
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
: This Systematic Review focused on the use of Bifidobacterium infantis and Bifidobacterium breve in neurodegenerative diseases. The study of metabolomics has demonstrated the relationship between dysbiosis and neuronal death. The modulation of the gut microbiota in the intervention of these pathologies has become a promising area of study, showing beneficial results such as the improvement of synaptic function, the reduction of neuroinflammation, as well as the improvement of associated neuropsychiatric symptoms. Several studies have addressed the administration of Bifidobacterium infantis and Bifidobacterium breve, alone, in conjunction or in combination with other strains, in the intervention of neurodegenerative diseases. This Systematic Review includes the results of 17 research articles (n=5 in humans and n=12 in animal models) and the main result was that the use of these Bifidobacterium strains, alone, in conjunction or in combination with other strains, produced improvements in neurodegenerative disease-related symptomatology and degenerative cellular processes. However, some studies shown neutral results in certain variables. These data suggest that the use of these probiotics has a neuroprotective effect that may delay the progression of the disease. This research provides relevant and updated information on the use of these Bifidobacterium strains in neurodegenerative pathologies such as Alzheimer's disease and Parkinson's disease.
Keywords:
neurodegenerative disease
; probiotics
; Alzheimer´s disease
; Parkinson´s disease
; neurodegenerative process
; Bifidobacterium infantis
; Bifidobacterium longum subsp. infantis
; Bifidobacterium breve
1. Introduction
Currently, it is known that there are numerous neurodegenerative diseases (more than 100 have been described to date), as well as medical conditions related to neurodegenerative processes [1,2]. These conditions include oxidative stress and mild cognitive impairment (MCI), which appear to be associated with the aging process. In addition, neurodegenerative diseases also include highly prevalent pathologies such as Alzheimer’s disease (AD) and Parkinson’s disease (PD) [3,4].
Oxidative stress is related to cognitive impairment and is caused by the toxic effects on cells produced by the oxidation of proteins, lipids, carbohydrates and nucleotides [5,6,7]. These processes, which take milliseconds, promote the accumulation of intracellular aggregates, and produce excitotoxicity, mitochondrial dysfunction and apoptosis [8,9]. Furthermore, MCI refers to a heterogeneous entity of impaired cognitive functions in a single or multiple domains that can be diagnosed as the onset of dementia or the early symptoms of AD [10,11,12,13].
AD is a neurodegenerative disease characterized by the deposition of extracellular senile plaques, and intracellular neurofibrillary tangles (NFTs), synapse reduction and gliosis develop [14,15,16,17,18]. Synapse loss is one of the earliest pathological events in AD, strongly correlating with cognitive decline and impaired memory function [19]. Gliosis, the reactive change in glial cells in response to damage, plays a critical role in the disease’s progression. Astrocytes and microglia, the primary glial cells involved, become activated and contribute to synaptic dysfunction and loss through the release of pro-inflammatory cytokines and other neurotoxic factors [20]. This combined reduction in synaptic connections and increase in gliosis disrupts neural communication and accelerates neurodegenerative processes, establishing these phenomena as central mechanisms in AD’s etiology [21]. Its progressive deterioration is complex in the later stages of the disease and therefore, it is essential to define interventions that regulate the progression of AD in early stages [22]. PD is other neurodegenerative disease characterized by premature destruction of dopaminergic neurons in the midbrain, impaired communication between the prefrontal cortex and subcortical areas and damage to the basal ganglia [23,24,25,26]. Basal ganglia dysfunction is prevalent in various neurological disorders, including Parkinson’s disease, Huntington’s disease, and Tourette syndrome, affecting millions worldwide. For instance, Parkinson’s disease alone affects approximately 1% of individuals over the age of 60, highlighting the widespread impact of basal ganglia-related pathologies [27,28].
In recent years, alterations in the gut microbiota (GM) have been increasingly associated with the onset and progression of neurodegenerative diseases such as Alzheimer’s disease and Parkinson’s disease [29,30,31]. Dysbiosis, or the loss of homeostasis in the GM, is related to these neurodegenerative conditions [32,33]. This imbalance in the microbial community can influence the disease process through various mechanisms, including the modulation of endocrine pathways, immune signals, and neurological factors [32,34,35]. Therapeutic proposals aimed at regulating the GM have been explored, particularly through the use of probiotics. These interventions have shown significant benefits by modulating the levels of bacterial families colonizing the human GM [30,36,37]. Probiotic treatments can reduce systemic inflammation, modulate the expression of genes associated with neurodegeneration, and improve both motor and non-motor symptoms of these pathologies [30,36,37]. This therapeutic approach holds promise for improving the symptoms associated with neurodegenerative processes.
Among the most studied probiotics, those composed mainly of Bifidobacterium and Lactobacillaceae were found. Lactobacillaceae is a family of long, curved or straight, gram-positive, anaerobic, lactic acid-forming bacilli. Bifidobacteriaceae is a genus of gram-positive, anaerobic bacilli, and they utilize the fructose-6-phosphate phosphoketolase pathway [38,39]. Of these families, the Bifidobacterium longum and Bifidobacterium bifidum species, and the Lactoplantibacillus plantarum, Lactobacillus paracasei and Lactobacillus casei species, respectively, are the ones that have shown the most scientific evidence in the intervention of neurodegenerative diseases [32,40].
Several studies suggest that the administration of different strains of Bifidobacterium, alone or in combination, significantly improve the neuropsychiatric symptoms present in neurodegenerative processes [41,42,43,44], although it is still complex to identify the exact mechanisms that give probiotics their neuroprotective action. Due to the diversity of comparing the data obtained in studies dealing with the use of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis, and Bifidobacterium breve, it is necessary to review the main research obtained in this line. Bifidobacterium infantis has been widely studied for its beneficial effects on gut health, particularly in preterm infants, showing significant promise in reducing gastrointestinal issues [45]. Similarly, Bifidobacterium longum subsp. infantis has been noted for its potential in alleviating symptoms of irritable bowel syndrome and other gastrointestinal disorders [46]. Bifidobacterium breve has also been explored extensively, demonstrating beneficial outcomes in both children and adults when used as a dietary supplement [47]. These findings provide a comprehensive understanding of the role of these probiotics in promoting gut health and mitigating gastrointestinal disorders.
Therefore, addressing these types of interventions is a challenge and an opportunity for the scientific and health community [29,48,49,50]. The data reported on the use of probiotics, and other techniques aimed at modulating the gut microbiota (e.g., fecal microbiota transplantation), have shown that it is essential to re-examine interventions for neurodegenerative diseases through the regulation of GM [51,52].
The previous Systematic Review conducted by our research group analyzed the impact of Lactiplantibacillus plantarum, alone or in combination, on the symptomatology associated with neurodegenerative processes. The results obtained have shown that the use of this probiotic had a potential benefit in reversing the progression of the pathology. The use of metabolomics allows the identification of altered metabolic markers [44,53,54,55]. Thus, for example, we found that amino acid and phospholipid alterations in the hippocampus are associated with memory deficits in AD patients, among other variables. These data suggest interventions aimed at restoring the levels of these markers with the use of probiotics. It is important to note that Bifidobacterium infantis is also known as Bifidobacterium longum subsp. infantis by scientific community; infantis is often classified as a subspecies of Bifidobacterium longum due to high genetic similarity observed in genome sequencing. Therefore, both terms are used in this study to avoid losing relevant data on this probiotic.
The main objective of this Systematic Review was to collect all significant findings on the impact of the administration of Bifidobacterium infantis (or Bifidobacterium longum subps. Infantis) and Bifidobacterium breve, alone, in conjunction or in combination with other strains, in the treatment of neurodegenerative diseases. The focus will be on metabolic and cognitive modifications in neurodegenerative processes. This review will provide current data on the use of these Bifidobacterium strains for a better understanding of the advances in this line of research.
2. Methods
2.1. Literature Search
The Systematic Review followed the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) statement [56,57] and we followed the methodology of our previous RS in conducting this research [58]. We searched for articles published between 2000 and 2024, in the following databases: ScienceDirect, Scopus, Wiley, PubMed and Web of Science (WoS), and the following search terms were used: [Bifidobacterium infantis OR Bifidobacterium longum subsp. infantis OR Bifidobacterium breve] AND [Neurodegenerative disease OR Parkinson Disease OR Alzheimer Disease]. Two authors independently conducted the literature search in January 2024 (A.I.B.-V and M.R.), the initial review of titles and abstracts, and the evaluation of retrievable articles for comprehensive review.
2.2. Study Selection
In both human and animal, the studies were screened for inclusion by the following eligibility criteria: i) that the use of Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve as a therapeutic target in the process of neurodegeneration was addressed, ii) that the studies included any of the pathologies studied in this Systematic Review, which involve neurodegeneration, AD, PD, and oxidative stress, iii) the full text was accessible online, iv) the article was written in English. Exclusion criteria included: i) studies primarily focused on the food industry or the use of Bifidobacterium strains in food products without a specific focus on neurodegenerative diseases or oxidative stress; ii) studies lacking a direct investigation into the therapeutic effects of the mentioned Bifidobacterium strains on neurodegenerative conditions; and iii) articles not available in full text or not written in English.
A total of 858 articles were retrieved (Figure 1), of which only 19 articles were identified as meeting the eligibility criteria. Once duplicates were removed, the titles and abstracts of all articles were examined for eligibility. Studies that not included intervention with study strains of Bifidobacterium (n=2) were excluded. Following this detailed review, a total of 17 studies were included. After the screening phase, all full texts of the selected studies were retrieved for comprehensive review, based on the established inclusion criteria.
2.3. Data Extraction
The data extracted from the included studies were as described below (Table 1):
- ▪
- Type of study (human or animal model).
- ▪
- Type of probiotic used (Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve, alone, in conjunction or in combination).
- ▪
- Neurodegenerative pathology addressed in the research.
- ▪
- Population (description).
- ▪
- Methodology of the research carried out.
- ▪
- Intervention (dose administered, time).
- ▪
- Results obtained after the intervention with Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve.

Table 1.
Effects of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis and Bifidobacterium breve, alone or in combination on neurodegenerative diseases (in human and animal models).
Table 1.
Effects of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis and Bifidobacterium breve, alone or in combination on neurodegenerative diseases (in human and animal models).
| Probiotics | Population | Methodology | Intervention | Results | References | ||
| Alzheimer’s Disease | |||||||
| HUMAN STUDIES | Mixture probiotic: Bifidobacterium longum subsp. infantis BLI-02, Bifidobacterium breve Bv-889, and Bifidobacterium animalis subsp. lactis CP-9 | 40 patients | Probiotic vs Sham | Probiotic (1 × 1010 CFU/capsule for 12 weeks) | BDNF levels in the treatment group increased significantly from a baseline value of 7115.1 ± 4461.9 pg/mL to an endpoint of 9678.5 ± 6652.9 pg/mL, with ** p = 0.005; The fold change of cortisol decrease was significantly larger in treatment group as compared with the active control group (119.4% vs. 94.3%, *p = 0.039) | (63) | 10.3390/nu16010016 |
| MCI (Mild Cognitive Impairment) | |||||||
| Bifidobacterium breve A1 | 117 patients | Probiotic vs Sham | Bifidobacterium breve A1 (2 capsules daily > 1×1010 CFU) for 12 weeks | Significant difference between B. breve A1 and placebo groups in subscale ‘immediate memory’ of RBANS and MMSE total score (#p < 0.05 between the treatment and placebo groups at baseline; †p < 0.05 between the treatment and placebo groups at 12-weeks examination); No significant intergroup difference was observed in terms of changes in scores from the baseline scores | (59) | 10.3920/BM2018.0170 | |
| Bifidobacterium breve MCC1274 | 115 patients | Probiotic vs Sham | Bifidobacterium breve MCC1274 (A1) (2×1010 CFU) daily for 24 weeks | A significant intergroup difference was observed in the changes from baseline of GM (gray matter atrophy in the whole brain) extent score (p = 0.013) | (60) | 10.3233/JAD-220148 | |
| Lactobacillus plantarum BioF-228, Lactococcus lactis BioF-224, Bifidobacterium lactis CP-9, Lactobacillus rhamnosus Bv-77, Lactobacillus johnsonii MH-68, Lactobacillus paracasei MP137, Lactobacillus salivarius AP-32, Lactobacillus acidophilus TYCA06, Lactococcus lactis LY-66, Bifidobacterium lactis HNO19, Lactobacillus rhamnosus HNO01, Lactobacillus paracasei GL-156, Bifidobacterium animalis BB-115, Lactobacillus casei CS-773, Lactobacillus reuteri TSR332, Lactobacillus fermentum TSF331, Bifidobacterium infantis BLI-02, and Lactobacillus plantarum CN2018 | 42 patients | Probiotic vs Sham | Mixture probiotic (>2*1010 CFU/g) probiotics daily for 12 weeks | Cognitive function and sleep quality were improved. Mini-Mental State Examination-MMSE (24.75 ± 2.47); Montreal Cognitive Assessment Scale-MoCA (22,05 ± 2,14 vs 20,10 ± 1,45); Pittsburgh Sleep Quality Index-PSQI (5,35 ± 2,78 vs 8,40 ± 1,76, p < 0,001) | (61) | 10.1016/j.gerinurse.2023.03.006 | |
| Oxidative stress | |||||||
| VSL#3® - Bifidobacterium infantis DSM 24737, Bifidobacterium longum DSM 24736, Bifidobacterium breve DSM 24732, Lactobacillus acidophilus DSM 24735, Lactobacillus delbrückii ssp. bulgaricus DSM 24734, Lactobacillus paracasei DSM 24733, Lactobacillus plantarum DSM 24730, and Streptococcus thermophilus DSM 24731 | 62 patients | Probiotic vs Sham | VSL#3® supplementation (2 capsules daily) for 56 days | Arm B did not significantly affect cholesterol or glucose, however it reduced ESR (p =0.05) and was associated with significant increases in serum folate (p=0.007) and serum vitamin B12 (p=0.001), and a decrease in plasma homocysteine (p < 0.001). Both diet alone and diet plus VSL#3® were associated with an increase in glutathione-S-transferase activity | (62) | 10.1016/j.clnu.2014.09.023 | |
| Alzheimer’s Disease | |||||||
| ANIMAL STUDIES | Bifidobacterium breve A1 | Male 10-week-old ddY mice | Probiotic vs Sham | Bifidobacterium breve A1 (1×109 CFU in 0.2 ml, starting 2 days before Aβ injection) daily for 10 days | B. breve A1 prevents Aβ-induced cognitive dysfunction; suppresses Aβ-induced changes in gene expression in the hippocampus; B. breve A1 and acetate partially ameliorate behavioral deficits (p < 0.05, ††p < 0.01 vs. control (sham). *p < 0.05, **p < 0.01 vs. Aβ (+)) | (66) | 10.1038/s41598-017-13368-2 |
| Mixture probiotic: Lactobacillus reuteri, Lactobacillus rhamnosus, and Bifidobacterium infantis | 50 male Wistar rats | Probiotic vs Sham | Mixture probiotic 2 g (1010 CFU) daily for 10 weeks | Aβ-treated group had longer time latency in comparison with the control and sham groups in MWM training phase (p < 0.001). Administration of probiotics promoted spatial memory and learning in comparison with Aβ-treated group (p < 0.01). Administration of probiotics mixture demonstrated a significant decrement in MDA level in comparison with Aβ-treated group (p < 0.001) | (67) | 10.29252/ibj.24.4.220 | |
| Bifidobacterium breve-five strains (B. breve NMG, B. breve MY, B. breve CCFM1025, B. breve XY, and B. breve WX) | 63 male C57BL/6J mice (8 weeks old) | Individual 5 strains of Bifidobacterium breve | Bifidobacterium breve-5 strains (B. breve NMG, B. breve MY, B. breve CCFM1025, B. breve XY, and B. breve WX) (1 × 109 CFU/mL) daily for 6 weeks | B. breve NMG and CCFM1025 administration led to significant improvements in alternation behavior and increases in total arm entries. However, the administration of the other three B. breve strains failed to improve working memory; CCFM1025, XY, and WX to Aβ1-42-treated mice significantly reduced the hippocampal accumulation of Aβ1-42 (Control vs. model: # p < 0.05 by unpaired student’s t-test; * p < 0.05, ** p < 0.01 by one-way ANOVA for all groups) | (68) | 10.3390/nu13051602 | |
| Bifidobacterium breve MCC1274 | 40 C57BL/6J mice (2-month-old) | Probiotic vs Sham | Bifidobacterium breve MCC1274 (1 × 109 CFU/mL) five times/week for four months | Significant decrease in soluble Aβ1-42 levels in the hippocampal extracts of probiotic mice vs to those of mice that received Sham; p-Akt and p-GSK-3β protein levels were significantly increased in the hippocampus of the probiotic group vs Sham group (* p < 0.05); B. breve MCC1274 significantly increased the protein levels of SYT and syntaxin, and showed a tendency to increase the protein levels of SYP and PSD-95 in hippocampal extracts (** p < 0.01) | (69) | 10.3390/nu14122543 | |
| Bifidobacterium breve A1 | 52 App knock-in (KI) mice (AppNL-G-F)-3-month-old | Probiotic vs Sham | Bifidobacterium breve A1 (1×109 CFU in 0.2 ml, starting 2 days before Aβ injection) daily for 10 days | Memory impairment: the probiotic group had a significantly increased exploration time for the novel object compared with the familiar object; the discrimination index (DI) was higher in the probiotics group vs Sham group; B. breve MCC1274 supplementation suppresses Aβ fibril formation; significantly upregulated ADAM10 and PS1 in the hippocampus, whereas AβPP and BACE1 levels did not change (p < 0.05) | (70) | 10.3233/JAD-215025 | |
| Vitalon Probiotics (VP) powder: Bacillus natto, Bacillus coagulans, Lactobacillus casei, Lactobacillus acidophilus, Bifidobacterium longum, Bifidobacterium breve | 9–15 mice/group- APP transgenic mouse line J20 & Wild-type (WT) littermate mice (control) | Prebiotic vs Synbiotic | 3.6 g/kg/day prebiotics (WT/P and APP/P)-2.5% inulin; or 4.1 g/kg/day synbiotics (WT/S and APP/S)-Vitalon, intragastrically for 2 months | The level of Aβ1-42 was significantly decreased in APP/S mice compared with APP/C mice; synbiotic treatment significantly reduced TNF-α levels (*p < .05; ***p < .001; ****p < .0001) | (71) | 10.1002/iub.2589 | |
| Bifidobacterium breve CCFM1025 | 40 Male adult C57BL/6J mice (8 weeks old) | Probiotic + EE vs Protiobic | Bifidobacterium breve CCFM1025 (1 × 109 CFU/mL) + daily for 6 weeks vs Bifidobacterium breve CCFM1025 (1 × 109 CFU/mL) daily for 6 weeks | EE + B. breve CCFM1025 showed improved working memory in the Y-maze; accumulation of hippocampal Aβ1-42 was significantly decreased in the EE-treated groups, with the EE + B. breve CCFM1025 (Control vs. Model: #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001) | (72) | 10.3389/fimmu.2022.1013664 | |
| Bifidobacterium breve CCFM1025 | 24 Male adult C57BL/6J mice (8 weeks old) | Probiotic vs Sham | Bifidobacterium breve CCFM1025 (5 × 109 CFU/mL) vs Veh-sterile 10% skimmed milk | The levels of L-tyrosine and tryptophan in the model group were restored by Bifidobacterium breve CCFM1025 treatment (p < 0.05); compared with the control mice, 36 metabolites were statistically altered in the hippocampal tissues of the model mice (p < 0.05) vs CCFM1025; CCFM1025 restored the level of the serum metabolite phenylalanine and L-glutamine levels in the hippocampus (* p < 0.05; ** p < 0.01) | (73) | 10.3390/nu14040735 | |
| Bifidobacterium breve HNXY26M4 | Male mice (16-week-old) | Probiotic vs Sham | Bifidobacterium breve HNXY26M4 (1 × 109 CFU/mL) daily for 12 weeks | Treatment with B. breve HNXY26M4 led to dramatically lower levels of Aβ1−42 than those in APP/PS1 mice; B. breve HNXY26M4 Supplementation Ameliorates Neuroinflammation, Oxidative Damage, and Synaptic Impairment in the Brains of APP/PS1 Mice; the levels of acetate and butyrate were increased in samples from B. breve HNXY26M4-treated mice, and a more substantial increase in acetate relative to butyrate (*p < 0.05, **p < 0.01, and ***p < 0.001) | (74) | 10.1021/acs.jafc.3c00652 | |
| Bifidobacterium breve HNXY26M4 | 40 Male adult C57BL/6J mice (8 weeks old) | Probiotic+EE vs Protiobic | Bifidobacterium breve HNXY26M4 (1 × 109 CFU/mL) daily + EE vs Bifidobacterium breve HNXY26M4 (1 × 109 CFU/mL) daily for 6 weeks | Mice that received only EE (ADEE) or EE combined with B. breve treatment (AD+BBEE) had significantly lower concentrations of Aβ1-42 in the hippocampus than ADSE mice (*p < 0.05, **p < 0.01, ***p < 0.001) | (75) | 10.26599/FSHW.2022.9250084 | |
| Parkinson’s Disease | |||||||
| Bifidobacterium breve A1-MCC1274 | 156 Male C57BL/6 mice (7–8 weeks old) | Probiotic vs Sham | Bifidobacterium breve MCC1274 (A1) (1 × 109 CFU/mL) daily for 4 days | B. breve A1 prevented the reduction of spine density in PD mice and maintained it at the same level as that in control mice (* p < 0.01 vs. Control + Saline and † p < 0.05: between MPTP + Saline and MPTP + B. breve A1); B. breve A1 did not show any significant effects on hippocampal cAMP levels in PD and control mice (* p < 0.01 vs. Control + Saline) | (64) | 10.3390/biomedicines9020167 | |
| Bifidobacterium breve CCFM1067 | 40 Male C57BL/6 mice (6 weeks old) | Probiotic vs Sham | Bifidobacterium breve CCFM1067 (109 CFU/200 μL saline) daily on days 8-41 | B. breve CCFM1067 improves MPTP-induced motor impairments: Both L-DOPA and B. breve CCFM1067 therapies substantially reduced MPTP-induced motor impairments in the PT (F (2,21) = 56.94, p < 0.0001), NBT (F (2,21) = 33.72, p < 0.0001), and RTR (F (2,18) = 21.99, p < 0.0001); B. breve CCFM1067 reduced the increase in striatal TNF-α (F (2,9) = 91.49, p < 0.0001), IL-1β (F (2,9) = 29.53, p < 0.0001), and IL-6 (F (2,9) = 24.61, p = 0.0002) | (65) | 10.3390/nu14214678 | |
3. Results
3.1. Human Studies
Five studies were conducted in humans. One addressed Alzheimer’s disease and the impact of the use of Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve in the intervention of neurodegenerative processes in AD patients (31) (n=1); and four of them explored MCI or Oxidative stress in neurodegenerative processes [59,60,61,62] (n=4).
3.1.1. Alzheimer’s Disease
Hsu et al. have conducted a recent study in 2024 in which they addressed the role of a probiotic composed of Bifidobacterium longum subsp. infantis BLI-02, Bifidobacterium breve Bv-889, Bifidobacterium animalis subsp. lactis CP-9, Bifidobacterium bifidum VDD088 and Lactobacillus plantarum PL-02 in AD treatment (1×1010 CFU/capsule). 40 patients participated in this study, and after 12 weeks of intervention with this probiotic mixture, a 36% increase in serum BDNF (brain-derived neurotrophic factor) (*p = 0.005) and an increase in SOD (antioxidant superoxide dismutase) (*p = 0.012) was observed in the experimental group vs. the control group. In addition, a reduction in IL-1β (*p = 0.041) and a significantly reduction in cortisol levels (119.4% vs. 94.3%, *p = 0.039) were as well observed (63). [*Main method of analysis (Student’s t-test)].
3.1.2. Mild Cognitive Impairment
The 2019 study by Kobayashi et al. included 117 older adults (50-80 years) and analyzed the impact of Bifidobacterium breve A1 on cognitive dysfunction associated with neurodegeneration. Patients received 2 capsules daily (>1×1010 CFU) for 12 weeks. The results showed significant improvements in MMSE-Mini-Mental State Examination, and RBANS-Assessment of Neuropsychological Status scores (#p < 0.05 between treatment and placebo groups at baseline; †p < 0.05 between treatment and placebo groups at the 12-week examination). However, no significant differences were observed between groups in terms of changes in scores from baseline scores [59]. [*Main method of analysis (Student’s t-test)].
Other study conducted by Asaoka et al. in 2022 addressed the role of Bifidobacterium breve MCC1274 (A1) in the treatment of older adults with chronological age-associated MCI. The study involved 115 patients (65-88 years) who received (2×1010 CFU) daily for 24 weeks. MMSE scores improved (< 25) compared to the placebo group (“orientation in time” and “writing” subscales). In the probiotic-treated group, there was a tendency to suppress the progression of progressive brain atrophy in some subjects (VOI Z-score ≥1.0- Z-score of gray matter atrophy in the volume of interest-VOI). Specifically, a significant intergroup difference was observed in the changes from baseline of GM (gray matter atrophy in the whole brain) extent score (p = 0.013) [60]. [*Main method of analysis (Student’s t-test)].
The study conducted by Fei et al. in 2023 addressed the use of a probiotic mixture including Lactobacillus plantarum BioF-228, Lactococcus lactis BioF-224, Bifidobacterium lactis CP-9, Lactobacillus rhamnosus Bv-77, Lactobacillus johnsonii MH-68, Lactobacillus paracasei MP137, Lactobacillus salivarius AP-32, Lactobacillus acidophilus TYCA06, Lactococcus lactis LY-66, Bifidobacterium lactis HNO19, Lactobacillus rhamnosus HNO01, Lactobacillus paracasei GL-156, Bifidobacterium animalis BB-115, Lactobacillus casei CS-773, Lactobacillus reuteri TSR332, Lactobacillus fermentum TSF331, Bifidobacterium infantis BLI-02, and Lactobacillus plantarum CN2018, in 42 volunteer participants (over 60 years of age). Patients took (>2×1010 CFU/g) probiotic daily for 12 weeks. The results showed that cognitive function and sleep quality were improved: MMSE (24.75 ± 2.47); Montreal Cognitive Assessment Scale-MoCA (22.05 ± 2.14 vs. 20.10 ± 1.45); Pittsburgh Sleep Quality Index-PSQI (5.35 ± 2.78 vs. 8.40 ± 1.76, p < 0.001) (61). [*Student’s t-test and analysis of variance (ANOVA) were used for comparative analyses].
3.1.3. Oxidative Stress
Valentini et al. in 2015 conducted a study analyzing the impact of VSL#3® (Bifidobacterium infantis DSM 24737, Bifidobacterium longum DSM 24736, Bifidobacterium breve DSM 24732, Lactobacillus acidophilus DSM 24735, Lactobacillus delbrückii ssp. bulgaricus DSM 24734, Lactobacillus paracasei DSM 24733, Lactobacillus plantarum DSM 24730, and Streptococcus thermophilus DSM 24731) in 62 patients (65 -85 years). VSL#3® were administered daily (2 capsules) for 56 days. VSL#3® did not significantly affect cholesterol or glucose levels; however, it did reduce the Erythrocyte Sedimentation Rate (ESR) (p = 0.05), indicating a decrease in systemic inflammation. This arm was also associated with significant increases in serum folate (p = 0.007) and serum vitamin B12 (p = 0.001), as well as a decrease in plasma homocysteine levels (p < 0.001). These biochemical changes suggest a potential reduction in oxidative stress, as elevated homocysteine is a known risk factor for oxidative damage and inflammation. Both diet alone and diet plus VSL#3® were associated with an increase in glutathione-S-transferase activity, an enzyme involved in detoxification processes and protection against oxidative stress (62). These findings collectively indicate that the interventions may help mitigate oxidative stress, likely through enhanced antioxidant defenses and improved nutrient status. [*Chi-squared test and Mann–Whitney U-test were used for biochemical parameters; ANOVA and ANCOVA were used for differences in inflammatory parameters before and after dietary interventions].
In conclusion, the evidence in human studies suggests that the use of Bifidobacterium infantis or Bifidobacterium longum subsp. infantis, alone or in combination with other strains, improved BDNF levels, reduced cortisol and ESR-Erythrocyte Sedimentation Rate levels, and increased folate and vitamin B12 levels. Moreover, the administration of Bifidobacterium breve MCC1274 (A1) showed an improvement on cognitive function (MMSE and RBANS test), and significant modifications in GM (gray matter atrophy in the whole brain) extent score [60,61,63]. In the study conducted by Valentini et al. in 2015, an increase in glutathione S-transferase activity was found to be associated with reactive oxygen species-ROS neutralization [62]. It is important to mention that the study by Kobayashi et al. (2019) did not show significant differences between groups in terms of changes in scores with respect to baseline scores [59].
3.2. Animal Studies
A total of 12 studies were conducted using different animal models. Of these, 10 focused on AD (29,38,39,41-44,48-50) (n=10); and 2 focused on PD [64,65] (n=2). All of them addressed the intervention with Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve for the improvement of degeneration associated with AD or PD.
3.2.1. Alzheimer’s Disease
Kobayashi et al. conducted a study in 2017 about the impact of Bifidobacterium breve A1 in an AD model-male (10-week-old) ddY mice. The probiotic was administered daily for 10 days (1×109 CFU in 0.2 ml, starting 2 days before Aβ injection). The results have shown that Bifidobacterium breve A1 prevents Aβ-induced cognitive dysfunction and suppresses Aβ-induced changes in gene expression in the hippocampus. Moreover, Bifidobacterium breve A1 and acetate partially ameliorate behavioral deficits (p < 0.05, ††p < 0.01 vs. Sham. *p < 0.05, **p < 0.01 vs. Aβ (+)) [66]. [*One-way analysis of variance followed by Student’s t or Mann–Whitney U post hoc tests].
The research conducted by Mehrabadi & Sadr, in 2020, used an AD model-Wistar rat with Aβ1-40 intra-hippocampal injection (n=50), and administered a 2 ml probiotic mixture composed of Lactobacillus reuteri, Lactobacillus rhamnosus, and Bifidobacterium infantis (1010 CFU daily for 10 weeks). Results shown that the Aβtreated group had a longer time latency compared to the control and Sham groups in the MWM-Morris water maze training phase (p < 0.001). Probiotic administration promoted spatial memory and learning compared to the Aβ-treated group (p < 0.01). Administration of the probiotic mixture showed a significant decrease in the level of malondialdehyde-MDA compared to the Aβ-treated group (p < 0.001) and a reduction in the levels of inflammation biomarkers IL-1β (p < 0.01) and TNF-α (p < 0.01) [67]. [*ANOVA, followed by post hoc Tukey’s test were used for comparative analyses].
In 2021, Zhu et al. conducted a study analyzing the impact of five strains of Bifidobacterium breve (B. breve NMG, B. breve MY, B. breve CCFM1025, B. breve XY, and B. breve WX) individually (1×109 CFU/mL) in 63 male AD model-C57BL/6J mice (8 weeks old), daily for 6 weeks. In this research, Bifidobacterium breve NMG and CCFM1025 administration led to significant improvements in alternation behavior and increases in total arm entries in the Morris water maze. However, the administration of the other three Bifidobacterium breve strains failed to improve working memory; CCFM1025, XY, and WX to Aβ1-42-treated mice significantly reduced the hippocampal accumulation of Aβ1-42 (Control vs. model: # p < 0.05 by unpaired Student’s t-test; * p < 0.05, ** p < 0.01 by one-way ANOVA for all groups) [68]. [*One-way analysis of variance (ANOVA) or the Kruskal–Wallis test with post hoc comparison].
Abdelhamid et al. (2022) conducted a study with AD model-C57BL/6J mice (2-month-old) (n=40). The animals were treated with Bifidobacterium breve MCC1274 (1×109 CFU/mL) five times/week for four months vs. Sham. Significant decrease in soluble Aβ1-42 levels in the hippocampal extracts of probiotic mice vs. Sham; p-Akt and p-GSK-3β protein levels were significantly increased in the hippocampus of the probiotic group vs. Sham group (*p < 0.05); B. breve MCC1274 significantly increased the protein levels of SYT and syntaxin, and showed a tendency to increase the protein levels of SYP and PSD-95 in hippocampal extracts (**p < 0.01) [69]. [*Two-tailed unpaired Student’s t-test (for normally distributed variables)].
Other study conducted for Abdelhamid et al. in 2022, with similar methodology, addressed the impact of Bifidobacterium breve MCC1274 (A1) in an AD model-App knock-in (KI) mice (AppNL-G-F)-3-month-old (n=52). Bifidobacterium breve A1 (1×109 CFU in 0.2 ml, starting 2 days before Aβ injection), were administered daily for 10 days. The probiotic group had a significantly increased exploration time for the novel object compared with the familiar object, improving the memory impairment. In addition, the discrimination index (DI) was higher in the probiotics group vs. Sham group; B. breve MCC1274 supplementation suppresses Aβ fibril formation; significantly upregulated ADAM10 and PS1 in the hippocampus, whereas AβPP and BACE1 levels did not change (p < 0.05) [70]. [*Main method of analysis (Student’s t-test)].
Deng et al., in 2022, conducted a study analyzing the effect of 2.5% inulin (3.6 g/kg/day) compared with a synbiotic (4.1 g/kg/day), daily for 2 months on APP transgenic mouse line J20 & Wild-type (WT) littermate mice (control). This synbiotic contained the inulin prebiotic and Vitalon Probiotics (VP) powder: Bacillus natto, Bacillus coagulans, Lactobacillus casei, Lactobacillus acidophilus, Bifidobacterium longum and Bifidobacterium breve. The results showed that the level of Aβ1-42 was significantly decreased in the experimental group compared with control mice; the synbiotic treatment significantly reduced TNF-α levels (*p < .05; ***p < .001; ****p < .0001) [71]. [*Differences between multiple means were assessed by one-way or two-way ANOVA].
Zhu et al. conducted a study in 2022 analyzing the impact of Bifidobacterium breve CCFM1025 in 40 male adult AD model-C57BL/6J mice (8 weeks old). The animals received the probiotic (1×109 CFU/mL) alone or in combination with enriched environment-EE, daily for 6 weeks. EE + Bifidobacterium breve CCFM1025 showed improved working memory in the Y-maze and accumulation of hippocampal Aβ1-42 was significantly decreased in the EE-treated groups, with the EE + B. breve CCFM1025 (Control vs. Model: #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001) [72]. [*A one-way analysis of variance (ANOVA) with Holm–Sidak test or Student’s t-test was used for the parametric analysis of differences between groups; Kruskal–Wallis test followed by Dunn’s test or Welch’s t-test was performed for nonparametric data variance analysis].
Other study conducted by the research group of Zhu et al. (2022) evaluated the impact of Bifidobacterium breve CCFM1025 in 24 adult male AD model-C57BL/6J mice (8 weeks old). The animals received the probiotic (1×109 CFU/mL) alone or Sham-sterilized 10% skim milk, daily for 6 weeks. The results have shown that the levels of L-tyrosine and tryptophan in the model group were restored by Bifidobacterium breve CCFM1025 treatment (p < 0.05); compared with the control mice, 36 metabolites were statistically altered in the hippocampal tissues of the model mice (p < 0.05) vs. CCFM1025; and restored the level of the serum metabolite phenylalanine and L-glutamine levels in the hippocampus (*p < 0.05; **p < 0.01) [73]. [*Main method of analysis (Principal Component Analysis-PCA) of all samples and the Pearson correlation coefficient between QC samples].
The study conducted by Zhu et al. [74] in an AD model-male mice (16-week-old) addressed the effects of Bifidobacterium breve HNXY26M4 vs. Sham. The animals received Bifidobacterium breve HNXY26M4 (1×109 CFU/mL) daily for 12 weeks and the results indicated that treatment with Bifidobacterium breve HNXY26M4 led to dramatically lower levels of Aβ1−42 than those in APP/PS1 mice; ameliorates neuroinflammation, oxidative damage, and synaptic impairment in the brains of APP/PS1 mice; the levels of acetate and butyrate were increased in samples from Bifidobacterium. breve HNXY26M4-treated mice, and a more substantial increase in acetate relative to butyrate (*p < 0.05, **p < 0.01, and ***p < 0.001) [74]. [*One-way ANOVA].
A recent study conducted by Zhu et al. in 2024, addressed the use of Bifidobacterium breve HNXY26M4 (1×109 CFU/mL) daily in male adult AD model-C57BL/6J mice (8 weeks old) (n=40), alone or administered with EE, for 6 weeks. The results indicated that this probiotic reduces cognitive impairment and modulates glutamine metabolism, improving brain function. Specifically, mice that received only Environmental Enrichment (EE, ADEE) or EE combined with Bifidobacterium breve treatment (AD+BBEE) had significantly lower concentrations of Aβ1-42 in the hippocampus than AD-SE mice (*p < 0.05, **p < 0.01, ***p < 0.001) [75]. This indicates that both environmental enrichment and the combination of EE with Bifidobacterium breve treatment are effective in reducing Aβ1-42 levels, a marker associated with Alzheimer’s disease pathology. It is important to note that while EE alone was effective, the addition of B. breve did not diminish the beneficial effects, suggesting that the probiotic may complement the environmental intervention rather than acting independently. The combined treatment may offer a synergistic effect, enhancing overall outcomes. [*Student’s t-test and analysis of variance (ANOVA) were used for comparative analyses].
In conclusion, the results obtained in AD model-animal studies have shown that Bifidobacterium breve (A1)- CCFM1025 could prevent cognitive disfunction, improve memory impairment, and ameliorate behavioral deficits [60,64,66]. Moreover, A1 suppressed Aβ fibril formation and p-Akt and p-GSK-3β protein levels were significantly increased in the hippocampus [63,64,66]. In addition, Bifidobacterium breve CCFM1025 (A1) restored the levels of L-tyrosine and tryptophan, the level of the serum metabolite phenylalanine, and L-glutamine levels in the hippocampus [73].
The studies that addressed the treatment with Bifidobacterium breve HNXY26M4 concluded that this probiotic produced a significative decrease of Aβ1-42 level in hippocampus, ameliorated neuroinflammation, oxidative damage and synaptic impairment in brain [74,75]. In the same line, the studies that administered a mixture probiotic or individual strains of Bifidobacterium breve showed that the use of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis or Bifidobacterium breve, improve the mnesic function, decrease the MDA levels [61]; reduced the Aβ1-42 level in hippocampus and reduce TNF-α levels [68,71].
3.2.2. Parkinson’s Disease
Ishii et al. conducted research in 2021 and Bifidobacterium breve A1 was evaluated in a PD model-male C57BL/6 mice (7–8 weeks old) (n=156). The probiotic was administered vs. Sham, in the same method: Bifidobacterium breve MCC1274 (A1) (1×109 CFU/mL), daily for 4 days. In this study, the results indicated that Bifidobacterium breve A1 prevented the reduction of spine density in PD mice and maintained it at the same level as that in control mice (*p < 0.01 vs. Control + Saline (†p < 0.05): between MPTP + Saline and MPTP + Bifidobacterium breve A1; Bifidobacterium breve A1 did not show any significant effects on hippocampal cAMP-adenylyl cyclase (AC)–cyclic AMP levels in PD and control mice (*p < 0.01 vs. Control + Saline) [64]. [*Tukey’s post-hoc test or Kruskal–Wallis analysis and Mann–Whitney U tests based on the analyses of homoscedasticity using Levene’s test].
In 2022, Li et al. conducted a study which addressed the impact of Bifidobacterium breve CCFM1067 (109 CFU/200 μL saline) daily on days 8-41. The authors used a PD model-male C57BL/6 mice (6 weeks old) (n=40). This study concluded that Bifidobacterium breve CCFM1067 improves MPTP-induced motor impairments: Both L-DOPA and Bifidobacterium breve CCFM1067 therapies substantially reduced MPTP-induced motor impairments in the PT-pole test (F (2,21) = 56.94, p < 0.0001), NBT-narrow-beam test (F (2,21) = 33.72, p < 0.0001), and RTR-rotarod test (F (2,18) = 21.99, p < 0.0001); Bifidobacterium. breve CCFM1067 reduced the increase in striatal TNF-α (F (2,9) = 91.49, p < 0.0001), IL-1β (F (2,9) = 29.53, p < 0.0001), and IL-6 (F (2,9) = 24.61, p = 0.0002) (65). [*D’Agostino–Pearson test, Student’s unpaired t-test or one-way analysis of variance (ANOVA) with Dunnett’s test was used to compare two or multiple datasets. The Mann-Whitney test or Kruskal-Wallis with Dunn’s test was conducted to compare two or multiple datasets, respectively].
Evidence suggests that the use of Bifidobacterium breve MCC1274 (A1) exerts a neuroprotector function prevented the reduction of spine density in PD model. It is interesting the result about the absence of modification in the hippocampal cAMP-adenylyl cyclase (AC)–cyclic AMP levels [64]. The use of Bifidobacterium breve CCFM1067 showed improvement in motor impairments and reduced the increase in striatal TNF-α (65).
4. Discussion
The main objective of this Systematic Review was to compile all significant findings from human and animal model studies investigating the impact of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis or Bifidobacterium breve, alone, in conjunction or in combination with other strains, on the improvement of symptomatology and cellular damage associated with neurodegenerative disease. The results obtained indicated that these probiotics improve both motor symptoms and neuropsychiatric impairments and reduce neuronal damage alone and in combination with other bacterial strains. In comparison, Lactiplantibacillus (Lactobacillus) plantarum has also shown significant potential in alleviating neurological symptoms. Studies have demonstrated that Lactiplantibacillus plantarum can improve neuropsychiatric disorders, sleep disorders, and autonomic dysfunction in patients with neurological conditions [58]. Furthermore, Lactiplantibacillus plantarum has been found to enhance cognitive function and mitigate neuronal damage when used in combination with other treatments, such as memantine [29]. Both Bifidobacterium strains and Lactiplantibacillus plantarum exhibit beneficial effects on the gut-brain axis, supporting their use as psychobiotics for improving both motor and neuropsychiatric symptoms [38].
Analyzing the impact of Bifidobacterium infantis, Bifidobacterium longum subsp. infantis or Bifidobacterium breve, alone, in conjunction or in combination with other strains on neurodegenerative diseases, was observed that both human and animal studies have mainly focused on AD, PD, oxidative stress and MCI, as shown in Table 1.
Regarding AD, it was observed that the use of Bifidobacterium strains studies presented a neuroprotective action, reducing cortisol levels and increasing BDNF levels, and reduced neuroinflammation. In addition, these probiotics showed improvements in memory function and learning, and up-regulation of acetate and butyrate levels, as well as the level of the serum metabolite phenylalanine and L-glutamine levels in the hippocampus [63,66,67,68,69,70,71,72,73,74,75].
Recent studies suggested that β-amyloid (Aβ) peptide is the main cause of cognitive impairment and synaptic dysfunction in AD [70]. Also, these studies suggested that Aβ decreases BDNF by reducing the CREB proteins. Therefore, BDNF increase and regulation of this signaling pathway (CREB-BDNF) is presented as a potential intervention in this pathology, regulating Aβ toxicity to delay disease progression [76,77,78].
Addressing the use of probiotics in AD, recent studies have shown a beneficial effect of Lactiplantibacillus plantarum, alone or in combination. Specifically, the previous Systematic Review conducted by our research group showed that the use of this probiotic has a neuroprotective action, regulating ATPase processes and improving glucose metabolism in the brain. In addition, the use of Lactiplantibacillus plantarum exerted a protective role on synaptic function and neuronal survival. Finally, an improvement in memory function was observed [58].
Other studies have analyzed the impact of different strains of Bifidobacterium on AD. Interventions with Bifidobacterium lactis and Bifidobacterium longum have shown positive effects in both human and animal models [79,80]. For example, a study conducted by Kim et al. in 2021 showed that a mixture of Bifidobacterium bifidum BGN4 and Bifidobacterium longum BORI administered for 12 weeks produced a significant increase in serum BDNF levels (p < 0.05) [81].
Regarding PD, both studies analyzed the impact of different strains of Bifidobacterium breve. Specifically, administration of Bifidobacterium breve MCC1274 (A1) showed protective effects by preventing the reduction of spine density [64]. Moreover, the use of Bifidobacterium breve CCFM1067 showed improvements in motor symptomatology, and a reduction of striatal TNF-α [65]. It is widely accepted that damage to the striatum is responsible for the motor symptoms observed in PD [82,83,84]. Motor symptomatology can emerge with as little as a 30% reduction in dopamine (DA) levels, although patients are typically diagnosed with an 80% loss of DA in the putamen due to compensatory mechanisms [85,86,87].
Addressing the use of probiotics in PD, previous studies have shown that administration of Lactobacillus and Bifidobacterium improved motor symptoms (Movement Disorder Society’s Unified Parkinson’s Disease Rating Scale scores). Among others, the researches addressing the use of Lactobacillus acidophilus, Lactobacillus fermentum and Lactobacillus reuteri showed neuroprotective effects and a reduction of oxidative stress. In addition, this genus is useful for the improvement of neuropsychiatric symptomatology associated with PD for its ability to modulate mood, or the quality of sleep, among others [88].
Analyzing the impact of different strains of Bifidobacterium, previous studies have shown their beneficial effects on this pathology. For example, Rezaeisasl, Salami & Sepehri in 2019 used a probiotic mixture composed of Bifidobacterium bifidum, Bifidobacterium longum, and Lactobacillus acidophilus. After a 6-week treatment, they found a protective effect on the serum profile and improvements in learning. However, this study is interesting because probiotic administration did not result in any improvement in memory function [89].
Regarding mild cognitive impairment, it was observed that the use of Bifidobacterium breve MCC1274 (A1) improved memory deficits. However, in the study conducted by Kobayashi et al. [59], no significant differences were found in the pre-post comparison of MMSE and RBANS scores [59]. Asaoka et al. [60] found that this probiotic improved gut microbiota (GM) levels compared to the control group, suggesting better gut health and reduced dysbiosis (60). In addition, in the 2023 study conducted by Fei et al. [61], a probiotic mixture containing various strains of Lactoplantibacillus and Bifidobacterium was used. The study found significant improvements in MMSE and MoCA (Montreal Cognitive Assessment) scores, and the sleep quality measured by PSQI (Pittsburgh Sleep Quality Index) [61].
Previous studies have analyzed the impact of probiotics on MCI. For instance, Lactiplantibacillus plantarum had shown to improve memory function, reduce neuroinflammation, and improve synaptic dysfunction [90,91]. Other probiotic, Lactobacillus plantarum DR7, had demonstrated improvements in memory and neuroprotective effects. Additionally, this probiotic reduced anxious-depressive symptoms [92].
In a 2020 study conducted by Xiao et al., Bifidobacterium breve was administered to older adults with MCI for 16 weeks. The results showed that those who took Bifidobacterium breve had significantly better scores on MMSE and RBANS compared to the control group [93]. In their study, Yang et al. in 2020 (94) used a probiotic mixture consisting of Bifidobacterium lactis, Lactobacillus casei, Bifidobacterium bifidum, and Lactobacillus acidophilus on animals for 12 weeks. The results showed a significant improvement in memory deficits and a decrease in previous cognitive dysfunction. Furthermore, an improvement of synaptic function and neuronal functioning were observed [94].
Regarding oxidative stress, the study conducted by Valentini et al. (2015), found an increase in glutathione-S-transferase activity [62]. There are various antioxidant systems, including antioxidants that prevent the formation of ROS [95]. These systems include SOD-superoxide dismutase, transferrin, glutathione peroxidase, and glutathione S-transferase. The latter plays a role in reducing ROS, regulating membrane permeability, and participating in the synthesis of proteins, DNA, and RNA, among others [96,97].
The scientific community has recently studied the use of probiotics on oxidative stress. For example, Lin et al. in 2022 [98] conducted an animal study on the effects of a probiotic mixture containing Bifidobacterium animalis subsp. infantis BLI-02, Bifidobacterium breve Bv889, Bifidobacterium bifidum VDD088, Bifidobacterium animalis subsp. lactis CP-9, and Lactobacillus plantarum PL-02. The study found that this probiotic mixture had a neuroprotective effect and increased butyrate levels, which are directly associated with inflammatory and oxidative processes [98].
In 2020, Ton et al. [99] conducted a study analyzing the use of a prebiotic kefir mixture composed of Acetobacter aceti, Acetobacter spp., Lactobacillus delbrueckii delbrueckii, Lactobacillus fermentum, Lactobacillus fructivorans, Enterococcus faecium, Leuconostoc spp., Lactobacillus kefiranofaciens, Candida famata, and Candida krusei in patients with oxidative stress and dementia. The study findings indicate an enhancement in cognitive function through mechanisms related to cell damage, organ inflammation, and oxidative stress [99].
The results reported in this research have shown that probiotic intervention may improve symptoms associated with cognitive impairment in neurodegenerative diseases, such as memory, spatial orientation, and learning, anxious level, among others. Additionally, Bifidobacterium probiotics have demonstrated neuroprotective properties and the ability to modulate several metabolic processes associated with neurodegeneration. While acknowledging certain limitations, these data serve as a valued step for addressing degenerative processes and neurodegenerative diseases.
5. Limitations and Future Research
One limitation that should be noted is the small number of registered clinical trials. The identification of biological markers is an active research field, although the use of metabolomics is a recent development. Nonetheless, the data collected in this Systematic Review are relevant. All of this data allows us to identify starting points for future research with a solid and rigorous empirical basis.
It should be noted that the databases consulted contain a limited number of full free articles. It is important to consider the possibility of validating results with studies that are not accessible to the scientific community.
It is important to consider another limitation, which is that the evaluated interventions are heterogeneous in terms of the administered dose and intervention duration. Additionally, some probiotics are used alone while others are used in combination with different strains, species, and even families, making it difficult to generalize the beneficial results obtained from their administration. Further study of the mechanisms of action of each administered probiotic is necessary.
This Systematic Review is the first to report and highlight the effects of Bifidobacterium infantis and Bifidobacterium breve, alone or in combination, on neurodegenerative diseases. The collected empirical evidence supports the development of interventions that incorporate the use of analyzed Bifidobacterium strains to improve motor, cognitive, and behavioural symptoms related to these pathologies, as well as to delay related-neurodegenerative processes.
6. Practical Applications
The review highlights the significant role of gut microbiota in the development and progression of neurodegenerative diseases. Changes in the diversity and composition of the gut microbiota, often leading to dysbiosis, are associated with brain pathologies such as Alzheimer’s and Parkinson’s diseases. Dysbiosis can disrupt cell homeostasis and contribute to neuroinflammation, a key factor in neurodegeneration. Probiotics, particularly certain strains of Bifidobacterium, have shown promise in reversing dysbiosis and improving cognitive function in both animal models and human studies. These live microorganisms, known as psychobiotics, may offer psychological benefits by modulating the gut-brain axis. The therapeutic potential of probiotics lies in their ability to enhance gut health, reduce inflammation, and possibly slow the progression of neurodegenerative diseases. However, further research is needed to fully understand the mechanisms involved and to develop targeted interventions that can be effectively integrated into clinical practice. The findings underscore the importance of an interdisciplinary approach to transform these scientific discoveries into practical applications that can benefit patients with neurodegenerative conditions.
7. Conclusions
The results from both human and animal studies indicate that Bifidobacterium infantis, Bifidobacterium longum subsp. infantis, and Bifidobacterium breve probiotic strains can improve neuropsychiatric symptoms, enhance cognitive functions, and reduce neuroinflammation and oxidative stress markers. Specifically, the administration of these Bifidobacterium strains was associated with improvements in memory, spatial orientation, and learning abilities, alongside a decrease in anxiety levels and enhanced motor function. These findings support the hypothesis that targeted probiotic interventions can positively modulate gut-brain interactions, potentially offering a neuroprotective effect that delays the progression of neurodegenerative diseases. Therefore, incorporating these probiotics into therapeutic protocols presents a promising avenue for enhancing the quality of life for individuals suffering from conditions such as Alzheimer’s and Parkinson’s diseases.
Author Contributions
Conceptualization, all authors; methodology, M.R., A.I.B.-V and V.J.C.-S. a; writing—original draft preparation, all authors; writing—review and editing, all authors; visualization, all authors; supervision, S.U., A.I.B.-V and V.J.C.-S; project administration, A.I.B-V, V.J.C.-S and S.U. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not Applicable.
Informed Consent Statement
Not Applicable.
Data Availability Statement
Not applicable.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dugger, B.N.; Dickson, D.W. Pathology of neurodegenerative diseases. Cold Spring Harb. Perspect. Biol. 2017, 9(7), a028035. [Google Scholar] [CrossRef] [PubMed]
- Chi, H.; Chang, H.Y.; Sang, T.K. Neuronal cell death mechanisms in major neurodegenerative diseases. Int. J. Mol. Sci. 2018, 19(10), 3082. [Google Scholar] [CrossRef] [PubMed]
- Litke, R.; Garcharna, L.C.; Jiwani, S.; Neugroschl, J. Modifiable risk factors in Alzheimer disease and related dementias: a review. Clin. Ther. 2021, 43(6), 953–965. [Google Scholar] [CrossRef] [PubMed]
- Agnello, L.; Ciaccio, M. Neurodegenerative diseases: from molecular basis to therapy. Int. J. Mol. Sci. 2022, 23, 21–12854. [Google Scholar] [CrossRef] [PubMed]
- McCord, J.M. The evolution of free radicals and oxidative stress. Am. J. Med. 2000, 108(8), 652–659. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative stress: a key modulator in neurodegenerative diseases. Molecules 2019, 24(8), 1583. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.T. Oxidative stress and mitochondrial dysfunction-linked neurodegenerative disorders. Neurol. Res. 2017, 39(1), 73–82. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, Y.; Yarjanli, Z.; Pakniya, F.; Bidram, E.; Łos, M.J.; Eshraghi, M.; Klionsky, D.J.; Ghavami, S.; Zarrabi, A. Targeting autophagy, oxidative stress, and ER stress for neurodegenerative disease treatment. J. Control Release 2022, 345, 147–175. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An overview of oxidative stress, neuroinflammation, and neurodegenerative diseases. Int. J Mol. Sci. 2022, 23(11), 5938. [Google Scholar] [CrossRef]
- Kasper, S.; Bancher, C.; Eckert, A.; Förstl, H.; Frölich, L.; Hort, J.; Korczyn, A.D.; Kressig, R.W.; Levin, O.; Palomo, M.S.M. Management of mild cognitive impairment (MCI): the need for national and international guidelines. World J. Biol. Psychiatry 2020, 21(8), 579–594. [Google Scholar] [CrossRef]
- Baiano, C.; Barone, P.; Trojano, L.; Santangelo, G. Prevalence and clinical aspects of mild cognitive impairment in Parkinson’s disease: a meta-analysis. Mov. Disord. 2020, 35(1), 45–54. [Google Scholar] [CrossRef]
- Jongsiriyanyong, S.; Limpawattana, P. Mild cognitive impairment in clinical practice: a review article. Am. J. Alzheimers Dis. Other Demen. 2018, 33(8), 500–507. [Google Scholar] [CrossRef] [PubMed]
- Blackman, J.; Swirski, M.; Clynes, J.; Harding, S.; Leng, Y.; Coulthard, E. Pharmacological and non-pharmacological interventions to enhance sleep in mild cognitive impairment and mild Alzheimer’s disease: a systematic review. J Sleep Res. 2021, 30(4), e13229. [Google Scholar] [CrossRef]
- Zhang, C. Etiology of Alzheimer’s Disease. Discov. Med. 2023, 35(178), 757. [Google Scholar] [CrossRef] [PubMed]
- Breijyeh, Z.; Karaman, R. Comprehensive review on Alzheimer’s disease: causes and treatment. Molecules 2020, 25(24), 5789. [Google Scholar] [CrossRef]
- Ando, K.; Brion, J.P.; Stygelbout, V.; Suain, V.; Authelet, M.; Dedecker, R.; Chanut, A.; Lacor, P.; Lavaur, J.; Sazdovitch, V.; et al. Clathrin adaptor CALM/PICALM is associated with neurofibrillary tangles and is cleaved in Alzheimer’s brains. Acta Neuropathol. 2013, 125(6), 861–878. [Google Scholar] [CrossRef] [PubMed]
- Allen, N.J. Astrocyte regulation of synaptic behavior. Annu. Rev. Cell Dev. Biol. 2014, 30, 439–463. [Google Scholar] [CrossRef]
- De Strooper, B.; Karran, E. The cellular phase of Alzheimer’s disease. Cell 2016, 164(4), 603–615. [Google Scholar] [CrossRef] [PubMed]
- Hulshof, L.A.; van Nuijs, D.; Hol, E.M.; Middeldorp, J. The role of astrocytes in synapse loss in Alzheimer’s disease: a systematic review. Front. Cell. Neurosci. 2022, 16, 899251. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, J.; Savage, J.C.; Tremblay, M.È. Synaptic loss in Alzheimer’s disease: mechanistic insights provided by two-photon in vivo imaging of transgenic mouse models. Front. Cell Neurosci. 2020, 14, 592607. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.; Grant, S.G. Synapse pathology in Alzheimer’s disease. Semin. Cell Dev. Biol. 2023, 139, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Scheltens, P.; Blennow, K.; Breteler, M.M.; de Strooper, B.; Frisoni, G.B.; Salloway, S.; Van der Flier, W.M. Alzheimer’s disease. The Lancet 2016, 388(10043), 505–517. [Google Scholar] [CrossRef] [PubMed]
- Meoni, S.; Cury, R.G.; Moro, E. New players in basal ganglia dysfunction in Parkinson’s disease. Prog. Brain Res. 2020, 252, 307–327. [Google Scholar] [PubMed]
- Adler, A.F.; Cardoso, T.; Nolbrant, S.; Mattsson, B.; Hoban, D.B.; Jarl, U.; Wahlestedt, J.N.; Grealish, S.; Björklund, A.; Parmar, M. hESC-derived dopaminergic transplants integrate into basal ganglia circuitry in a preclinical model of Parkinson’s disease. Cell Rep. 2019, 28(13), 3462–3473.e5. [Google Scholar] [CrossRef] [PubMed]
- Costa, H.N.; Esteves, A.R.; Empadinhas, N.; Cardoso, S.M. Parkinson’s disease: a multisystem disorder. Neurosci. Bull. 2023, 39(1), 113–124. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R. Parkinson’s disease: from pathogenesis to pharmacogenomics. Int. J. Mol. Sci. 2017, 18(3), 551. [Google Scholar] [CrossRef] [PubMed]
- Leisman, G.; Braun-Benjamin, O.; Melillo, R. Cognitive-motor interactions of the basal ganglia in development. Front. Syst. Neurosci. 2014, 8, 16. [Google Scholar] [CrossRef]
- Schmahmann, J.D.; Pandya, D.N. Disconnection syndromes of basal ganglia, thalamus, and cerebrocerebellar systems. Cortex 2008, 44, 1037–1066. [Google Scholar] [CrossRef] [PubMed]
- Quigley, E.M.M. Microbiota-brain-gut axis and neurodegenerative diseases. Curr. Neurol. Neurosci. Rep. 2017, 17(12), 94. [Google Scholar] [CrossRef] [PubMed]
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 2016, 167(6), 1469–1480.e12. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Li, G.; Huang, P.; Liu, Z.; Zhao, B. The gut microbiota and Alzheimer’s disease. J. Alzheimers Dis. 2017, 58(1), 1–15. [Google Scholar] [CrossRef] [PubMed]
- Uceda, S.; Echeverry-Alzate, V.; Reiriz-Rojas, M.; Martínez-Miguel, E.; Pérez-Curiel, A.; Gómez-Senent, S.; Beltrán-Velasco, A.I. Gut microbial metabolome and dysbiosis in neurodegenerative diseases: psychobiotics and fecal microbiota transplantation as a therapeutic approach—A comprehensive narrative review. Int. J. Mol. Sci. 2023, 24(17), 13294. [Google Scholar] [CrossRef] [PubMed]
- Pluta, R.; Ułamek-Kozioł, M.; Januszewski, S.; Czuczwar, S.J. Gut microbiota and pro/prebiotics in Alzheimer’s disease. Aging 2020, 12(6), 5539–5550. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Huang, S.Y.; Chen, S.D.; Zhang, Y.R.; Huang, Y.Y.; Yu, J.T. Investigating casual associations among gut microbiota, metabolites, and neurodegenerative diseases: a mendelian randomization study. J. Alzheimers Dis. 2022, 87(1), 211–222. [Google Scholar] [CrossRef]
- Chen, Z.; Maqbool, J.; Sajid, F.; Hussain, G.; Sun, T. Human gut microbiota and its association with pathogenesis and treatments of neurodegenerative diseases. Microb. Pathog. 2021, 150, 104675. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Luo, Y.; Ray Chaudhuri, K.; Reynolds, R.; Tan, E.K.; Pettersson, S. The role of gut dysbiosis in Parkinson’s disease: mechanistic insights and therapeutic options. Brain 2021, 144(9), 2571–2593. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Gao, J.; Zhu, M.; Liu, K.; Zhang, H.L. Gut microbiota and dysbiosis in Alzheimer’s disease: implications for pathogenesis and treatment. Mol. Neurobiol. 2020, 57(12), 5026–5043. [Google Scholar] [CrossRef] [PubMed]
- Socała, K.; Doboszewska, U.; Szopa, A.; Serefko, A.; Włodarczyk, M.; Zielińska, A.; Poleszak, E.; Fichna, J.; Wlaź, P. The role of microbiota-gut-brain axis in neuropsychiatric and neurological disorders. Pharmacol. Res. 2021, 172, 105840. [Google Scholar] [CrossRef] [PubMed]
- Aizawa, E.; Tsuji, H.; Asahara, T.; Takahashi, T.; Teraishi, T.; Yoshida, S.; Koga, N.; Hattori, K.; Ota, M.; Kunugi, H. Bifidobacterium and Lactobacillus counts in the gut microbiota of patients with bipolar disorder and healthy controls. Front. Psychiatry. 2019, 9, 730. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, C.; do Carmo, M.; Melo, B.; Alves, M.; dos Santos, C.; Monteiro, S.; Bomfim, M.R.Q.; Fernandes, E.S.; Monteiro-Neto, V. In vitro antimicrobial activity and probiotic potential of Bifidobacterium and Lactobacillus against species of Clostridium. Nutrients 2019, 11(2), 448. [Google Scholar] [PubMed]
- Mohammadi, G.; Dargahi, L.; Peymani, A.; Mirzanejad, Y.; Alizadeh, S.A.; Naserpour, T.; Nassiri-Asl, M. The effects of probiotic formulation pretreatment (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) on a lipopolysaccharide rat model. J. Am. Coll. Nutr. 2019, 38(3), 209–217. [Google Scholar] [PubMed]
- Mohammadi, G.; Dargahi, L.; Naserpour, T.; Mirzanejad, Y.; Alizadeh, S.A.; Peymani, A.; Nassiri-Asl, M. Probiotic mixture of Lactobacillus helveticus R0052 and Bifidobacterium longum R0175 attenuates hippocampal apoptosis induced by lipopolysaccharide in rats. Int. Microbiol. 2019, 22(3), 317–323. [Google Scholar]
- Zhang, C.; Zhang, Y.; Li, H.; Liu, X. The potential of proteins, hydrolysates and peptides as growth factors for Lactobacillus and Bifidobacterium: current research and future perspectives. Food Funct. 2020, 11(3), 1946–1957. [Google Scholar] [PubMed]
- Ni, Y.; Zhang, Y.; Zheng, L.; Rong, N.; Yang, Y.; Gong, P.; Yang, Y.; Siwu, X.; Zhang, C.; Zhu, L.; Fu, Z. Bifidobacterium and Lactobacillus improve inflammatory bowel disease in zebrafish of different ages by regulating the intestinal mucosal barrier and microbiota. Life Sci. 2023, 324, 121699. [Google Scholar] [CrossRef] [PubMed]
- Batta, V.K.; Rao, S.C.; Patole, S.K. Bifidobacterium infantis as a probiotic in preterm infants: a systematic review and meta-analysis. Pediatr. Res. 2023, 94(6), 1887–1905. [Google Scholar] [PubMed]
- Dylag, K.; Hubalewska-Mazgaj, M.; Surmiak, M.; Szmyd, J.; Brzozowski, T. Probiotics in the mechanism of protection against gut inflammation and therapy of gastrointestinal disorders. Curr. Pharm. Des. 2014, 20, 1149–1155. [Google Scholar] [CrossRef] [PubMed]
- Bozzi Cionci, N.; Baffoni, L.; Gaggìa, F.; Di Gioia, D. Therapeutic microbiology: the role of Bifidobacterium breve as food supplement for the prevention/treatment of paediatric diseases. Nutrients 2018, 10(11), 1723. [Google Scholar] [PubMed]
- Zhao, Z.; Ning, J.; Bao, X.Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced Parkinson’s disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9(1), 226. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xu, J.; Ling, Y.; Wang, F.; Gong, T.; Yang, C.; Ye, S.; Ye, K.; Wei, D.; Song, Z.; Chen, D.; Liu, J. Fecal microbiota transplantation alleviated Alzheimer’s disease-like pathogenesis in APP/PS1 transgenic mice. Transl. Psychiatry 2019, 9(1), 189. [Google Scholar] [CrossRef] [PubMed]
- Vendrik, K.E.W.; Ooijevaar, R.E.; de Jong, P.R.C.; Laman, J.D.; van Oosten, B.W.; van Hilten, J.J.; Ducarmon, Q.R.; Keller, J.J.; Kuijper, E.J.; Contarino, M.F. Fecal microbiota transplantation in neurological disorders. Front. Cell. Infect. Microbiol. 2020, 10, 98. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; Dhalaria, R.; Guleria, S.; Cimler, R.; Sharma, R.; Siddiqui, S.A.; Valko, M.; Nepovimova, E.; Dhanjal, D.S.; Singh, R.; et al. Anti-oxidant potential of plants and probiotic spp. in alleviating oxidative stress induced by H2O2. Biomed. Pharmacother. 2023, 165, 115022. [Google Scholar] [CrossRef] [PubMed]
- Morella, I.; Negro, M.; Dossena, M.; Brambilla, R.; D’Antona, G. Gut-muscle-brain axis: molecular mechanisms in neurodegenerative disorders and potential therapeutic efficacy of probiotic supplementation coupled with exercise. Neuropharmacology 2023, 240, 109718. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.F.; Cheng, Y.F.; You, S.T.; Kuo, W.C.; Huang, C.W.; Chiou, J.J.; Hsu, C.C.; Hsieh-Li, H.M.; Wang, S.; Tsai, Y.C. Lactobacillus plantarum PS128 alleviates neurodegenerative progression in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced mouse models of Parkinson’s disease. Brain Behav. Immun. 2020, 90, 26–46. [Google Scholar] [PubMed]
- Mallikarjuna, N.; Praveen, K.; Yellamma, K. Role of Lactobacillus plantarum MTCC1325 in membrane-bound transport ATPases system in Alzheimer’s disease-induced rat brain. BioImpacts 2016, 6(4), 203–209. [Google Scholar] [PubMed]
- Bonfili, L.; Cecarini, V.; Gogoi, O.; Berardi, S.; Scarpona, S.; Angeletti, M.; Rossi, G.; Eleuteri, A.M. Gut microbiota manipulation through probiotics oral administration restores glucose homeostasis in a mouse model of Alzheimer’s disease. Neurobiol. Aging 2020, 87, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Shamseer, L.; Clarke, M.; Ghersi, D.; Liberati, A.; Petticrew, M.; Shekelle, P.; Stewart, L.A.; PRISMA-P Group. Preferred reporting items for systematic review and meta-analysis protocols (PRISMA-P) 2015 statement. Syst. Rev. 2015, 4(1), 1. [Google Scholar] [CrossRef] [PubMed]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Moher, D. Updating guidance for reporting systematic reviews: development of the PRISMA 2020 statement. J. Clin. Epidemiol. 2021, 134, 103–112. [Google Scholar] [CrossRef]
- Beltrán-Velasco, A.I.; Reiriz, M.; Uceda, S.; Echeverry-Alzate, V. Lactiplantibacillus (Lactobacillus) plantarum as a complementary treatment to improve symptomatology in neurodegenerative disease: a systematic review of open access literature. Int. J. Mol. Sci. 2024, 25(5), 3010. [Google Scholar]
- Kobayashi, Y.; Kuhara, T.; Oki, M.; Xiao, J.Z. Effects of Bifidobacterium breve A1 on the cognitive function of older adults with memory complaints: a randomised, double-blind, placebo-controlled trial. Benef. Microbes 2019, 10(5), 511–520. [Google Scholar] [PubMed]
- Asaoka, D.; Xiao, J.; Takeda, T.; Yanagisawa, N.; Yamazaki, T.; Matsubara, Y.; Sugiyama, H.; Endo, N.; Higa, M.; Kasanuki, K.; et al. Effect of probiotic Bifidobacterium breve in improving cognitive function and preventing brain atrophy in older patients with suspected mild cognitive impairment: Results of a 24-week randomized, double-blind, placebo-controlled trial. J. Alzheimers Dis. 2022, 88(1), 75–95. [Google Scholar] [PubMed]
- Fei, Y.; Wang, R.; Lu, J.; Peng, S.; Yang, S.; Wang, Y.; Zheng, K.; Li, R.; Lin, L.; Li, M. Probiotic intervention benefits multiple neural behaviors in older adults with mild cognitive impairment. Geriatr. Nurs. 2023, 51, 167–175. [Google Scholar] [CrossRef]
- Valentini, L.; Pinto, A.; Bourdel-Marchasson, I.; Ostan, R.; Brigidi, P.; Turroni, S.; Hrelia, S.; Hrelia, P.; Bereswill, S.; Fischer, et al. Impact of personalized diet and probiotic supplementation on inflammation, nutritional parameters and intestinal microbiota – The “RISTOMED project”: Randomized controlled trial in healthy older people. Clin. Nutr. 2015, 34(4), 593–602. [Google Scholar] [CrossRef] [PubMed]
- Hsu, Y.C.; Huang, Y.Y.; Tsai, S.Y.; Kuo, Y.W.; Lin, J.H.; Ho, H.H.; Chen, J.F.; Hsia, K.C.; Sun, Y. Efficacy of probiotic supplements on brain-derived neurotrophic factor, inflammatory biomarkers, oxidative stress and cognitive function in patients with Alzheimer’s dementia: a 12-week randomized, double-blind active-controlled study. Nutrients 2023, 16(1), 16. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Furuoka, H.; Kaya, M.; Kuhara, T. Oral administration of probiotic Bifidobacterium breve improves facilitation of hippocampal memory extinction via restoration of aberrant higher induction of neuropsin in an MPTP-induced mouse model of Parkinson’s disease. Biomedicines 2021, 9(2), 167. [Google Scholar] [PubMed]
- Li, T.; Chu, C.; Yu, L.; Zhai, Q.; Wang, S.; Zhao, J.; Zhang, H.; Chen, W.; Tian, F. Neuroprotective effects of Bifidobacterium breve CCFM1067 in MPTP-induced mouse models of Parkinson’s disease. Nutrients 2022, 14(21), 4678. [Google Scholar] [PubMed]
- Kobayashi, Y.; Sugahara, H.; Shimada, K.; Mitsuyama, E.; Kuhara, T.; Yasuoka, A.; Kondo, T.; Abe, K.; Xiao, J.Z. Therapeutic potential of Bifidobacterium breve strain A1 for preventing cognitive impairment in Alzheimer’s disease. Sci. Rep. 2017, 7(1), 13510. [Google Scholar] [PubMed]
- Mehrabadi, S.; Sadr, S.S. Assessment of probiotics mixture on memory function, inflammation markers, and oxidative stress in an Alzheimer’s disease model of rats. Iran. Biomed. J. 2020, 24(4), 220–228. [Google Scholar] [CrossRef]
- Zhu, G.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Administration of Bifidobacterium breve improves the brain function of Aβ1-42-treated mice via the modulation of the gut microbiome. Nutrients 2021, 13(5), 1602. [Google Scholar] [PubMed]
- Abdelhamid, M.; Zhou, C.; Jung, C.G.; Michikawa, M. Probiotic Bifidobacterium breve MCC1274 mitigates Alzheimer’s disease-related pathologies in wild-type mice. Nutrients 2022, 14(12), 2543. [Google Scholar] [PubMed]
- Abdelhamid, M.; Zhou, C.; Ohno, K.; Kuhara, T.; Taslima, F.; Abdullah, M.; Jung, C.G.; Michikawa, M. Probiotic Bifidobacterium breve prevents memory impairment through the reduction of both amyloid-β production and microglia activation in APP knock-in mouse. J. Alzheimers Dis. 2022, 85(4), 1555–1571. [Google Scholar] [PubMed]
- Deng, S.; Chen, C.; Lin, H.; Cheng, I.H. The beneficial effect of synbiotics consumption on Alzheimer’s disease mouse model via reducing local and systemic inflammation. IUBMB Life 2022, 74(8), 748–53. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Guo, M.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Bifidobacterium breve intervention combined with environmental enrichment alleviates cognitive impairment by regulating the gut microbiota and microbial metabolites in Alzheimer’s disease mice. Front. Immunol. 2022, 20, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Guo, M.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Integrative metabolomic characterization reveals the mediating effect of Bifidobacterium breve on amino acid metabolism in a mouse model of Alzheimer’s disease. Nutrients 2022, 14(4), 735. [Google Scholar] [PubMed]
- Zhu, G.; Zhao, J.; Wang, G.; Chen, W. Bifidobacterium breve HNXY26M4 attenuates cognitive deficits and neuroinflammation by regulating the gut–brain axis in APP/PS1 mice. J. Agric. Food Chem. 2023, 71(11), 4646–55. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Guo, M.; Zhao, J.; Zhang, H.; Wang, G.; Chen, W. Environmental enrichment in combination with Bifidobacterium breve HNXY26M4 intervention amplifies neuroprotective benefits in a mouse model of Alzheimer’s disease by modulating glutamine metabolism of the gut microbiome. Food Sci. Hum. Wellness 2024, 13(2), 982–92. [Google Scholar]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.; Zhu, X.; Si, J. Angelica polysaccharide ameliorates memory impairment in Alzheimer’s disease rat through activating BDNF/TrkB/CREB pathway. Exp. Biol. Med. 2020, 245(1), 1–10. [Google Scholar] [CrossRef] [PubMed]
- Pláteník, J.; Fišar, Z.; Buchal, R.; Jirák, R.; Kitzlerová, E.; Zvěřová, M.; Raboch, J. GSK3β, CREB, and BDNF in peripheral blood of patients with Alzheimer’s disease and depression. Prog. Neuropsychopharmacol Biol. Psychiatry 2014, 50, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Nimgampalle, M. Anti-Alzheimer properties of probiotic, Lactobacillus plantarum MTCC 1325 in Alzheimer’s disease induced albino rats. J. Clin. Diagn. Res. 2017, 11(8), KC01–KC05. [Google Scholar] [PubMed]
- Chen, Z.; Zhong, C. Oxidative stress in Alzheimer’s disease. Neurosci. Bull. 2014, 30(2), 271–81. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Cha, L.; Sim, M.; Jung, S.; Chun, W.Y.; Baik, H.W.; Shin, D.M. Probiotic supplementation improves cognitive function and mood with changes in gut microbiota in community-dwelling older adults: a randomized, double-blind, placebo-controlled, multicenter trial. J. Gerontol. A Biol. Sci. Med. Sci. 2021, 76(1), 32–40. [Google Scholar] [CrossRef] [PubMed]
- Bernheimer, H.; Birkmayer, W.; Hornykiewicz, O.; Jellinger, K.; Seitelberger, F. Brain dopamine and the syndromes of Parkinson and Huntington clinical, morphological and neurochemical correlations. J. Neurol. Sci. 1973, 20(4), 415–55. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yang, P.; Knight, W.; Guo, Y.; Perlmutter, J.S.; Benzinger, T.L.S.; Morris, J.C.; Xu, J. The interactions of dopamine and oxidative damage in the striatum of patients with neurodegenerative diseases. J. Neurochem. 2020, 152(2), 235–251. [Google Scholar] [CrossRef]
- Cheng, Q.; Wang, J.; Li, M.; Fang, J.; Ding, H.; Meng, J.; Zhang, J.; Fang, X.; Liu, H.; Ma, C.; et al. CircSV2b participates in oxidative stress regulation through miR-5107-5p-Foxk1-Akt1 axis in Parkinson’s disease. Redox Biol. 2022, 56, 102430. [Google Scholar] [CrossRef] [PubMed]
- Cheng, H.; Burke, R.E. The WldS mutation delays anterograde, but not retrograde, axonal degeneration of the dopaminergic nigro-striatal pathway in vivo. J. Neurochem. 2010, 113(3), 683–91. [Google Scholar] [PubMed]
- Hornykiewicz, O. Dopamine miracle: from brain homogenate to dopamine replacement. Mov. Disord. 2002, 17, 501–508. [Google Scholar] [CrossRef]
- Troshev, D.; Berezhnoy, D.; Kulikova, O.; Abaimov, D.; Muzychuk, O.; Nalobin, D.; Stvolinsky, S.; Fedorova, T. The dynamics of nigrostriatal system damage and neurobehavioral changes in the rotenone rat model of Parkinson’s disease. Brain Res. Bull. 2021, 173, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ghyselinck, J.; Verstrepen, L.; Moens, F.; Van Den Abbeele, P.; Bruggeman, A.; Said, J.; Smith, B.; Barker, L.A.; Jordan, C.; Leta, V.; et al. Influence of probiotic bacteria on gut microbiota composition and gut wall function in an in-vitro model in patients with Parkinson’s disease. Int. J. Pharm. X 2021, 3, 100087. [Google Scholar] [CrossRef] [PubMed]
- Rezaeiasl, Z.; Salami, M.; Sepehri, G. The effects of probiotic Lactobacillus and Bifidobacterium strains on memory and learning behavior, long-term potentiation (LTP), and some biochemical parameters in β-amyloid-induced rat’s model of Alzheimer’s disease. Prev. Nutr. Food Sci. 2019, 24(3), 265–73. [Google Scholar] [PubMed]
- Huang, H.J.; Chen, J.L.; Liao, J.F.; Chen, Y.H.; Chieu, M.W.; Ke, Y.Y.; Hsu, C.C.; Tsai, Y.C.; Hsieh-Li, H.M. Lactobacillus plantarum PS128 prevents cognitive dysfunction in Alzheimer’s disease mice by modulating propionic acid levels, glycogen synthase kinase 3 beta activity, and gliosis. BMC Complement. Med. Ther. 2021, 21, 259. [Google Scholar] [CrossRef]
- Lin, S.W.; Tsai, Y.S.; Chen, Y.L.; Wang, M.F.; Chen, C.C.; Lin, W.H.; Fang, T.J. Lactobacillus plantarum GKM3 promotes longevity, memory retention, and reduces brain oxidation stress in SAMP8 mice. Nutrients 2021, 13(8), 2860. [Google Scholar] [PubMed]
- Zaydi, A.I.; Lew, L.C.; Hor, Y.Y.; Jaafar, M.H.; Chuah, L.O.; Yap, K.P.; Azlan, A.; Azzam, G.; Liong, M.T. Lactobacillus plantarum DR7 improved brain health in aging rats via the serotonin, inflammatory and apoptosis pathways. Benef. Microbes 2020, 11(8), 753–66. [Google Scholar]
- Xiao, J.; Katsumata, N.; Bernier, F.; Ohno, K.; Yamauchi, Y.; Odamaki, T.; Yoshikawa, K.; Ito, K.; Kaneko, T. Probiotic Bifidobacterium breve in improving cognitive functions of older adults with suspected mild cognitive impairment: a randomized, double-blind, placebo-controlled trial. J. Alzheimers Dis. 2020, 77(1), 139–47. [Google Scholar] [PubMed]
- Yang, X.; Yu, D.; Xue, L.; Li, H.; Du, J. Probiotics modulate the microbiota–gut–brain axis and improve memory deficits in aged SAMP8 mice. Acta Pharm. Sin. B 2020, 10(3), 475–87. [Google Scholar] [CrossRef] [PubMed]
- Averill-Bates, D.A. The antioxidant glutathione. Vitam. Horm. 2023, 121, 109–141. [Google Scholar] [PubMed]
- Domingues, I.; Gravato, C. Oxidative stress assessment in zebrafish larvae. Methods Mol. Biol. 2018, 1797, 477–486. [Google Scholar] [PubMed]
- Hellou, J.; Ross, N.W.; Moon, T.W. Glutathione, glutathione s-transferase, and glutathione conjugates, complementary markers of oxidative stress in aquatic biota. Environ. Sci. Pollut. Res. Int. 2012, 19(6), 2007–23. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.Y.; Lin, J.H.; Kuo, Y.W.; Chiang, P.F.R.; Ho, H.H. Probiotics and their metabolites reduce oxidative stress in middle-aged mice. Curr. Microbiol. 2022, 79(4), 104. [Google Scholar] [CrossRef] [PubMed]
- Ton, A.M.M.; Campagnaro, B.P.; Alves, G.A.; Aires, R.; Côco, L.Z.; Arpini, C.M.; Oliveira, T.G.E.; Campos-Toimil, M.; Meyrelles, S.S.; Pereira, T.M.C.; et al. Oxidative stress and dementia in Alzheimer’s patients: effects of synbiotic supplementation. Oxid. Med. Cell. Longev. 2020, 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Flow diagram of the search and selection process. 17 studies met the eligibility criteria and investigated the effects of Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve alone or in combination on neurodegenerative disease (5 studies in human and 12 studies in animal model).
Figure 1.
Flow diagram of the search and selection process. 17 studies met the eligibility criteria and investigated the effects of Bifidobacterium infantis or Bifidobacterium longum subsp. infantis or Bifidobacterium breve alone or in combination on neurodegenerative disease (5 studies in human and 12 studies in animal model).
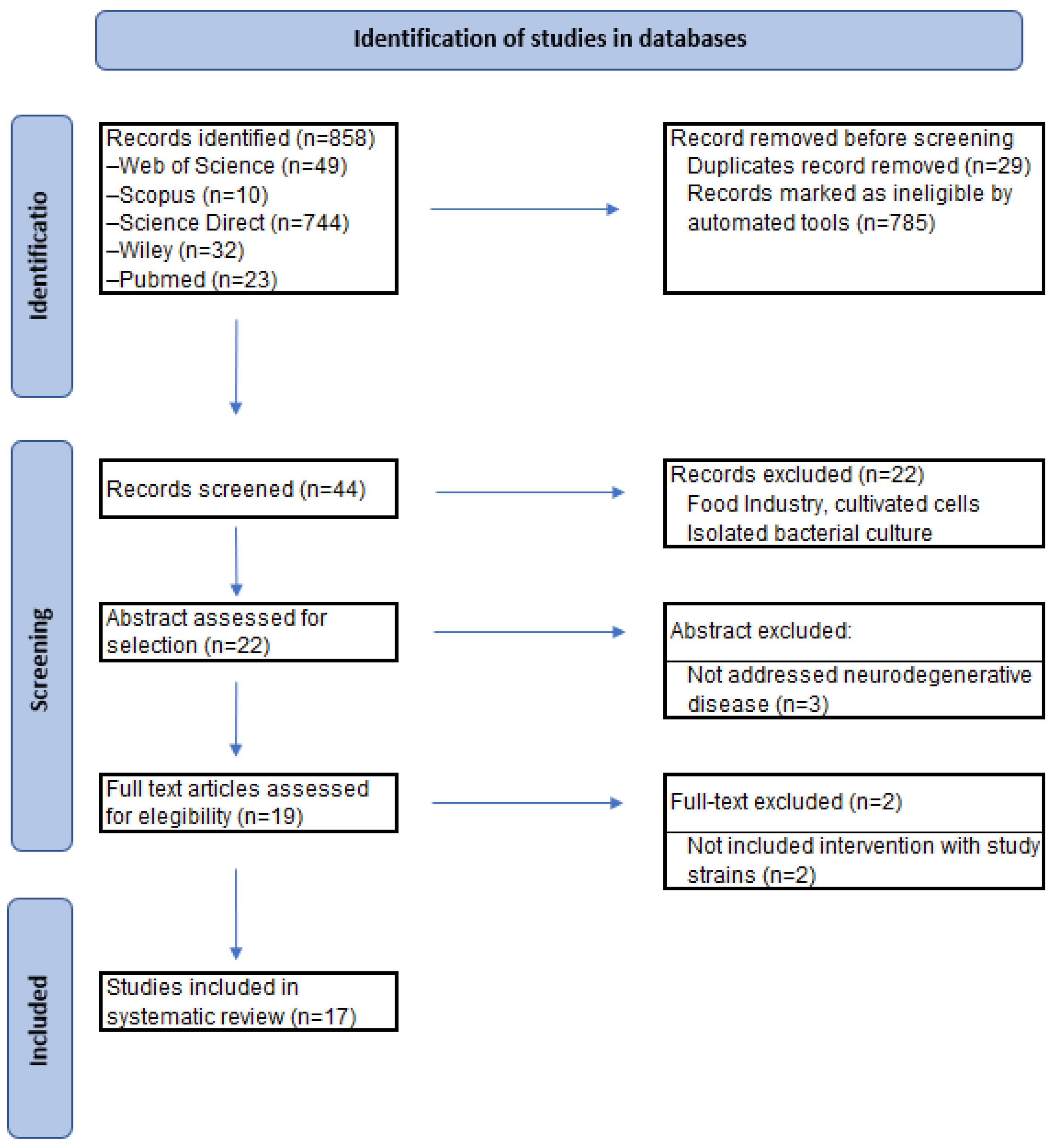
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



