
Submitted:
30 May 2024
Posted:
31 May 2024
You are already at the latest version
Abstract
Commercially cultivated Prunus species are commonly grown in adjacent or mixed orchards, and can be infected with unique or commonly shared viruses. Apple (Malus domestica), another member of the Rosacea and distantly related to Prunus, can share the same growing regions and common pathogens. Pollen can be a major route for virus transmission, and analysis of the pollen virome in tree fruit orchards can provide insights into these virus pathogen complexes from mixed production sites. Commercial honey bee (Apis mellifera) pollination is essential for improved fruit set and yield in tree fruit production systems. Here we describe the metagenomics-based detection of plant viruses through bee and pollen samples collected during the spring bloom period from a mixed Prunus and Malus orchard. Twenty-one unique viruses were detected in samples collected during apricot (Prunus armeniaca), sweet cherry (Prunus avium), peach (Prunus persica), and apple blooms. Ilarviruses prune dwarf virus (PDV) and prunus necrotic ringspot virus (PNRSV), Secoviridae family members tomato necrotic ringspot virus, tobacco necrotic ringspot virus, and prunus virus F, and Capillovirus cherry virus A (CVA) were detected in all time points, while other viruses were detected with restricted ranges. These results demonstrate the benefits of area-wide monitoring through bee pollination activities, and provide new insights into the diversity of viruses in tree fruit pollination ecosystems.
Keywords:
metagenomics
; virus
; pollen
; pollen transmission
; Prunus
; Honey bee
; Apricot
; Cherry
; Peach
; Apple
; Virus diversity
; Virus spillover
Introduction
The Rosaceae family contains over 100 genera and 3000 individual species, including many important fruit tree crops [1]. Prunus species including apricots (Prunus armeniaca), sweet cherry [Prunus avium (L.)], and peach (Prunus persica) are often grown in close proximity in commercial farming operations. Apples (Malus domestica) are commonly grown near Prunus orchards, but have a wider habitat and production regions than Prunus spp. Tree fruit production is a major sector in Canada, with apples accounting for 20.1% of total fruit farm gate values, followed by sweet cherries (5.7%), peaches (3.4%), and apricots (0.2%), together accounting for over 350 million Canadian (CDN) dollars in farm gate values across Canada in 2021 [2]. Viruses are a major threat to tree fruit production, and can result in severe losses [3]. The diversity, distribution, transmission, and host range of many of these viruses is poorly understood, especially at a whole-farm ecosystem level [4,5]. Effective monitoring approaches are required to fully appreciate the distribution of viruses in these systems, and the potential for virus transmission between tree fruit species and varieties.
Over 163 viruses have been reported to infect major woody fruit trees, of which at least six infect both Prunus species and apples including: apple mosaic virus (ApMV, genus Ilarvirus), apple chlorotic leaf spot virus (ACLSV; genus Trichovirus), cherry leaf roll virus (CLRV, genus Nepovirus), tomato ringspot virus (ToRSV, genus Nepovirus), and prunus necrotic ringspot virus (PNRSV, genus Ilarvirus) [3,6,7,8,9,10]. Common viruses of Prunus trees include prune dwarf virus (PDV; genus Ilarvirus) and cherry virus A (CVA; genus Capillovirus), which are not known to infect apples [11,12]. Other viruses have a more restricted Prunus host range, such as nectarine stem pitting virus (NSPaV, genus Luteovirus) which has only been reported in peach and nectarine [13,14]. At least twenty-one viruses are known to infect apple trees, including apple stem grooving virus (ASGV, genus Capillovirus), ACLSV, and apple stem pitting virus (ASPV, genus Foveavirus)[3]. Citrus concave gum associated virus (CCGaV; genus Phlebovirus) is a recently described virus infecting apple and citrus, but is not known to infect Prunus [3,10,15]. Viral host ranges can be quite complex, and are often not well defined [16,17].
Transmission pathways can impose restraints on plant virus evolution, and plays a role in determining the natural host range [17,18,19,20]. Plant viruses can be transmitted mechanically, through vectors, seed, or vegetative propagation. Pollen is another major route of transmission in tree fruits for some viruses, both horizontal and vertical [21,22]. Of the ~46 known pollen transmitted viruses, only 18 are horizontally transmitted, suggesting different mechanisms and restrictions to pollen-mediated virus transmission [21,22]. Viruses can also have direct negative effects on flowering and fruit production. PNRSV, for example, has been observed on the surface and within apricot pollen grains, and can negatively affect pollen germination [23,24]. In some cases, pollen germination and stigma penetration is essential for horizontal transmission, as demonstrated for raspberry bushy dwarf virus [25]. Other viruses can be potentially transmitted by pollinators like bees during foraging activities [26,27]. However, the relationship between the pollen virome and the orchard-wide plant virome is poorly understood [5,28,29]. Fruit tree orchards can be complex mixtures of closely related fruit tree species infected by multiple virus species with overlapping viral host ranges, but horizontal transmission could be restrained through pollen-pistil compatibility factors.
Some tree fruit species require pollen donors for successful fruit production. While peaches and many apricot varieties are self-compatible, many cherry varieties are self-incompatible, further complicating aspects of pollen transmission of viruses [30,31]. The risk of virus transmission between Prunus species through pollen is generally regarded to be low, while transmission between Prunus and Malus species would be even less likely. In addition to pollen-pistil compatibility, Prunus species and varieties do not necessarily flower at the same time, creating additional temporal barriers to pollen-associated virus transmission. Commercial honey bee (Apis mellifera) pollination services are widely used in tree fruit production systems to improve fruit set and fruit quality [32]. Honey bees forage intensively within a 1.5 km radius from their hives in agricultural settings, depending on flower availability [33,34]. Bees can also passively transmit plant viruses associated with pollen, or through direct contact with flowers [21,26,27,28].
Bees and pollen can be useful in monitoring agricultural systems for pathogen presence or ecosystem health [4,5,35,36,37]. Foraging honey bees are potentially carrying pathogen-contaminated pollen and nectar which could provide information on the presence of these pathogens in nearby flowering plants [5,36,37]. Here, we take advantage of bee-based area-wide metagenomics monitoring approaches to better understand viral species and variant diversity in tree fruit production systems, with a specific focus on apricots, cherries, peaches, and apples.
Materials and Methods
Farm Site and Sampling Details
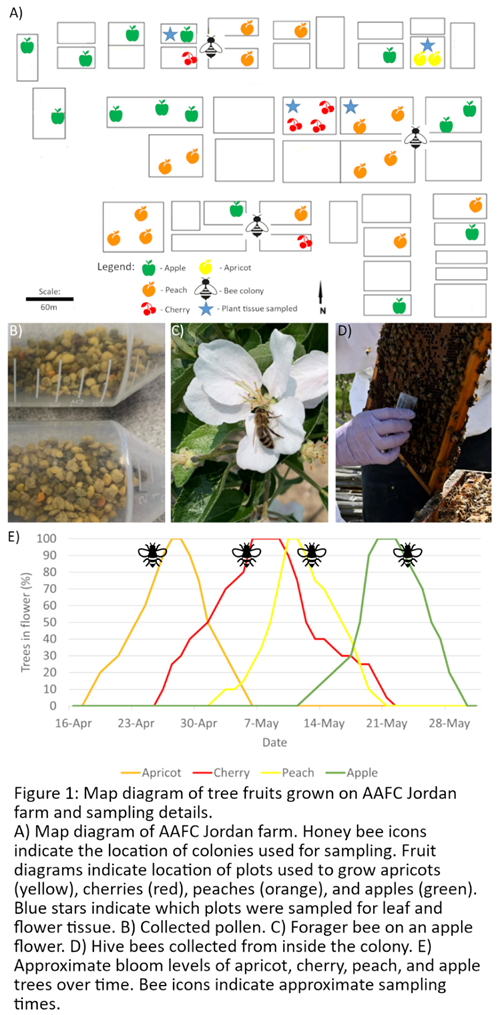
A long-established research farm containing multiple different tree fruit species in Jordan, Ontario (Jordan Farm) was selected as a study site because of the diversity and ages of tree fruit crops present. Three honey bee colonies were placed at various locations on the farm during the tree fruit bloom period in spring 2020, with each colony sampled independently at each time point, representing biological replicates for the site (Figure 1A). Three sample types were collected including ~25 Forager bees (collected outside colonies returning from foraging trips, with visible signs of pollen on their corbiculae), ~25 hive bees (collected from the brood nest of each hive, with no visible signs of pollen), corbiculae), ~25 hive bees (collected from the brood nest of each hive, with no visible signs of pollen), and ~5 mL pollen collected using pollen traps (ApiHex, Guelph, Canada) installed on the front of each hive (Figure 1B-D). Samples were collected at four time points during the spring of 2020 corresponding with peak bloom levels for apricots (April 23rd), cherries (May 4th), peaches (May 12th), and apples (May 26th)(Figure 1E). In addition, two plant sample replicates consisting of leaf and flower tissue were collected from 10 random individuals located near the bee colony. One plot was sampled for leaf/flower tissue from each representative crop species examined (apricot, cherry, peach, and apple; Figure 1A).The most prominent tree fruit species grown on Jordan farm at the time of sampling were peaches and apples, followed by cherries and apricots (Figure 1A; Table 1). Trees were of mixed age, variety and rootstock combinations (Table 1). One plot each of nectarines (Prunus persica v . nucipersica) and plums (Prunus domestica) were also grown on site, and in flower during the period of sample collection, but were not analyzed as a part of this study. Other crops grown at this site included garlic (Allium sativum), strawberries (Fragaria x ananassa), grape (Vitis vinifera), and hops (Humulus lupulus) but were not flowering during the sample collection period (not shown).
RNA Extraction and Sequencing
Sample processing, extraction and sequencing were performed as in Smadi et al., 2024 [5,36]. Briefly, total RNA (totRNA) was extracted using the spectrum total plant RNA extraction kit (Sigma Aldrich, ON, Canada) while dsRNA was extracted from composite plant samples following Kesanakurti et al. (2016)[12]. Extracted totRNA was treated with an rRNA depletion step using the RiboMinusTM Plant Kit for RNA-Seq (Invitrogen, Waltham, MA, USA) as per the manufacturer’s instructions. Ribo depleted totRNA and dsRNA HTS libraries were generated using the Illumina TruSeq Stranded mRNA Library Prep kit, following the manufacturer’s protocol, starting after the mRNA selection steps [12]. Libraries were dual indexed using the IDT for Illumina TruSeq RNA UD Indexes (Illumina, San Diego, CA, USA), normalized and sequenced using a NextSeq500 high output kit v2.5, 75 cycles (Illumina) which generated between 10 and 16 million reads, on average for each sample. DsRNA samples were also sequenced using a NextSeq500, but with approximately four to six million reads on average for each sample. RNAseq files were uploaded to the Sequence Read Archive and individual virus CP sequences were uploaded to genbank (Supplemental table 1; supplemental file 1).
Bioinformatics and Phylogenetics
HTS sample files were imported into Virtool (www.virtool.ca) for sample management, quality control (QC) and data analysis. Reads passing QC using FASTQC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/, accessed on September 2021), were mapped to plant virus databases updated December 2021 using the Pathoscope 2 pipeline [38]. Reads were aligned to representative isolates of all known plant viruses pulled from Genbank in September 20th, 2021. Viruses with representative isolates receiving at least one mapped read then had reads mapped against all of their known isolates. Bowtie2 2.3.2 [39] was used in local mode for both rounds of read mapping with minimum score (–min-score) set to “L, 20, 1.0”, seed length (-L) to 15, and mismatches per seed (-N) set to 0. In the second round, the maximum number of alignments returned (-k) was set to 100. Reads matching viruses were also mapped to a host reference genome. Except for a 0 value for mismatches per seed (-N), default Bowtie2 parameters were used for mapping. Reads were eliminated from the analysis if they had a greater or equal alignment score to the host versus the virus. Multi-mapping reads were handled using a refactored derivative of the Pathoscope2 identification module, which exactly matches the output of the published module. Read values are fractionally reassigned from least likely source genomes to most likely [40]. Virus identification based on Virtool was used to create sample-specific pathogen profiles. A minimum of 10% genome coverage was required for a virus species to be considered a positive detection from both totRNA and dsRNA samples. Sample profiles were combined to create a site-specific profile which included calculating the average frequency of detection for each sample type, and the average frequency of detection across all samples, average genome coverage, and Viral reads per million (VRPM) for each virus detected across all samples (Supplemental file 1). VRPM is similar to Transcripts Per Million, and was calculated from the total number reads mapping to each individual virus, dividing by genome length of the virus in kilobasepairs (Kbps), and then normalized for the total number of reads in the sequencing run, per million. Since Pathoscope 2 assigns reads aligning to multiple genomes proportionally, this can cause some inconsistencies with VRPM values when reads map to more than one closely related virus species [39]. Due to inconsistencies of viroid detections, read counts and VRPM were manually annotated using Geneious prime version 11.0.14.1 (Biomatters inc, CA, USA).
Using host genome-subtracted de novo assembled contigs for each sample, sequences were aligned to the PDV, PNRSV, and CVA reference sequence using Geneious Prime (Supplemental table 1). Samples with full coat protein nucleotide sequence coverage were used for pairwise and phylogenetic analysis, along with genbank reference sequences. Pairwise nucleotide distance comparisons were constructed using Geneious prime. Maximum likelihood phylogenetic trees were constructed using MEGA 11 with 1000 bootstrap replications [41].
Results
Virus Detection during Tree Fruit Bloom
Viral profiles from each sample were used to calculate the average frequency of detection in each sample type replicate, at each time point. The total average genome coverage and average VRPM across all samples was also calculated (Table 2; Supplemental file 1). A total of 21 virus species were identified from bee and pollen samples: PDV was most commonly detected (64% ), followed by CVA (58%), PNRSV (58%), prunus virus F (44 %; PVF; genus Fabavirus), ToRSV (44 %), and tobacco ringspot virus (44 %; TRSV; genus Nepovirus) (Table 2). The Ilarvirus genus was best represented, including PDV, PNRSV, apple mosaic virus (11%), blackberry chlorotic ringspot virus (8%), tobacco streak virus (8%), and tomato necrotic shock virus (ToNSV; 6%; Table 2). The six most frequently detected viruses (PDV, CVA, PNRSV, PVF, TRSV, and ToRSV) were detected at all four sampling time points, and in all sample types (Figure 2). CVA had the highest average genome coverage (70%), while PDV had the highest normalized read counts (VRPM; Table 2).

Virus Species Diversity in Different Sample Types and Sampling Times
To understand the distribution of viruses across sample types and time points, the average number of virus species identified in each sample type was calculated (Figure 2A). Four viruses were identified in the majority of samples, with a range of two to eight viruses identified per sample (Figure 2A). When examining the number of viruses detected among sample types irrespective of time point, a significant main effect was detected (One-way ANOVA, p = 0.007, F = 5.80, df = 2), with differences observed between the number of virus types identified in pollen and hive bee samples (Tukey’s HSD, p = 0.005. When comparing among sampling times, no significant difference was observed between the number of viruses identified (One-way ANOVA, p = 0.71, F = 0.40, df = 3; Figure 2A). CVA, PVF, BMV, TRSV, ToRSV, PDV, PNRSV, and PLMVd were detected at all time points. TSV was associated with apricot and cherry time points, CCGaV and ASGV were unique to the peach time point, ToNSV, AHHVd peanut stunt virus (PSV; genus Cucumovirus) and citrus virus A (CiVA; genus Coguvirus) to the apple time point, and raphanus sativus cryptic virus 3 (RSCV3), ApMV, and PZSV with both peach and apple time points (Figure 2B). PDV, PNRSV, and CVA were the three most prominent viruses detected overall, and they were most frequently detected during the cherry time point (Figure 2C). The greatest number of virus species were identified in pollen and forager bee samples (Figure 3A). No unique virus species could be associated with hive bees, with viruses identified in hive bees also identified in pollen and forager bee samples (n = 7), including CVA, PDV, PNRSV, TRSV, ToRSV, PVF, and BMV (Figure 3A).
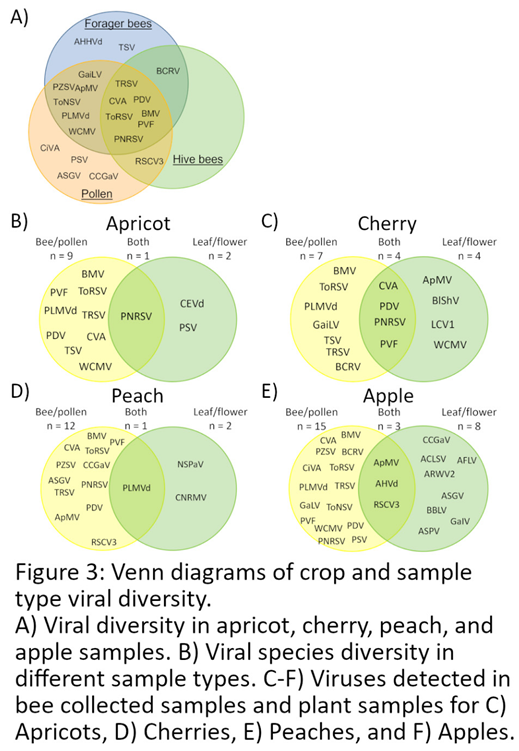
To correlate viruses identified in bee-collected samples with viruses infecting the targeted tree fruit species, two replicates of composite leaf and flower samples were collected from ~10 random individuals from one apricot, cherry, peach, and apple plot located near the bee colonies, at the same time as when the bee samples were collected (Figure 1A). As opposed to totRNA extracted bee/pollen samples, dsRNA was extracted from leaf/flower samples to preferentially isolate replicating viruses (Table 3). During the apricot time point fewer viruses were detected in leaf/flower (n = 3) compared to bee/pollen samples (n = 10), with only PNRSV identified in both sample sets from this time point (Figure 3B). In cherries, seven viruses were unique to bee/pollen samples [BMV, ToRSV, PLMVd, TSV, TRSV, BCRV, and Gaillardia latent virus (GaiLV; genus Carlavirus)], four viruses (PDV, CVA, PNRSV, and PVF) were identified in both bee/pollen and leaf/flower samples (Figure 3C), while four viruses were unique to leaf/flower samples (ApMV, BlShV, LCV1, and WCMV). In peaches, 13 viruses were identified in bee/pollen samples compared with three from leaf/flower samples; nectarine stem pitting associated virus (genus Luteovirus) and cherry necrotic rusty mottle virus (genus Robigovirus) were detected only in leaf/flower samples, while PLMVd detected in both sample types (Table 3; Figure 3D). Many viruses were identified in apple leaf/flower samples (n = 8), but not in bee/pollen samples including ACLSV, apple flat limb virus (AFLV; genus Rubodvirus), apple rubbery wood virus 2 (ARWV2; genus Rubodvirus), ASPV, ASGV, and CCGaV (Figure 3E). Viruses identified in both sample types from apples included ApMV, AHHVd, and RSCV3 (unclassified Partitiviridae family) (Figure 3D).
Coat Protein Sequence Diversity of CVA, PDV, and PNRSV
Pairwise and phylogenetic analysis of the PDV, CVA, and PNRSV coat protein (CP) nucleotide sequences were undertaken to investigate virus sequence diversity. Only sample data with full coverage of the CP open reading frame (ORF) for one or more of the three viruses were used. The PNRSV CP ORF sequences were the most diverse ranging from 89-100 % identity, while CVA and PDV CP sequences ranged from 95.3 - 98.5 % and 96 – 100 % identity, respectively (Figures 4A, 5A, and 6A). In total, 11 samples had full CVA CP coverage, including two leaf/flower samples collected from cherries (ONJF1-CH-1T1 and ONJF1-CH-1T2; Figure 4; Supplemental table 1). Six CP sequences were obtained from cherry time point samples, three from apricot, and two from apple (Figure 4B). All CVA CP sequences clustered closely together with a previously reported CVA isolate from Jordan farm (MF062118), and grouped within phylogroup II as defined in Gao et al., 2017 (Figure 4B)[42,43].
Complete PDV CP sequences were obtained from 26 sample data sets, including plant tissue samples from cherry (n = 2) (Figure 5A; Supplemental table 1). Two branches of sequences derived from these samples were identified, all within PDV phylogroup II as defined in Kinoti et al., 2018 (Figure 5B)[44]. The first branch contained nine sequences derived from peach (n = 4), cherry (n = 3, two from leaf/flower samples), and apricot (n = 2), and clustered closely with the reference sequence (NC_008038). Identities within this branch ranged from 97.9-100 % (Figure 5B). The second branch contained eight sequences with identities ranging from 99.2-100 %, and clustered closely with an isolate from a Bulgarian sweet cherry sample (MK139682; Figure 5)[45]. Sequence identity was typically over 98 % within each branch, and less than 97.6 % identical between branches (Figure 5A). Five other sequences derived from apricot and cherry samples branched more independently (Figure 5B).
In total, 23 PNRSV CP sequences were recovered from pollen, bee and plant sample data sets (Figure 6A). Of these, 17 clustered within the PV32 phylogroup along with the reference sequence (NC_004363), and shared a high degree of identity (> 98.5 %)(Figure 6) [46]. These sequence data were derived from all four time points, including only one isolate from the apple time point (ONJF1-AP-2P; Figure 6B). Two isolates derived from a cherry and apricot leaf/flower samples were included in this group. One sequence derived from an apricot hive bee sample branched more closely with the PV96 phylogroup (ONJF1-AT-3H). One sequence derived from an apple pollen sample (ONJF1-AP-3P) was distantly related to all other sequences from this study (90.4 – 91.9 %) and did not associate with any particular phylogroup.


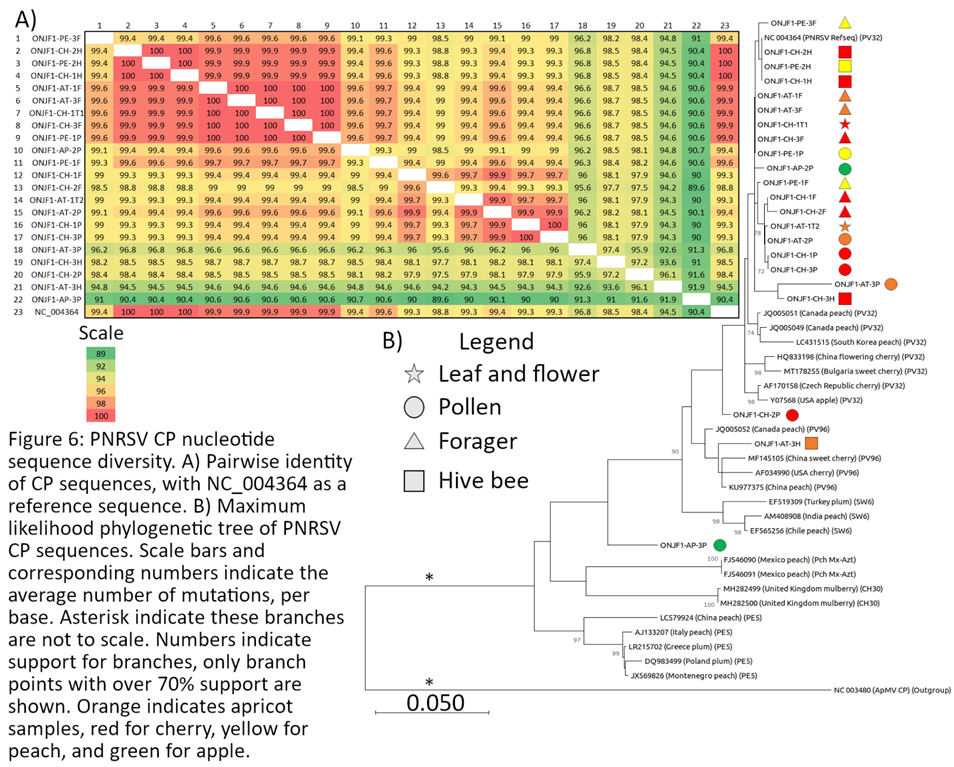

Table 3.
Plant virus detection from apricot, cherry, peach, and apple leaf/flower tissue.
| Virus species | Genus or family | Frequency (%) | Total Detections(n) | Average frequency of detection (%) | Average Genome Coverage (%) | Average VRPM | |||
| Apricot | Cherry | Peach | Apple | ||||||
| Plant tissue | |||||||||
| n=2 | n=2 | n=2 | n=2 | ||||||
| Prunus necrotic ringspot virus | Ilarvirus | 100 | 50 | 3 | 38 | 70.8 | 2041 | ||
| Apple mosaic virus | Ilarvirus | 50 | 50 | 2 | 25 | 20.6 | 0 | ||
| Cherry virus A | Capillovirus | 100 | 2 | 25 | 99.6 | 1181 | |||
| Prune dwarf virus | Ilarvirus | 100 | 2 | 25 | 15.7 | 1 | |||
| Apple chlorotic leaf spot virus | Trichovirus | 100 | 2 | 25 | 78.7 | 500 | |||
| Apple flat limb virus | Rubodvirus | 100 | 2 | 25 | 39.65 | 4 | |||
| Apple rubbery wood virus 2 | Rubodvirus | 100 | 2 | 25 | 91.5 | 107 | |||
| Apple stem pitting virus | Foveavirus | 100 | 2 | 25 | 67.6 | 225 | |||
| Prunus virus F | Fabavirus | 100 | 2 | 25 | 97.1 | 67 | |||
| Little cherry virus 1 | Velarivirus | 100 | 2 | 25 | 99.95 | 1314 | |||
| Nectarine stem pitting-associated virus | Luteovirus | 100 | 2 | 25 | 82.2 | 223 | |||
| Cherry necrotic rusty mottle virus | Robigovirus | 100 | 2 | 25 | 95.05 | 58 | |||
| Raphanus sativus cryptic virus 3 | Unclassified Partitiviridae | 50 | 1 | 13 | 18.9 | 0 | |||
| Apple hammerhead viroid | Pelamoviroid | 50 | 1 | 13 | 100 | 2410 | |||
| Apple stem grooving virus | Capillovirus | 50 | 1 | 13 | 99.6 | 7528 | |||
| Citrus concave gum associated virus | Coguvirus | 50 | 1 | 13 | 99.2 | 417 | |||
| Peach latent mosaic viroid | Pelmaviroid | 50 | 1 | 13 | 99.6 | 27 | |||
| Blueberry latent virus | Amalgavirus | 50 | 1 | 13 | 13.2 | 4 | |||
| Blueberry shock virus | Ilarvirus | 50 | 1 | 13 | 11.1 | 7 | |||
| Citrus excordis viroid | Pospiviroidae | 50 | 1 | 13 | 96.4 | 169 | |||
| Grapevine associated ilarvirus | Ilarvirus | 50 | 1 | 13 | 24.6 | 1 | |||
| Peanut stunt virus | Cucumovirus | 50 | 1 | 13 | 16.9 | 0 | |||
| White clover mosaic virus | Potexvirus | 50 | 1 | 13 | 26.7 | 0 | |||
Discussion
Metagenomics Based Detection of Plant Viruses through Pollen
Honey bees are capable of foraging from a broad diversity of plants in various landscapes [49]. Individual honey bees choose floral resources based on exploiting profitable flowers in the vicinity of their hive and exhibit considerable fidelity to a crop or floral patch during successive flights, only switching when pollen becomes scarce or unattractive [47]. Colony-level pollen foraging decisions are affected by landscape and closely follow changes in the seasonal availability of floral resources [48,49,50]. Flowering plant species including those of agricultural importance, have overlapping bloom periods which can result in a complex of pollens from multiple species being collected by honey bees at any one time. While each time point represented the peak bloom period for each species we examined, other tree fruit and naturally occurring plant species were in bloom which could have contributed to the diversity of pollens collected, resulting in a complex mixture of pollens from multiple potential plant species (Figure 2B). Consistent with this, CVA, PDV, and PNRSV coat protein sequences did not assort together based on time of sampling, suggesting broad floral inputs into bee and pollen samples. It is therefore difficult to accurately ascertain the origins of viruses identified from pollen-based environmental sampling [50]. Viruses could be originating from closely related Prunus and Malus species or potential weedy host reservoirs.
The identification of plant species from pollen samples through DNA metabarcoding could help to define the limits of pollen-based virus identification and better understand the potential for alternate hosts for viruses of concern [51,52,53,54]. Combining work-flows to include multiple environmental-health indicators including bacterial and fungal pathogens, pesticide and chemical monitoring, invasive plant species, and even honey bee pathogens and parasites, could create a powerful one-health monitoring tool [5,37,51,52,54,55,56,57,58]. Regardless, the most frequent detections of plant viruses (ex. PDV and PNRSV) were viruses known to cause disease in these systems, and identified a striking amount of nucleotide sequence diversity, demonstrating the accuracy and sensitivity of pollen-mediated metagenomics based detection of plant viruses in tree fruit orchard systems [5,59].
Viral Diversity in Bee-Collected Pollen Samples
Many viruses are associated with pollen transmission including members of the Alphacryptovirus, Ilarvirus, Nepovirus, and Potyvirus genera [21,59,60]. Ilarviruses in particular were widely prevalent in this study, with representatives from three of the four Ilarvirus subgroups identified [60,61]. ToNSV from subgroup 1, BCRV, TSV, and PDV from subgroup 3, and PNRSV and ApMV from subgroup 4 were detected, demonstrating a large diversity of Ilarviruses at this site. BCRV and ToNSV are not normally associated with Prunus hosts, and while BCRV has been reported to infect apple this virus was not detected in our survey of leaf/flower samples from apples [62,63]. BCRV is typically associated with cane fruits like blackberries (Rubus allegheniensis) or other Rubus plants [63,64] Primary hosts for ToNSV are Solanaceous plants [65], yet no Solanaceous crops were grown at the time on Jordan farm. Both viruses were detected in low frequencies, which highlights the sensitivity of this approach and complexities of interpretation. Three other viruses in the Bromoviridae family were identified including BMV, PSV, and pelargonium zonate spot virus (genus Anulavirus). BMV, PSV, BCRV, and many other viruses identified in this study were also identified in a recent survey of the pollen virome of wild plants, confirming a strong association with pollen, and suggests they could be widespread in the endemic plant pollen ecosystem near Jordan farm [4]. Understanding the host origins of viruses in bee and pollen samples can be difficult, but can inform targeted follow up studies in commercial fruit crops [66].
Viruses in the Secoviridae family were also detected in all time points, including nepoviruses ToRSV and TRSV, and Fabavirus PVF [67,68]. Nepoviruses are commonly transmitted via nematodes, seed and pollen [67], while known regarding the transmission of PVF. Fabaviruses have a broad host range and are associated with aphid transmission but no vector has been identified for PVF to date, while this and our previous study strongly associate PVF with pollen in tree fruit systems [5,69]. ToRSV and TRSV were not identified in leaf/flower samples, but were present in all other bee and pollen samples. ToRSV was more frequent during apricot and cherry blooms while TRSV was more frequent during apple bloom, but these three viruses were not identified in leaf/flower samples. It is possible they could have been missed in our limited random sampling of leaf/flower tissue, or are infecting other plant species in the area.
The number of viruses identified per sample was largely consistent from apricot through to the peach bloom, with slightly more viruses detected in apple pollen samples, despite substantial increases in the number of individual trees in flower as the bloom period progressed (Table 1). However, the viral species diversity increased during peach and apple blooms correlated with increased numbers of individual in flower. PDV, PNRSV, and CVA were the most frequently detected viruses, present across all time points and sample types, yet were more prevalent during the cherry time bloom. Virus profiles taken during flowering of the three Prunus species were more similar, whereas the profiles obtained during the apple flowering time point included detections of apple-infecting viruses/viroids like CiVA, ApMV, and AHHVd. Apple-infecting viruses like ASGV and CCGaV were only identified in the peach time point, suggesting potential crossover from apple or other pome pollen. Two apple-infecting coguviruses (CCGaV and CiVA) were detected in one sample from peach and apple, respectively. These two viruses were also prominent in apple pollen samples from our previous study, which could suggest a previously unreported association with pollen [5]. Coguviruses can be graft-transmissible, and CCGaV was recently reported to be seedborne [70,71].
Sequence Diversity of Identified Viruses
The known cherry-infecting viruses CVA, PDV and PNRSV were the most common viruses detected during the cherry time point, each with a unique pattern of observed nucleotide sequence diversity. All CVA CP sequences clustered with a previously described cherry isolate from this site [45,72]. In this study, aside from detection in bee/pollen samples from all time points, CVA was identified in cherry leaf/flower tissue with high read counts, but not from other Prunus or apple leaf/flower samples consistent with this virus originating primarily from cherry trees (Figure 4). Consistent with previous studies, CVA nucleotide sequences were highly variable despite grouping together in phylogenetic analysis [44,73]. CVA is transmitted via grafting and no other major transmission route has been widely reported; further studies could investigate the high association of CVA with pollen [74].
PDV isolates cluster into three major phylogroups, with all isolates identified in this study belonging to phylogroup II [44]. Phylogenetic and pairwise sequence analysis identified two distinct sub-groupings within phylogroup II, one associated with peach bee/pollen samples and two cherry leaf/flower samples, and another detected in all four sampling time points. Additional minor isolates were identified including one closely related to an isolate from cherries in British Columbia (MZ220994), demonstrating multiple variants of PDV present at this site. The high variability and distinct groupings of isolates in this study could be due to the wide variety of Prunus species, varieties, and ages of plants. Further studies could investigate the host range and pathogenicity of these variants, which could help to identify priorities for management.
PNRSV variants present in the samples were more diverse than CVA or PDV, with CP nucleotide sequence identities ranging from 89.6 – 99.7 %. This was greater than previous reports from this site (95.1 - 100 %, or in other related studies of PNRSV pathology (94 – 100 %)[43,75]. The PNRSV sequence data separated into two major clusters, one associated with apricot leaf/flower samples, and the second from cherry leaf/flower samples, suggesting unique variants of PNRSV associated with these hosts. A third, unique variant of PNRSV with the lowest identity to all other samples (89.6-91.9 %) was detected from the apple time point, possibly arising from apple trees, or other host flowering at this site at the same time. Data from this study demonstrate a broad diversity of sequences within the PV32 phylogroup, one isolate associated with PV96 [29,75], and one isolate from an apple pollen sample distantly related to other groups. PV32 and PV96 have been described as mild isolates of PNRSV, but can still result in suppressed growth, necrotic spots and chlorosis on Prunus tomentosa [76].
Conclusion
Bee-mediated plant virus monitoring can provide an overview of the pollen-associated virome, and potential viral pathogens in the immediate area [5,36,37,48]. Bee-mediated sampling can greatly facilitate sampling from multiple individuals at a commercial farm site which can often have thousands of trees, but determining the exact origins of viruses detected or understanding the dangers associated with transmission is still poorly understood. Compared to virus identification from plant samples based on visual symptoms, bee/pollen-mediated surveys of viral diversity could provide many advantages including reduced sampling labour, capturing greater levels of viral diversity, and identifying emerging viruses before they become widespread. PDV, PNRSV and CVA were widely prevalent in bee and pollen samples from most time points, with the highest frequency of detection during cherry bloom. Viruses detected during apple bloom were consistent with apple-infecting viruses like AHHVd, ApMV, and CiVA. A broad diversity of PDV and PNRSV variants were present at this site based on CP nucleotide sequence comparisons, demonstrating many advantages to bee-mediated metagenomic based virus monitoring.
Competing Interests
The authors declare no competing interests.
Author Contributions
Conceptualization J.S.G.; sample collection R.V. and J.S.G.; Sample processing: J.P.; Data curation R.V.; Sequencing M.R.; Data analysis R.V. and J.S.G. Writing – original draft preparation, J.S.G., writing – review and editing, all authors. Funding acquisition, J.S.G., M.R., M.M.G., S.F.P., and G.B.
Funding
This research was funded by the Government of Canada through Agriculture and Agri-Food Canada) and the Canadian Food Inspection Agency (CFIA) interdepartmental project funding to JSG, MR, MMG, GB and SFP (AAFC Project ID# J-002323, CFIA project ID# N-000212), as well as CFIA GRDI funding to GB and MR [CFIA 2546 GRDI-SID-P-1903].
Acknowledgements
We wish to thank Graham B. Thurston, Canadian Food Inspection Agency, for help in developing ideas related to this manuscript. We also wish to thank the farm staff at Jordan farm for ongoing maintenance and cultivation of orchards at this site.
References
- Soundararajan, P.; Won, S.Y.; Kim, J.S. Insight on Rosaceae Family with Genome Sequencing and Functional Genomics Perspective. BioMed Res. Int. 2019, 2019, 1–12, . [CrossRef]
- AAFC - Agriculture and Agri-Food Canada. Statistical overview of the Canadian Fruit Industry 2021. Horticulture Section, Crops and Horticulture Division (2022). https://agriculture.canada.ca/sites/default/files/documents/2022-12/Fruit%20Report_2021_ENG.pdf.
- Umer, M.; Liu, J.; You, H.; Xu, C.; Dong, K.; Luo, N.; Kong, L.; Li, X.; Hong, N.; Wang, G.; et al. Genomic, Morphological and Biological Traits of the Viruses Infecting Major Fruit Trees. Viruses 2019, 11, 515, . [CrossRef]
- Fetters, A.M.; Cantalupo, P.G.; Wei, N.; Robles, M.T.S.; Stanley, A.; Stephens, J.D.; Pipas, J.M.; Ashman, T.-L. The pollen virome of wild plants and its association with variation in floral traits and land use. Nat. Commun. 2022, 13, 1–11, . [CrossRef]
- Smadi, M.; Lee, E.; Phelan, J.; Wang, A.; Bilodeau, G.J.; Pernal, S.F.; Guarna, M.M.; Rott, M.; Griffiths, J.S. Plant virus diversity in bee and pollen samples from apple (Malus domestica) and sweet cherry (Prunus avium) agroecosystems. Front. Plant Sci. 2024, 15, 1335281, . [CrossRef]
- Chandel V, Rana T, Handa A, Thakur PD, Hallan V, Zaidi AA. Incidence of Prunus necrotic ringspot virus on Malus domestica in India. J. Phytopath., 2008, 156, 382-384.
- Hu, G.J.; Dong, Y.F.; Zhang, Z.P.; Fan, X.D.; Ren, F.; Li, Z.N.; Zhou, J. First Report of Prunus necrotic ringspot virus Infection of Apple in China. Plant Dis. 2016, 100, 1955, . [CrossRef]
- Kinoti, W.M.; Constable, F.E.; Nancarrow, N.; Plummer, K.M.; Rodoni, B. Generic Amplicon Deep Sequencing to Determine Ilarvirus Species Diversity in Australian Prunus. Front. Microbiol. 2017, 8, 1219, . [CrossRef]
- Çelik, A.; Ertunç, F. First report of prunus necrotic ringspot virus infecting apple in Turkey. J. Plant Pathol. 2019, 101, 1227–1227, . [CrossRef]
- Xiao, H.; Hao, W.; Storoschuk, G.; MacDonald, J.L.; Sanfaçon, H. Characterizing the Virome of Apple Orchards Affected by Rapid Decline in the Okanagan and Similkameen Valleys of British Columbia (Canada). Pathogens 2022, 11, 1231, . [CrossRef]
- Kozieł, E.; Bujarski, J.J.; Otulak, K. Molecular Biology of Prune Dwarf Virus—A Lesser Known Member of the Bromoviridae but a Vital Component in the Dynamic Virus–Host Cell Interaction Network. Int. J. Mol. Sci. 2017, 18, 2733, . [CrossRef]
- Kesanakurti, P.; Belton, M.; Saeed, H.; Rast, H.; Boyes, I.; Rott, M. Screening for plant viruses by next generation sequencing using a modified double strand RNA extraction protocol with an internal amplification control. J. Virol. Methods 2016, 236, 35–40, . [CrossRef]
- Bag, S.; Al Rwahnih, M.; Li, A.; Gonzalez, A.; Rowhani, A.; Uyemoto, J.K.; Sudarshana, M.R. Detection of a New Luteovirus in Imported Nectarine Trees: A Case Study to Propose Adoption of Metagenomics in Post-Entry Quarantine. Phytopathology® 2015, 105, 840–846, . [CrossRef]
- Villamor, D.E.V.; Mekuria, T.A.; Pillai, S.S.; Eastwell, K.C.; Ho, T.; Al Rwahnih, M.; Martin, R.R.; Tzanetakis, I.E.; Green, K.J.; Mollov, D.; et al. High-Throughput Sequencing Identifies Novel Viruses in Nectarine: Insights to the Etiology of Stem-Pitting Disease. Phytopathology® 2016, 106, 519–527, . [CrossRef]
- Wright, A.A.; Cross, A.R.; Harper, S.J. A bushel of viruses: Identification of seventeen novel putative viruses by RNA-seq in six apple trees. PLOS ONE 2020, 15, e0227669, . [CrossRef]
- McLeish MJ, Fraile A, Farcia-Arsenal F. Ecological complexity in plant virus host range evolution. Adv. Virus Res., 2018, 101: 293-339.
- McLeish, M.J.; Fraile, A.; García-Arenal, F. Evolution of plant–virus interactions: host range and virus emergence. Curr. Opin. Virol. 2019, 34, 50–55, . [CrossRef]
- Hamelin, F.M.; Allen, L.J.; Prendeville, H.R.; Hajimorad, M.R.; Jeger, M.J. The evolution of plant virus transmission pathways. J. Theor. Biol. 2016, 396, 75–89, . [CrossRef]
- Gallet, R.; Michalakis, Y.; Blanc, S. Vector-transmission of plant viruses and constraints imposed by virus–vector interactions. Curr. Opin. Virol. 2018, 33, 144–150, . [CrossRef]
- McLaughlin, A.A.; Hanley-Bowdoin, L.; Kennedy, G.G.; Jacobson, A.L. Vector acquisition and co-inoculation of two plant viruses influences transmission, infection, and replication in new hosts. Sci. Rep. 2022, 12, 1–13, . [CrossRef]
- Card, S.D.; Pearson, M.N.; Clover, G.R.G. Plant pathogens transmitted by pollen. Australas. Plant Pathol. 2007, 36, 455–461, . [CrossRef]
- Jones RAC. Plant and insect viruses in managed and natural environments: novel and neglected transmission pathways. Adv. Virus Res., 2018, 101: 149-187.
- Amari, K.; Burgos, L.; Pallas, V.; Sanchez-Pina, M.A. Prunus necrotic ringspot virusEarly Invasion and Its Effects on Apricot Pollen Grain Performance. Phytopathology® 2007, 97, 892–899, . [CrossRef]
- Amari, K.; Burgos, L.; Pallás, V.; Sánchez-Pina, M.A. Vertical transmission of Prunus necrotic ringspot virus: hitch-hiking from gametes to seedling. J. Gen. Virol. 2009, 90, 1767–1774, . [CrossRef]
- Isogai, M.; Yoshida, T.; Nakanowatari, C.; Yoshikawa, N. Penetration of pollen tubes with accumulated Raspberry bushy dwarf virus into stigmas is involved in initial infection of maternal tissue and horizontal transmission. Virology 2014, 452-453, 247–253, . [CrossRef]
- Shipp, J.; Buitenhuis, R.; Stobbs, L.; Wang, K.; Kim, W.; Ferguson, G. Vectoring of Pepino mosaic virus by bumble-bees in tomato greenhouses. Ann. Appl. Biol. 2008, 153, 149–155, . [CrossRef]
- Levitzky, N.; Smith, E.; Lachman, O.; Luria, N.; Mizrahi, Y.; Bakelman, H.; Sela, N.; Laskar, O.; Milrot, E.; Dombrovsky, A. The bumblebee Bombus terrestris carries a primary inoculum of Tomato brown rugose fruit virus contributing to disease spread in tomatoes. PLOS ONE 2019, 14, e0210871, . [CrossRef]
- Tayal, M.; Wilson, C.; Cieniewicz, E. Bees and thrips carry virus-positive pollen in peach orchards in South Carolina, United States. J. Econ. Èntomol. 2023, 116, 1091–1101, . [CrossRef]
- Oliver, J.E.; Freer, J.; Andersen, R.L.; Cox, K.D.; Robinson, T.L.; Fuchs, M. Genetic Diversity of Prunus necrotic ringspot virus Isolates Within a Cherry Orchard in New York. Plant Dis. 2009, 93, 599–606, . [CrossRef]
- Abdallah, D.; Baraket, G.; Perez, V.; Hannachi, A.S.; Hormaza, J.I. Self-compatibility in peach [Prunus persica (L.) Batsch]: patterns of diversity surrounding the S-locus and analysis of SFB alleles. Hortic. Res. 2020, 7, 170, . [CrossRef]
- Piri S, Kiani E, Sedaghathoor S. Study on fruitset and pollen-compatibility status in sweet cherry (Prunus avium L.) cultivars. Erwebs-Obstbau, 2022, 64: 165-170.
- Delaplane K S, Mayer D F. Crop Pollination by Bees. CABI Publishing: New York. 2000.
- Beekman, M.; Ratnieks, F.L.W. Long-range foraging by the honey-bee, Apis mellifera L.. Funct. Ecol. 2000, 14, 490–496, . [CrossRef]
- Steffan-Dewenter, I.; Kuhn, A. Honeybee foraging in differentially structured landscapes. Proc. R. Soc. B: Biol. Sci. 2003, 270, 569–575, . [CrossRef]
- Roberts, J.M.K.; Ireland, K.B.; Tay, W.T.; Paini, D. Honey bee-assisted surveillance for early plant virus detection. Ann. Appl. Biol. 2018, 173, 285–293, . [CrossRef]
- Lee, E.; Vansia, R.; Phelan, J.; Lofano, A.; Smith, A.; Wang, A.; Bilodeau, G.J.; Pernal, S.F.; Guarna, M.M.; Rott, M.; et al. Area Wide Monitoring of Plant and Honey Bee (Apis mellifera) Viruses in Blueberry (Vaccinium corymbosum) Agroecosystems Facilitated by Honey Bee Pollination. Viruses 2023, 15, 1209, . [CrossRef]
- Cunningham, M.M.; Tran, L.; McKee, C.G.; Polo, R.O.; Newman, T.; Lansing, L.; Griffiths, J.S.; Bilodeau, G.J.; Rott, M.; Guarna, M.M. Honey bees as biomonitors of environmental contaminants, pathogens, and climate change. Ecol. Indic. 2021, 134, 108457, . [CrossRef]
- Hong, C.; Manimaran, S.; Shen, Y.; Perez-Rogers, J.F.; Byrd, A.L.; Castro-Nallar, E.; A Crandall, K.; Johnson, W.E. PathoScope 2.0: a complete computational framework for strain identification in environmental or clinical sequencing samples. Microbiome 2014, 2, 33–33, . [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359, . [CrossRef]
- Xiang, Y.; Belton, M.; Saeed, H.; Hayes, S.; Lawrence, T.; Birch, C.; Bhagwat, B.; et al. Application of Next Generation Sequencing for Diagnostic Testing of Tree Fruit Viruses and Viroids. Plant Dis. 2017, 101, 1489–1499, . [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027, . [CrossRef]
- Gao, R.; Xu, Y.; Candresse, T.; He, Z.; Li, S.; Ma, Y.; Lu, M. Further insight into genetic variation and haplotype diversity of Cherry virus A from China. PLOS ONE 2017, 12, e0186273, . [CrossRef]
- Simkovich, A.J.; Li, Y.; Kohalmi, S.E.; Griffiths, J.S.; Wang, A. Molecular Identification of Prune Dwarf Virus (PDV) Infecting Sweet Cherry in Canada and Development of a PDV Full-Length Infectious cDNA Clone. Viruses 2021, 13, 2025, . [CrossRef]
- Kinoti, W.M.; Constable, F.E.; Nancarrow, N.; Plummer, K.M.; Rodoni, B. The Incidence and Genetic Diversity of Apple Mosaic Virus (ApMV) and Prune Dwarf Virus (PDV) in Prunus Species in Australia. Viruses 2018, 10, 136, . [CrossRef]
- Kamenova, I.; Borisova, A.; Popov, A. Incidence and genetic diversity of Prune dwarf virus in sweet and sour cherry in Bulgaria. Biotechnol. Biotechnol. Equip. 2019, 33, 980–987, . [CrossRef]
- Glassa M, Betinova E, Kudela O, Subr Z. Biological and molecular characterization of Prunus necrotic ringspot virus isolates and possible approaches to their phylogenetic typing. Ann. App. Biol., 2002, 140, 215-329.
- Free, J.B. The Flower Constancy of Honeybees. J. Anim. Ecol. 1963, 32, 119–131, . [CrossRef]
- Danner, N.; Molitor, A.M.; Schiele, S.; Härtel, S.; Steffan-Dewenter, I. Season and landscape composition affect pollen foraging distances and habitat use of honey bees. Ecol. Appl. 2016, 26, 1920–1929, . [CrossRef]
- Jones, L.; Lowe, A.; Ford, C.R.; Christie, L.; Creer, S.; de Vere, N. Temporal Patterns of Honeybee Foraging in a Diverse Floral Landscape Revealed Using Pollen DNA Metabarcoding of Honey. Integr. Comp. Biol. 2022, 62, 199–210, . [CrossRef]
- Lowe, A.; Jones, L.; Brennan, G.; Creer, S.; Christie, L.; de Vere, N. Temporal change in floral availability leads to periods of resource limitation and affects diet specificity in a generalist pollinator. Mol. Ecol. 2022, 32, 6363–6376, . [CrossRef]
- Tremblay, .D.; Duceppe, M.; Thurston, G.B.; Gagnon, M.; Côté, M.; Bilodeau, G.J. High-resolution biomonitoring of plant pathogens and plant species using metabarcoding of pollen pellet contents collected from a honey bee hive. Environ. DNA 2019, 1, 155–175, . [CrossRef]
- McKinnon, A.C.; Collins, L.; Wood, J.L.; Murphy, N.; Franks, A.E.; Steinbauer, M.J. Precision Monitoring of Honey Bee (Hymenoptera: Apidae) Activity and Pollen Diversity during Pollination to Evaluate Colony Health. Insects 2023, 14, 95, . [CrossRef]
- Leponiemi, M.; Freitak, D.; Moreno-Torres, M.; Pferschy-Wenzig, E.-M.; Becker-Scarpitta, A.; Tiusanen, M.; Vesterinen, E.J.; Wirta, H. Honeybees’ foraging choices for nectar and pollen revealed by DNA metabarcoding. Sci. Rep. 2023, 13, 1–15, . [CrossRef]
- Leontidou, K.; Vokou, D.; Sandionigi, A.; Bruno, A.; Lazarina, M.; De Groeve, J.; Li, M.; Varotto, C.; Girardi, M.; Casiraghi, M.; et al. Plant biodiversity assessment through pollen DNA metabarcoding in Natura 2000 habitats (Italian Alps). Sci. Rep. 2021, 11, 1–12, . [CrossRef]
- Hasiow-Jaroszewska B, Boezen D, Zwart MP. Metagenomic studies of viruses in weeds and wild plants: a powerful approach to characterize variable virus communities. Viruses, 2021, 13: 1939.
- Bell, K.L.; de Vere, N.; Keller, A.; Richardson, R.T.; Gous, A.; Burgess, K.S.; Brosi, B.J.; Van der Bank, M.; Adamowicz, S.J.; Chain, F.J.; et al. Pollen DNA barcoding: current applications and future prospects. Genome 2016, 59, 629–640, . [CrossRef]
- Milla, L.; Schmidt-Lebuhn, A.; Bovill, J.; Encinas-Viso, F. Monitoring of honey bee floral resources with pollen DNA metabarcoding as a complementary tool to vegetation surveys. Ecol. Solutions Évid. 2022, 3, e12120, . [CrossRef]
- Hoffmann, V.; Paul, B.; Falade, T.; Moodley, A.; Ramankutty, N.; Olawoye, J.; Djouaka, R.; Lekei, E.; de Haan, N.; Ballantyne, P.; et al. A one health approach to plant health. CABI Agric. Biosci. 2022, 3, 1–7, . [CrossRef]
- Fetters, A.M.; Ashman, T. The pollen virome: A review of pollen-associated viruses and consequences for plants and their interactions with pollinators. Am. J. Bot. 2023, 110, e16144, . [CrossRef]
- Pallas V, Aparicio F, Herranz MC, Sanches-Navarro JA, Scott SW. The molecular biology of ilarviruses. Adv. Virus Res., 2013, 87: 139-81.
- Pallas, V.; Aparicio, F.; Herranz, M.C.; Amari, K.; Sanchez-Pina, M.A.; Myrta, A.; Sanchez-Navarro, J.A. Ilarviruses of Prunus spp.: A Continued Concern for Fruit Trees. Phytopathology® 2012, 102, 1108–1120, . [CrossRef]
- Tzanetakis IE, Gergerich RC, Martin RR. A new Ilarvirus found in rose. Plant Pathol., 2006, 55(4): 568–568.
- Tzanetakis IE, Postman JD,, Martin RR. First report of blackberry chlorotic ringspot virus in Rubus sp. in the United States. Plant dis., 2007, 91: 463.
- Poudel, B.; Ho, T.; Laney, A.; Khadgi, A.; Tzanetakis, I.E. Epidemiology of Blackberry chlorotic ringspot virus. Plant Dis. 2014, 98, 547–550, . [CrossRef]
- Bratsch, S.A.; Grinstead, S.; Creswell, T.C.; Ruhl, G.E.; Mollov, D. Characterization of Tomato Necrotic Spot Virus, a Subgroup 1 Ilarvirus Causing Necrotic Foliar, Stem, and Fruit Symptoms in Tomatoes in the United States. Plant Dis. 2019, 103, 1391–1396, . [CrossRef]
- Bonilla, F.O.R.; Cieniewicz, E. Distribution and Diversity of Prunus Necrotic Ringspot Virus, Prune Dwarf Virus, and Peach Latent Mosaic Viroid in Wild Prunus spp. in South Carolina and Georgia. Phytofrontiers™ 2022, 2, 363–370, . [CrossRef]
- Fuchs, M.; Hily, J.-M.; Petrzik, K.; Sanfaçon, H.; Thompson, J.R.; van der Vlugt, R.; Wetzel, T.; ICTV Report Consortium ICTV Virus Taxonomy Profile: Secoviridae 2022. J. Gen. Virol. 2022, 103, 001807, . [CrossRef]
- Thompson JR, Dasgupta I, Fuchs M, Iwanami T, Karasev AV, Petrzik K, Sanfacon H, Tzanetakis I, van der Vlugt R, Wetzel T, Yoshikana N, and ICTV Report Consortium. ICTV virus taxonomy profile: Secoviridae. J. Gen. Virol., 2017, 98: 529-531.
- Koloniuk, I.; Sarkisova, T.; Petrzik, K.; Lenz, O.; Přibylová, J.; Fránová, J.; Špak, J.; Lotos, L.; Beta, C.; Katsiani, A.; et al. Variability Studies of Two Prunus-Infecting Fabaviruses with the Aid of High-Throughput Sequencing. Viruses 2018, 10, 204, . [CrossRef]
- Sasaya, T.; Palacios, G.; Briese, T.; Di Serio, F.; Groschup, M.H.; Neriya, Y.; Song, J.-W.; Tomitaka, Y. ICTV Virus Taxonomy Profile: Phenuiviridae 2023. J. Gen. Virol. 2023, 104, 001893, . [CrossRef]
- Wunsch, A.; Hoff, B.; Sazo, M.M.; van Zoeren, J.; Lamour, K.H.; Hurtado-Gonzales, O.P.; Fuchs, M. Viruses of Apple Are Seedborne but Likely Not Vertically Transmitted. Viruses 2024, 16, 95, . [CrossRef]
- Simkovich, A.; Kohalmi, S.E.; Wang, A. First Report of Little Cherry Virus 1 Infecting Sweet Cherry in Ontario, Canada. Plant Dis. 2021, 105, 4173–4173, . [CrossRef]
- Kesanakurti, P.; Belton, M.; Saeed, H.; Rast, H.; Boyes, I.; Rott, M. Comparative analysis of cherry virus A genome sequences assembled from deep sequencing data. Arch. Virol. 2017, 162, 2821–2828, . [CrossRef]
- Beaver-Kanuya, E.; Harper, S. Detection and quantification of four viruses in Prunus pollen: Implications for biosecurity. J. Virol. Methods 2019, 271, 113673, . [CrossRef]
- Cui H, Hong N, Wang G, Wang A. Detection and genetic diversity of Prunus necrotic ringspot virus in the Niagara fruit belt, Canada. Can. J. plant pathol., 2012, 34, 104-113.
- Kamenova, I.; Borisova, A. Biological and molecular characterization of Prunus necrotic ringspot virus isolates from sweet and sour cherry. Biotechnol. Biotechnol. Equip. 2021, 35, 567–575, . [CrossRef]
Table 1.
Summary of varieties, rootstocks, total number, and age of Prunus and Malus trees grown on Jordan Farm.
Table 1.
Summary of varieties, rootstocks, total number, and age of Prunus and Malus trees grown on Jordan Farm.
| Crop | Number of Plots | Total Area (ha) | Variety | Rootstock | Total number of trees | Age of trees (years) |
|---|---|---|---|---|---|---|
| Apricot | 1 | 0.57 | Haroblush, Harlayne | Krymsk 1, Krymsk 86, Krymsk 99 | 208 | 2 |
| Cherry | 3 | 0.96 | Vista, Vogue, Hedelfingen, Tehranivee, Stella, Vandalay, Montmorency | Mazzard, Mahaleb, Gisela 5 | 317 | 14-50 |
| Peach | 10 | 3.849 | Redhaven, Vivid, Nectarine, Harrow Diamond, Cresthaven | Halford, Bailey, TLC, Bailey Field | 2832 | 2-17 |
| Apple | 11 | 3.83 | Brookfield Gala, Empire, Red Fuji, Ambrosia, Honey Crisp, Silken, Morspur McIntosh, Golden Delicious, Royal Gala, Courtland, Delicious Red Chief, McIntosh Marshall, Jonagold, McIntosh | G41, M26, BUD 9, M9, Mark | 3123 | 3-26 |
Table 2.
Plant virus detections in Bee and Pollen samples from AAFC Jordan farm during apricot, cherry, peach, and apple blooms.
Table 2.
Plant virus detections in Bee and Pollen samples from AAFC Jordan farm during apricot, cherry, peach, and apple blooms.
| Virus | Genus | Frequency of detection (%) | Total detections(n) | Average frequency of detection (%) | Average Genome Coverage (%) | Average VRPM | |||||||||||
| Apricot | Cherry | Peach | Apple | ||||||||||||||
| Forager | Hive | Pollen | Forager | Hive | Pollen | Forager | Hive | Pollen | Forager | Hive | Pollen | ||||||
| n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | n=3 | ||||||
| Prune dwarf virus | Ilarvirus | 33 | 33 | 100 | 67 | 100 | 100 | 33 | 100 | 67 | 33 | 100 | 23 | 64 | 62 | 32548 | |
| Cherry virus A | Capillovirus | 67 | 67 | 100 | 67 | 100 | 100 | 33 | 33 | 33 | 100 | 21 | 58 | 70 | 5042 | ||
| Prunus necrotic ringspot virus | Ilarvirus | 67 | 33 | 67 | 100 | 100 | 100 | 67 | 33 | 33 | 100 | 21 | 58 | 54 | 26401 | ||
| Prunus virus F | Fabavirus | 33 | 33 | 67 | 67 | 100 | 100 | 33 | 100 | 16 | 44 | 63 | 2920 | ||||
| Tobacco ringspot virus | Nepovirus | 33 | 33 | 67 | 67 | 67 | 100 | 100 | 67 | 16 | 44 | 47 | 11657 | ||||
| Tomato ringspot virus | Nepovirus | 67 | 67 | 100 | 33 | 33 | 33 | 100 | 67 | 33 | 16 | 44 | 36 | 4000 | |||
| Brome mosaic virus | Bromovirus | 67 | 67 | 100 | 33 | 33 | 67 | 33 | 67 | 33 | 15 | 42 | 29 | 1743 | |||
| Peach latent mosaic viroid | Pelamoviroid | 67 | 67 | 67 | 100 | 67 | 11 | 31 | 37 | 0 | |||||||
| Apple mosaic virus | Ilarvirus | 67 | 67 | 4 | 11 | 33 | 135 | ||||||||||
| White clover mosaic virus | Potexvirus | 33 | 33 | 67 | 4 | 11 | 13 | 343 | |||||||||
| Blackberry chlorotic ringspot virus | Ilarvirus | 33 | 33 | 33 | 3 | 8 | 34 | 5762 | |||||||||
| Pelargonium zonate spot virus | Anulavirus | 33 | 67 | 3 | 8 | 23 | 1449 | ||||||||||
| Tobacco streak virus | Ilarvirus | 67 | 33 | 3 | 8 | 16 | 117 | ||||||||||
| Gaillardia latent virus | Carlavirus | 33 | 33 | 2 | 6 | 18 | 4 | ||||||||||
| Raphanus sativus cryptic virus 3 | Unclassified Partitiviridae | 33 | 33 | 2 | 6 | 47 | 53 | ||||||||||
| Tomato necrotic spot virus | Ilarvirus | 33 | 33 | 2 | 6 | 40 | 296 | ||||||||||
| Apple hammerhead viroid | Pelamoviroid | 33 | 1 | 3 | 20 | 0 | |||||||||||
| Apple stem grooving virus | Capillovirus | 33 | 1 | 3 | 66 | 279 | |||||||||||
| Citrus concave gum associated virus | Coguvirus | 33 | 1 | 3 | 67 | 1408 | |||||||||||
| Citrus virus A | Coguvirus | 33 | 1 | 3 | 28 | 39 | |||||||||||
| Peanut stunt virus | Cucumovirus | 33 | 1 | 3 | 10 | 0 | |||||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



