
Submitted:
28 May 2024
Posted:
29 May 2024
You are already at the latest version
Abstract
Traditional asexual propagation of hops limits its phytosanitary quality and the concomitant plant availability. This scenario worsens with the global climate change. In this study we develop protocols for in vitro propagation of Humulus lupulus var. mapuche. Two explant types were evaluated: shoot tips, from field material collected in Río Negro province (Argentina), and nodal segments, from vitroplants. Different growth regulator, such as 6-Benzylaminopurine, gibberellic acid, indole-3-butyric acid and indole-3-acetic acid, was used to induce desired morphogenic responses. Additionally, the impact of abscisic acid application on acclimatization was assessed, and resulted in robust growth and reduced plant stress symptoms during ex vitro adaptation. Also, the study develops a protocol for synthetic seeds production, with storage at 4°C to enhance conservation and plant material transport, ensuring a constant supply of female plants. This comprehensive approach addresses current producer demands and emerging climate challenges. The findings highlight the fundamental role of in vitro multiplication of hops in improving plant material availability and quality, with significant implications for the brewing industry and related sectors.
Keywords:
hops
; in vitro propagation
; phytosanitary quality
; acclimatization
; encapsulation
; climate challenges
1. Introduction
Humulus lupulus L. is a species of economic relevance, especially in countries located between latitudes 35° and 55° north and south of the equator [1], where environmental conditions favor its optimal development. However, the specific combination of climate and soil required by hops increases its vulnerability to extreme weather events, which are expected to become more frequent in the current global context. The expansion of craft breweries has driven the demand for hops worldwide [2], posing new challenges and opportunities for producers in terms of expanding the area under cultivation, improving product quality, and achieving socioeconomic and environmental sustainability. Additionally, global climate change (GCC) is altering weather patterns and water availability, significantly affecting different agricultural sectors, especially crops with specific environmental requirements, such as hops.
In recent years, different studies have highlighted the adverse effects of GCC on hop production, impacting both quantity and quality [3,4]. In a recent study, Mozny et al. [2] analyzed historical data from major European hop-producing regions and revealed that drought and high temperatures during the growth stage negatively affected hop crops, advancing their maturation and reducing the production and the concentration of alpha acids content, responsible for the bitter taste of beer.
In addition, to the GCC impacts, the spread of pathogens like downy mildew, powdery mildew, and different viruses through contaminated material poses a significant threat to the brewing industry, evidenced by considerable economic and yield losses observed [5,6,7,8,9,10]. For instance, in the 1990s, hop production in the United States suffered significant losses and crop damage due to powdery mildew (Podosphaera macularis) [11]. However, the lack of updated global figures makes it challenging to accurately assess the actual impact of pests and pathogens on the hop market.
The exclusive demand for female plants by the brewing industry and the dioecious nature of the species present challenges for its propagation by seeds, as this could result in individuals of both sexes. Therefore, producers resort to vegetative propagation by rhizomes or cuttings [12]. However, these practices sustained over time may cause certain disadvantages in terms of phytosanitary quality, as they favor the accumulation and dispersal of pathogens. In this regard, Agehara et al. [5] reported that the propagation of hop plants from in vitro culture has been shown not to cause significant problems with diseases, according to observations made in an experimental field. This underscores the importance of the cultivation and regeneration of meristems to obtain material of phytosanitary quality, which represents an effective solution for the prevention and control of pathogens in hop cultivation. Thus, micropropagation emerges as an effective alternative due to its multiple advantages; however, the success of these biotechniques depends largely on the efficiency of the acclimatization period, critical for correcting anatomical-functional abnormalities related to in vitro conditions. During this process, it is crucial to carefully address stomatal dysfunctions, high transpiration rates, and the presence of a thin cuticle, which can lead to leaf and/or plant wilting and senescence [13,14,15]. Several studies have linked these issues to the limited capacity of plants to produce ABA [16,17]. This phytohormone plays a key role in inducing drought tolerance, as well as other types of biotic and abiotic stress [18], and participates in various plant physiological processes, including stomatal closure [19,20]. In this regard, Dias et al. [21] reported reduced water loss due to transpiration and prevention of oxidative damage in the acclimatization of Ulmus minor plants after treatment with a foliar application of ABA.
The objective of this study was to evaluate the response to in vitro culture and the ex vitro acclimatization of Humulus lupulus var. mapuche, as an alternative to traditional asexual propagation. We have established an efficient propagation protocol that guarantees the phytosanitary quality and availability of plants in a short period of time, and facilitates the transport and exchange of material. On the other hand, numerous studies have addressed plant tissue culture and explored various pathways of hop regeneration, and have evaluated the response of different types of explants and varieties [22,23,24,25,26,27,28,29,30,31,32]. In addition, other studies have been conducted on organogenic callus production and regeneration [33,34,35], virus elimination by thermotherapy and meristem culture [36], and encapsulation of nodal segments and shoot tips to obtain synthetic seeds [26,37,38]. These techniques offers several outstanding advantages in modern agriculture, including the possibility of conserving plant species through low-temperature storage, facilitating the transfer and exchange of high-quality germplasm, as well as pathogen-free and genetically modified material [39,40,41,42,43,44,45,46,47,48]. Additionally, somatic or synthetic seeds offer a crucial advantage in the case of hops, as they guarantee the multiplication of female plants. Such seeds are obtained from specific plant tissues, ensuring clonal propagation and, therefore, the maintenance of desirable characteristics, such as the female sex, of the resulting plants. However, despite the many advantages of this biotechnique in terms of crop genetic diversification and the preservation of agricultural biodiversity, its application and study have been limited to a small number of plant species [49,50,51,52].
In conclusion, this research addresses various aspects of hop plant tissue culture, including the micropropagation of different explant types, improvements in ex vitro acclimatization, and the production of synthetic seeds. With this approach, we aim to provide a more comprehensive understanding of the possibilities and challenges in the application of various biotechniques in hop cultivation, with a view to offering innovative solutions for the brewing industry in the context of current climate challenges.
2. Results
2.1. Effect of Plants Growth Regulators on Shoot Tip Cultivation
The addition of growth regulators was crucial to induce morphogenesis since significant differences were found in terms of explant regeneration and elongation, compared to the control (Table 1; Figure 1). In this sense, at the end of the assay, shoot tips cultivated in M1 initiated a process of oxidation and subsequent necrosis. The differentiation and regeneration of shoot tips were started after four weeks of cultivation (Figure 1 b-c), whereas complete plants with approximately five nodes, were obtained after 14 weeks. On the other hand, explants cultivated in M2 and M3 exhibited callus formation in the cutting zone (4.44 ± 3.2% and 6.67 ± 2.3%, respectively), along with isolated rooting events through indirect organogenesis (6.67 ± 1.50% in both treatments).
2.2. Rooting of Uninodal Segments
Although auxinic media are generally used for root induction, the highest percentage of rooting of the uninodal segments obtained occurred in the absence of these regulators. Furthermore, in these cases, rooting was through the direct path (Figure 2a), whereas the roots obtained in media provided with auxins were produced by indirect organogenesis, originating from calli formed at the basal part of the explant (Figure 2b; Table 2). In contrast, to the results obtained for shoot tip regeneration, the nodal segments produced complete plants with similar characteristics (five nodes) after six weeks of cultivation.
2.3. ABA Effects in the Acclimatization Process
The acclimatization and rustication (Figure 3) of complete plants obtained from
in vitro culture is a critical step for their adaptation prior to exposure to the stressfull ex vitro environment. Usually, plant survival is impared by water loss and senescence. The phytohormone ABA play a key role in plant adaptation to stress. However, after 21 days of transplantation into substrate in the ex vitro environment, it was observed that the percentage of surviving plants did not vary between ABA treated plants (50 µM and 100 µM) respect to mock (data not shown). Nevertheless, differences were observed in relation to other phenotypes, such as seedling growth, number of green leaves and number of senescent leaves (Figure 4). For instance, data showed an increment in growth and number of green leaves in plants treated with 50 µM ABA respect to mock and 100 µM ABA (Figure 4d-e). Moreover, the number of senescent leaves decrease in ABA treated seedlings (50 µM, 100 µM) respect to mock (Figure 4f).
2.4. Synthetic Seeds
The conversion rate of synthetic seeds (Figure 5a) exhibited no significant differences (p>0.05) between ½ MS with BAP (0.88 µM) and GA3 (2.89 µM) and ½ MS (mock). However, after 3 weeks of incubation at 21±2°C, in the medium supplemented with BAP and GA3, only elongation of pre-existing axillary buds was observed, whereas in the medium without plant growth regulators, complete plants were obtained (figure 5b). Consequently, this medium was selected for the cultivation of cold-treated synthetic seeds.
The results obtained in the different treatments demonstrate that cold exposure had a negative impact on the conversion of synthetic seeds into seedlings. The synthetic seeds stored at 4ºC showed significant differences in the evaluated parameters (conversion, rooting, and viability) compared to the control, after three weeks of incubation at 21±2°C. The conversion and rooting percentages varied according to the duration of exposure to cold, ranging between 46.67% and 76.67%, and 26.67% and 46.67%, respectively (Table 3). Additionally, a high percentage of encapsulated explants were viable.
3. Discussion
This research has demonstrated the feasibility of two in vitro multiplication methods for hops: nodal segment culture and shoot tip regeneration. Both alternatives present different advantages that must be considered in the context of the brewing industry and the challenges associated with global climate change.
Nodal segment cultivation emerges as an economical and fast alternative, as it does not require the use of growth regulators. This could address the unmet demand of the brewing industry by offering a cost-effective and efficient way to obtain plant material for hop production.
On the other hand, shoot tip propagation, despite slower and more expensive due to the need of growth regulators, ensures the healthiness of the obtained material. This technique is widely used for cloning pathogen-free plants and could be crucial for maintaining the quality and productivity of hop in the context of climate change. In this regard, global changes in rainfall patterns may increase the incidence of fungal diseases in hop crops. Establishing an in vitro propagation system before this problem arises could ensure continued productivity by guaranteeing the availability of high sanitary quality plant material.
Our findings are in agreement with previous research emphasizing the importance of growth regulators in hop shoot tip cultivation. The addition of BAP and GA3 was crucial for shoot differentiation and regeneration from shoot tips, which could interfere with the differentiation and regeneration process of these tissues into shoots. In this regard, Adams [36] reported the necessity of using growth regulators such as BAP and GA3 for successful regeneration of hop meristems. Similarly, the observed events of rhizogenesis obtained in the absence of auxins and callogenesis could be associated with endogenous auxin levels and the balance between endogenous hormones and added growth regulators, respectively [10,52,53,54,55,56,57]. These results highlight the complexity of plant tissue responses to hormonal stimuli and underscore the importance of adjusting cultivation conditions for each type of explant, as well as the concentration and combination of growth regulators, aspects that could significantly influence the success of the technique and the obtaining of complete plants for clonal multiplication.
Moreover, successful rooting of hop uninodal segments was achieved through direct organogenesis, without the addition of plant growth regulators to the culture medium. These findings are of great importance, as the absence of callogenesis limits somaclonal variation events, a desirable feature to avoid variability and ensure clonal multiplication of female hop plants. Our results are consistent with previous research that has reported direct rhizogenesis of nodal segments of this species [58]. However, we found some discrepancies with other authors regarding the combination and concentration of auxins required to induce rhizogenesis [22,32]. Additionally, differences were observed in the time required to obtain a complete plant from nodal segments compared to shoot tip cultivation, which can be attributed to both, the inherent characteristics of the tissues and the hormonal concentration of the explants used and to the mapuche cultivar specific properties. While nodal segments rooting offers the potential to large-scale production in a reduced space and time, the shoot tip method may require a considerably longer period to achieve comparable results; however, it offers the additional advantage of providing phytosanitary quality material. These differences in development times can have significant implications in planning large-scale hop production, depending on the demands and specific objectives of each hop producer.
The results of ex vitro plant acclimatization showed that foliar application of ABA reduces plant stress, improving growth, green leaf number and reducing the number of senescent leaves. However, survival rate was not affected. These findings are consistent with previous studies reporting the beneficial effects of ABA use on plant growth and development during acclimatization in Ulmus minor [21]. Additionally, this phytohormone could attenuate leaf stress, reducing senescence, which would have beneficial effects on subsequent plant development. Our results regarding ABA application in ex vitro acclimatization underscore the importance of considering post-culture management to ensure a successful plant adaptation to the ex vitro environment.
Regarding synthetic seed production, we observed that the cold treatment had a negative impact on the conversion process as the duration of exposure to cold increased, consistent with findings described by Engelmann [59]. However, the percentages of viable explants, close to 100%, indicate that low temperatures did not significantly affect the final survival of the obtained seedlings. In this regard, Sharma et al. [60] pointed out that the critical factors for achieving successful commercialization of this technique and high viability of the synthetic seeds are the storage conditions and the storage period during preservation and transport. It should be noted that the encapsulation methodologies of nodal segments proposed here have previously been applied to shoot tips, resulted in higher conversion percentages for seeds stored for one and two weeks in cold (90%), compared to what was observed in this study [38]. However, shoot tip rooting percentages were lower (between 0 and 10%), compared to those obtained in this study (between 26.67% to 46.67%). These studies suggest that nodal segments are the most suitable explants for encapsulation, compared to shoot tips, since they showed lower susceptibility to treatment and a greater capacity to develop and produce a complete plant, even after a period of cold storage. Although other authors have reported successful encapsulation in other hop varieties [26,37], they did not evaluate conservation at 4°C and also required matrices supplemented with growth regulators or sucrose, aspects that increase the costs of the procedures, compared to our study.
Taken together, our results support the importance of developing appropriate cultivation and management protocols to enhance the efficiency and viability of hop plant production, from obtaining and surviving complete plants to synthetic seeds production. In particular, we have demonstrated that shoot tip cultivation offers an efficient way to propagate plants of high phytosanitary quality, whereas the use of nodal segments offers a rapid and economical alternative for clonal multiplication.
Furthermore, our results on synthetic seeds production reveal both challenges and opportunities in short and medium-term conservation, as well as in the efficiency of hop seed production. The application of these methodologies not only ensures a constant supply of female plants through cloning but also facilitates the transportation of plant material and germplasm exchange. This contributes to diversifying current production matrices and enhancing the availability of quality raw materials used in the brewing industry.
Finally, our findings provide a comprehensive perspective on the possibilities and challenges in hop in vitro multiplication. This comes at a crucial time when the adoption of advanced technologies can enhance productivity and sustainability in the agricultural sector, addressing the increasing demand and global climate challenges.
4. Materials and Methods
4.1. Plant Material and Explant Conditioning
Humulus lupulus L. var. mapuche, was used as a source of explants. The plants were kindly provided by a hop farm located in the province of Río Negro, Argentina (40° 48' 17.236'' S; 63° 9' 10.584'' W). Uninodal cuts were obtained from these plants to begin the plant tissue culture.
The plant material underwent a disinfection process involving immersion in 70% v/v ethanol (5 minutes), followed by 20 minutes immersion in a solution of 10% v/v commercial bleach (containing 35 gCl/l) and Tween 20® (0.01% v/v). Subsequently, it was rinsed several times with autoclaved distilled water. Finally, the shoot tips were isolated, which contained the meristems (0.8 ± 0.3 mm long) with the meristematic dome and two leaf primordia. Additionally, uninodal segments of two-month-old in vitro plants were utilized for rooting assays.
4.1.1. Culture Conditions
The shoot tips were incubated in glass tubes (40 ml) whereas the nodal segments were cultivated in glass jar (225 ml). In both cases, 5 explants per container were placed. On the other hand, the synthetic seeds were stored at 4°C in the fridge in petri dishes, which contained 10 synthetic seeds per dish. Then, when synthetic seeds were cultivated in a growth room, glass jar (450 ml) was used.
The pH of each medium was adjusted to 5,8 with NaOH or HCl. The culture was incubated in a growth room at 21±2°C according to the optimal thermal requirements reported by Nievas et al. [61], with a photoperiod of 16 h (50 μmol/m2seg from white fluorescent lamps).
4.2. Induction of Morphogenic Responses
4.2.1. Shoot Tip Cultivation
The explants were cultivated in sterile Murashige and Skoog [62] medium at a half concentration (½ MS), solidified with Sigma agar (0.65%) and added with sucrose (3%) and different plant growth regulators (PGR; auxins, cytokinins and gibberellin). The controls were established in their absence.
For shoot tips, culture medium was supplemented with PGR according to the methodology proposed for this species by Adams [36]. It comprised ½ MS without growth regulators (M1), and supplemented with 0.88 µM BAP (M2) and 2.89 µM of GA3 (M3). After one month, the following organogenic responses were assessed: presence of new shoots and/or its elongation, appearance of callus and root formation.
Each treatment was composed of 15 explants and the experiments were repeated three times.
4.2.2. Rooting Unimodal Segments
The uninodal segments intended for the rooting test were isolated from 2-month-old in vitro plants. Maintaining sterile conditions, the explants were extracted from the middle part of the seedling, and the leaves were removed. These segments were cultivated in ½ MS solidified with Sigma agar (0.65%), and supplemented with sucrose (3%) and IBA and IAA in various concentrations and combinations, as suggested by Roy et al. [22]. Four treatments were conducted: T1 (without PGR), T2 (IAA 5.7 µM), T3 (IBA 4.9 µM), and T4 (IAA 5.7 µM + IBA 4.9 µM). After two months of culture, the presence of callus, root formation, and the regeneration pathway (direct or indirect organogenesis) were evaluated.
Each treatment was composed of 15 explants and the experiments were repeated three times.
4.3. Acclimatization to Ex Vitro Conditions and ABA Treatment
The complete seedlings obtained from the in vitro culture of shoot tips and uninodal segments initiated the acclimatization process following the methodologies suggested by Dias et al. [21] and Pizzio et al. [63] with modifications. Briefly, the day before transplanting into soil, the seedlings (n=15) were treated with ABA solution at concentrations of 50 µM and 100 µM via spraying on adaxial and abaxial sides of leaves, whereas those mock treated served as the control group. Subsequently, the plants (N=45) were transferred to pots (220 ml) filled with Growmix ® sustrate and covered with a film, then incubated at room temperature with a photoperiod of 16 hours. Over the initial three weeks period, the containers were gradually opened, and after this duration, the film was completely removed. The parameters assessed included plant survival, attained height, and the number of green and senescent leaves. Finally, the surviving plants were transplanted into 0.5 liter pots and maintained in an outdoors environment. Three biological replicate were performed with similar results.
4.4. Synthetic Seeds
Uninodal segments of in vitro plants were encapsulated in a 2% (w/v) sodium alginate matrix, which was sterilized before, and then collected in drops using a micropipette. Subsequently, they were immersed in a 100 mM calcium chloride (CaCl2) solution for 3 to 5 minutes under constant agitation. Finally, the synthetic seeds obtained were placed on sterile tissue paper to remove excess CaCl2.
To determine the suitable medium for its culture, the synthetic seeds were cultivated in: ½ MS without PGR and ½ MS supplemented with BAP 0.88 µM and GA3 2.89 µM. Both were solidified with Sigma agar (0.65%), and supplemented with sucrose (3%). The cultures were maintained under controlled light and temperature conditions (16 h light, 21±2ºC).
Additionally, in order to assess the effects of short-term cold storage, synthetic seeds were stored for one, two and three weeks at 4ºC in the dark (TA, TB and TC, respectively), under controlled light and temperature conditions (16 h light,). During cold storage, the synthetic seeds were placed on agarose medium, whereas for conversion, they were transferred to ½ MS solidified with Sigma agar (0.65%), and supplemented with sucrose (3%). The synthetic seeds which were not subjected to storage at 4°C and were directly incubated at 21±2ºC served as the control group (T0).
Each treatment was comprised three replicates of 10 synthetic seeds, and observations were conducted over a three-week period to asses root appearance, percentage of viable explants, where viability was considered when the encapsulated explants presented a green appearance, without yellowing and without necrotic parts [64], and percentage of synthetic seeds conversion, considered as the percentage of synthetic seeds where the encapsulation matrix ruptured, leading to the emergence of shoots [42].
4.5. Statistical Analysis
All data were subjected to analysis of variance and comparisons of means were made with the least significant difference test at the 5% level of probability. The statistical difference between treatments were performed using one way ANOVA. Those data that did not present a normal distribution were analyzed using the Kruskal-Wallis test and suitable non-parametric post-hoc Tukey's test.
Author Contributions
Conceptualization, Gastón Pizzio and Patricia Boeri; Investigation, Luciana Di Sario, María Zubillaga, Claudio Zepeda Moreno, Gastón Pizzio and Patricia Boeri; Methodology, Luciana Di Sario, Gastón Pizzio and Patricia Boeri; Project administration, Patricia Boeri; Supervision, Gastón Pizzio and Patricia Boeri; Writing – original draft, Luciana Di Sario and Patricia Boeri; Writing – review & editing, Luciana Di Sario, María Zubillaga, Claudio Zepeda Moreno, Gastón Pizzio and Patricia Boeri.
Funding
This research received no external funding.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dodds, K. Hops—A Guide for New Growers; NSW Department of Primary Industries: Orange, NSW, Australia, 2017; ISBN 978-1-76058-007-0. [Google Scholar]
- Mozny, M.; Trnka, M.; Vlach, V.; Zalud, Z.; Cejka, T.; Hajkova, L. ... Büntgen, U. Climate-induced decline in the quality and quantity of European hops calls for immediate adaptation measures. Nat. Commun. 2023, 14, 6028. [Google Scholar] [CrossRef]
- El cambio climático podría duplicar el coste de una cerveza. Available online: https://ee-ip.org/es/article/el-cambio-climatico-podria-duplicar-el-coste-de-una-cerveza-1187 (accesses on 25 February 2024).
- Potopová, V.; Lhotka, O.; Možný, M.; Musiolková, M. Vulnerability of hop-yields due to compound drought and heat events over European key-hop regions. Int. J. Climatol. 2021, 41, E2136–E2158. [Google Scholar] [CrossRef]
- Agehara, S.; Acosta-Rangel, A.; Gallardo, M.; Vallad, G. Selection and Preparation of Planting Material for Successful Hop Production in Florida: HS1381, 9/2020. EDIS: Gainesville, FL, 2020; Volume 2020 (5). [CrossRef]
- Eastwell, K.C.; Barbara, D.J. Apple Mosaic Virus. In Field Guide for Integrated Pest Management in Hops, 3rd ed.; O‘Neal, S.D., Walsh, D.B., Gent, D.H., Eds.; U.S. Hop Industry Plant Protection Committee: Pullman, WA, 2015; Volume 1, p. 39. [Google Scholar]
- 7. Eastwell, K. C.; Barbara, D.J. Carlavirus Complex: American Hop Latent Virus, Hop Latent Virus, and Hop Mosaic Virus. In Field Guide for Integrated Pest Management in Hops, 3rd ed.; O‘Neal, S.D., Walsh, D.B., Gent, D.H., Eds.; U.S. Hop Industry Plant Protection Committee: Pullman, WA, 2015; Volume 1, p. 38. [Google Scholar]
- Gent, D.H.; Johnson, D.A.; Gevens, A.J.; Hausbeck, M.K. Downy Mildew. In Field Guide for Integrated Pest Management in Hops, 3rd ed.; O‘Neal, S.D., Walsh, D.B., Gent, D.H., Eds.; U.S. Hop Industry Plant Protection Committee: Pullman, WA, 2015; Volume 1, pp. 15–21. [Google Scholar]
- Gent, D.H.; Nelson, M.E.; Gadoury, D.M.; Gevens, A.J.; Hausbeck, M.K. Powdery Mildew. In Field Guide for Integrated Pest Management in Hops, 3rd ed.; O‘Neal, S.D., Walsh, D.B., Gent, D.H., Eds.; U.S. Hop Industry Plant Protection Committee: Pullman, WA, 2015; Volume 1, pp. 25–29. [Google Scholar]
- Levitus, G.; Echenique, V.; Rubinstein, C.; Hopp, E.; Mroginski, L. Biotecnología y mejoramiento vegetal II, 2nd ed.; Ediciones Instituto Nacional de Tecnología Agropecuaria (INTA): Buenos Aires, Argentina, 2010; p. 650. [Google Scholar]
- Gent, D.H.; Nelson, M.E.; George, A.E.; Grove, G.G.; Mahaffee, W.F.; Ocamb, C.M. . Turechek, W.W. A decade of hop powdery mildew in the Pacific Northwest. Plant health prog. 2008, 9, 33. [Google Scholar] [CrossRef]
- Faragó, J.; Psenácová, I.; Faragová, N. The use of biotechnology in hop (Humulus lupulus L.) improvement. Nova Biotech. 2009, 9, 279–293. [Google Scholar] [CrossRef]
- Osório, M.L.; Gonçalves, S.; Coelho, N.; Osório, J.; Romano, A. Morphological, physiological and oxidative stress markers during acclimatization and field transfer of micropropagated Tuberaria major plants. PCTOC 2013, 115, 85–97. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, G.; Santos, C. Acclimatization of micropropagated plantlets induces an antioxidative burst: a case study with Ulmus minor Mill. Photosynthetica, 2011, 49, 259–266. [Google Scholar] [CrossRef]
- Pospíšilová, J.; Synková, H.; Haisel, D.; Baťková, P. Effect of abscisic acid on photosynthetic parameters during ex vitro transfer of micropropagated tobacco plantlets. Biol. Plant. 2009, 53, 11–20. [Google Scholar] [CrossRef]
- Aguilar, M.L.; Espadas, F.L.; Coello, J.; Maust, B.E.; Trejo, C.; Robert, M.L.; Santamaria, J.M. The role of abscisic acid in controlling leaf water loss, survival and growth of micropropagated Tagetes erecta plants when transferred directly to the field. J. Exp. Bot. 2000, 51, 1861–1866. [Google Scholar] [CrossRef]
- Santamaria, J.M.; Davies, W.J.; Atkinson, C.J. Stomata of micropropagated Delphinium plants respond to ABA, CO2, light and water potential, but fail to close fully. J. Exp. Bot. 1993, 44, 99–107. [Google Scholar] [CrossRef]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Aroca, R.; Vernieri, P.; Ruiz-Lozano, J.M. Mycorrhizal and non-mycorrhizal Lactuca sativa plants exhibit contrasting responses to exogenous ABA during drought stress and recovery. J. Exp. Bot. 2008, 59, 2029–2041. [Google Scholar] [CrossRef]
- Wasilewska, A.; Vlad, F.; Sirichandra, C.; Redko, Y.; Jammes, F.; Valon, C. ... Leung, J. An update on abscisic acid signaling in plants and more…. Mol. Plant 2008, 1, 198–217. [Google Scholar] [CrossRef]
- Dias, M.C.; Correia, C.; Moutinho-Pereira, J.; Oliveira, H.; Santos, C. Study of the effects of foliar application of ABA during acclimatization. PCTOC 2014, 117, 213–224. [Google Scholar] [CrossRef]
- Roy, A.T.; Leggett, G.; Koutoulis, A. Development of a shoot multiplication system for hop (Humulus lupulus L.). In Vitro Cell. Dev. Biol.-Plant 2001, 37, 79–83. [Google Scholar] [CrossRef]
- Smýkalová, I.; Ortová, M.; Lipavská, H.; Patzak, J. Efficient in vitro micropropagation and regeneration of Humulus lupulus on low sugar, starch-Gelrite media. Biol. Plant. 2001, 44, 7–12. [Google Scholar] [CrossRef]
- Mafakheri, M.; Hamidoghli, Y. Micropropagation of hop (Humulus lupulus L.) via shoot tip and node culture. In IV International Humulus Symposium 1236, 1st. ed.; Matthews, P., Stevens, F., Eds.; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2019; Volume 1, pp. 154–196. [Google Scholar]
- Trojak-Goluch, A.; Kawka, M.; Czarnecka, D. The effects of explant source and hormone content on plant regeneration and induction of tetraploids in Humulus lupulus L. In Vitro Cell. Dev. Biol.-Plant, 2015, 51, 152-159. [CrossRef]
- Liberatore, C.M.; Rodolfi, M.; Beghè, D.; Fabbri, A.; Ganino, T.; Chiancone, B. Adventitious shoot organogenesis and encapsulation technology in hop (Humulus lupulus L.). Sci. Hortic. 2020, 270, 109416. [Google Scholar] [CrossRef]
- Clapa, D.; Hârța, M. Establishment of an efficient micropropagation system for Humulus lupulus L. cv. cascade and confirmation of genetic uniformity of the regenerated plants through dna markers. Agronomy. 2021, 11, 2268. [Google Scholar] [CrossRef]
- Hirakawa, T.; Tanno, S.; Tanno, S. In vitro propagation of Humulus lupulus through the induction of axillary bud development. Plants 2022, 11, 1066. [Google Scholar] [CrossRef]
- Lagos, F.S.; Zuffellato-Ribas, K.C.; Deschamps, C. Vegetative propagation of hops (Humulus lupulus L.): historical approach and perspectives. Semina: Cien. Agra. 2022, 43, 1373–1394. [Google Scholar] [CrossRef]
- Di Sario, L. Evaluación del cultivo in vitro de variedades nacionales de lúpulo (Humulus lupulus) como una alternativa a la propagación agámica tradicional. Tesis de grado, Universidad Nacional de Río Negro, Villa Regina (Río Negro, Argentina), 2023. http://rid.unrn.edu.ar/handle/20.500.12049/10188. 1204. [Google Scholar]
- Ha, T.H.; Lyu, J.I.; Lee, J.H.; Ryu, J.; Park, S.H.; Kang, S.Y. Studies on growth characteristics and propagation method of introduced hop (Humulus lupulus L.) cultivars. Korean J. Plant Res. 2023, 36, 181–190. [Google Scholar] [CrossRef]
- Iacuzzi, N.; Salamone, F.; Farruggia, D.; Tortorici, N.; Vultaggio, L.; Tuttolomondo, T. Development of a New Micropropagation Protocol and Transfer of In Vitro Plants to In Vivo Conditions for Cascade Hop. Plants 2023, 12, 2877. [Google Scholar] [CrossRef]
- Batista, D.; Sousa, M.J.; Pais, M.S. Plant regeneration from stem and petiole-derived callus of Humulus lupulus L.(Hop) clone Braganca and var. Brewer’s Gold. In Vitro–Plant 1996, 32, 37–41. [Google Scholar] [CrossRef]
- Gurriarán, M.J.; Revilla, M.A.; Tamés, R.S. Adventitious shoot regeneration in cultures of Humulus lupulus L.(hop) cvs. Brewers Gold and Nugget. Plant Cell Rep. 1999, 18, 1007–1011. [Google Scholar] [CrossRef]
- Peredo, E.L.; Revilla, M.Á.; Arroyo-García, R. Assessment of genetic and epigenetic variation in hop plants regenerated from sequential subcultures of organogenic calli. J. Plant Physiol. 2006, 163, 1071–1079. [Google Scholar] [CrossRef]
- Adams, A.N. Elimination of viruses from the hop (Humulus lupulus) by heat therapy and meristem culture. J. Hortic. Sci. 1975, 50, 151–160. [Google Scholar] [CrossRef]
- Martinez, D.; Tamés, R.S.; Angeles Revilla, M. Cryopreservation of in vitro-grown shoot-tips of hop (Humulus lupulus L.) using encapsulation/dehydration. Plant Cell Rep. 1999, 19, 59–63. [Google Scholar] [CrossRef]
- Di Sario, L.; Zubillaga, M.F.; Sharry, S.E.; Boeri, P.A. Semillas sintéticas: una estrategia para la conservación de lúpulo (Humulus lupulus L.). In Biotecnología productiva y sostenible, 1st ed; Gutiérrez Mora, A., Ed.; Editorial CIATEJ: Guadalajara, México, 2023; Volume 1, pp. 101–112. [Google Scholar]
- Danso, K.E.; Ford-Lloyd, B.V. Encapsulation of nodal cuttings and shoot tips for storage and exchange of cassava germplasm. Plant Cell Rep. 2003, 21, 718–725. [Google Scholar] [CrossRef]
- Naik, S.K.; Chand, P.K. Nutrient-alginate encapsulation of in vitro nodal segments of pomegranate (Punica granatum L.) for germplasm distribution and exchange. Sci. Hortic., 2006, 108, 247–252. [Google Scholar] [CrossRef]
- Rai, M.K.; Jaiswal, V.S.; Jaiswal, U. Alginate-encapsulation of nodal segments of guava (Psidium guajava L.) for germplasm exchange and distribution. J. Hortic. Sci. Biotechnol. 2008, 83, 569–573. [Google Scholar] [CrossRef]
- Ray, A.; Bhattacharya, S. Storage and plant regeneration from encapsulated shoot tips of Rauvolfia serpentina—an effective way of conservation and mass propagation. S AFR J BOT 2008, 74, 776–779. [Google Scholar] [CrossRef]
- Rai, M.K.; Asthana, P.; Singh, S.K.; Jaiswal, V.S.; Jaiswal, U. The encapsulation technology in fruit plants—a review. Biotechnol. Adv. 2009, 27, 671–679. [Google Scholar] [CrossRef]
- Parveen, S.; Shahzad, A. Encapsulation of nodal segments of Cassia angustifolia Vahl. for short-term storage and germplasm exchange. Acta Physiol. Plant. 2014, 36, 635–640. [Google Scholar] [CrossRef]
- Ahmed, M.R.; Anis, M.; Al-Etta, H.A. Encapsulation technology for short-term storage and germplasm exchange of Vitex trifolia L. Rend. Fis. Acc. Lincei 2015, 26, 133–139. [Google Scholar] [CrossRef]
- Gantait, S.; Kundu, S.; Ali, N.; Sahu, N.C. Synthetic seed production of medicinal plants: a review on influence of explants, encapsulation agent and matrix. Acta Physiol. Plant. 2015, 37, 98. [Google Scholar] [CrossRef]
- Alatar, A.A.; Ahmad, N.; Javed, S.B.; Abdel-Salam, E.M.; Basahi, R.; Faisal, M. Two-way germination system of encapsulated clonal propagules of Vitex trifolia L.: an important medicinal plant. J. Hortic. Sci. Biotechnol. 2017, 92, 175–182. [Google Scholar] [CrossRef]
- Rihan, H.Z.; Kareem, F.; El-Mahrouk, M.E.; Fuller, M.P. Artificial seeds (principle, aspects and applications). Agronomy 2017, 7, 71. [Google Scholar] [CrossRef]
- Sharma, A.; Kaur, R.; Sharma, N. In vitro morphogenic response of different explants of Gentiana kurroo Royle from Western Himalayas—an endangered medicinal plant. Physiol. Mol. Biol. Plants 2014, 20, 249–256. [Google Scholar] [CrossRef]
- Hasan, S.M.Z.; Takagi, H. Alginate-coated nodal segments of yam (Dioscorea spp.) for germplasm exchange and distribution. Plant Genet. Resour. Newsl. 1995, 103, 32–35. [Google Scholar]
- Pattnaik, S.; Chand, P.K. Morphogenic response of the alginate-encapsulated axillary buds from in vitro shoot cultures of six mulberries. PCTOC 2000, 60, 177–185. [Google Scholar] [CrossRef]
- Soneji, J.; Rao, P.; Mhatre, M. Germination of synthetic seeds of pineapple (Ananas comosus L. Merr.). Plant Cell Rep. 2002, 20, 891–894. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin–cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Fortes, A.M.; Pais, M.S. Organogenesis from internode-derived nodules of Humulus lupulus var. Nugget (Cannabinaceae): histological studies and changes in the starch content. Am. J. Bot. 2000, 87, 971–979. [Google Scholar] [CrossRef]
- Villarreal, B. ¿Cómo se forman las nuevas plantas in vitro? - Morfogénesis in vitro. Organogénesis. Embriogénesis somática. In Plantas de probeta, 1st ed.; Sharry, S., Adema, M., Abedini, W., Eds.; Editorial de la Universidad de La Plata: La Plata, Argentina, 2015; Volume 1, pp. 92-97.
- Segura, J. Introducción al desarrollo. Concepto de hormona vegetal. In Fundamentos de fisiología vegetal, 2nd ed.; Azcon, J., Talon, M., Eds.; McGraw-Hill Interamericana de España: Barcelona, España, 2008; pp. 349–376. [Google Scholar]
- Krikorian, A.D.; Kelly, K.; Smith, D.L. Hormones in Tissue Culture and Micro-Propagation. In Plant Hormones and their Role in Plant Growth and Development; Davies, P.J., Eds.; Springer, Dordrecht, 1987; pp.
- Peredo, E.L.; Arroyo-Garcia, R.; Revilla, M.Á. Epigenetic changes detected in micropropagated hop plants. J. Plant Physiol. 2009, 166, 1101–1111. [Google Scholar] [CrossRef]
- Engelmann, F. In vitro conservation of tropical plant germplasm - a review. Euphytica 1991, 57, 227–243. [Google Scholar] [CrossRef]
- Sharma, S.; Shahzad, A.; da Silva, J.A.T. Synseed technology—a complete synthesis. Biotechnol. Adv. 2013, 31, 186–207. [Google Scholar] [CrossRef]
- Nievas, W.E.; Villarreal, P.; Rosati, A.; Rodriguez, A.B.; Lago, J. El cultivo del lúpulo. Aspectos agroambientales y económicos para el Alto Valle del río Negro, 1st ed.; Ediciones Instituto Nacional de Tecnología Agropecuaria (INTA): Buenos Aires, Argentina, 2021; p. 82. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Pizzio, G.A.; Mayordomo, C.; Lozano-Juste, J.; Garcia-Carpintero, V.; Vazquez-Vilar, M.; Nebauer, S.G. ... Rodriguez, P.L. PYL1-and PYL8-like ABA receptors of Nicotiana benthamiana play a key role in ABA response in seed and vegetative tissue. Cells 2022, 11, 795. [Google Scholar] [CrossRef]
- Micheli, M.; Hafiz, I.A.; Standardi, A. Encapsulation of in vitro-derived explants of olive (Olea europaea L. cv. Moraiolo): II. Effects of storage on capsule and derived shoots performance. Sci. Hortic. 2007, 113, 286–292. [Google Scholar] [CrossRef]
Figure 1.
Morphogenetic responses observed in shoot tip cultivation after four weeks of incubation in: (a) ½ MS; (b) ½ MS + BAP (0.88 µM); (c) ½ MS + GA3 (2.89 µM).
Figure 1.
Morphogenetic responses observed in shoot tip cultivation after four weeks of incubation in: (a) ½ MS; (b) ½ MS + BAP (0.88 µM); (c) ½ MS + GA3 (2.89 µM).

Figure 2.
Rooting observed after six weeks in nodal segment cultivation: (a) Direct organogenesis; (b) Indirect organogenesis.
Figure 2.
Rooting observed after six weeks in nodal segment cultivation: (a) Direct organogenesis; (b) Indirect organogenesis.

Figure 3.
Acclimatization and rustication of in vitro plants: (a) Plant after 21 days in ex vitro enviroment; (b) Plant after six months in ex vitro enviroment.
Figure 3.
Acclimatization and rustication of in vitro plants: (a) Plant after 21 days in ex vitro enviroment; (b) Plant after six months in ex vitro enviroment.

Figure 4.
Phenotypes evaluated in the acclimatization and rustication in plant after 21 days in ex vitro enviroment: (a) A representative mock plant; (b) Sprayed with ABA 50µM; and (c) ABA 100µM; quantification of (d) Plant growth; (e) Number of green leaves; (f) Number of senescent leaves. The asterisk indicates: * p ≤ 0.05 (Student’s t-test or ANOVA followed by Dunnet’s test).
Figure 4.
Phenotypes evaluated in the acclimatization and rustication in plant after 21 days in ex vitro enviroment: (a) A representative mock plant; (b) Sprayed with ABA 50µM; and (c) ABA 100µM; quantification of (d) Plant growth; (e) Number of green leaves; (f) Number of senescent leaves. The asterisk indicates: * p ≤ 0.05 (Student’s t-test or ANOVA followed by Dunnet’s test).
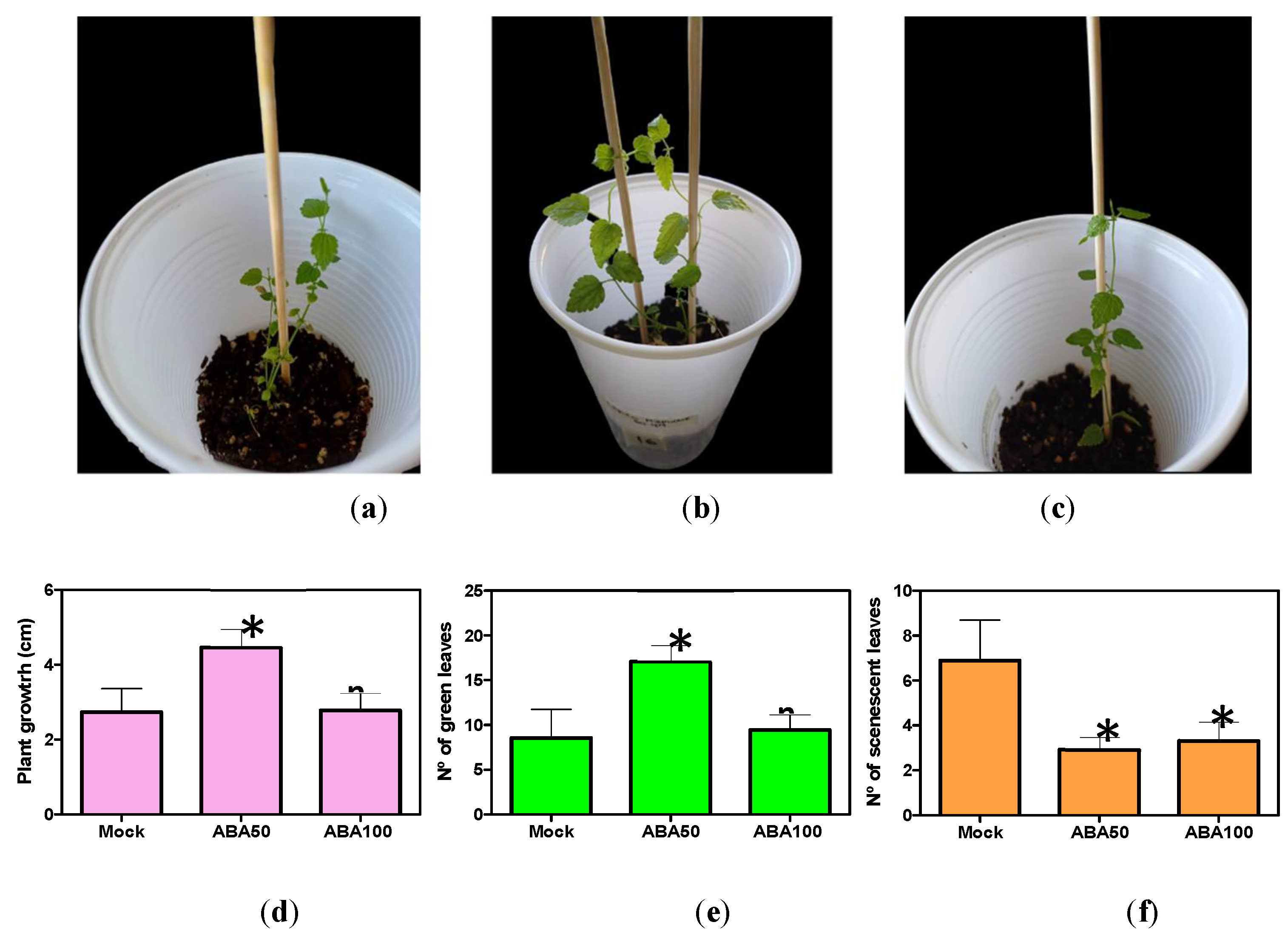
Figure 5.
Synthetic seeds: (a) Conversion at seven days after incubation at 21±2°C; (b) Complete plant after four weeks incubation at 21±2°C. .
Figure 5.
Synthetic seeds: (a) Conversion at seven days after incubation at 21±2°C; (b) Complete plant after four weeks incubation at 21±2°C. .
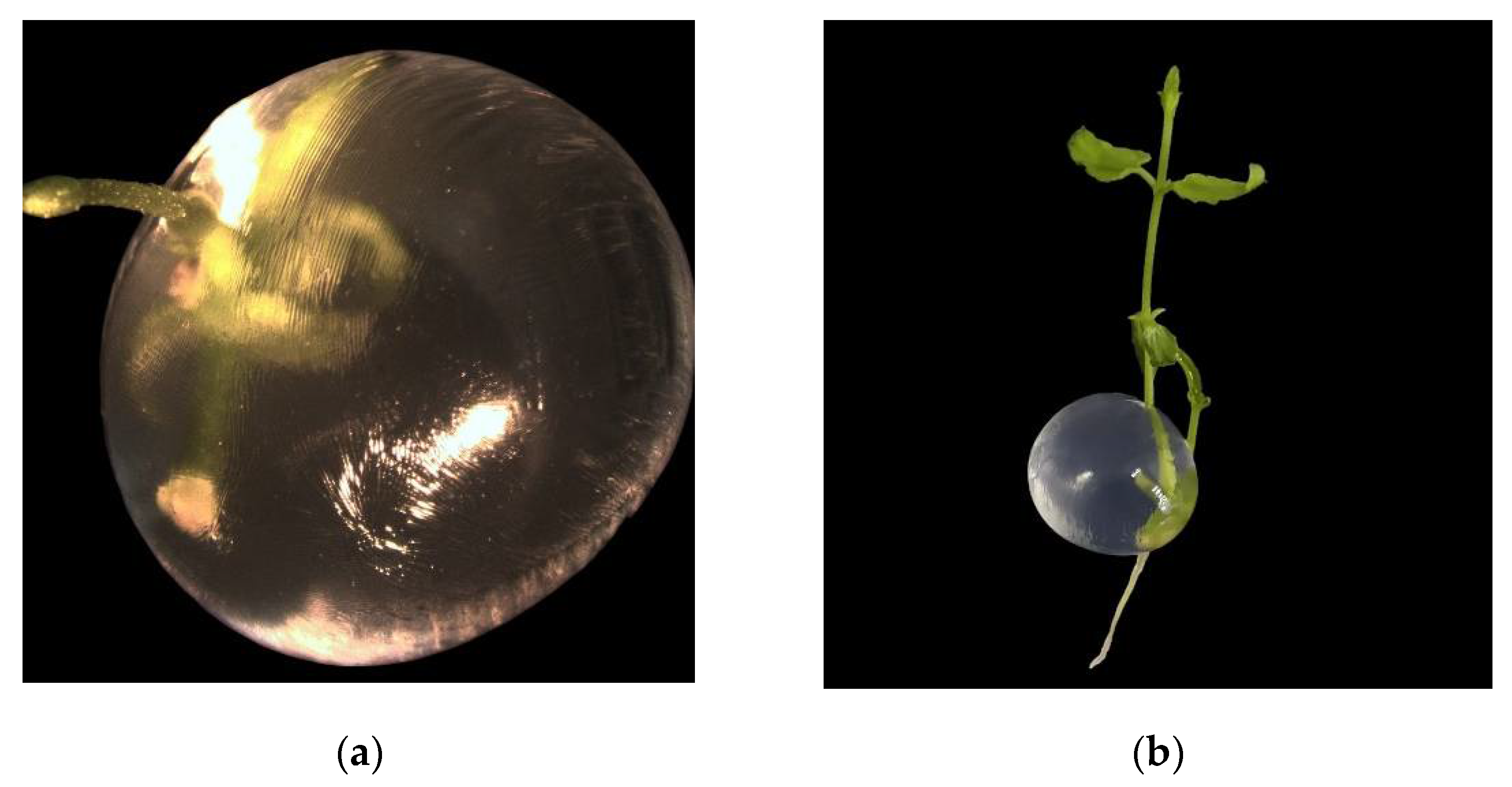

Table 1.
Effects of plant growth regulators on meristem differentiation and shoot elongation.
| Culture medium | BAP (µM) | GA3 (µM) | Regenerated shoot tips [%] |
Elongation [%] |
|---|---|---|---|---|
| M1 | 0 | 0 | 0 ± 0 a | 0 ± 0 a |
| M2 | 0.88 | 0 | 75.56 ± 12.2 b | 42.22 ± 9.8 b |
| M3 | 0 | 2.89 | 86.67 ± 10 b | 35.56 ± 10.6 b |
Data are expressed as the mean ± standard deviation (SD). Values with the same letter within the same column are not significantly different (p > 0.05) according to the Kruskal-Wallis test. M1: ½ MS; M2: ½ MS + BAP (0.88 µM); M3: ½ MS + GA3 (2.89 µM).
Table 2.
Morphogenic responses evaluated in the culture of uninodal segments.
| Morphogenetic responses (%) | ||||||
|---|---|---|---|---|---|---|
| IAA (µM) | IBA (µM) | Rooting | Direct rooting | Indirect rooting | Callus | |
| T1 | 0 | 0 | 91.11 ± 8.45 b | 100 ± 0 b | 0 ± 0 a | 0 ± 0 a |
| T2 | 5.7 | 0 | 60 ± 14.14 a | 32.41 ± 14.7 a | 67.59 ± 14.7 b | 84.44 ± 8.82 b |
| T3 | 0 | 4.9 | 57.78 ± 21.08 a | 45.37 ± 27.99 a | 54.63 ± 27.99 b | 91.11 ± 8.53 bc |
| T4 | 5.7 | 4.9 | 57.78 ± 15.63 a | 22.22 ± 18.16 a | 77.78 ± 18.16 b | 100 ± 0 c |
Data are expressed as the mean ± standard deviation (SD). Values with the same letter within the same column are not significantly different (p > 0.05) according to the Kruskal-Wallis test.
Table 3.
Responses observed in the production of synthetic seeds stored at 4°C.
| Evaluated responses (%) | ||||
|---|---|---|---|---|
| Week(s) at 4°C | Conversion | Rooting | Viability | |
| T0 | 0 (control) | 90 ± 10 c | 56.67 ± 12.11 c | 100 ± 0.00 b |
| TA | 1 | 76.67 ± 16.33 bc | 46.67 ± 19.66 bc | 86.67 ± 5.16 a |
| TB | 2 | 66.67 ± 15.06 ab | 26.67 ± 5.16 ab | 90.00 ± 0.00 a |
| TC | 3 | 46.67 ± 8.16 a | 33.33 ± 12.11 a | 86.67 ± 5.16 a |
Data are expressed as the mean ± standard deviation (SD). Values with the same letter within the same column are not significantly different (p > 0.05) according to the Kruskal-Wallis test.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



