
Submitted:
31 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
Glioblastoma (GBM) is an aggressive brain cancer characterized by significant molecular and cellular heterogeneity, complicating treatment efforts. Current standard therapies, including surgical resection, radiation, and temozolomide (TMZ) chemotherapy, often fail to achieve long-term remission due to tumor recurrence and resistance. A pro-oxidant environment is in-volved in glioma progression, with oxidative stress contributing to the genetic instability that leads to gliomagenesis. However, the generation of reactive oxygen species (ROS) results in the dysregulation of antioxidant defenses in GBM cells, creating a vulnerability that can be thera-peutically exploited. Evaluating pro-oxidant therapies in brain tumors is crucial due to their po-tential to selectively target and eradicate cancer cells by exploiting the elevated oxidative stress levels inherent in these malignant cells, thereby offering a novel and effective strategy for overcoming resistance to conventional therapies. This study investigates the therapeutic potential of doxorubicin (DOX) and photodynamic therapy (PDT) with Me-ALA, focusing on their effects on redox homeostasis. Basal ROS levels and antioxidant gene expression (NFE2L2, CAT, GSR) were quantitatively assessed across GBM cell lines, revealing significant variability probably linked to genetic differences. DOX and PDT treatments, both individually and in combination, were ana-lyzed for their efficacy in inducing oxidative stress and cytotoxicity. An in silico analysis further explored the relationship between gene mutations and oxidative stress in GBM patients, providing insights into the molecular mechanisms underlying treatment responses. Our findings suggest that pro-oxidant therapies, such as DOX and PDT in combination, could selectively target GBM cells, highlighting a promising avenue for improving therapeutic outcomes in GBM.
Keywords:
photodynamic therapy
; chemotherapy
; glioblastoma
; adjuvant treatment
; oxidative therapy
; TP53 mutation
; PTEN mutation
1. Introduction
According to the World Health Organization (WHO), glioblastoma (GBM) is a particularly aggressive type of cancer that accounts for 48.6% of malignant tumors in the brain and 14.5% of all central nervous system (CNS) tumors [1]. The cellular source of this glioma, which is classified as Grade 4, is still a topic of discussion, emphasizing its significant variability and cellular hierarchies that mainly arise from stem or progenitor cells and that confer great intra- and intertumoral heterogeneity [2].
The current treatment for GBM often involves a step-by-step process starting with surgical removal of the tumor, followed by additional treatment with radiation and chemotherapy with temozolomide (TMZ). Nonetheless, the molecular diversity inherent in GBM and the formidable barrier presented by the blood-brain barrier (BBB) pose significant challenges in achieving complete eradication of tumor cells. Adverse prognostic outcomes in GBM patients often arise from the extensive intertumoral and intratumoral heterogeneity, as well as the occurrence of postoperative relapses and development of treatment resistance, phenomena observed in approximately half of the patients undergoing TMZ therapy [3]. Hence, there is a critical imperative to explore alternative therapeutic avenues, as conventional treatments seldom result in a cure.
Oxidative stress plays a crucial role in GBM cells, influencing both tumor progression and therapeutic response [4]. GBM cells typically exhibit elevated levels of reactive oxygen species (ROS) due to their high metabolic activity and inherent genetic instability [5,6]. Oxidative stress relies in large part on a mismatch between the cellular antioxidant defense system and excess formation of ROS, including free radicals, which are molecules with unpaired electrons at the outer orbitals. This increased oxidative stress contributes to the aggressive nature of the tumor by promoting DNA damage, mutagenesis, and cellular proliferation. Additionally, the dysregulation of antioxidant defenses in GBM cells creates a vulnerability that can be therapeutically exploited [5,7]. While initially appearing counterintuitive due to the association of ROS with cellular damage and malignant transformation, the selective induction of oxidative stress within cancer cells holds therapeutic promise, particularly in the context of aggressive malignancies such as GBM [4,5]. By further increasing ROS levels through pro-oxidant therapies, it is possible to push the oxidative stress beyond the threshold that cancer cells can tolerate, leading to their selective eradication while sparing normal cells [8]. Understanding the dynamics of oxidative stress in GBM cells is therefore essential for developing effective treatments and improving patient outcomes.
Recent trends in preclinical [3,9] and clinical studies [10] of GBM have focused on the utilization of doxorubicin (DOX), one of the earliest chemotherapy agents (CTA) in cancer treatment. Doxorubicin is considered a pro-oxidant therapy due to its ability to generate ROS within cancer cells. This CTA induces oxidative stress through various mechanisms. It can undergo redox cycling in the presence of cellular reductases, resulting in the production of superoxide anions and other ROS. Additionally, it can also enhance the fenton reaction by chelating iron and interfere with the electron transport chain in the mitochondria, leading to increased ROS production [11,12]. Conversely, Photodynamic Therapy (PDT) has emerged as a promising alternative therapeutic modality for GBM, owing to its ability to selectively target cancerous cells while minimizing damage to surrounding healthy tissue by generating oxidative stress [13]. Ongoing clinical trials aim to further elucidate the efficacy of PDT in GBM treatment [14,15]. An advantage of utilizing photosensitizers (PS) in PDT for GBM is their capacity to function as a photodiagnostic agents for evaluating the existence of any residual cells after surgery [16]. Moreover, PDT exhibits the capability to synergize with other treatment modalities within a combination framework [17]. These combination therapies often entail the administration of multiple anticancer agents in reduced dosages compared to individual administration, thereby enhancing efficacy, reducing toxicity, and mitigating the development of drug resistance by targeting diverse cancer pathways.
Key mutations found in GBM patients, including the amplification of the epidermal growth factor receptor (EGFR), as well as mutations in the genes encoding phosphatase and tensin homolog (PTEN) and tumor protein 53 (TP53), have been linked to the response to treatments and the accumulation of ROS, which play a significant role in gain-of-function (GOF) activities in cancer cells [18,19]. Accumulating evidence underscores the pivotal role of these GOF activities in promoting tumor progression and conferring resistance to various anticancer therapies, thereby underscoring the imperative to comprehend their underlying mechanisms in the context of therapeutic interventions [20]. Moreover, there is growing evidence suggesting that cells harboring mutations such as in TP53 may exhibit heightened susceptibility to pro-oxidant drugs compared to those with the wild-type form of this gene [21,22,23,24]. This heightened sensitivity facilitates the accumulation of deleterious ROS, ultimately leading to cellular damage and demise of cancer cells.
To evaluate the impact of pro-oxidant treatments, such as DOX chemotherapy and PDT with Me-ALA contemplating GBM heterogeneity, we propose examining their effects on various human GBM cell lines with different mutation conditions. Our study will encompass both individual treatment approaches and combined therapeutic regimens. Furthermore, an in silico study was conducted to investigate the correlation between specific gene mutations and the abnormal regulation of oxidative stress in patients with GBM. This investigation aimed to provide an explanation for the observed behavior in various cell line models.
2. Results
2.1. The Impact of Pro-Oxidant Therapies on the Cellular Viability of GBM Cells as Monotherapies
To assess the therapeutic efficacy of DOX, monocultures of GBM cells (LN229, U87MG, and T98G) were cultured in 96-well plates and exposed to varying concentrations of CTA for 24 hours. U87MG cells exhibited the highest sensitivity to the cytotoxic effect of DOX, with an IC50 of 0.14 ± 0.1 µM. In this regard, DOX significantly reduced cell viability even at the lowest concentration of 0.1 µM. Conversely, the LN229 cell line demonstrated the greatest resistance to DOX treatment, with an IC50 of 6.88 ± 0.6 µM compared to the IC50 of 0.5 ± 0.15 µM observed for T98G (Figure 1A).
A statistical comparison of dose-response curves obtained by nonlinear regression fitting (Hill equation) was conducted among the cell lines. LN229 exhibited the highest level of resistance, and the other two cell lines displayed comparable behavior. These results obtained through the assessment of metabolic activity with MTT are supported by observed cellular morphological changes consistent with cell death (Figure S1). Upon comparing the effects induced by the use of CTA, lesser morphological changes are observed for the LN229 cell line in comparison to U87MG and T98G at equivalent concentrations of DOX.
On the other hand, the prooxidant effect of PDT with different Me-ALA concentrations were assayed (0.01 - 2 mM) using a light dose of 1 J/cm2. Me-ALA functions as a prodrug, undergoing bioconversion to the endogenous photosensitizer PpIX [25]. Both Me-ALA in concentrations up to 2 mM and irradiation alone with light doses up to 20 J/cm2 did not induce cell death alone (Figure S2). When comparing the concentration of 0.1 mM using 1 J/cm2, LN229 showed no decrease in cellular viability, while U87MG and T98G showed a decrease of 5% and 40% respectively. At a concentration of 0.5 mM, the cellular viability percentages were 64%, 36%, and 28% for LN229, U87MG, and T98G, respectively. The study found that the LN229 cell line exhibited the highest level of resistance to PDT with Me-ALA, whereas the T98G and U87MG cell lines were the most susceptible (Figure 1B). The IC50 calculated were 0.8; 0.33 and 0.3 mM for LN229, U87MG and T98G respectively. Results were supported by morphological changes observed after PDT treatment (Figure S3).
By analyzing the fluorescence emission of DOX and PpIX subsequent to the incubation with Me-ALA, flow cytometry was employed to quantify the cellular uptake of CTA and the production of PpIX. The results are shown in Figure 2 for the three GBM cell lines. An increase in the concentration of DOX exposure resulted in a corresponding rise in cellular uptake. Furthermore, an increase in PpIX production was noted in each of the three GBM cell lines in a manner dependent on the concentration of Me-ALA (Figure 2B). The LN229 cell line exhibited the highest incorporation of both DOX and PpIX, followed by the T98G cell line, with the U87MG cell line demonstrating the smallest values, mainly in the highest concentrations for DOX (1.5 μM: p <0,0001 and 10 μM: p <0,0001) and highest concentrations for Me-ALA (0.1 μM: p <0,0001 and 0.5 μM: p <0,0001).
2.2. The Impact of Pro-Oxidant Therapies on ROS Production in GBM Cells as Monotherapies
Since both DOX and PDT cause oxidative stress to initiate cell death, the decision was made to assess the level of oxidative stress by using the DCFDA probe [26] in the three GBM cell lines treated with each therapy individually. The fluorescence intensity histograms (Figure S4) were examined and normalized using the autofluorescence values of the control cells [26]. The findings are presented in Figure 3.
An increase in ROS production was observed in GBM cells treated with both DOX CTA and PDT in vitro. After being exposed to DOX, the three cell lines exhibited a significant increase in ROS levels in comparison to the control group. This suggests that the CTA, particularly at concentrations of 1.5 and 10 μM, induced significant oxidative stress. The disparities were marginally more noticeable in LN229 and U87MG cells when compared to T98G cells (Figure 3A). Similarly, PDT with Me-ALA treatment led to a significant rise in ROS production, reflecting the prooxidant nature of this therapeutic modality. In this case, the levels of ROS were higher than those achieved with DOX treatment at the tested concentrations (0.1 and 0.5 mM) (p <0,0001). The results of ROS production are consistent with those of PpIX bioproduction and cell viability assessment where there is a relationship between the endogenous PS production and the cytotoxic effect induced by ROS-mediated PDT irradiation. It was found that PDT using Me-ALA induced oxidative stress in all cell lines, with higher levels observed in the T98G and U87MG cell lines compared to DOX treatment (Figure 3B). Moreover, upon comparing among cell lines, it was noted that U87MG exhibited significantly higher values of DCF MFI, followed by T98G and, lastly, LN229. Therefore, it can be inferred that the U87MG cell line experiences the highest level of stress after being treated with PDT, as evidenced by the lethal impact of the therapy. LN229, in contrast, demonstrated the highest level of resistance to the cytotoxic prooxidant effects of PDT. This was evident from its minimal production of oxidative stress, which was significantly lower than the other two cell lines. These findings emphasize that both DOX and PDT have the capability to induce oxidative stress in GBM cells, indicating their potential as effective treatments for specifically targeting tumor cells through ROS-mediated mechanisms. However, when we analyze the two cell lines that experienced notable increases in ROS in both treatment methods, their cell viability showed contrasting patterns. The LN229 cell line, despite showing increased levels of ROS in response to therapeutic interventions, displays significant resistance to both treatments.
2.3. N-Acetylcysteine (NAC) Rescues Pro-Oxidant Cytotoxicity Induced by DOX and PDT Therapies
Cysteine acts as a precursor to the antioxidant glutathione, providing protection against ROS that trigger oxidative stress and lead to cell death [8]. In order to investigate the cytoprotective action of NAC and particularly reversing the deleterious effect of ROS generated by both pro-oxidant therapies, we decided to use NAC at a non-cytotoxic concentration (5 μM) prior to conducting both monotherapies. NAC pretreatment was found to improve cell viability under both pro-oxidant therapy (Figure 4) and reduced the cytotoxic effects of three concentrations of DOX in U87MG cells and in PDT treatment with 0.1 and 0.25 mM Me-ALA. As previously observed, the U87MG cell line was the most sensitive to both therapies, with a higher production of oxidative stress post-treatment. Therefore, the cytoprotection result with NAC supports the notion that the cytotoxic mechanism mediated by ROS is relevant in this cell line.
Conversely, the negative impact of the treatments was only reversed in the other two cell lines when greater concentrations of DOX were used (1 and 5 µM for T98G and 5 μM for LN229). These concentrations notably elevated oxidative stress in the DCF experiment. In the particular case of PDT with Me-ALA, the reversal effect was not as pronounced as with DOX treatment, likely due to the higher levels of ROS triggered by this therapy, probably surpassing the cellular antioxidant mechanisms including those provided by NAC.
2.4. Synergistic Prooxidant Combination of PDT and DOX Treatments
A therapeutic combination study was done to identify potential synergistic effects between the drugs being evaluated. By combining a chemotherapy regimen that targets tumor cells and their surrounding environment with the cytotoxic effects of selective PDT, it could serve as a supplementary therapeutic option to surgery. This integrated treatment approach was developed to address the current difficulties in treating GBM. The combination treatment involved exposing three GBM cell lines to varying concentrations of DOX, followed by the addition of varied amounts of Me-ALA in a radiation regimen similar to that used in monotherapy (Figure S6 and S7). The concentrations evaluated were based on previous monotherapy experiments and are near the IC50.
The combination of both treatments in LN229, as indicated by the matrix (Figure S5), showed that the highest percentages of inhibition were achieved with the highest concentrations of both Me-ALA and DOX (60-90%). In this sense, an additive effect in the combination of treatments for this cell line could be determined (Figure 5A), along with the possibility of therapeutic synergy for the combined treatment with the highest tested concentrations of 5 mM Me-ALA and 5 μM DOX. The synergy score for the LN229 cell line was 7.132.
The therapeutic combination for the U87MG cell line generated greater cytotoxic effects than those observed for the monotherapies of DOX and PDT. Upon analyzing the combination matrix, it was observed that percentages of inhibition above 60% were obtained for more combinations of both treatments compared to the other cell lines. The synergy score demonstrated a synergistic effect of the therapeutic combination, reflected in a score of 12.5 (Figure 5B).
Finally, the cell inhibition percentages corresponding to the combination of PDT with DOX presented in the matrix (Figure 5C) suggest that for any combination of these two treatments using a concentration equal to or greater than 0.1 mM of Me-ALA, cell inhibition percentages greater than 88% are achieved. The same occurs for any combination with the highest concentration of DOX, for which over 95% cell inhibition was obtained. The synergy score was -7.4, indicating an additive effect. These results were reflected in the levels of oxidative stress achieved with both therapies in combination.
A methodology was developed to evaluate the oxidative stress elicited by the treatment combinations in a synergistic modality. It included exposure to 0.5 μM DOX for 24 hours, followed by incubation with 0.1 mM Me-ALA and irradiation of 1 J/cm2. Following the administration of both therapies, the degree of oxidative stress was assessed utilizing the DCF probe. Oxidative stress levels increased in all three cell lines when both combination treatments were used, as opposed to when PDT was used alone (Figure 6). The studied combination exhibited significantly higher levels in the U87MG and T98G cell lines compared to lower values in the LN229 cell line.
2.5. Reactive Oxidative Homeostasis Evaluation in GBM Cell Lines
GBM cancer cells exhibit a notable characteristic of having both high amounts of ROS and excessive production of intracellular antioxidant enzymes, which distinguishes them from normal cells [8,27,28]. In order to examine the function of basal oxidative stress homeostasis in GBM cells, we analyzed the basal ROS level and mRNA expression levels of the most pertinent antioxidant enzymes and related genes in the three GBM cell lines. The objective was to determine if these factors could be used as biomarkers to predict the effectiveness of ROS-based therapies. Basal ROS levels and the expression of genes related to the oxidative response were evaluated among GBM cell lines. Results are presented in Figure 7. Our results indicated significant heterogeneity in basal ROS levels across the cell lines, with U87MG exhibiting the highest ROS levels, followed by LN229 and T98G, which demonstrated intermediate and low ROS levels, respectively (Figure 7A). In parallel, the mRNA levels of key antioxidant response genes, including NFE2L2 (NRF2), glutathione superoxide reductase (GSR), and catalase (CAT), were quantitatively analyzed by 2^-ΔΔCt method (Figure 7B, 7C and 7D). Notably, T98G and LN229 showed markedly elevated mRNA levels of these genes compared to U87MG, suggesting an improved compensatory upregulation in response to oxidative stress. Conversely, U87MG, with the highest ROS levels, exhibited the least expression of antioxidant response mRNA. These findings underscore the variability in oxidative stress management among GBM cell lines and highlight the potential for differential sensitivity to ROS-targeted therapies.
2.6. Comprehensive Analysis of Oxidative Stress-Related Genes in Glioblastoma and Their Relationship with TP53 and PTEN Mutations
To investigate the role of oxidative stress in GBM, a comprehensive analysis of oxidative stress-related genes was undertaken. The gene list was curated from the Gene Ontology (GO) term GO:0045454 ("cell redox homeostasis") (https://amigo.geneontology.org/amigo/term/GO:0045454) and supplemented with relevant literature findings. An in silico gene expression analysis was conducted using the TCGA Pan-Cancer Atlas for GBM patient data obtained from cBioPortal. Considering the documented involvement of PTEN and TP53 mutations in oxidative stress modulation [21,22,23,24,29,30,31], the 145 GBM patients were stratified into four groups based on their mutation status (Figure 8A):
- TP53 WT - PTEN WT (n = 64): Patients with wild-type (WT) TP53 and PTEN genes.
- TP53 WT - PTEN mut (n = 30): Patients with WT TP53 and mutated (mut) PTEN genes.
- TP53 mut - PTEN WT (n = 31): Patients with mutated TP53 and WT PTEN genes.
- TP53 mut - PTEN mut (n = 20): Patients with mutated TP53 and PTEN genes.
To visualize relationships within the data, a heatmap was generated (Figure 8B). Rows were centered and scaled to ensure all features were on an equal footing. Hierarchical clustering was performed using correlation distance and average linkage on both rows and columns. This analysis identified two distinct clusters. Interestingly, the clustering appeared to segregate samples primarily based on the mutational status of the TP53 gene. Group 1 contained samples with wild-type TP53 regardless of PTEN mutation status, while Group 2 included samples with mutated TP53, again with PTEN mutation playing a secondary role in cluster assignment.
Out of the 43 genes analyzed, 6 exhibited statistically significant differences between some of the groups studied. Patients with double mutations (TP53 mut and PTEN mut) showed upregulation of ERO1A (Figure 8C) and TXNRD1 (Figure 8D). ERO1A, encoded by the Ero1a gene, is crucial for protein folding but can also generate ROS as a byproduct, contributing to oxidative stress [32]. TXNRD1 is an oxidoreductase associated with elevated ROS levels and poor prognosis in cancer [33]. This pattern suggests a potential shift towards a pro-oxidant state in patients with combined TP53 and PTEN mutations. This may explain the susceptibility of the T98G cell line (double mutant) to pro-oxidant antitumor therapies.
Interestingly, within TP53 mutants, the presence of a PTEN mutation is linked to an upregulation of the prooxidant ERO1A and a downregulation of the antioxidant GIT1 (Figure 8E). GIT1 is known for its role in regulating the Keap1-Nrf2 signaling pathway, crucial for orchestrating the cellular antioxidant response [34]. In PTEN wild-type patients, an overexpression of GIT1 was observed in the presence of TP53 mutations. However, this heightened expression coincided with a decrease in the expression of key antioxidant genes such as HVCN1 (Figure 8F), NFE2L2 (Figure 8G), and Selenos (Figure 8H), indicating a compromise in the overall antioxidant defense system. Selenos, recognized for its antioxidant and anti-inflammatory functions, helps maintain endoplasmic reticulum (ER) morphology and regulates ER stress [35]. NRF2, transcription factor encoded by the gene nfe2l2, acting as the master regulator of the cellular antioxidant response, binds to antioxidant response elements (ARE) within the promoter regions of numerous cytoprotective genes, including phase 2 detoxifying enzymes, thereby facilitating their expression and mitigating the effects of reactive electrophiles [36]. Additionally, recent insights suggest that the regulation of ROS production by VSOP/Hv1, protein encoded by hvcn1 gene, can be pro or antioxidant, depending on age [37]. These findings could shed light on why the LN229 cell line exhibits the greatest resistance to prooxidant therapies. This cell line carries PTEN WT and P53 mut P98L (C293T); the mutation on TP53, which does not affect its DNA binding ability, results in partial protein functionality—an effect that is not fully understood (functioning similarly to WT) [38]. This resistance would be further supported by its consistent overexpression of protective antioxidant systems.
These findings underscore the intricate interplay between TP53 and PTEN mutations in modulating the oxidative stress response in GBM. TP53 mutations seem to play a dominant role, potentially influencing the expression of both pro-oxidant and antioxidant genes. Further studies are needed to confirm these observations and explore the therapeutic potential of targeting specific components of the oxidative stress pathway in GBM based on TP53 and PTEN mutation status.
3. Discussion
GBM cells utilize moderate amounts of ROS and reactive nitrogen species (RNS) to facilitate their proliferation and invasion [39]. It should be mentioned that certain mutations can lead to increased production of ROS and for this reason, tumor cells consistently generate a greater amount of ROS compared to normal cells. However, the resulting oxidative stress can be counteracted by an increased overall antioxidant capacity, which is achieved through the buildup of antioxidant molecules as a result of an advanced adaptation [40]. GBM cells rely heavily on these systems to neutralize the harmful effects of ROS generation. Therefore, overwhelming the cells with excessive ROS production could be a potential strategy to eliminate tumor cells [8,41].
The impact of prooxidant therapies on GBM cell lines as monotherapies was assessed through a series of assays, shedding light on their effectiveness and underlying mechanisms. These cell lines are frequently employed in vitro models for GBM, exemplifying certain characteristics of the disease's intrinsic intratumoral and intertumoral heterogeneity [42]. For instance, the T98G and LN229 cell lines harbor mutations in TP53, a gene frequently dysregulated across various tumor types and prevalent in both primary and secondary GBMs (30% and 65% incidence, respectively). In contrast, the U87MG cell line expresses the wild-type functional form of TP53. Cells with perturbations in the p53 pathway exhibit diverse capacities to mitigate oxidative stress and have been implicated in processes such as invasion, migration, proliferation, and evasion of apoptosis [43,44,45]. Notably, the M237I p53 mutation, found in the T98G cell line, has been associated with the acquisition of resistance to standard CTA [46,47]. It is evident that the basal oxidative stress varies among these cell lines, resulting in varied amounts of basal ROS (Figure 7A). These varying ROS levels are associated with distinct antioxidant responses. In this study, the antioxidant response was assessed by examining the gene expression of specific molecules involved in the antioxidant response. For instance, NRF2 is the primary transcription factor that controls the antioxidative response [36], and its expression levels differed among GBM cell lines (Figure 7B). Currently, many treatment approaches are being examined to inhibit this biological pathway [23,48].
DOX monotherapy revealed varying sensitivity among GBM cell lines (LN229, U87MG, T98G), with U87MG cells displaying the highest susceptibility and LN229 cells exhibiting the greatest resistance to DOX. These observations were supported by discernible cellular morphological changes indicative of apoptotic cell death. Conversely, PDT with Me-ALA exhibited divergent efficacy among the cell lines, with LN229 cells demonstrating notable resistance and U87MG and T98G cells displaying heightened susceptibility. PDT presents itself as a compelling approach to provoke cell death in tumor cells by generating ROS and thereby causing a redox imbalance. In this study, we opted to utilize the prodrug Me-ALA, which is a methylated variant that possesses greater lipophilicity compared to ALA. ALA is one of the primary PS examined in PDT for GBM [14,49]. Previously, we have demonstrated greater resistance to PDT with PS based on conjugated polymer nanoparticles in T98G cells, which coincided with elevated expression levels of antioxidant enzymes and may be related to the mutational status of TP53 [8]. In this study, PDT with Me-ALA induced effective cell death with an elevation of ROS surpassing the antioxidant levels possessed by the cell line, possibly due to the higher production of PpIX observed in these cells. Based on the outcomes of post-treatment cell viability assessments, including the generation of endogenous PS and cellular incorporation of DOX, it can be deduced that the LN229 cell line exhibits the greatest resistance to both treatments when used as monotherapies, despite having the highest level of cellular uptake.
Both DOX and PDT Me-ALA treatments led to increased ROS production in GBM cells, with higher ROS levels observed with PDT compared to DOX. U87MG cells exhibited the highest ROS levels post-treatment, indicating their heightened susceptibility to oxidative stress-induced cell death. Meanwhile, LN229, although it shares a mutation of the TP53 gene, it is not the same mutation that the U87MG line has. Furthermore, the mutation status of the PTEN gene is different between both cell lines. This may suggest the differential behavior between both cell lines. Pre-treatment with NAC rescued cytotoxicity induced by DOX and PDT in U87MG cells, emphasizing the role of ROS in their cytotoxic mechanism.
Combining PDT with DOX resulted in synergistic cytotoxic effects in GBM cell lines, with U87MG cells showing the highest sensitivity to the combined treatment. Analysis of oxidative stress levels following combination therapy revealed increased stress levels across all cell lines, particularly pronounced in U87MG and T98G cells. The level of oxidative stress within cells reflects a balance between the rate of ROS production and the activity of detoxifying systems that neutralize them [50]. The increased basal oxidative stress in transformed cells, particularly in tumor cells harboring mutations in tumor suppressor genes such as TP53 and PTEN throughout the carcinogenesis process, renders them highly dependent on their antioxidant systems to counteract the harmful effects of ROS [21]. Interestingly, this dependence could potentially be a vulnerability for cancer cells that have a mutated TP53 gene. For instance, this vulnerability is demonstrated by the increased sensitivity of these cells to treatment with H2O2 [51]. However, previous research has also demonstrated that mutated p53 enhances the synthesis of antioxidant enzymes as a defensive reaction to the oxidative circumstances induced by the GOF activities of the mutant protein [22,43].
Besides, PTEN has a crucial role in cellular metabolic regulation and response to oxidative stress. The main function of this is to protect cells from cell death caused by oxidative stress by regulating the PI3K/AKT signaling pathway to limit the production of oxidative stress [29,52]. However, when PTEN is lost, it disrupts the equilibrium of redox processes, leading to increased oxidative stress. This has the potential to affect the response of GBM cells to pro-oxidant treatments as well. A prior investigation has demonstrated that GBM U87MG cells harboring a mutant form of PTEN, which leads to its loss of function, display elevated levels of oxidative stress in comparison to those identical cells in which PTEN functionality has been reinstated by overexpressing the wild-type variant [29]. Therefore, we considered the mutational status of both PTEN and TP53 in the three cell lines employed in this study. U87MG and T98G presented mutations in PTEN with loss of function of its enzymatic activity, meanwhile LN229 had the wildtype PTEN gene.
Mutational analysis of TP53 and PTEN genes revealed distinct gene expression profiles associated with oxidative stress response. Patients with combined TP53 and PTEN mutations showed upregulation of prooxidant genes like ERO1A and TXNRD1, suggesting a prooxidant shift in these cases. TP53 mutations appeared to influence the expression of both prooxidant and antioxidant genes, with potential implications for treatment response. LN229 cells, characterized by PTEN wild-type and TP53 mutant status, exhibited resistance to prooxidant therapies, potentially due to enhanced expression of protective antioxidant systems.
These findings highlight the complex interplay between prooxidant therapies, ROS production, and genetic factors in GBM, underscoring the importance of personalized treatment strategies targeting specific molecular pathways involved in oxidative stress response. Insufficient antioxidant defenses lead to oxidative stress, resulting in cell death when the harmful effects of ROS are not reduced. Further research is warranted to validate these observations and explore their clinical implications for GBM management.
4. Materials and Methods
4.1. Cell Lines and Culture Conditions
In this study, three distinct human GBM tumor cell lines were employed. The first cell line, U87MG (ATCC® HTB-14™) with wild-type TP53 and mutant PTEN was originated from a GBM specimen obtained from an adult male patient. The second cell line, T98G (ATCC® CRL-1690™), with mutated TP53 (M237I) and PTEN genes, was derived from a GBM specimen obtained from a 61-year-old male patient. Finally, the third cell line, LN229 (ATCC® CRL-2611™), was isolated from the right frontal parieto-occipital cortex of a 60-year-old Caucasian patient diagnosed with GBM. In contrast to the first two cell lines, this cell line harbors mutant TP53 (P98L) and wild-type PTEN. All GBM cell lines were cultured in Dulbecco's Modified Eagle's Medium (DMEM, Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS, Internegocios, S.A, Buenos Aires, Argentina).
4.2. Cell Uptake Analysis of DOX and Bioproduction of PpIX
GBM cell lines were cultured in 24-well plates with 20,000 cells per well and incubated at a temperature of 37°C in a CO2 incubator for one night. Subsequently, the media was substituted with DOX at different concentrations (0.1, 0.5, 1.5, and 10 μM). The cells were then incubated for 24 hours, and the fluorescence of the CTA in the cells was assessed using flow cytometry in the red-B detector channel. On the other hand, to induce intracellular formation of PpIX, the three cell lines were seeded onto 24-well plates at a density of 20,000 cells. Subsequently, cells were exposed to media containing Me-ALA at concentrations of 0.01, 0.1, and 0.5 mM for a duration of 4 hours. In order to quantify PpIX formation, cell fluorescence was subsequently analyzed by flow cytometry in the red-B channel.
4.3. Assessment of the Efficacy of DOX as a Single Treatment in GBM Cell Lines
The cell suspensions were prepared at a concentration of 200,000 cells per milliliter and seeded onto 96-well plates at a density of 20,000 cells per well. The cells were cultured for 24 hours at a temperature of 37°C in a CO2 incubator. Subsequently, various concentrations of DOX were examined within a range of 0.1 - 102.4 μM, with a duration of exposure of 24 hours. After that time, the CTA medium was extracted from the plate. A solution of MTT in complete DMEM (0.5 mg/mL) was prepared, and 100 μL of this solution was added to each well. The plate was then incubated in darkness, under the same conditions as before, for a duration of 3 hours. Next, the medium was disposed of and 100 μL of DMSO was introduced to dissolve the formazan crystals. Ultimately, the measurement of absorption was conducted utilizing a multiplate reader (Multiskan, Thermo Scientific) at a specific wavelength of 570 nm. The absorption values of the treated groups were compared to the control group, which did not receive any treatment. The values were transformed into percentages, representing cell viability relative to the control, which was considered to have 100% viability.
4.4. Assessment of the Impact of TFD with Me-ALA on the Survival of GBM Cells
Cell viability was assessed following exposure to varying doses of Me-ALA in the three GBM cell lines. To achieve this, a 100 mM solution of the photosensitizer Me-ALA was prepared. Cell suspensions were prepared at a concentration of 200,000 cells per milliliter and seeded onto 96-well plates at a density of 20,000 cells per well. Afterwards, solutions of Me-ALA were prepared in concentrations of 0.01, 0.1, 0.5, 1, and 2 mM in serum-free DMEM. The same medium was used for the control samples. The complete DMEM that was originally in the plates was removed, and two washes were performed with PBS 1X. The prepared Me-ALA solutions were then added to the plates, and the samples were incubated in the dark for 4 hours. Following this period, irradiation was performed utilizing a multiLED system emitting light at a wavelength of 635 ± 15 nm. The light dosage applied was 1 J/cm2, with a luminous output of 21.25 mW/cm2. Subsequently, the media was replaced with DMEM containing 10% SFB and the cells were incubated for 24 hours. The following day, the feasibility of the cells was assessed using the MTT colorimetric test, as previously reported.
4.5. Reversal of Prooxidant Cytotoxic Effects in Monotherapy with DOX and PDT through N-Acetylcysteine Intervention
The three GBM cell lines were seeded in 96-well plates at a density of 10,000 cells per well. On the next day, a solution containing N-acetylcysteine (NAC) in 10% FBS DMEM (5 µM) was introduced to the specified wells and left to incubate for 24 hours. Following this period, the cells were subjected to DOX treatment at concentrations of 0.1, 1, and 5 µM for an additional 24 hours. Certain wells were administered chemotherapy exclusively, without the inclusion of NAC. In addition, GBM cell lines were subjected to pre-incubation with Me-ALA (0.1 – 0.25 - 0.5 mM) for a duration of 24 hours before being incubated with NAC as described previously. The cells were exposed to 1 J/cm2 of light after being incubated with Me-ALA for 4 hours. Cell viability was assessed 24 hours after treatments using MTT, and a comparison was made between wells that were incubated with NAC (NAC +) and those that were not (NAC -).
4.6. Citotoxicity effect of Combination of Doxorubicin and PDT with Me-ALA
We performed an experiment to evaluate the effect of combining DOX and PDT with Me-ALA on the viability of three different cell lines (U87MG, T98G, and LN229). Cells were placed at a concentration of 20,000 cells per well in 96-well plates for this experiment. After 24 hours of culturing, the cells were treated with different concentrations of DOX close to the IC50 values (0.05, 0.5, and 5 µM for T98G and LN229, and 0.01, 0.1, and 1 µM for U87MG) for 24 hours. After removing DOX, cells were treated with Me-ALA at concentrations of 0.01, 0.1, and 0.5 mM for 4 hours, then subjected to irradiation of 1 J/cm2. Cell viability in each treatment was evaluated using an MTT assay after 24 hours.
4.7. Determination of Synergy between Doxorubicin and PDT with Me-ALA
The interaction of DOX and PDT with Me-ALA was assessed using SynergyFinder, a tool available at https://synergyfinder.fimm.fi/. The analysis was conducted on May 2024. SynergyFinder is a web-based application that assesses the synergy between a combination of medications using different reference models. In order to do this, we utilized the Highest Single Agent (HSA) model. This model measures the level of synergy by comparing it to the greatest response of a single agent. It suggests that the predicted combined impact is equal to the maximum response of a single drug at the corresponding concentrations. Summarized synergy scores represent the mean additional reaction resulting from pharmacological interactions. For example, a synergy score of 15 equates to a 15% response over the anticipated level. The acquired synergy scores were categorized as synergistic (synergy score > 10), additive (synergy score ranging from -10 to 10), or antagonistic (synergy score < -10).
4.8. Evaluation of Intracellular Oxidative Stress after Mono and Combo Treatment
GBM cell lines were seeded into 24-well plates with 500 μL of a prepared cell suspension containing 100,000 cells/mL per well, resulting in a total of 50,000 cells in each well. These plates were then incubated for 24 hours at 37°C with 5% CO2.
Following the incubation period, the medium was aspirated from all wells, and distinct concentrations of DOX (0.1, 0.5, 1.5 and 10 μM), approximating the IC50 value, were applied for 24 hours. Additionally, three concentrations of Me-ALA near the IC50 (0.01, 0.1 and 0.5 mM) were administered for 4 hours, followed by irradiation with a light dose of 1 J/cm2. Immediately after these separate treatments, the medium was aspirated, and a solution of CM-H2DCFDA in PBS at a concentration of 10 μM was introduced.
For the combined treatment, cells were seeded as previously described and incubated with DOX at 0.5 μM for 24 hours, followed by incubation with Me-ALA at 0.1 mM, and irradiation with a light dose of 1 J/cm2. DCF fluorescence was assessed immediately after PDT treatment. To serve as a positive control, certain wells were treated with a solution of hydrogen peroxide (30% v/v, diluted 1/200 in PBS) and incubated for 20 minutes at 37°C. Following a 30-minute incubation period with the oxidative stress probe, the medium was aspirated, and complete medium was added for a further incubation period of 10 minutes. Subsequently, the medium was removed, wells were rinsed with 1X PBS, and cells were detached using 1% trypsin.
The samples underwent analysis using flow cytometry (Guava easycyte 6 2L), with the green-B detector utilized to analyze the fluorescence of the DCF probe.
4.9. Gene Expression Analysis of Antioxidant Response in GBM Cell Lines
A quantitative polymerase chain reaction (qPCR) was conducted to assess the gene expression of several molecules involved in the antioxidant response of GBM cell lines. RNA was isolated from U87MG, LN229, and T98G cells cultured under growth conditions using TRIzol reagent (Invitrogen, Thermo Fisher Scientific). The extracted RNA was then reverse transcribed using M-MLV reverse transcriptase (Invitrogen, Thermo Fisher Scientific) following the manufacturer's instructions. The qPCR measurement was conducted using the SYBR Green qPCR Master Mix from Agilent Technologies, CA, USA. Each reaction contained 10 ng of cDNA and was performed on Agilent's Stratagene Mx3000PRO equipment. [53]. ACTB gene expression was used as housekeeping gene to normalize the expression of target genes and the 2-ΔΔCT method was used to calculate relative levels of gene expression using the Strata gene MxPro QPCR software v3.00 tool (Stratagene, Agilent Technologies). The quality of amplification was confirmed using a typical melting-curve cycle. The samples were subjected to triplicate analysis, and the experiment was replicated two times. Forward and reverse primers for the genes of interest are shown in Table 1.
4.10. Expression Analysis of Cell Redox Homeostasis Genes in GBM Patient Samples
In silico analysis leveraged the TCGA Pan-Cancer Atlas data accessible through cBioPortal (accessed May 2024) [54]. This analysis focused on 145 glioblastoma (GBM) patients. Mutation status of TP53 and PTEN genes, along with mRNA expression of relevant genes, was retrieved for diploid samples. Venn diagrams were generated using InteractiVenn [55], while heatmaps were constructed using the online platform ClustVis [56]. Patients were stratified into four groups based on TP53 and PTEN mutation status: TP53 WT - PTEN WT (n = 94): patients with wild-type (WT) TP53 and PTEN genes; TP53 WT - PTEN mut (n = 30): patients with WT TP53 and mutated (mut) PTEN genes; TP53 mut - PTEN WT (n = 31): patients with mutated TP53 and WT PTEN genes; TP53 mut - PTEN mut (n = 20): patients with mutated TP53 and PTEN genes. The analysis focused on mRNA expression of genes associated with the Gene Ontology (GO) term "cell redox homeostasis" (GO:0045454) (https://www.ebi.ac.uk/QuickGO/term/GO:0045454) and was further supplemented by relevant literature findings on oxidative stress.
4.11. Statistical Analysis
The data collected from the cell uptake, ROS production, and cytotoxicity experiments were analyzed using Graph Prism software. One-way ANOVA was used to determine the significant differences in cell viability for cells treated with a single treatment of DOX and PDT with Me-ALA. The fluorescence intensity of DOX uptake and PpIX bioproduction was measured for each treatment group and normalized to the autofluorescence of untreated cells. The acquired data was subsequently analyzed using Graph Prism software. Two-way ANOVA was used to determine the significant difference in cell uptake and intracellular bioproduction of PpIX among various treatment groups considering different concentrations and the behavior of cell lines. The experiments were conducted with a sample size of n=6 and repeated twice.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Figure S1: title; Table S1: title; Video S1: title.
Author Contributions
Conceptualization, L.E.I.; methodology, B.A.C., M.D.C., M.J.L. and L.E.I.; software, B.A.C.; validation, M.J.L. and L.E.I.; formal analysis, B.A.C., M.D.C., M.J.L. and L.E.I.; investigation, B.A.C., M.D.C., M.J.L. and L.E.I.; resources, M.J.L. and L.E.I.; data curation, B.A.C., M.D.C., M.J.L. and L.E.I.; writing—original draft preparation, B.A.C., M.J.L. and L.E.I.; writing—review and editing, M.J.L. and L.E.I.; visualization, L.E.I.; supervision, L.E.I.; project administration, L.E.I.; funding acquisition, L.E.I. All authors have read and agreed to the published version of the manuscript.”
Funding
This research was funded by grants from Agencia Nacional de Promoción Científica y Tecnológica (PICT) (PICT-2020-SERIEA-00051;), SECyT, UNRC and CONICET (PIBAA 2022–2023 28720210100004CO) to L.E.I.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
M.D.C thanks CONICET for PhD scholarship. B.A.C. thanks Consejo Interuniversitario (CIN) for student scholarship. M.J.L. and L.E.I. are members of the Scientific Researcher Career at CONICET and faculty at UNRC.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: a summary. Neuro. Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Ah-Pine, F.; Khettab, M.; Bedoui, Y.; Slama, Y.; Daniel, M.; Doray, B.; Gasque, P. On the origin and development of glioblastoma: multifaceted role of perivascular mesenchymal stromal cells. Acta Neuropathol. Commun. 2023 111 2023, 11, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Oraiopoulou, M.E.; Tzamali, E.; Psycharakis, S.E.; Tzedakis, G.; Makatounakis, T.; Manolitsi, K.; Drakos, E.; Vakis, A.F.; Zacharakis, G.; Papamatheakis, J.; et al. The Temozolomide–Doxorubicin paradox in Glioblastoma in vitro–in silico preclinical drug-screening. Sci. Reports 2024 141 2024, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ostrowski, R.P.; Pucko, E.B. Harnessing oxidative stress for anti-glioma therapy. Neurochem. Int. 2022, 154, 105281. [Google Scholar] [CrossRef] [PubMed]
- Olivier, C.; Oliver, L.; Lalier, L.; Vallette, F.M. Drug Resistance in Glioblastoma: The Two Faces of Oxidative Stress. Front. Mol. Biosci. 2021, 7, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Ramiro, A.; Ramírez-Ortega, D.; de La Cruz, V.P.; Hérnandez-Pedro, N.Y.; González-Esquivel, D.F.; Sotelo, J.; Pineda, B. Role of redox status in development of Glioblastoma. Front. Immunol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Du, S.; Du, Y.; Ren, J.; Ying, G.; Yan, Z. Glutathione reductase mediates drug resistance in glioblastoma cells by regulating redox homeostasis. J. Neurochem. 2018, 144, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Caverzán, M.D.; Beaugé, L.; Chesta, C.A.; Palacios, R.E.; Ibarra, L.E. Photodynamic therapy of Glioblastoma cells using doped conjugated polymer nanoparticles: An in vitro comparative study based on redox status. J. Photochem. Photobiol. B Biol. 2020, 212, 112045. [Google Scholar] [CrossRef]
- Dhungel, L.; Rowsey, M.E.; Harris, C.; Raucher, D. Synergistic Effects of Temozolomide and Doxorubicin in the Treatment of Glioblastoma Multiforme: Enhancing Efficacy through Combination Therapy. Mol. 2024, Vol. 29, Page 840 2024, 29, 840. [Google Scholar] [CrossRef]
- Butt, O.H.; Zhou, A.Y.; Huang, J.; Leidig, W.A.; Silberstein, A.E.; Chheda, M.G.; Johanns, T.M.; Ansstas, G.; Liu, J.; Talcott, G.; et al. A phase II study of laser interstitial thermal therapy combined with doxorubicin in patients with recurrent glioblastoma. Neuro-Oncology Adv. 2021, 3, 1–10. [Google Scholar] [CrossRef]
- Peter, S.; Alven, S.; Maseko, R.B.; Aderibigbe, B.A. Doxorubicin-Based Hybrid Compounds as Potential Anticancer Agents: A Review. Mol. 2022, Vol. 27, Page 4478 2022, 27, 4478. [Google Scholar] [CrossRef]
- Cappetta, D.; De Angelis, A.; Sapio, L.; Prezioso, L.; Illiano, M.; Quaini, F.; Rossi, F.; Berrino, L.; Naviglio, S.; Urbanek, K. Oxidative stress and cellular response to doxorubicin: A common factor in the complex milieu of anthracycline cardiotoxicity. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef]
- Ibarra, L.E.; Camorani, S.; Agnello, L.; Pedone, E.; Pirone, L.; Chesta, C.A.; Palacios, R.E.; Fedele, M.; Cerchia, L. Selective Photo-Assisted Eradication of Triple-Negative Breast Cancer Cells through Aptamer Decoration of Doped Conjugated Polymer Nanoparticles. Pharmaceutics 2022, 14, 626. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, L.E.; Vilchez, M.L.; Caverzán, M.D.; Milla Sanabria, L.N. Understanding the glioblastoma tumor biology to optimize photodynamic therapy: From molecular to cellular events. J. Neurosci. Res. 2021, 99, 1024–1047. [Google Scholar] [CrossRef]
- Bhanja, D.; Wilding, H.; Baroz, A.; Trifoi, M.; Shenoy, G.; Slagle-Webb, B.; Hayes, D.; Soudagar, Y.; Connor, J.; Mansouri, A. Photodynamic Therapy for Glioblastoma: Illuminating the Path toward Clinical Applicability. Cancers 2023, Vol. 15, Page 3427 2023, 15, 3427. [Google Scholar] [CrossRef]
- Akimoto, J.; Fukami, S.; Ichikawa, M.; Mohamed, A.; Kohno, M. Intraoperative photodiagnosis for malignant glioma using photosensitizer talaporfin sodium. Front. Surg. 2019, 6, 421595. [Google Scholar] [CrossRef]
- Songca, S.P. Combinations of Photodynamic Therapy with Other Minimally Invasive Therapeutic Technologies against Cancer and Microbial Infections. Int. J. Mol. Sci. 2023, Vol. 24, Page 10875 2023, 24, 10875. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Dube, C.; Gibert, M.; Cruickshanks, N.; Wang, B.; Coughlan, M.; Yang, Y.; Setiady, I.; Deveau, C.; Saoud, K.; et al. The p53 pathway in glioblastoma. Cancers (Basel). 2018, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Ortiz, E.; de la Cruz-López, K.G.; Becerril-Rico, J.; Sarabia-Sánchez, M.A.; Ortiz-Sánchez, E.; García-Carrancá, A. Mutant p53 Gain-of-Function: Role in Cancer Development, Progression, and Therapeutic Approaches. Front. Cell Dev. Biol. 2021, 8, 607670. [Google Scholar] [CrossRef]
- Wang, W.; Cheng, B.; Miao, L.; Me, Y.; Wu, M. Mutant p53-R273H gains new function in sustained activation of EGFR signaling via suppressing miR-27a expression. Cell Death Dis. 2013 44 2013, 4, e574–e574. [Google Scholar] [CrossRef]
- Cordani, M.; Butera, G.; Pacchiana, R.; Masetto, F.; Mullappilly, N.; Riganti, C.; Donadelli, M. Mutant p53-Associated Molecular Mechanisms of ROS Regulation in Cancer Cells. Biomol. 2020, Vol. 10, Page 361 2020, 10, 361. [Google Scholar] [CrossRef] [PubMed]
- Lisek, K.; Campaner, E.; Ciani, Y.; Walerych, D.; Sal, G. Del; Lisek, K.; Campaner, E.; Ciani, Y.; Walerych, D.; Del Sal, G. Mutant p53 tunes the NRF2-dependent antioxidant response to support survival of cancer cells. Oncotarget 2018, 9, 20508–20523. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.S.; Duong, C.P.; Haupt, S.; Montgomery, K.G.; House, C.M.; Azar, W.J.; Pearson, H.B.; Fisher, O.M.; Read, M.; Guerra, G.R.; et al. Inhibiting the system xC−/glutathione axis selectively targets cancers with mutant-p53 accumulation. Nat. Commun. 2017 81 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Achanta, G.; Huang, P. Role of p53 in Sensing Oxidative DNA Damage in Response to Reactive Oxygen Species-Generating Agents. Cancer Res. 2004, 64, 6233–6239. [Google Scholar] [CrossRef] [PubMed]
- Lamberti, M.J.; Morales Vasconsuelo, A.B.; Ferrara, M.G.; Rumie Vittar, N.B. Recapitulation of Hypoxic Tumor–stroma Microenvironment to Study Photodynamic Therapy Implications. Photochem. Photobiol. 2020, 96, 897–905. [Google Scholar] [CrossRef] [PubMed]
- Caverzán, M.D.; Oliveda, P.M.; Beaugé, L.; Palacios, R.E.; Chesta, C.A.; Ibarra, L.E. Metronomic Photodynamic Therapy with Conjugated Polymer Nanoparticles in Glioblastoma Tumor Microenvironment. Cells 2023, 12, 1541. [Google Scholar] [CrossRef] [PubMed]
- Hardiany, N.S.; Mulyawan, W.; Wanandi, S.I. Correlation between oxidative stress and tumor grade in glioma cells from patients in Jakarta. Med. J. Indones. 2012, 21, 122–127. [Google Scholar] [CrossRef]
- Faraji-Rad, M.; Khajavi, M.; Arjmand, M.H.; Shajari, E.; Alamdari, D.H. Pro-oxidant-antioxidant balance in patients with high grade glioblastoma multiform. Middle East J. Cancer 2015, 6, 79–83. [Google Scholar]
- Luo, S.; Lei, K.; Xiang, D.; Ye, K. NQO1 is regulated by PTEN in glioblastoma, mediating cell proliferation and oxidative stress. Oxid. Med. Cell. Longev. 2018, 2018. [Google Scholar] [CrossRef]
- Lee, J.J.; Kim, B.C.; Park, M.J.; Lee, Y.S.; Kim, Y.N.; Lee, B.L.; Lee, J.S. PTEN status switches cell fate between premature senescence and apoptosis in glioma exposed to ionizing radiation. Cell Death Differ. 2011 184 2010, 18, 666–677. [Google Scholar] [CrossRef]
- Bankoglu, E.E.; Tschopp, O.; Schmitt, J.; Burkard, P.; Jahn, D.; Geier, A.; Stopper, H. Role of PTEN in Oxidative Stress and DNA Damage in the Liver of Whole-Body Pten Haplodeficient Mice. PLoS One 2016, 11, e0166956. [Google Scholar] [CrossRef] [PubMed]
- Appenzeller-Herzog, C.; Riemer, J.; Christensen, B.; Sørensen, E.S.; Ellgaard, L. A novel disulphide switch mechanism in Ero1α balances ER oxidation in human cells. EMBO J. 2008, 27, 2977–2987. [Google Scholar] [CrossRef] [PubMed]
- Leone, A.; Roca, M.S.; Ciardiello, C.; Costantini, S.; Budillon, A. Oxidative Stress Gene Expression Profile Correlates with Cancer Patient Poor Prognosis: Identification of Crucial Pathways Might Select Novel Therapeutic Approaches. Oxid. Med. Cell. Longev. 2017, 2017. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Gao, P.; Huang, Y.; Wu, M.; Yi, J.; Zhou, Z.; Zhao, X.; Jiang, T.; Liu, H.; Qin, T.; et al. Git1-PGK1 interaction achieves self-protection against spinal cord ischemia-reperfusion injury by modulating Keap1/Nrf2 signaling. Redox Biol. 2023, 62, 102682. [Google Scholar] [CrossRef] [PubMed]
- Gorini, F.; Vassalle, C. Selenium and Selenoproteins at the Intersection of Type 2 Diabetes and Thyroid Pathophysiology. Antioxidants 2022, Vol. 11, Page 1188 2022, 11, 1188. [Google Scholar] [CrossRef] [PubMed]
- Rojo de la Vega, M.; Chapman, E.; Zhang, D.D. NRF2 and the Hallmarks of Cancer. Cancer Cell 2018, 34, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Okochi, Y.; Ozaki, T.; Imura, Y.; Koizumi, S.; Yamazaki, M.; Abe, M.; Sakimura, K.; Yamashita, T.; Okamura, Y. Unconventional role of voltage-gated proton channels (VSOP/Hv1) in regulation of microglial ROS production. J. Neurochem. 2017, 142, 686–699. [Google Scholar] [CrossRef] [PubMed]
- Paulmurugan, R.; Afjei, R.; Sekar, T. V.; Babikir, H.A.; Massoud, T.F.; Paulmurugan, R.; Afjei, R.; Sekar, T. V.; Babikir, H.A.; Massoud, T.F. A protein folding molecular imaging biosensor monitors the effects of drugs that restore mutant p53 structure and its downstream function in glioblastoma cells. Oncotarget 2018, 9, 21495–21511. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, K.; Asthana, S.; Kumar, D. Role of Oxidative Stress in Metabolic Reprogramming of Brain Cancer. Cancers 2023, Vol. 15, Page 4920 2023, 15, 4920. [Google Scholar] [CrossRef]
- Ramírez-Expósito, M.J.; Martínez-Martos, J.M. The Delicate Equilibrium between Oxidants and Antioxidants in Brain Glioma. Curr. Neuropharmacol. 2018, 17, 342–351. [Google Scholar] [CrossRef]
- Popa-Wagner, A.; Mitran, S.; Sivanesan, S.; Chang, E.; Buga, A.M. ROS and brain diseases: The good, the bad, and the ugly. Oxid. Med. Cell. Longev. 2013. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.J.; Seo, H.W.; Baek, J.H.; Lim, S.H.; Hwang, S.G.; Kim, E.H. Gene expression profiling of glioblastoma cell lines depending on TP53 status after tumor-treating fields (TTFields) treatment. Sci. Reports 2020 101 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Torrens-Mas, M.; Cordani, M.; Mullappilly, N.; Pacchiana, R.; Riganti, C.; Palmieri, M.; Pons, D.G.; Roca, P.; Oliver, J.; Donadelli, M. Mutant p53 induces SIRT3/MnSOD axis to moderate ROS production in melanoma cells. Arch. Biochem. Biophys. 2020, 679, 108219. [Google Scholar] [CrossRef] [PubMed]
- Wong, R.P.C.; Tsang, W.P.; Chau, P.Y.; Co, N.N.; Tsang, T.Y.; Kwok, T.T. p53-R273H gains new function in induction of drug resistance through down-regulation of procaspase-3. Mol. Cancer Ther. 2007, 6, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Olafson, L.R.; Gunawardena, M.; Nixdorf, S.; McDonald, K.L.; Rapkins, R.W. The role of TP53 gain-of-function mutation in multifocal glioblastoma. J. Neurooncol. 2020, 147, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Pedrote, M.M.; Motta, M.F.; Ferretti, G.D.S.; Norberto, D.R.; Spohr, T.C.L.S.; Lima, F.R.S.; Gratton, E.; Silva, J.L.; de Oliveira, G.A.P. Oncogenic Gain of Function in Glioblastoma Is Linked to Mutant p53 Amyloid Oligomers. iScience 2020, 23. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Chen, J.X.; Liu, J.P.; You, C.; Liu, Y.H.; Mao, Q. Gain of function of mutant TP53 in glioblastoma: Prognosis and response to temozolomide. Ann. Surg. Oncol. 2014, 21, 1337–1344. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.R.D.V.; Pour Khavari, A.; Rizzo, M.; Sakamoto-Hojo, E.T.; Haghdoost, S. Targeting NRF2, Regulator of Antioxidant System, to Sensitize Glioblastoma Neurosphere Cells to Radiation-Induced Oxidative Stress. Oxid. Med. Cell. Longev. 2020, 2020. [Google Scholar] [CrossRef]
- Quach, S.; Schwartz, C.; Aumiller, M.; Foglar, M.; Schmutzer, M.; Katzendobler, S.; El Fahim, M.; Forbrig, R.; Bochmann, K.; Egensperger, R.; et al. Interstitial photodynamic therapy for newly diagnosed glioblastoma. J. Neurooncol. 2023, 162, 217–223. [Google Scholar] [CrossRef]
- Waris, G.; Ahsan, H. Reactive oxygen species: Role in the development of cancer and various chronic conditions. J. Carcinog. 2006, 5. [Google Scholar] [CrossRef]
- Cordani, M.; Butera, G.; Dando, I.; Torrens-Mas, M.; Butturini, E.; Pacchiana, R.; Oppici, E.; Cavallini, C.; Gasperini, S.; Tamassia, N.; et al. Mutant p53 blocks SESN1/AMPK/PGC-1α/UCP2 axis increasing mitochondrial O2ˉ· production in cancer cells. Br. J. Cancer 2018 1198 2018, 119, 994–1008. [Google Scholar] [CrossRef] [PubMed]
- Kitagishi, Y.; Matsuda, S. Redox regulation of tumor suppressor PTEN in cancer and aging (Review). Int. J. Mol. Med. 2013, 31, 511–515. [Google Scholar] [CrossRef] [PubMed]
- Ibarra, L.E.; Beaugé, L.; Arias-Ramos, N.; Rivarola, V.A.; Chesta, C.A.; López-Larrubia, P.; Palacios, R.E. Trojan horse monocyte-mediated delivery of conjugated polymer nanoparticles for improved photodynamic therapy of glioblastoma. Nanomedicine 2020, 15, 1687–1707. [Google Scholar] [CrossRef] [PubMed]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Heberle, H.; Meirelles, V.G.; da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinformatics 2015, 16, 1–7. [Google Scholar] [CrossRef]
- Metsalu, T.; Vilo, J. ClustVis: a web tool for visualizing clustering of multivariate data using Principal Component Analysis and heatmap. Nucleic Acids Res. 2015, 43, W566–W570. [Google Scholar] [CrossRef]
Figure 1.
Cell viability after DOX and PDT treatments as monotherapies. Quantification of cell viability by MTT assay 24 h after DOX (A) or PDT (B) treatments for U87MG, T98G and LN229 cell lines as a function of DOX concentration and Me-ALA concentration and light dose of 1 J/cm2 respectively. Cell viability percentages were normalized to control cells not exposed to treatments. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks ** p < 0.01, *** p < 0.001 and ns: no statistically significant differences.
Figure 1.
Cell viability after DOX and PDT treatments as monotherapies. Quantification of cell viability by MTT assay 24 h after DOX (A) or PDT (B) treatments for U87MG, T98G and LN229 cell lines as a function of DOX concentration and Me-ALA concentration and light dose of 1 J/cm2 respectively. Cell viability percentages were normalized to control cells not exposed to treatments. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks ** p < 0.01, *** p < 0.001 and ns: no statistically significant differences.
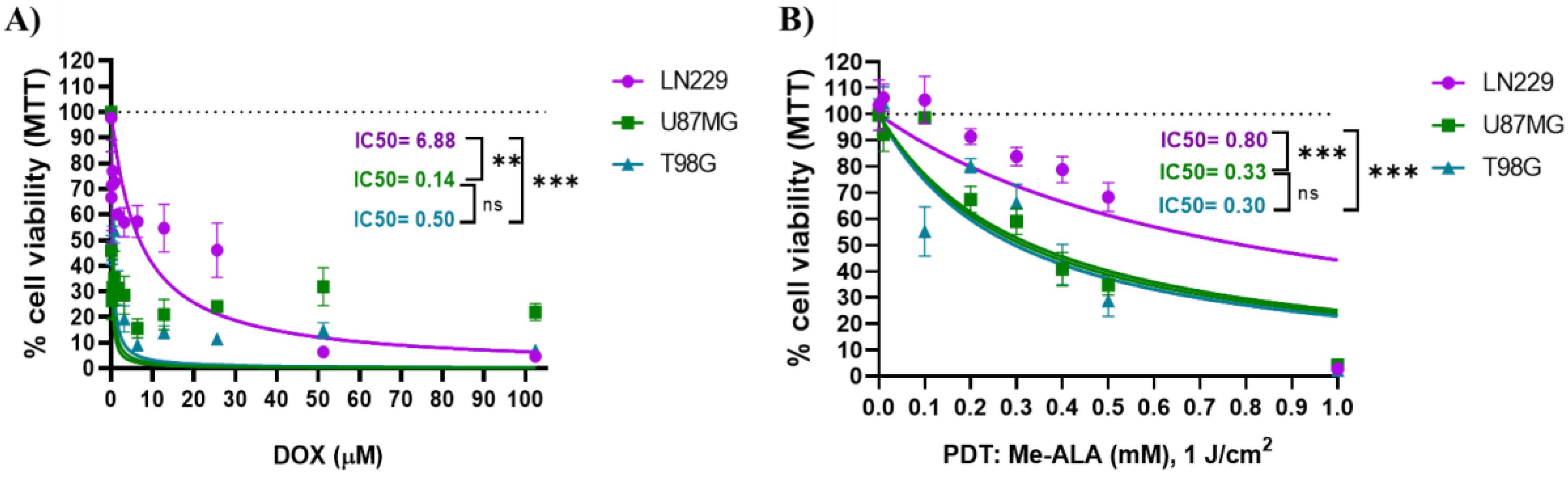
Figure 2.
DOX incorporation and PpIX bioproduction. A) DOX uptake evaluation by flow cytometry in GBM cell lines after 24 hours of incubation with different DOX concentration. B) PpIX bioproduction evaluation by flow cytometry in GBM cell lines after 4 hours of incubation time. Both DOX uptake and PpIX intracellular levels were represented by fluorescence intensity fold increment (MFI, arbitrary units A.U. compared with control cells) in the red channel of FC. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
Figure 2.
DOX incorporation and PpIX bioproduction. A) DOX uptake evaluation by flow cytometry in GBM cell lines after 24 hours of incubation with different DOX concentration. B) PpIX bioproduction evaluation by flow cytometry in GBM cell lines after 4 hours of incubation time. Both DOX uptake and PpIX intracellular levels were represented by fluorescence intensity fold increment (MFI, arbitrary units A.U. compared with control cells) in the red channel of FC. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
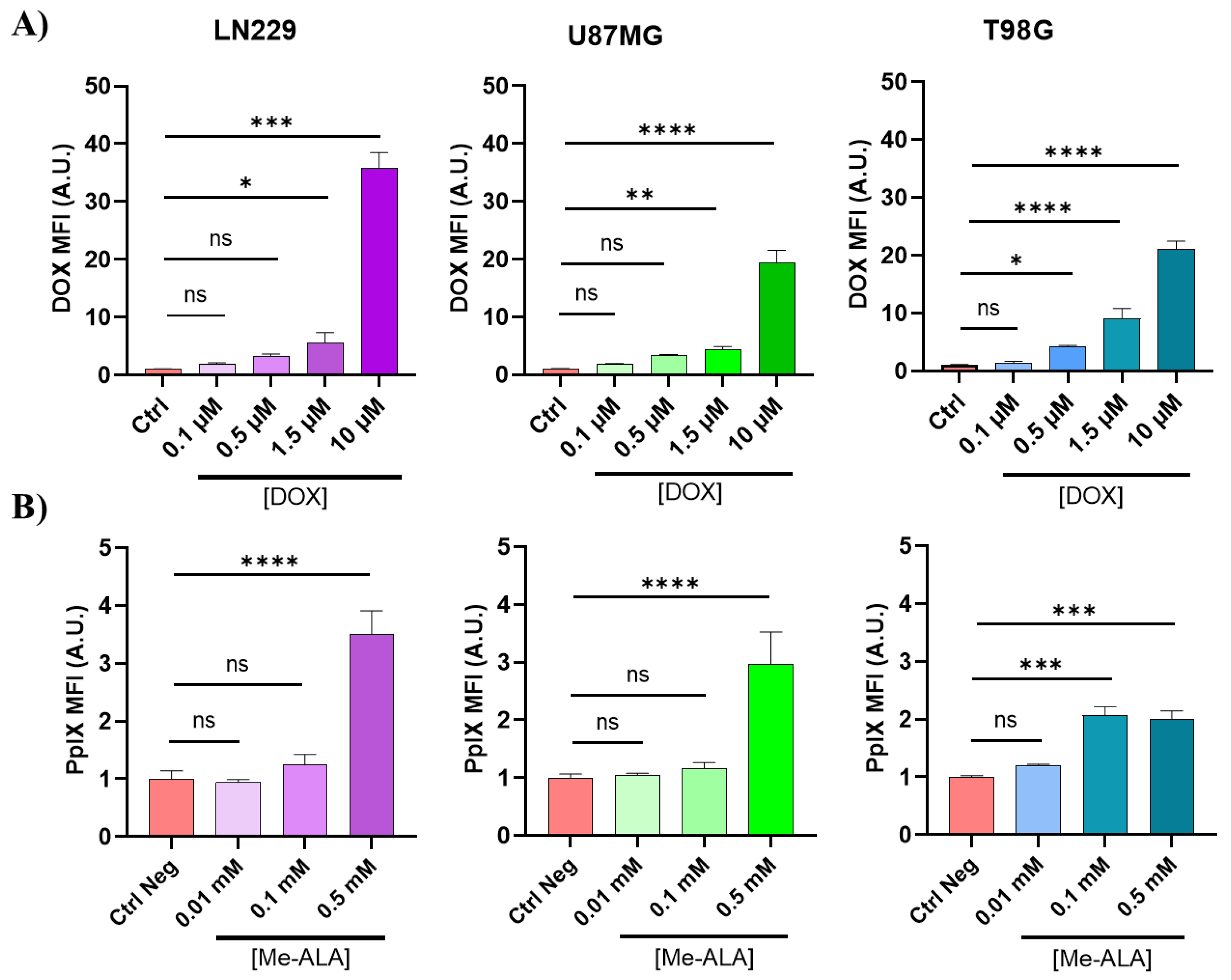
Figure 3.
ROS production evaluation after monotherapies. Geometric mean fluorescence intensity quantification relative to autofluorescence of control group for LN229, U87MG and T98G after DOX treatment (A) and PDT with Me-ALA (B). ROS levels were determined immediately after each treatment with DCFDA assay using flow cytometry. Statistical analysis was performed using two-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
Figure 3.
ROS production evaluation after monotherapies. Geometric mean fluorescence intensity quantification relative to autofluorescence of control group for LN229, U87MG and T98G after DOX treatment (A) and PDT with Me-ALA (B). ROS levels were determined immediately after each treatment with DCFDA assay using flow cytometry. Statistical analysis was performed using two-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
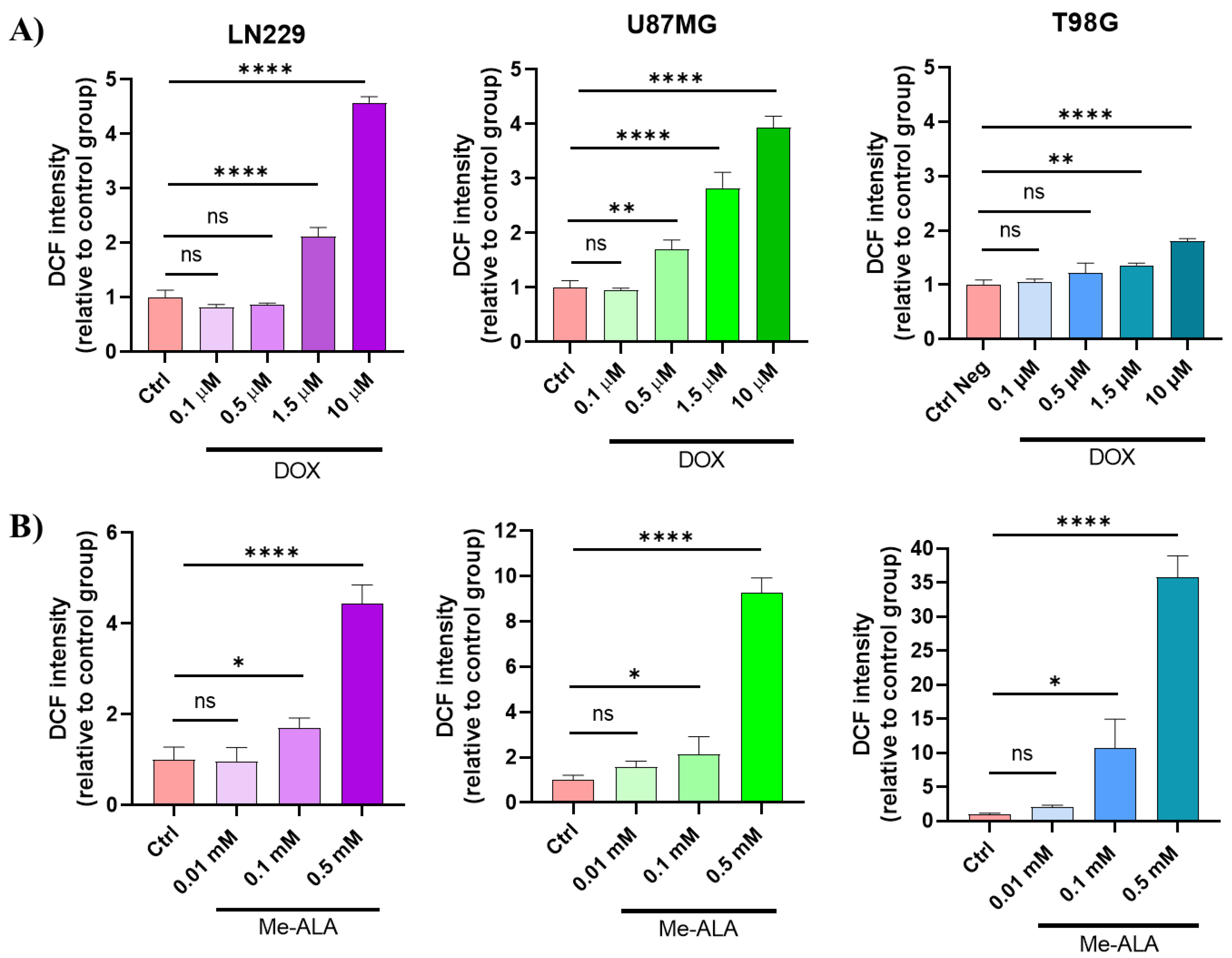
Figure 4.
Evaluation of cytoprotective effects of NAC pretreatment against prooxidant treatments. Effect of protective effect of NAC against prooxidant-induced cell death in LN229 (A), U87MG (B) and T98G (C) GBM cell lines. Cells were incubated with 5 µM NAC for 24 hours before exposure to DOX or PDT with Me-ALA. Cytotoxicity was determined with the MTT-based assay. Statistical analysis was performed using two-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
Figure 4.
Evaluation of cytoprotective effects of NAC pretreatment against prooxidant treatments. Effect of protective effect of NAC against prooxidant-induced cell death in LN229 (A), U87MG (B) and T98G (C) GBM cell lines. Cells were incubated with 5 µM NAC for 24 hours before exposure to DOX or PDT with Me-ALA. Cytotoxicity was determined with the MTT-based assay. Statistical analysis was performed using two-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001, ns: no statistically significant differences.
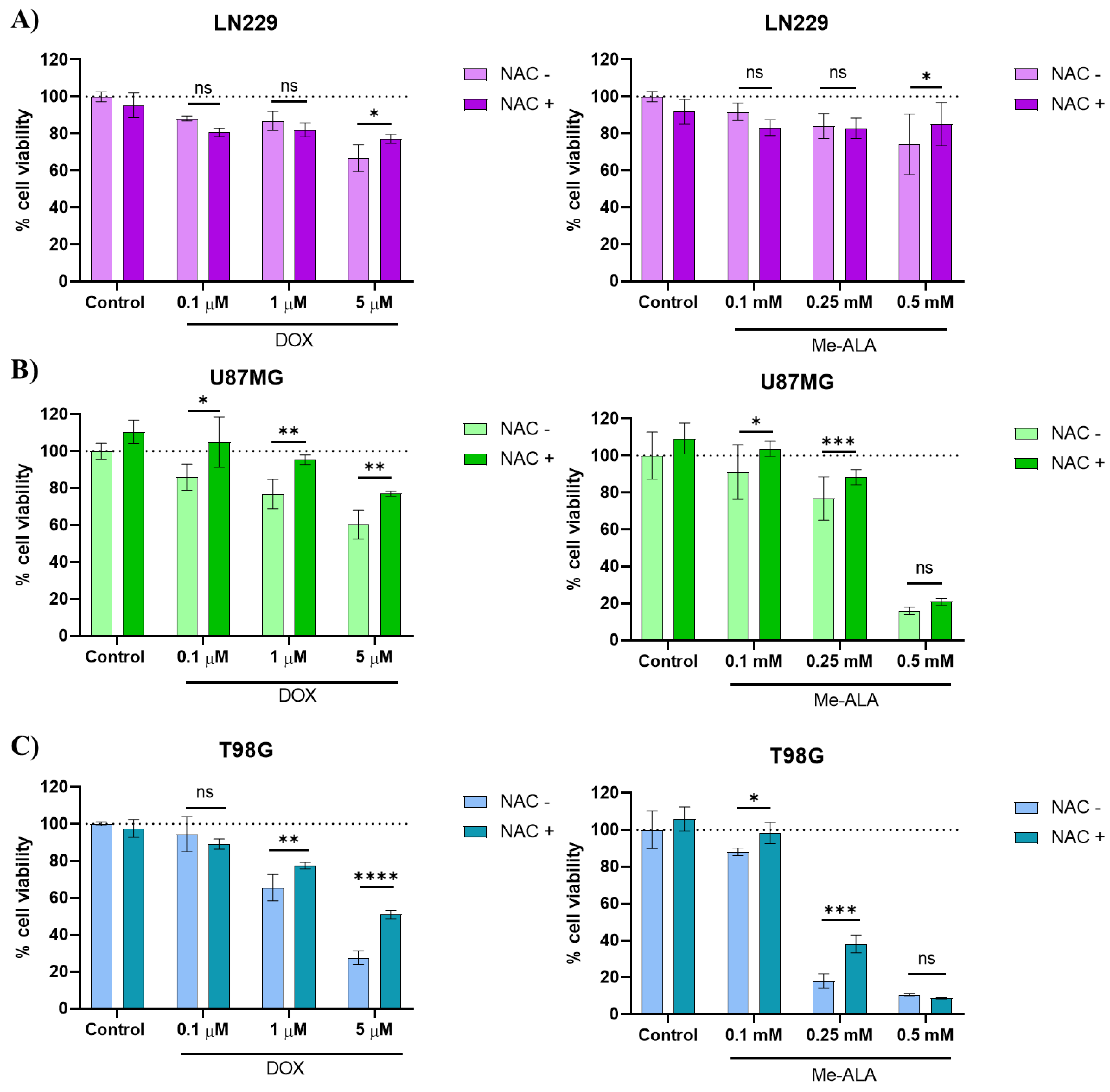
Figure 5.
Evaluation of the synergistic combination of PDT and DOX treatments. 3D images showing synergy between the combination of DOX and PDT with Me-ALA based on HSA score for different GBM cell lines; LN229 (A), U87MG (B) and T98G (C). The graph was generated using synergy finder, an online-based tool to determine synergy.
Figure 5.
Evaluation of the synergistic combination of PDT and DOX treatments. 3D images showing synergy between the combination of DOX and PDT with Me-ALA based on HSA score for different GBM cell lines; LN229 (A), U87MG (B) and T98G (C). The graph was generated using synergy finder, an online-based tool to determine synergy.
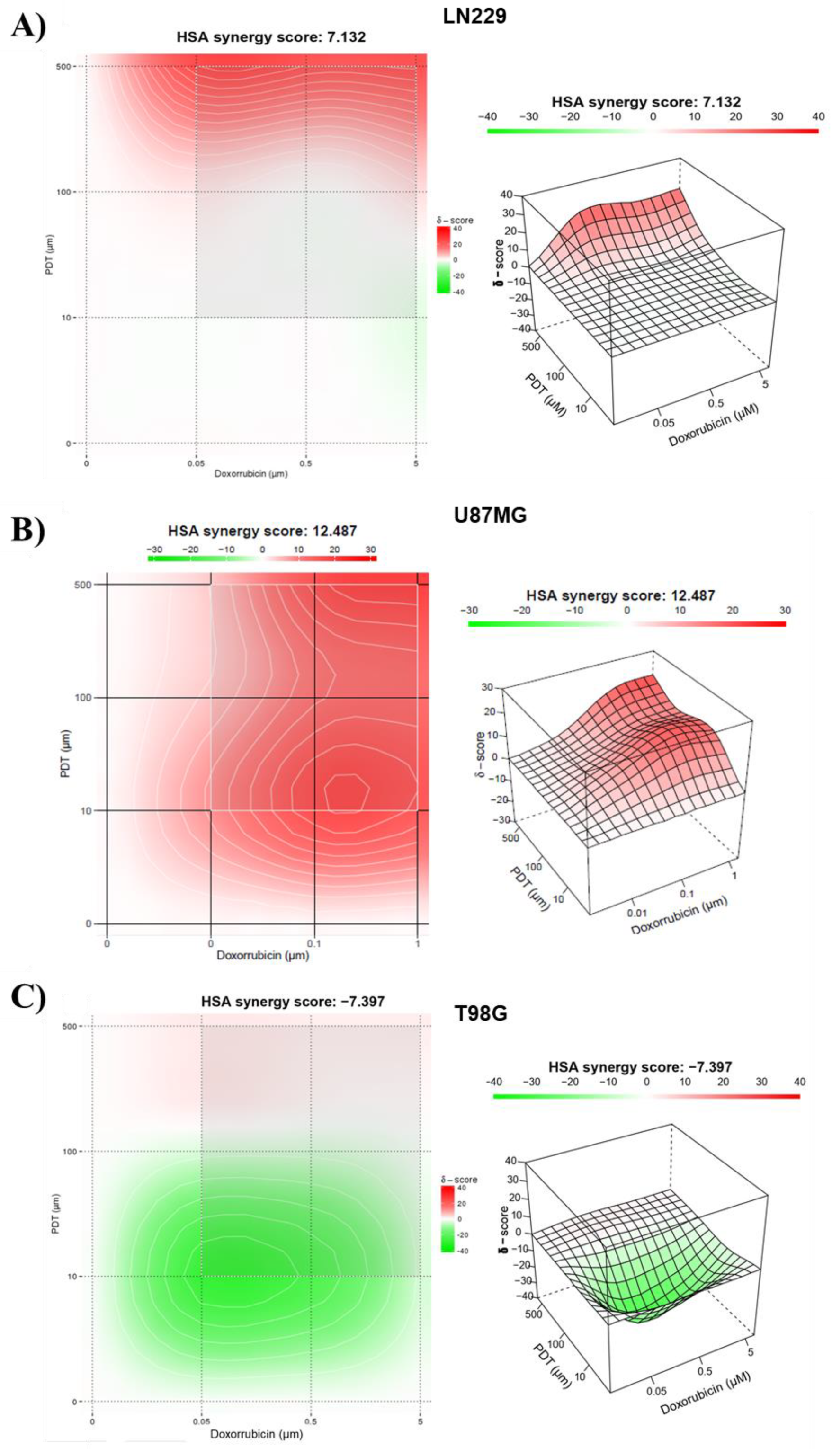
Figure 6.
ROS production evaluation after combination therapy. Geometric mean fluorescence intensity quantification relative to autofluorescence of control group for LN229, U87MG and T98G after DOX and PDT treatments. ROS levels were determined immediately after PDT treatment with DCFDA assay using flow cytometry. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, *** p < 0.001 and **** p < 0.0001.
Figure 6.
ROS production evaluation after combination therapy. Geometric mean fluorescence intensity quantification relative to autofluorescence of control group for LN229, U87MG and T98G after DOX and PDT treatments. ROS levels were determined immediately after PDT treatment with DCFDA assay using flow cytometry. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, *** p < 0.001 and **** p < 0.0001.
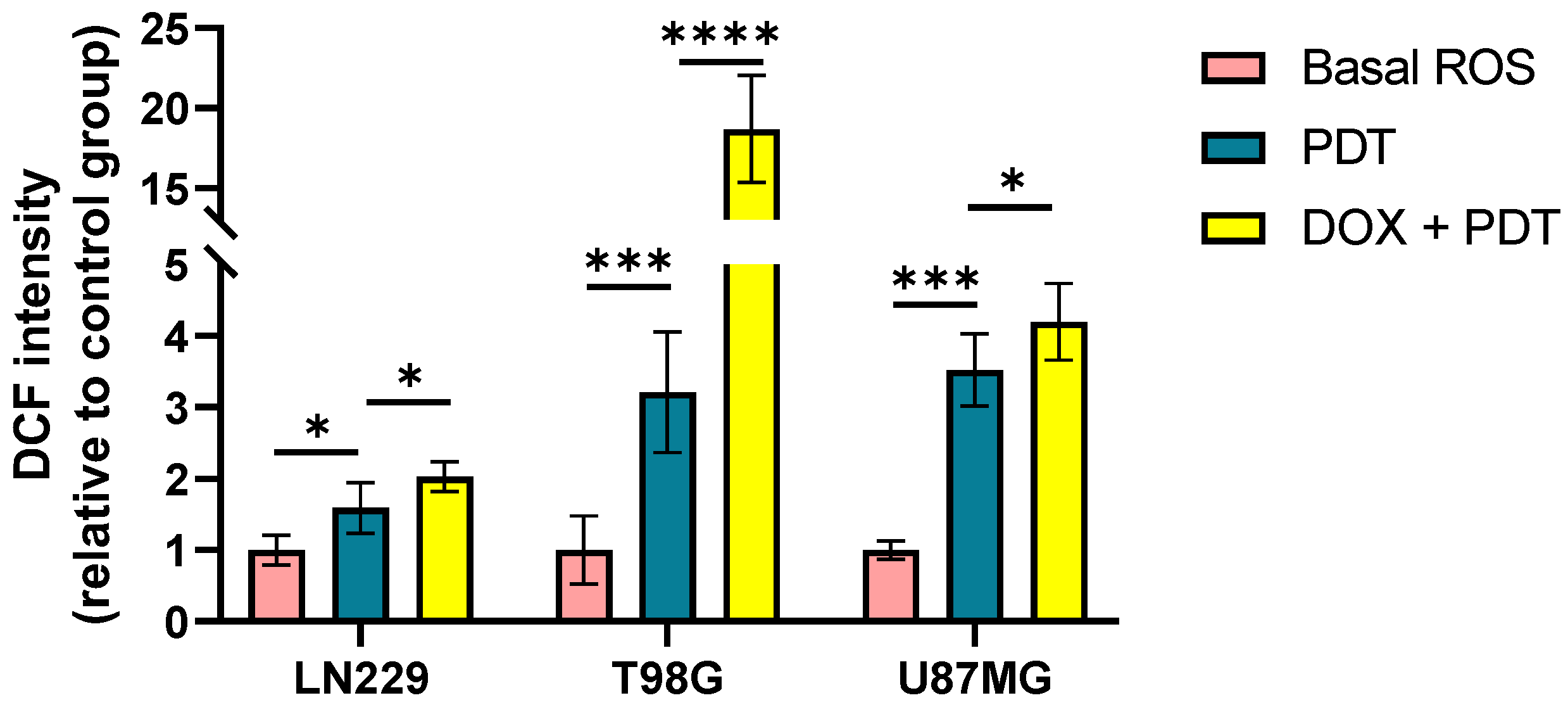
Figure 7.
Basal redox homeostasis in GBM cell lines. A) Geometric mean DCF fluorescence intensity quantification in GBM cell lines under basal growth conditions. Relative quantification of NRF2 (B), glutathione superoxide reductase (GSR) (C) and catalase (D) gene expression. Relative expressions across all cell lines were normalized by ACTB and relativized to the gene expression in U87MG. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01 and *** p < 0.001.
Figure 7.
Basal redox homeostasis in GBM cell lines. A) Geometric mean DCF fluorescence intensity quantification in GBM cell lines under basal growth conditions. Relative quantification of NRF2 (B), glutathione superoxide reductase (GSR) (C) and catalase (D) gene expression. Relative expressions across all cell lines were normalized by ACTB and relativized to the gene expression in U87MG. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks * p < 0.05, ** p < 0.01 and *** p < 0.001.
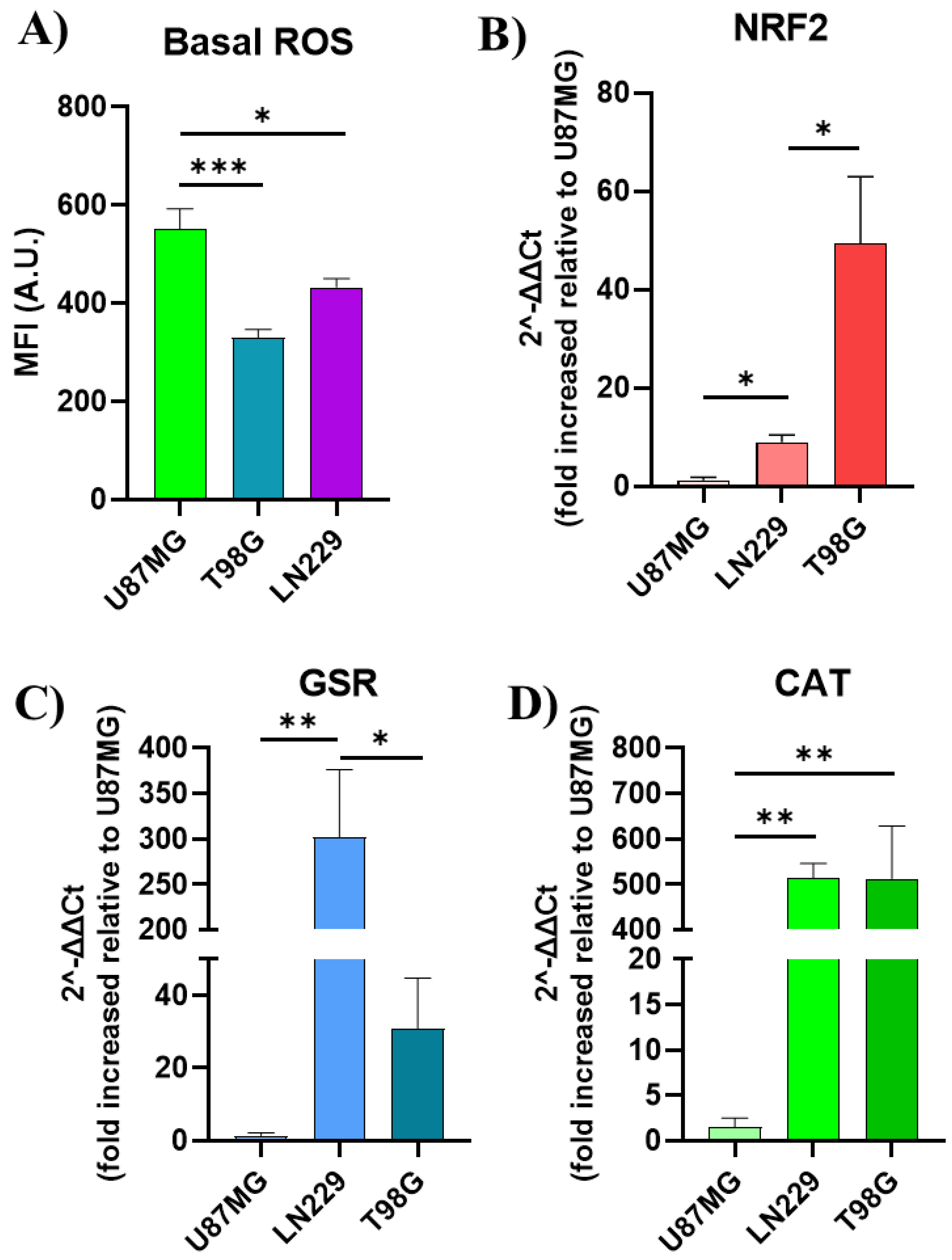
Figure 8.
In silico analysis of oxidative stress-related genes in GBM. A) Venn diagram, created using InteractiVenn, illustrates the distribution of patients across groups based on their TP53 and PTEN mutation status. The overlapping areas depict the number of patients harboring mutations in both genes or only one of them. This figure provides a visual representation of the patient stratification used in the subsequent analyses. B) Heatmap depicts the expression patterns of oxidative stress-related genes across GBM patient groups classified based on TP53 and PTEN mutation status. The analysis was performed using ClustVis. The color intensity represents the relative expression level of each gene across samples. Red indicates high expression, while blue represents low expression. C-H) Dot plots showing statistically significant differences in mRNA expression levels of oxidative stress-related genes across patient groups categorized based on TP53 and PTEN mutation status. The groups include: TP53 WT - PTEN WT (n = 94 patients): Patients with wild-type (WT) TP53 and PTEN genes; TP53 WT - PTEN mut (n = 30 patients): Patients with WT TP53 and mutated (mut) PTEN genes; TP53 mut - PTEN WT (n = 31 patients): Patients with mutated TP53 and WT PTEN genes; TP53 mut - PTEN mut (n = 20 patients): Patients with mutated TP53 and PTEN genes. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks: p < 0.001 (**) and p < 0.05 (*).
Figure 8.
In silico analysis of oxidative stress-related genes in GBM. A) Venn diagram, created using InteractiVenn, illustrates the distribution of patients across groups based on their TP53 and PTEN mutation status. The overlapping areas depict the number of patients harboring mutations in both genes or only one of them. This figure provides a visual representation of the patient stratification used in the subsequent analyses. B) Heatmap depicts the expression patterns of oxidative stress-related genes across GBM patient groups classified based on TP53 and PTEN mutation status. The analysis was performed using ClustVis. The color intensity represents the relative expression level of each gene across samples. Red indicates high expression, while blue represents low expression. C-H) Dot plots showing statistically significant differences in mRNA expression levels of oxidative stress-related genes across patient groups categorized based on TP53 and PTEN mutation status. The groups include: TP53 WT - PTEN WT (n = 94 patients): Patients with wild-type (WT) TP53 and PTEN genes; TP53 WT - PTEN mut (n = 30 patients): Patients with WT TP53 and mutated (mut) PTEN genes; TP53 mut - PTEN WT (n = 31 patients): Patients with mutated TP53 and WT PTEN genes; TP53 mut - PTEN mut (n = 20 patients): Patients with mutated TP53 and PTEN genes. Statistical analysis was performed using one-way ANOVA with Tukey's post-hoc test. Significance levels are denoted by asterisks: p < 0.001 (**) and p < 0.05 (*).
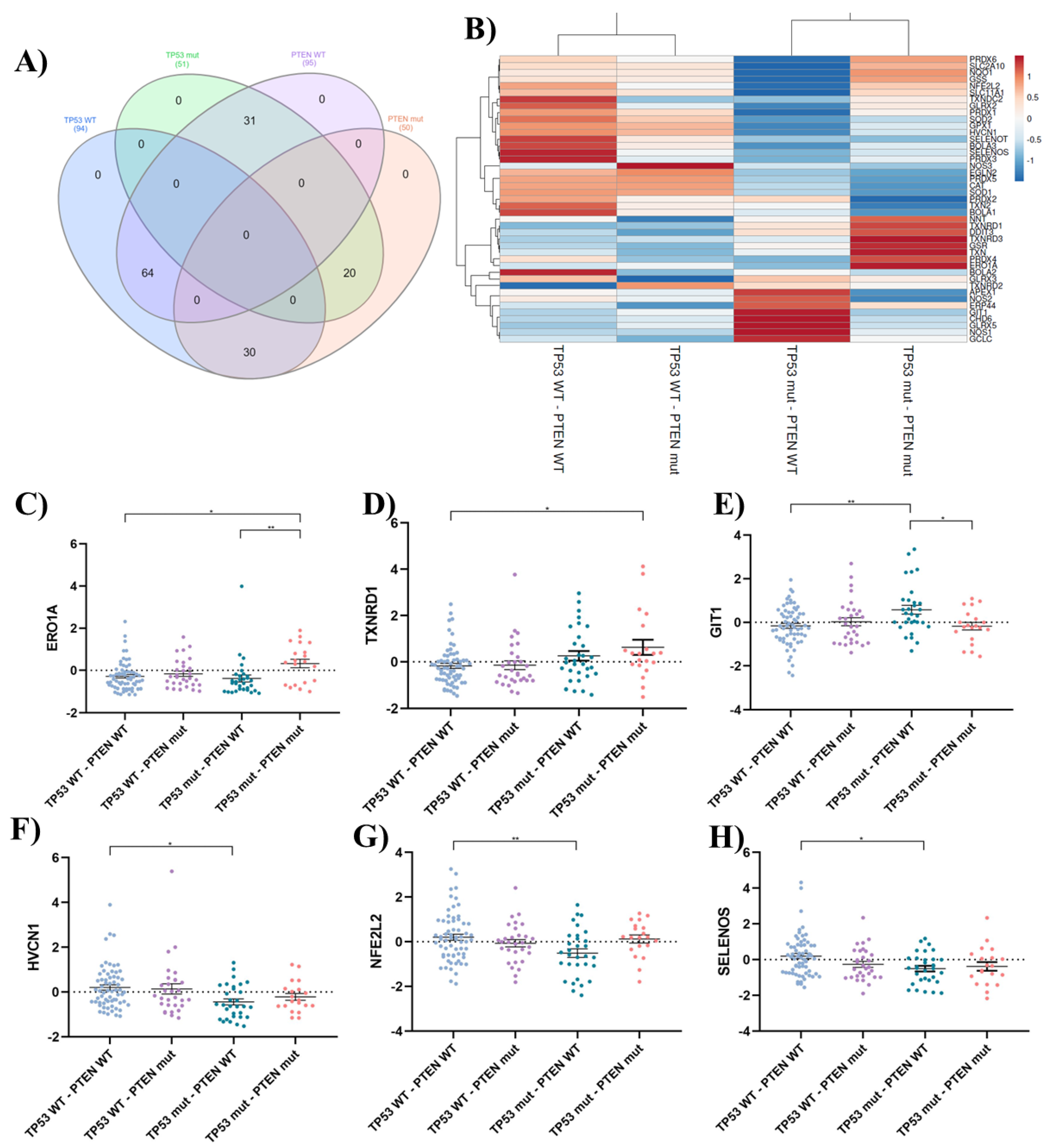

Table 1.
qPCR primers designed on Primer-BLAST and verified on BLAST-N (NCBI).
| Gene | Forward 5´- 3´ | Reverse 3´- 5´ | Product length (bp) | NM |
|---|---|---|---|---|
| ACTB | ATTGCCGACAGGATGCAGAA | GCTGATCCACATCTGCTGGAA | 150 | NM_001101.5 |
| GSR | TGGCACTTGCGTGAATGTTG | CACATAGGCATCCCGCTTTTC | 157 | NM_001195102.3 |
| NFE2L2 | TCAGCGACGGAAAGAGTATGA | CCACTGGTTTCTGACTGGATGT | 174 | NM_006164.5 |
| CAT | GGCGAGGCAGCTTGAGTTAA | CACCGCCTCGGCTTGTC | 331 | NM_001752.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



