
Submitted:
28 May 2024
Posted:
29 May 2024
You are already at the latest version
Abstract
In the present study, we aimed to observed hair follicle development and describe Wnt10b/β-catenin signaling pathways related to hair follicle development in different ages of Rex rabbits. Six Rex rabbits (3 male and 3 female) of 15, 30, 60, 90, 120 and 150 days of age were selected and skin samples were collected for hair follicle development and Wnt/β-catenin signaling pathway genes expresion detection. The results showed that the the values for body weight, coat length, skin thickness, skin area, and skin weight of Rex rabbits were increased significantly with age (p < 0.05). The total hair follicle density, secondary hair follicle density and the ratio of the secondary hair follicle density to the primary hair follicle density in 120 days of Rex rabbits was higher than that in the other ages (p < 0.05), and the major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation. Furthermore, the expression of the Wnt10b, CTNNB1, adenomatosis polyposis protein (APC), Disheveled 2 (DVL2), glycogen synthase kinase 3β(GSK-3β), Lymphoid enhancer-binding factor 1 (LEF1), dickkopf-1 (DKK1) and transforming growth factor beta 1 (TGFβ-1) genes in the skin tissue of Rex rabbits at different age was significantly differences (p < 0.05), and the expression of the Wnt10b, CTNNB1, DVL2, GSK-3β, and TGFβ-1 genes at 90 days was higher than that in the other days, the expression of the LEF1 and DKK1 genes at 60 days was higher than that in the other days. As western blotting indicated, the β-catenin protein phosphorylation (P-CTNNB1) levels in the skin tissue of Rex rabbits shows the trend of first increasing, then decreasing and then increasing, 60 days, 120 days and 150 days significantly higher than 15 days, 30 days or 90 days (p < 0.05). With the increasing of ages, the GSK-3β protein phosphorylation (P-GSK-3β) levels in the skin tissue of Rex rabbits were increasing, and maximum at 120 days of age (p < 0.05). These results show that ages could influence on the fur quality and hair follicle traits of Rex rabbits, and Wnt10b/β-catenin signaling pathways is involved in the biological process of periodic hair follicle development in Rex rabbits. Considering the rule of fur growth, the slaughter age of Rex rabbits should not be earlier than 120 days.
Keywords:
age
; rabbit
; Wnt10b/β-catenin
; fur quality
; hair follicle development
1. Introduction
Hair Follicle is a kind of adjunct organ of skin, which exists in dermis and subcutaneous tissue [1]. Hair follicles are the base of hair growth and provide hair fixation and protection, and wool animals providing important textile materials for human beings. The hair follicle of mammal after birth has the periodic regeneration function, including hair shaft active growth (anagen phase), apoptosis (catagen phase) and hair follicle relative static (telogen phase) 3 stages [2]. Hair follicles depend on periodic cycle growth to realize the growth and renewal of hair, and their periodic changes determine the periodic growth and shedding of hair, and the periodic development of hair follicles is affected by many factors, regulated by a series of signaling molecules [3–5]. The Wnt/β-catenin signaling pathway is a group of signal transduction pathways regulated by Wnt protein and β-catenin gene (CTNNB1), these proteins include Wnt, β-catenin, frizzled, dishevelled and low density lipoprotein receptor related protein 5/6 proteins (LRP5/6) [6]. In addition, a complex of glycogen synthase kinase 3β (GSK-3β), adenomatosis polyposis protein (APC), axin and casein kinase 1 also plays a key regulatory role, the polyprotein complex can phosphorylate β-catenin residues and eventually mediate the degradation of β-catenin ubiquitination. Wnt protein ligands activate the Wnt signaling pathway by binding to receptors, and the Wnt-frizzled/lipoprotein receptor complex can induce β-catenin phosphorylation, thereby enabling β-catenin and T-cell factor (TCF)/ lymphoid enhancer factor (LEF) /β-catenin-mediated gene expression to regulate the proliferation and differentiation of hair follicle cells [7,8,9]. The canonical Wnt/β-catenin signaling pathway not only regulates embryonic growth and development, tumorigenesis and formation, but also plays a key regulatory role in hair follicle development and hair shaft growth [10,11,12]. Rex rabbits are important fur rabbits, while the effect of the Wnt/β-catenin signaling pathway on fur rabbits has rarely been reported, and the mechanism of Wnt/β-catenin signaling pathway influence on hair follicle development remains unclear. In this study, the gene expression and protein phosphorylation level of Wnt/β-catenin signaling pathway in the skin of different age from Rex rabbits was studied to determine the relationship between Wnt/β-catenin and hair follicle development. In addition, an appropriate slaughter age of Rex rabbits was also determined.
2. Materials and Methods
2.1. Experimental Design
Six Rex rabbits (3 male and 3 female) of 15, 30, 60, 90, 120 and 150 days of age under the same feed, the same house, the same environment were selected, and the experimental rabbits were sacrificed via cervical dislocation. The skin samples were collected and stored in a cryopreservation tube and 4% paraformaldehyde solution respectively. The expression of Wnt/β-catenin signaling pathway genes was detected by fluorescence after reverse transcription, and changes of β-catenin and GSK-3β phosphorylation level. The fixed samples were made into paraffin sections, stained by hematoxylin-eosin (HE) staining, the longitudinal samples were used to observe the hair follicle location period, and the transverse samples were used to count the hair follicle density.
2.2. Determination of Indicators and Methods
2.2.1. Body Weight and Visceral Development
The experimental rabbits were fasted 12 hours prior to slaughter and weighed. Before slaughter, the animals were stunned by electric shock and then slaughtered by bloodletting, the heart, lung, liver and kidney were carefully peel and also weighed, then heart index, lung index, liver index and kidney index were calculated by dividing their weights by the live weight before slaughter.
2.2.2. Fur Quality
The fur quality index in different ages of Rex rabbit detection method as described previously [13]. The adhesive tissue and fat was removed from fur, and weighed, then cut in the middle of the abdomen, and the length and width of the skin were measured using a soft ruler. The skin area was calculated by skin length*skin width. The thickness of the skin was measured using vernier calipers and the length of the coat was measured using a straightedge. The hair follicle density was observed with paraffin section and HE staining. After HE staining, photos were taken under ordinary light conditions with a microscope (Nikon, ECLIPSE 80i, Japan), and Image-Pro Plus 6.0 analysis software was used to calculate the hair follicle density (primary hair follicle density, secondary hair follicle density, and total hair follicle density), and the ratio of the secondary hair follicle density to the primary hair follicle density was also calculated.
2.2.3. RNA Isolation and Real-Time PCR Analysis
SYBR Green I method was used to detect gene expression in rabbits' skin tissues. Total RNA extraction and real-time PCR were performed as described previously [14]. RNA quality was determined by agarose gel electrophoresis and purity was assessed by nucleic acid spectrophotometer (DENOVIX, DS-11 Spectrophotometer, USA). The ratios of A260/A280 of all samples were in the range of 1.8-2.0, and 1.0% agarose gel electrophoresis with ethidium bromide (Sigma-Aldrich, Germany) staining detected good integrity of RNA bands. Sequences of primers in this experiment are shown in Table 1. Using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as the normalized gene, the mRNA relative amount of the gene was calculated by 2 delta-delta CT method [15].
2.2.4. Western Blotting
The methods for total protein extraction and sodium dodecyl sulfate (SDS) - polyacrylamide gel electrophoresis (PAGE) was performed according to Liu et al. [16]. Total protein was extracted from skin tissue using a radioimmunoprecipitation lysis buffer (Beyotime, Shanghai, China) and protein concentration was determined using the BCA Protein Assay Kit (Conwin, Beijing, China). The extracted protein (50 ng/sample) was dissolved in 40 mL of SDS loading buffer (Solarbio, Beijing, China) and then electrophoresed on a 12.5% SDS-PAGE gel (Bio-Rad, Richmond, USA) and transferred to a polyvinylidene fluoride membrane (Millipore, Billerica, USA). Protein molecular mass standard markers were purchased from Thermo (USA). The membranes were blocked with 5% skim milk in PBS (Solarbio, China) at 4 °C overnight and incubated with 1:1,000 dilution primary antibodies (ACTIN, Servicebio, Wuhan, China; P-CTNNB1, BIOSS, Beijing, China; P-GSK-3β antibody, Servicebio, Wuhan, China). The membranes were then washed with Tris-buffered saline containing Tween (Solarbio, Beijing, China) and incubated with a 1:3000 dilution of a horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG antibody (Servicebio, Wuhan, China) at 37 °C for 1 h. The proteins were visualized using Beyo ECL reagent (Beyotime, Shanghai, China). Band intensity was quantified using a AIWBwellTM Bioimage Analysis System (Servicebio, Wuhan, China). The relative content of samples is the ratio of index gray value to internal parameter gray value.
2.3. Data Processing
The SPSS 26.0 statistical software was used for the one-way ANOVA analysis of the data, and Duncan’s test was used for multiple comparisons. The results are presented as the mean value and standard error of means (SEM), and p < 0.05 was considered to be a significant difference.
3. Results
3.1. Growth and Development of Internal Organs
The effects of different ages on growth and development of internal organs in Rex rabbits are shown in Table 2. As indicated, the values for body weight, heart weight, lung weight, liver weight and kidney weight of Rex rabbits were increased significantly with age (p < 0.05), and the age of 150 days was significantly higher than that of 15 days, 30 days, 60 days or 90 days. However, heart index, lung index, and kidney index decreased with significantly age (p < 0.05), and the age of 15 days was higher than that in higher days of age (60 days, 90 days, 120 days or 150 days.). For the liver index, it increased first and then decreased with the increase of age, and reached the maximum at 60 days of age (p < 0.05).
3.2. Fur Quality
The effects of different ages on the fur quality of Rex rabbits are shown in Table 3. As indicated, the values for coat length, skin thickness, skin area, and skin weight of Rex rabbits were increased significantly with age (p < 0.05), and the age of 150 days was significantly higher than that of 15 days. However, there was no significant difference in relative weight of skin of Rex rabbits at different ages (p > 0.05).
3.3. Hair Follicle Development and Density
The hair follicle development in dorsal skin were observed via paraffin vertical sectioning and HE staining, and it was found that the major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation (Figure 1). Besides, the hair follicle density in dorsal skin were observed via paraffin transverse sectioning and HE staining is shown in Figure 2. After statistical analysis found that the total hair follicle density, secondary hair follicle density and the ratio of the secondary hair follicle density to the primary hair follicle density in 120 days of Rex rabbits was higher than that in the other ages (p < 0.05). However, the primary hair follicle density did not change significantly with the ages (p > 0.05; Table 4).
3.4. Gene Expression of Hair Follicle Development
The effects of different ages on the gene expression of hair follicle development in Rex rabbits are shown in Table 5. The expression of the Wnt10b, CTNNB1, APC, DVL2, GSK-3β, LEF1, DKK1 and TGF-β1 genes in the skin tissue of Rex rabbits at different age was significantly differences (p < 0.05), and the expression of the Wnt10b, CTNNB1, DVL2, GSK-3β, and TGF-β1 genes at 90 days was higher than that in the other days, the expression of the LEF1 and DKK1 genes at 60 days was higher than that in the other days. There was no significant difference in Fzd4 and TCF3 gene expression of Rex rabbits at different ages (p > 0.05).
3.5. Protein Phosphorylation Level
The effects of different ages on the protein phosphorylation levels of hair follicle development in Rex rabbits are shown in Figure 3. As indicated, the β-catenin protein phosphorylation (P-CTNNB1) levels in the skin tissue of Rex rabbits shows the trend of first increasing, then decreasing and then increasing, 60 days, 120 days and 150 days significantly higher than 15 days, 30 days or 90 days (p < 0.05, Figure 3a). With the increasing of ages, the GSK-3β protein phosphorylation (P-GSK-3β) levels in the skin tissue of Rex rabbits were increasing, and maximum at 120 days of age (p < 0.05, Figure 3c).
4. Discussion
The rabbit has the characteristics of fast growth in the early stage and slow growth in the late stage, and the growth rate of Tianfu black rabbit and New Zealand rabbit showed an upward trend at the age of 30-75 days, and a downward trend at the age of 75 days [17]. Qin et al [18] reported that the average weight of 15, 30, 45, 60, 75 and 90 days old German white rabbits were 64.3 g, 194.6 g, 373.3 g, 495.0 g, 648.4 g and 732.2 g respectively. The growth peak of rabbits was from 30 to 75 days old, and the growth rate decreased after 90 days. As a local variety, the growth rate of southwest Minxinan black rabbit is slow, and the best slaughter time is later than that of the commercial rabbit (the appropriate age is 110-130 days) [19]. Besides, 84 days of age of slaughter age is appropriate for Laiwu black rabbits according to muscle quality and potential commercial value [20]. In the actual production, it was determined that the slaughter of medium-sized meat rabbit at the age of 2.5-3 months was economical and the best economic benefit could be obtained. The internal organs are the basic facilities of animal life and the carrier of animal physiological function. As indicated in this study (Table 2), the values for body weight, heart weight, lung weight, liver weight and kidney weight of Rex rabbits were increased with age. However, heart index, lung index, and kidney index decreased with age. For the liver index, it increased first and then decreased with the increase of age, and reached the maximum at 60 days of age. Therefore, the growth and development of Rex rabbits internal organs have nonlinear characteristics.
Rex rabbits is a typical type of rabbit for skin, whose skin has important economic value. The early stage (30-90 days old) of Rex rabbit is mainly individual growth, after 90 days old fur began to mature [21]. In this work, the values for coat length, skin thickness, skin area, and skin weight of Rex rabbits were increased with age. However, the relative weight of skin of Rex rabbits were similar between different ages (Table 3). Hair follicle is a kind of adjunct organ of skin, which exists in dermis and subcutaneous tissue. Hair follicles are complex suborgan structures that are formed during embryonic development by the interaction of neuroectoderm and mesenchyme, and can be divided into primary and secondary hair follicles, primary follicles develop from a variety of cell taxa and secondary follicles from primary follicles [22]. Wnt protein is a kind of secretory glycoproteins rich in cysteine, which can act by paracrine or autocrine, it binds to the frizzled receptor family and the low-density lipoprotein receptor related protein (LRP) on the cell membrane, stabilizes and accumulates β-catenin in the cytoplasm [23]. The mammalian genome encodes 17 Wnt proteins, including Wnt 3, Wnt 4, Wnt 5a, Wnt 6, Wnt 7a, Wnt 7b, Wnt 10a, Wnt 10b, Wnt 11 and Wnt 16 [24]. Beta-catenin is one of the important molecules in Wnt signaling pathway, which is involved in many biological processes such as embryogenesis and ontogeny. When Wnt binds to frizzled and LRP5/6, the cytosolic protein dishevelled (DVL) is activated, resulting in the phosphorylation of LRP5/6 by casein kinase 1 alpha and glycogen synthase kinase 3 beta (GSK-3β), the binding of Axin to LRP5/6, and the accumulation of free β-catenin in the cytoplasm without phosphorylation. β-catenin enters the nucleus, displaces tansducin-like enhancer while binding to the lymphoid enhancer factor/ T-cell factor, recruits two histone modifiers, cAMP-response element binding protein binding protein (CBP) and Brahma-related gene-1 (BRG1), as well as coactivators such as B-cell lymphoma 9 (BCL9) and Pygopus, and activates the expression of target genes [25]. Wnt signaling requires the sequestration of GSK-3β inside multivesicular endosomes [26]. In this experiment, with the increasing of ages, the GSK-3β protein phosphorylation (P-GSK-3β) levels in the skin tissue of Rex rabbits were increasing, and maximum at 120 days of age (Figure 3c), which suggests that GSK-3β as an important member of Wnt/β-catenin pathway signaling participates in the biological process of Rex rabbit hair follicle development. Dickkopf-associated protein 1 (DKK1) is an important antagonist of the Wnt signaling pathway. Transgenic mice overexpressing the Wnt extracellular inhibitor DKK1 block the Wnt/β-catenin signaling pathway, thereby affecting hair follicle initiation and development and even ultimately leading to deletion of hair follicles in postnatal mice [27]. During embryogenesis, conditional knockout of the β-catenin gene in the epidermis results in a blocked formation of the hair substrate, thereby blocking hair follicle morphogenesis [28]. DKK1 can inhibit the Wnt signaling pathway by inhibiting β-catenin activity, leading to hair follicle degeneration [29]. The hair follicle development of Rex rabbit mainly concentrated in 20-26 gestational age, the most active development was at 23-24 gestational age, and the basic development was formed at 25-26 gestational age [30]. Activation of the Wnt/β-catenin signaling pathway is a critical initial step in hair basal plate formation during hair follicle development in the embryonic period [31,32]. After birth, the hair of Rex rabbits has a certain growth period, growing to the end of maturity because of undifferentiated cells at the bottom of the hair follicle gradually slow differentiation, and finally stop growing; This process of growth, decay and repose of rabbit hair and its replacement by new hair is called reflexing [33]. The basis of Rex rabbit hair replacement is the growth period, the decline period and the resting period of hair follicle development. According to the morphological characteristics of hair follicle, the hair follicle cycle can be divided into anagen, catagen and telogen. The growth process of hair follicles goes through growing period, declining period and resting period, hair follicles undergo a cycle of growth, decline, and rest before re-entering the growth cycle, a process known as hair follicle periodic regeneration [34]. In this study, we found that there are periodic changes in the development of Rex rabbits hair follicles, the major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 days and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation (Figure 1). The hair follicle density in 120 days of Rex rabbits was higher (Table 4; Figure 2), this is consistent with previous reports.
Lei et al [35] results demonstrated that overexpression of mouse whisker Wnt10b protein can activate the canonical Wnt signaling pathway, promoting the proliferation of hair stromal cells, thereby inducing hair follicles to enter the growth phase; Adenovirus-mediated overexpression of Wnt10b protein induces hair follicles to grow from quiescent phase. In contrast, siRNA-mediated knockout of the Wnt10b gene can prevent hair follicles from entering the growth phase [36]. Wnt10b-mediated aberrant activation of the Wnt pathway increases the number of CD34 + hair follicle stem cells in a proliferative state, resulting in enlargement of the hair bulb, hair shaft, and dermal papilla [37]. Beta-catenin is transiently expressed in the epidermis of adult mice, and the normal hair follicle cycle is interrupted to enter the growth phase and to form new hair follicles de novo [38] , these results suggest that β-catenin-dependent Wnt pathway is sufficient to induce the proliferation of hair follicle stem cells and subsequent growth of new hair follicles. High expression of β-catenin induces hair follicle stem cells to differentiate into hair follicle-forming cells, and when β-catenin is low expression or absent, hair follicle stem cells no longer participate in hair self-renewal, instead, they differentiate in the direction of epithelial keratinocytes [39]. Dasgupta et al [40] , upon the occurrence of the above phenomena, re-overexpressed β-catenin, a process that reverses, that is, cells that would otherwise have differentiated in the epithelial direction differentiate in the direction of forming hair follicle structures, allowing hair growth to continue. In addition, studies have shown that increasing the content of stable β-catenin can induce the formation of new hair follicles in adult epidermis [41]. When Wnt signaling pathway is activated, β-catenin dephosphorylates and stably exists, which makes cytosolic β-catenin enter nucleus, initiates expression of downstream genes, and regulates hair follicle periodic growth. However, when local epidermal activity of β-catenin is overexpressed, hair follicles are periodically dysregulated, which can lead to suppression of folliculogenesis [42,43,44,45]. In this study, the β-catenin protein phosphorylation (P-CTNNB1) levels in the skin tissue of Rex rabbits shows the trend of first increasing, then decreasing and then increasing, 60 days, 120 days and 150 days higher than 15 days, 30 days or 90 days (Figure 3a), which suggests that the P-CTNNB1 levels in the skin tissue of Rex rabbits related to the biological process of hair follicle development. Besides, transforming growth factor-β (TGF-β) can regulate the quiescence or activation state of hair follicle stem cells, and hair follicles can advance into the growth phase by injection of TGF-β2 recombinant protein [46]. Dermal papilla cells can secrete TGF-β2 after knockout of the TGF-β receptor in hair follicle stem cells, inability to receive TGF-β2 signaling, which activates hair follicle stem cells to transition from resting to growth phase [47]. Further, their existence of a growth/rest switching mechanism in the hair follicle that is based on an Eng-dependent feedback cross-talk between Wnt/β-catenin and Bmp/Smad signals [48]. The expression of the Wnt10b, CTNNB1, APC, DVL2, GSK-3β, LEF1, DKK1 and TGF-β1 genes in the skin tissue of Rex rabbits at different age was differences, and the expression of the Wnt10b, CTNNB1, DVL2, GSK-3β, and TGF-β1 genes at 90 days was higher than that in the other days (Table 5). Therefore, Wnt10b/β-catenin signaling pathways is involved in the biological process of periodic hair follicle development in different ages of Rex rabbits.
5. Conclusions
There are periodic changes in the development of Rex rabbit hair follicles, the major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 days and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation. The hair follicle density in 120 days of Rex rabbits was higher, and Wnt10b/β-catenin signaling pathways is involved in the biological process of periodic hair follicle development in different ages of Rex rabbits. Considering the rule of fur growth, the slaughter age of Rex rabbit should not be earlier than 120 days.
Author Contributions
Ce Liu and Gongyan Liu conceived and planned experiments; Yin Zhang methodology; Shuxia Gao, Haitao Sun, Liya Bai carried out experimental part; Ce Liu, Liping Yang and Gongyan Liu carried out molecular analysis; Yin Zhang carried out statistical analysis; Ce Liu prepared the manuscript draft, Gongyan Liu, Shuxia Gao and Haitao Sun were funding acquisition. All authors provided critical feedback and helped share the research, analysis, and manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been financially supported by funds from the Natural Science Foundation of Shandong Province (ZR2023QC017); Action Plan for Rural Revitalization of Science and Technology Innovation, Key Research and Development Program of Shandong Province (2023TZXD044); Shandong Province Modern Agricultural Industry Technology System (SDAIT-21-09); Earmarked Fund for Modern Agro-industry Technology Research System (CARS-43-G-7).
Institutional Review Board Statement
The experimental procedures were approved by the Institute of Animal Husbandry and Veterinary, Shandong Academy of Agricultural Sciences Animal Care and Use Committee (IASVM-2022-08).
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Schneider, M.R.; Schmidt-Ullrich, R.; Paus, R. The hair follicle as a dynamic miniorgan. Curr. Biol. 2009, 19, R132–R142. [Google Scholar] [CrossRef]
- Hsu, Y.C.; Li, L.; Fuchs, E. Transit-amplifying cells orchestrate stem cell activity and tissue regeneration. Cell 2014, 157, 935–949. [Google Scholar] [CrossRef]
- Millar, S. E. Molecular mechanisms regulating hair follicle development. J. Invest. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef]
- Lee, J.; Tumbar, T. Hairy tale of signaling in hair follicle development and cycling. Semin. Cell Dev. Biol. 2012, 23, 906–916. [Google Scholar] [CrossRef]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Qureshi, W.M.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef]
- Xiang, M. Gene regulation by Wnt signaling pathway in the oriented differentiation of hair follicle stem cells. J Tissue Eng. Reconstr. Surg. 2011, 7, 290–294. [Google Scholar]
- Lin, C.; Yuan, Y.; Chen, X.; Li, H.; Cai, B.; Liu, Y.; Zhang, H.; Li, Y.; Huang, K. Expression of Wnt/β-catenin signaling, stem-cell markers and proliferating cell markers in rat whisker hair follicles. J. Mol. Histol. 2015, 46, 233–240. [Google Scholar] [CrossRef]
- González-Sancho, J.M.; Brennan, K.R.; Castelo-Soccio, L.A.; Brown, A.M.C. Wnt proteins induce dishevelled phosphorylation via an LRP5/6-independent mechanism: irrespective of their ability to stabilize b-catenin. Mol. Cell. Biol. 2004, 24, 4757–4768. [Google Scholar] [CrossRef]
- Enshell-Seijffers, D.; Lindon, C.; Kashiwagi, M.; Morgan, B.A. beta-catenin activity in the dermal papilla regulates morphogenesis and regeneration of hair. Develop. Cell 2010, 18, 633–642. [Google Scholar] [CrossRef]
- Majidinia, M.; Aghazadeh, J.; Jahanban-Esfahlani, R.; Yousefi, B. The roles of Wnt/beta-catenin pathway in tissue development and regenerative medicine. J. Cell Physiol. 2018, 233, 5598–5612. [Google Scholar] [CrossRef]
- Tian, H. , Wang, X.; Lu, J.; Tian, W.; Chen, P. MicroRNA-621 inhibits cell proliferation and metastasis in bladder cancer by suppressing Wnt/beta-catenin signaling. Chem. Biol. Interact. 2019, 308, 244–251. [Google Scholar] [CrossRef]
- Wu, Z.; Zhu, Y.; Liu, H.; Liu, G.; Li, F. Wnt10b promotes hair follicles growth and dermal papilla cells proliferation via Wnt/β-Catenin signaling pathway in Rex rabbits. Bioscience Rep. 2020, 40, BSR20191248. [Google Scholar] [CrossRef]
- Liu, G.; Liu, C.; Zhang, Y.; Sun, H.; Yang, L.; Bai, L.; Gao, S. Hair follicle development of Rex rabbits is regulated seasonally by Wnt10b/β-Catenin, TGFβ-BMP, IGF1, and EGF signaling pathways. Animals 2023, 13, 3742. [Google Scholar] [CrossRef]
- Liu, G.Y.; Wu, Z.Y.; Zhu, Y.L.; Liu, L.; Li, F.C. Effects of dietary vitamin B6 on the skeletal muscle protein metabolism of growing rabbits. Anim. Prod. Sci. 2017, 57, 2007–2015. [Google Scholar] [CrossRef]
- LiLivak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2 -ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, G.; Li, S.; Liu, H.; Zhu, Y.; Bai, L.; Sun, H.; Gao, S.; Jiang, W.; Li, F. The functions of ocu-miR-205 in regulating hair follicle development in Rex rabbits. BMC Dev. Biol. 2020, 20, 1–13. [Google Scholar] [CrossRef]
- Lai, S.; Yang, G.; Li, Y. A comparison between Tianfu black rabbit and New Zealand rabbit in growth rate and feed conversion efficiency. J. Sichuan Agricul. University 2002, 20, 362–363+366. [Google Scholar]
- Qin, Y.; Li, L.; Du, Y. Comparison of early growth and feed conversion efficiency between German White Rabbit and New Zealand Rabbit. Chinese Rabbit Breeding 2000, 2, 18–20. [Google Scholar]
- Liu, G.; Li, M.; Liu, M.; Sun, H.; Bai, L.; Liu, C.; Yang, L.; Jiang, W.; Wang, P.; Gao, S. Effects of different slaughter ages on slaughter performance, fat deposition and muscle quality of Minxinan black rabbit. Chinese J. Anim. Husbandry 2022, 58, 224–228. [Google Scholar]
- Zhou, B.; Zhang, Y.; Yang, L.; Gao, S.; Liu, G.; Zhang, Y.; Zhang, H.; Bai, L. Comparative analysis of production performance and muscle quality of Laiwu black rabbits at different slaughter ages. Chinese J. Anim. Nutr. 2023, 35, 5322–5332. [Google Scholar]
- Yurniati, Y.; Raharjo, Y.; Kusmajadi, S. The effect of restricted feeding and different of slaughtering age on production of Rex rabbit pelt. J. Indonesian Trop. Anim. Agric. 2014, 35, 192–196. [Google Scholar] [CrossRef]
- Alonso, L. , Fuchs, E. Stem cells in the skin: waste not, Wnt not. Genes Dev. 2003, 17, 1189–1200. [Google Scholar] [CrossRef] [PubMed]
- Clevers, H.; Nusse, R. Wnt/beta catenin signaling and diseease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef]
- Augustin, I. Wnt signaling in skin homeostasis and pathology. J. Dtsch. Dermatol. Ges. 2015, 13, 302–306. [Google Scholar] [CrossRef]
- Saito-Diaz, K.; Chen, T.W.; Wang, X.; Thorne, C.A; Wallace, H. A.; Page-Mc, C. A.; Lee, E. The way Wnt works: components and mechanism. Growth Factors 2013, 31, 1–31. [Google Scholar] [CrossRef]
- Yuan, Y.P.; Huang, K.; Xu, Y.M.; Chen, X.C.; Li, H.H.; Zhang, H.; Li, Y.; Lin, C.M. Canonical and non-canonical Wnt signaling control the regeration of amputated rodent vibrissae follicles. J. Mol. Histol. 2016, 47, 47,1–8. [Google Scholar] [CrossRef]
- Taelman, V.F.; Dobrowolski, R.; Plouhinec, J.L.; Fuentealba, L.C. , Vorwald, P.P., Gumper, I., Sabatini, D.D.; Robertis, E.M. Wnt signaling requires sequestration of glycogen synthase kinase 3 inside multivesicular endosomes. Cell 2010, 143, 1136–1148. [Google Scholar] [CrossRef]
- Lim, X.; Nusse, R. Wnt signaling in skin development, homeostasis, and disease. Cold Spring Harb. Perspect. Biol. 2013, 5, a008029. [Google Scholar] [CrossRef]
- Kwack, M.H.; Kim, M.K.; Kim, J.C.; Sung, Y.K. Dickkopf 1 promotes regression of hair follicles. J. Invest. Dermatol. 2012, 132, 1554–1560. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Li, B.; Zhu, Y.; Wang, C.; Li, F. Differential gene expression profiles in foetal skin of Rex rabbits with different wool density. World Rabbit Sci. 2016, 24, 223–231. [Google Scholar] [CrossRef]
- Fu, J.; Hsu, W. Epidermal Wnt controls hair follicle induction by orchestrating dynamic signaling crosstalk between the epidermis and dermis. J. Invest. Dermatol. 2013, 133, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Chen D, Jarrell A, Guo, C. Dermal beta catenin activity in response to epidermal Wntligands is required for fibroblast proliferation and hair follicle initiation. Development 2012, 139, 1522–1533. [Google Scholar]
- Wu, Z.; Sun, L.; Liu, G.; Liu, H.; Liu, H.; Yu, Z.; Xu, S.; Li, F.; Qin, Y. Hair follicle development and related gene and protein expression of skins in Rex rabbits during the first 8 weeks of life. Asian Austral. J. Anim. 2019, 32, 477–484. [Google Scholar] [CrossRef]
- Woo, C.; Eleanor, W.; Bruce, A. Morgan dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline. Develop. Stem Cells 2013, 140, 1676–1683. [Google Scholar]
- Lei, M.; Lai, X.; Bai, X.; Qiu, W.; Yang, T.; Liao, X.; Chuong, C.M.; Yang, L.; Lian, X.; Zhong J., L. Prolonged overexpression of Wnt10b induces epidermal keratinocyte transformation through activating EGF pathway. Histochem. Cell Biol. 2015, 144, 209–221. [Google Scholar] [CrossRef]
- Li, Y.H.; Zhang, K.; Yang, K.; Ye, J.X.; Xing, Y.Z.; Guo, H.Y.; Deng, F.; Lian, X.H.; Yang, T. Adenovirus-mediated Wnt10b overexpression induces hair follicle regeneration. J. Invest. Dermatol. 2013, 133, 42–48. [Google Scholar] [CrossRef]
- Lei, M.; Guo, H.; Qiu, W.; Lai, X.; Yang, T.; Widelitz R., B.; Chuong, C.M.; Lian, X.; Yang, L. Modulating hair follicle size with Wnt10b/DKK1 during hair regeneration. Exp. Dermatol. 2014, 23, 407–413. [Google Scholar] [CrossRef]
- Lichtenberger, B.M.; Mastrogiannaki, M.; Watt, F.M. Epidermal beta-catenin activation remodels the dermis via paracrine signalling to distinct fibroblast lineages. Nat. Commun. 2016, 7, 10537. [Google Scholar] [CrossRef]
- Huelsken, J.; Vogel, R.; Erdmann, B.; Cotsarelis, G.; Birchmeier, W. Beta catenin controls hair follicle morphogenesis and stem cell differentiation in the skin. Cell 2001, 105, 533–545. [Google Scholar] [CrossRef]
- Dasgupta, R.; Fuchs, E. Multiple roles for activated LEF/TCF transcription complexes during hair follicle development and differentiation. Develop. 1999, 126, 4557–4568. [Google Scholar] [CrossRef]
- Niezgoda, A.; Niezgoda, P.; Nowowiejska, L.; Białecka, A.; Męcińska-Jundziłł, K.; Adamska, U.; Czajkowski, R. Properties of skin stem cells and their potential clinical applications in modern dermatology. Eur. J. Dermatol. 2017, 27, 227–236. [Google Scholar] [CrossRef]
- Mastrogiannaki M, Lichtenberger B M, Reimer A, Collins, C. A.; Driskell, R. R.; Watt, F. M. Beta-catenin stabilization in skin fibroblasts causes fibrotic lesions by preventing adipocyte differentiation of the reticular dermis. J. Invest. Dermatol. 2016, 136, 1130–1142. [Google Scholar] [CrossRef] [PubMed]
- Jahoda, C.; Gilmore, A.C. What lies beneath: Wnt/beta-catenin signaling and cell fate in the lower dermis. J. Invest. Dermatol. 2016, 136, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.J.; Lin, G.Y.; Zhu, J.Y. , Yin, G.Q.; Huang, D.; Yan Y.Y. LncRNA-PCAT1 maintains characteristics of dermal papilla cells and promotes hair follicle regeneration by regulating miR-329/Wnt10b axis. Exp. Cell Res. 2020, 394, 112031. [Google Scholar] [CrossRef]
- Rishikaysh, P.; Dev, K.; Diaz, D.; Qureshi, W.M.S.; Filip, S.; Mokry, J. Signaling involved in hair follicle morphogenesis and development. Int. J. Mol. Sci. 2014, 15, 1647–1670. [Google Scholar] [CrossRef] [PubMed]
- Foitzik, K.; Paus, R.; Doetschman, T. The TGF-beta 2 isoform is both a required and sufficient inducer of murine hair follicle morphogenesis. Deve.Biol. 1999, 212, 278–289. [Google Scholar] [CrossRef] [PubMed]
- Oshimori, N.; Fuchs, E. Paracrine TGF-beta signaling counterbalances BMP-mediated repression in hair follicle stem cell activation. Cell Stem Cell. 2012, 10, 63–75. [Google Scholar] [CrossRef]
- Calvo-Sanchez, M.I.; Fernandez-Martos, S.; Carrasco, E.; Bernabéu, C.; Quintanilla, M.; Jesús, E.J. A role for the Tgf-beta/bmp co-receptor endoglin in the molecular oscillator that regulates the hair follicle cycle. J. Mol. Cell Biol. 2019, 11, 39–52. [Google Scholar] [CrossRef]
Figure 1.
Longitudinal incision of skin and hair follicle tissue of Rex rabbits, (a) skinning in 15 days; (b) skinning in 30 days; (c) skinning in 60 days; (d) skinning in 90 days; (e) skinning in 120 days; (f) skinning in 150 days. The major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation. Scale bars =500 μm.
Figure 1.
Longitudinal incision of skin and hair follicle tissue of Rex rabbits, (a) skinning in 15 days; (b) skinning in 30 days; (c) skinning in 60 days; (d) skinning in 90 days; (e) skinning in 120 days; (f) skinning in 150 days. The major hair follicles were in the growing period at 15 days and 30 days of age, some of them had already entered the degeneration period at 60 days of age, and most of them entered the degeneration period at 90 days of age, the presence of quiescent hair follicles and the enlargement of primary hair follicles were also observed. At 120 and 150 days of age, the back hair follicles entered the growing period, and the secondary hair follicles had obvious differentiation. Scale bars =500 μm.
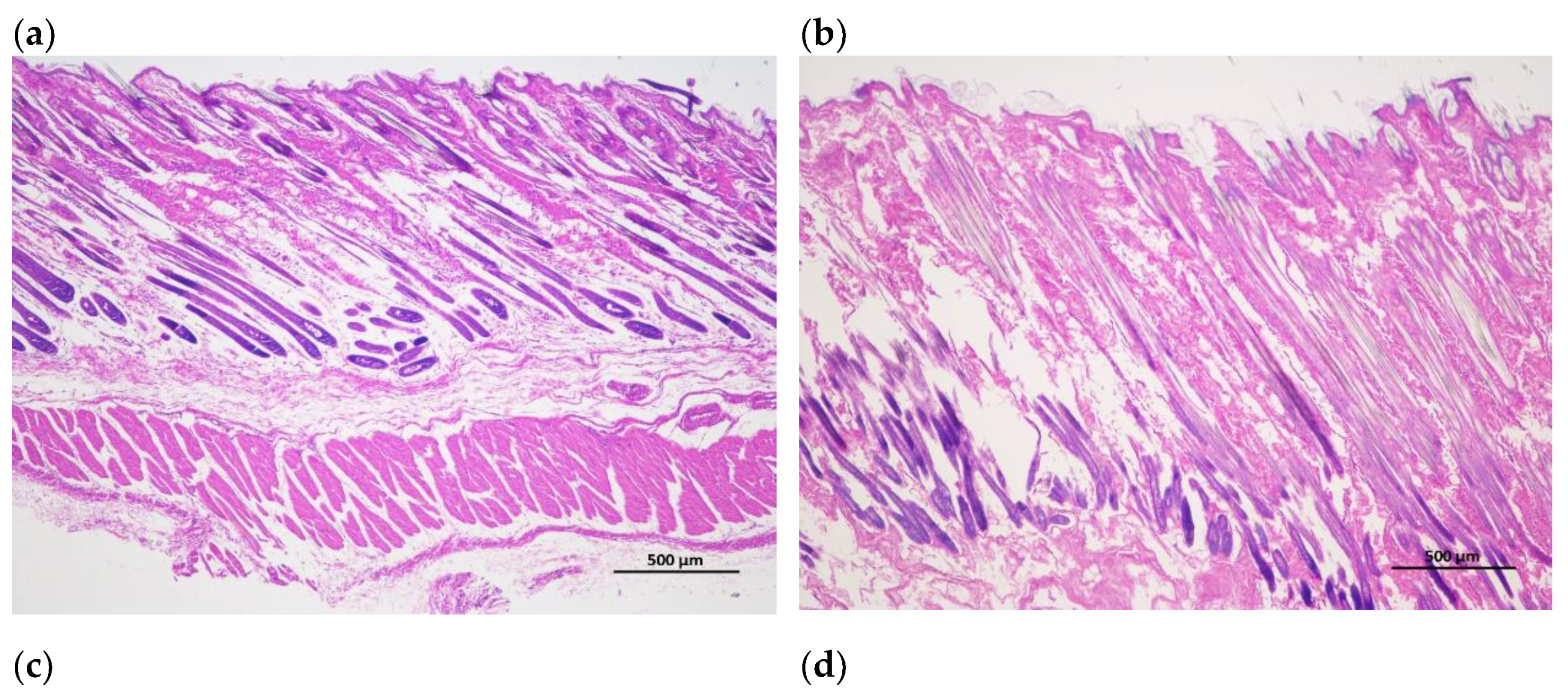
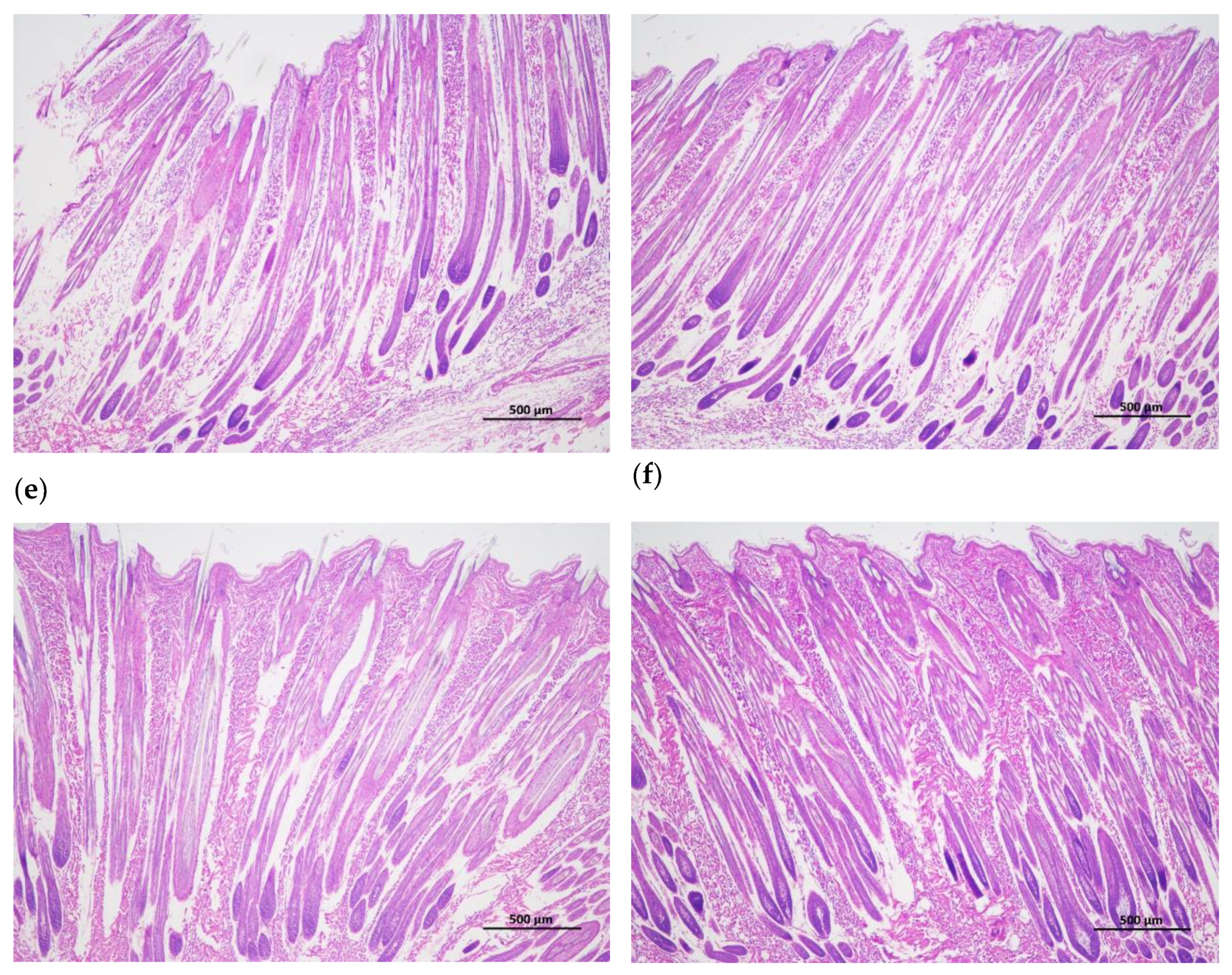
Figure 2.
Cross-cutting of skin and hair follicle tissue of Rex rabbits, (a) skinning in 15 days; (b) skinning in 30 days; (c) skinning in 60 days; (d) skinning in 90 days; (e) skinning in 120 days; (f) skinning in 150 days. The total hair follicle density, secondary hair follicle density and the ratio of the secondary hair follicle density to the primary hair follicle density in 120 days of Rex rabbits was higher than that in the other ages. Scale bars =500 μm.
Figure 2.
Cross-cutting of skin and hair follicle tissue of Rex rabbits, (a) skinning in 15 days; (b) skinning in 30 days; (c) skinning in 60 days; (d) skinning in 90 days; (e) skinning in 120 days; (f) skinning in 150 days. The total hair follicle density, secondary hair follicle density and the ratio of the secondary hair follicle density to the primary hair follicle density in 120 days of Rex rabbits was higher than that in the other ages. Scale bars =500 μm.
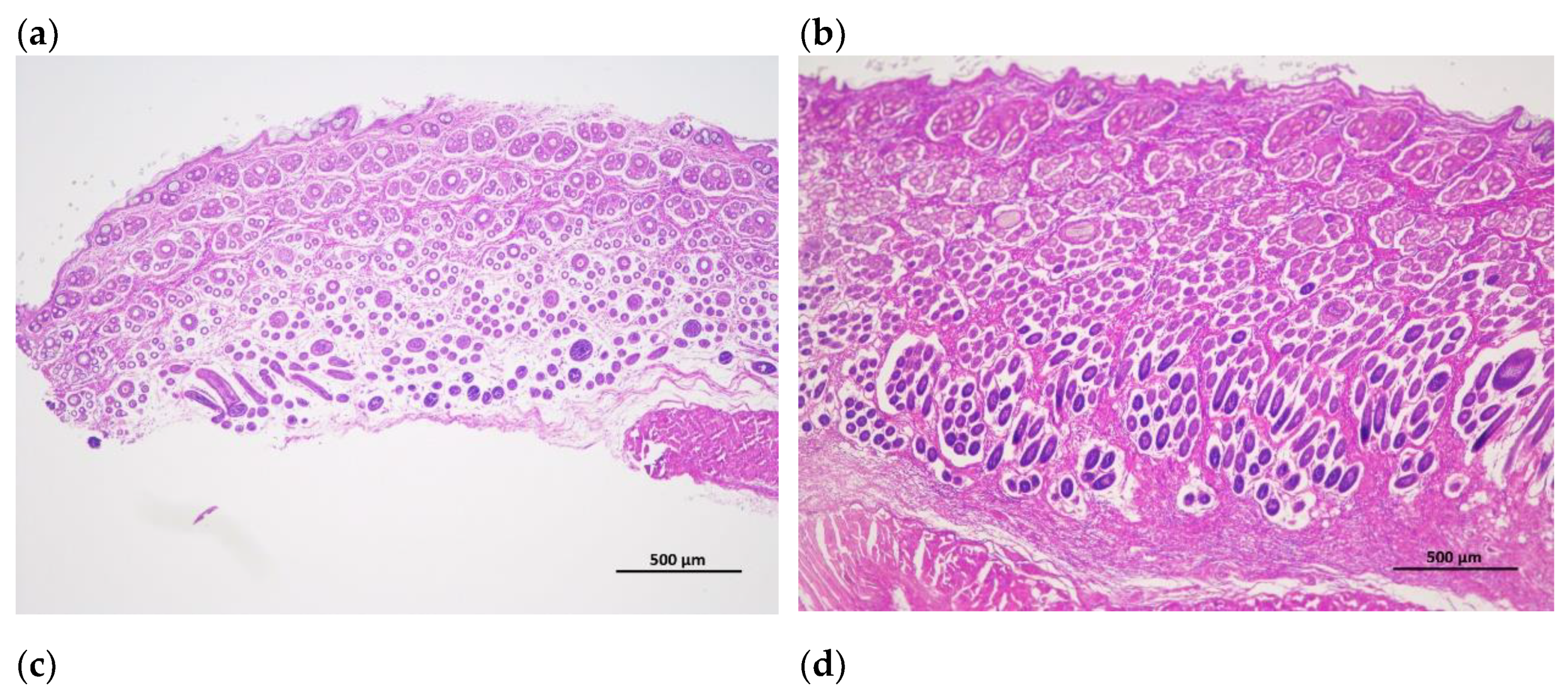
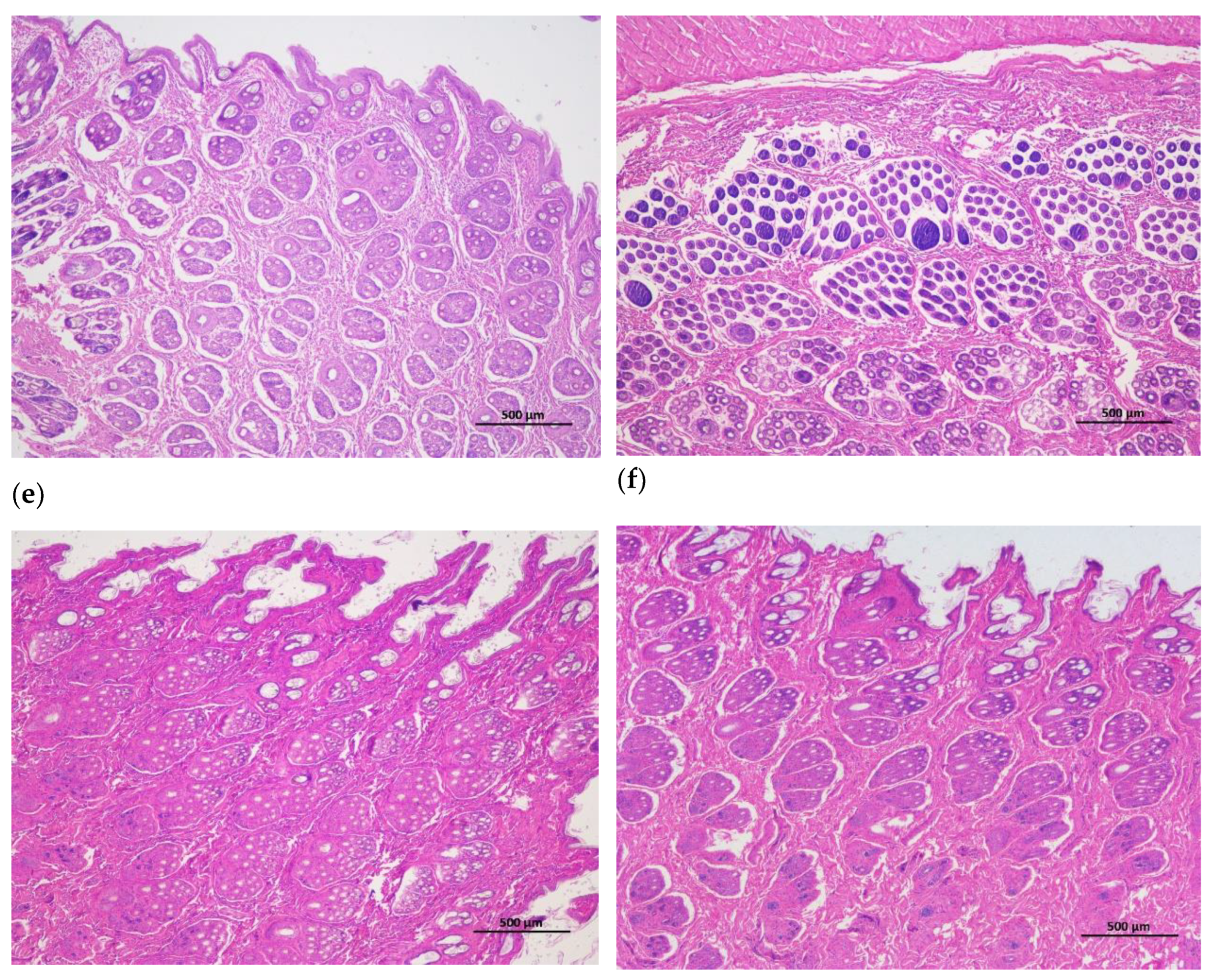
Figure 3.
Effects of different ages on the protein phosphorylation levels in Rex rabbits. (a) β-catenin protein phosphorylation (P-CTNNB1) levels in skin tissue; (b) Blotting strip of P-CTNNB1 in skin tissue; (c) GSK-3β protein phosphorylation (P-GSK-3β) levels in skin tissue; (d) Blotting strip of P-GSK-3β in skin tissue; Means with different superscripts are significantly different (p < 0.05), n=6. .
Figure 3.
Effects of different ages on the protein phosphorylation levels in Rex rabbits. (a) β-catenin protein phosphorylation (P-CTNNB1) levels in skin tissue; (b) Blotting strip of P-CTNNB1 in skin tissue; (c) GSK-3β protein phosphorylation (P-GSK-3β) levels in skin tissue; (d) Blotting strip of P-GSK-3β in skin tissue; Means with different superscripts are significantly different (p < 0.05), n=6. .
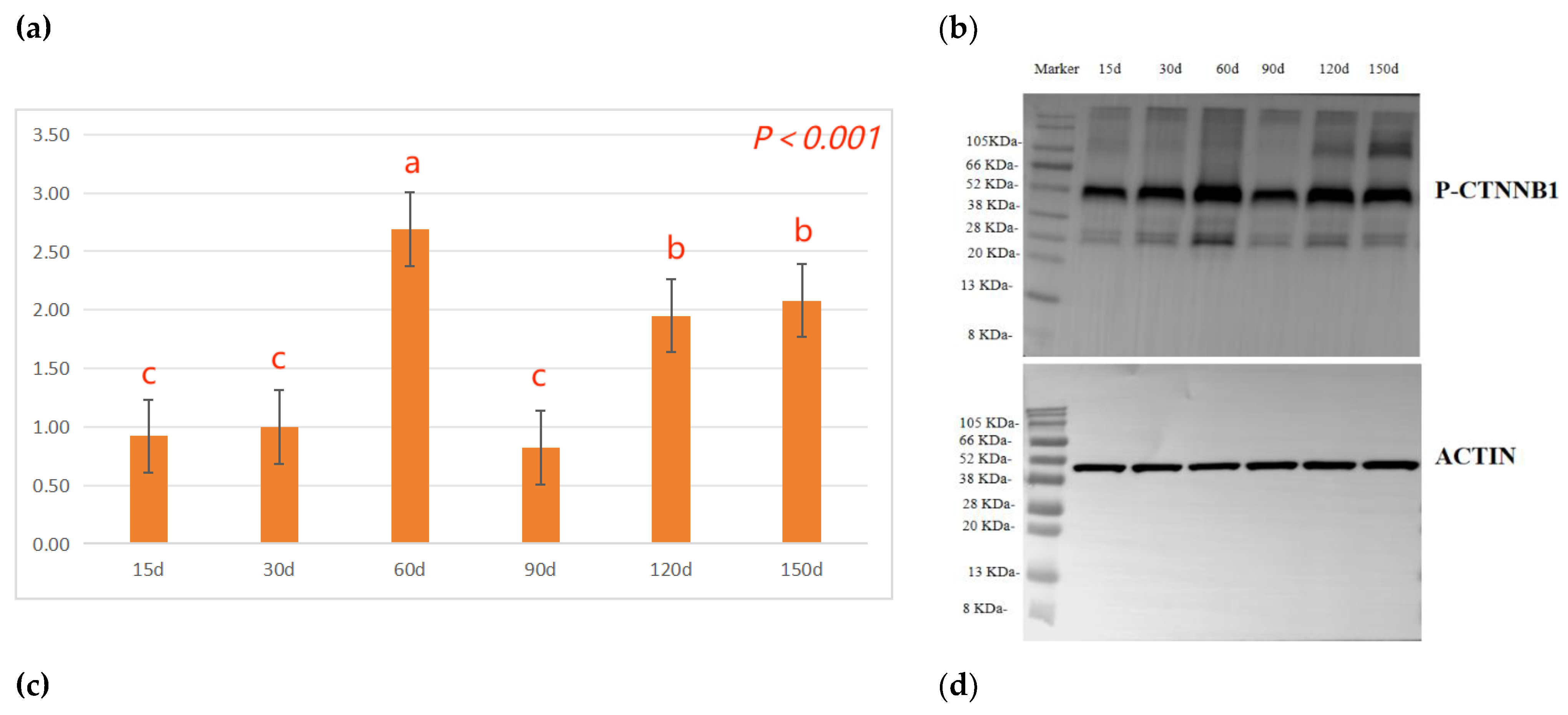
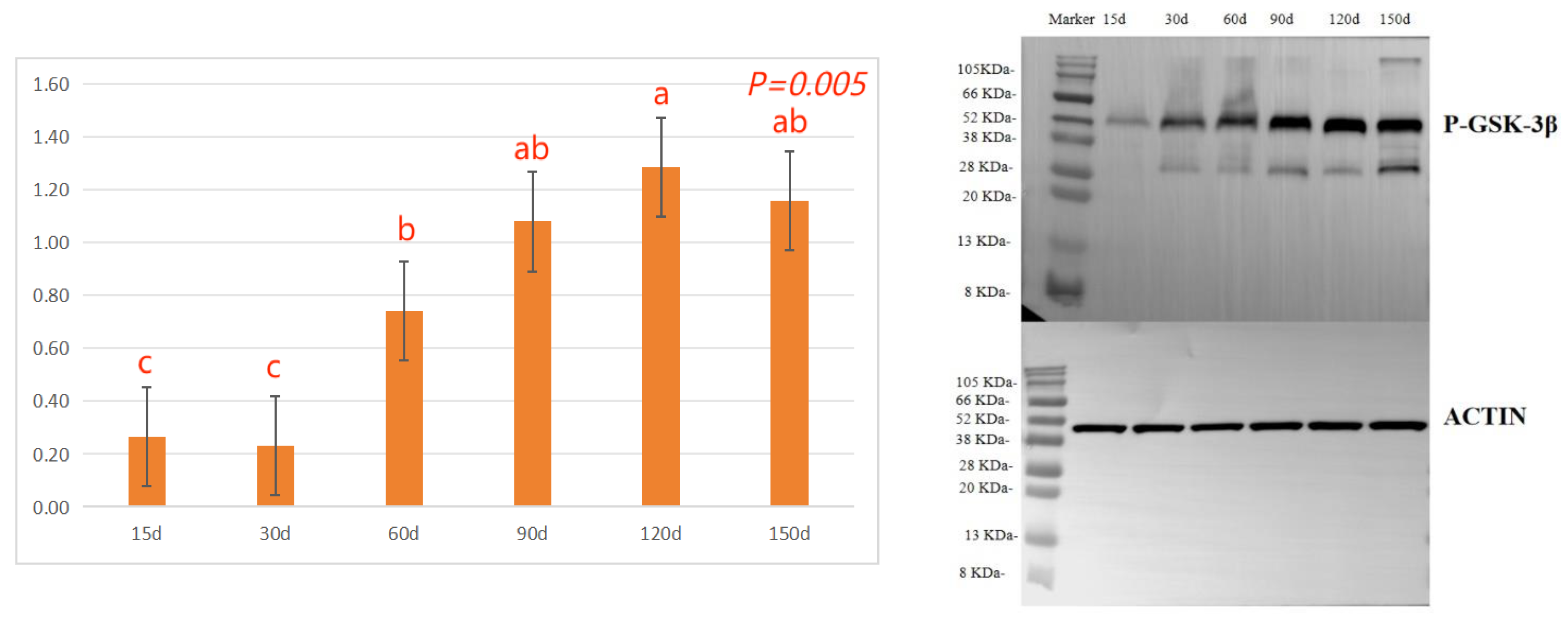

Table 1.
Information on primers.
| Gene | Accession number | Primer sequence (5′-3′) | Product length, bp |
|---|---|---|---|
| GAPDH | NM_001082253.1 | F: TTCCAGTATGATTCCACCCACG | 232 |
| R: GGGCTGAGATGATGACCCTTTT | |||
| Wnt10b | XM_002711076.4 | F: GGCGAGAATGAGAATCCATAACAA | 196 |
| R: GTTGTGGGTGTCAATGAAGATGG | |||
| Fzd4 | XM_002708648 | F: AAGTGGGTCAGATGGTCCTG | 117 |
| R: CCCGATGAAGTGAAACTGGT | |||
| CTNNB1 | XM_051852655.1 | F: TGGATACCTCCCAAGTCCTGTA | 207 |
| R: CCAGACGCTGAACATTAGTAGGAT | |||
| APC | XM_008248401 | F: GACTCCAGGCTTCTGGTTTG | 121 |
| R: TAGTGCTCTGGTGGGCTCTT | |||
| DVL2 | XM_008270807 | F: ACTCCACCATGTCCCTCAAC | 117 |
| R: CGATGTAGATGCCTCCGTCT | |||
| GSK-3β | XM_017347066.1 | F: TGAGGTCTATCTTAATCTGGTGCTG | 183 |
| R: TGTGGTTTAATATCCCGATGGC | |||
| TCF3 | NM_001171390 | F: CGGGAGATAGAGCAGGTGAA | 127 |
| R: GGTAGTCATCGCCGTAGGAG | |||
| LEF1 | XM_008267508 | F: GCGTCCACACCTGTAACCTT | 122 |
| R: CTCTTCCTCAAATCCCTCCA | |||
| DDK1 | NM_001082737.2 | F: ATGGGTATTCCCGCAGAACC | 150 |
| R: CCTTGAGGACGGGCTTACAG | |||
| TGF-β1 | XM_008249704.2 | F: CTGCTGTGGCTCCTAGTGTTGA | 134 |
| R: AGCCGCAGTTTGGACAGGAT |
GAPDH: glyceraldehyde-3-phosphate dehydrogenase; Fzd4: Frizzled 4; CTNNB1: catenin beta 1; APC: Adenomatosis polyposis protein; DVL2: Disheveled 2; GSK-3β: glycogen synthase kinase 3 beta; TCF3: Transcription factor 3; Lef1: Lymphoid enhancer-binding factor 1; DKK1: dickkopf-1; TGF-β1: transforming growth factor beta 1; F: forward primer; R: reverse primer.
Table 2.
Effects of different ages on growth and development of internal organs in Rex rabbits.
| Items | Age (days) | SEM | p-value | |||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 60 | 90 | 120 | 150 | |||
| Body weight (g) | 340.0 f | 785.0 e | 1455.0 d | 2073.3 c | 2601.7 b | 3178.3 a | 169.15 | <0.001 |
| Heart weight (g) | 1.73 e | 3.15 d | 5.48 c | 6.97 b | 7.18 b | 8.62 a | 0.441 | <0.001 |
| Lung weight (g) | 3.30 d | 7.28 c | 10.13 bc | 11.58 b | 10.27 bc | 17.20 a | 0.877 | <0.001 |
| Liver weight (g) | 11.25 d | 27.43 c | 58.28 b | 60.30 b | 66.98 b | 80.22 a | 4.174 | <0.001 |
| Kidney weight (g) | 4.27 e | 8.13 d | 11.43 cd | 13.77 bc | 17.48 ab | 18.65 a | 0.985 | <0.001 |
| Heart index (g/kg) | 5.21 a | 3.98 b | 3.79 bc | 3.37 bc | 2.75 c | 2.71 c | 0.200 | <0.001 |
| Lung index (g/kg) | 9.95 a | 9.26 ab | 6.98 abc | 5.65 bc | 3.96 c | 5.37 bc | 0.609 | 0.018 |
| Liver index (g/kg) | 33.02 b | 34.53 b | 41.62 a | 28.12 c | 25.81 c | 25.24 c | 1.129 | <0.001 |
| Kidney index (g/kg) | 12.56 a | 10.32 ab | 7.89 bc | 6.64 c | 6.81 c | 5.87 c | 0.515 | <0.001 |
Means with different superscripts in a row are significantly different (p < 0.05), n=6. SEM: standard error of means.
Table 3.
Effects of different ages on the fur quality of Rex rabbits.
| Items | Age (days) | SEM | p-value | |||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 60 | 90 | 120 | 150 | |||
| Coat length (cm) | 1.81 b | 1.82 b | 1.88 ab | 1.86 ab | 1.87 ab | 1.95 a | 0.013 | 0.026 |
| Skin thickness (mm) | 4.67 b | 6.62 a | 6.87 a | 6.92 a | 7.25 a | 7.28 a | 0.174 | <0.001 |
| Skin area (cm2) | 367.50 f | 649.67 e | 996.50 d | 1354.75 c | 1699.00 b | 1937.80 a | 169.145 | <0.001 |
| Skin weight (g) | 55.0 f | 118.8 e | 227.6 d | 322.9 c | 435.1 b | 545.4 a | 29.928 | <0.001 |
| Relative weight of skin (%) | 16.19 | 14.26 | 15.59 | 15.56 | 16.64 | 17.16 | 0.316 | 0.119 |
Means with different superscripts in a row are significantly different (p < 0.05), n=6. SEM: standard error of means.
Table 4.
Effects of different ages on the hair follicle density of Rex rabbits.
| Items | Age (days) | SEM | p-value | |||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 60 | 90 | 120 | 150 | |||
| Total hair follicle density (count/mm2) | 232.8 d | 287.8 c | 308.7 bc | 334.9 b | 370.8 a | 319.4 bc | 8.508 | <0.001 |
| Primary hair follicle density (count/mm2) | 12.02 | 14.36 | 12.17 | 13.58 | 13.11 | 14.04 | 0.291 | 0.089 |
| Secondary hair follicle density (count/mm2) | 220.8 d | 273.4 c | 296.5 bc | 321.3 b | 357.7 a | 305.4 bc | 8.421 | <0.001 |
| Secondary hair follicle/ Primary hair follicle ratio | 18.47 d | 19.46 cd | 24.52 ab | 23.85 b | 27.42 a | 21.87 bc | 0.667 | <0.001 |
Means with different superscripts in a row are significantly different (p < 0.05), n=6. SEM: standard error of means.
Table 5.
Effects of different ages on the gene expression of hair follicle development in Rex rabbits.
Table 5.
Effects of different ages on the gene expression of hair follicle development in Rex rabbits.
| Items | Age (days) | SEM | p-value | |||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 60 | 90 | 120 | 150 | |||
| Wnt10b | 1.00 b | 2.43 b | 2.54 b | 4.24 a | 1.85 b | 1.29 b | 0.273 | 0.003 |
| Fzd4 | 1.00 | 1.12 | 2.53 | 1.30 | 2.06 | 1.69 | 0.169 | 0.052 |
| CTNNB1 | 1.00 b | 3.05 a | 1.39 b | 2.68 a | 0.59 b | 0.52 b | 0.220 | <0.001 |
| APC | 1.00 b | 1.68 b | 5.38 a | 1.89 b | 5.46 a | 3.66 a | 0.376 | <0.001 |
| DVL2 | 1.00 c | 0.87 c | 1.39 bc | 2.10 a | 1.74 ab | 1.45 bc | 0.103 | 0.001 |
| GSK-3β | 1.00 b | 2.61 a | 0.98 b | 2.21 a | 0.66 b | 0.82 b | 0.178 | 0.001 |
| TCF3 | 1.00 | 1.11 | 1.13 | 1.64 | 1.38 | 1.45 | 0.089 | 0.281 |
| LEF1 | 1.00 c | 1.64 bc | 3.93 a | 3.42 ab | 3.27 ab | 2.57 abc | 0.311 | 0.037 |
| DDK1 | 1.00 b | 0.35 c | 1.83 a | 0.43 bc | 0.45 bc | 0.56 b | 0.112 | <0.001 |
| TGF-β1 | 1.00 bc | 1.40 ab | 1.04 bc | 2.04 a | 1.47 ab | 0.53 c | 0.115 | 0.001 |
Means with different superscripts in a row are significantly different (p < 0.05), n=6. SEM: standard error of means. Fzd4: Frizzled 4; CTNNB1: catenin beta 1; APC: Adenomatosis polyposis protein; DVL2: Disheveled 2; GSK-3β: glycogen synthase kinase 3 beta; TCF3: Transcription factor 3; Lef1: Lymphoid enhancer-binding factor 1; DKK1: dickkopf-1; TGF-β1: transforming growth factor beta 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



