
Submitted:
29 May 2024
Posted:
30 May 2024
You are already at the latest version
Abstract
Introduction: A central role for neuroinflammation in epileptogenesis has recently been suggested by several investigations. This systematic review explores the role of inflammatory mediators in epileptogenesis, its association with seizure severity, and its correlation with drug-resistant epilepsy (DRE). Material and Methods: The study analysed articles published in JCR journals from 2019 to 2024, including retrospective, prospective, case-control, or cross-sectional studies. It combined MESH and free terms for "Epileptogenesis" and "Neuroinflammation" and included searches for biomarkers previously reported in narrative reviews. Results: The study analysed 243 articles related to epileptogenesis and neuroinflammation, with 356 from selective searches by biomarker type. After eliminating duplicates, 324 articles were evaluated, with 272 excluded and 52 evaluated by authors. 21 articles were included in the qualitative evaluation, including 18 case-control studies, 2 case series, and 1 prospective study. This systematic review provides acceptable support for five biomarkers: TNF-a and some of its soluble receptors (sTNFr2), HMGB1 and TLR-4, CCL2 and IL-33. Discussions: Certain receptors, cytokines, and chemokines are examples of neuroinflammation-related biomarkers that may be crucial for the early diagnosis of refractory epilepsy or may be connected to the control levels of epileptic patients. Their value will be better defined by future studies.
Keywords:
Epileptogenesis
; Neuroinflammation
; Biomarkers
; HMGB1
; TNF-a
; TLR-4
; sTNFr2
; CCL2
; IL-33
; Drug Resistant Epilepsy
1. Introduction
Neuroinflammation refers to the inflammatory response of the Central Nervous System (CNS) to deviations from homeostasis that cannot be reversed by homeostatic mechanisms alone [1]. Today it is known that it is one of the pathophysiological processes that is transversally involved in multiple pathologies of the CNS [2]. Its role has been shown to be very relevant in the pathogenesis of diseases such as Multiple Sclerosis [3], the most paradigmatic example, but it is present in a wide variety of other diseases with special attention to neurodegenerative diseases [4,5]
Residents innate immune cells (particularly microglia and astrocytes) are involved in this CNS inflammatory response, and cytokines and their receptors are involved, which act to promote the migration of leukocytes to the site of inflammation and endothelial adhesion. Understanding the processes that occur between the immune system and CNS is crucial, especially in the age of personalized medicine, since many of the elements identified can become diagnostic or prognostic biomarkers of the disease or even become therapeutic targets [6,7,8].
In recent years, several studies have addressed the role that neuroinflammation may play in epileptogenesis [9,10,11]. Although the molecular mechanisms underlying these pathophysiological processes are not yet fully understood, it has been speculated that inflammatory mediators may cause abnormal angiogenesis and impairment of permeability of the blood-brain barrier (BBB), a circumstance that is closely related to epileptogenesis [11,12]. On the other hand, the unregulated focal or systemic inflammatory processes themselves lead to the formation of aberrant neuronal connections and hyperexcitable neural networks as well as an altered response to neurotransmitters, thus participating in the process of epileptogenesis. For both reasons, over the past two decades there has been growing evidence of both clinical and basic studies providing strong support for the conclusion that neuroinflammation is involved in epileptogenesis [13,14,15,16,17,18,19,20,21].
Finally, the role of cytokines as potential pro-inflammatory mediators in the neuropathology of epilepsy may also contribute to elucidating the process of epileptogenesis [22,23,24,25]. Thus, the overexpression of some of these inflammatory mediators in the hippocampus and neocortex of patients with epilepsy indicates the activation of multiple pro- and anti-epileptogenic immune pathways.
We know that between 25 and 30% of epileptic patients are resistant to available anti-seizure medications (ASMs) [26] and some studies have shown that the levels of inflammatory mediators may be elevated in these patients [7,16,27]. This data could open the door to the use of these molecules as biomarkers that can be used as predictors of the inflammatory response and for the development of new treatments in drug-resistant epilepsy [28].
The purpose of this systematic review is to know the state of art in the comprehension of the role of inflammatory mediators in epileptogenesis, to know if elevated levels of inflammatory mediators in serum and CSF can be associated with seizure severity and recurrence, and to know which of them can be most correlated with DRE and could therefore be explored as biomarkers of DRE or epileptogenesis.
2. Methods
2.1. Search Strategy and Databases: A Systematic Search Was Carried Out Using the Following Inclusion Criteria
- Articles published in journals indexed in JCR in the last 5 years (January 2019 to February 2024) using the MEDLINE database.
- Articles that included retrospective, prospective, case-control or cross-sectional studies.
- Searches were carried out by combining the following MESH and free terms: “Epileptogenesis” and “Neuroinflammation”
Specifically, the molecules included in this additional search were:
- High mobility group box 1/HMGB1”,
- Toll-Like-Receptor 4/ TLR-4”,
- Interleukin-1/IL-1”,
- Interleukin-6/IL-6”,
- Transforming growth factor beta/TGF-β” and
- Tumour necrosis factor-alpha/TNF-α”.
- 5.
- Articles published in English and/or Spanish.
The review protocol followed the declaration of Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA).
2.2. Exclusion Criteria
- Duplicate articles, editorials, letters to the editor, or reviews (both narrative and systematics).
- Articles that include basic research studies on tissues or animal models.
- Articles on studies not related to any type of epilepsy or focused on acute symptomatic seizures due to infectious, traumatic, vascular or oncological processes.
2.3. Study Selection
Each of the selected articles was evaluated by reading the title and abstract and their keywords by two different reviewers (MJA and PSC) applying the eligibility and exclusion criteria mentioned above. In case of discrepancy between the two reviewers, the aid of a third reviewer (PCG) was needed. The selected articles were read in their entirety to evaluate the degree of scientific evidence. The included studies were assessed based on the criteria discussed in the Standards for Reporting of Diagnostic Accuracy (STARD) checklist [33].
2.4. Data Extraction
2.5. Flowchart 2020 PRISMA
3. Results and Discussion
A total of 243 articles resulted from the search “epileptogenesis and neuroinflammation” and 356 from selective searches by biomarker type. Specifically, the results by type of biomarker were:
- HMGB1: 31 results
- TLR-4: 38 results
- IL-1b: 69 results
- IL-6: 101 results
- TGF-β: 29 results
- TNF-a: 91 results
After eliminating duplicates, 324 articles were evaluated by reading their titles and abstracts and applying the defined inclusion and exclusion criteria. A total of 272 articles were excluded and another 52 were evaluated by the authors through reading the full text. Finally, 21 articles were included in the qualitative evaluation. See Figure 1 for PRISMA 2020 Flowchart. By type of articles, our selection included 18 case-control studies, 2 case series, and 1 prospective population-based study.
3.1. Interleukin 1β (IL-1β)
Our review has found studies with conflicting results. The one with the highest level of evidence is, a prospective study by Wang et al. [35] that does not show elevated levels of this biomarker in people with epilepsy. Other case-control studies also report similar results [36,37]. Choi et al. [38], conversely, in a retrospective case-control study found an elevation of serum and CSF IL-1β in children who had suffered epileptic seizures in the previous 48 h. Kamaşak et al.[30] studied 28 children with DRE of at least three years of evolution, demonstrating significant differences in IL-1β levels. Based on these results, it has been proposed as a biomarker of drug-resistance. Other studies, however, report decreased levels of this cytokine in DRE [39]. Our conclusion is that there is no conclusive evidence in this regard although it is possible that IL-1β could be a valid marker for the detection of recent seizures.
3.2. Interleukin 6 (IL-6)
3.3. Interleukin 17 (IL-17)
A Prospective Study on patients with TLE [35] and 3 other case-control studies included in this systematic review [37,40,41] determined this IL but only the study by Alvin et al. [40] showed a decrease in their serum levels of patients with epilepsy compared to healthy controls. We consider that there is insufficient evidence to defend its role as a useful biomarker in clinical practice.
3.4. Other Interleukins
A significant additional number of ILs have eventually been studied, by case-control studies. We must highlight the study by Alvim et al. [40], which shows a decrease in serum levels of IL-2, IL-4, IL-6 or IL-10 in patients with epilepsy versus healthy controls. In the study by Ethemoglu et al. [43] IL-33 level was found higher in all the patients with epilepsy.
3.5. TNF-α
In total, 11 publications that are a part of our systematic review contain this molecule. Most of them did not demonstrate any appreciable distinctions between epilepsy sufferers and healthy controls. However, three of them—the TLE study by Wang et al. [35]., the Drug Resistant Epilepsy (DRE) study by Sokolova et al. [44]., and the chronic epilepsy study by Kothur et al. [36].—do indicate an increase in these levels in these individuals. According to this last study, there was an increase in CSF TNF-α during the acute stage of status epilepticus or FIRE-type encephalitis, and a decrease during the chronic phases.
3.6. Transforming Growth Factor Beta (TGF-β)
Several investigations have been carried out to establish a connection between these experimental findings and drug-resistant epilepsy. In a case-control study, Yu et al. [45]. demonstrated that individuals with resistant epilepsy had higher TGFβ levels in their CSF than patients with controlled seizures. Since this article was published in 2014, it has not been incorporated into the systematic review. There are no other studies on this biomarker included in the Review, so we cannot prove its validity in patients with epilepsy.
3.7. Toll Like Receptor 4 (TLR-4)
Three of the case-control studies included in this review have demonstrated the association of high levels of this biomarker with drug-resistant epilepsy [30,46,47] as well as its correlation with the number of seizures [46]. So, it can be proposed as a biomarker not only of refractoriness but also of the control of epilepsy.
3.8. HMGB1
In our review we have found up to 6 case-control studies that have shown that HMGB1 is increased in the blood of patients with epilepsy compared to healthy individuals [46,47,48,49,50]. One of these studies analyses concomitantly the levels of this molecule in CSF and serum and finds elevation of its levels also in CSF but without correlation with serum values [49].
A further question some research address is whether the increase just happens immediately following a seizure or if it lasts for hours afterwards. Blood samples were taken by Nass et al. [50]. at the start of the investigation, as well as 2, 6, and 24 hours following a generalised tonic-clonic seizure. The findings verified that HMGB1 was raised right away and continued to be raised for six hours.
Some studies attempt to characterize this biomarker in DRE. So, Yue et al. [46]. found that serum HMGB1 levels were higher in patients with DRE compared to drug-responsive epilepsy. In addition, there was a positive correlation between HMGB1 expression and seizure frequency.
Walker et al. [48] discovered that DRE patients’ serum had higher amounts of HMGB1 than did healthy controls and patients with drug-responsive epilepsy who had not experienced a seizure in at least six months. Still, no meaningful correlation was found with the epilepsy subtype or with the length or frequency of seizures during the preceding month. Kan et al. [47] also found a relationship between seizure frequency and duration.
Conflicting results were observed in only one study that was part of our review. Panina et al. [51]. discovered that, in contrast to a control group, patients with both controlled and refractory temporal lobe epilepsy had decreased serum HMGB1 levels. The authors contend that because their sample of patients had a wide range of epilepsy types and durations, their results are inconsistent with those of other studies.
These studies suggest that HMGB1 can be used as a biomarker of drug resistance in patients with epilepsy with a greater number of published studies supporting its use than other inflammatory markers included in this review.
3.9. Chemokines
In several studies of our review [36,41,42,52,53] some chemokines have been found to be increased. The one that has been identified in the largest number of studies is CCL2, a potent chemoattractant protein for monocytes, and hence is alternatively referred to as monocyte chemoattractant protein-1 (MCP-1). Elevated levels of CCL2 expression in an inflamed brain were associated with activation and recruitment of macrophages/microglia to the injury sites [54]. Česká et al. [42] compared cytokine levels in children with DRE with healthy control and control patients in both CSF and blood plasma, finding elevated levels of CCL2.
3.10. Soluble TNF-α Receptors
According to the study of Alvim et al. [40], individuals with epilepsy had greater plasma levels of sTNFr2 than controls, but lower levels of TNFα and sTNFr1. Accordingly, sTNFr2 plasma levels are a promising indicator of epilepsy activity.
4. Discussion
Our systematic review’s initial finding is that there isn’t any research with a high degree of evidence. Out of the twenty-one selected articles, nineteen are case-control studies, while one is a prospective study [35], and one is an uncontrolled case series study [53]. Though two of them analyse CSF at the same time [42,49], the great majority concentrate on serum analysis and another only analyses CSF [36].
On the other hand, there is significant variation across the patient cohorts utilised in the various studies regarding the type of epilepsies included in them, the patient inclusion criteria, and the total number of patients employed. Because of this, there is a lot of variation in the findings, which makes comparing these researches challenging.
Cross-cutting phenomena such as neuroinflammation can arise in autoimmune, post-traumatic, viral, structural, or even hereditary epilepsies. The idea that neuroinflammation may be one of the triggers through a cascade of release of inflammatory and peripheral mediators is strongly supported by the data gathered over the past 20 years [10,11,55,56,57].
Our work serves to summarize the state of research as well as the potential for use in clinical practice of inflammatory biomarkers in plasma and CSF from patients with epilepsy and especially their relationship with DRE.
This systematic review provides acceptable support for five biomarkers: TNF-a and some of its soluble receptors (sTNFr2), HMGB1 and its TLR-4, CCL2 and IL-33 (Table 2).
Certain autoimmune illnesses have been linked to increased amounts of the nuclear non-histone protein HMGB-1. Its two main receptors, TLR-4 and RAGE, mediate a biological process that stimulates inflammation [58] by starting the IL-1R/TLR-4 cascade, which produces some types of cytokines and raises the hyperexcitability of neurons [59]. Furthermore, because it stimulates the phosphorylation of the N-methyl-D-aspartate receptor (NMDAR) and raises the Ca2+ channel’s permeability, the HMGB1-TLR-4 axis is essential for neuronal hyperexcitability. The breakdown of the blood-brain barrier (BBB) can also result from the activation of the HMGB1-TLR-4 axis, allowing the permeation of activated lymphocytes, antibodies, inflammatory cytokines, or albumin from peripheral blood to the CNS, aggravating inflammation. Elevated HMGB1 levels have been associated with several CNS disorders [60,61,62,63]. The results of our review position this biomarker as the most supported by recent studies, especially in the characterization of DRE [46,48] and as a biomarker of recent epileptic activity [50].
TLR-4 is an innate immune system receptor of HMGB1 y that has been proposed as a promoter of epileptogenesis [64]. The results of our systematic review allow us to propose this molecule as a biomarker of DRE.
Another molecule of interest in our work is the Pro-inflammatory cytokine TNF-α. This molecule is secreted by activated astrocytes and microglia; it raises microglial glutamate levels and promotes Ca2+ entry into the cell similarly to IL-1β. However, TNF-α also causes the GABA receptor to endocytose, which depletes membrane inhibitory receptors and has an inhibitory effect. These conflicting effects of TNF-α on excitatory and inhibitory receptors may be a factor in neuronal hyperexcitability and, consequently, epileptogenesis [25]. Among the cytokines most researched in relation to epilepsy is TNF-α. Our systematic review finds several studies that correlate this serum biomarker with DRE or chronic epilepsy [35,36,44].
Perhaps of greater interest, it has the identification of some of the TNF-a receptors as biomarkers. There are two types of TNFα receptors: membrane-bound (TNFr1 and TNFr2) and soluble (sTNFr1 and sTNFr2). Some studies suggest that sTNFr1 and sTNFr2 better reflect TNFα activity because they are more stable than TNFα. The role of TNFα in epileptogenesis is a matter of debate. Some authors have suggested a proconvulsive role for this molecule [11]. The results of our review corroborate that sTNFr2 could be a promising biomarker related to epileptic activity [65].
Astrocytes, microglia, and endothelial cells secrete a family of proteins known as chemokines. By controlling neurotransmitter-releasing, voltage-dependent, or G-protein-dependent channels, they modulate neuronal excitability and are crucial in the entry of immune cells into the brain via certain G-protein-coupled receptors (GPCRs) [66,67]. This accumulating evidence suggests that chemokines and downstream signalling pathways mediate the interaction between neuroinflammation and epileptogenesis. Some chemokines such as CCL2, CCL3, lCCL4, Fractalkine (CX3CL1), CXCL13, and the corresponding chemokine receptor CCR2, CCR5, C-X-C receptor 4 (CXCR4), CXCR5, have been found elevated in the hippocampus in animal models of epilepsy [68]. Elevated levels of CCL2 expression in an inflamed brain were associated with activation and recruitment of macrophages/microglia to the injury sites [54]. The results of our systematic review only allow us to propose CCL2 as a biomarker of epilepsy [41,42,53], a potent chemoattractant protein for monocytes, and hence is alternatively referred to as monocyte chemoattractant protein-1 (MCP-1).
Even if some of the other molecules under examination were preceded by encouraging experimental trials, there doesn’t seem to be enough information about them at this moment to consider them as prospective biomarkers for persistent epilepsies.
So, Il-1b is a cytokine produced by the resident cells of central innate immunity. The binding of IL-1β to its receptor (IL-1R) activates nuclear factor κB (NF-κB) and three mitogen-activated protein kinase (MAPK) signaling pathways, all of which are involved in the production of other cytokines and in the upregulation of genes related to inflammation and generation of reactive oxygen species. These signaling pathways lead to the activation of TLR-4 [32,69]. On the other hand, IL-1β influences the entry of calcium through the Nmethyl-Aspartate Receptor (NMDA), reduces glutamate uptake by astrocytes, and increases glutamate release by glial cells [70]. Both TLR-4 and IL-1β are expressed at low levels within the brain under basal conditions but can increase rapidly during acute pathological conditions. Thus, elevated levels of IL-1β have been found after febrile seizures [71] and its role in epileptogenesis has been postulated [72]. However, our systematic review has found contradictory results that prevent us from including this molecule as a consolidated biomarker in epileptogenesis right now. Further studies will be needed to decide the true role of this biomarker.
IL-6 plays a fundamental role in regulating the inflammatory response and activating adaptive immunity [73]. Astrocytes and neurons synthesize this substance, which can be released by perivascular and brain endothelial cells in response to inflammatory and infectious stimuli. Some older studies not included in our review suggested that IL-6 could be elevated in CSF and plasma immediately after tonic-clonic seizures [74,75] or in patients with focal epilepsies [76], but our systematic review fails to corroborate the results of these studies.
Of the rest of the Interleukins, only the study by Ozlem et al. [43] showed us that IL-33 level was found higher in all the patients with epilepsy. IL-33, a novel member of the cytokine family associated with IL-1 is widely expressed from a wide range of cell types and tissues, including astrocytes, neurons, microglia and oligodendrocytes.
Finally, TGF-b is involved in cell proliferation, growth, and differentiation. Its role in neuroinflammation, however, is still unclear. The TGFβ type I receptor is more highly expressed in the cytoplasm of astrocytes, according to research on neocortical temporal lobe tissue from thirty epileptic patients [77]. It has also been suggested that blocking this receptor in vivo reduces the likelihood of epileptogenesis [78]. Our systematic review does not allow us to propose it as a biomarker of epilepsy in view of the current evidence.
The role of inflammatory markers as biomarkers of epilepsy has already been discussed in previous revisions, such as Zhang et al. [64]. The connection between two signalling pathways, the HMGB1/TLR4 and IL-1β/IL-1R1 pathways and epilepsy is the main topic of this review. The study also looks at mediators’ roles and their pathways of sequestration and acknowledges the need for more research on the possibility of cell-to-cell interference in neuroinflammation, including activated microglia, astrocytes, and neurons, as well as the function of pro-inflammatory substances like chemokines, cytokines, bioactive lipids and growth factors.
One of the main consequences of our study is the possible consideration of the TNF-a/sTNFr2 and HMGB1/TLR-4 axes as therapeutic targets in DRE. There is a previous systematic review on this topic [79]. Although the evidence found by these authors was also scarce, they conclude that the use of anti-IL-1, anti-IL-6, and anti-CD20 agents in patients with drug-resistant epilepsy and refractory status epilepticus has shown promising results and a good safety profile and that, concerning research perspectives, there is increasing interest in the potential use of anti-chemokine and anti-HMGB-1 agents. With regard to anti-TNF-a drugs, experience was limited to published case series in the treatment of Rasmussen’s encephalitis with Adalimumab [80,81]. Indeed, there is an ongoing clinical trial trying to evaluate the benefit of Adalimumab in patients with Rasmussen Encephalitis (ClinicalTrials.gov Identifier: NCT04003922).
Glycyrrhizin, an HMGB1 inhibitor medication, was noted as a possible treatment agent in the narrative review our group [11] published on the same subject because it has shown neuroprotective and antiepileptic benefits in many animal models of epilepsy [82,83] Nevertheless, this medication has not been evaluated in epilepsy clinical studies.
5. Conclusions
Despite the lack of strong scientific backing for the studies that are currently available, they do enable us to identify a number of neuroinflammation-related biomarkers that could be important in the early detection of DRE or could be connected to the level of control experienced by epilepsy patients. Some DAMP molecules like HMGB1 and its membrane receptors (TLR-4) are among them, as are cytokines like TNF-a and some of its soluble receptors like sTNFr2 or IL-33, as well as chemokines like CCL2. These biomarkers may occasionally be suggested as potential treatment targets for particular forms of epilepsy. Future research will enable us to better characterise its actual value.
Author Contributions
Conceptualization: MJAC, PJSC, MNP and GET X.X.; methodology, MJAC and PJSC.; software, MJAC, PJSC, NLCP, BOM and MNP.; validation, PCG, YLM and GGM.; formal analysis, MJAC and PJSC; investigation, MJAC and PJSC; resources, PJSC.; data curation, MJAC and PJSC.; writing—original draft preparation, MJAC and PJSC; writing—review and editing, MJAC and PJSC; visualization, PCG, GGM, YLM, BOM, GET and MNP.; supervision, PJSC; project administration, PJSC.; funding acquisition, PJSC. All authors have read and agreed to the published version of the manuscript.” Please turn to the CRediT taxonomy for the term explanation. Authorship must be limited to those who have contributed substantially to the work reported.
Funding
This research was funded by Alianza Andalucía Neuro-RECA – Roche en Neurología Médica de Precisión; Andalusian Network of Clinical and Translational Research in Neurology (Neuro-RECA) of the Ministry of Health and Families of Andalusia (Code: RIC-0111-2019) and Project “Personalized medicine in neurological diseases through the application of biomarkers for improving the diagnosis, prognosis and treatment of the patient”. Research projects with public-private collaboration. Fundacion Progreso y Salud. Ministry of Health and Families of Andalusia. (Code: PIP-0123-2022). GET is under a contract of the “Nicolás Monardes” programme from the Andalusian Health Service, Andalusian Regional Ministry of Health and Consumption.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue Homeostasis and Inflammation. Annu Rev Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef] [PubMed]
- Cervellati, C.; Trentini, A.; Pecorelli, A.; Valacchi, G.; Valacchi, G.; Valacchi, G. Inflammation in Neurological Disorders: The Thin Boundary between Brain and Periphery. Antioxidants Redox Signal. 2020, 33, 191–210. [Google Scholar] [CrossRef]
- Govindarajan, V.; De Rivero Vaccari, J.P.; Keane, R.W. Role of inflammasomes in multiple sclerosis and their potential as therapeutic targets. J Neuroinflammation. 2020, 17, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Frank-Cannon, T.C.; Alto, L.T.; McAlpine, F.E.; Tansey, M.G. Does neuroinflammation fan the flame in neurodegenerative diseases? Mol Neurodegener. 2009, 4, 47. [Google Scholar] [CrossRef] [PubMed]
- Adamu, A.; Li, S.; Gao, F.; Xue, G. The role of neuroinflammation in neurodegenerative diseases: current understanding and future therapeutic targets. Front Aging Neurosci. 2024, 16. [Google Scholar] [CrossRef]
- Solleiro-Villavicencio, H.; Rivas-Arancibia, S. Effect of chronic oxidative stress on neuroinflammatory response mediated by CD4+T cells in neurodegenerative diseases. Front Cell Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Jiang, J.X.; Fewings, N.; Dervish, S.; Fois, A.F.; Duma, S.R.; Silsby, M.; et al. Novel Surrogate Markers of CNS Inflammation in CSF in the Diagnosis of Autoimmune Encephalitis. Front Neurol. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Wesselingh, R.; Butzkueven, H.; Buzzard, K.; Tarlinton, D.; O’Brien, T.J.; Monif, M.; et al. Innate Immunity in the Central Nervous System: A Missing Piece of the Autoimmune Encephalitis Puzzle? Front Immunol. 2019, 10, 1–14. [Google Scholar] [CrossRef]
- Wang, S.; Guan, Y.; Li, T. The Potential Therapeutic Role of the HMGB1-TLR Pathway in Epilepsy. Curr Drug Targets. 2021, 22, 171–182. [Google Scholar] [CrossRef]
- Matin, N.; Tabatabaie, O.; Falsaperla, R.; Lubrano, R.; Pavone, P.; Mahmood, F.; et al. Epilepsy and innate immune system: A possible immunogenic predisposition and related therapeutic implications. Hum Vaccin Immunother. 2015, 11, 2021–2029. [Google Scholar] [CrossRef]
- Aguilar-Castillo, M.J.; Cabezudo-García, P.; Ciano-Petersen, N.L.; García-Martin, G.; Marín-Gracia, M.; Estivill-Torrús, G.; et al. Immune Mechanism of Epileptogenesis and Related Therapeutic Strategies. Biomedicines. 2022, 10, 716. [Google Scholar] [CrossRef] [PubMed]
- Marchi, N.; Granata, T.; Janigro, D. Inflammatory pathways of seizure disorders. Trends Neurosci. 2014, 37, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Rüegg, S. Introduction. Immunity and Inflammation in Epilepsy. Epilepsia. 2011, 52 Suppl 3, 1–4. [Google Scholar] [CrossRef]
- Vezzani, A.; Rüegg, S. The pivotal role of immunity and inflammatory processes in epilepsy is increasingly recognized: introduction. Epilepsia. 2011, 52, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Friedman, A.; Dingledine, R.J. The role of inflammation in epileptogenesis. Neuropharmacology. 2013, 69, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Villasana-Salazar, B.; Vezzani, A. Neuroinflammation microenvironment sharpens seizure circuit. Neurobiol Dis. 2023, 178. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A. Epilepsy and Inflammation in the Brain: Overview and Pathophysiology. Epilepsy Curr. 2014, 14, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Orsini, A.; Foiadelli, T.; Costagliola, G.; Michev, A.; Consolini, R.; Vinci, F.; et al. The role of inflammatory mediators in epilepsy: Focus on developmental and epileptic encephalopathies and therapeutic implications. Epilepsy Res. 2021, 172. [Google Scholar] [CrossRef] [PubMed]
- Patterson, K.P.; Brennan, G.P.; Curran, M.; Kinney-Lang, E.; Dubé, C.; Rashid, F.; et al. Rapid, Coordinate Inflammatory Responses after Experimental Febrile Status Epilepticus: Implications for Epileptogenesis. eneuro. 2015, 2, ENEURO.0034–152015. [Google Scholar] [CrossRef]
- Rana, A.; Musto, A.E. The role of inflammation in the development of epilepsy. J Neuroinflammation. 2018, 15, 144. [Google Scholar] [CrossRef]
- Sanz, P.; Rubio, T.; Garcia-Gimeno, M.A. Neuroinflammation and Epilepsy: From Pathophysiology to Therapies Based on Repurposing Drugs. Int J Mol Sci. 2024, 25, 4161. [Google Scholar] [CrossRef] [PubMed]
- Kamali, A.N.; Zian, Z.; Bautista, J.M.; Hamedifar, H.; Hossein-Khannazer, N.; Hosseinzadeh, R.; et al. The Potential Role of Pro-Inflammatory and Anti-Inflammatory Cytokines in Epilepsy Pathogenesis. Endocrine, Metab Immune Disord - Drug Targets. 2021, 21, 1760–1774. [Google Scholar]
- Soltani Khaboushan, A.; Yazdanpanah, N.; Rezaei, N. Neuroinflammation and Proinflammatory Cytokines in Epileptogenesis. Mol Neurobiol. 2022, 59, 1724–1743. [Google Scholar] [CrossRef]
- Hulkkonen, J.; Koskikallio, E.; Rainesalo, S.; Keränen, T.; Hurme, M.; Peltola, J. The balance of inhibitory and excitatory cytokines is differently regulated in vivo and in vitro among therapy resistant epilepsy patients. Epilepsy Res. 2004, 59(2–3):199–205.
- Banote, R.K.; Akel, S.; Zelano, J. Blood biomarkers in epilepsy. Acta Neurol Scand. 2022 Apr. [CrossRef]
- Janmohamed, M.; Brodie, M.J.; Kwan, P. Pharmacoresistance – Epidemiology, mechanisms, and impact on epilepsy treatment. Neuropharmacology. 2020 May;168. [CrossRef]
- Zhang, Z.; Liu, Q.; Liu, M.; Wang, H.; Dong, Y.; Ji, T.; et al. Upregulation of HMGB1-TLR4 inflammatory pathway in focal cortical dysplasia type, I.I. J Neuroinflammation. 2018, 15(1). [CrossRef]
- Bazhanova, E.D.; Kozlov, A.A.; Litovchenko A, V. Mechanisms of Drug Resistance in the Pathogenesis of Epilepsy: Role of Neuroinflammation. A Literature Review. Brain Sci. 2021, 11(5). [CrossRef]
- Chen, Y.; Nagib, M.M.; Yasmen, N.; Sluter, M.N.; Littlejohn, T.L.; Yu, Y.; et al. Neuroinflammatory mediators in acquired epilepsy: an update. Inflamm Res. 2023, 72, 683–701. [Google Scholar] [CrossRef] [PubMed]
- Kamaşak, T.; Dilber, B.; Yaman, S.Ö.; Durgut, B.D.; Kurt, T.; Çoban, E.; et al. HMGB-1, TLR4, IL-1R1, TNF-α, and IL-1β: novel epilepsy markers? Epileptic Disord. 2020, 22, 183–193. [Google Scholar] [CrossRef] [PubMed]
- French, J.A.; Cole, A.J.; Faught, E.; Theodore, W.H.; Vezzani, A.; Liow, K.; et al. Safety and Efficacy of Natalizumab as Adjunctive Therapy for People With Drug-Resistant Epilepsy. Neurology. 2021, 97, e1757–67. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Maroso, M.; Balosso, S.; Sanchez, M.A.; Bartfai, T. IL-1 receptor/Toll-like receptor signaling in infection, inflammation, stress and neurodegeneration couples hyperexcitability and seizures. Brain Behav Immun. 2011, 25, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.F.; Korevaar, D.A.; Altman, D.G.; Bruns, D.E.; Gatsonis, C.A.; Hooft, L.; et al. STARD 2015 guidelines for reporting diagnostic accuracy studies: explanation and elaboration. [CrossRef]
- Miller, J. The Scottish Intercollegiate Guidelines Network (SIGN). Br J Diabetes Vasc Dis. 2002, 2, 47–49. [Google Scholar] [CrossRef]
- Wang, B.; Li, Q.; Wang, H.; Du, X.; Lai, Q.; Li, X.; et al. TNF-α: A serological marker for evaluating the severity of hippocampal sclerosis in medial temporal lobe epilepsy? J Clin Neurosci. 2024, 123, 123–129. [Google Scholar] [CrossRef]
- Kothur, K.; Bandodkar, S.; Wienholt, L.; Chu, S.; Pope, A.; Gill, D.; et al. Etiology is the key determinant of neuroinflammation in epilepsy: Elevation of cerebrospinal fluid cytokines and chemokines in febrile infection-related epilepsy syndrome and febrile status epilepticus. Epilepsia. 2019, 60, 1678–1688. [Google Scholar] [CrossRef]
- Gledhill, J.M.; Brand, E.J.; Pollard, J.R. ; St. Clair, R.D.; Wallach, T.M.; Crino, P.B. Association of Epileptic and Nonepileptic Seizures and Changes in Circulating Plasma Proteins Linked to Neuroinflammation. Neurology. 2021, 96, E1443–52. [Google Scholar]
- Choi, J.; Kim, S.Y.; Kim, H.; Lim, B.C.; Hwang, H.; Chae, J.H.; et al. Serum α-synuclein and IL-1β are increased and correlated with measures of disease severity in children with epilepsy: potential prognostic biomarkers? BMC Neurol. 2020, 20, 85. [Google Scholar] [CrossRef] [PubMed]
- Saengow, V.E.; Chiangjong, W.; Khongkhatithum, C.; Changtong, C.; Chokchaichamnankit, D.; Weeraphan, C.; et al. Proteomic analysis reveals plasma haptoglobin, interferon-γ, and interleukin-1β as potential biomarkers of pediatric refractory epilepsy. Brain Dev. 2021, 43, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Alvim, M.K.M.; Morita-Sherman, M.E.; Yasuda, C.L.; Rocha, N.P.; Vieira, É.L.; Pimentel-Silva, L.R.; et al. Inflammatory and neurotrophic factor plasma levels are related to epilepsy independently of etiology. Epilepsia. 2021, 62, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Milano, C.; Montali, M.; Barachini, S.; Burzi, I.S.; Pratesi, F.; Petrozzi, L.; et al. Increased production of inflammatory cytokines by circulating monocytes in mesial temporal lobe epilepsy: A possible role in drug resistance. J Neuroimmunol. 2024, 386, 578272. [Google Scholar] [CrossRef] [PubMed]
- Česká, K. ; Papež J, Ošlejšková, H. ; Slabý, O.; Radová, L.; Loja, T.; et al. CCL2/MCP-1, interleukin-8, and fractalkine/CXC3CL1: Potential biomarkers of epileptogenesis and pharmacoresistance in childhood epilepsy. Eur J Paediatr Neurol. 2023, 46, 48–54. [Google Scholar]
- Ethemoglu, O.; Calık, M.; Koyuncu, I.; Ethemoglu, K.B.; Göcmen, A.; Güzelcicek, A.; et al. Interleukin-33 and oxidative stress in epilepsy patients. Epilepsy Res. 2021, 176, 106738. [Google Scholar] [CrossRef]
- Sokolova T, V. , Zabrodskaya, Y. M.; Litovchenko A V., Paramonova, N.M.; Kasumov, V.R.; Kravtsova S V., et al. Relationship between Neuroglial Apoptosis and Neuroinflammation in the Epileptic Focus of the Brain and in the Blood of Patients with Drug-Resistant Epilepsy. Int J Mol Sci. 2022, 23, 12561. [Google Scholar]
- Yu, W.; Zou, Y.; Du, Y.; Luo, J.; Zhang, M.; Yang, W.; et al. Altered cerebrospinal fluid concentrations of TGFβ1 in patients with drug-resistant epilepsy. Neurochem Res. 2014, 39, 2211–2217. [Google Scholar] [CrossRef] [PubMed]
- Yue, Z.; Tang, J.; Peng, S.; Cai, X.; Rong, X.; Yang, L. Serum concentration of high-mobility group box 1, Toll-like receptor 4 as biomarker in epileptic patients. Epilepsy Res. 2023, 192, 107138. [Google Scholar] [CrossRef]
- Kan, M.; Song, L.; Zhang, X.; Zhang, J.; Fang, P. Circulating high mobility group box-1 and toll-like receptor 4 expressions increase the risk and severity of epilepsy. Brazilian J Med Biol Res. 2019, 52(7). [CrossRef]
- Walker, L.E.; Sills, G.J.; Jorgensen, A.; Alapirtti, T.; Peltola, J.; Brodie, M.J.; et al. High-mobility group box 1 as a predictive biomarker for drug-resistant epilepsy: A proof-of-concept study. Epilepsia. 2022, 63(1). [CrossRef]
- Wang, N.; Liu, H.; Ma, B.; Zhao, T.; Chen, Y.; Yang, Y.; et al. CSF high-mobility group box 1 is associated with drug-resistance and symptomatic etiology in adult patients with epilepsy. Epilepsy Res. 2021, 177, 106767. [Google Scholar] [CrossRef] [PubMed]
- Nass, R.D.; Wagner, M.; Surges, R.; Holdenrieder, S. Time courses of HMGB1 and other inflammatory markers after generalized convulsive seizures. Epilepsy Res. 2020, 162. [Google Scholar] [CrossRef] [PubMed]
- Panina, Y.S.; Timechko, E.E.; Usoltseva, A.A.; Yakovleva, K.D.; Kantimirova, E.A.; Dmitrenko D, V. Biomarkers of Drug Resistance in Temporal Lobe Epilepsy in Adults. Metabolites. 2023, 13, 83. [Google Scholar] [CrossRef] [PubMed]
- Gakharia, T.; Bakhtadze, S.; Lim, M.; Khachapuridze, N.; Kapanadze, N. Alterations of Plasma Pro-Inflammatory Cytokine Levels in Children with Refractory Epilepsies. Children. 2022, 9, 1506. [Google Scholar] [CrossRef] [PubMed]
- Bronisz, E.; Cudna, A.; Wierzbicka, A.; Kurkowska-Jastrzębska, I. Serum Proteins Associated with Blood–Brain Barrier as Potential Biomarkers for Seizure Prediction. Int J Mol Sci. 2022, 23, 14712. [Google Scholar] [CrossRef] [PubMed]
- Selenica M-LB, Alvarez, J. A.; Nash, K.R.; Lee, D.C.; Cao, C.; Lin, X.; et al. Diverse activation of microglia by chemokine (C-C motif) ligand 2 overexpression in brain. J Neuroinflammation. 2013, 10, 856.
- Oby, E.; Janigro, D. The blood-brain barrier and epilepsy. Epilepsia. 2006, 47, 1761–1774. [Google Scholar] [CrossRef] [PubMed]
- Webster, K.M.; Sun, M.; Crack, P.; O’Brien, T.J.; Shultz, S.R.; Semple, B.D. Inflammation in epileptogenesis after traumatic brain injury. J Neuroinflammation. 2017, 14(1). [CrossRef]
- Singh, S.; Singh, T.G.; Rehni, A.K. An Insight into Molecular Mechanisms and Novel Therapeutic Approaches in Epileptogenesis. CNS Neurol Disord Drug Targets. 2020, 19, 750–779. [Google Scholar] [CrossRef] [PubMed]
- Terrone, G.; Balosso, S.; Pauletti, A.; Ravizza, T.; Vezzani, A. Inflammation and reactive oxygen species as disease modifiers in epilepsy. Neuropharmacology. 2020, 167. [Google Scholar] [CrossRef]
- Maroso, M.; Balosso, S.; Ravizza, T.; Liu, J.; Bianchi, M.E.; Vezzani, A. Interleukin-1 type 1 receptor/Toll-like receptor signalling in epilepsy: The importance of IL-1beta and high-mobility group box 1. Journal of Internal Medicine. J Intern Med; 2011, 319–326.
- Ravizza, T.; Terrone, G.; Salamone, A.; Frigerio, F.; Balosso, S.; Antoine, D.J.; et al. High Mobility Group Box 1 is a novel pathogenic factor and a mechanistic biomarker for epilepsy. Brain Behav Immun. 2018, 72, 14–21. [Google Scholar] [CrossRef]
- Paudel, Y.N.; Semple, B.D.; Jones, N.C.; Othman, I.; Shaikh, M.F. High mobility group box 1 (HMGB1) as a novel frontier in epileptogenesis: from pathogenesis to therapeutic approaches. J Neurochem. 2019, 151, 542–557. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Yang, L.; Liu, X.; Feng, Y.; Pang, Z.; Lin, Y. Hmgb1/cxcl12-mediated immunity and TH17 cells might underlie highly suspected autoimmune epilepsy in elderly individuals. Neuropsychiatr Dis Treat. 2020, 16, 1285–1293. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, S.; Wales, E.; Zaben, M. The Role of HMGB1 in Traumatic Brain Injury—Bridging the Gap Between the Laboratory and Clinical Studies. Curr Neurol Neurosci Rep. 2021, 21, 75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, F.; Zhai, F.; Liang, S. Role of HMGB1/TLR4 and IL-1β/IL-1R1 Signaling Pathways in Epilepsy. Front Neurol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Alvim MMKM, Morita-Sherman, M. E.M.; Yasuda, C.L.; Rocha, N.N.P.; Vieira, É.É.L.; Pimentel-Silva, L.R.; et al. Inflammatory and neurotrophic factor plasma levels are related to epilepsy independently of etiology. Epilepsia. 2021, 62, epi17023.
- Yung, S.C.; Farber, J.M. Chemokines. Second Edi. Elsevier Inc.; 2013. [CrossRef]
- Ono, S.J.; Nakamura, T.; Miyazaki, D.; Ohbayashi, M.; Dawson, M.; Toda, M. Chemokines: Roles in leukocyte development, trafficking, and effector function. J Allergy Clin Immunol. 2003, 111, 1185–1199. [Google Scholar] [CrossRef] [PubMed]
- Arisi, G.M.; Foresti, M.L.; Katki, K.; Shapiro, L.A. Increased CCL2, CCL3, CCL5, and IL-1β cytokine concentration in piriform cortex, hippocampus, and neocortex after pilocarpine-induced seizures. J Neuroinflammation. 2015, 12, 129. [Google Scholar] [CrossRef] [PubMed]
- Vezzani, A.; Viviani, B. Neuromodulatory properties of inflammatory cytokines and their impact on neuronal excitability. Neuropharmacology. 2015, 96, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Viviani, B.; Bartesaghi, S.; Gardoni, F.; Vezzani, A.; Behrens, M.M.; Bartfai, T.; et al. Interleukin-1β Enhances NMDA Receptor-Mediated Intracellular Calcium Increase through Activation of the Src Family of Kinases. J Neurosci. 2003, 23, 8692–8700. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Chen, R.; Zhang, H.; Jiang, C.; Gong, J. Cerebrospinal fluid neuron specific enolase, interleukin-1β and erythropoietin concentrations in children after seizures. Child’s Nerv Syst. 2017, 33, 805–811. [Google Scholar] [CrossRef]
- Li, G.; Bauer, S.; Nowak, M.; Norwood, B.; Tackenberg, B.; Rosenow, F.; et al. Cytokines and epilepsy. Seizure. 2011, 20, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Hurst, S.M.; Wilkinson, T.S.; McLoughlin, R.M.; Jones, S.; Horiuchi, S.; Yamamoto, N.; et al. IL-6 and Its Soluble Receptor Orchestrate a Temporal Switch in the Pattern of Leukocyte Recruitment Seen during Acute Inflammation. Immunity. 2001, 14, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Peltola, J.; Palmio, J.; Korhonen, L.; Suhonen, J.; Miettinen, A.; Hurme, M.; et al. Interleukin-6 and Interleukin-1 receptor antagonist in cerebrospinal fluid from patients with recent tonic–clonic seizures. Epilepsy Res. 2000, 41, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Peltola, J.; Hurme, M.; Miettinen, A.; Keränen, T. Elevated levels of interleukin-6 may occur in cerebrospinal fluid from patients with recent epileptic seizures. Epilepsy Res. 1998, 31, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Alapirtti, T.; Rinta, S.; Hulkkonen, J.; Mäkinen, R.; Keränen, T.; Peltola, J. Interleukin-6, interleukin-1 receptor antagonist and interleukin-1beta production in patients with focal epilepsy: A video–EEG study. J Neurol Sci. 2009, 280(1–2):94–7.
- Lu, Y.; Xue, T.; Yuan, J.; Li, Y.; Wu, Y.; Xi, Z.; et al. Increased expression of TGFβ type I receptor in brain tissues of patients with temporal lobe epilepsy. Clin Sci. 2009, 117, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Ivens, S.; Kaufer, D.; Flores, L.P.; Bechmann, I.; Zumsteg, D.; Tomkins, O.; et al. TGF-β receptor-mediated albumin uptake into astrocytes is involved in neocortical epileptogenesis. Brain. 2007, 130, 535–547. [Google Scholar] [CrossRef] [PubMed]
- Costagliola, G.; Depietri, G.; Michev, A.; Riva, A.; Foiadelli, T.; Savasta, S.; et al. Targeting Inflammatory Mediators in Epilepsy: A Systematic Review of Its Molecular Basis and Clinical Applications. Front Neurol. 2022, 13. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, S.; Villeneuve, N.; Trébuchon, A.; Kaphan, E.; Lepine, A.; McGonigal, A.; et al. Anti-tumor necrosis factor alpha therapy (adalimumab) in Rasmussen’s encephalitis: An open pilot study. Epilepsia. 2016, 57, 956–966. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, S.; Boucraut, J.; Bartolomei, F. Medical treatment of Rasmussen’s Encephalitis: A systematic review. Rev Neurol (Paris). 2022. [CrossRef]
- Paudel, Y.N.; Khan, S.U.; Othman, I.; Shaikh, M.F. Naturally Occurring HMGB1 Inhibitor, Glycyrrhizin, Modulates Chronic Seizures-Induced Memory Dysfunction in Zebrafish Model. ACS Chem Neurosci. 2020, 12(18). [CrossRef]
- Li Y jun, Wang, L. ; Zhang, B.; Gao, F.; Yang, C.M. Glycyrrhizin, an HMGB1 inhibitor, exhibits neuroprotective effects in rats after lithium-pilocarpine-induced status epilepticus. J Pharm Pharmacol. 2019, 71, 390–399.
- Walker, L.E.; Sills, G.J.; Jorgensen, A.; Alapirtti, T.; Peltola, J.; Brodie, M.J.; et al. High-mobility group box 1 as a predictive biomarker for drug-resistant epilepsy: A proof-of-concept study. Epilepsia. 2022, 63, e1–6. [Google Scholar] [CrossRef] [PubMed]
- Kamaşak, T.; Dilber, B.; Yaman, S.Ö.; Durgut, B.D.; Kurt, T.; Çoban, E.; et al. HMGB-1, TLR4, IL-1R1, TNF-α, and IL-1β: novel epilepsy markers? Epileptic Disord. 2020, 22, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Kegler, A.; Pascotini, E.T.; Caprara, A.L.F.; Arend, J.; Gabbi, P.; Duarte, M.M.; et al. Relationship between seizure type, metabolic profile, and inflammatory markers in blood samples of patients with epilepsy. Epileptic Disord. 2021, 23, 74–84. [Google Scholar] [CrossRef] [PubMed]
- Mochol, M.; Taubøll, E.; Aukrust, P.; Ueland, T.; Andreassen, O.A.; Svalheim, S. Interleukin 18 (IL-18) and its binding protein (IL-18BP) are increased in patients with epilepsy suggesting low-grade systemic inflammation. Seizure. 2020, 80, 221–225. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
PRISMA 2020 Flowchart. *Consider, if feasible to do so, reporting the number of records identified from each database or register searched (rather than the total number across all databases/registers). **If automation tools were used, indicate how many records were excluded by a human and how many were excluded by automation tools. From: Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021;372:n71. doi: 10.1136/bmj.n71.
Figure 1.
PRISMA 2020 Flowchart. *Consider, if feasible to do so, reporting the number of records identified from each database or register searched (rather than the total number across all databases/registers). **If automation tools were used, indicate how many records were excluded by a human and how many were excluded by automation tools. From: Page MJ, McKenzie JE, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 2021;372:n71. doi: 10.1136/bmj.n71.
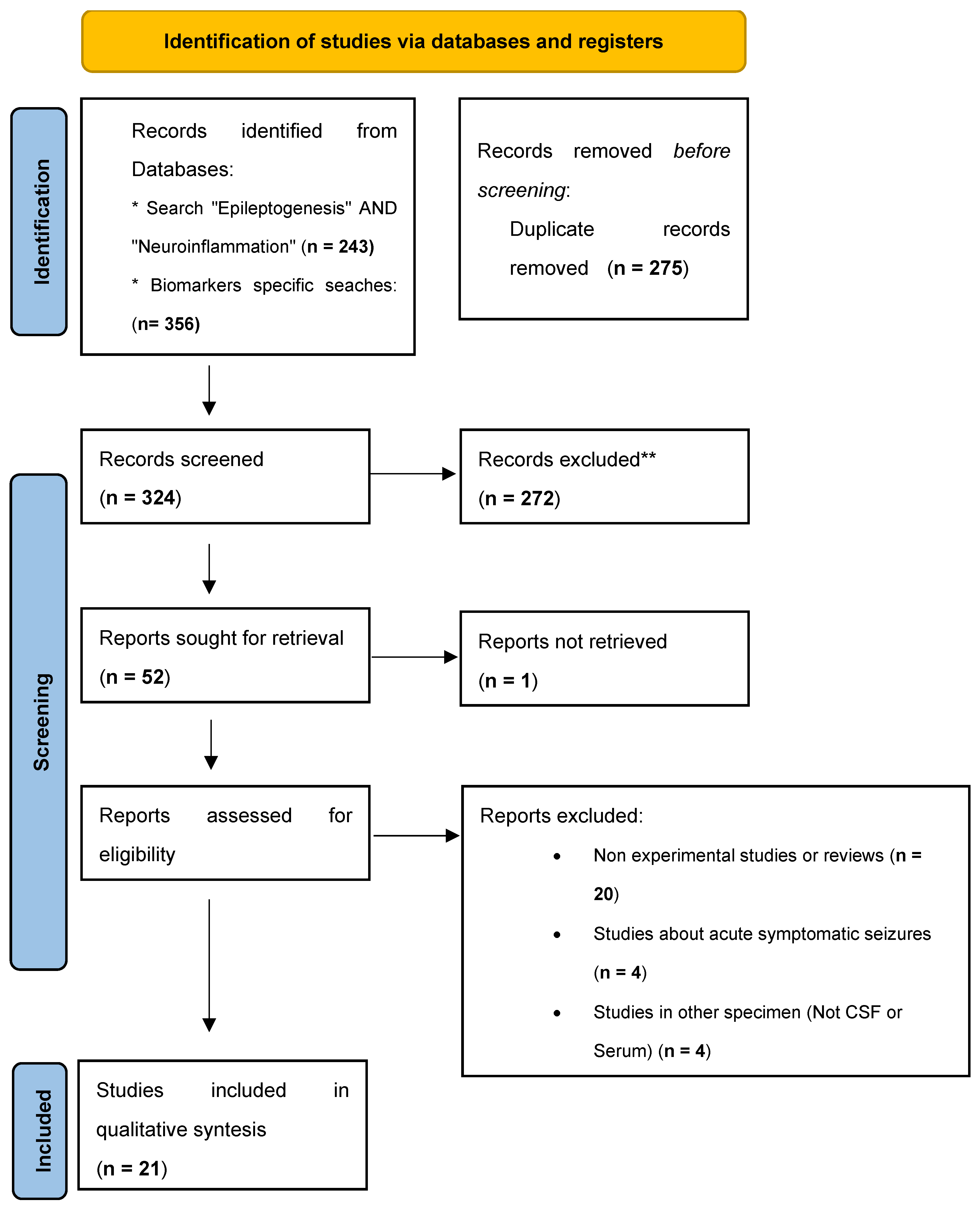

Table 1.
Summary of the studies included in the Systematic Review for qualitative analysis.
| Reference | Biomarkers studied | Sample Type | Type of study | Results | N | Level of evidence |
|---|---|---|---|---|---|---|
| Kothur et al. [36] | IL-1ra, GM-CSF, IL-1β, TNF-α, IL-2,IL-4, IL-6, IL-8, IL-10, IL-13, IL-17A, IFN-γ, CCL2/MCP-1, CCL5/RANTES, CXCL1/GRO, CXCL10/IP-10,CCL3/MIP-1a, CCL4/MIP-1b, IL-12 (p40), IL-12 (p70), IFN-α, G-CSF, CCL11/eotaxin. IL-21, IL-23, CXCL13/BCA-1, CCL17/TARC, CCL21/6Ckine, CXCL12/SDF-1. CXCL9/MIG, CXCL11/I-TAC, and CCL19/MIP-3b | CSF | Case-control | TNF-α and CCL19 were mildly elevated in chronic epilepsy. | Patients with FIRES/FIRES-related disorders (FRD; n = 6), FSE (n = 8), afebrile status epilepticus (ASE; n = 8), and chronic epilepsy (n = 21) | 2- |
| Yue et al. [46] | HMGB1 y TLR4 | Serum | Case-control | HMGB1 y TLR4 were elevated in chronic epilepsy | 72 epilepsy patients diagnosed with epilepsy vs 43 healthy controls | 2 |
| Jieun et al. [38] | α-synuclein, IFN-β, IFN-γ, IL-1β, IL-6, IL-10 and TNF-α | Serum | Case-control | α-synuclein levels were significantly increased in children with epilepsy. Serum IL-1β levels showed significant correlation only with drug resistance in children with epilepsy. |
115 epilepsy patients having afebrile seizure attacks within the last 48 h vs vs 146 healthy controls. | 2+ |
| Saengow et al. [39] | gamma (IFN-c), IL-1β, and TNF-a | Serum | Case-control | IL-1β level was significantly decreased in patients with DRE. IFN-c level was significantly increased in patients with DRE. TNF-a showed no statistical change between groups. |
65 patients with drug-resistant epilepsy vs 6 healthy controls | 2- |
| Walker et al. [84] | HMGB1 | Serum | Case-control | Patients with drug-resistant epilepsy had higher levels of HMGB1 than both healthy controls and patients with drug-responsive epilepsy | 65 patients with drug-resistant epilepsy vs 74 healthy controls |
2+ |
| Kamaşak et al. [85] | HMGB-1, TLR-4, IL-1R1, TNF-a, IL-1β | Serum | Case-control | Significantly higher levels of HMGB-1, TLR-4, TNF-a and IL-1β in the severe epilepsy group than in the other two groups | 28 children with DRE vs 29 children with controlled epilepsy vs 27 healthy controls | 2 |
| Aline et al. [86] | TNF-a, Caspase and Lipid factors | Serum | Case-control | No significant differences. But patients with generalized epilepsy demonstrated a significant correlation between TNF-α and caspase 8, caspase 3, and Picogreen. | 43 epileptic patients vs 41 healthy controls | 2 |
| Alvim et al. [40] | IL-1, IL-2, IL-4, IL-6, IL-10, IL-17, IFNγ, TNF-α, soluble TNF receptor 1 (sTNFr1), sTNFr2, BDNF, neurotrophic factor 3 (NT3), NT4/5, ciliary neurotrophic factor (CNTF), nerve growth factor (NGF), and glial cell line-derived neurotrophic factor (GDNF). | Serum | Case-control | The plasma levels of BDNF, NT3, NGF, and sTNFr2 were higher, whereas IL-2, IL-4, IL-6, IL-10, IL-17, IFNγ, TNFα, CNTF, and sTNFr1 were lower in patients than controls. The molecule sTNFr2 was the best marker to discriminate patients from controls also differing between patients with frequent and infrequent seizures. |
446 patients with epilepsy vs 166 healthy controls. | 2+ |
| Minchen et al. [47] | HMGB1 and TLR4 | Serum | Case-control | HMGB1 and TLR4 levels were higher in epilepsy patients compared with controls HMGB1 and TLR4 expressions were correlated with higher possibility of drugs resistance. | 105 epilepsy patients vs 100 healthy controls | 2 |
| Ethemoglu et al. [43] . | IL-33 | Serum | Case-control | IL-33 level was found higher in all the patients with epilepsy compared to the control group. | 60 patients with epilepsy (21 patients with treatment-resistant epilepsy and 39 patients with well-controlled epilepsy) vs 35 control subjects | 2 |
| Panina et al. [51] | BDNF, TNF-a, HMGB1 and NTRK2 | Serum | Case-control | A decrease in the concentration of BDNF, TNF-a, and HMGB1 was registered in the group of patients with TLE compared with the control group. |
166 patients with epilepsy (49 with treatment-resistant epilepsy and 117 patients with well-controlled epilepsy) vs 203 controls. | 2 |
| Wang et al. [35] | IL-1β, IL-5, IL-6, IL-8, IL-17, IFN-γ and TNF-α | Serum | Prospective, population-based study | The level of TNF-α in the mTLE-HS-P group was significantly higher than that of the patients in the mTLE-HS-N and healthy control groups, and the level of TNF-α in the patients in the mTLE-HS-N group was significantly higher than that of the patients in the healthy control group. | 71 patients with medial TLE vs 20 controls | 2++ |
| Milano et al. [41] | IL-6, TNF-α, IL-33, IL-8, CCL2, IL-13, IL-1β, IFN-γ, IL-1Ra, CCL3, IL-4, CCL4, IL-5, IL-1α, IL-17 A, IL-18, IL-33r, IL-1RII, IL-1RI | Serum | Case-control | Levels of CCL2, CCL3 and IL-8 were elevated in the serum of patients with epilepsy compared to healthy controls, without differences between drugs-resistant and drug-sensitive patients. | 47 patients diagnosed with MTLE vs 25 healthy controls | 2 |
| Sokolova et al. [44] | IL-1RA, interferon IFN-, IL-10 IL-2, IL-8, IL-7, TNF-α, IL-4, sCD40L | Serum | Case-control. | The level of the immunoregulatory cytokine IL-2 and the chemoattractant proinflammatory IL-8 was decreased in DRE patients. Proinflammatory cytokines (TNF-α, IL-4, sCD40L) was increased. |
6 DRE patients vs 5 healthy controls. | 2- |
| Wang et al. [49] | HMGB1 | CSF and Serum | Case-control | The CSF HMGB1 concentrations were significantly higher in the DRE vs the other groups. Patients with symptomatic etiology showed significantly high levels of CSF HMGB1. Patients without remission expressed elevated levels of CSF HMGB1 at one-year follow-up. CSF HMGB1 levels were positively associated with seizure frequency. |
27 DRE patients, 56 Newly diagnosed epileptic patients and 22 other non-inflammatory neurological disorders | 2- |
| Mochol et al. [87] | IL-18; Interleukine 18 binding protenin (IL-18BP) | Serum | Case-control | Increased serum levels of IL-18 and IL-18BP in epilepsy patients. | 119 patients with epilepsy and 80 healthy controls | 2 |
| Nass et al. [50] | c-reactive protein (CRP), HMGB1, S100, RAGE, ICAM1 and MMP9 | Serum | Case Series | Rapid postictal increase of HMGB1 and S100. |
28 patients with Epilepsy with Generalized Seizures. | 3 |
| Gakharia et al. [52] | CCL2, CCL4 and CCL11 and PGE2 | Serum | Case-control | High CCL11 and PGE2 levels correlated with their seizure frequency and epilepsy severity. | 40 epileptic patients (20 DRE and 20 Controlled epilepsy) vs 16 healthy controls. | 2 |
| Bronisz et al. [53] | MMP-9, MMP-2, CCL-2, S100B, TIMP-1, TIMP-2, ICAM-1, TSP-2, P-selectin) |
Serum | Case series | Levels of MMP-2, MMP-9, and CCL-2 were found to influence seizure count in 1, 3, 6, and 12 months of observation. | 49 patients with epilepsy. | 3 |
| Gledhill et al. [37] | CRP, calbindin, cytokeratin-8, eotaxin, eotaxin-2, eotaxin-3, granulocyte-macrophage colony-stimulating factor, ICAM-1, IFN–γ, IL-1β, IL-1α, IL-2, IL-4, IL-5, IL-6, IL-7, IL-8, IL-10, IL-12/IL-23 p40, IL-12 p70, IL-13, IL-15, IL-16, IL-17, IFN-γ-inducible protein 10, macrophage colony-stimulating factor (M-CSF), monocyte chemoattractant protein (MCP)–1, MCP-2, MCP-4, macrophage-derived chemokine, macrophage migration inhibitory factor, macrophage inflammatory protein (MIP)–1β, MIP-1α, MIP-5, matrix metalloproteinase (MMP)–1, MMP-3, MMP-9, Nectin-4, Osteoactivin, osteonectin, P-cadherin, serum amyloid protein A, stem cell factor (SCF), thymus and activation regulated chemokine, TNF–α, TNF-β, TNF–r1, TNF–r2 (R2), TNF–related apoptosis-inducing ligand (TRAIL), vascular cell adhesion molecule 1, and vascular endothelial growth factor A |
Serum | Case-control | TRAIL, ICAM-1, MCP-2, and TNF-r1 were elevated in epilepsy within 24 hours after seizure | 137 patients with epilepsy vs 29 controls. | 2+ |
| Česká et al. [42] | IL-6, IL-8, IL-10, IL-18, CXCL10/IP-10, CCL2/MCP-1, BLC, TNF-α, C-X3-X and fractalquine (CXC3CL1) | CSF and Serum | Case-control | Significant elevation of CCL2/MCP-1 in CSF and serum. Higher levels of fractalkine/CXC3CL1 in serum of pharmacoresistant patients than in controls |
26 patients with epilepsy (22 DRE, 4 non-DRE) vs 9 healthy controls. | 2- |
Table 2.
Summary of biomarkers that have shown some evidence in the systematic review without contradictory results.
Table 2.
Summary of biomarkers that have shown some evidence in the systematic review without contradictory results.
| Biomarkers | Number of studies with positive results | Sample | Quality of the evidence | Conclusions of the studies |
|---|---|---|---|---|
| HMGB1 | 6 Case-control studies | Serum & CSF | 2+ | Possible biomarker of DRE. Possible biomarker of seizure frequency. Temporal Relationship with Generalized Tonic-Clonic Seizures. |
| TNF-a | 2 Case-control studies & 1 Prospective population-based study | Serum & CSF | 2+ | Possible biomarker of DRE. |
| TLR-4 | 3 Case-control studies | Serum | 2 | Possible biomarker of DRE. Possible biomarker of seizure frequency. |
| rTNFr2 | 1 Case-control study | Serum | 2- | Possible biomarker of seizure frequency. |
| CCL2/MCP-1 | 1 Case-control study | Serum & CSF | 2- | Possible biomarker of DRE |
| IL-33 | 1 Case-control study | Serum | 2- | Possible biomarker of epilepsy. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



