
Submitted:
27 May 2024
Posted:
28 May 2024
You are already at the latest version
Abstract
The conversion of native habitats into anthropogenic ones compromises the original composition and configuration of the landscapes, influencing ecological dynamics and affecting biodiversity. Increasingly, landscape ecology has shown that these effects can only be understood if accessed at adequate spatial scales, as the scale which landscape structure is evaluated influences species responses. Here, we investigated how three variables of landscape composition (proportion of forest, coffee crop and pasture) and two of configuration (number of fragments and mean nearest neighbor distance) interfere in the richness and composition of medium and large-sized mammals considering a multiscale approach. We recorded medium and large-sized mammal species in 13 landscapes with predominantly matrices of coffee and pasture in Minas Gerais, Brazil. Then, we built distance-based linear models to identify the scale of effect of each landscape variable for both response variables considering eight scales (from 250 m to 2000 m). Finally, we verified the influence of the landscape on the richness and composition of mammals, considering the landscape variables in their respective scales of effect. We found 67% of the probable species occurring in the region. The scales of effect varied among landscape variables probably due to the different variables affect different aspects of organisms' ecological requirements. The proportion of pasture in the landscape explained the variation in species richness, while the proportion of forest explained the variation in species composition. In addition, the proportion of pasture in the landscape had a positive influence on species richness, indicating that this matrix may favor the presence of generalist species of habitat and/or this result is due to the higher concentration of species in the fragments immersed in this matrix. These results suggest that considering different responses to biodiversity is important to understand different aspects of the landscape's influence on biodiversity. In addition, the composition of the landscape is fundamental for the perpetuation of species and, therefore, both forest cover and types of matrices in the landscape must be considered to improve species conservation strategies. Finally, generalizing a spatial scale can lead to misinterpretations about the influence of the landscape on biodiversity.
Keywords:
matrix
; habitat loss
; habitat fragmentation
; scale of effect
; landscape ecology
1. Introduction
A large part of terrestrial ecosystems is impacted due to the conversion of natural habitats into anthropic habitats [1]. The conversion of natural habitats into anthropogenic ones changes the composition (e.g., types and proportions of land uses) and configuration (e.g., spatial arrangement of landscape elements) of the landscape, causing the loss of native habitats and fragmentation of the remnants [2]. Thus, habitat loss and fragmentation are the main causes of defaunation, including vertebrates of larger body size such as medium and large-sized mammals [3,4,5,6].
Habitat amount is commonly evaluated to explain the species richness in forest fragments [7,8,9]. However, the matrix around the forest remnants also influences the occurrence of species and ecological dynamics [10,11,12,13]. The effects that the matrix can have on species are mainly related to species movement and dispersion, availability of resources and changes in abiotic characteristics. Therefore, the magnitude of the impact that the types and quantities of matrices in the landscape exert on populations depends on characteristics that influence the life history of the species (e.g., ability to move, diet, habitat requirement) [14]. In this way, anthropogenic matrices can have ambivalent effects on the impacts of habitat loss and habitat fragmentation on biodiversity.
Habitat fragmentation per se changes the original landscape configuration, increasing the number of forest fragments of different sizes and shapes, dispersed in the landscape and under greater edge effect [15,16]. The effects of fragmentation on biodiversity have been discussed [12,16,17,18,19] and most of the effects are considered as negative [18,20,21], but also positive or neutral [16,17]. However, many studies evaluated the effect of habitat fragmentation without distinguishing it from the effect of habitat loss and without considering an appropriate scale [17]. In this way, fragmentation must be accessed from metrics related to landscape configuration since it is characterized by the division of habitat, independent of habitat loss [15,17].
One of the challenges of studies at the landscape level is to define the best scale at which the landscape composition and configuration variables should be collected [22,23,24]. This is not a trivial question, as species’ responses to landscape variables are shaped by ecological processes - such as metapopulation dynamics, dispersion, inter and intra-specific interactions - that act at different scales [16,24,25,26]. A solution to this challenge is to define the scale of effect of each landscape variable, that is, the scale in which the landscape variable has the greatest effect under the evaluated response variable and consider the multiscale approach, which each variable is evaluated in their respective effect scale [22,24].
The Atlantic Forest and the Cerrado are Brazilian domains with high species endemism and are highly threatened due to, mainly, conversion of habitat to agriculture, livestock and urbanization and, therefore, they are considered biodiversity hotspots [27,28,29,30]. The forest cover of the contemporary Atlantic Forest is restricted to only 12% of its original cover, with approximately 84% of the remainder represented by forest fragments smaller than 50 hectares [31]. While approximately 50% of the native vegetation cover in the Cerrado has been transformed into agriculture and pasture [30]. Consequently, the biological populations present in the forest remnants are under the influence of the landscape characteristics to persist in these fragmented landscapes.
Our main goal was to compare how landscape configuration and composition affect medium and large-sized mammal communities in agricultural ecosystems. We also evaluated the scale of effect of each landscape variable for both mammal richness and composition, and we compared the influence of matrix types on the mammal communities. We hypothesized that landscape composition variables affect more the mammal communities than the landscape configuration ones. Also, we expected that the scales of effect vary among variables and the more different the matrix to the native habitat, the greater the influence of the matrix on the mammal communities.
2. Materials and Methods
2.1. Study Area
We sampled medium and large-sized mammals in 13 agricultural landscapes (1267 ha each) of Minas Gerais state, southeast of Brazil (Figure 1). This region is in a transitional zone between Atlantic Forest and Cerrado domains, both hotspots for biodiversity conservation [27]. The study region has a subtropical climate with the dry and colder season from April to September and the wet and warmer season from October to March and mean annual temperature and precipitation of 20.4°C and 1,460 mm, respectively [32,33].
We selected the landscape size (two km of radius that result in 1267 ha of area for each landscape) based on similar studies that detected the influence of the landscape in medium and large-sized mammals in landscapes smaller than or equal to two km in radius [34,35]. We studied landscapes composed by small and dispersed fragments of semideciduos forest [31] immerse in an agricultural matrix dominated by coffee crops and pasture (Figure 1). We choose the agricultural landscapes according to a gradient of forest cover in landscapes, considering the central point of the landscape in a forest fragment. As the study region is quite fragmented, the sampled forest cover gradient varies from 10 to 47% in landscapes of 1267 ha. However, we believe that this gradient is sufficient to evaluate the process that shapes the studied communities, since Beca et al. (2017) identified a minimum threshold of 45% of forest cover in landscapes of the same size to detect specialist forest species.
2.2. Data Sampling
We recorded mammal species using one camera trap (Bushnell HD, Bushnell Outdoor, Products, California, USA) per landscape from December 2017 to August 2018. We installed the cameras in the central forest fragment (focal point) of each landscape and they were spaced at least four kilometers apart from each other in order to avoid overlapping the landscapes and thus maintain data independence. Also, we programmed the cameras to record three consecutive photos at a three-second interval when activated, and to work 24 hours for four months in each landscape, totalizing 1483 hours of sampling effort for all landscapes. We considered as independent record the photos of the same species with one hour of interval [36].
We followed the patch-landscape approach to classify the landscape variables, that is, we collected the response variables in a focal fragment and the landscape variables at landscape level around the sampling point [37]. We collected data in eight spatial scales from buffers around focal point with radius sizes ranging from 250 to 2000 meters with 250 m of interval (Figure 2). In each spatial scale we collected five landscape variables: forest cover, coffee cover, pasture cover, number of fragments (NP) and mean distance among forest fragments (NNDis) (Table 1). We collected landscape data using high resolution imagens with a reference scale of 1:5,000 (ArcGIS 10.5 basemap imagery, Digital Globe satellites 2014-2017) and the V-Late extension (2013) in the software ArcGIS 10.5.
2.3. Data Analysis
We considered just the terrestrial and native mammal species for the analysis, excluding eventual records from arboreal and exotic species [38,39]. First, we tested for spatial correlation between species composition and the geographical distance matrix by the Mantel Test. For this, we correlated the matrix of presence and absence of species, using the Jaccard similarity, with the matrix of geographic coordinate points, using the Euclidean distance, from the mantel function of the Vegan package with 999 permutations in the software R (R version 3.4 and RStudio 1.0.136). There was no evidence of spatial correlation between the sampling points (r = - 0.13 p < 0.05). Also, we evaluated the sampling effort per landscape from the 137 accumulation curves performed by the specaccum function of the vegan package in R (R version 3.4 138 and RStudio 1.0.136). The curves were constructed based on the species richness per day of 139 sampling. In general, the curves did not reach the asymptote (Supplementary Figure 1). Then, we 140 compared the observed richness with that estimated by the first-order Jackknife estimator (Jackknife 1) [40] (Supplementary Table 1). We found a mean sampling sufficiency of 70%, which we considered enough to characterize mammals’ communities in the landscapes.
For the selection of the scale of effect and the influence of landscape variables in both response variables (species richness and composition), we used distance-based linear models (DistLM) performed in Primer 6 Permanova+ software [41,42]. We selected the scales of effect from the construction of univariate distance-based linear models using the determination coefficient (R² = proportion of variation explained by the model) as a selection criteria. We selected according to the highest explanation value independently of the significance of the model, once, as the species richness and composition are composed of species with different ecological requirements, the effect of a specific scale may not be evident [24]. So, we built eight univariate distance-based linear models, one for each scale, for each landscape variable and response variable, totalizing 80 models. We did this process for each response variable, species richness and composition, considering the Euclidean distance and Sørensen’s dissimilarity index, respectively. Finally, we verified the meaning of the influence of each explanatory variable on their respective scales of effect on species richness based on scatter plots generated in the R software (R version 3.4 and RStudio 1.0.136).
After selecting the scales of effect, we tested for collinearity of the explanatory variables of each response variable by Pearson’s correlation test from the rcorr function in the vegan package of R. We found that the forest cover and pasture cover were correlated for species richness and composition (Pearsons ‘r = -0.78; Pearsons’ r = -0.73, respectively). Then, we maintained the significant variable or the one with the greatest explanatory power (R²) in the DistLM if none of them were significant. Therefore, for the general species richness model, we maintained the pasture cover and, for the general species composition model, we maintained the forest cover.
Thus, we built a general model for each response variable, including the non-correlated explanatory variables in their respective scales of effect to access the influence of landscape structure on mammal communities. We built the general models using the stepwise strategy and selected from the adjusted R² with 999 permutations and a significance level of P <0.05.
3. Results
We obtained 454 independent records of 24 species of medium and large-sized mammals from nine families, including 18 native terrestrial species, two native arboreal species (Callithrix penicillata and Sapajus nigritus) and four exotic species (Bos taurus, Canis familiaris, Equus caballus and Sus scrofa) (Supplementary Table 2). On average, we recorded six species per landscape, ranging from four to 11 species. Dasypys novencinctus, Nasua nasua and Leopardus pardalis were the most recorded species, occurring in nine of the 13 landscapes (about 70%), while Puma concolor and Chrysocyon brachyurus occurred in only one landscape.
The scales of effect varied among landscape and responses variables. Pasture cover in the landscape was the only variable with effect on the smallest tested scale (250 m) for the two response variables. For species richness, the scales of effect for forest cover and the number of fragments were at 500 m, while for the coffee cover and NNDist at 1500 and 1250 m, respectively. For species composition, the scale of effect for forest cover was at 750 m, for coffee cover and number of fragments at 500 m and for NNDist at 1250 m (Figure 3 and Supplementary Table 3).
In addition, considering the landscape variables in their respective scales of effect, we found that the forest and coffee covers influenced negatively the species richness. While the pasture cover, number of fragments and mean distance among the forest fragments influenced positively the mammal richness (Figure 4).
Mammals Response to Landscape Variables
The variation in species richness was explained only by the pasture cover (adjusted R² = 33%, P = 0.024; Table 2), there is, the pasture in the landscape influences the number of medium and large-sized mammals species in the forest fragments. While the variation in species composition was explained by forest cover (R² = 16%, P = 0.022), coffee cover (R² = 11%, P = 0.142) and the mean distance among the forest fragments (R² = 11%, P = 0.175; Table 3). The three variables together explained 17% of the variation in species composition. However, the coffee cover and the mean distance among the forest fragments were not significant (P> 0.05), suggesting that the inclusion of these variables did not add substantially explanation to the model. So, the forest cover in the landscape interferes in which medium and large-sized mammal species will be in the forest fragments.
4. Discussion
The scales of effect varied among the landscape and response variables, indicating that the landscape variables affect large and medium-sized mammal species in different aspects, highlighting the importance of evaluating different measures to understand the effect of landscape structure on biodiversity. We found that the landscape composition (forest, pasture and coffee covers) influences more the richness and composition of medium and large-sized mammal than the landscape configuration (number of forest fragments and mean distance among forest fragments) and the pasture in the landscape has a positive influence on species richness while the forest cover influences species composition.
The landscape composition and configuration influence ecological processes and can have different effects depending on the community structure and the spatial scale [25,26]. Different variables affect different dimensions of the ecological requirements of organisms leading to scale-dependent responses. As, for example, ecological processes such as metapopulation dynamics can shape ecological responses on larger spatial scales than processes such as competition, predation, resource availability, and anthropogenic impacts such as hunting [43,44]. In addition, different species can have different responses from the same variable at the same scale. That is, the quantity of a given agricultural matrix can be a barrier for the movement of species from the interior of the forest [10,45,46]. However, for generalist species, some agriculture matrix can function as an alternative foraging site, or even displacement corridor [47,48]. Thus, the scale of effect resulting from the response of species communities is the combination of the variation in the influence that each independent variable has on each species.
In our study, the pasture cover was the only variable that explained the variation in species richness, with a high explanation support (33%) for a positive relationship between the pasture cover and species richness. Although this result is contrary to the expected positive relation between richness and native habitat cover in agricultural landscapes [7,49,50], agroecosystems can favor the presence of generalist species from open areas and/or typical of the Cerrado. Studies have shown that landscapes altered by anthropic processes tend to favor the occurrence of species with low dependence on intact ecosystems or forest interior [51,52]. More than 50% of the species registered in our study have wide geographical distribution, varied diet and high movement capacity [53]. That is, species that can use different types of habitats and that can be favored in modified landscapes [54]. In addition, some studies have also found no relationship between the richness of medium and large-sized mammals and the amount of native habitat [8,21,55,56] or others have recorded a high species richness in landscapes with strong urban [58,59] or agricultural influence [8,49,50]. In this way, species with a wide ecological niche can be favored by these modified landscapes and, probably, maintain their populations in agricultural landscapes [60].
On the other hand, the positive relationship between species richness and the pasture cover in the landscape may be due to the use of the almost exclusive remaining fragments by the species, since the proportion of pasture is inversely proportional to that of forest. In addition, we found, for example, that the landscape with the highest number of species (landscape P (e), Supplementary Table 4) has a high pasture cover and a low frequency of individuals of the sampled species. Consequently, the sampled fragment can be used by species present in the landscape as a foraging site, reproduction or as stepping stones, mainly due to the reduced forest cover in the landscape [61,62]. In general, species with large home range, as is the case of most medium and large-sized mammals sampled, which persist in modified landscapes depend on the remaining fragments present in the landscape [63], highlighting the importance of keeping small fragments in agricultural landscapes [17].
Our results also show that despite forest cover does not explain the variation in species richness, it influences species composition, explaining part of the variation between the sampled communities. Some authors [64,65] developed and improved the metapopulation theory based on the analogy to island biogeography theory [66], where the fragment size and isolation in a terrestrial landscape correspond to the size and isolation of oceanic islands. Since then, studies on habitat fragmentation include variables to represent this analogy and explain the relationship between species-area in continental environments [67,68]. However, the matrices around the fragments affect the biodiversity differently than the ocean, since fragmented landscapes are not binary like oceanic islands [45,69]. Nevertheless, Fahrig [16] argued that both effects of fragment size and isolation are shaped by the effect of the sampling area. That is, in a landscape of a certain size, the amount of forest habitat around the sampled forest fragment would be a better predictor for species richness [16] and other ecological responses [9,70] than fragment size and isolation [16]. In addition, Fahrig [16] adds that the matrix effect is secondary to the amount of habitat and that the landscape configuration should have little or no effect on the species. Although we have not corroborated this hypothesis for species richness, we found that the amount of native habitat (i.e., forest cover) was the variable with the greatest effect on the mammal composition, followed, and with minor importance, by the coffee cover and finally by mean distance among forest fragments.
As landscape composition has a greater effect on medium and large-sized mammals than landscape configuration, more heterogeneous and structurally more complex agricultural landscapes can favor greater connectivity between the fragments and increase the diversity of species [71]. In addition, the role and management of the matrix in order to contribute to the conservation of biodiversity is one of the main strategies for the permanence of species in modified landscapes [14]. However, matrix management involves economic and social aspects in addition to ecological ones [72] and, therefore, an interdisciplinary approach is necessary for conservation measures in agroecosystems to be applied and effective.
Finally, exploring different responses to biodiversity can be important to understand different aspects of landscape influence on species. As well, the scale in which variables of landscape composition and configuration are measured can interfere with the species-landscape relationship. Therefore, we emphasize the importance of future studies on the relationship between landscape structure and biodiversity to consider different biodiversity responses and a multiscale approach with a wide range of scales. Also, with the advancement of geoprocessing tools and development of free software, such studies are currently more viable. Considering landscape composition and configuration variables on an appropriate spatial scale allows us to expand our knowledge about the relationship between landscape and biodiversity and thus improve management and conservation measures in agricultural ecosystems.
Author Contributions
RFP and MP contributed to the conception and study design; RFP and MP conducted the fieldwork and data analysis; RFP, CR and MP contributed to the interpretation, writing and revision of the manuscript.
Funding
This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001, and by Fundação de Amparo à Pesquisa de Minas Gerais (FAPEMIG). MP was supported by the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Acknowledgments
We thank to F. Vilas Boas, M. Melo-Dias, A. Soto-Werschitz, F.F. Siqueira, and M. Siqueira for the field help. We are grateful for NKG – Fazenda da Lagoa, Fazenda Pinhal, Fazenda Bom Jardim and all landowners who allowed the data sampling. A special thanks to R. Monserrat from NKG – Fazenda da Lagoa for the assistance during the data collection and to F. F. Siqueira for the support with image processing.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Contribution to the Field Statement
Habitat loss and forest fragmentation are the main causes of biodiversity loss and the main reason is the conversion of native habitats to agricultural areas. These degradation processes alter the original composition and configuration of the landscape and consequently affect the ecological dynamics of the species due to changes in conditions and resource availability. However, the effects of habitat loss and forest fragmentation on biodiversity are often not distinguished and the scale at which they are assessed is often not adequate, making it difficult to understand the main impacts and limitations of how the conversion of native habitats affects biodiversity. Thus, our study provides new insights into the importance of landscape composition for medium and large sized mammals and how the species responses to the landscape variables may differ according to the scale at which the variables are assessed. In addition, we found that the pasture cover in the landscape positively influenced the mammal richness, a different result from the expected, but which allows further investigations in the field area. Likewise, our study emphasizes the importance of considering the different land uses for biodiversity conservation measures and a multiscale approach to assess the influence of landscape on species.
References
- Kissinger, G., M. Herold, V. De Sy. Drivers of Deforestation and Forest Degradation: A Synthesis Report for REDD+ Policymakers. Lexeme Consulting, Vancouver Canada, 2012, 46p.
- Dunning, J.B.; Danielson, B.J.; Pulliam, H.R. Ecological processes that affect populations in complex landscapes. Oikos. 1992, 65, 169–175. [Google Scholar] [CrossRef]
- Redford, K.H. The empty forest. BioScience. 1992, 42, 412–422. [Google Scholar] [CrossRef]
- Chiarello, A.G. Effects of fragmentation of the Atlantic forest on mammal communities in south-eastern Brazil. Biological Conservation. 1999, 89, 71–82. [Google Scholar] [CrossRef]
- Bello, C.; Galetti, M.; Pizo, M.A.; Magnago, L.F. S.; Rocha, M.F. , et al. Defaunation affects carbon storage in tropical forests. Science advances. 2005, 1, 1–10 e1501105. [Google Scholar]
- Bogoni, J.A.; Pires, J.S. R.; Graipel, M.E.; Peroni, N.; Peres, C.A. Wish you were here: How defaunated is the Atlantic Forest biome of its medium-to large-bodied mammal fauna? ., PloS one. 2018, 13, e0204515. [Google Scholar] [CrossRef] [PubMed]
- Regolin, A.L.; Cherem, J.J.; Graipel, M.E.; Bogoni, J.A.; Ribeiro, J.W.; et al. Forest cover influences occurrence of mammalian carnivores within Brazilian Atlantic Forest. Journal of Mammalogy. 2017, 98, 1721–1731. [Google Scholar] [CrossRef]
- Beca, G.; Vancine, M.H.; Carvalho, C.S.; Pedrosa, F.; Alves, R.S. C.; et al. High mammal species turnover in forest patches immersed in biofuel plantations. Biological Conservation. 2017, 210, 352–359. [Google Scholar] [CrossRef]
- Gestich, C.C.; Arroyo-Rodríguez, V.; Ribeiro, M.C.; da Cunha, R.G.; Setz, E.Z. Unraveling the scales of effect of landscape structure on primate species richness and density of titi monkeys (Callicebus nigrifrons). Ecological Research, 2019, 34, 150–159. [Google Scholar] [CrossRef]
- Prugh, L.R.; Hodges, K.E.; Sinclair, A.R.; Brashares, J.S. Effect of habitat area and isolation on fragmented animal populations. Proceedings of the National Academy of Sciences. 2008, 105, 20770–20775. [Google Scholar] [CrossRef]
- Kupfer, J.A.; Malanson, G.P.; Franklin, S.B. Not seeing the ocean for the islands: the mediating influence of matrix-based processes on forest fragmentation effects. Global ecology and biogeography. 2006, 15, 8–20. [Google Scholar] [CrossRef]
- Didham, R.K.; Kapos, V.; Ewers, R.M. Rethinking the conceptual foundations of habitat fragmentation research. Oikos. 2012, 121(2), 161–170. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Vieira, M.V. Does the type of matrix matter? A quantitative review of the evidence. Biodiversity and Conservation. 2010, 19, 1205–1223. [Google Scholar] [CrossRef]
- Driscoll, D.A.; Banks, S.C.; Barton, P.S.; Lindenmayer, D.B.; Smith, A.L. Conceptual domain of the matrix in fragmented landscapes. Trends in Ecology & Evolution. 2013, 28, 605–613. [Google Scholar]
- McGarigal, K.; Cushman, S.A. Comparative evaluation of experimental approaches to the study of habitat fragmentation effects. Ecological Applications. 2002, 12, 335–345. [Google Scholar] [CrossRef]
- Fahrig, L. Rethinking patch size and isolation effects: the habitat amount hypothesis. Journal of Biogeography. 2013, 40, 1649–1663. [Google Scholar] [CrossRef]
- Fahrig, L. Ecological responses to habitat fragmentation per se. Annual Review of Ecology, Evolution, and Systematics. 2017, 48, 1–23. [Google Scholar] [CrossRef]
- Hanski, I. Habitat fragmentation and species richness. Journal of Biogeography. 2015, 42(5), 989–993. [Google Scholar] [CrossRef]
- Fletcher Jr, R.J.; Reichert, B.E.; Holmes, K. The negative effects of habitat fragmentation operate at the scale of dispersal. Ecology. 2018, 99, 2176–2186. [Google Scholar] [CrossRef] [PubMed]
- Villard, M.A.; Metzger, J.P. Beyond the fragmentation debate: a conceptual model to predict when habitat configuration really matters. Journal of Applied Ecology. 2014, 51, 309–318. [Google Scholar] [CrossRef]
- Püttker, T. , Indirect effects of habitat loss via habitat fragmentation: A cross-taxa analysis of forest-dependent species. Biological Conservation. 2020, 241. [Google Scholar] [CrossRef]
- Brennan, J.M.; Bender, D.J.; Contreras, T.A.; Fahrig, L. Focal patch landscape studies for wildlife management: Optimizing sampling effort. In: Liu, J; Tailor W.W. Integrating landscape ecology into natural resource management. Cambridge: Cambridge University Press. 2002. [Google Scholar]
- Jackson, H.B.; Fahrig, L. What size is a biologically relevant landscape? . Landscape Ecology. 2012, 27, 929–941. [Google Scholar] [CrossRef]
- Jackson, H.B.; Fahrig, L. Are ecologists conducting research at the optimal scale? . Global Ecology and Biogeography. 2015, 24, 52–63. [Google Scholar] [CrossRef]
- Miguet, P.; Jackson, H.B.; Jackson, N.D.; Martin, A.E.; Fahrig, L. What determines the spatial extent of landscape effects on species? Landscape Ecology. 2016, 31, 1177–1194. [Google Scholar] [CrossRef]
- Suárez-Castro, A.F.; Simmonds, J.S.; Mitchell, M.G.; Maron, M.; Rhodes, J.R. The scale-dependent role of biological traits in landscape ecology: a review. Current Landscape Ecology Reports. 2018, 3, 12–22. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature. 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Galindo-Leal, C.; Câmara, I.D. G. Atlantic Forest hotspot status: an overview. The Atlantic Forest of South America: biodiversity status, threats, and outlook. 1 edition, Island Press. USA, 2003, 408p.
- Laurance, W.F. Conserving the hottest of the hotspots. Biological Conservation. 2009, 142, 1137. [Google Scholar] [CrossRef]
- Strassburg, B.B.; Brooks, T.; Feltran-Barbieri, R.; Iribarrem, A.; Crouzeilles, R. , et al. Moment of truth for the Cerrado hotspot. Nature Ecology & Evolution. 2017, 1, 1–3. [Google Scholar]
- Ribeiro, M.C.; Metzger, J.P.; Martensen, A.C.; Ponzoni, F.J.; Hirota, M.M. The Brazilian Atlantic Forest: How much is left, and how is the remaining forest distributed? Implications for conservation. Biological Conservation. 2009, 142, 1141–1153. [Google Scholar]
- Schiffler, G. (2003). Fatores determinantes da riqueza local de espécies de Scarabaeidae (Insecta Coleoptera) em fragmentos de floresta estacional semidecídua. [master’s thesis] [Lavras (MG)]: Universidade Federal de Lavras.
- Dantas, A.A. A.; Carvalho, L.G. D.; Ferreira, E. Classificação e tendências climáticas em Lavras, MG. Ciência e Agrotecnologia. 2007, 31, 1862–1866. [Google Scholar] [CrossRef]
- Andrade-Núñez, M.J.; Aide, T.M. Effects of habitat and landscape characteristics on medium and large mammal species richness and composition in northern Uruguay. Zoologia. 2010, 27, 909–917. [Google Scholar] [CrossRef]
- Lyra-Jorge, M.C.; Ribeiro, M.C.; Ciocheti, G.; Tambosi, L.R.; Pivello, V.R. Influence of multi-scale landscape structure on the occurrence of carnivorous mammals in a human-modified savanna, Brazil. European Journal of Wildlife Research. 2010, 56, 359–368. [Google Scholar] [CrossRef]
- Srbek-Araujo, A.C.; Chiarello, A.G. Is camera-trapping an efficient method for surveying mammals in Neotropical forests? A case study in south-eastern Brazil. Journal of Tropical Ecology. 2005, 121–125. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Fahrig, L. Why is a landscape perspective important in studies of primates? American Journal of Primatology. 2014, 76, 901–909. [Google Scholar] [CrossRef] [PubMed]
- Paglia, A.P.; Da Fonseca, G.A.; Rylands, A.B.; Herrmann, G.; Aguiar, L.M.; Chiarello, A. G, et al. Lista Anotada dos Mamíferos do Brasil 2ª Edição/Annotated Checklist of Brazilian Mammals. Occasional papers in conservation biology. 2012, 6, 1–82 Arlington: Conservation International. [Google Scholar]
- Quintela, F.; Rosa, C.A.; Feijo, A. Updated and annotated checklist of recent mammals from Brazil. Anais da Academia Brasileira de Ciências. 2020, 92.
- Smith, C.D.; Pontius, J.S. Jackknife estimator of species richness with S-PLUS. Journal of Statistical Software 15: 1–12. 2006.
- Legendre, P.; Anderson, M.J. Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments. Ecological Monographs. 1999, 69(1), 1–24. [Google Scholar] [CrossRef]
- McArdle, B.H.; Anderson, M.J. Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology. 2001, 82, 290–297. [Google Scholar] [CrossRef]
- Arroyo-Rodríguez, V.; Mandujano, S. Conceptualization and measurement of habitat fragmentation from the primates’ perspective. International Journal of Primatology. 2009, 30, 497–514. [Google Scholar] [CrossRef]
- Marsh, C.; Link, A.; King-Bailey, G.; Donati, G. Effects of fragment and vegetation structure on the population abundance of Ateles hybridus, Alouatta seniculus and Cebus albifrons in Magdalena Valley, Colombia. Folia Primatologica. 2016, 87, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Ricketts, T.H. The matrix matters: effective isolation in fragmented landscapes. The American Naturalist. 2001, 158, 87–99. [Google Scholar] [CrossRef]
- Prevedello, J.A.; Gotelli, N.J.; Metzger, J.P. A stochastic model for landscape patterns of biodiversity. Ecological Monographs. 2016, 86, 462–479. [Google Scholar] [CrossRef]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biological Reviews. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Caryl, F.M.; Quine, C.P.; Park, K.J. Martens in the matrix: the importance of nonforested habitats for forest carnivores in fragmented landscapes. Journal of Mammalogy. 2012, 93, 464–474. [Google Scholar] [CrossRef]
- Gheler-Costa, C.; Vettorazzi, C.A.; Pardini, R.; Verdade, L.M. The distribution and abundance of small mammals in agroecosystems of southeastern Brazil. Mammalia. 2012, 76(2), 185–191. [Google Scholar] [CrossRef]
- Dotta, G.; Verdade, L.M. Medium to large-sized mammals in agricultural landscapes of south-eastern Brazil. Mammalia. 2011, 75, 345–352. [Google Scholar] [CrossRef]
- Ferraz, K.M. P. B.; Ferraz, S.F. B.; Moreira, J.R.; Couto, H.T. Z.; Verdade, L.M. Capybara (Hydrochoerus hydrochaeris) distribution in agroecosystems: A cross-scale habitat analysis. Journal of Biogeography. 2007, 34(2), 223–230. [Google Scholar] [CrossRef]
- Calvão, L.B.; Juen, L.; de Oliveira Junior, J.M. B.; Batista, J.D.; Júnior, P.D. M. Land use modifies Odonata diversity in streams of the Brazilian Cerrado. Journal of Insect Conservation. 2018, 22(5-6), 675-685.
- Reis, N.; Peracchi, A.; Pedro, W.; Lima, I. Mamíferos do Brasil. Londrina, Brasil, 2006, 437p.
- Sánchez-Zapata, J.A.; Calvo, J.F. Raptor distribution in relation to landscape composition in semi-arid Mediterranean habitats. Journal of Applied Ecology. 1999, 36, 254–262. [Google Scholar] [CrossRef]
- Pardo, L.E.; Campbell, M.J.; Edwards, W.; Clements, G.R.; Laurance, W.F. Terrestrial mammal responses to oil palm dominated landscapes in Colombia. PloS one. 2018, 13, e0197539. [Google Scholar] [CrossRef] [PubMed]
- Soto-Werschitz, A. ; Mandujano, S; Passamani, M. Influence of forest type on the diversity, abundance, and naïve occupancy of the mammal assemblage in the southeastern Brazilian Atlantic Forest. Therya. 2023, 14.
- Dias, W.A. F.; Tezori, R.F. F.; Oliveira, A.K. Registro de mamíferos de médio e grande porte em dois fragmentos florestais no município de São Carlos, Estado de São Paulo. Multiciência. 2012, 11, 277–293. [Google Scholar]
- Melo-Dias, M.; Passamani, M. Mamíferos de médio e grande porte no campus da Universidade Federal de Lavras, sul do estado de Minas Gerais, Brasil. Oecologia Australis. 2018, 22. [Google Scholar] [CrossRef]
- González-Suárez, M.; Gómez, A.; Revilla, E. Which intrinsic traits predict vulnerability to extinction depends on the actual threatening processes. Ecosphere. 2013, 4, 1–16. [Google Scholar] [CrossRef]
- Fahrig, L.; Merriam, G. Conservation of fragmented populations. Conservation Biology. 1994, 8, 50–59. [Google Scholar] [CrossRef]
- Sampaio, R.; Lima, A.P.; Magnusson, W.E.; Peres, C.A. Long-term persistence of midsized to large-bodied mammals in Amazonian landscapes under varying contexts of forest cover. Biodiversity and Conservation. 2010, 19, 2421–2439. [Google Scholar] [CrossRef]
- Holt, R.D. Food webs in space: an island biogeographic perspective. Food webs. Boston: Springer.
- Levins, R. Some demographic and genetic consequences of environmental heterogeneity for biological control. American Entomologist. 1969, 15, 237–240. [Google Scholar] [CrossRef]
- Hanski, I. Single-species metapopulation dynamics: concepts, models and observations. Biological Journal of the Linnean Society. 1991, 42, 17–38. [Google Scholar] [CrossRef]
- MacArthur, R.H. , & Wilson, E.O. The theory of island biogeography (Vol. 1). Princeton University Press. USA, 2001. 203p.
- Haila, Y. A conceptual genealogy of fragmentation research: from island biogeography to landscape ecology. Ecological Applications. 2002, 12, 321–334. [Google Scholar]
- Laurance, W.F. Theory meets reality: how habitat fragmentation research has transcended island biogeographic theory. Biological Conservation. 2008, 141, 1731–1744. [Google Scholar] [CrossRef]
- Anderson, J.; Rowcliffe, J.M.; Cowlishaw, G. Does the matrix matter? A forest primate in a complex agricultural landscape. Biological Conservation. 2007, 135, 212–222. [Google Scholar]
- Rabelo, R.M.; Aragón, S.; Bicca-Marques, J.C.; Nelson, B.W. Habitat amount hypothesis and passive sampling explain mammal species composition in Amazonian river islands. Biotropica. 2019, 51, 84–92. [Google Scholar] [CrossRef]
- Perović, D.J.; Gurr, G.M.; Raman, A.; Nicol, H.I. Effect of landscape composition and arrangement on biological control agents in a simplified agricultural system: a cost–distance approach. Biological Control. 2010, 52, 263–270. [Google Scholar] [CrossRef]
- Phalan, B.; Onial, M.; Balmford, A.; Green, R.E. Reconciling food production and biodiversity conservation: land sharing and land sparing compared. Science. 2011, 333, 1289–1291. [Google Scholar] [CrossRef]
- Boesing, A.L.; Nichols, E.; Metzger, J.P. Land use type, forest cover and forest edges modulate avian cross-habitat spillover. Journal of Applied Ecology. 2018, 55, 1252–1264. [Google Scholar] [CrossRef]
- Lang, S.; Blaschke, T. 2009. Análise da paisagem com SIG, São Paulo: Oficina de textos, Brazil, 2009, 424p.
Figure 1.
Study area in Minas Gerais, Brazil. The white circles in the image represent the largest buffers (2 km radius) around the focal fragment where the trap camera was installed.
Figure 1.
Study area in Minas Gerais, Brazil. The white circles in the image represent the largest buffers (2 km radius) around the focal fragment where the trap camera was installed.
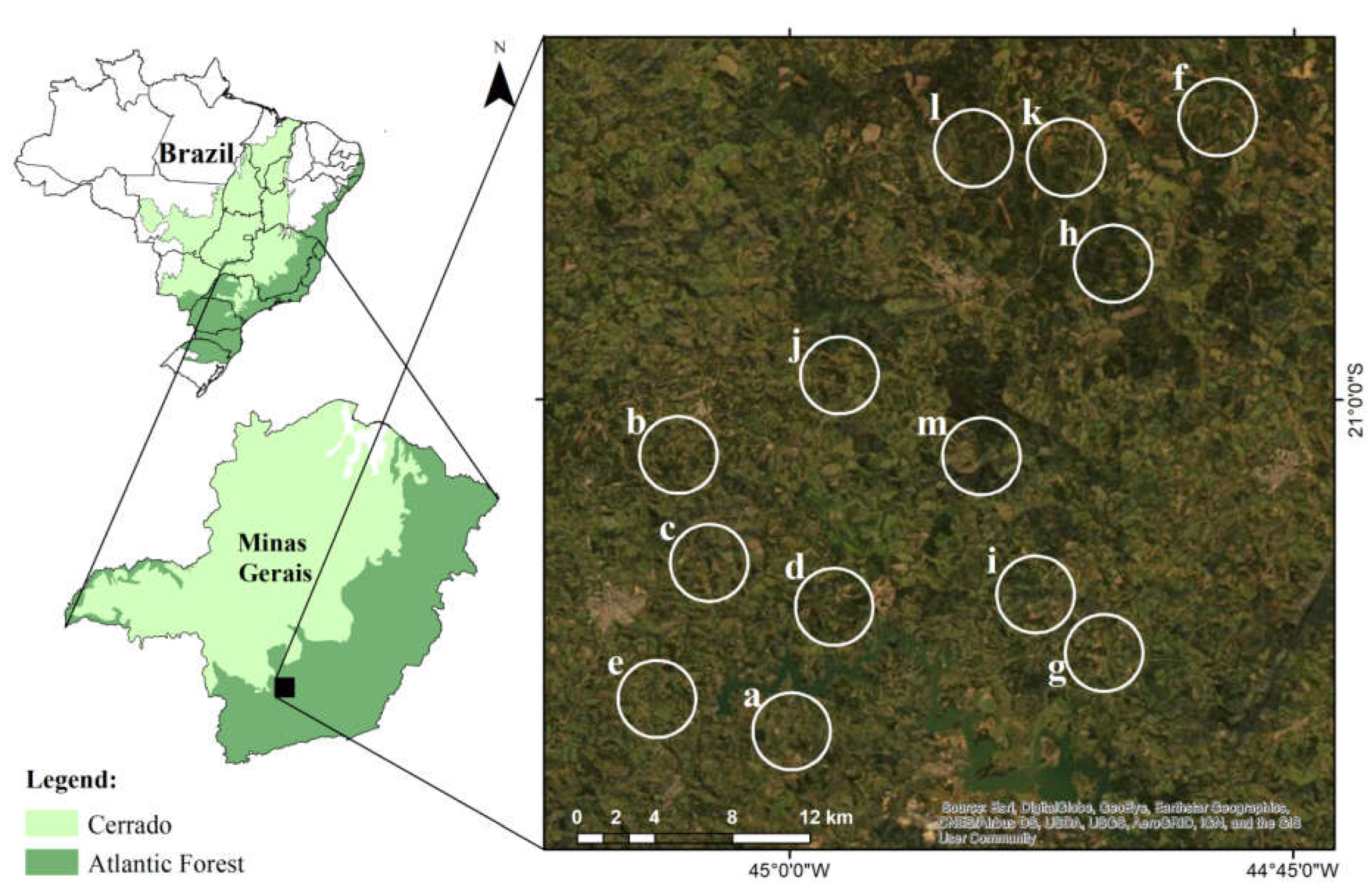
Figure 2.
Sampling landscapes in increasing order (“a” to “m”) of forest cover. The eight buffers around the focal point, where each camera was installed, represent the scales evaluated for each landscape variable.
Figure 2.
Sampling landscapes in increasing order (“a” to “m”) of forest cover. The eight buffers around the focal point, where each camera was installed, represent the scales evaluated for each landscape variable.

Figure 3.
The scales of effect of each landscape variable for each response variable. R² = determination coefficient. Filled points = selected scales of effect.
Figure 3.
The scales of effect of each landscape variable for each response variable. R² = determination coefficient. Filled points = selected scales of effect.
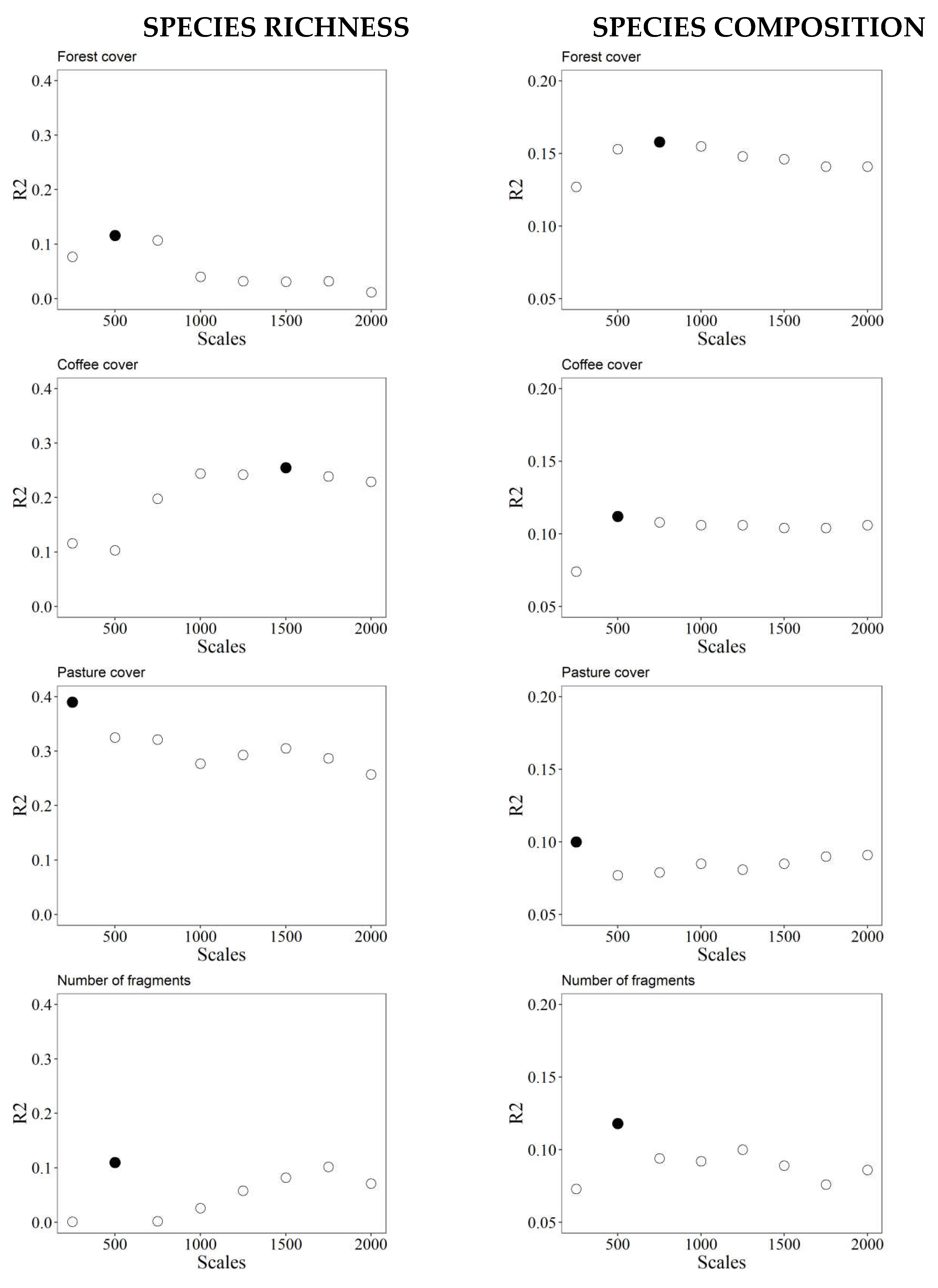
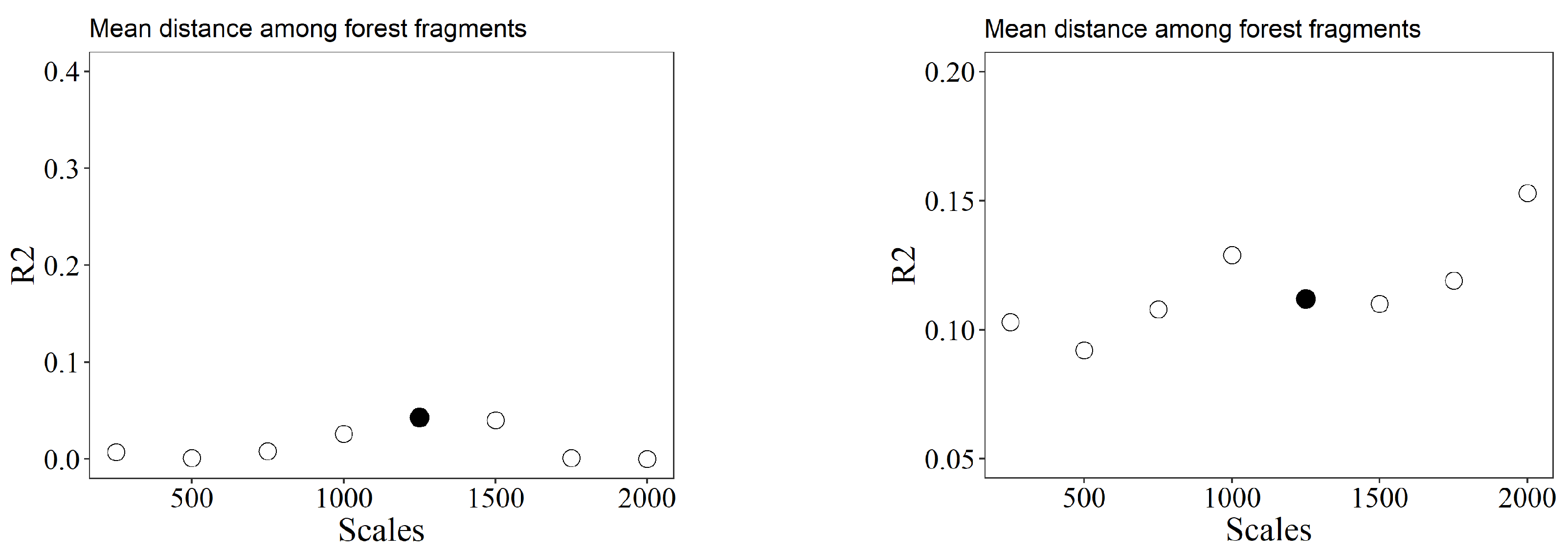
Figure 4.
Scatter plots for species richness and landscape variables analyzed in their respective scale of effect.
Figure 4.
Scatter plots for species richness and landscape variables analyzed in their respective scale of effect.
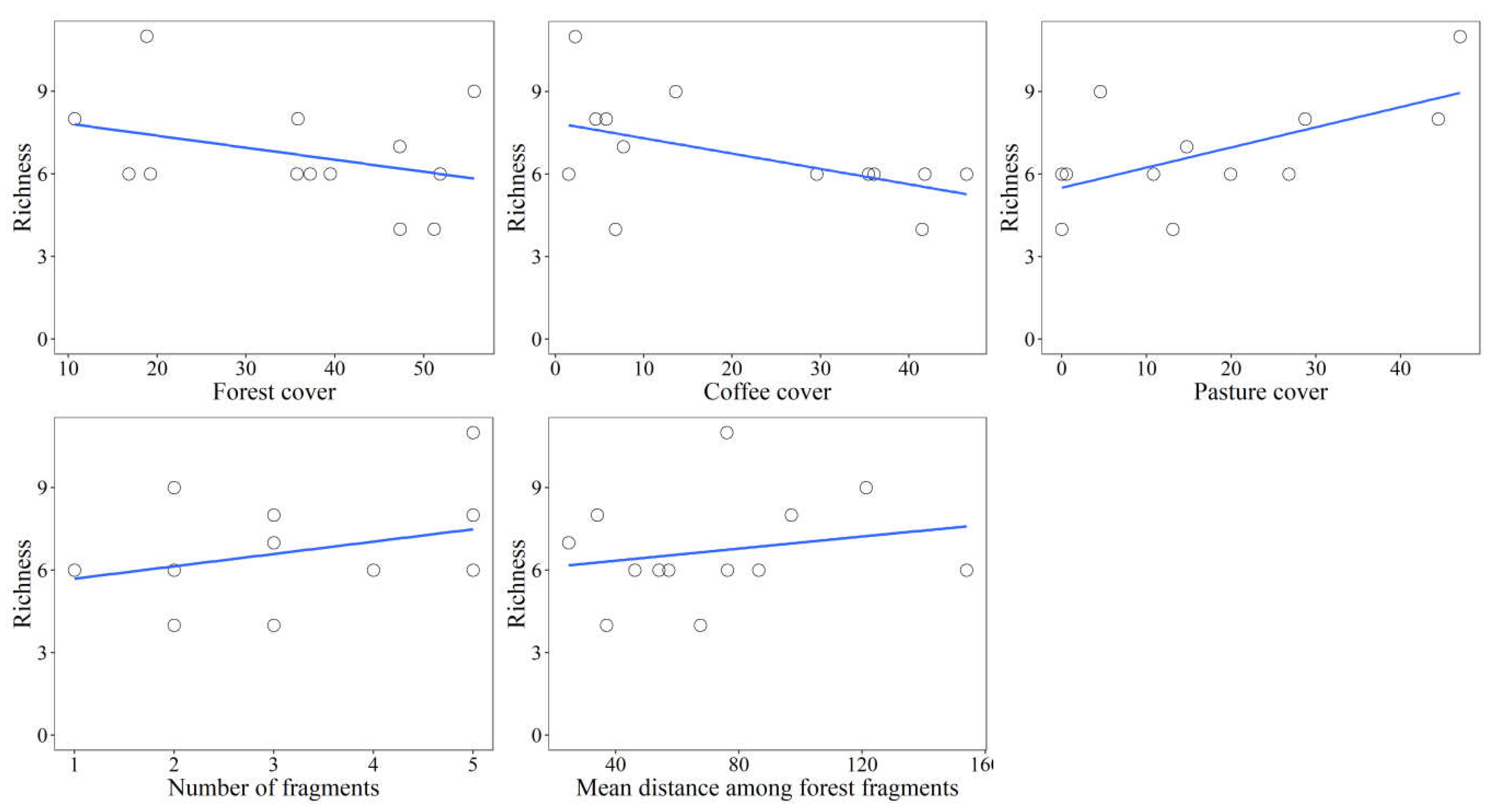

Table 1.
Landscape variables selected to evaluate the influence of landscape structure on medium and large sized mammals in agroecosystems in Minas Gerais, Brazil.
Table 1.
Landscape variables selected to evaluate the influence of landscape structure on medium and large sized mammals in agroecosystems in Minas Gerais, Brazil.
| Variable | Meaning | Ecological interpretation | Reference |
|---|---|---|---|
| Composition landscape variables | |||
| Forest cover | Proportion of area covered by forest |
Proxy for habitat amount in the landscape |
[16] |
| Coffee cover | Proportion of area covered by coffee |
More permeable matrix for forest species |
[73] |
| Pasture cover | Proportion of area covered by pasture |
Less or not permeable matrix for forest species |
[50] |
| Configuration landscape variables | |||
| NP | Number of forest fragments in landscape |
More fragments indicates more fragmentation |
[17] |
| NNDist |
Mean distance among forest fragments (m) |
The more isolated the fragments indicate more fragmentation |
[74] |
Table 2.
Sequential test of the distance-based linear model for species richness. Only uncorrelated variables were considered.
Table 2.
Sequential test of the distance-based linear model for species richness. Only uncorrelated variables were considered.
| Variable (scale) | Adj R^2 | Pseudo-F | P | R² | Cumul. | res.df |
| +Pasture cover (250 m) |
0.3344 | 7.0298 | 0.024 | 0.389 | 0.389 | 11 |
Legend: Adj R² = proportion of the explained variation adjusted by the number of variables included in the model; R² = proportion of the variance explained by each variable; Cumul = cumulative proportion of the variance explained by multiple variables; res.def = degrees of freedom of the residues.
Table 3.
Sequential test of the distance-based linear model for species composition. Only uncorrelated variables were considered.
Table 3.
Sequential test of the distance-based linear model for species composition. Only uncorrelated variables were considered.
| Variable (scale) | Adj R^2 | Pseudo-F | P | Prop. | Cumul. | res.df |
| +Forest cover (750 m) | 0.0816 | 2.0668 | 0.022 | 0.1582 | 0.15817 | 11 |
| +Coffee cover (500 m) | 0.1226 | 1.5141 | 0.142 | 0.1107 | 0.26887 | 10 |
| +NNDist (2000 m) | 0.1658 | 1.5171 | 0.175 | 0.10547 | 0.37434 | 9 |
Legend: Adj R² = proportion of the explained variation adjusted by the number of variables included in the model; R² = proportion of the variance explained by each variable; Cumul = cumulative proportion of the variance explained by multiple variables; res.def = degrees of freedom of the residues; NNDist = mean distance among forest fragments.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



