholar] [CrossRef]
Figure 1.
Percentage of fruits in four developmental stages (green, yellow, cherry and overripe) in samples collected from five C. arabica plants (A) and distribution of the cherry fruits in six ripeness stages according to their redness intensity (B).
Figure 1.
Percentage of fruits in four developmental stages (green, yellow, cherry and overripe) in samples collected from five C. arabica plants (A) and distribution of the cherry fruits in six ripeness stages according to their redness intensity (B).

Figure 2.
Anthocyanin concentration in arabica coffee coat according to red ripeness classes.
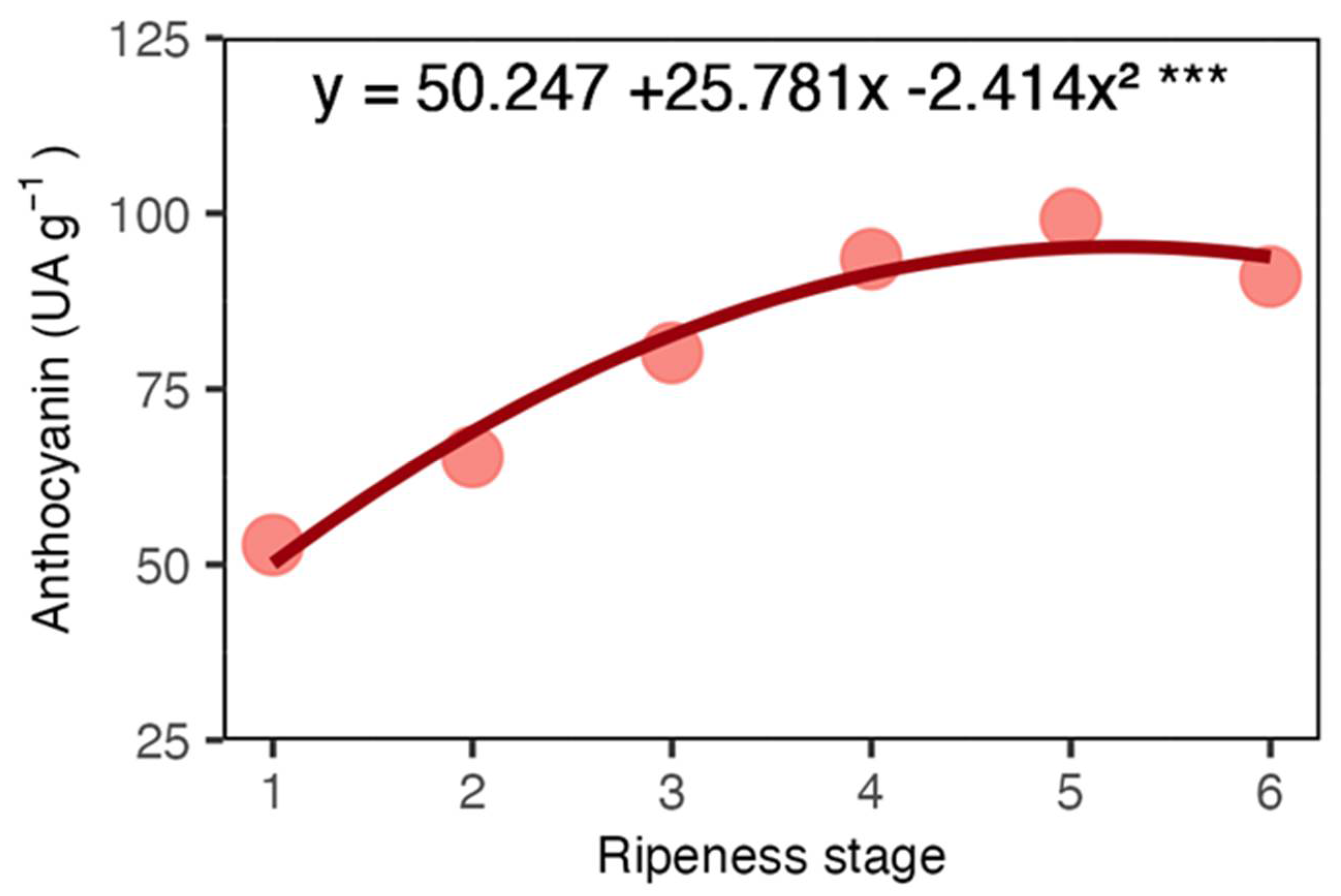
Figure 3.
Total soluble sugar (TSS) concentration in the coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
Figure 3.
Total soluble sugar (TSS) concentration in the coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
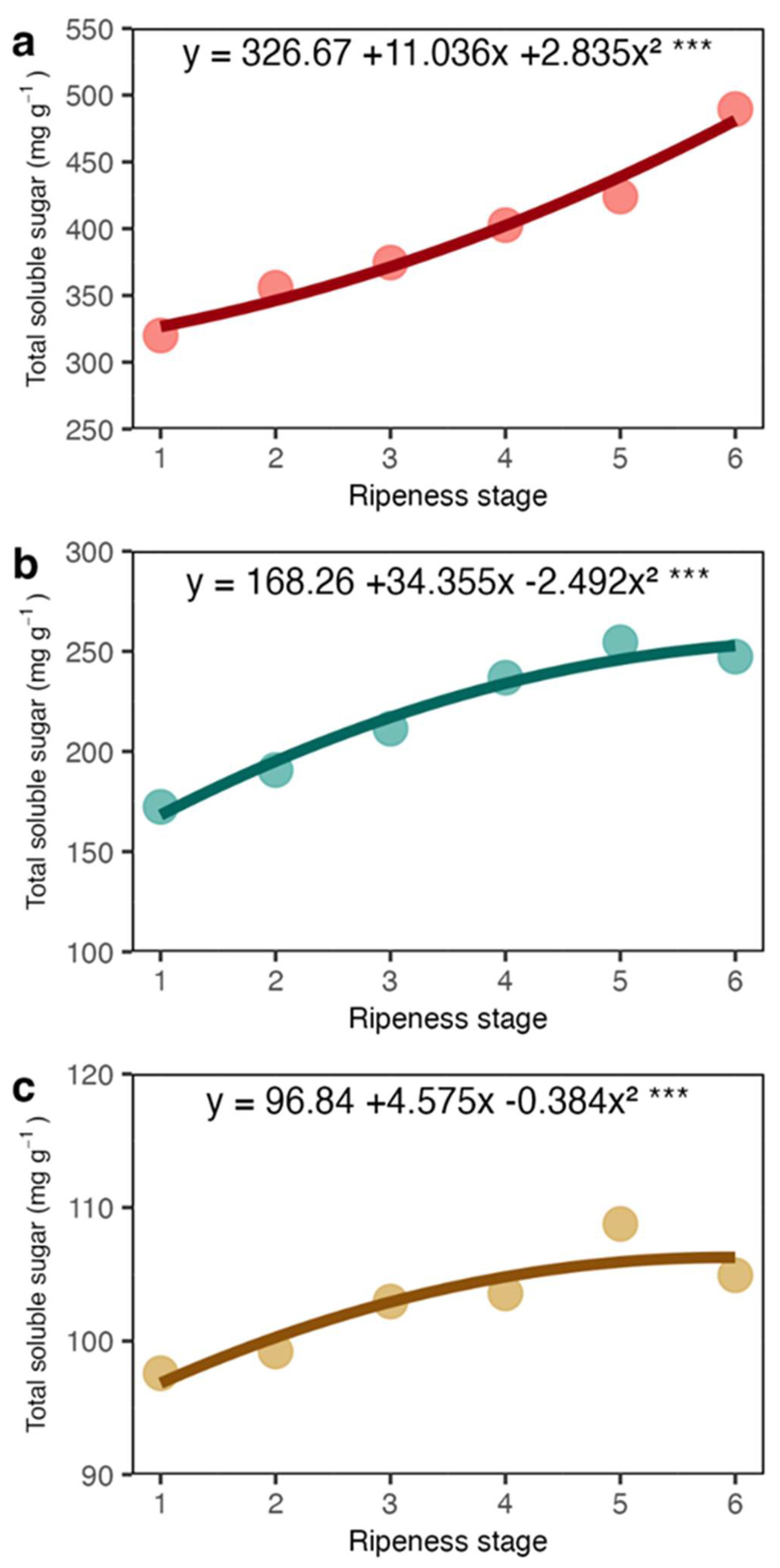
Figure 4.
Sucrose concentration in coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
Figure 4.
Sucrose concentration in coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
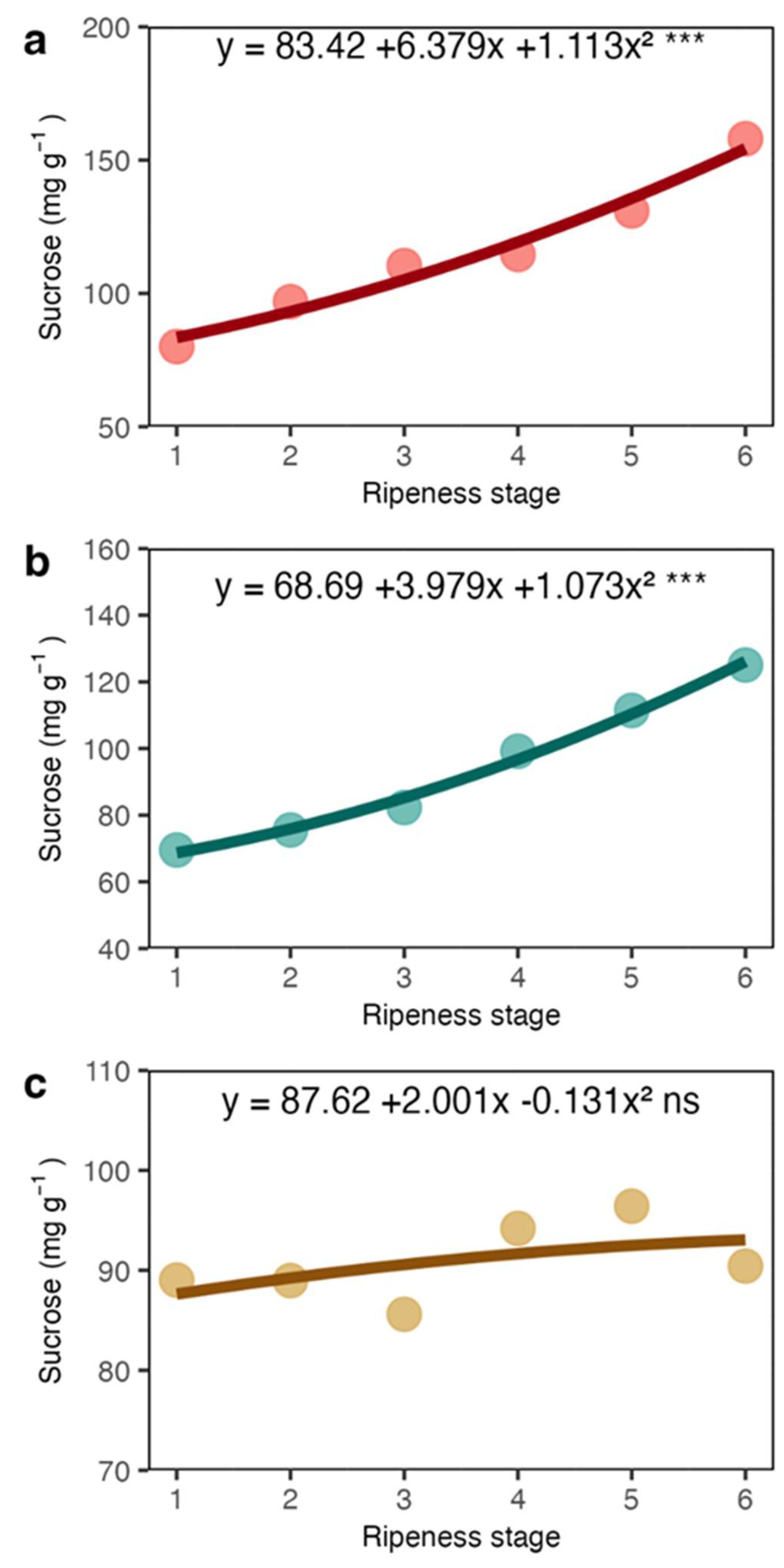
Figure 5.
Reducing sugar concentration in the coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
Figure 5.
Reducing sugar concentration in the coat (a), juice (b) and beans (c) of arabica coffee according to the red ripeness classes.
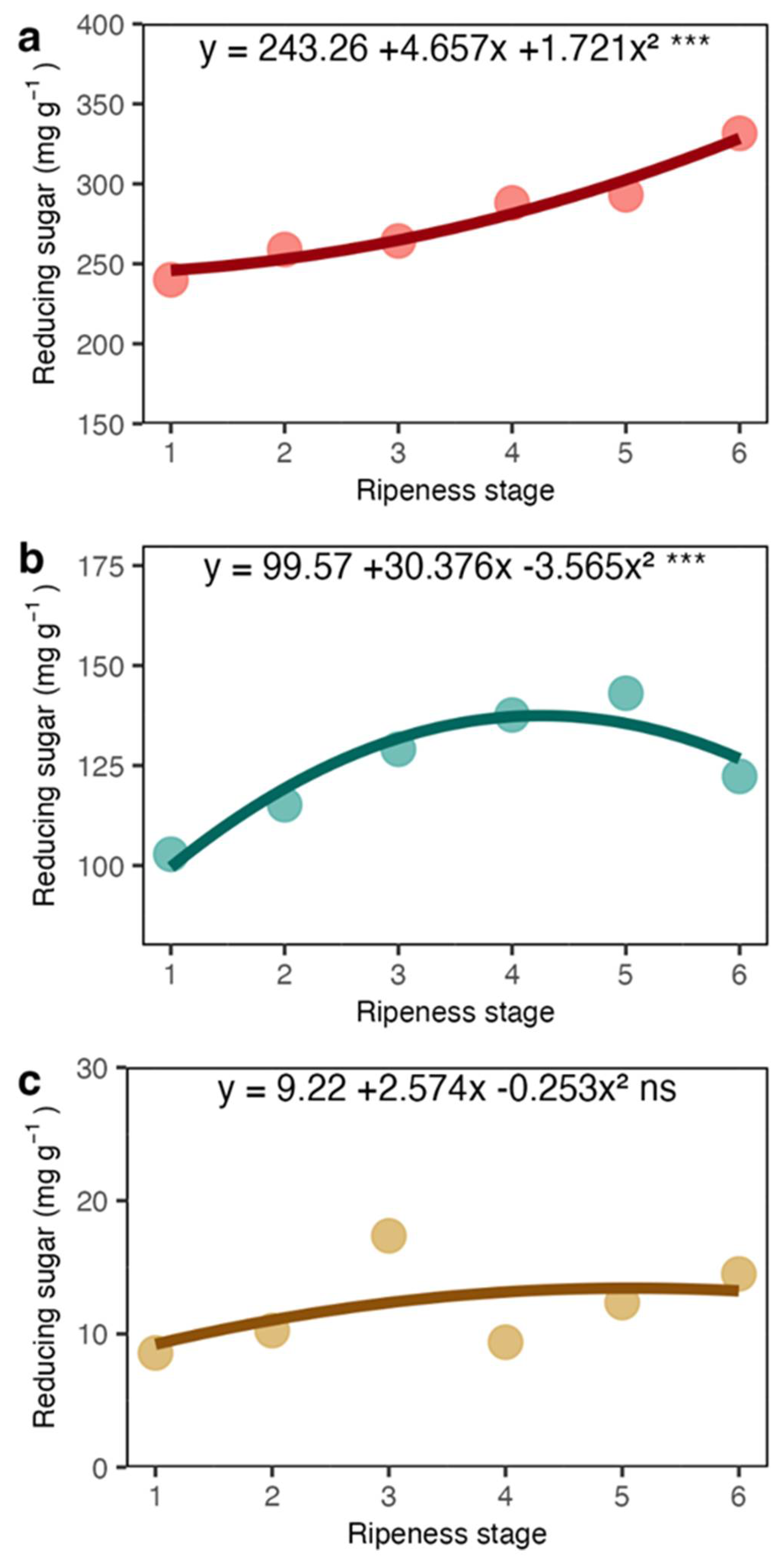
Figure 6.
Starch concentration in coat (a) and beans (b) of arabica coffee according to the red ripeness classes.
Figure 6.
Starch concentration in coat (a) and beans (b) of arabica coffee according to the red ripeness classes.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



