
Submitted:
31 May 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
Klebsiella pneumoniae, a member of the ESKAPE pathogen group, is a prominent cause of hospital-acquired infections. The WHO recognized carbapenem-resistant K. pneumoniae as a critical-one priority pathogen. These resilient superbugs have the ability to form biofilms and present a significant global threat. In the present study, we isolated and characterized a bacteriophage SAKp02, from hospital sewage, infectious to carbapenem-resistant K. pneumoniae patient isolates. SAKp02 could infect 43 of 72 clinical isolates, indicating a broad host spectrum. Whole genome analysis classified SAKp02 within the family Casjensviridae, with a 59,343 bp genome encoding 82 ORFs. Comparative genomic analysis revealed significant differences between SAKp02 and its closest viruses, indicating a distinct genetic makeup positioning it as a novel phage strain within the lineage. The SAKp02 genome comprises bacteriolytic enzymes, including holin, endolysin, and phage depolymerase, crucial for bacterial lysis and biofilm disruption. It reduced biofilm biomass by over threefold compared to the control and eradicated 99% of viable cells within a 4-hour treatment period. Scanning electron microscopy corroborated the ability of the phage to dismantle biofilm matrices and lyse bacterial cells. Safe and effective treatments are warranted, and hence, the fully characterized lytic phages with therapeutic potential against drug-resistant clinical isolates of bacteria are needed. Our study is the first to report the antibacterial and antibiofilm activity of Casjensviridae phages, and our discovery of a novel K. pneumoniae phage broadens the arsenal against the bacteria.
Keywords:
antimicrobial resistance
; biofilm bacteriophage
; K. pneumoniae
; alternative to antibiotics
; antibiofilm strategy
1. Introduction
Klebsiella pneumoniae is an opportunistic oropharyngeal pathogen associated with nosocomial and community-based infections [1,2,3]. It belongs to the Enterobacteriaceae family, and WHO has listed carbapenem-resistant K. pneumoniae as a priority one critical pathogen group [4]. Pathogenic K. pneumoniae can be categorized into three groups: healthcare-associated opportunistic or classic K. pneumoniae (cKp), a community-acquired infection causing hypervirulent K. pneumoniae (hyKp) and the global threat superbugs, i.e., multi-drug resistant (MDR) K. pneumoniae [5]. Studies showed that the MDR strains possess a greater risk of infection than the hyKp strains, as the MDR strains are more likely to acquire virulence compared to the propensity of hyKp strains to acquire drug resistance genes [6,7]. There are several contributory factors attributing to the pathogenicity of K pneumoniae, such as capsule lipopolysaccharides and polysaccharides that help in evading the host immune system by hiding the bacteria from phagocytosis; fimbrial adhesin proteins that help in the adhesion of the bacterial cell to host tissues and importantly, the ability to form biofilms on tissue or device surfaces [8,9,10]. Klebsiella pneumoniae demonstrates remarkable proficiency in biofilm formation, readily adhering to implanted medical devices and various host tissues [11,12]. Moreover, a correlation has been identified between the presence of antibiotic resistance genes and the biofilm formation ability of the bacterial strains. A study conducted by Yang et al. underscored a significant correlation between the presence of extended-spectrum beta-lactamase (ESBL) in clinical K. pneumoniae isolates and their propensity for biofilm formation [13]. Further exacerbating the clinical challenge, research by Anderl et al. revealed the inefficacy of antibiotics in disrupting K. pneumoniae biofilms. Even following prolonged exposure, ampicillin, and ciprofloxacin exhibited negligible impact on biofilm integrity [14]. Particularly concerning was the inability of ampicillin to penetrate biofilms harbouring beta-lactamase-producing K. pneumoniae strains. The escalating emergence of antibiotic resistance coupled with the persistent resistance of bacterial biofilms to conventional antibiotic treatment underscores the urgent need for alternative therapeutic strategies. With the dearth of novel antibiotics in development pipelines, researchers are compelled to explore non-traditional avenues to combat K. pneumoniae infections effectively.
Bacteriophage, being the nature-built dedicated scavenger of bacteria, has gained a resurgent interest as an alternative approach to control bacterial infection. Phage discovery predates antibiotics by 13 years; however, antibiotics garnered widespread attention thereafter, owing to their potent killing activity and the subsequent surge in new antibiotic development. Over the past century, escalating drug resistance-associated disease burdens have spurred intensified research into phages as therapeutic agents. Bacteriophages emerge as indispensable antibacterial tools due to their ability to co-evolve with bacteria, ensuring that bacteria cannot evade phage, a limitation often encountered with antibiotics [15]. Studies indicate that developing resistance to phages may incur fitness costs to bacteria and reverse antibiotic resistance, potentially prolonging the efficacy of existing antibiotics [16,17]. Furthermore, bacteriophages possess inherent genetic mechanisms enabling them to hijack bacterial hosts, thus exhibiting innate bacteriolytic capabilities crucial for parasitizing bacteria and propagating within their populations. Moreover, bacteriophages exhibit the ability to penetrate and disrupt biofilms—a feature extensively documented in various studies [18]. However, several significant challenges confront phage therapy, foremost among them being the high specificity of phages to particular bacterial strains. Continuous efforts in isolating and characterizing novel phages remain imperative. Additionally, numerous parameters influencing the interaction between phages and bacteria, as well as factors contributing to the resurgence of phage resistance, necessitate ongoing scientific inquiry for a comprehensive understanding of these dynamics. In this study, we successfully isolated a lytic bacteriophage capable of targeting a multidrug-resistant clinical strain of K. pneumoniae from hospital sewage water. We extensively examined its bacteriolytic activity, host range, and ability to combat biofilms. The isolated phage was thoroughly characterized, including its efficiency in host cell attachment, the time a single phage takes to complete an infection (latency period), the number of progenies produced during the replication cycle (burst size), and the stability of the isolated phage. Additionally, we performed transmission electron microscopy to gain insight into the phage's morphology and conducted a whole genome sequence analysis to understand its genetic composition. Moreover, an alternative to the antibiotic treatment approach is becoming a priority in combating drug-resistant bacterial infections, necessitating fully characterized lytic phages with therapeutic potential. For the first time, our study highlights the antibacterial and antibiofilm properties of Casjensviridae phages, showcasing their capability against resistant clinical isolates. Notably, our discovery of a novel Klebsiella pneumoniae phage significantly expands the arsenal available to target these extremely drug-resistant bacteria,
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
The extensively drug-resistant (XDR) Klebsiella pneumoniae isolates procured from Madras Medical Mission Hospital (MMMH) Chennai, India, were cultivated at 37 °C in Luria-Bertani Broth (LB, Himedia India (M1245)) under shaking at 150 rpm until otherwise mentioned. To determine viable cell counts, serially diluted cultures were plated on sterile LB agar plates and incubated overnight at 37 °C. The colony forming units/mL were estimated. All the experiments were done in triplicates.
2.2. Isolation and Purification of Bacteriophage
Bacteriophage was isolated from the sewage water collected from the Madras Medical Mission Hospital (MMMH) Chennai, India, as described by Oliveira et al. [19], with slight modifications. Briefly, 5 ml LB was mixed with an equal volume of hospital sewage and 5 ml of mid-log phase XDR K. pneumoniae clinical isolate B3768 strain, grown to OD600 ~ 0.3 - 0.6. The mixture was incubated overnight at 37 °C shaking at 150 rpm. The mixture was centrifuged at 12,000× g for 10 minutes at room temperature (RT). Spot test was done for preliminary confirmation of the presence of lytic bacteriophage in the supernatant, where 10 µl of the supernatant was dropped on the top of the lawn of the bacterial isolate of interest and allowed to dry. The plate was incubated at 37 °C overnight. A clear zone of lysis was considered as positive bacteriolytic activity. The purification of a single phage was carried out using a double agar overlay technique by Kropinski et al. [20] with slight modifications. Briefly, a tenfold serial dilution of the supernatant was prepared. 100 µl of diluted supernatant was inoculated in 500 µl of an overnight culture of K. pneumoniae, and the whole mixture was added into 3 ml of 0.5% soft agar (Himedia India GRM026) and poured over 1.5% nutrient agar (NA) plate and incubated overnight at 37 °C. The next day, a single plaque was picked and inoculated in early log phase culture and kept for overnight incubation. The suspension was centrifuged at 12,000× g for 10 minutes at RT, and a double agar overlay technique was performed with the supernatant. The procedure was repeated thrice to get a single purified phage. Finally, the supernatant was treated with chloroform in 1: 10 ratio for 15 minutes, after chloroform treatment the suspension was centrifuged and filtered through a 0.22 μm syringe driven filter (Himedia India TPP-99722) and stored at 4 °C
2.3. Host Adsorption Assay
Adsorption assays were performed according to the procedure described by Avni et al. [21] with slight modifications. Briefly, 6.0 × 107 cells/ml of K. pneumoniae B3768 strain was mixed with 6.0 × 106 SAKp02 phage particles with a Multiplicity of Infection (MOI) of 0.1 in different microcentrifuge tubes and incubated at 37 °C in static condition for 30 minutes. At 5-minute intervals, microcentrifuge tubes were removed and centrifuged for 4 minutes at 12,000× g, and unabsorbed phages present in the supernatant were determined by the double agar overlay technique.
2.4. Determination of Phage Life Cycle by One Step Growth Curve
A one-step growth curve to determine the latency and burst size of the phage was performed as described by Amarillas et al. [22] with slight modifications. K. pneumoniae B3768 strain was grown to a mid-exponential phase (OD: 0.6). Bacteriophage SAKp02 was mixed with an MOI 0.1 and kept for 10 minutes (according to previously evaluated adsorption time) at 37 °C in static condition. After incubation, the phage-bacteria suspension was centrifuged at 12,000× g for 10 minutes, and the supernatant containing unabsorbed phages was discarded. The pellet was resuspended in 10 ml of LB and incubated at shaking for 150 rpm for 2 hours. 100 µl Samples were collected at every 20-minute interval, and titration was done to determine the phage number.
2.5. Host Range of the Bacteriophage
Spot test was done to evaluate the host spectrum of SAKp02. Lawn was prepared on the LB NA plates with the bacterial strains using mid-log phase cultures. Subsequently, 2.5 µl of purified phage SAKp02 was spotted on the lawn. The spots were allowed to dry, and the plates were incubated overnight at 37 °C. The appearance of clear lysis zones indicated susceptible bacterial strains, whereas the absence of plaques indicated phage-immune strains.
2.6. Assessment of Bacteriolytic Activity
A bacterial challenge test was performed to evaluate bacteriolytic activity with different MOIs. Briefly, the log phase culture of K. pneumoniae B3768 strain was grown up to exponential phase (OD: 0.600) (Bacterial cell number against particular OD was predetermined to adjust the MOI). The culture was centrifuged at 12,000× g for 10 minutes, and the supernatant was discarded. The pallet was resuspended in fresh medium and aliquoted into microcentrifuge tubes. Bacteriophage was added according to MOI 0.5, 1, 10, and 100. 200 µl of phage-bacteria mixture of each MOI was dispensed in 96 well microtiter plates in triplicates. The plates were placed in a BioteK Epoch-2 microtiter plate reader (Agilent Technologies, USA) for 12 hours. OD of each well was taken at 20-minute intervals.
2.7. Phage Stability
Phage stability was determined according to the method described by Sarkar et al. with slight modification [23]. Solvent Stability: 1 × 109/ml Phage particles were mixed with PBS, chloroform, and 0.9% saline in equal proportion and incubated at 37 °C, 150 rpm for 1 hour. Post incubation, the suspension was centrifuged at 12,000× g for 10 minutes, and the supernatant was used for phage titration. Thermal Stability: The exact amount of phage was kept at 25, 37, 55, 65, and 75 °C for 1 hour. Titration was done post-incubation to assess the stability. pH Stability: The pH of LB broth was adjusted to 2 to 14. Phage lysate with a phage number 1 × 109/ml was mixed equally and kept for one hour at 37 °C. Titration was done post-incubation to assess the pH stability.
2.8. Transmission Electron Microscopy
On a clean piece of Parafilm, 100 µl of SAKp02 suspension with a concentration of ~109 PFU/ml was dispensed. Prior to this, the Parafilm was cleaned by wiping it with 70% ethanol. Next, a 300 mesh Carbon Coated Copper grid (Electron Microscopy Sciences, USA, EMS200-Cu) was gently positioned on the phage suspension sample and left undisturbed for 25 to 30 minutes. After the designated time, the grid was carefully lifted off from the samples. Excess liquid surrounding the grid was removed by gently dabbing it with filter paper, ensuring not to disturb the mesh. Subsequently, the grids were flipped over, and the sample side was placed down onto a droplet of 2% aqueous (wt/vol) phosphotungstic acid (Himedia, India, GRM398) solution (pH 6.8) for 3 minutes. To remove excess liquid, the grids were blotted dry using filter paper. They were then placed over another piece of filter paper and left to air-dry under a Petri plate, positioned upside down to prevent contact with the moist surface. After drying, the sample was imaged under Thermo Scientific™ Talos L120C TEM (20-120 kV) at All India Institute of Medical Sciences, New Delhi, India.
2.9. DNA Extraction
50 ml of crude bacteriophage sample was sent to Redcliffe Lifetech Pvt. Ltd. Noida, India Ltd for DNA extraction and genome sequence by Illumina Hiseq platform. Phage DNA was exposed to RNase at a concentration of 3 μg/mL and DNase at a concentration of 1 μg/mL. This treatment occurred overnight at a temperature of 37 °C. Subsequently, the phage suspension was subjected to incubation at 80 °C for 15 minutes to deactivate the enzymes. Following the deactivation step, the purified phages were treated with proteinase K at a concentration of 50 μg/mL. This treatment took place at a temperature of 56 °C for 1 hour. Additionally, SDS was present in the treatment at a concentration of 0.5%. Then it was mixed with an equal volume of a phenol/chloroform/isoamyl alcohol solution (25:24:1 ratio). The resulting mixture was then centrifuged at 13,000× g for 5 minutes at 4 °C. After centrifugation, the supernatant was mixed with an equal volume of isopropanol and stored at -20 °C for 1 hour. Following this, another round of centrifugation took place. To wash the pellet, 1 mL of 75% ethanol was added and allowed to sit at room temperature for 1 minute. Subsequently, the mixture underwent centrifugation at 12,000× g for 10 minutes at 4 °C. The resulting pellets were air-dried and then dissolved in 30 µL of ddH2O. Finally, they were stored at -20 °C for future use. Samples Concentration and Quality was estimated using Qubit 4.0 Fluorometer. All extracted DNA Samples were run & checked on 0.8% Agarose Gel made in 1× TAE Buffer.
2.10. Bacteriophage Whole Genome Sequencing, Annotation and Bioinformatics
Phage whole genome sequencing was performed in the Illumina Hiseq platform. Denovo genome assembly was done by the St. Petersburg genome assembler (SPAdes). It is an assembly toolkit containing various assembly pipelines. SPAdes support paired-end reads, mate-pairs, and unpaired reads. It can take as input several paired-end and mate-pair libraries simultaneously. SPAdes was initially designed for small genomes. It was tested on bacterial (both single-cell MDA and standard isolates), fungal and other small genomes. We assembled the data and extracted the phage sequence using phaster and annotated the same using Prokka (1.14.6). (Ref: http://cab.spbu.ru/software/spades/). Quality control was done by fastp (0.20.1) The Fastp tool is designed to provide fast all-in-one pre-processing for FastQ files. This tool is developed in C++ with multithreading supported to afford high performance. Quality control and pre-processing of FASTQ files are essential to providing clean data for downstream analysis (Ref: https://github.com/OpenGene/fastp). The phage sequence was annotated using the prokka tool. Prokka is a command line tool for the rapid annotation of bacterial, archaeal, and viral genomes. Further annotation and Genome Map was constructed using RAST version 2 (https://rast.nmpdr.org/rast.cgi), BLASTp server (https://blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE=Proteins), PHASTER (https://phaster.ca/) PhaGAA (http://phage.xialab.info/predict), genomevx (http://wolfe.ucd.ie/GenomeVx/) and Geneious Prime® 2023.0.4, for t-RNA in the genome was done by tRNAscan-SE v. 2.0 (http://lowelab.ucsc.edu/tRNAscan-SE/). Phylogenetic tree analysis was done in MEGA11 software based on the whole genome sequence as well as the large terminase subunit. EasyFigure (https://mjsull.github.io/EasyFigure/files.html) was used to compare the related viral genomes. An essential criterion for phages to be used as therapeutics that it should not be carrying any drug-resistance genes or virulent factors. We have used ResFinder 4.1 (https://cge.food.dtu.dk/services/ResFinder/) developed by Centre for Genomic Epidemiology to find drug resistance genes in a genome, and presence of virulent factor was examined in Virulence Factor Database (VFDB) (http://www.mgc.ac.cn/VFs/search_VFs.htm).
2.11. Assessment of Anti-Biofilm Activity of SAKp02
The biofilm inhibition assays were conducted according to the method outlined by Chauhan et al. [24] in 2014, with slight modifications. Overnight cultures of the K. pneumoniae B3768 strain were diluted 1:100 in fresh media and inoculated into a 96-well microtiter plate. The plate was then incubated at 37 °C without shaking for 24 hours to allow the formation of mature biofilms. Subsequently, the planktonic bacteria were gently removed by pipetting, and the wells were rinsed three times with PBS. Next, the 24-hour-old mature biofilms were treated with SAKp02. A 100 μL solution of the phage suspension was added to the wells at MOI 1 and left in the wells for specific time intervals (4 hours, 8 hours, and 12 hours) at 37 °C under static conditions. Control wells were left untreated and inoculated with PBS. After the respective treatment period, the phage suspension was removed, and the wells were rinsed three times with PBS. The experiment was conducted in two sets with four replicates each. One set was used for viable cell count, and the other set was used for biomass estimation using crystal violet (CV) staining. For the viable cell count, the wells were vigorously scraped to remove any remaining biofilm, and the resulting mixture was homogenized with PBS. Viable cell count was determined by plating serial dilutions on LB agar medium and incubating the plates at 37 °C. For the crystal violet assay, after washing the wells with PBS following the treatment, the wells were stained with 0.1% CV. After a 15-minute incubation period, the stain was carefully removed, and the wells were rinsed with PBS to remove any excess stain. To dissolve the CV stain adhered to the wells, a solution of acetone and ethanol was added. The solution was mixed thoroughly by pipetting and then transferred to a separate flat-bottom microtiter plate for absorbance measurement at a wavelength of 580 nm.
2.12. Scanning Electron Microscopy
Scanning Electron Microscopy (SEM) of the biofilm was performed as described by Chauhan et al. 2016 with slight modification [25]. Briefly, one cm2 glass coverslips were placed in individual wells of 6-well plates, and an O/N culture of K. pneumoniae B3768 diluted to 1:100 was inoculated in 6-well plates to allow the biofilms to form on the coverslips. The plate was incubated at 37 °C under static conditions for 24 hours. Post incubation, the coverslips were washed thrice with PBS to remove any loosely adhered bacteria. The biofilms were treated with phage SAKp02 and incubated for a required time at 37 °C under static conditions. The untreated wells were taken as controls. Coverslips from the wells were taken out and dipped in the fixative solution (2.5% glutaraldehyde solution in PBS) overnight. After overnight fixation, the slides were washed with PBS and dehydrated with 10, 20, 30, 40, 50, 60, 70, 80, 90, and 100% ethanol for 10 minutes each. The slides were air-dried, gold-sputtered, and observed under Sigma 300, Carl Zeiss Field Emission Scanning Electron Microscope, Tripura University, Agartala, India.
2.13. Statistical Analysis
The results presented in this study are based on a minimum of three independent experiments, ensuring reproducibility. To determine statistical differences, an unpaired t test was conducted using GraphPad Prism version 8.0. Statistical significance was defined as p-values less than 0.05.
3. Results
3.1. Phage SAKp02 Forms Depolymerase-Induced Bull’s Eye Plaques with a Short Adsorption Time and a 20-Minute Latency
Bacteriophage SAKp02 was isolated against K. pneumoniae XDR clinical strain B3768 from hospital sewage of MMMH. The phage could produce clear, medium-sized plaque approximately 1 cm in diameter with depolymerase activity. A typical bull’s eye plaque morphology in the bacterial lawn was observed (Figure 1A). TEM analysis revealed that the length of phage SAKp02 is 349 nm with a capsid size of 138 nm. The tail of SAKp02 is 211 nm long, and the phage consists of a neck, a contractile tail, and a central tube (Figure 1B). The adsorption assay clarified the time required for a virus to attach to the host receptor, which is a crucial parameter for analysing the infectivity of the virus. The average adsorption time necessary for a phage to initiate an infection has been reported as 10 to 15 minutes. However, phage SAKp02 showed a relatively shorter adsorption time of 5 minutes (Figure 1C), and 95% of the phages could adsorb to the host. One-step growth curve data indicated that the phage SAKp02 has an eclipse period of approximately 20 minutes and a rise period of 40 minutes with a burst size of approximately 202 pages per bacterial cell (Figure 1D).
3.2. Phage SAKp02 Is Strongly Bacteriolytic with a Broad Host Range
To evaluate the lytic activity of bacteriophage SAKp02, a bacterial challenge test was performed with different MOIs (MOI; 0.001, 0.01, 0.1, 1, 10 & 100). Assessment of bacteriolytic activity is crucial to determine the nature of the phage, whether the phage is lytic or lysogenic. It is reported that the temperate phage enters into lysogeny soon after the initiation of infection and does not restrict the growth of bacteria for longer. Phage SAKp02 could control the bacterial growth in different MOIs for 12 hours (Figure 2A & Figure 2B). Clinical isolates of K. pneumoniae isolated from MMM hospital were used to evaluate the host range of SAKp02, and a total of 72 clinical isolates were tested for the host range. Among the clinical isolates, phage SAKp02 could infect ~60% of clinical isolates with a clear lysis zone (Figure 2C).
3.3. Phage SAKp02 Is Stable at Different pH, Temperature and Solvent
When considering a therapeutic approach, phage stability is a critical concern. Thermal stability, the effect of various pH on the virus, and stability with different solvents are also significant concerns. Hence, the impact of pH, temperature, PBS solution, 0.9% saline solution, chloroform, and ethyl alcohol were assessed. Most of the eukaryotic as well as prokaryotic viruses are most stable in the neutral pH; phage SAKp02 showed a similar pattern, and the most significant decrease in phage titer was observed from pH 12 and in pH 1, 3 and 14, no plaque was observed (Figure 3A). Phage SAKp02 showed stability up to 55 °C. However, a significant decrease in phage titer was observed at 65 °C and 85 °C, and at 100 °C, no plaque was observed (Figure 3B). Phage SAKp02 was adequately stable in PBS, 0.9% saline, and chloroform (Figure 3C).
3.4. SAKp02 Belongs to the Casjensviridae Family of Viruses
The genome of phage SAKp02 is 59,343 base pairs (BP) long with a GC content of 56.1%, comprising 82 Open Reading Frame (ORF) and no tRNA genes (Figure 4A). Out of the total ORFs, 38 show significant homology with functional proteins, leaving 54% of the protein as hypothetical function; this suggests that a significant portion of the predicted proteins still need to be characterized, with their roles and functions in phage biology still being determined (Table 1). The annotated genome map of phage SAKp02 is depicted in Figure 4B. The SAKp02 genome is compact, linear, and has only 6.1% intragenic space. The longest gene in SAKp02 is 4301 nucleotides, with its functional features yet to be identified. The smallest gene is 113 nucleotide long encoding tape measure protein, which facilitates the transit of the viral DNA in host cytoplasm during infection. The whole genome of SAKp02 shows a significant similarity with Klebsiella phage vB_KpnS_MAG26fr, belonging to family Casjensviridae and the class Caudoviricetes. Phylogenetic analysis based on the terminase large subunit indicates a homology with Klebsiella phage Soft, also a member of Casjensviridae (Figure 4C). Comparative genomic analysis of phage SAKp02 with two Klebsiella phages showed high similarity (Figure 4D). A BLAST search revealed a high similarity in nucleotide percent identity between Klebsiella phage vB_KpnS_MAG26fr and phage SAKp02, while the phylogenetic analysis of the large terminase subunit indicated a closer similarity with Klebsiella phage Soft. VIRDIC analysis of intergenomic similarity of phage SAKp02 with Klebsiella phage soft and Klebsiella phage vB_KpnS_MAG26fr has shown a high similarity, indicating the clustering of the phages in the similar family, however, are distinct novel phages (Figure 4E). We found the phage genome was without any antibiotic-resistant genes or virulent factors, which is ideal for therapeutic use. The genome of SAKp02 has been deposited to NCBI, accession number OR290970.1, https://blast.ncbi.nlm.nih.gov/Blast.cgi#alnHdr_2523361381.
3.5. SAKp02 Effectively Dismantle K. pneumoniae Biofilm and Reduce Viable Cell Count
The scanning electron microscopy (SEM) analysis provided valuable insights into the impact of bacteriophage SAKp02 on 24-hour-old biofilm. The control slide displayed a well-established biofilm structure, characterized by densely packed bacteria coated with extracellular polymeric substances (EPS) (Figure 5A). However, exposure of the biofilm to SAKp02 resulted in significant disruption of the biofilm. The biofilms were visibly disintegrated, with degraded EPS and cell debris remaining on the coverslips. This visual observation indicated that bacteriophage SAKp02 had an eliminatory effect on the biofilm. (Figure 5B–E). To further investigate the antibiofilm activity, a crystal violet assay was conducted to measure the optical density at 580 nm (OD580) of the solubilized crystal violet, which serves as an indicator of biofilm biomass. The results demonstrated that after SAKp02 treatment, a significant reduction in biofilm biomass was achieved. SAKp02. Twelve hours post-treatment with bacteriophage SAKp02, a substantial 7-fold decrease in optical density compared to the control indicated the effective reduction of biofilm formation by the phage (Figure 5E). To validate these findings, bacterial colony counting was performed before and after the introduction of bacteriophage. This quantitative analysis provided evidence of the impact of phage treatment on biofilm-residing bacterial cells. After 4 hours of exposure to SAKp02, there was a 99.3% reduction in bacterial cells. However, the most pronounced reduction in viable bacterial cells in biofilm, amounting to a 99.999% reduction, was observed at the 12-hour post-infection with SAKp02 (Figure 5F). These results further confirmed the antibiofilm activity of bacteriophage SAKp02 and highlighted its effectiveness in reducing K. pneumoniae biofilm.
4. Discussion
Due to the high emergence of drug resistance, the treatment outcomes for Klebsiella pneumoniae have been extremely poor, prompting the World Health Organization (WHO) to designate this microorganism as a "priority-one critical pathogen" or the research and development of new antibiotics and new alternative treatments in 2017 [26]. A follow-up report revealed the insufficiency in the development of newer antibiotics. Besides, the ability of MDR K. pneumoniae to form biofilms further compounds the difficulties of treating patients [11,12]. Recent studies indicate that utilizing bacteriophages shows promise as an effective strategy against biofilms [27]. However, one of the major challenges in implementing phages as a clinical therapy is their host specificity, as most phages exhibit high specificity for their particular host strains; hence, it is crucial to isolate and fully characterize the bacteriophages, infectious against drug-resistant strains. It will expand the phage library against AMR strains, which can be further utilized for therapeutic purposes. Moreover, the accumulation of experimental data will be useful for the progression of phage therapy. This study isolated phage SAKp02, infectious against several drug-resistant clinical strains of K. pneumoniae, from hospital sewage water. A clear plaque indicates the virulent activity of the phage SAKp02 against the respective clinical strains. The plaques developed a halo, a semi-transparent clearing zone encircling the transparent core lysis zone (Figure 1A). Halos are formed due to the diffusion of phage-encoded enzymes through the bacterial lawn. Halos appeared after 24 hours of incubation, indicating a strong depolymerase activity. The genomic analysis identifies the tail fiber protein ORF36 (Table 1) reported for the depolymerase activity, which supports the plaque formation pattern of SAKp02 on the bacterial lawn. Electron microscopic image revealed the SAKp02 as a tailed phage (Figure 1B). Whole genome analysis revealed that the phages belong to the class of tailed bacteriophages, Caudoviricetes and family Casjensviridae. The adsorption rate, period, and time required to replicate and produce phage particles per host cell are important for phage infectivity. The time required from adsorption to bursting out of progeny virus after one infection cycle is attributed as eclipse or latency period. We found that the phage SAKp02 is relatively quick in getting adsorbed to the host compared to the average adsorption period of reported phages [28]. SAKp02 could adsorb to bacterial host as soon as 5 minutes (Figure 1C). To complete the replication cycle, phage SAKp02 took 40 minutes (Figure 1D). In approximately 40 minutes, phage SAKp02 could produce approximately 202 phage particles per bacterial cell. The bacteriolytic activity of the phages is the most crucial parameter to consider when using the respective phage in therapy. Several studies have reported the development of phage-resistant post-phage treatment within hours [29]. Resistance can be developed in several ways: modulation in host receptor, thus inhibiting the phage attachment, blocking phage DNA from entering inside bacterial cells, or nuclease mediated [30]. However, the development of phage resistance can interfere with the efficacy of phage therapy. Here, we have evaluated the bacteriolytic efficacy of phage SAKp02 with different MOIs (Figure 2A & B). Several studies have reported MOI-dependent lytic activity and phage resistance resurgence. However, with a different range of MOIs, phage SAKp02 could control bacterial growth for 12 hours, and the emergence of phage resistance has not been reported for any of the MOIs. This indicates a potent bacteriolytic activity of SAKp02. The host range of the phage is also an important factor to be included in the arsenal of phage therapy. SAKp02 could infect several drug-resistant clinical strains (Figure 2C), possibly indicating a common cell membrane protein required for the phage attachment. More studies are required to explore this possibility, which is one of the major concerns of phage therapy. The stability study revealed that the phage is substantially stable at 37ºC and range of pH (Figure 3A & B). One of the major concerns of any therapeutics containing live biological content is pharmaceutical excipients. Pharmaceutical excipients can also impact the stability of several vaccines. Phages, being the virus, need to be administrated and formulated in a constituent; thus, solvent stability is one of the important parameters regarding the longevity of the phage as a therapeutic constituent. SAKp02 has shown stability in PBS and 0.9% saline, the most common solvents used in experiments, besides 0.9% saline, which is the most commonly used intravenous fluid given to patients. Genomic analysis revealed that the phage SAKp02 belongs to the family Casjensviridae and the genus Yonseivirus. The whole genome sequences have identified several important proteins associated with phage lysis: holins, endolysin, tail fiber (spike) protein, or depolymerase (Figure 4B) (Table 1). Experimental data on the bacteriolytic activity has shown the efficient lytic activity of the phage SAKp02, indicating the efficient activity of the lytic enzymes. SAKp02 shows high nucleotide homogeneity with Klebsiella phage vB_KpnS_MAG26fr and Klebsiella phage soft SAKp02. EasyFigure was used to analyse the genomic similarities of phage SAKp02 with previously reported Klebsiella bacteriophages Vb_KpnS_MAG26fr and soft. BLAST comparison between three phages indicates a high nucleotide identity of several CDS regions ranging from as low as 65% to complete homogeneity. However, despite a high nucleotide identity, we can observe a dynamic genomic rearrangement between these genomes (Figure 4D), indicating strong evolutionary footsteps. Data on Casjensviridae phages are limited; hence, comparisons about bacteriolytic or antibiofilm properties could not be made. Moreover, this is the first extensive study on Casjensviridae phages. We found that phage SAKp02 does not carry the genes associated with pathogenicity or drug resistance; hence, it is safe for therapeutic use.
SAKp02 could significantly reduce the biofilm formed by drug-resistant K. pneumoniae strains after 4 hours of treatment. The most crucial component of the biofilm is EPS; in the control slide of scanning microscopic image (Figure 5A), we could observe that the bacterial cells are densely packed and embedded in the side EPS layer. Right after 4 hours of SAKp02 treatment, a reduction in the EPS layer could be seen (Figure 5B). The SEM data reflect the data of biofilm biomass reduction (Figure 5E) where, after 4 hours of treatment, approximately a 3-fold reduction in OD could be seen. The highest 12 hours of treatment was given to biofilm because studies have shown that there is a chance of the probable emergence of a phage-resistant bacterial population after a prolonged period of phage exposure. Extensive research on the emergence of phage resistance and the impact of phage resistance on bacterial fitness is required. Several researchers recommend a combination of antibiotics with phages to reduce the chance of developing phage and antibiotic resistance. The application of phage cocktails could be another approach to minimize phage resistance. Considering the rapidly emerging drug resistance, a safe and efficacious treatment alternative is needed to be established. Phages are nature-assigned scavengers of bacteria, equipped with bacteriolytic enzymes and genetic makeup to parasitize bacterial replication machinery to use against bacteria itself. Several studies have been done on the safety of the bacteriophages, and it has been reported that the phages are safe to use. Most importantly, being nature-made brings an advantage of evolutionary modifications, one of the most essential criteria for tackling ever-evolving bacteria. Selection pressure-driven evaluations make phages indispensable antimicrobial agents and a never-ending arsenal against bacteria.
Author Contributions
SC: data curation and writing—original draft; SJP and AR: data curation and reviewing; A.C: conceptualization, supervision, and writing—editing and review. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by UGC-BSR (F.30-487/2019(BSR)) and SERB-CRG (CRG/2021/001974).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author/s.
Acknowledgments
SC would like to thank ICMR for fellowship support.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| ESKAPE | Enterococcus faecium, Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumannii, Pseudomonas aeruginosa, and Enterobacter spp |
| WHO | World Health Organization |
| MMMH | Madras Medical Mission Hospital |
| ORF | Open Reading Frame |
| CDS | Coding Sequence |
| cKp | Classic K. pneumoniae |
| hyKp | Hypervirulent K. pneumoniae |
| EPS | Extracellular Polymeric Substance |
| BP | Base Pair |
| ESBL | Extended-Spectrum Beta-lactamase |
| MDR | Multi Drug Resistance |
| XDR | Extremely Drug Resistance |
| TEM | Transmission Electron Microscopy |
| SEM | Scanning Electron Microscopy |
| OD | Optical Density |
References
- Podschun, R.; Ullmann, U. Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clinical Microbiology Reviews 1998, 11, 589–603. [Google Scholar] [CrossRef]
- Luo, K.; Tang, J.; Qu, Y.; Yang, X.; Zhang, L.; Chen, Z.; Kuang, L.; Su, M.; Mu, D. Nosocomial infection by Klebsiella pneumoniae among neonates: a molecular epidemiological study. The Journal of Hospital Infection 2021, 108, 174–180. [Google Scholar] [CrossRef]
- Mohd Asri, N.A.; Ahmad, S.; Mohamud, R.; Mohd Hanafi, N.; Mohd Zaidi, N.F.; Irekeola, A.A.; Shueb, R.H.; Yee, L.C.; Mohd Noor, N.; Mustafa, F.H.; Yean, C.Y.; Yusof, N.Y. Global Prevalence of Nosocomial Multidrug-Resistant Klebsiella pneumoniae: A Systematic Review and Meta-Analysis. Antibiotics 2021, 10, 1508. [Google Scholar] [CrossRef] [PubMed]
- Mancuso, G.; Midiri, A.; Gerace, E.; Biondo, C. Bacterial Antibiotic Resistance: The Most Critical Pathogens. Pathogens 2021, 10, 1310. [Google Scholar] [CrossRef]
- Chang, D.; Sharma, L.; Dela Cruz, C.S.; Zhang, D. Clinical Epidemiology, Risk Factors, and Control Strategies of Klebsiella pneumoniae Infection. Frontiers in Microbiology 2021, 12, 750662. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Ferrer, S.; Peñaloza, H.F.; Budnick, J.A.; Bain, W.G.; Nordstrom, H.R.; Lee, J.S.; Van Tyne, D. Finding Order in the Chaos: Outstanding Questions in Klebsiella pneumoniae Pathogenesis. Infection and Immunity 2021, 89, e00693-20. [Google Scholar] [CrossRef]
- Wyres, K.L.; Wick, R.R.; Judd, L.M.; Froumine, R.; Tokolyi, A.; Gorrie, C.L.; Lam MM, C.; Duchêne, S.; Jenney, A.; Holt, K.E. Distinct evolutionary dynamics of horizontal gene transfer in drug resistant and virulent clones of Klebsiella pneumoniae. PLoS Genetics 2019, 15, e1008114. [Google Scholar] [CrossRef]
- Zhu, J.; Wang, T.; Chen, L.; Du, H. Virulence Factors in Hypervirulent Klebsiella pneumoniae. Frontiers in Microbiology 2021, 12, 642484. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.; Sanghrajka, K.; Gajjar, D. High Adhesion and Increased Cell Death Contribute to Strong Biofilm Formation in Klebsiellapneumoniae. Pathogens 2019, 8, 277. [Google Scholar] [CrossRef]
- Guerra, M.E.S.; Destro, G.; Vieira, B.; Lima, A.S.; Ferraz, L.F.C.; Hakansson, A.P.; Darrieux, M.; Converso, T.R. Klebsiella pneumoniae Biofilms and Their Role in Disease Pathogenesis. Frontiers in Cellular and Infection Microbiology 2022, 12, 877995. [Google Scholar] [CrossRef]
- Ochońska, D.; Ścibik, Ł.; Brzychczy-Włoch, M. Biofilm Formation of Clinical Klebsiella pneumoniae Strains Isolated from Tracheostomy Tubes and Their Association with Antimicrobial Resistance, Virulence and Genetic Diversity. Pathogens 2021, 10, 1345. [Google Scholar] [CrossRef] [PubMed]
- Stahlhut, S.G.; Struve, C.; Krogfelt, K.A.; Reisner, A. Biofilm formation of Klebsiella pneumoniae on urethral catheters requires either type 1 or type 3 fimbriae. FEMS Immunology and Medical Microbiology 2012, 65, 350–359. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, Z. Biofilm-forming Klebsiella pneumoniae strains have greater likelihood of producing extended-spectrum beta-lactamases. The Journal of Hospital Infection 2008, 68, 369–371. [Google Scholar] [CrossRef] [PubMed]
- Anderl, J.N.; Franklin, M.J.; Stewart, P.S. Role of antibiotic penetration limitation in Klebsiella pneumoniae biofilm resistance to ampicillin and ciprofloxacin. Antimicrobial Agents and Chemotherapy 2000, 44, 1818–1824. [Google Scholar] [CrossRef] [PubMed]
- Koskella, B.; Brockhurst, M.A. Bacteria-phage coevolution as a driver of ecological and evolutionary processes in microbial communities. FEMS Microbiology Reviews 2014, 38, 916–931. [Google Scholar] [CrossRef]
- Mangalea, M.R.; Duerkop, B.A. Fitness Trade-Offs Resulting from Bacteriophage Resistance Potentiate Synergistic Antibacterial Strategies. Infection and Immunity 2020, 88, e00926-19. [Google Scholar] [CrossRef]
- Chan, B.; Sistrom, M.; Wertz, J.; et al. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci Rep 2016, 6, 26717. [Google Scholar] [CrossRef]
- Chang, C.; Yu, X.; Guo, W.; Guo, C.; Guo, X.; Li, Q.; Zhu, Y. Bacteriophage-Mediated Control of Biofilm: A Promising New Dawn for the Future. Frontiers in Microbiology 2022, 13, 825828. [Google Scholar] [CrossRef]
- Oliveira, A.; Sillankorva, S.; Quinta, R.; Henriques, A.; Sereno, R.; Azeredo, J. Isolation and characterization of bacteriophages for avian pathogenic E. coli strains. Journal of Applied Microbiology 2009, 106, 1919–1927. [Google Scholar] [CrossRef]
- Kropinski, A.M.; Mazzocco, A.; Waddell, T.E.; Lingohr, E.; Johnson, R.P. Enumeration of bacteriophages by double agar overlay plaque assay. Methods in Molecular Biology 2009, 501, 69–76. [Google Scholar] [CrossRef]
- Sinha, A.; Eniyan, K.; Manohar, P.; Ramesh, N.; Bajpai, U. Characterization and genome analysis of B1 sub-cluster mycobacteriophage PDRPxv. Virus Research 2020, 279, 197884. [Google Scholar] [CrossRef]
- Amarillas, L.; Rubí-Rangel, L.; Chaidez, C.; González-Robles, A.; Lightbourn-Rojas, L.; León-Félix, J. Isolation and Characterization of phiLLS, a Novel Phage with Potential Biocontrol Agent against Multidrug-Resistant Escherichia coli. Frontiers in Microbiology 2017, 8, 1355. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Das, M.; Bhowmick, T.S.; Koley, H.; Atterbury, R.; Chakrabarti, A.K.; Sarkar, B.L. Isolation and Characterization of Novel Broad Host Range Bacteriophages of Vibrio cholerae O1 from Bengal. Journal of Global Infectious Diseases 2018, 10, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Lebeaux, D.; Ghigo, J.M.; Beloin, C. Full and broad-spectrum in vivo eradication of catheter-associated biofilms using gentamicin-EDTA antibiotic lock therapy. Antimicrobial Agents and Chemotherapy 2012, 56, 6310–6318. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, A.; Ghigo, J.M.; Beloin, C. Study of in vivo catheter biofilm infections using pediatric central venous catheter implanted in rat. Nature Protocols 2016, 11, 525–541. [Google Scholar] [CrossRef] [PubMed]
- Mitiku, A.; Aklilu, A.; Tsalla, T.; Woldemariam, M.; Manilal, A.; Biru, M. Magnitude and antimicrobial susceptibility profiles of Gram-Negative bacterial isolates among patients suspected of urinary tract infections in Arba Minch General Hospital, southern Ethiopia. PloS one 2022, 17, e0279887. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Lu, H.; Zhang, S.; Shi, Y.; Chen, Q. Phages against Pathogenic Bacterial Biofilms and Biofilm-Based Infections: A Review. Pharmaceutics 2022, 14, 427. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Bacteriophage Adsorption: Likelihood of Virion Encounter with Bacteria and Other Factors Affecting Rates. Antibiotics 2023, 12, 723. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Zheng, P.; Ji, W.; Fu, Q.; Wang, H.; Yan, Y.; Sun, J. SLPW: A Virulent Bacteriophage Targeting Methicillin-Resistant Staphylococcus aureus In vitro and In vivo. Frontiers in Microbiology 2016, 7, 934. [Google Scholar] [CrossRef]
- Li, N.; Zeng, Y.; Wang, M.; Bao, R.; Chen, Y.; Li, X.; Pan, J.; Zhu, T.; Hu, B.; Tan, D. Characterization of Phage Resistance and Their Impacts on Bacterial Fitness in Pseudomonas aeruginosa. Microbiology Spectrum 2022, 10, e0207222. [Google Scholar] [CrossRef]
Figure 1.
Plaque and structural morphology of phage SAKp02 (A). Plaque morphology of phage SAKp02 on the lawn of K. pneumoniae B3768 strain. Depolymerase activity is marked with an arrow (B). TEM Image of phage. Adsorption rate and time of SAKp02 and single step growth curve; (C). SAKp02 took 5 minutes to adsorb to the host with a rate of 95% of total phage particles. (D). Phage SAKp02 has shown a 20-minute latency period and a burst size of ~202 phage particles per host cell.
Figure 1.
Plaque and structural morphology of phage SAKp02 (A). Plaque morphology of phage SAKp02 on the lawn of K. pneumoniae B3768 strain. Depolymerase activity is marked with an arrow (B). TEM Image of phage. Adsorption rate and time of SAKp02 and single step growth curve; (C). SAKp02 took 5 minutes to adsorb to the host with a rate of 95% of total phage particles. (D). Phage SAKp02 has shown a 20-minute latency period and a burst size of ~202 phage particles per host cell.
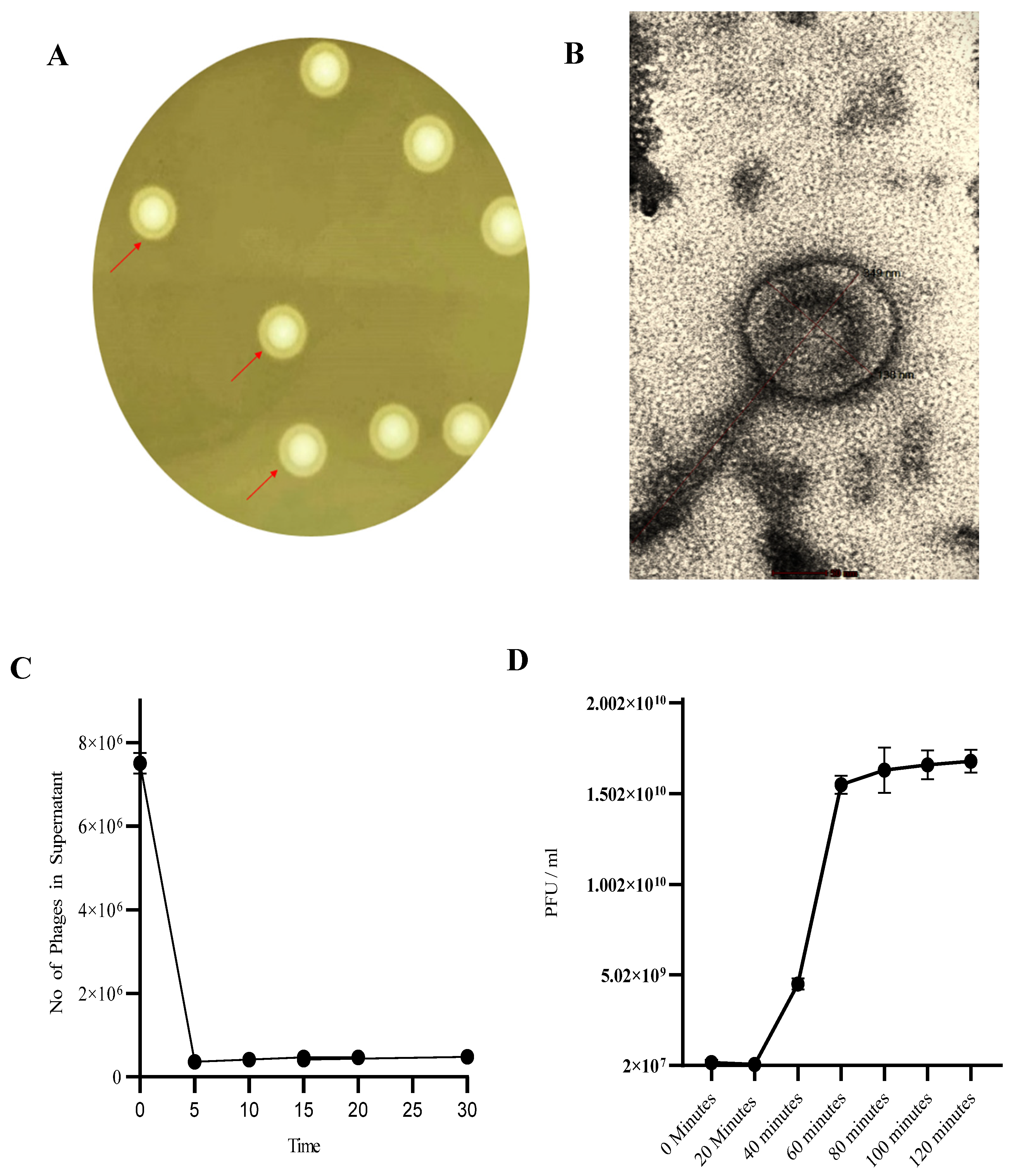
Figure 2.
Killing kinetics and Host range of phage SAKp02; (A). Phage SAKp02 could restrict the growth of clinical strain B3768 in comparison to the control (Bacteria + LB) at different MOI (B). Endpoint OD of the control and phage treatment with different MOIs after 12 hours post-phage treatment. (C). Phage SAKp02 could infect 43 K pneumoniae clinical isolates among 72 tested strains.
Figure 2.
Killing kinetics and Host range of phage SAKp02; (A). Phage SAKp02 could restrict the growth of clinical strain B3768 in comparison to the control (Bacteria + LB) at different MOI (B). Endpoint OD of the control and phage treatment with different MOIs after 12 hours post-phage treatment. (C). Phage SAKp02 could infect 43 K pneumoniae clinical isolates among 72 tested strains.
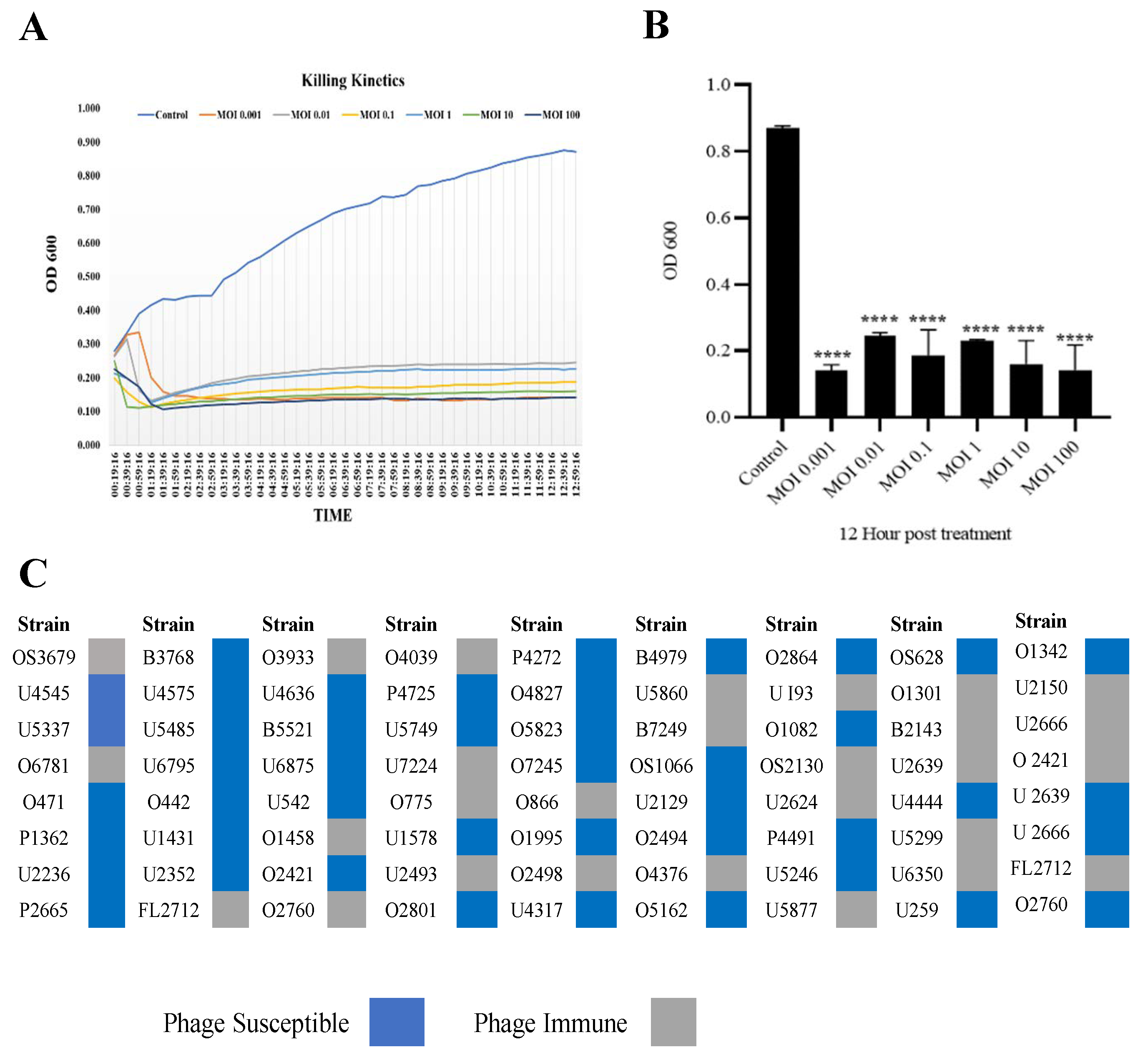
Figure 3.
Stability studies of phage SAKp02; (A). At pH 1, 3 and 14 complete degradation of phage SAKp02 was observed. (B). At 65 °C, a significant ~ 6-fold reduction was observed, and complete denaturation of phage particles at 100 °C was seen. (C). SAKp02 showed stability in PBS, chloroform, and 0.9% saline.
Figure 3.
Stability studies of phage SAKp02; (A). At pH 1, 3 and 14 complete degradation of phage SAKp02 was observed. (B). At 65 °C, a significant ~ 6-fold reduction was observed, and complete denaturation of phage particles at 100 °C was seen. (C). SAKp02 showed stability in PBS, chloroform, and 0.9% saline.
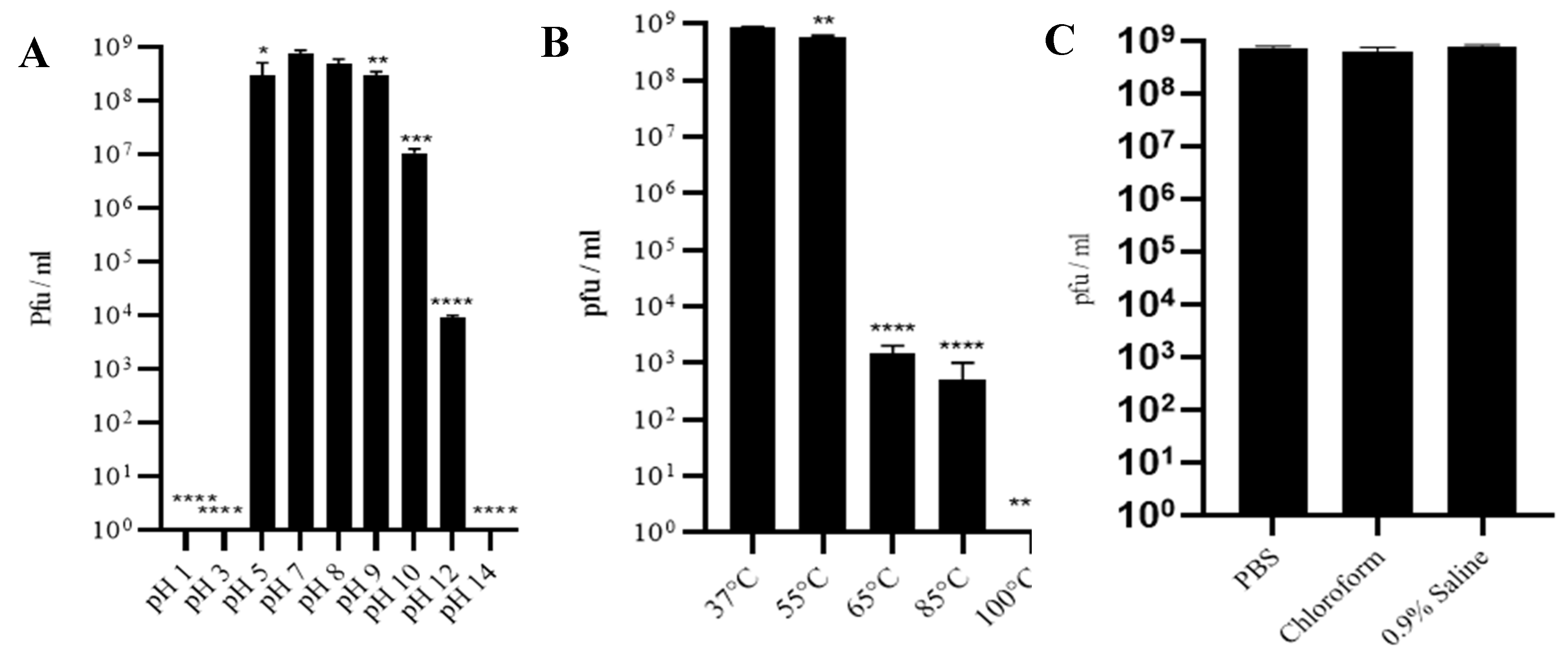
Figure 4.
Overview of whole genome of phage SAKp02 and bioinformatic analysis; (A). The GC content and genome alignment of phage SAKp02 constructed by GC Content calculator, the blue skew indicates the GC content according to the nucleotide regions. Mean GC content is marked as a straight black line. Calculation shows that the GC content of SAKp02 is 56.1%. (B). The genome map of phage SAKp02 generated by the genomevx. ORFs coding for hypothetical proteins is denoted in grey. Genes with predicted functions are coloured; orange-coloured genes are indicated as phage enzyme associated with replication process at different stages of viral life cycle, green coloured CDSs are denoted as structural proteins of the virus, and yellow coloured CDSs are proteins associated with regulatory activities. (C). Neighbour-joining phylogenetic tree of terminase large subunits from bacteriophage SAKp02 and the related phages. Sequences were downloaded from NCBI. (D). EasyFigure was used to create a linear representation and comparative analysis, visualizing alignments phage soft (top DNA), SAKp02 (middle DNA), and phage vB_KpnS_MAG26fr (bottom DNA). Arrows denote the transcription direction of predicted ORFs and, are categorized as grey (hypothetical proteins), green (structural proteins), orange (viral enzymes), and yellow (regulatory proteins). Shaded regions indicate well-conserved segments with 65-100% amino acid similarity, with darker grey regions showing higher identity. The scale is in 10 kbp increments. (E) A heatmap generated by VIRIDIC illustrates the intergenomic similarities on the right half, using color-coding to provide a quick visual comparison of the ten phage genomes most closely related to SAKp02 (NC-048805.1: Phage soft, OR290970.1: SAKp02, OP558002.1: Klebsiella phage vB_KpnS_MAG26fr).
Figure 4.
Overview of whole genome of phage SAKp02 and bioinformatic analysis; (A). The GC content and genome alignment of phage SAKp02 constructed by GC Content calculator, the blue skew indicates the GC content according to the nucleotide regions. Mean GC content is marked as a straight black line. Calculation shows that the GC content of SAKp02 is 56.1%. (B). The genome map of phage SAKp02 generated by the genomevx. ORFs coding for hypothetical proteins is denoted in grey. Genes with predicted functions are coloured; orange-coloured genes are indicated as phage enzyme associated with replication process at different stages of viral life cycle, green coloured CDSs are denoted as structural proteins of the virus, and yellow coloured CDSs are proteins associated with regulatory activities. (C). Neighbour-joining phylogenetic tree of terminase large subunits from bacteriophage SAKp02 and the related phages. Sequences were downloaded from NCBI. (D). EasyFigure was used to create a linear representation and comparative analysis, visualizing alignments phage soft (top DNA), SAKp02 (middle DNA), and phage vB_KpnS_MAG26fr (bottom DNA). Arrows denote the transcription direction of predicted ORFs and, are categorized as grey (hypothetical proteins), green (structural proteins), orange (viral enzymes), and yellow (regulatory proteins). Shaded regions indicate well-conserved segments with 65-100% amino acid similarity, with darker grey regions showing higher identity. The scale is in 10 kbp increments. (E) A heatmap generated by VIRIDIC illustrates the intergenomic similarities on the right half, using color-coding to provide a quick visual comparison of the ten phage genomes most closely related to SAKp02 (NC-048805.1: Phage soft, OR290970.1: SAKp02, OP558002.1: Klebsiella phage vB_KpnS_MAG26fr).
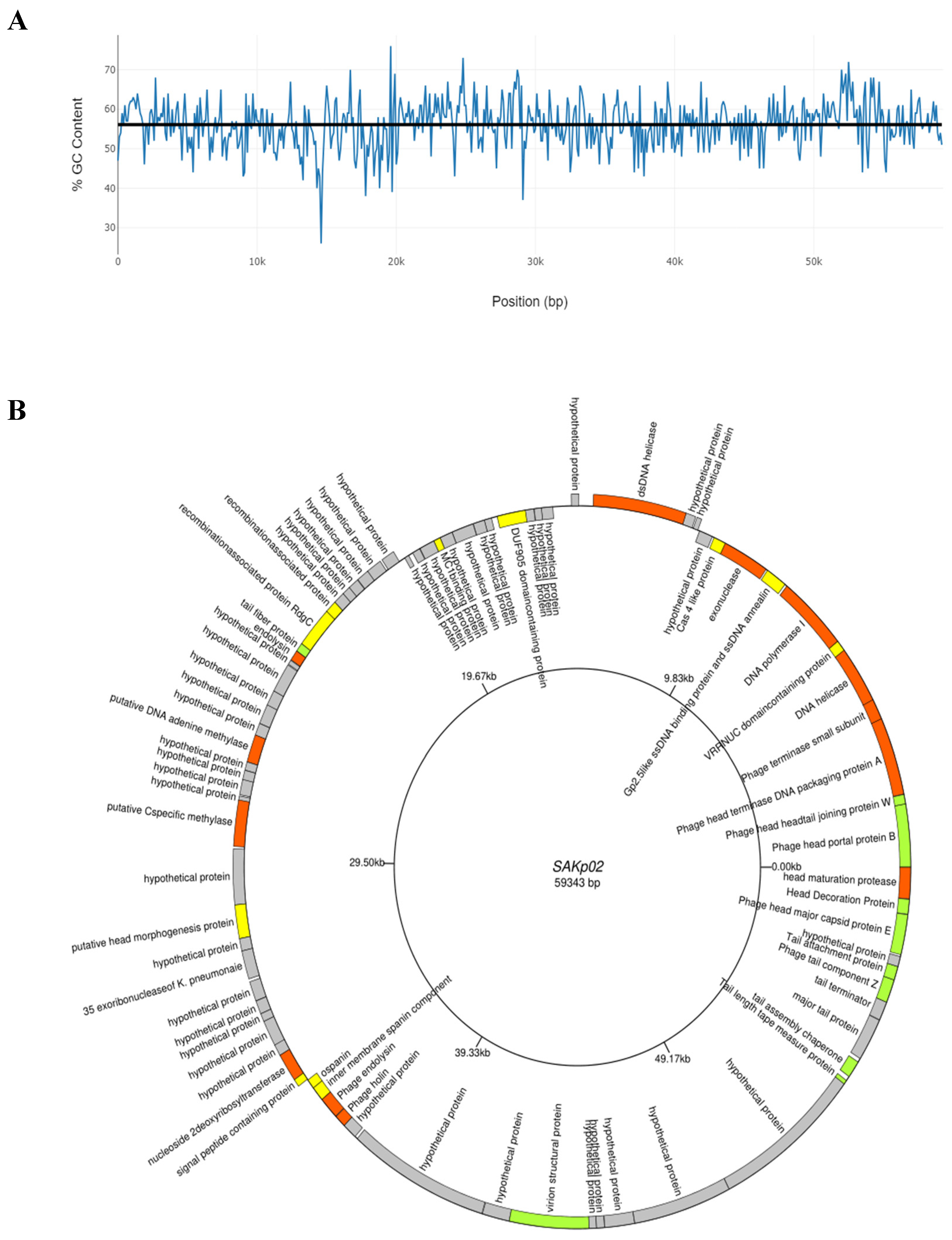
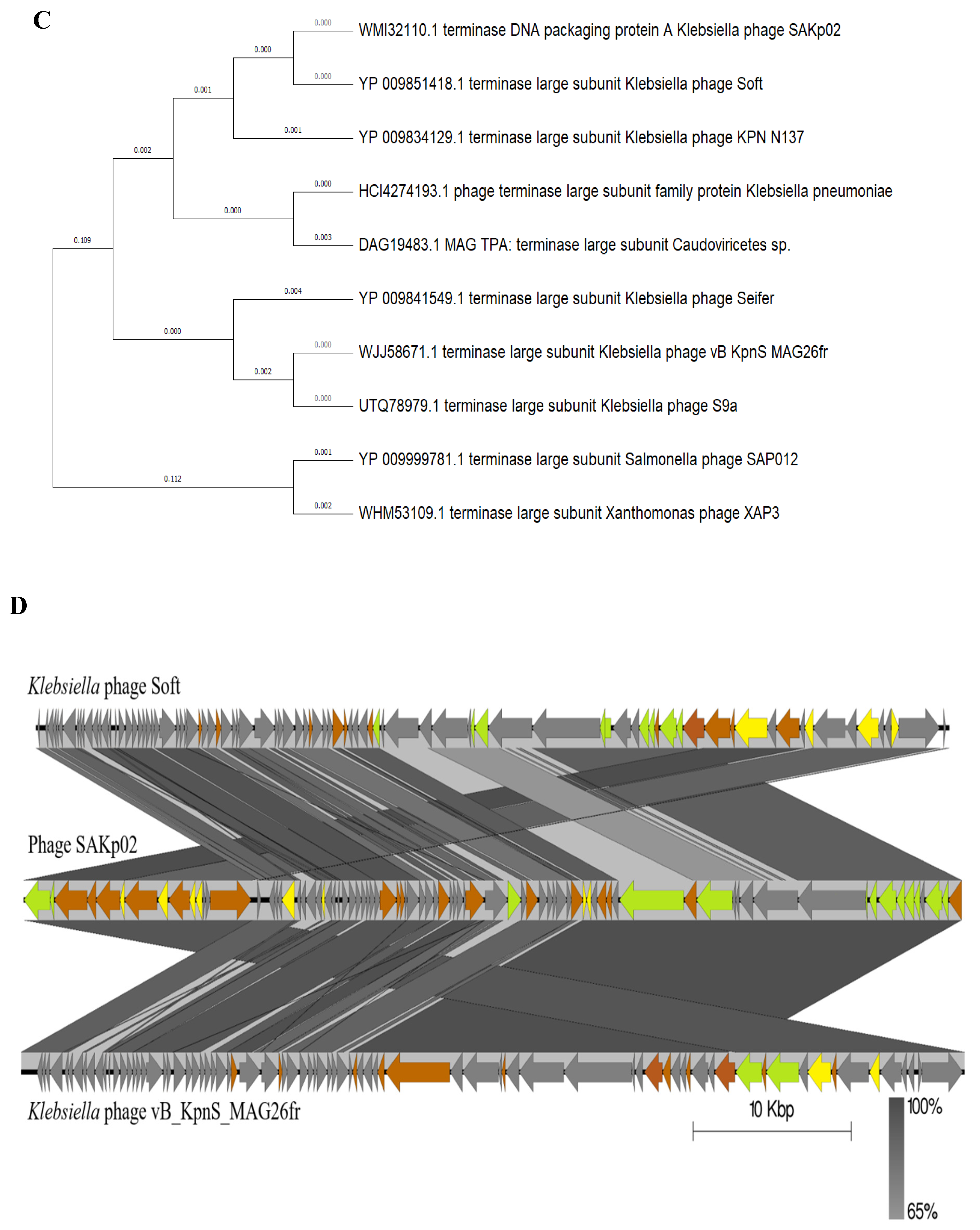
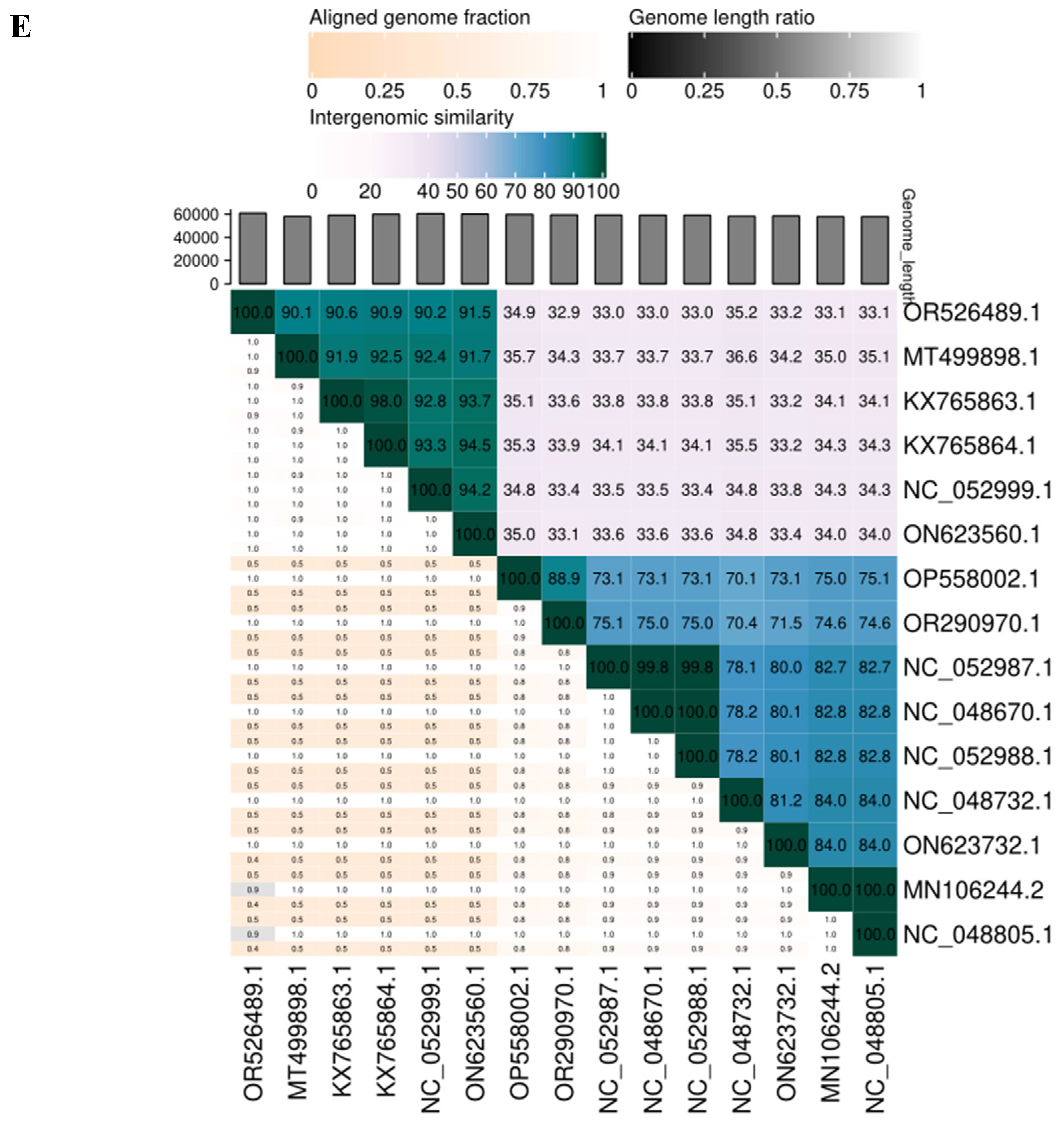
Figure 5.
Assessment of antibiofilm activity of SAKp02. (A). The Scanning Electron Microscopy (SEM) analysis revealed that K. pneumoniae exhibited a compact bacterial community covered by an extracellular polymeric substance (EPS) layer (white arrows). This observation indicates strong biofilm formation by K. pneumoniae MDR clinical strains. (B). 4-hour treatment with SAKp02 could effectively degrade the EPS layers and reduced the density of the bacteria. (C). & (D). After 8 and 12 hours of treatment with phage SAKp02; there was a noticeable degradation of the biofilm matrix and bacterial cell. Cellular debris (red arrow) on the slides indicates a lytic phage infection. (E). Crystal violet assay indicated an effective demolition of biofilm biomass after 4 hours of phage treatment; 12 hours of phage treatment showed the lowest biofilm biomass; a 7-fold decrease in OD compared to control is seen. (F). Effective lysis of biofilm viable cells was seen with SAKP02 infection; only <0.001 percent of bacteria survived after 12 hours of phage treatment.
Figure 5.
Assessment of antibiofilm activity of SAKp02. (A). The Scanning Electron Microscopy (SEM) analysis revealed that K. pneumoniae exhibited a compact bacterial community covered by an extracellular polymeric substance (EPS) layer (white arrows). This observation indicates strong biofilm formation by K. pneumoniae MDR clinical strains. (B). 4-hour treatment with SAKp02 could effectively degrade the EPS layers and reduced the density of the bacteria. (C). & (D). After 8 and 12 hours of treatment with phage SAKp02; there was a noticeable degradation of the biofilm matrix and bacterial cell. Cellular debris (red arrow) on the slides indicates a lytic phage infection. (E). Crystal violet assay indicated an effective demolition of biofilm biomass after 4 hours of phage treatment; 12 hours of phage treatment showed the lowest biofilm biomass; a 7-fold decrease in OD compared to control is seen. (F). Effective lysis of biofilm viable cells was seen with SAKP02 infection; only <0.001 percent of bacteria survived after 12 hours of phage treatment.
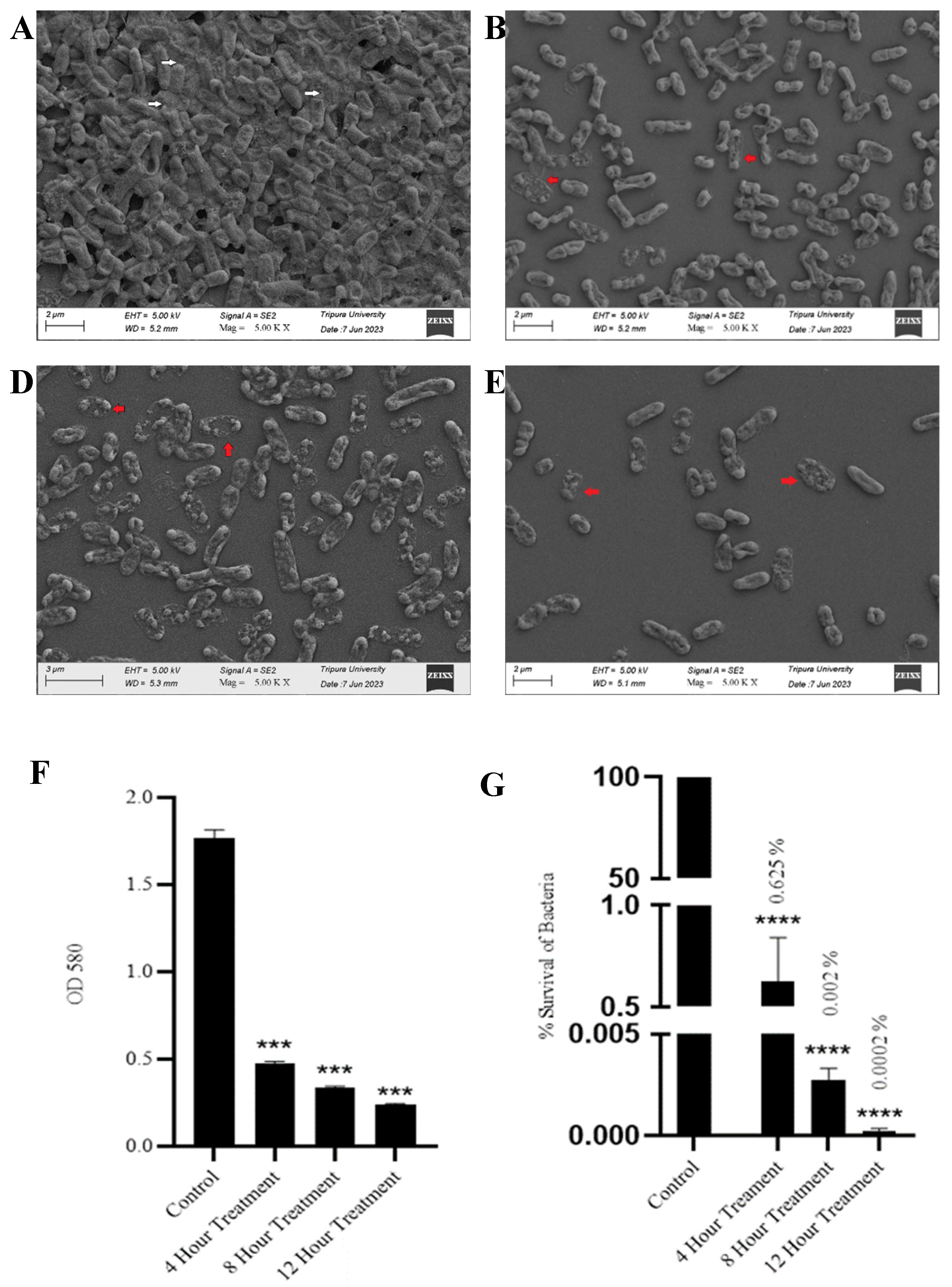

Table 1.
Predicted ORFs in SAKp02 genome.
| Sr. | Start | Stop | Size(nt) | Strand | Function |
| 1 | 1665 | 1 | 1664 | - | Phage head, portal protein B |
| 2 | 1916 | 1665 | 251 | - | Phage head, head-tail joining protein W |
| 3 | 4003 | 1928 | 2075 | - | Phage head, terminase DNA packaging protein A |
| 4 | 4562 | 3993 | 569 | - | Phage terminase, small subunit |
| 5 | 6087 | 4546 | 1541 | - | DNA helicase |
| 6 | 6376 | 6080 | 296 | - | VRR-NUC domain-containing protein |
| 7 | 8418 | 6376 | 2042 | - | DNA polymerase I |
| 8 | 9105 | 8485 | 620 | - | Gp2.5-like ssDNA binding protein and ssDNA annealing protein |
| 9 | 10501 | 9152 | 1349 | - | Exonuclease |
| 10 | 10844 | 10494 | 350 | - | Cas 4 like protein |
| 11 | 11291 | 10899 | 392 | - | Hypothetical protein |
| 12 | 11344 | 11481 | 137 | + | Hypothetical protein |
| 13 | 11521 | 11802 | 281 | + | Hypothetical protein |
| 14 | 11799 | 14390 | 2591 | + | dsDNA helicase |
| 15 | 14793 | 15005 | 212 | + | Hypothetical protein |
| 16 | 15860 | 15531 | 329 | - | Hypothetical protein |
| 17 | 16063 | 15857 | 206 | - | Hypothetical protein |
| 18 | 16326 | 16075 | 251 | - | Hypothetical protein |
| 19 | 17131 | 16316 | 815 | - | DUF905 domain-containing protein |
| 20 | 17510 | 17298 | 212 | - | Hypothetical protein |
| 21 | 17833 | 17507 | 326 | - | Hypothetical protein |
| 22 | 18472 | 17843 | 629 | - | Hypothetical protein |
| 23 | 18840 | 18475 | 365 | - | Hypothetical protein |
| 24 | 19040 | 18843 | 197 | - | MC1-binding protein |
| 25 | 19477 | 19040 | 437 | - | Hypothetical protein |
| 26 | 19714 | 19490 | 224 | - | Hypothetical protein |
| 27 | 19976 | 19848 | 128 | - | Hypothetical protein |
| 28 | 20158 | 20535 | 377 | + | Hypothetical protein |
| 29 | 20595 | 21044 | 449 | + | Hypothetical protein |
| 30 | 21022 | 21366 | 344 | + | Hypothetical protein |
| 31 | 21379 | 21681 | 302 | + | Hypothetical protein |
| 32 | 21674 | 21907 | 233 | + | Hypothetical protein |
| 33 | 21904 | 22218 | 314 | + | Hypothetical protein |
| 34 | 22215 | 22547 | 332 | + | Hypothetical protein |
| 35 | 22537 | 23607 | 1070 | + | Exonuclease |
| 36 | 23609 | 23836 | 227 | + | Tail fiber protein |
| 37 | 23838 | 24110 | 272 | + | Endolysin |
| 38 | 24126 | 24242 | 116 | + | Hypothetical protein |
| 39 | 24266 | 25033 | 767 | + | Hypothetical protein |
| 40 | 25030 | 25437 | 407 | + | Hypothetical protein |
| 41 | 25427 | 25936 | 509 | + | Hypothetical protein |
| 42 | 25942 | 26262 | 320 | + | Hypothetical protein |
| 43 | 26255 | 26965 | 710 | + | Putative DNA adenine methylase |
| 44 | 26968 | 27183 | 215 | + | Hypothetical protein |
| 45 | 27187 | 27450 | 263 | + | Hypothetical protein |
| 46 | 27428 | 27820 | 392 | + | Hypothetical protein |
| 47 | 27855 | 27980 | 125 | + | Hypothetical protein |
| 48 | 27958 | 29142 | 1184 | + | Putative C-specific methylase |
| 49 | 29203 | 30630 | 1427 | + | Hypothetical protein |
| 50 | 30630 | 31493 | 863 | + | Putative head morphogenesis protein |
| 51 | 31495 | 31815 | 320 | + | Hypothetical protein |
| 52 | 31812 | 32546 | 734 | + | 3'-5' exoribonuclease of K. pneumonaie |
| 53 | 32621 | 33133 | 512 | + | Hypothetical protein |
| 54 | 33137 | 33484 | 347 | + | Hypothetical protein |
| 55 | 33487 | 33696 | 209 | + | Hypothetical protein |
| 56 | 33698 | 34381 | 683 | + | Hypothetical protein |
| 57 | 34365 | 34664 | 299 | + | Hypothetical protein |
| 58 | 34661 | 35404 | 743 | + | Nucleoside 2-deoxyribosyltransferase |
| 59 | 35394 | 35594 | 200 | + | Signal peptide containing protein |
| 60 | 35881 | 35618 | 263 | - | O-Spanin |
| 61 | 36267 | 35878 | 389 | - | Hypothetical protein |
| 62 | 36851 | 36267 | 584 | - | Phage endolysin |
| 63 | 37196 | 36864 | 332 | - | Phage holin |
| 64 | 37615 | 37196 | 419 | - | Hypothetical protein |
| 65 | 41762 | 37695 | 4067 | - | Hypothetical protein |
| 66 | 42560 | 41781 | 779 | - | Hypothetical protein |
| 67 | 44839 | 42563 | 2276 | - | Vrion structural protein |
| 68 | 45051 | 44839 | 212 | - | Hypothetical protein |
| 69 | 45278 | 45051 | 227 | - | Hypothetical protein |
| 70 | 46139 | 45288 | 851 | - | Hypothetical protein |
| 71 | 48969 | 46144 | 2825 | - | Hypothetical protein |
| 72 | 53274 | 48973 | 4301 | - | Hypothetical protein |
| 73 | 53380 | 53267 | 113 | - | Tail length tape measure protein |
| 74 | 53964 | 53506 | 458 | - | Tail assembly chaperone |
| 75 | 55210 | 54071 | 1139 | - | Major tail protein |
| 76 | 55727 | 55224 | 503 | - | Tail terminator |
| 77 | 56344 | 55724 | 620 | - | Phage tail, component Z |
| 78 | 56709 | 56344 | 365 | - | Tail attachment protein |
| 79 | 56980 | 56711 | 269 | - | Hypothetical protein |
| 80 | 58095 | 57031 | 1064 | - | Phage head, major capsid protein E |
| 81 | 58497 | 58108 | 389 | - | Phage head, head-DNA stabilization protein D |
| 82 | 59343 | 58510 | 833 | - | Phage head, head-tail preconnector protease C |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



