
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
Fragile X syndrome (FXS) is caused by the full mutation in the FMR1 gene on the Xq27.3 chromosome region. It is the most common monogenic cause of Autism Spectrum Disorder (ASD) and inherited intellectual disability (ID). Besides ASD, ID and other issues, individuals with FXS may exhibit sleep problems and impairment of circadian rhythm (CR). The Drosophila melanogaster models of FXS, such as dFMR1B55, present excellent model for research in the FXS field. During this study, sleep pattern and CR in dFMR1B55 mutants were analyzed, using a new platform based on continuous high-resolution videography in integration with a highly-customized version of software. This methodology provides more sensitive results, which could be crucial for all further research in this model of fruit flies. The study revealed that dFMR1 B55 male mutants can be considered weak rhythmic flies rather than totally arrhythmic, and present a good alternative animal model of genetic disorder, which includes impairment of CR and sleep behavior. The combination of videography and software used in the current study should be recommended as a gold-standard methodology for such further research.
Keywords:
Drosophila melanogaster model of fragile X syndrome
; FMR1 gene
; Fragile X syndrome
; Circadian rhythm
; Sleep.
1. Introduction
Fragile X syndrome (FXS) is a neurodevelopmental disorder caused by the full mutation (FM) in the Fragile X Messenger Ribonucleoprotein 1 (FMR1) gene on the Xq27.3 chromosome region [1]. The FM of the FMR1 gene is caused by an expansion of the CGG trinucleotide repeats (more than 200 CGGs) in the 5’ untranslated region [2]. The FMR1 gene codes FMR1 protein (FMRP) which is an RNA binding protein important for translation of certain mRNAs involved primarily in brain development, neuronal synapse formation and synaptic plasticity [3]. FMRP is known as a translation repressor which limits various protein synthesis in the brain. FMR1 transcription is suppressed by methylation of the FMR1 gene, occurring in the FM. In the absence of FMRP, neuronal circuit formation and higher cognitive functions can be impaired [4]. FMRP has a crucial role in synaptic function due to regulation of the translation of the metabotropic Glutamate Receptor 5 (mGluR5) in astrocytes and myelin production in oligodendrocytes [5,6,7]. On the other hand, FMRP can regulate transcription and RNA synthesis by interaction with transcription factors and chromatin modifying enzymes [4]. Finally, FMRP can interact with some proteins such as ion channels and regulate their function [4]. FMRP through the interaction with the large conductance Ca2+-activated K+ (BK) channels, modulates action potential duration and consequently regulates neurotransmitter release and short-term plasticity in CA3 hippocampal pyramidal neurons [3]. All these factors together contribute to the clinical presentation of FXS.
FXS is the most common monogenic cause of Autism Spectrum Disorder (ASD) and inherited intellectual disability (ID) [8]. IQ levels in FXS patients can be correlated with gender, methylation status and FMRP abundance [9,10]. Approximately 85% of males and 25-30% of females with FXS have ID [6]. On the other side, more than half (50–60%) of males and 20% of females with FXS are diagnosed with ASD (reviewed in: Protic et al., 2022 [11]). Generally, females with FXS have less severe symptomatology due to compensatory activation of the unaffected X [11,12].
Besides ASD, patients with FXS exhibit ID, sleep problems, changes in circadian rhythm (CR), delay in the development of motor functions, of speech and of language in early childhood, and often attention deficit hyperactivity disorder (ADHD) [13]. FXS can be also characterized by other comorbidities such as neurological seizures, endocrinological problems like obesity, cardiac abnormalities (mitral valve prolapse), and macroorchidism in puberty [14,15]. Recurrent otitis media infections in childhood and strabismus are also present in patients with FXS. Physical appearance can also be characterized by an elongated face, prominent ears, mandibular prognathism, and joint hypermobility [14]. As mentioned above, sleep problems are often present in individuals with FXS and manifest mostly as hardly falling asleep, impaired sleep quality and waking up many times during the night. Sleep problems tend to affect quality of patient’s life and have a negative impact on the whole family [11,16].
FXS preclinical research on animals is limited to a few models, such as the Fmr1 knock-out (KO) mice, the Fmr1 KO zebrafish and the Drosophila melanogaster model of FXS (Fmr1, FBgn0028734 [17]; herein named dFMR1) such as dFMR1B55 mutants [18,19,20]. The dFMR1B55 allele is one example of the Drosophila melanogaster model of FXS. In dFMR1B55, the 2.5 kb deletion including exons 2, 3, and 4 of the dFMR1 gene was generated by an imprecise excision of the EP(3)3422 P-element [21]. The Drosophila melanogaster models of FXS, such as dFMR1B55, show impairment in CR, climbing abilities, social interaction, olfactory learning and memory etc. [22,23]. Tracking of locomotor activity enables sleep and CR analysis in the Drosophila melanogaster models of FXS. Until now, CRs have been studied in Drosophila mostly with infrared-based beam-crossing methods [21,23,24,25,26,27]. These methods may lead to under-estimation of the total activity of flies throughout the day, which may, in turn, affect estimation of critical CR and sleep statistics [28,29].
The aim of this study is to analyze sleep pattern and CR in dFMR1B55 mutants as an animal model for further research in the field of FXS, using a new platform based on continuous high resolution video monitoring in integration with customized version of the open-source app VANESSA.
2. Results
2.1. CR Rhythm Analysis in the w1118, per01 and dFMR1B55 Groups
The CR of white1118 (w1118), period01 (per01) and dFMR1B55 flies was analyzed using video monitoring and VANESSA application.
A graphical representation of the activity patterns (CRs) of selected flies of the white1118 (w1118), period01 (per01) and dFMR1B55 groups in the dark-dark (DD) period (from 3rd to 10th experimental day) and their synchronization with environmental cues (light) is presented in actograms within Figure 1.
In the control w1118 groups (N = 50 for males and N = 50 for females), the rhythmicity was observed in 84% (42/50) of females and in 96% (48/50) of male flies. Comparison between sexes showed a significantly higher frequency of rhythmic flies in males (p = 0.04). The mean period for rhythmic individuals was 23.70 ± 0.17 h for females (N = 42) and 23.78 ± 0.27 h for males (N = 48) and there was no statistically significant difference in period values (p = 0.14) between sexes. In addition, w1118 rhythmic females had significantly higher (p < 0.0001, both) median and mean values of rhythm power (median = 263.90, with range: 68.85 - 423.60; mean ± SD: 252.60 ± 100.90) in comparison with w1118 rhythmic males (median = 110.90, with range: 21.17 - 385.00, mean ± SD: 125.60 ± 72.49).
In the per01 groups, the rhythmicity was observed in 62% (31/50) of females and in 62% (31/50) of male flies. Comparison between sexes showed the same frequency of rhythmic flies (p > 0.99). The mean period for rhythmic individuals was 23.60 ± 0.31 h for females (N = 31) and 23.86 ± 0.27 h for males (N = 31). Furthermore, for rhythmic females, the median value of the power of the rhythm was 145.00 (range: 17.99 - 395.50) and the mean value was 157.30 ± 102.50. In addition, the median and mean values for rhythmic males were 68.49 (range: 15.10 - 373.80 and 109.60 ± 102.70, respectively. Interestingly, male per01 rhythmic individuals had a significantly longer period (p = 0.0002), while per01 rhythmic females had a significantly higher power of the rhythm than the rhythmic per01 males (p = 0.0180).
Finally, in the experimental dFMR1B55 groups, rhythmicity was observed in 98% (49/50) of females and in 74% (37/50) of male flies indicating a significantly lower frequency of rhythmic flies in males (p = 0.0005). The mean period for rhythmic individuals was 23.66 ± 0.36 h for females (N = 49) and 23.69 ± 0.49 h for males (N = 37) without statistically significant difference in period values (p = 0.51) between sexes. The median values of the power of the rhythm were 128.90 (range: 14.18 - 314.80) for rhythmic females and 82.65 (range: 15.12 - 292.20) for rhythmic males; while the mean values of the same parameter were 135.60 ± 68.81 for females and 90.18 ± 65.16 for males. Like w118 and per01 females, dFMR1B55 rhythmic females had a significantly higher power of rhythm than the rhythmic dFMR1B55 males (p = 0.001).
The described values of periods and power of the CRs, in individual rhythmic and arhythmic w1118, per01, and dFMR1B55 males, are visualized within selected chi-square periodograms in Figure 2.
Comparisons of examined variables related to CR (frequency of rhythmicity, period and power of rhythm) among included groups of flies revealed that the lowest frequencies of rhythmic flies of both sexes were noticed in per01 (61% in both, males and females). However, a significantly lower frequency of rhythmicity was shown for dFMR1B55 males compared to w1118 males (p = 0.002). Interestingly, dFMR1B55 females had a higher frequency of rhythmicity compared to w1118 of the same sex (p = 0.01). On the other hand, there were not statistically significant differences between dFMR1B55 and per01 males (p = 0.20) in term of frequency of rhythmic flies, while for females, dFMR1B55 showed a higher frequency of rhythmicity compared to per01 (p < 0.0001). Finally, for both sexes, w1118 had a higher frequency of rhythmicity compared to per01 (females: p = 0.01; males: p < 0.0001).
Periods for rhythmic individuals were not statistically different for w1118 vs per01 vs dFMR1B55 (females: p = 0.25; males: p = 0.10). Multiple comparisons among included groups divided by sex did not show statistically significant differences in period values (p > 0.05, all). Violin plots of periods, presented in Figure 3, show that there were approximately the same period values in rhythmic males of all examined groups (w1118, per01 and dFMR1B55).
The power of the rhythm comparison among w1118, per01 and dFMR1B55 revealed statistically significant differences among them (females: p < 0.0001; males: p = 0.02). Specifically, for both sexes, dFMR1B55 had significantly lower power than w1118 (females: p < 0.0001; males: p = 0.03). There were no significant differences between dFMR1B55 and per01 in power values (both sexes: p > 0.9999).
Bar plots of frequency of rhythmicity and power of rhythm in w1118, per01 and dFMR1B55 are presented in Figure 4.
2.2. Sleep Analysis in the w1118, per01 and dFMR1B55 Groups
The sleep patterns of white1118 (w1118), period01 (per01) and dFMR1B55 flies was analyzed using video monitoring and VANESSA application.
In the control w1118 groups, the total sleeping time during DD (3rd to 10th day) was 66.95 ± 50.12 h (median: 61.03 h with range: 3.52 - 172.30 h) for females and 104.70 ± 53.05 h (median: 100.40 h with range: 17.27 - 228.30 h) for males. Females had a total of 177.30 ± 106.60 sleeping episodes (median: 174.50 with a range 17 - 443) during the whole DD period, while males had 272.60 ± 104.90 (median: 267.50 with a range 63.00 - 493.00). Comparison between sexes showed that w1118 males had a significantly longer total sleeping time in DD (p = 0.0005) with a larger number of sleeping episodes compared to w1118 females (p < 0.0001).
In the per01 groups, the total sleeping time during DD (3rd to 10th day) was 81.48 ± 48.48 h (median: 72.90 h with range: 12.42 - 244.50 h) for females and 123.30 ± 53.25 h (median: 124.60 h with range: 19.47 - 285.70 h) for males. Females had a total of 132.20 ± 74.96 sleeping episodes (median: 106.50 with range 31.00 - 289.00) during whole DD while males had 191.90 ± 68.29 (median: 182.00 with range 39.00 - 344.00). Comparison between sexes showed that per01 males had a significantly longer total sleeping time in DD (p < 0.0001) with a larger number of sleeping episodes (p < 0.0001).
Finally, in the experimental dFMR1B55 groups, the total sleeping time during DD (3rd to 10th day) was 142.70 ± 67.53 h (median: 146.50 h with range: 16.32 - 279.50 h) for females and 212.50 ± 85.32 h (median: 243.50 h with range: 40.38 - 318.80 h) for males. Females had a total of 218.50 ± 84.95 sleeping episodes (median: 215.50 with range 56.00 - 402.00) during whole DD, while males had 304.90 ± 83.44 (median: 314.00 with range 130.00 - 496.00). Comparison between sexes showed that dFMR1B55 males had a significantly longer total sleeping time in DD (p < 0.0001) with a larger number of sleeping episodes (p < 0.0001).
Comparison of examined variables related to sleep (total sleeping time during DD and number of sleeping episodes during DD) revealed statistically significant differences between those three groups, both in males and females (p < 0.0001, all). Longest total sleeping time during DD is noticed in dFMR1B55 in both sexes and average values and medians were twice larger than in w1118 and per01 flies (p < 0.0001, all). On the other hand, there were not statistically significant differences between w1118 and per01 in the total sleeping time during DD (females: p = 0.63; males: p = 0.40). Compared to per01, both dFMR1B55 males and females showed a statistically significant larger number of sleeping episodes in DD (p < 0.0001).
3. Discussion
The current study is the first study that used high-resolution continuous video monitoring for a detailed analysis of circadian rhythm and sleep patterns in Drosophila Fmr1 mutants based on their locomotor activity. This study revealed a lower percentage of rhythmic males in the dFMR1B55 group in comparison to the control wild-type male flies. This observation may be based on the duration of their sleep, which revealed that dFMR1B55 generally sleeps more compared to the w1118 and per01 phenotypes. Although there is a high frequency of rhythmic flies in the dFMR1B55 group in both sexes, it is important to emphasize that they mostly had a significantly lower power of rhythm than w1118. On the other hand, their power of rhythm is similar to rhythmic per01 flies. All investigated groups of flies had a similar period of CR, which was around 24 hours.
CR changes through the measurement of locomotor activity have been previously tested in a few Drosophila FXS models. All these studies showed dFMR1 mutants as heterogeneous populations consisting of both rhythmic and arrhythmic individuals [21,23,25,26]. In accordance with our study, previous studies that investigated CR in dFMR1B55 mutants also found differences in frequency of rhythmicity between dFMR1B55 and wild-type of flies. All previously described types [21,26]. However, their results were independent of sexes, while groups of flies were divided by sexes in the current study. Although the frequency of rhythmicity of the dFMR1B55 mutants found in previous studies [21,26] was much lower than in the current study, these results cannot be compared due to different experimental designs. Specifically, in the current study, the experimental design was based on video tracking of flies in subgroups divided by sex. However, similarly to our results, Inoue et al. (2002) calculated the period for the dFMR1B55 mutants as 23.80 ± 0.50 h [21].
Per is a key gene in CR regulation [30] and per01 was included as a control group in this study based on results in previously published articles that reported the majority of dFMR1B55 males as arrhythmic [21]. Interestingly, according to the results obtained in the current study using high-resolution videography and VANESSA software, the majority of these flies were rhythmic with a low power of rhythm. Clock per01 mutants have a non-functional PER protein and cannot sustain rhythmicity properly [28]. However, there are studies where weak CR components were detected in per01 [31,32]. Klarsfeld et al. (2003) classified more than a third of per01 individuals as rhythmic [28], with weak power of rhythm, which is in concordance with the results obtained in the present study. In the current study, the frequency and power of dFMR1B55 males` rhythmicity did not differ from per01 males, suggesting that regarding CR, dFMR1B55 males are more similar to the per01 line, where weak rhythm was detected, and completely different than w1118 same sex. Thus, the main research question is could scientists consider that dFMR1B55 males are rhythmic with weak power of rhythmicity rather than they are being completely arhythmic? Based on our findings, it is evident that most dFMR1B55 males exhibit weak rhythmicity compared to the control group (w1118). This distinction in rhythmicity is supported by the methodology employed in our research. However, the same conclusion cannot be drawn for dFMR1B55 females, as our results indicate that almost all of them displayed rhythmicity. Also, w1118 females could not be a proper control group for comparison since, unexpectedly, the frequency of rhythmicity is lower in w1118 females than in the dFMR1B55 of the same sex.
Other studies investigating CR rhythm in the Drosophila FXS model used different strains (dFXRΔ113 and dFMR3), and also showed differences in frequency of rhythmicity between FXS flies models and wild-type flies [23,25]. The power of rhythm as a CR parameter of dFMR3 was only reported in the study conducted by Dockendorff et al (2002) using Fast Fourier Transform (FFT) analysis. Similar to our results, they showed much lower power of rhythmicity in dFMR1 male mutants compared to wild-type [23]. In addition, previous sleep study showed longer total sleep duration and a higher number of sleeping episodes in the Drosophila FXS model compared to the wild-type, which is consistent with our results [24]. In all these studies, no measures were made separately on females and males as done in the present study.
All previously described CR studies of Drosophila FXS models used the infrared-based beam crossing method by Drosophila Activity Monitoring (DAM) system [21,23,24,25,26]. The previously used data analysis software was different in different studies. High-resolution continuous video monitoring with the Zantiks MWP Unit used in the current study is different from the most widely used infrared-based beam crossing method. While the infrared-based method considers the number of crossing beams events during time and is limited to movements near beams, the results of videography are presented as crossed distances. Micromovements of Drosophila happening anywhere in the well can also be detected using this tracking system. Some scientists reported their concerns about the under- or over-estimation of infrared detection of some fly movements. These limitations are linked to fly positions in the system [33]. In the current study, frequent short grooming bouts followed by short, crossed distances are being noticed in dFMR1B55 mutants. In other words, the video monitoring used in the current study can detect those micromovements anywhere in the system. These micromovements could be probably the main reason of higher frequency of rhythmic flies in per01 groups and general less total sleep values. Observed difference in methodology between DAN system and video-based monitoring may be also the main reason for variable results on CR in dFMR1 mutants. Video monitoring as a more sensitive method could be useful especially in mutants with reduced overall motility [34]. Bolduc et al. (2010) reported lower locomotor activity in dFMR1B55 compared to w1118 [22], which is similar to the current study and could be related to significantly higher total sleeping in dFMR1B55 mutants. In addition, video recording always shows less sleep than DAM systems [35]. Micromovements less than 3 mm, i. e. activity records less than 1 body length of flies, will be considered for exclusion in the next iteration of our work. However, based on current and previous results, video monitoring used in our study could present a recommended methodology for sleep research and CR analyses in dFMR1 mutants.
Zantiks MWP Unit is a fully automated system with no need for additional interventions during the whole experimental period [36]. The system includes software, is controlled via a web browser and has full customer support. On the other hand, challenges are associated with software for CR and sleep analyses because almost all available software is adapted to the DAM system. However, VANESSA is an open-source, user-friendly application for CR and sleep data acquisition, analysis, and visualization [37,38], and it was adapted for analyses of fruit fly video tracking recordings in our research. The VANESSA apps have Graphical User Interfaces (GUIs), are also hosted on a server and can be directly used from a browser (available on https://cryptodice.shinyapps.io/vanessa-dam-cra/ and https://cryptodice.shinyapps.io/vanessadam-sa/) [38]. Neither Zantiks MWP Unit nor VANESSA apps require high additional engineering or programming knowledge, which is an important advantage for research laboratories. Finally, the combination of Zantiks MWP and VANESSA presents modern, useful technology that provides a more sensitive methodology in the field of chronobiology.
4. Materials and Methods
4.1. Flies
The dFMR1B55 mutants, were created by Inoue et al in 2002 through an imprecise excision of the EP(3)3422 P-element. This process resulted in a deletion of the dFMR1 genomic DNA, which included exons 2, 3, and 4, thereby creating a protein null allele B55 [21]. The control groups for this study consisted of the wild-type w1118 and period01 per01, period mutants.
Flies were reared under a 12 h light/12 h dark cycle on standard cornmeal/molasses/agar medium at 25°C and 60% relative humidity [39]. Virgin flies were collected 8 hours after eclosion. Each male was kept separately in its own vial, while females were kept in groups of five on standard cornmeal/molasses/agar medium. After flies reached the age of 3-4 days, video monitoring of locomotor activity started at 7 a.m.
All analyzed groups of flies were divided based on phenotype and sex, and each group finally contained 50 flies in total.
4.2. Video Monitoring of Locomotor Activity
Video monitoring and data collection of locomotor activity was performed in the Zantiks MWP Unit (Zantiks Ltd, Cambridge UK). Demo scripts for Zantiks MWP Unit are available on Zanscript - Script library Zantiks. A total of 96 flies (virgin males and females) were individually housed in each well of a 96-well-plate. Each well was partly filled with a transparent sugar/agar medium. Animals were video tracked under standard conditions of 25°C and 55-65% humidity. Locomotor activity rhythms were measured as distance traveled (mm) every 30 seconds under light-dark cycles of 12 hours (LD12:12) for 2 days followed by constant darkness (dark-dark, DD) for 8 days.
4.3. CR and Sleep Pattern Analyses
Analyses of parameters was done by VANESSA, an open-source R-based set of applications for CR and sleep data acquisition, analysis, and visualization (available on https://github.com/orijitghosh/VANESSADAM). Free running periods and powers of the CR were estimated by the chi-square periodograms in the highly customized version of VANESSA made to analyze data from Zantiks MWP. The fly was considered rhythmic if the peak of the periodogram appeared above the alpha = 0.05 confidence level. Males and females were analyzed separately.
4.4. Statistical Analyses
Statistical analyses were performed in GraphPad Prism 8. Parametric and non-parametric statistical tests were used to compare the period and power between groups. Significance was defined as p ≤ 0.05.
5. Conclusions
Circadian rhythm and sleep analyses by new tools consisting of high-resolution videography and a highly customized version of software provide more sensitive results in the analysis of dFMR1 mutants, which could be crucial for all further research in this model of fruit flies. Specifically, this study revealed that dFMR1 male mutants can be considered weak rhythmic flies rather than totally arrhythmic and present a good experimental model for further behavioral research in the field of FXS. Finally, dFMR1B55 male mutants could be an important tool for pharmacological initial drug screening as an example of an alternative animal model of genetic disorder, which includes impairment of CR and sleep behavior. The combination of videography and software used in the current study should be recommended as a gold-standard methodology for such further research.
Author Contributions
Conceptualization, D.P.; methodology, S.M., A.G. and M.S.; software, S.M. and A.G.; validation, D.P, and S.M.; formal analysis, S.M.; investigation, S.M, V.M. and M.S.; resources, D.P; data curation, S.M., V.M. and M.S.; writing—original draft preparation, S.M. and V.M.; writing—review and editing, D.P, M.C, and D.B.; visualization, S.M, and D.P.; supervision, D.P and D.B.; project administration, D.P.; funding acquisition, D.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Science Fund of the Republic of Serbia, Program IDEA, GRANT No. 7673781. The authors of the publication are responsible for the content of this publication and this content does not express the attitudes of the Science Fund of the Republic of Serbia. Project No. 451-03-66/2024-03/200110 financially supported by The Ministry of Science, Technological Development and Innovation of the Republic of Serbia.
Institutional Review Board Statement
Ethical review and approval were waived for this study. The study includes only alternative models, i. e., Drosophila Fragile X model. According to the National Centre for the Replacement Refinement & Reduction of Animal in Research, partial replacement includes the use of some animals that, based on current scientific thinking, are not considered capable of experiencing suffering. This includes invertebrates such as Drosophila, nematode, worms, social amoebae, and immature forms of vertebrates (https://www.nc3rs.org.uk/the-3rs). According to Directive 2010/63/EU of the European Parliament and of the Council of 22 September 2010 on the protection of animals used for scientific purposes (https://eur-lex.europa.eu/eli/dir/2010/63/oj) and Serbia Law on animals’ welfare (Sl. list RS 41/09 at: https://www.paragraf.rs/propisi/zakon_o_dobrobiti_zivotinja.html), ethical review permissions is not needed for scientific research which include alternative models such as Drosophila.
Informed Consent Statement
Not applicable
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors on request.
Acknowledgments
Acknowledgements to the whole Zantiks Ltd. team for providing ready support with the equipment whenever needed. We would like to express our gratitude to Prof. Dr. Mark Wu, who is affiliated with the Department of Neurology and the Department of Neuroscience at Johns Hopkins University School of Medicine (Baltimore, MD, USA) for all his consultations, suggestions, and support during this study.
Conflicts of Interest
Over the past 24 months, Dejan B. Budimirovic, MD, has received funding from Zynerba Pharmaceuticals (ZYN2-CL-017, ZYN2-CL-033) and Tetra Discovery Partners, Inc. (BPN14770-CNS-204, BPN14770-CNS-301, BPN14770-CNS-302) as an investigator on the listed five clinical trials in individuals with fragile X syndrome (FXS). He has also received funding from CDC (RFA-DD-21-002). All the above funding has been directed to the Kennedy Krieger Institute/the Johns Hopkins Medical Institutions, meaning that Dr. Budimirovic receives no personal funds, and the Institute has no relevant financial interest in any of the commercial entities listed. All other authors declare no conflicts of interest.
References
- Verkerk, A.J.M.H.; Pieretti, M.; Sutcliffe, J.S.; Fu, Y.-H.; Kuhl, D.P.A.; Pizzuti, A.; Reiner, O.; Richards, S.; Victoria, M.F.; Zhang, F.; et al. Identification of a Gene (FMR-1) Containing a CGG Repeat Coincident with a Breakpoint Cluster Region Exhibiting Length Variation in Fragile X Syndrome. Cell 1991, 65, 905–914. [Google Scholar] [CrossRef] [PubMed]
- Feng, G.; Deák, P.; Chopra, M.; Hall, L.M. Cloning and Functional Analysis of TipE, a Novel Membrane Protein That Enhances Drosophila Para Sodium Channel Function. Cell 1995, 82, 1001–1011. [Google Scholar] [CrossRef] [PubMed]
- Weiler, I.J.; Irwin, S.A.; Klintsova, A.Y.; Spencer, C.M.; Brazelton, A.D.; Miyashiro, K.; Comery, T.A.; Patel, B.; Eberwine, J.; Greenough, W.T. Fragile X Mental Retardation Protein Is Translated near Synapses in Response to Neurotransmitter Activation. Proc. Natl. Acad. Sci. U.S.A. 1997, 94, 5395–5400. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.D.; Zhao, X. The Molecular Biology of FMRP: New Insights into Fragile X Syndrome. Nat Rev Neurosci 2021, 22, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Higashimori, H.; Morel, L.; Huth, J.; Lindemann, L.; Dulla, C.; Taylor, A.; Freeman, M.; Yang, Y. Astroglial FMRP-Dependent Translational down-Regulation of mGluR5 Underlies Glutamate Transporter GLT1 Dysregulation in the Fragile X Mouse. Human Molecular Genetics 2013, 22, 2041–2054. [Google Scholar] [CrossRef] [PubMed]
- Salcedo-Arellano, Ma.J.; Hagerman, R.J.; Martínez-Cerdeño, V. Fragile X syndrome: clinical presentation, pathology and treatment. GMM 2023, 156, 3599. [Google Scholar] [CrossRef]
- Wang, H. Developmentally-Programmed FMRP Expression in Oligodendrocytes: A Potential Role of FMRP in Regulating Translation in Oligodendroglia Progenitors. Human Molecular Genetics 2003, 13, 79–89. [Google Scholar] [CrossRef] [PubMed]
- Devitt, N.; Gallagher, L.; Reilly, R. Autism Spectrum Disorder (ASD) and Fragile X Syndrome (FXS): Two Overlapping Disorders Reviewed through Electroencephalography—What Can Be Interpreted from the Available Information? Brain Sciences 2015, 5, 92–117. [Google Scholar] [CrossRef] [PubMed]
- Huddleston, L.B.; Visootsak, J.; Sherman, S.L. Cognitive Aspects of Fragile X Syndrome. WIRES Cognitive Science 2014, 5, 501–508. [Google Scholar] [CrossRef]
- Tassone, F.; Hagerman, R.J.; Iklé, D.N.; Dyer, P.N.; Lampe, M.; Willemsen, R.; Oostra, B.A.; Taylor, A.K. FMRP Expression as a Potential Prognostic Indicator in Fragile X Syndrome. Am J Med Genet 1999, 84, 250–261. [Google Scholar] [CrossRef]
- Protic, D.D.; Aishworiya, R.; Salcedo-Arellano, M.J.; Tang, S.J.; Milisavljevic, J.; Mitrovic, F.; Hagerman, R.J.; Budimirovic, D.B. Fragile X Syndrome: From Molecular Aspect to Clinical Treatment. IJMS 2022, 23, 1935. [Google Scholar] [CrossRef] [PubMed]
- Thurman, A.J.; McDuffie, A.; Hagerman, R.; Abbeduto, L. Psychiatric Symptoms in Boys with Fragile X Syndrome: A Comparison with Nonsyndromic Autism Spectrum Disorder. Research in Developmental Disabilities 2014, 35, 1072–1086. [Google Scholar] [CrossRef] [PubMed]
- Fragile X Syndrome and Premutation Disorders: New Developments and Treatments; Hagerman, R.J., Hagerman, P.J., Eds.; Mac Keith Press: London, 2020; ISBN 978-1-911612-38-4. [Google Scholar]
- Saldarriaga, W.; Tassone, F.; González-Teshima, L.Y.; Forero-Forero, J.V.; Ayala-Zapata, S.; Hagerman, R. Fragile X Syndrome. Colomb Med (Cali) 2014, 45, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Protic, D.; Salcedo-Arellano, M.J.; Dy, J.B.; Potter, L.A.; Hagerman, R.J. New Targeted Treatments for Fragile X Syndrome. CPR 2019, 15, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Budimirovic, D.B.; Protic, D.D.; Delahunty, C.M.; Andrews, H.F.; Choo, T.; Xu, Q.; Berry-Kravis, E.; Kaufmann, W.E.; for the FORWARD Consortium. Sleep Problems in Fragile X Syndrome: Cross-sectional Analysis of a Large Clinic-based Cohort. American J of Med Genetics Pt A 2022, 188, 1029–1039. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, V.K.; Larkin, A.; Thurmond, J. The FlyBase Consortium Using FlyBase: A Database of Drosophila Genes and Genetics. In Drosophila; Dahmann, C., Ed.; Methods in Molecular Biology; Springer US: New York, NY, 2022; Vol. 2540, pp. 1–34. ISBN 978-1-07-162540-8. [Google Scholar]
- Dahlhaus, R. Of Men and Mice: Modeling the Fragile X Syndrome. Front. Mol. Neurosci. 2018, 11, 41. [Google Scholar] [CrossRef] [PubMed]
- Drozd, M.; Bardoni, B.; Capovilla, M. Modeling Fragile X Syndrome in Drosophila. Front. Mol. Neurosci. 2018, 11, 124. [Google Scholar] [CrossRef]
- Trajković, J.; Makevic, V.; Pesic, M.; Pavković-Lučić, S.; Milojevic, S.; Cvjetkovic, S.; Hagerman, R.; Budimirovic, D.B.; Protic, D. Drosophila Melanogaster as a Model to Study Fragile X-Associated Disorders. Genes 2022, 14, 87. [Google Scholar] [CrossRef]
- Inoue, S.B.; Shimoda, M.; Nishinokubi, I.; Siomi, M.C.; Okamura, M.; Nakamura, A.; Kobayashi, S.; Ishida, N.; Siomi, H. A Role for the Drosophila Fragile X-Related Gene in Circadian Output. Current Biology 2002, 12, 1331–1335. [Google Scholar] [CrossRef]
- Bolduc, F.V.; Valente, D.; Nguyen, A.T.; Mitra, P.P.; Tully, T. An Assay for Social Interaction in Drosophila Fragile X Mutants. Fly 2010, 4, 216–225. [Google Scholar] [CrossRef]
- Dockendorff, T.C.; Su, H.S.; McBride, S.M.J.; Yang, Z.; Choi, C.H.; Siwicki, K.K.; Sehgal, A.; Jongens, T.A. Drosophila Lacking Dfmr1 Activity Show Defects in Circadian Output and Fail to Maintain Courtship Interest. Neuron 2002, 34, 973–984. [Google Scholar] [CrossRef] [PubMed]
- Bushey, D.; Tononi, G.; Cirelli, C. The Drosophila Fragile X Mental Retardation Gene Regulates Sleep Need. Journal of Neuroscience 2009, 29, 1948–1961. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Hiesinger, P.R.; Schroeder, A.J.; Kume, K.; Verstreken, P.; Jackson, F.R.; Nelson, D.L.; Hassan, B.A. Drosophila Fragile X Protein, DFXR, Regulates Neuronal Morphology and Function in the Brain. Neuron 2002, 34, 961–972. [Google Scholar] [CrossRef] [PubMed]
- Sekine, T.; Yamaguchi, T.; Hamano, K.; Siomi, H.; Saez, L.; Ishida, N.; Shimoda, M. Circadian Phenotypes of Drosophila Fragile X Mutants in Alternative Genetic Backgrounds. Zoological Science 2008, 25, 561–571. [Google Scholar] [CrossRef] [PubMed]
- Tataroglu, O.; Emery, P. Studying Circadian Rhythms in Drosophila Melanogaster. Methods 2014, 68, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Klarsfeld, A.; Leloup, J.-C.; Rouyer, F. Circadian Rhythms of Locomotor Activity in Drosophila. Behavioural Processes 2003, 64, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Shafer, O.T.; Keene, A.C. The Regulation of Drosophila Sleep. Current Biology 2021, 31, R38–R49. [Google Scholar] [CrossRef]
- Dubowy, C.; Sehgal, A. Circadian Rhythms and Sleep in Drosophila melanogaster. Genetics 2017, 205, 1373–1397. [Google Scholar] [CrossRef] [PubMed]
- Helfrich, C.; Engelmann, W. Evidences for Circadian Rhythmicity in the Per° Mutant of Drosophila Melanogaster. Zeitschrift für Naturforschung C 1987, 42, 1335–1338. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. The Locomotor Activity Rhythm of Drosophila Melanogaster Is Controlled by a Dual Oscillator System. Journal of Insect Physiology 2001, 47, 877–887. [Google Scholar] [CrossRef]
- Andrew, D.R.; Moe, M.E.; Chen, D.; Tello, J.A.; Doser, R.L.; Conner, W.E.; Ghuman, J.K.; Restifo, L.L. Spontaneous Motor-Behavior Abnormalities in Two Drosophila Models of Neurodevelopmental Disorders. J Neurogenet 2021, 35, 1–22. [Google Scholar] [CrossRef]
- Dubowy, C.; Sehgal, A. Circadian Rhythms and Sleep in Drosophila Melanogaster. Genetics 2017, 205, 1373–1397. [Google Scholar] [CrossRef] [PubMed]
- Jagannathan, S.R.; Jeans, T.; Van De Poll, M.N.; Van Swinderen, B. Multivariate Classification of Multichannel Long-Term Electrophysiology Data Identifies Different Sleep Stages in Fruit Flies. Sci. Adv. 2024, 10, eadj4399. [Google Scholar] [CrossRef] [PubMed]
- Godino-Gimeno, A.; Thörnqvist, P.-O.; Chivite, M.; Míguez, J.M.; Winberg, S.; Cerdá-Reverter, J.M. Obesity Impairs Cognitive Function with No Effects on Anxiety-like Behaviour in Zebrafish. IJMS 2023, 24, 12316. [Google Scholar] [CrossRef]
- Geissmann, Q.; Garcia Rodriguez, L.; Beckwith, E.J.; Gilestro, G.F. Rethomics: An R Framework to Analyse High-Throughput Behavioural Data. PLoS ONE 2019, 14, e0209331. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Sheeba, V. VANESSA—Shiny Apps for Accelerated Time-Series Analysis and Visualization of Drosophila Circadian Rhythm and Sleep Data. J Biol Rhythms 2022, 37, 222–231. [Google Scholar] [CrossRef]
- Simon, A.F.; Chou, M.-T.; Salazar, E.D.; Nicholson, T.; Saini, N.; Metchev, S.; Krantz, D.E. A Simple Assay to Study Social Behavior in Drosophila: Measurement of Social Space within a Group 1. Genes Brain and Behavior 2012, 11, 243–252. [Google Scholar] [CrossRef]
Figure 1.
Presentation of individual actograms from the 3rd to the 10th experimental day during dark-dark (DD) period for: (a) rhythmic w1118 male; (b) arhythmic per01 male; (c) arhythmic dFMR1B55 male. Actograms of femailes are not presented due to less evident differences among them. Gray bars indicate subjective days under DD.
Figure 1.
Presentation of individual actograms from the 3rd to the 10th experimental day during dark-dark (DD) period for: (a) rhythmic w1118 male; (b) arhythmic per01 male; (c) arhythmic dFMR1B55 male. Actograms of femailes are not presented due to less evident differences among them. Gray bars indicate subjective days under DD.
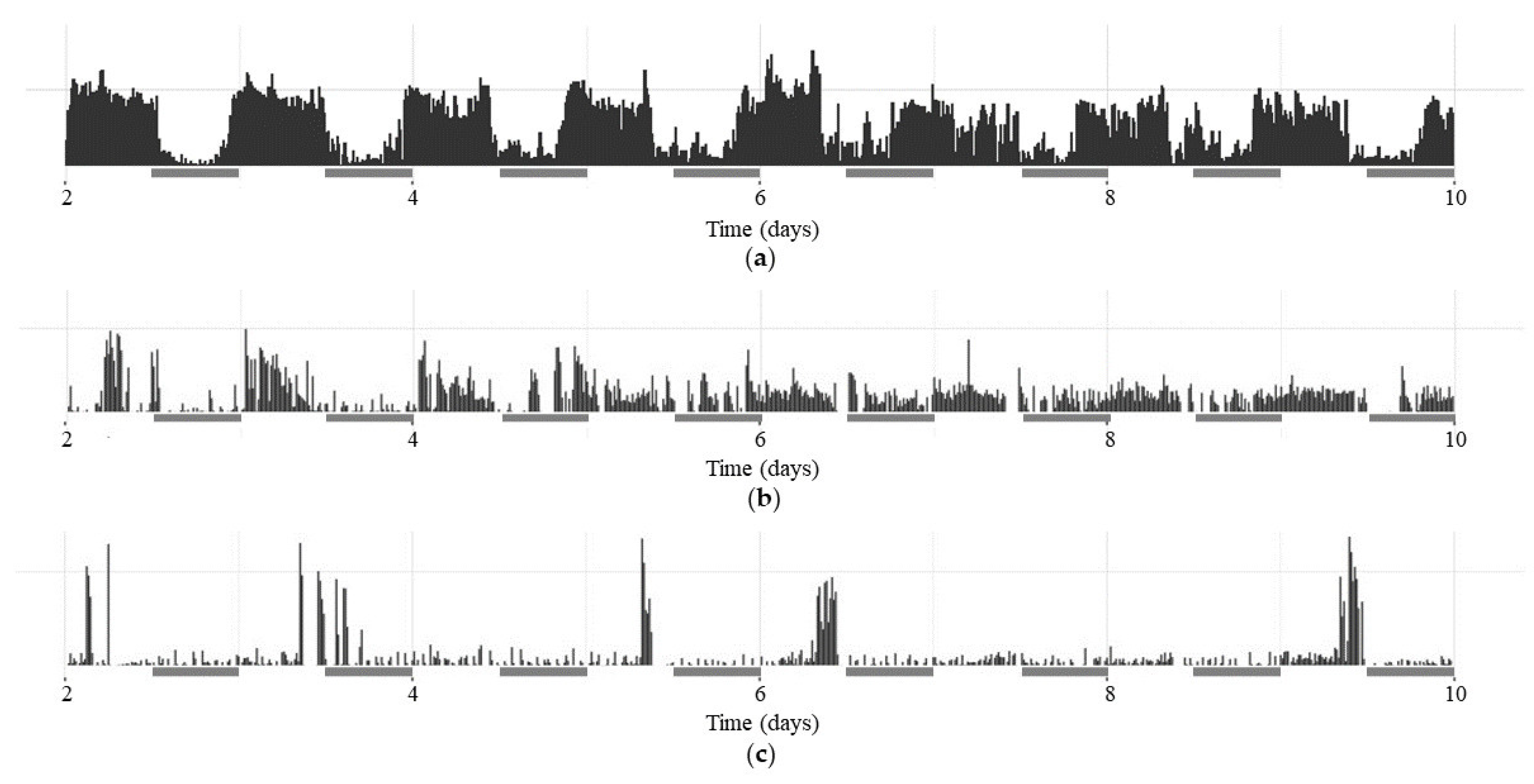
Figure 2.
Presentation of representative individual chi-square periodograms for: (a) rhythmic w1118 male; (b) rhythmic per01 male; (c) rhythmic dFMR1B55 male; (d) arhythmic per01 male and (e) arhythmic dFMR1B55 male. The red horizontal line depicts alpha = 0.05 threshold.
Figure 2.
Presentation of representative individual chi-square periodograms for: (a) rhythmic w1118 male; (b) rhythmic per01 male; (c) rhythmic dFMR1B55 male; (d) arhythmic per01 male and (e) arhythmic dFMR1B55 male. The red horizontal line depicts alpha = 0.05 threshold.
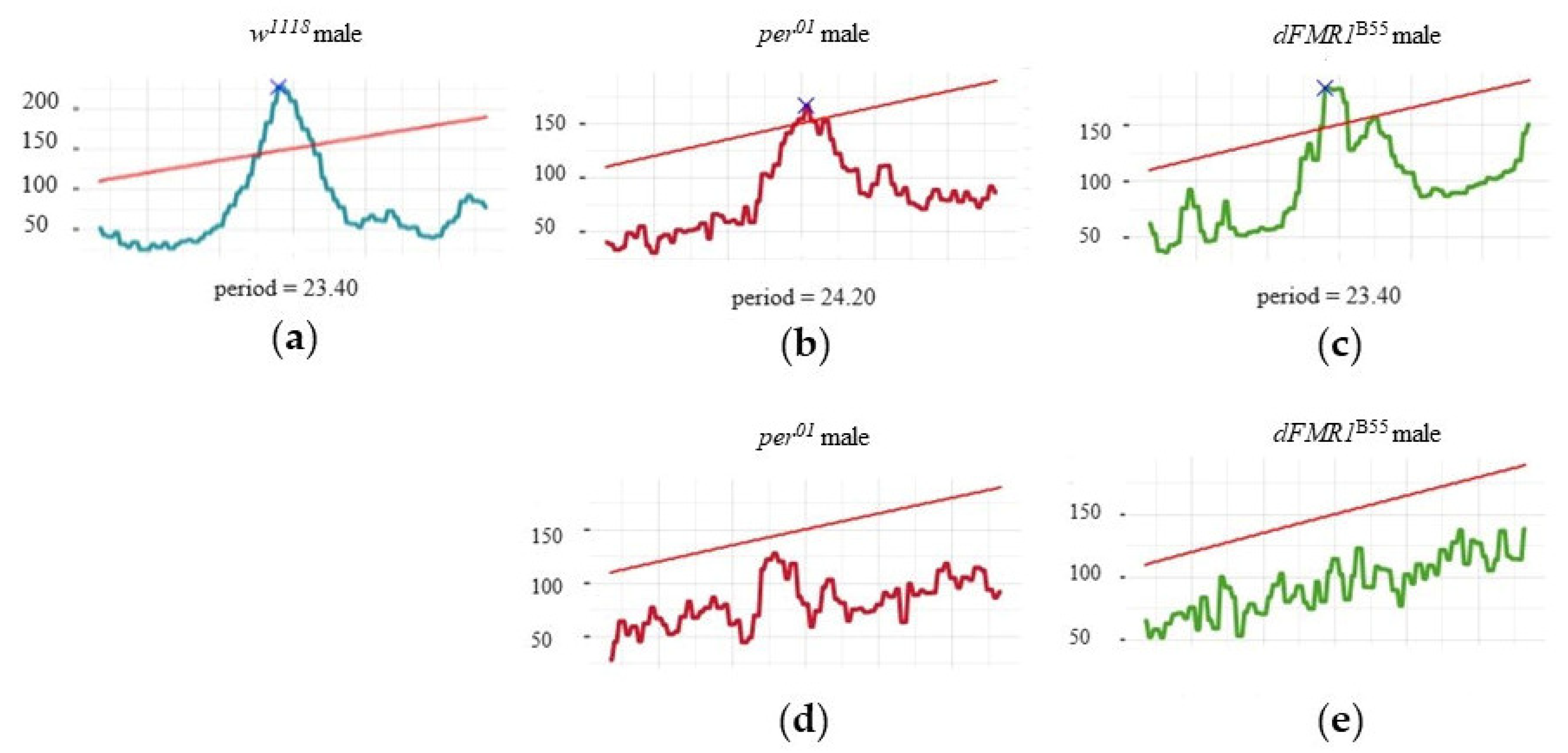
Figure 3.
Violin plots showing the distribution of periods of: (a) rhythmic w1118 males; (b) rhythmic per01 males; (c) rhythmic dFMR1B55 males.
Figure 3.
Violin plots showing the distribution of periods of: (a) rhythmic w1118 males; (b) rhythmic per01 males; (c) rhythmic dFMR1B55 males.
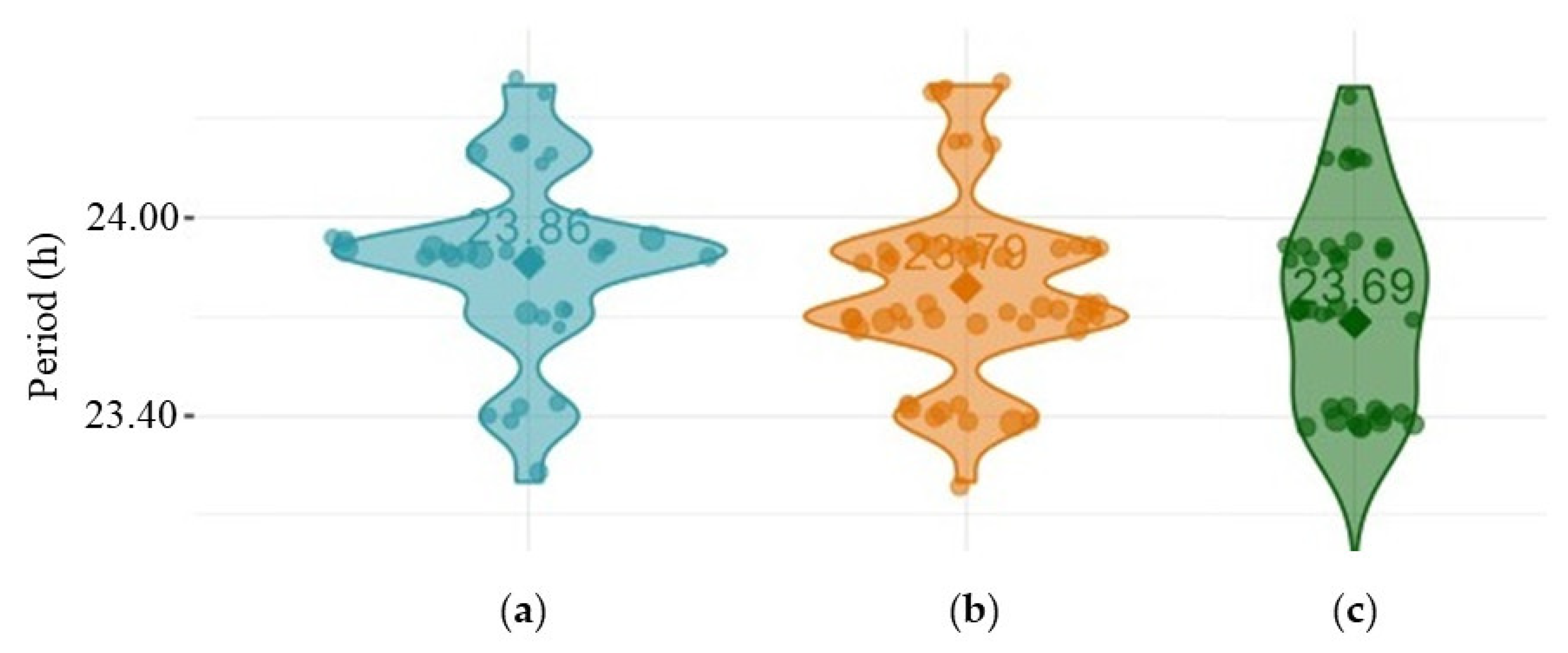
Figure 4.
Bar plots of CR parameters in w1118, per01 and dFMR1B55 rhythmic flies. (a) frequency of rhythmicity in females; (b) frequency of rhythmicity in males; (c) power of rhythm in females; (d) power of rhythm in males. Error bars, as graphical representations of the variability of data, represent the standard deviations of data sets relative to the mean.
Figure 4.
Bar plots of CR parameters in w1118, per01 and dFMR1B55 rhythmic flies. (a) frequency of rhythmicity in females; (b) frequency of rhythmicity in males; (c) power of rhythm in females; (d) power of rhythm in males. Error bars, as graphical representations of the variability of data, represent the standard deviations of data sets relative to the mean.
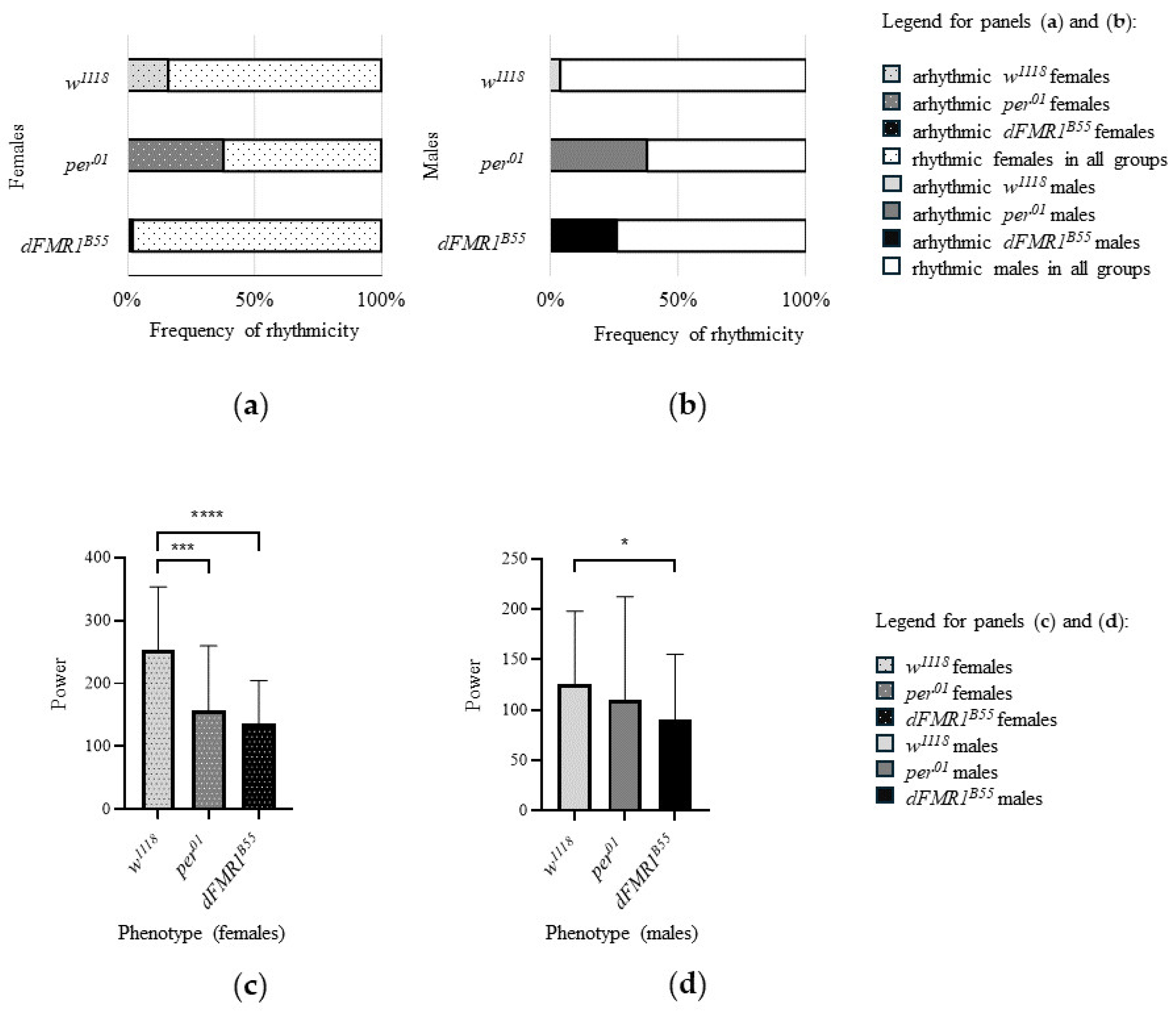
Figure 5.
Bar plots of total sleeping time during dark-dark (DD) periods in: (a) females and (b) males; and number of sleeping episodes during DD periods in: (c) females and (d) males.
Figure 5.
Bar plots of total sleeping time during dark-dark (DD) periods in: (a) females and (b) males; and number of sleeping episodes during DD periods in: (c) females and (d) males.
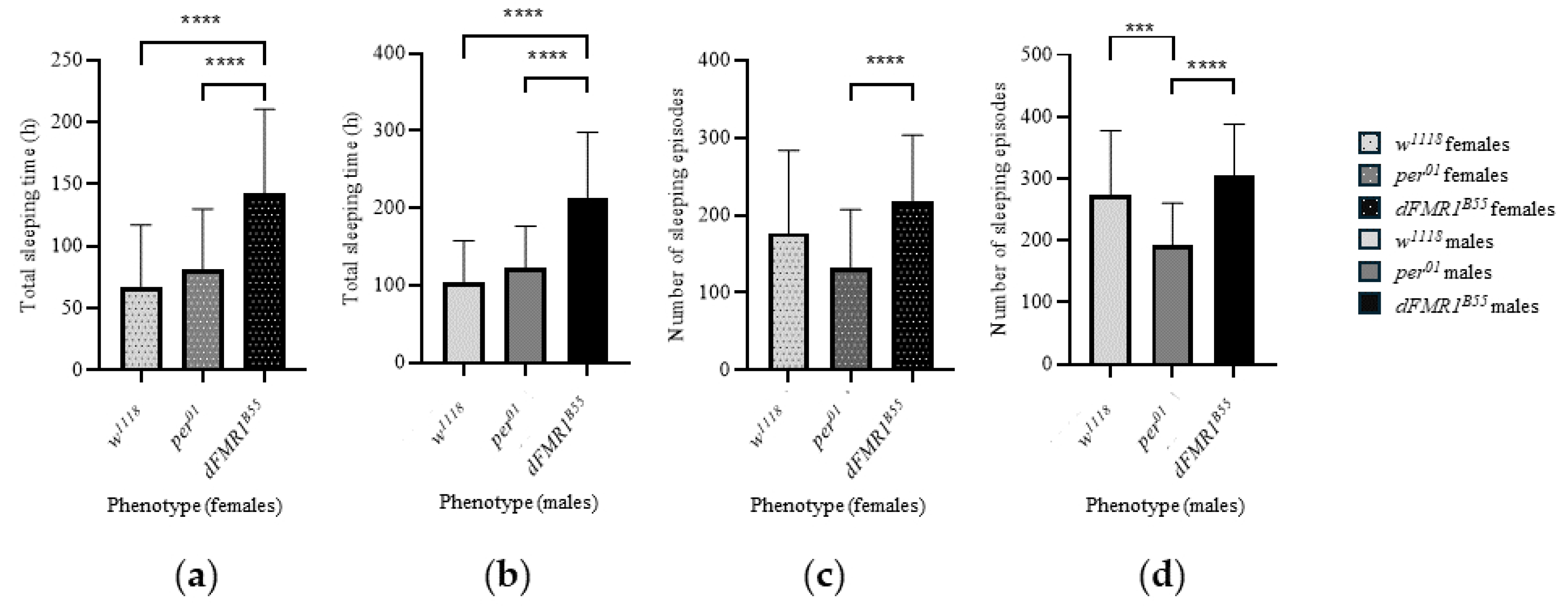
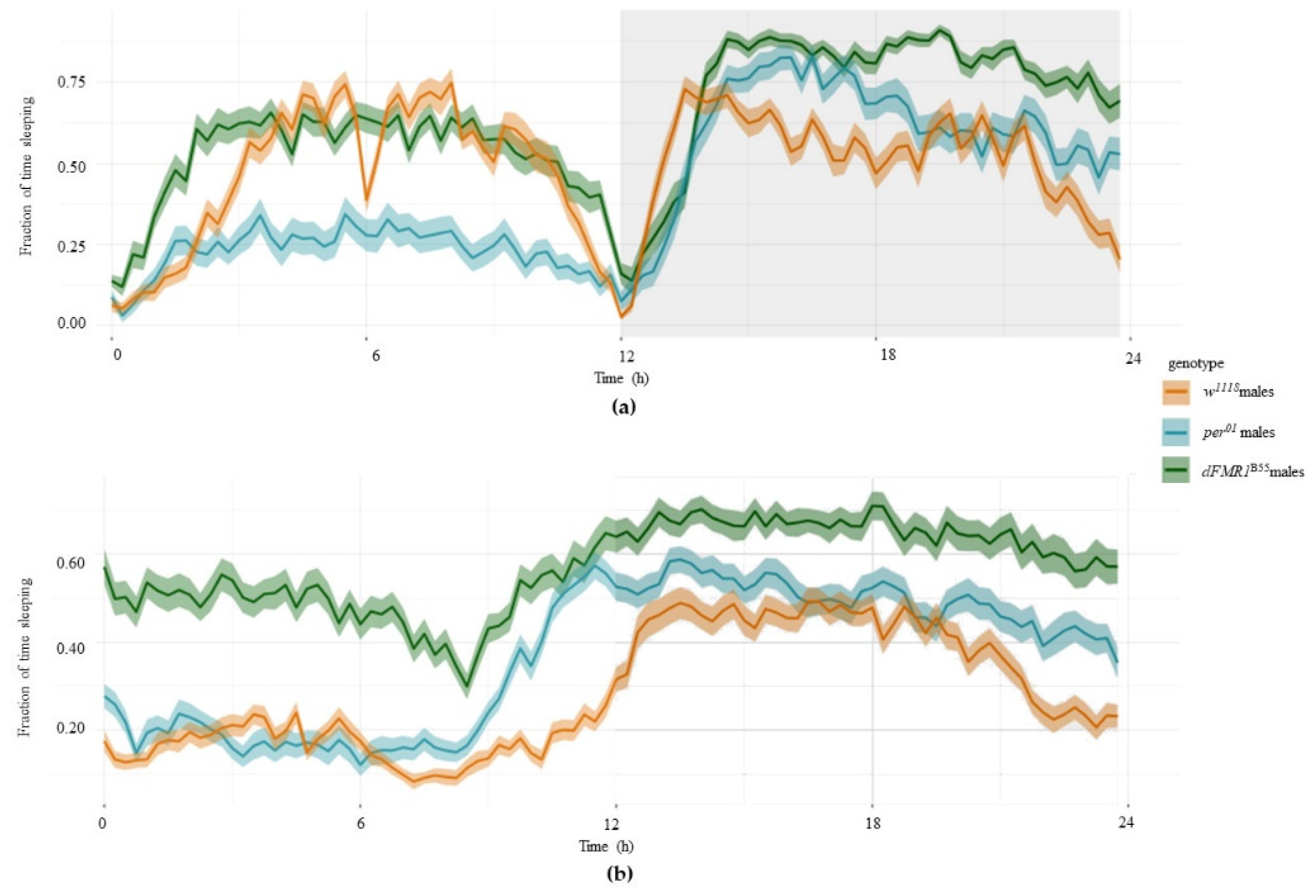
Figure 6.
Average sleep profiles for: (a) light-dark (LD) cycles (1st to 2nd day); (b) dark-dark (DD) cycles (3rd to 10th day).
Figure 6.
Average sleep profiles for: (a) light-dark (LD) cycles (1st to 2nd day); (b) dark-dark (DD) cycles (3rd to 10th day).
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



