
Submitted:
31 May 2024
Posted:
03 June 2024
You are already at the latest version
Abstract
Bioactive peptides are sequences of amino acids, cut from the parent protein with en-zymes or high pressure processing that can impart a tertiary health benefit to the con-sumer that goes beyond basic human nutrition. This study explores mesopelagic fish in-cluding Maurolicus muelleri and Benthosema glaciale as sources of peptides with an-ti-inflammatory, heart health beneficial, antioxidant and other bioactivities. Bioactive peptides were generated post-catch using different enzymatic hydrolysis processes ap-plied to mesopelagic species caught from experimental cruises targeting these fish. Maurolicus muelleri and Benthosema glaciale were the dominant species processed. Bi-oactive hydrolysates included a Maurolicus muelleri Alcalase hydrolysate and a Maurolicus muelleri autolysis. Hundreds of peptide sequences were identified using mass spectrome-try from active hydrolysates. In silico analysis was used to rank identified peptide se-quences in terms of overall bioactivity using Peptide Ranker, anti-inflammatory activity using PrepAIP, ability to impart Umami flavours with Umami-MRNN and dipeptidyl peptidase IV inhibitory potential using AntiDMPpred . Thirty peptides were selected fol-lowing in silico analysis. Several of the selected peptides had sequence homology with peptides derived from other fish proteins including Atlantic salmon (peptide QCPLHR-PWAL) and thin-lipped mullet (peptide FDAFLPM). Seven peptides predicted to have an-ti-inflammatory, anti-type 2 diabetes or Umami potential using in silico strategies were chemically synthesized and their anti-inflammatory activities confirmed using in vitro bioassays with COX-1 and COX-2 enzymes. The peptide QCPLHRPWAL inhibited COX-1 and COX-2 by 82.90% (+/-0.54) and 53.84% respectively and had a selectivity index greater than 10. This peptide warrant further research as a novel anti-inflammatory/pain relief peptide. Other bioactivities associated with DPP-IV inhibition and Umami flavours de-velopment are discussed. Identified peptides offer potential for use in functional foods or topical agents to prevent pain and inflammation associated with diseases like psoriasis, arthritis and inflammatory bowel disease, however further research in vivo is required.
Keywords:
peptides
; anti-inflammatory
; heart health
; anti-diabetic
; Type-2-diabetes
; in silico analysis
; mesopelagic fish
; hydrolysates
; circular economy
1. Introduction
Mesopelagic fish occur in the twilight zone at depths of between 200-1000 meters and are largely underexplored and not exploited currently as food, feed and as a source of bioactive compounds. Mesopelagic species include Maurolicus muelleri (Mueller’s Pearlside) and Meganyctiphanes norvegica (Northern Krill) [1]. Recent work in the MEESO and SUMMER EU H2020 projects identified Angiotensin-1-converting enzymes (ACE-1) inhibitory, Dipeptidyl peptidase IV (DPP-IV) inhibitory and antimicrobial activities in hydrolysates and extracts generated from mixed mesopelagic fish trawls [2]. Additional studies by Aliyu and colleagues (2022) identified the antioxidant potential of hydrolysates generated from M. muelleri and Krill [3]. More recently, Medina and colleagues looked at on board processing of mesopelagic species including M. muelleri and Benthosema glaciale as a source of dietary lipids [4].
Mueller’s Pearlside is a small fish 4-5 cm in length and there is a significant population of this species in the upper mesopelagic zone. Due to its size and appearance, use of this resource is limited despite the abundant numbers of this species reported to exist with estimates of 11-25 Gt present in global waters [5]. Hydrolysis of difficult to process biomass with enzymes is a useful approach for the generation of high quality protein products. Enzymes impart selectivity and specificity to products and hydrolysis can add value to proteins by increasing digestibility, bioavailability and bioactivities of the protein biomass by creation of bioactive peptides. Additionally, hydrolysis can reduce the potential of proteins to cause allergy [6,7]. Bioactive peptides are sequences between 2-30 amino acids in length that may be derived from parent proteins using hydrolysis and have several known bioactivities [8]. A database of bioactive peptides known as BIOPEP-UWM contains over 4,800 bioactive peptides identified from natural resources. Bioactivities ordinarily reported in this database include ACE-1 inhibitors, DPP-IV inhibitors, antioxidative and antimicrobial peptides. Bioactive and functional food peptides can positively affect regulation of an individual’s health by preventing various ailments and by improving medical conditions related to lifestyle, metabolism, and immunity. Several therapeutic peptides are approved for use in the mitigation of blood disorders and diseases associated with metabolic syndrome such as type-2-diabetes (T2D). For example, the drug Liraglutide (Victoza) - a glucagon-like peptide-1 receptor (GLP-1R) agonist is approved for treatment of T2D and is a top-selling drug produced by Novo Nordisk [10]. Goserelin, with the commercial name Zoladex from Astra Zeneca is a gonadotropin-releasing hormone agonist (GnRH agonist) that suppresses specific sex hormones and is used to treat prostate and breast cancers [11]. Other companies including Amgen, Eli Lilly, Roche, and Pfizer have peptide drugs in development for treatment of various diseases. Peptides have high selectivity and potency, low toxicity, have several targets; they have low accumulation in tissue and high chemical diversity. Disadvantages of peptides can include poor solubility, oral bioavailability and metabolic stability [12].
Chronic inflammation is a prolonged, over-reactive, immune system response and if it goes un-checked, it keeps the body in a constant state of stress and high alert resulting in disease condition like arthritis, psoriasis and irritable bowel disease (IBD) [13]. Globally, 3 out of 5 people die due to chronic inflammatory diseases associated with respiration and heart issues as well as stroke, cancer, diabetes and obesity [14]. Psoriasis, caused also by systemic inflammation, affects approximately 125 million people worldwide and has the highest incidences of occurrence in Northern European countries like Norway and Ireland [15]. Chronic pain and systemic inflammation often occur in sufferers concurrently. Arthritis affects 350 million people worldwide [16]. Drugs including ibuprofen and aspirin act on targets for inflammation and pain including cyclooxygenase (COX) enzymes to reduce pain and treat chronic inflammation. They are non-steroidal anti-inflammatory drugs (NSAIDs) that are highly effective but their long-term use is linked to toxicity, high blood pressure and hepatic issues [17]. Worldwide, alternatives to NSAIDs are sought from natural sources. Cyclooxygenase (COX; E.C. 1.14. 99.1) enzymes, convert arachidonic acid into inflammatory prostaglandins (PGs). There are two types; COX-1, which, is expressed in the gastric epithelium where it provides protection to the gastric mucosa as it regulates the production of acid and mucus. COX-2 is involved in inflammation and is a source of the inflammatory mediator’s prostaglandin E2 (PGE2) and prostacyclin (PGI2) in the arachidonic acid pathway [18]. COX-2 inhibitors have anti-inflammatory, anti-pyretic and analgesic effects in diseases including rheumatoid and osteoarthritis and acute pain [19]. Patents concerning food derived ingredients and their use for preventing pain and inflammation exist. For example, a patent filed in 2005 details a dietary supplement containing natural COX inhibitors and methods for inhibiting pain and inflammation. The COX inhibitory ingredient consisted of a pain-relieving anthocyanin-enriched blend of berry extracts (elderberry, bilberry, tart cherries) [20].
Monoacylglycerol lipase (MAGL; EC 3.1. 1.23) is a rate-limiting enzyme in the degradation of Monoacylglycerol, which hydrolyses Monoacylglycerol to glycerol and fatty acids, with highest expression in brain, white adipose tissue and the liver. It is also an endocannabinoid-degrading enzyme converting 2-arachidonoylglycerol (2-AG), an endogenous ligand for the cannabinoid receptors CB1 and CB2, into arachidonic acid. MAGL terminates 2-AG signaling, and is the major source of arachidonic acid and proinflammatory prostaglandins in the liver or in the brain. It is considered an anti-inflammatory target [21]. Natural COX and MAGL inhibitory compounds exhibiting anti-inflammatory activity, which can be included in functional foods or within nutraceutical formulations, are a potentially better alternative to synthetic drugs for the prevention and treatment of inflammation and pain. Few anti-inflammatory peptides from food sources are reported in the literature. One example is the anti-inflammatory peptide with the sequence Pro-Pro-Tyr-Leu-Ser-Pro, Ile-Ile-Gly-Gly-Ala-Leu, and Phe-Leu-Pro-Pro-Val-Thr-Ser-Met-Gly derived from maize [22]. Dia, Wang, Oh, de Lumen, and de Mejia (2009) isolated lunasin, which is a chemopreventive peptide from defatted soybean flour, and determined the anti-inflammatory effect that lunasin decreased the production of NO and PGE2, and the expression of iNOS and COX-2 in LPS-stimulated RAW264.7 macrophages [23]. A number of anti-inflammatory peptides were identified from fish protein hydrolysates previously. Ahn isolated an anti-inflammatory tripeptide with the sequence Pro-Ala-Tyr (PAY) from the pectoral fin of salmon [24]. PAY exhibited anti-inflammatory action via inhibiting NO/iNOS and PGE2/COX-2 pathway. PAY inhibited IL-1β production by 64.89% at a concentration of 0.75 mM. Jang and colleagues isolated anti-inflammatory peptides from enzymatic hydrolysates of sandfish (Arctoscopus japonicus) protein [25]. Additionally, the high molecular weight fraction of a pepsin hydrolysate generated from blue mussel was found to inhibit iNOS and COX-2 gene expression [26]. No inhibitors of MAGL are reported in the literature to date, derived from fish sources.
Renin and DPP-IV inhibitors can help to modulate innate and adaptive immunity. Inhibition of DPP-IV has known beneficial effects concerning glycemic control in cardiovascular disease and renal issues. In addition, DPP-IV inhibitors can help reduce weight in obese individuals.
Herein, we describe the generation, characterisation, synthesis and confirmation of COX-1, COX-2 and MAGL inhibitory activities by peptides derived from hydrolysates generated from mesopelagic fish trawls including the species M. muelleri. Generated hydrolysates were also assessed for their antioxidant, renin and DPP-IV inhibitory activities. A strategy combining enzyme hydrolysis using different proteases including Alcalase® applied to M. muelleri; screening hydrolysates for different bioactivities using in vitro assay methods; enrichment of bioactive hydrolysates using molecular weight cut off (MWCO) filtration; enrichment of active hydrolysates using cartridge filtration to generate permeates, and subsequent mass spectrometry (MS) analysis of permeates was used to generate and identify a number of peptide sequences from Irish and Spanish mesopelagic species. Subsequently, in silico analysis using the programmes Peptide Ranker [27], PreAIP RF [28], BIOPEP-UWM and Umami-MRNN were used to characterise the bioactive potential of peptide sequences and their potential to affect inflammation, type-2-diabetes and flavours (umami). Following in silico analysis, eight peptides were selected for chemical synthesis and were subsequently assessed for their COX-1, COX-2 and MAGL inhibitory activities in vitro. The percentage inhibition of COX-1, COX-2 and MAGL was determined for the most active peptides and COX-1, COX-2 and MAGL EC50 values were assigned to three novel peptides identified from the different mesopelagic hydrolysates. Novel peptides discovered and not found in databases like BIOPEP-UWM include peptides with the sequences QCPLRPWAL, NVGEVVCIFLTALGLPEALI, and FDFLPM derived from the hydrolysis of Irish mesopelagic fish species M. muelleri with Alcalase®, as well as several other peptides described here.
2. Results
2.1. Hydrolysate Generation from Irish and Norwegian Mesopelagic Trawls and Proximate Compositional Analysis of Resulting Hydrolysates
The degree of hydrolysis for Irish hydrolysate samples made using Alcalase® applied to Irish mesopelagic fish trawls consisting of a mixture of the fish species Notocopelus elongtus kroyeri, Benthosema glaciale and Maurolicus muelleri as well as Blue whiting species ranged from 50-70%. Following generation of hydrolysates and stabilization using freeze-drying, hydrolysates were characterised for their protein, ash, lipid and moisture content using AOAC methods for protein, ash and lipid content, as described previously [30]. The protein content of generated hydrolysates ranged from 14.04% (hydrolysate generated from Notocopelus elongtus kroyeri) to 18.02% (hydrolysate generated from M. muelleri). The ash content of hydrolysates generated from Irish mesopelagic trawls using Alcalase® was between two (Notocopelus elongtus kroyeri) to 3.8% (M. muelleri) and the lipid content ranged from 0.46% (hydrolysate of Maurolicus muelleri with Alcalase®) to 9.92% (Benthosema glaciale hydrolysate with Alcalase®) of dry weight.
2.2. Hydrolysate Generation from Spanish Mesopelagic Fish Trawl
The yields of solids, oil, hydrolysates and bones generated using different enzymes applied to Spanish mesopelagic trawls consisting of M. muelleri are shown in Figure 1. The greatest yield of hydrolysate resulted from use of Alcalase and Papain applied to the mesopelagic biomass (M. muelleri solely) and the combination of Papain and Bromelain applied to the same biomass. The lowest hydrolysate yields were obtained using Bromelain and no added enzyme (control, with only endogenous enzymes) applied to M. muelleri biomass.
2.3. Molecular Weight Distribution and the Degree of Hydrolysates Generated from M. muelleri Captured on Spanish Trawls
The molecular size profile distribution of the peptides resulting from the enzymatic digestion of Spanish M. muelleri indicates that papain, the combination of papain and bromelain and Protamex (applied separately to mesopelagic biomass) produced more peptides less than 1-kDa in size compared to the peptides generated using other enzymes applied to the biomass. Bromelain alone was less efficient and produced more peptides greater than 3-kDa in size (Figure 2).
The degree of hydrolysis observed for the hydrolysates generated from M. muelleri using different commercial enzymes ranged from 50%-60%.
2.4. In Vitro Bioactivity Screening of Generated Mesopelagic Fish Hydrolysates
2.4.1. Cyclooxygenase (COX) Enzyme Inhibition by Generated Hydrolysates
Freeze-dried Irish, Norwegian and Spanish hydrolysates generated from mesopelagic species with different enzymes were assessed at concentrations of 1 mg/mL in dimethyl sulfoxide (DMSO) to determine their potential to prevent pain and inflammation via the COX-1 and COX-2 inhibition pathways (Figure 3 and Figure 4). M. muelleri hydrolysed with Alcalase 2.4 LG (MME02) and M. muelleri hydrolysed by endogenous enzymes (MMC019) were found to inhibit COX-1 enzyme by 39.42% and 37.53% when assayed at concentrations of 1 mg/mL compared to a positive control. The same extracts were found to inhibit COX-2 by 25.14% and 20.14% respectfully compared to Resveratrol, which inhibited COX-2, by 65.87% (the positive control). In addition, the Irish hydrolysate generated from Benthosema glaciale biomass with Alcalase® (labelled 2 corresponds to CE21004 Haul2) inhibited both COX-1 and COX-2 by 20-22% when assayed at 1 mg/mL.
2.4.2. Monoacylglycerol Lipase (MAGL) Enzyme Inhibition by Generated Hydrolysates
MAGL is a serine hydrolase that hydrolyses monoglycerides into glycerol and fatty acids. It links the endocannabinoid and eicosanoid systems through degradation of endocannabinoid 2-arachidaoylglycerol into arachidonic acid, the precursor of prostaglandins and other inflammatory mediators. MAGL inhibitors are known anti-nociceptive, anxiolytic, anti-inflammatory agents [31]. Irish mesopelagic species hydrolysates were assessed for their ability to inhibit the MAGL enzyme. M. muelleri biomass hydrolysed with Alcalase® (labelled sample 23) and passed through a 3-kDa molecular weight cut-off (MWCO) filter inhibited MAGL by 63.19% when assayed at a concentration of 1 mg/mL in 1% DMSO and water. The positive control, the compound JZL 195, inhibited MAGL by 98.16% when assayed at a concentration of 4.4 µM. All other extracts did not significantly inhibit MAGL when assayed at a similar concentration. These hydrolysates were selected for further characterisation using mass spectrometry and in silico analysis.
2.4.3. 2,2′-Azino-bis-3-ethylbenzthiazoline-6-sulphonic Acid Total Antioxidant Capacity (ABTS) of Mesopelagic Hydrolysates
The hydrolysates generated from Spanish mesopelagic fish - M. muelleri hydrolyzed with Alcalase 2.4 L (MME02 and MMD02) and M. muelleri hydrolyzed by endogenous enzymes (MMC019) displayed antioxidant activities as well as the Irish hydrolysate generated from Benthosema glaciale hydrolyzed with Alcalase® (sample 2, Figure 5). These hydrolysates quenched the ABTS radical maximally compared to the positive control Resveratrol® and had ABTS-TAC values of 325 (± 24.79) µM TE/mg; 330 µM TE/mg and 210 µM TE/mg, respectively. Resveratrol quenched the radical by 330 µM TE/mg (Figure 5). These hydrolysates were selected for further characterisation using mass spectrometry and in silico analysis.
2.4.4. Angiotensin-1-Converting Enzyme (ACE-1) and Renin Inhibition by Mesopelagic Hydrolysates
All hydrolysates generated from Irish mesopelagic fish hauls including Benthosema glaciale biomass hydrolysed with Alcalase®; Notocopelus elongtus kroyeri hydrolysed with Alcalase®; M.muelleri biomass hydrolysed with Alcalase® and M. muelleri mixed biomass hydrolysed with Alcalase® inhibited ACE-1 by > 95% when assayed at a concentration of 1 mg/mL compared to the positive control, assayed at a concentration of 0.5 mg/mL that inhibited ACE-1 by 100%. Specifically, for M. muelleri biomass hydrolysed with Alcalase® the ACE-I IC50 was 27 µg/ml. These results are quite promising compared to other published ACE inhibition bioactivities of marine hydrolysates as, for instance, salmon protein hydrolysate (IC50 of 20-140 µg/mL), manila clams (IC50 of 420 µg/mL) or tuna frame protein hydrolysate (IC50 of 2 mg/mL) . Inhibition of the enzyme Renin by mesopelagic hydrolysates is shown in Figure 6. Mesopelagic hydrolysates inhibited Renin by between 20-59% (Figure 6) when assayed at a concentration of 1 mg/mL. MMC19 (endogenous enzymes) and MMD02 (Alcalase 2.4 LG) gave results comparable to commercial controls SB and SP.
2.4.5. Dipeptidyl Peptidase IV Inhibition by Mesopelagic Hydrolysates
Generated mesopelagic hydrolysates were assessed for their ability to inhibit DPP-IV at concentrations of 1 mg/mL using Sitagliptin as a positive control. Only the extract generated from Benthosema glaciale biomass using Alcalase® was found to inhibit DPP-IV by 17.33% when assayed at a concentration of 1 mg/mL compared to the positive control Sitagliptin that inhibited DPP-IV by 68.79%. The active hydrolysate was assessed further using MS and in silico analysis.
2.5. Identification of Bioactive Peptides Using Mass Spectrometry and In Silico Analysis
Hydrolysates identified as having bioactivities using in vitro bioassays described in section 4 were characterised following hydrolysate clean-up using MS. Bioactive hydrolysates included CE21009 Haul 23 Maurolicus muelleri (Code 23) Alcalase hydrolysate, an Irish sample; Maurolicus muelleri (MMC019) Endogenous enzyme autolysis (Spanish sample), spray dried and Maurolicus muelleri (MME02 - Spanish haul) alcalase hydrolysate. 1724, 95 and 182 peptide sequences were identified from each hydrolysate respectively. These peptides had homology with several different proteins of different origins as shown in the supplementary data. Following identification, peptides were ranked in terms of their potential to be bioactive using an in silico approach expanded upon from previously published work by our group [32,33]. Peptide ranker was used to predict potential bioactivity of peptide sequences based on amino acid charge and peptide structure as described previously [27]. The freely available programme PreAIP available at http://kurata14.bio.kyutech.ac.jp/PreAIP/ was used to predict the collective anti-inflammatory activity of peptides identified in each hydrolysate and anti-inflammatory potential of individual peptides. A search for sequenced peptides in the database BIOPEP-UWM was used to confirm the novel nature of the mesopelagic fish species derived peptides and Umami-MRNN https://umami-mrnn.herokuapp.com/ was used to predict if peptides would impart umami flavor or not. The database service AntiDMPpred available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl was used to identify the anti-diabetes type 2 activity of individual peptides [34]. The 10 most bioactive peptides found in each hydrolysate are shown in Table 1. The hydrolysate generated from Maurolicus muelleri using Alcalase (Irish mesopelagic fish sample) was found to contain peptides from several different proteins. Only three peptide sequences in this hydrolysate predicted to be anti-inflammatory and to impart Umami flavor were derived from fish proteins. These include the peptide FDAFLPM with a Peptide Ranker score of 0.955, predicted to be a medium anti-inflammatory agent and with no Umami potential derived from Thin-lipped mullet proteins. The peptide QCPLHRPWAL identified as having high potential anti-inflammatory activity (PreAIP RF value of 0.499) has amino acid homology with a protein derived from Atlantic salmon (Salmo salar) and the peptide NVGEVVCIFLTAALGLPEALI predicted to be highly anti-inflammatory (PreAIP RF value of 0.612) and also predicted to have anti-diabetic potential using AntiDMPpred (probability value of 0.8) has sequence homology with a protein from Makaira nigricans commonly known as Atlantic Blue Marlin. The anti-inflammatory potential of the 10 most bioactive peptides generated in this hydrolysate was also predicted to be significant with a PreAIP RF value of 0.482 (high confidence anti-inflammatory). The hydrolysate generated from Maurolicus muelleri (MMC019) with only endogenous enzyme autolysis (Spanish sample) and subsequently spray dried also has predicted anti-inflammatory activities with a combined PreAIP value of 0.514 for the 10 most bioactive active peptides identified using Peptide Ranker. Individually, these peptides had predicted anti-inflammatory potential values ranging from 0.540 (identified using PreAIP RF) for the peptide IAGFEIFDFNSLEQLC, derived from fish proteins from the species Cypinus carpio (Common carp) to 0.284 (not anti-inflammatory according to PreAIP RF) for the peptide GFAGDDAPR derived from fish proteins identified with homology to proteins derived from Takifuga rubipes (Japanese puffer fish). The hydrolysate generated from Maurolicus muelleri (MME02 - Spanish haul) using Alcalase enzyme had a medium confidence AIP with a PreAIP RF value of 0.446. The peptide SDNAYQFMLT identified as having sequence homology to proteins derived from Common Carp is predicted to have medium anti-inflammatory activity with a PreAIP value of 0.412. In addition, this peptide was predicted to impart Umami flavor with an umami, predicted threshold of 34.065384 mmol/L (Table 1).
2.6. Chemical Synthesis and Confirmation of Anti-Inflammatory Activity of Peptides Using In Vitro COX and MAGL Inhibition Bioassays
Peptide IAGFEIFDFNSLEQLC predicted to have anti-inflammatory and Umami imparting bioactivities and with sequence homology to a protein derived from Cypinus carpio (Common carp); QCPLHRPWAL with homology to a protein from Atlantic salmon and predicted anti-inflammatory activities, SDNAYQFMLT with predicted anti-inflammatory activity and sequence homology to a fish protein from Common carp, as well as the peptides PSRILYG, FDFLPM, GFNPPDLDIM and GFAGDDAPR were chemically synthesised by GenScript Biotech (Leiden, The Netherlands). GenScript also verified the purity of the peptides by analytical RP-HPLC–MS. COX-1 and COX-2 percentage (%) inhibition values, COX half-maximal Inhibitory Concentration (IC50) values and the selectivity index of selected peptides were determined for these synthesised peptides and values compared to the control, a commercially available COX inhibitor drug Diclofenac. The peptide IAGFEIFDFNSLEQLC inhibited COX-1 by 53.26% (+/-5.26) when assayed at a concentration of 1 mg/mL with a COX-1 IC50 value of 189.76 µg/mL (102.79 µM). This peptide inhibited COX-2 by 47.46% when assessed at a concentration of 1 mg/mL with a COX-2 IC50 value of 366.48 µM and a selectivity index value of 0.52. Peptide QCPLHRPWAL inhibited COX-1 by 82.90% (+/-0.54) with a COX-1 IC50 value of 2386.64 µM. This peptide inhibited COX-2 by 53.84% when assayed at a concentration of 1 mg/mL and displayed a COX-2 IC50 value of 220.66 µM with a selectivity index value of 10.81. Selectivity index (SI) refers to the ratio of the toxic concentration of a sample against its effective bioactive concentration. A SI value ≥ 10 shows potential that the peptide is non-toxic but bioactive and warrants further investigation [35]. Peptide SDNAYQFMLT inhibited COX-1 by 53.72% when assayed at a concentration of 0.5 mg/mL and peptide FDAFLPM inhibited COX-1 by 34.49% at the same concentration. The peptide NVGEVVCIFLTAALGLPEALI derived from Irish M. muelleri hauls and hydrolyzed with Alcalase® was also chemically synthesized and assessed for its ability to inhibit the MAGL enzyme. This peptide inhibited MAGL by 53.05% (+/-5.25) when assayed at a concentration of 0.5 mg/mL compared to the positive control, JZL 195 and had a MAGL IC50 value of 13.61 µg/mL (6.35 µM).
3. Discussion
Recently Rivera-Jiménez and colleagues collated an excellent review concerning peptides and protein hydrolysates exhibiting anti-inflammatory activities and detailed different sources, structural features and modulation mechanisms for the limited number of anti-inflammatory hydrolysates from fish that are reported in the literature [36]. The review details peptides of fish origin derived from salmon skin and pectoral fin hydrolysates of the species Salmo salar as well as peptides from hydrolysates of herring millet, mollusk, abalone and oyster. All listed fish-derived anti-inflammatory peptides regulated inflammation by inhibition of nitric oxide (NO) production, inhibition of IL-6 and TNF-α [36]. Peptides identified from bivalve visceral mass and salmon bones were COX-2 inhibitors with inhibition values ranging from 45% -58% and NO IC50 values of 54.07 µg ml-1 reported for the peptide PFNEGTFAAS derived from mollusk abalone [37]. No anti-inflammatory peptides that work by inhibition of MAGL are reported in this paper or in databases like BIOPEP-UWM. In addition, confirmation of anti-inflammatory activities by synthesis of COX-2 peptides are not reported in this review.
Herein, we have identified seven novel bioactive peptides with unique amino acid sequences derived from mesopelagic fish M. muelleri and Benthosema sp. hydrolyzed with the enzyme Alcalase. COX-1, COX-2 and MAGL inhibitory activities were determined for all seven peptides. All synthesized peptides inhibited COX-1 and COX-2 by between 34-54% when assayed at 1 mg/mL concentrations and peptide IAGFEIFDFNSLEQLC had COX-1 and COX-2 values of 102.79 µM and 366 µM respectively. Only one peptide, peptide NVGEVVCIFLTAALGLPEALI derived from Irish M. muelleri hydrolyzed with Alcalase inhibited the enzyme MAGL and had an MAGL IC50 value of 6.35 µM. The COX and MAGL inhibitory potential of identified peptides compare favorably with previously identified anti-inflammatory peptides like LREMLSTMCTARGA, AVGPGPRG and VPWGPWPKG from a bromelain hydrolysate of sea cucumber that inhibited NO in murine macrophages with reported NO IC50 values of 572.096 mg/mL and 674.435 mg/mL respectively [38]. Previously, anti-inflammatory peptides from pure amaranth with the peptide sequences HGSEPFGPR and RPRYPWRYT were found to reduce LOX-1 expression and these peptides had IC50 values comparable to the IC50 values determined against COX-1 and COX-2 for the mesopelagic derived peptides [39]. The peptides are not as active as natural products derived from Chinese herbs such as the compounds Acetyl-11-keto-β-boswellic acid, β-boswellic acid, acetyl-α-boswellic acid, acetyl-β-boswellic acid, known COX-1 selective inhibitors with IC50 values of approximately 10 μM. The Chinese medicine derived compounds Senkyunolide O and cryptotanshinone, known COX-2 selective inhibitors were reported to have COX2- IC50 values of 5 μM and 22 μM, respectively previously [40]. QCPLHRPWAL inhibited COX-1 by 82.90% (+/-0.54) and COX-2 by 53.84% with a selectivity index value of 10.81. This peptide, along with peptide NVGEVVCIFLTALGLPEALI warrant further examination in in vivo trials based on their IC50 values, novelty and selectivity index values. The whole hydrolysates generated from mesopelagic fish species, independently, did not display COX-1 or COX-2 inhibitory activities greater than 50% when assayed at concentrations of 1 mg/mL. However, In silico analysis work applied to the generated hydrolysates using PreAIP suggest that they too are anti-inflammatory and this activity could be enhanced by generating molecular weight cut off (MWCO) fractions to enrich for the listed bioactive peptides in Table 1. The Maurolicus muelleri alcalase hydrolysate inhibited MAGL when assayed at a concentration of 1 mg/mL by 62% and was the only hydrolysate to do so. The MAGL inhibitory peptide NVGEVVCIFLTAALGLPEALI had an MAGL IC50 value of 6.35 µM. No fish derived peptide inhibitors of MAGL are reported in the literature or databases like BIOPEP-UWM currently. The discovery of this peptide and its IC50 value are novel and highly active. Previously, Jha and colleagues reported the discovery of two compounds with promising MAGL inhibitory potency as they had IC50 values below 50 µM [41]. In addition, this peptide was found to have potential to impact on the development of Type-2-diabetes as shown in Table 1, where it was the only peptide identified as having anti-diabetic potential using in silico analysis with the programme AntiDMPred. In addition, this peptide had an umami threshold value of 19.895546 mmol/L. A typical umami peptide such as –Lys-Gly-Asp-Glu-Glu-Ser-Leu-Ala (HKGDFFSLA) derived from beef gravy, has an umami threshold value of 36.730614 mmol/L when assessed using umami-MRNN [42]. Indeed, several of the peptides identified in the mesopelagic hydrolysates that are bioactive and have predicted anti-inflammatory and anti-diabetic activity also are predicted to impart umami flavours with values predicted for different peptides of between 1.6841059 mmol/L for peptide GFAGDDAPR to 37.667642 mmol/L for peptide LACNCNLHARRCRFNM. Umami, derived from the Japanese language, refers to a “delicious savory taste” and is the fifth primary taste sense [43]. In the development of pet foods, palatability is important as if dogs/cats won’t eat nutritionally balanced pet food, they will lack a healthy diet. Cats generally like meaty, umami flavours as they are obligate carnivores and research has shown that they possess glutamate receptors [44]. Identified novel peptides have potential for use in topical applications, in functional foods or supplements however, in vivo confirmation of bioactivities is required.
4. Materials and Methods
4.1. Supply and Processing of Raw Materials
The mesopelagic samples used in this work were supplied from different experimental cruises carried out by Norwegian (Pelagia), Irish (Marine Institute) or Spanish (AZTI and the Spanish Institute of Oceanography (IEO) partners, respectively as part of the MEESO (EU Horizon 2020; grant agreement No 817669) and SUMMER (EU Horizon 2020; grant agreement No 817806).
4.1.1. Irish and Norwegian Samples
Samples of Mueller’s Pearl side (Maurolicus muelleri Pearl side) and Blue Whiting species were supplied by the Irish Marine Institute and BIM and subsequently processed and analyzed by Teagasc. These hauls were composed of different mesopelagic species depending on the Haul date and were the result of the Western European Shelf Pelagic Acoustic Survey (WESPAS) and Blue Whiting oceanographic campaign conducted in March, April and May 2021 (Table 2).
Figure 7.
Haul samples from WESPAS survey containing Notocopelus elongtus kroyeri. Blue whiting (not analyzed as part of this work), Benthosema glaciale and Maurolicus muelleri.
Figure 7.
Haul samples from WESPAS survey containing Notocopelus elongtus kroyeri. Blue whiting (not analyzed as part of this work), Benthosema glaciale and Maurolicus muelleri.

Enzymatic Hydrolysis of Irish and Norwegian Samples
Hydrolysis of Irish mesopelagic species (Haul codes 2, 13, 14, and 23) were performed independently using a New Brunswick 1 L bioreactor (Mason Technology, Dublin, Ireland) with temperature, stirring (RPM) and pH control. The enzyme Alcalase CLEA, was added individually at an enzyme to substrate ratio of 1:100 (v:w) (1%) and the optimum conditions for the enzyme was maintained – temperature 60 °C, pH 7 (maintained using 1 M NaOH or 1 M HCL) and rpm 250 for 4 h. The enzyme was deactivated by heating the hydrolysates to 95 °C for 15 min in a water bath (Grant JB Aqua 12, Grant instruments, Cambridgeshire, UK). Hydrolysis was performed in triplicate. The degree of hydrolysis was determined by monitoring, the quantity of 1 M NaOH consumed during the course of the reaction and the equation:
Degree of hydrolysis percentage (DH %) =β×Nb×1/α×1/Mp×1/htot×100%, where;
β is the volume of base (mL) consumed; Nb is the normality of the base; α is the correction factor, which, refers to the average degree of dissociation of α-NH2 groups during the hydrolysis and was determined for each enzyme according to the Adler–Nissen method ; Mp represents the mass in g of protein; and the parameter htot refers to the total number of peptide bonds in a protein (meqv g− 1 protein). This value is determined from the amino acid composition of the protein and is 8.6 for fish protein.
Hydrolysates were generated from mesopelagic species caught in Norwegian waters using enzymes including Bromelain, Roholase, Corolase and MaxiPro using conditions optimized for each enzyme at Nofima. These hydrolysates were assessed for bioactivities as shown in later sections. The hydrolysis processes were performed at laboratory scale using a Symphony 7100 Bathless Dissolution Distek equipment (Distek Inc., North Brunswick, NJ, USA). In all the processes the temperature, time and stir speed were controlled and monitored. Following hydrolysis, enzymes were inactivated by heat treatment at 95°C for a period of 15 min.
Hydrolysate Enrichment Using Molecular Weight Cut-Off (MWCO) Filtration
Following hydrolysis of Irish mesopelagic species with Alcalase CLEA, 3-kDa fractions were generated using molecular weight cut-off (MWCO) filters 3-kDa (Millipore Carrigaline, Co. Cork, Ireland) with the Prep/Scale™-TFF Cartridge Holder system where, pressure was maintained at 20 psi at room temperature for 1 h. These fractions were labelled as 3-kDa permeates. All fractions were frozen, freeze-dried and stored at −20 °C until further use. Hydrolysates were freeze-dried for subsequent processing and analysis. Hydrolysates and permeates were freeze-dried in an industrial scale freeze-drier FD 80 model at Teagasc (Cuddon Engineering, New Zealand).
Proximate Compositional Analysis of Mesopelagic Species
The total protein content of whole mesopelagic fish, hydrolysates and permeates was determined using the LECO FP328 Nitrogen determination method (LECO Corporation) based on the Dumas method of nitrogen determination and according to the AOAC method 992.15, 1990 [45]. Ash content was determined using a carbolite muffle furnace according to the method of Pearson. The lipid content was quantified using AOAC Method 2008.06 with an Oracle Rapid NMR Fat Analyzer, (CEM Corporation, Matthews, NC, USA) as described previously [46].
4.1.2. Spanish Samples
Samples supplied and subsequently analyzed by AZTI were composed exclusively of Maurolicus muelleri species that resulted from two JUVENA oceanographic campaigns conducted in September 2019 and 2020 carried out by AZTI and the Spanish Institute of Oceanography (IEO) for the estimation of the stock of the anchovy’s fisheries in the Bay of Biscay. The samples were frozen and glazed on board the ship and different quantities of biological material were kept in separate bags, for compositional analysis, and hydrolysis assays. All samples were stored at -18 °C at AZTI until processing. Before processing fish samples were pooled to achieve enough quantity to perform all the hydrolysis trials with a same material composition.
Enzymatic Hydrolysis of Spanish Samples
The hydrolysis processes were performed at laboratory scale using a Symphony 7100 Bathless Dissolution Distek equipment (Distek Inc., North Brunswick, NJ, USA). In all the processes the temperature, time and stir speed were controlled and monitored. Following hydrolysis, enzymes were inactivated by heat treatment at 95°C for a period of 15 min. Bioreactor contents were sieved to separate the bones, and centrifuged (2650 ×g; 15 min; ambient temperature) to separate 3 different layers; 1) an oil-water emulsion (top-layer), 2) a water-based fraction (the protein hydrolysate – the middle layer) and 3) a solid pellet (bottom layer). Hydrolysates were freeze-dried for subsequent processing and analysis.
Four different enzymes, with different enzymatic activity, where tested to produce protein hydrolysates: Protamex® (NOVOZYMES, Bagsvaerd, DK) a broad-spectrum endo-protease, Alcalase® 2.4 L FG (NOVOZYMES, Bagsvaerd, DK) an endo-protease of the serine type, Papain P144GL/100 (90 – 110 u/g) (Biocatalysts Ltd., Cardiff, UK), Bromelain P523MDP/2000 (1,800 – 2,200GDU/g) (Biocatalysts Ltd., Cardiff, UK) and a combination of Papain plus Bromelain. A control (CTR) was added to the trials, which was carried out without enzyme addition at pH 6.0, 50 °C and 3 h.
The pH of each run of the experimental design was controlled manually and adjusted with NaOH 1 M in a final volume of 500 mL (250 g of fish + 250 g water). All the processes were carried out with 1% enzyme (protein based), for 3 h at 250 rpm and at the optimum pH and temperature of each enzyme or enzyme combinations (Table 1).
Analysis of Spanish Samples
Chemical analysis
The chemical composition of the fractions resulting from the hydrolysis was measured by applying the Association of Official Analytical Chemists (AOAC) Official Methods (AOAC, 2007), dry matter (method 934.01), ash (method 942.05) and nitrogen (method 955.04), using a nitrogen-to-protein conversion factor of N x 6.25 was applied for crude protein content determination.
Protein extraction yield (PEY %) was determined as follow (Eq. 1);
PEY (%) = (Protein content in the hydrolysate (g)/ (Protein content in the initial sample (g) x 100 (Eq. 1)
Degree of Hydrolysis
The O-phthaldialdehyde (OPA) method was applied to quantify the degree of hydrolysis (DH %) [46]. Briefly, 3.7% w/v Na tetraborate decahydrate and 0.58% w/v Nadodecyl-sulphate aqueous solution was mixed with 4% (w/v) methanolic solution of OPA in a proportion of 51.5:1. Then, 0.38% v/v of mercaptoethanol was incorporated into the solution. The sample solutions were standardized to 0.01 g protein/L. 60 L of sample and 180 L of OPA reagent were dosed in microplate wells. They were incubated at ambient temperature for five minutes. The determinations were done at 360 nm excitation wavelength and 460 nm emission wavelength. N_-Acetyl-L-lysine was used as standard. DH was calculated as described by Nielsen, Petersen and Dambmann (2001) [47]; Where h is the number of hydrolyzed bonds. htot, defined as the total number of peptide bonds per protein equivalent, was used as 8.6 mg equivalent/g protein, and α and β were 1.00 and 0.40.
DH = h/htot * 100 (Eq. 2)
h = (serine-NH2 - β)/ α megv/g protein (Eq. 3)
Molecular weight distribution
Size exclusion high performance liquid chromatography (SE-HPLC) was performed to assess the peptide molecular weight distribution. Samples were analysed using AdvanceBio SEC LC column (130 Å, 7.8 x 150 mm, 2.7 µm) (Agilent Technologies, Spain) connected to a diode array detector (DAD) (Agilent Technologies, Spain). 5 μL of sample were injected onto the column kept at room temperature. The mobile phase consisted in pH 7 sodium phosphate buffer (150 mM). Samples were eluted at a flow rate of 0.5 mL· min−1 and UV signal was measured at 220 nm. Each sample was filtered through 0.45 μm PVDF filter. AdvanceBio SEC 130 Å Protein Standard (Agilent Technologies, Spain) was used to determine the elution time depending on the weight of the molecules. Every sample was diluted to a protein concentration of 3 g·L-1.
4.2. Bioactivity Screening of Hydrolysates and Permeates
Irish, Norwegian and Spanish hydrolysates were assessed for different bioactivities that may potentially affect inflammation and pain, heart health, the development of type 2 diabetes (T2D) and antioxidant activities.
4.2.1. Cyclooxygenase (COX; EC E.C. 1.14. 99.1) Inhibition COX-1 and COX-2
The anti-pain and anti-inflammatory bioactivities of generated mesopelagic hydrolysates and permeates were assessed using a cyclooxygenase inhibition assay, targeting human COX-2 and ovine COX-1 enzymes, as described previously. COX (Prostaglandin H synthase) inhibition plays a positive role in prevention of a variety of health conditions and COX inhibitors have documented analgesic, anti-inflammatory, and antipyretic properties. They can influence pain and inflammation related to diseases like osteoarthritis and rheumatoid arthritis, musculoskeletal injury, migraines, and cancer. COX inhibitors are indicated for mild pain and are usually prescribed for short durations of time to patients. Generated hydrolysates were screened at concentrations of 1 mg/ml for their ability to inhibit COX-1 and COX-2 using the human COX-2 enzyme and ovine COX-1 enzyme and by measuring the peroxidase activity colour metrically using a spectrophotometer and the appearance of N,N,N’,N’-tetramethyl-p-phenylenediamine (TMPD) at 590 nm in the presence and absence of the hydrolysate and a positive control (Resveratrol – COX-1) and X (COX-2) assayed at recommended concentrations in a 96 well plate format using the assay kit – COX colorimetric inhibition screening assay kit (Item No 701050) from Cayman Chemicals (Cayman chemicals, Ellsworth Rd., Ann Arbor, MI, USA). Resveratrol (Sigma, Dublin, Ireland) is the positive control for COX-1 inhibition. COX inhibition was calculated using the equation:
% COX inhibition = (Corrected 100% initial
activity – Corrected Inhibitor activity)/ (Corrected 100% initial
activity)*100
Where: Corrected 100% initial activity was the absorbance value recorded for wells containing 150 µl 1X assay Buffer, 10 µL of either COX-1 or COX-2 enzyme and 10 µL of solvent (DMSO) minus the absorbance value recorded for the background well containing 160 µL of 1 X Assay Buffer and 10 µL of solvent.
4.2.2. Monoacylglycerol Lipase (MAGL; EC 3.1. 1. 23) Inhibition
Monoacylglycerol lipase (MAGL) is a serine hydrolase that catalyzes the hydrolysis of monoglycerides into glycerol and fatty acids and links the endocannabinoid and eicosanoid systems through degradation of 2-arachidaoylglycerol to arachidonic acid, a precursor of prostaglandins and other inflammatory mediators. MAGL inhibitors can have anti-nociceptive, anxiolytic, anti-inflammatory, and even anti-cancer benefits. The Cayman’s Monoacylglycerol lipase inhibition screening assay kit was used to measure inhibition of MAGL by generated hydrolysates and the positive control JZL 195 inhibitor. The assay was performed in compliance with the manufacturer’s instructions. MAGL hydrolyses 4-Nitrophenylacetate resulting in a yellow product, 4-nitrophenol that has an absorbance between 405-412 nm. The disappearance of the yellow colour is indicative of MAGL inhibition. MAGL inhibition was calculated using the equation:
% MAGL inhibition = (corrected 100% initial
activity – corrected inhibitor activity)/ (corrected 100% initial activity)
*100
Where: Corrected 100% initial activity was the absorbance value recorded for wells containing 150 µl 1X assay Buffer, 10 µL of MAGL enzyme and 10 µL of solvent minus the absorbance value recorded for the background well containing 160 µL of 1 X Assay Buffer and 10 µL of solvent (1.4% DMSO in water).
4.2.3. ACE-I Inhibition Assay
This assay was performed independently on Irish hydrolysates generated at Teagasc and Spanish hydrolysates generated by AZTI and assayed by the Leitat Technological Centre, using an ACE-I inhibitor assay kit and in accordance with the manufacturer’s instructions (ACE Kit – WST assay kit from Dojindo Laboratories, Maryland, USA). All fractions were assayed at a concentration of 1 mg/mL HPLC grade water in triplicate and means and SD were calculated. The known ACE-I inhibitor Captopril was used as a positive control at a concentration of 1 mg/mL. Absorbance was measured with a FLUOstar Omega microplate reader (BMG LABTECH GmbH) at 450 nm.
4.2.4. DPP-IV Inhibition Assay
This assay was carried out following using a DPP-IV inhibitor screening assay kit and in accordance with the manufacturer’s instructions (Cayman Chemicals, USA). All hydrolysates (Irish, Spanish and Norwegian) were assayed in triplicate, and means and SD were calculated. The known DPP-IV inhibitor Sitagliptin (Merck, Dublin, Ireland) was used as a positive control. Fluorescence intensity was recorded with a FLUOstar Omega microplate reader (BMG LABTECH GmbH) using an excitation wavelength of 355 nm and an emission wavelength of 460 nm.
4.2.5. Antioxidant Capacity: ABTS Assay
The ABTS (2, 2-azinobis-(3-ethylbenzthiazoline-6-sulfonic acid) assay adjusted to microplate volume was used to determine the antioxidant capacity of Spanish mesopelagic hydrolysates. The colorimetric results were measured in Varioskan™ LUX multimode microplate reader (Thermo Fisher Scientific, Waltham, MA, USA). In brief, 7 mM ABTS solution and 2.45 mM potassium persulfate were diluted in PBS (phosphate-buffered saline) for an absorbance of approximately 0.7 at 734 nm. The determination consisted in the decrease in absorbance at 734 nm of the reagent solution, 6 min after the sample was added in the micro-well in a ratio of 1:100 (v/v) (sample: ABTS solution), as the result of the reduction of the radical colored ABTS. Trolox was used as the standard, and the antioxidant capacity (AOC) was calculated as Trolox Equivalent Antioxidant Capacity (TEAC, µM TE/ mg of sample).
4.2.6. Renin Inhibition Activity
Renin Inhibitory activity of hydrolysates was carried out according to the manufacturers’ instructions. Briefly, prior to carrying out the assay, the renin assay buffer was diluted by adding 3 mL of 50 mM Tris-HCl, pH 8.0 buffer to 27 mL of HPLC grade water to give a final assay buffer concentration of 50 mM Tris-HCl (pH 8.0) containing 100 mM NaCl. The renin protein solution was diluted 20-fold with assay buffer before use, and the assay buffer was pre-warmed to 37 °C in accordance with the manufacturers’ instructions. Background wells were prepared by adding 20μL of substrate, 160μL of assay buffer, and 10μL of HPLC grade water. Negative control wells were prepared by adding 20μL of substrate, 150μL of assay buffer, and 10μL of HPLC grade water. Inhibitor wells were prepared by adding 20μL of substrate, 150μL of assay buffer, and 10μL of sample. The reaction was initiated by adding10μL of renin to the control and sample wells. Fluorescence intensity was recorded using an excitation wavelength of 340 nm and an emission wavelength of 490 nm. All hydrolysates were assayed at a concentration of 1 mg/mL in triplicate. The known specific renin inhibitor, Z-Arg-Arg-Pro-Phe-His-Sta-Ile-His-Lys-(Boc)-OMe, was used as a positive control. Percentage inhibition was calculated using the following equation:
Renin inhibition % = 100% initial activity (AF) - inhibitor (AF))/100% initial activity (AF)) 100)
Where; AF is the average fluorescence. Initial activity is the assay performed without the presence of an inhibitor.
4.3. Peptides Identified Using Mass Spectrometry
The most active hydrolysates identified using in vitro bioactivity assays included the Spanish Maurolicus muelleri hydrolysate generated with Alcalase 2.4 L FG (MME02) and the Spanish Maurolicus muelleri hydrolysate generated with endogenous enzymes (MMC019). Additionally, the Irish Maurolicus muelleri hydrolysed with Alcalase (23) displayed MAGL inhibitory activity. These hydrolysates were selected for subsequent mass spectrometry (MS) analysis. The 3-kDa MWCO permeate fractions generated from each bioactive hydrolysate were processed for mass spectrometry analysis using the Preomics Phoenix Clean-up Kit (96×), (Preomics, D-82152 Planegg/Martinsried, Germany) in accordance with the manufacturers’ instructions. Samples were acidified and hydrophobic and hydrophilic contaminants removed using a series of wash steps and peptides eluted from the cartridge and prepared in loading buffer for LC-MS/MS analysis. Peptides were identified using a mass spectrometer nanoESI qQTOF (6600 plus TripleTOF, AB SCIEX, Framingham, MA, USA) using liquid chromatography and tandem mass spectrometry (LC–MS/MS). A total of 1 μL of each permeate was loaded onto a trap column (3 µ C18-CL 120 Ᾰ, 350 μm × 0.5 mm; Eksigent, Redwood City, CA, USA) and desalted with 0.1% TFA (trifluoroacetic acid) at 5 µL/min for 5 min. The permeates were then loaded onto an analytical column (3 µ C18-CL 120 Ᾰ, 0.075 × 150 mm) equilibrated in 5% acetonitrile (ACN) 0.1% formic acid (FA). Elution was carried out with a linear gradient from 5 to 40% B in A for 20 min, where solvent A was 0.1% FA and solvent B was ACN with 0.1% FA) at a flow rate of 300 nL/min. The sample was ionized in an electrospray source of Optiflow < 1 μL Nano applying 3.0 kV to the spray emitter at 200 °C. Analysis was carried out in a data-dependent mode. Survey MS1 scans were acquired from 350 to 1400 m/z for 250 ms. The quadrupole resolution was set to ‘LOW’ for MS2 experiments, which were acquired from 100 to 1500 m/z for 25 ms in ‘high sensitivity’ mode. The following switch criteria were used: charge: 1+ to 4+; minimum intensity; 100 counts per second (cps). Up to 50 ions were selected for fragmentation after each survey scan. Dynamic exclusion was set to 15 s. The system sensitivity was controlled by analysing 500 ng of K562 protein extract digest (SCIEX); in these conditions, 1819 proteins were identified (FDR < 1percentage) in a 20 min gradient. Protein Pilot v 5.0. (SCIEX Framingham, MA, USA) default parameters were used to generate the peak list directly from 6600 plus TripleTOF wiff files. The Paragon algorithm of ProteinPilot v 5.0 was used to search the SwisProt and Uniprot database with the following parameters: no enzyme specificity, IAM cysalkylation, no taxonomy restriction, and the search effort set to throughout. Peptides were identified with a confidence of ≥ 95%.
4.4. Assessment of Bioactive Potential of Identified Peptides
The bioactive potential of each peptide was predicted using several in silico/AI methods, some of which were described previously by this group [48]. Briefly, Peptide Ranker tool (http://bioware.ucd.ie/~compass/biowareweb/ [49], accessed on 22 January 2024) and PreAIP (http://kurata1.bio.kyutech,ac.jp/PreAIP/) [50], were used to assess overall bioactivity and potential of the peptides to act as anti-inflammatory molecules. The novelty of identified peptides was determined by performing a literature and database search in BIOPEP-UWM (http://www.uwm.edu.pl/biochemia/index.php/en/biopep[51], accessed on 23rd of January 2024). The umami activity of selected peptides identified using mass spectrometry was assessed using Umami-MRNN https://umami-mrnn.herokuapp.com/ (accessed on 23/01/2024) and BIOPEP-UWM as described previously [50].
5. Conclusions
Result generated in this work corroborate the potential of mesopelagic fish as a source of nutrients but additionally as a reservoir for novel, bioactive peptide generation. Several unique, novel bioactive peptides were identified and their anti-inflammatory and heart health potential validated using in silico and in vitro bioassays. Additionally, the umami flavour potential of generated hydrolysates was assessed using in silico analysis of specific peptides. The bioactivities of generated hydrolysates are promising and offer potential for use in the treatment of inflammatory health conditions like psoriasis and others and may be commercially exploitable, however further in vivo work is required. The work demonstrates the potential of mesopelagic fisheries as a resource for generation of new, novel bioactive compounds for food and pharma applications.
Author Contributions
For research articles with several authors, a short paragraph specifying their individual contributions must be provided. The following statements should be used “Conceptualization, M.H., R.G.S. and C.B.; methodology, M.H., L.M., B.I., J. I., L. B., T. C., J. G., A. D., S.A., L. T., R. G.S; software, M.H validation, M.H. M.H.; formal analysis, M.H investigation, M.H resources, M.H data curation, M.H writing—original draft preparation, M.H writing—review and editing, M.H. visualization, M.H., supervision, M.H.; project administration, R.G.S; M.H.,.; funding acquisition, M.H., R. G. S., C. B. All authors have read and agreed to the published version of the manuscript.
Funding
This research has received funding from the European Union’s Horizon 2020 research and innovation programme under grant agreements No 817806 (SUMMER project) and No 817669 (MEESO project).
Institutional Review Board Statement
Not applicable as studies did not involve humans or animals.
Data Availability Statement
Further information is available on results from the MEESO database or directly from the corresponding author. All data is open access and available from the corresponding author.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Staby, A. , Røstad, A., Kaartvedt, S., Long-term acoustical observations of the mesopelagic fish Maurolicus muelleri reveal novel and varied vertical migration patterns. Marine Ecology Progress Series 2011, 441, 241–255. [Google Scholar]
- Naik, A.S., Whitaker, R.D., Albrektsen, S., Solstad, R.G., Thoresen, L., Hayes, M. Mesopelagic fish protein hydrolysates and extracts: A source of novel anti-hypertensive and anti-diabetic peptides. Front. Mar. Sci., 2021, 8, 1-9. [CrossRef]
- Abdulbasit, A. , Biochemical composition and bioactive properties of mesopelagic fish-derived protein hydrolysate from Northern Krill (Meganyctiphanes norvegica) and Muller’s Pearlside (Maurolicus muelleri) for possible utilization of novel marine resources, 2022, Master’s thesis in Biotechnology, Norwegian University of Science and Technology, Faculty of Natural Sciences, 1-70.
- Madina, M.A.; Grimaldo, E.; Grimsmo, L.; Toldnes, B.; Slizyte, R.; Carvajal, A.K.; Schei, M.; Selnes, M.; Falch, E. Exploring the Potential of Atlantic Mesopelagic Species Processed on Board Commercial Fishing Vessels as a Source of Dietary Lipids. Foods 2024, 13, 1094. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.H. , Kriest, I. , Getzlaff, J., Diving deeper: Mesopelagic fish biomass estimates comparison using two different models. Front. Mar. Sci. 2023, 10. [Google Scholar] [CrossRef]
- Calcinai L, Bonomini MG, Leni G, Faccini A, Puxeddu I, Giannini D, Petrelli F, Prandi B, Sforza S, Tedeschi, T. Effectiveness of enzymatic hydrolysis for reducing the allergenic potential of legume by-products. Sci Rep. 2022 Oct 7;12(1):16902. [CrossRef] [PubMed] [PubMed Central]
- Jiang, X.; Rao, Q. Effect of Processing on Fish Protein Antigenicity and Allergenicity. Foods 2021, 10, 969. [Google Scholar] [CrossRef]
- Gianfranceschi, G.L. , Gianfranceschi, G., Quassinti, L., Bramucci, M., Biochemical requirements of bioactive peptides for nutraceutical efficacy. Journal of Functional Foods, 2018; 47, 252–263Get rights and content. [Google Scholar] [CrossRef]
- Minkiewicz P, Iwaniak A, Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides. Current Opportunities. Int J Mol Sci. 2019, 20. [CrossRef] [PubMed] [PubMed Central]
- Lund, A., Knop, F.K., Vilsbøll, T., Glucagon-like peptide-1 receptor agonist for the treatment of type 2 diabetes: Differences and similarities. European Journal of Internal Medicine, 2014, 25, 407-414. [CrossRef]
- Magon, N. Gonadotropin releasing hormone agonists: Expanding vistas. Indian J Endocrinol Metab. 2011, 15, 261–267. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Rossino, G., Marchese, E., Galli, G., Verde, F., Finizio, M., Serra, M., Linciano, P., Collina, S. Peptides as Therapeutic Agents: Challenges and Opportunities in the Green Transition Era. Molecules. 2023, 28, 7165. [CrossRef] [PubMed]
- Chen, L., Deng, H., Cu,i H., Fang, J., Zuo, Z., Deng, J., Li, Y., Wang, X., Zhao, L. Inflammatory responses and inflammation-associated diseases in organs. Oncotarget. 2017, 9, 7204-7218. [CrossRef] [PubMed]
- Placha, D., Jampilek, J. Chronic Inflammatory Diseases, Anti-Inflammatory Agents and Their Delivery Nanosystems. Pharmaceutics. 2021, 13, 1, 64. [CrossRef] [PubMed] [PubMed Central]
- Bu, J., Ding, R., Zhou, L., Chen, X., Shen, E. Epidemiology of Psoriasis and Comorbid Diseases: A Narrative Review. Front Immunol. 2022, 10;13:880201. [CrossRef] [PubMed] [PubMed Central]
- Zhou, W.B.S., Meng, J., Zang, J. Does low-grade systemic inflammation have a role in chronic pain? Front. Mol. Neurosci., 2021, 14. [CrossRef]
- Bindu, S., Mazumder, S., Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem Pharmacol. 2020, 180, 114147. Epub 2020 Jul 10. [CrossRef] [PubMed]
- Smith, C.J., Zhang, Y., Koboldt, C.M., Muhammad, J., Zweifel, B.S., Shaffer, A., Talley, J.J., Masferrer, J.L., Seibert, K., Isakson, P.C. Pharmacological analysis of cyclooxygenase-1 in inflammation. Proc Natl Acad Sci U S A. 1998, 95, 13313-8. [CrossRef] [PubMed]
- Curtis, E., Fuggle, N., Shaw, S., Spooner, L., Ntani, G., Parsons, C., Corp, N., Honvo, G., Baird, J., Maggi, S., Dennison, E., Bruyère, O., Reginster, J.Y., Cooper, C. Safety of Cyclooxygenase-2 Inhibitors in Osteoarthritis: Outcomes of a Systematic Review and Meta-Analysis. Drugs Aging. 2019 36, 1, 25-44. [CrossRef] [PubMed]
- Seeram, N.P. , Momin, R.A., Nair, M.G., Bourquin, L.D. Cyclooxygenase inhibitory and antioxidant cyanidin glycosides in cherries and berries. Phytomedicine. 2001, 8, 362–9. [Google Scholar] [CrossRef] [PubMed]
- Allaire, M. , Al Sayegh, R., Mabire, M., Hammoutene, A., Siebert, M., Caër, C., Cadoux, M., Wan, J., Habib, A., Le Gall, M., de la Grange, P., Guillou, H., Postic, C., Paradis, V., Lotersztajn, S., Gilgenkrantz, H. Monoacylglycerol lipase reprograms hepatocytes and macrophages to promote liver regeneration. JHEP Rep. 2023, 8, 100794. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Liang, Q. , Chalamaiah, M., Liao, W., Ren, X., Ma, H., Wu, J. Zein hydrolysate and its peptides exert anti-inflammatory activity on endothelial cells by preventing TNF-α-induced NF-ĸβ activation. Journal of Functional Foods, 2022; 64, 103598. [Google Scholar] [CrossRef]
- Dia, V.P., Wang, W., Oh, V.L., de Lumen, B.O., Gonzalez de Mejia, E. Isolation, purification and characterisation of lunasin from defatted soybean flour and in vitro evaluation of its anti-inflammatory activity. Food Chem. 2009, 114, 1, 108-115. [CrossRef]
- Ahn, C.B., Cho, Y.S., Je, J.,Y. Purification and anti-inflammatory action of tripeptide from salmon pectoral fin byproduct protein hydrolysate. Food Chem. 2015, 1, 168:151-6. Epub 2014 Jun 2. 1, 168; 1. [CrossRef] [PubMed]
- Jang, H.L. , Shin, S.R., Yoon, K.Y. Hydrolysis conditions for antioxidant peptides derived from enzymatic hydrolysates of sandfish (Arctoscopus japonicus). Food Sci Biotechnol. 2017, 26, 5, 1191-1197. [CrossRef] [PubMed]
- Kemp, D.C. , Kwon, J.Y. Fish and Shellfish-Derived Anti-Inflammatory Protein Products: Properties and Mechanisms. Molecules, 2021, 26, 3225. [CrossRef] [PubMed]
- Mooney, C. , Haslam, N.J., Pollastri, G., Shields, D.C. Towards the Improved Discovery and Design of Functional Peptides: Common Features of Diverse Classes Permit Generalized Prediction of Bioactivity. PLoS ONE, 2012, 7, e45012. [CrossRef]
- Khatun, M.S. , Hasan, M.M., Kurata, H. PreAIP: Computational Prediction of Anti-inflammatory Peptides by Integrating Multiple Complementary Features. Front Genet. 2019, 129. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Qi, L., Du, J., Sun, Y., Xiong, Y., Zhao, X., Dan, D., Zhi, Y., Dang, Y., Gao, X. Umami-MRNN: Deep learning based prediction of umami peptide using RNN and MLP. Food Chem., 2023, 405, A, 134935. [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 1998. [Google Scholar]
- Deng, H., Li, W. Monoacylglycerol lipase inhibitors: modulators for lipid metabolism in cancer malignancy, neurological and metabolic disorders. Acta Pharm Sin B. 2020, 10, 4, 582-602. Epub 2019 Oct 18. [CrossRef] [PubMed]
- Hayes, M., Mora, L., Lucakova, S. Identification of Bioactive Peptides from Nannochloropsis oculata Using a Combination of Enzymatic Treatment, in Silico Analysis and Chemical Synthesis. Biomolecules. 2022, Dec 2;12(12):1806. [CrossRef] [PubMed]
- Lafarga, T. , Aluko, R.E., Rai, D.K., O’Connor, P., Hayes, M. Identification of bioactive peptides from a papain hydrolysate of bovine serum albumin and assessment of an antihypertensive effect in Spontaneously Hypertensive Rats. Food Research International 2016, 81, 91–99. [Google Scholar] [CrossRef]
- Chen, X., Huang, J., He, B. AntiDMPpred: a web service for identifying anti-diabetic peptides. PeerJ. 2022, 10:e13581. [CrossRef] [PubMed]
- Ju, Z., Li, M., Xu, J., Howell, D.C., Li, Z., Chen, F.E. Recent development on COX-2 inhibitors as promising anti-inflammatory agents: The past 10 years. Acta Pharm Sin B. 2022, 12, 2790-2807. Epub 2022 Jan 11. [CrossRef] [PubMed]
- Rivera-Jiménez, J. , Berraquero-García, C., Pérez-Gálve, R., García-Moreno, P.J., Espejo-Carpio, F.J., Guadix, A., and Guadix, E.M. Peptides and protein hydrolysates exhibiting anti-inflammatory activity: sources, structural features and modulation mechanisms. Food and Function, 2022, 13, 12510. [Google Scholar]
- Shin, T.-S. , Choi, K.S., Chun, J., Kho, K.H., Son, S.A., Shim, S-Y. Anti-inflammatory effect of Abalone (aliotis discus Hannai) Viscera via inhibition of ROS production in LPS stimulated RAW 267.4 cells. Microbiol. Biotechnol. Lett. 2022, 50, 1, 21–30. [Google Scholar] [CrossRef]
- Ghanbari, R. , Ebahimpour, M., Zarei, A., Ismail, A., Abdul-Hamid, A., and Saari, N., Purification and Characterisation of Nitric Oxide Inhibitory peptides from Actinopyga lecanora through enzymatic hydrolysis. Food Biotechnol., 2016, 30, 263–277. [Google Scholar]
- Grancieri, M. , Martino, H.S.D., Gonzalez de Mejia, E. Digested total protein and protein fractions from chia seed (Salvia hispanica L.) had high scavenging capacity and inhibited 5-LOX, COX-1-2, and iNOS enzymes. Food Chem., 2019, 289, 204–214. [Google Scholar] [PubMed]
- Cao, H. , Yu, R., Choi, Y., Ma, Z.Z., Zhang, H., Xiang, W., Lee, D.Y., Berman, B.M., Moudgil, K.D., Fong, H.H., van Breemen, R., B. Discovery of cyclooxygenase inhibitors from medicinal plants used to treat inflammation. Pharmacol Res. 2010, 61, 6, 519–24. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Jha, V., Biagi, M., Spinelli, V., Di Stefano, M., Macchia, M., Minutolo, F., Granchi, C., Poli, G., Tuccinardi, T. Discovery of Monoacylglycerol Lipase (MAGL) Inhibitors Based on a Pharmacophore-Guided Virtual Screening Study. Molecules. 2020 26;26(1):78. [CrossRef] [PubMed]
- Zhang, Y. , Venkitasamy, C., Pan, Z., Liu, W., Zhao, L. Novel Umami Ingredients: Umami peptides and their taste. Journal of Food Science 2017, 82, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Mei, S., Yao, S., Mo, J., Wang, Y., Tang, J., Li, W., Wu, T. Integration of cloud-based molecular networking and docking for enhanced umami peptide screening from Pixian douban. Food Chem X. 2023, 27, 21:101098. [CrossRef] [PubMed]
- Belloir, C. , Savistchenko, J., Neiers, F., Taylor, A.J., McGrane, S., Briand, L. Biophysical and functional characterization of the N-terminal domain of the cat T1R1 umami taste receptor expressed in Escherichia coli. PLoS ONE, 2017, 12(10): e0187051. [CrossRef]
- Association of Official Analytical Chemists. Official Methods of Analysis of AOAC International; AOAC International: Rockville, MD, USA, 1998. [Google Scholar]
- Hoyle, N.T.; Merritt, J.H. Quality of fish protein hydrolysates from herring (Clupea harengus). J. Food Sci. 1994, 59, 76–79. [Google Scholar] [CrossRef]
- Nielsen, P.H. , Petersen, D., Dambmann, C. Food Science, 2001, Improved method for determining food protein degree of hydrolysis. 66, 5, 642-646.
- Hayes, M.; Aluko, R.E.; Aurino, E.; Mora, L. Generation of Bioactive Peptides from Porphyridium sp. and Assessment of Their Potential for Use in the Prevention of Hypertension, Inflammation and Pain. Mar. Drugs 2023, 21, 422. [Google Scholar] [CrossRef]
- Mooney, C. , Haslam, N., Pollastri, G., Shields, D. Towards the improved discovery and design of functional peptides: common features of diverse classes permit generalized prediction of bioactivity. PloS One 2012, 7(10). [Google Scholar]
- Khatun MS, Hasan MM, Kurata, H. PreAIP: Computational Prediction of Anti-inflammatory Peptides by Integrating Multiple Complementary Features. Front Genet. 2019 Mar 5;10:129. [CrossRef] [PubMed]
- Minkiewicz, P. , Iwaniak, A., Darewicz, M., BIOPEP-UWM database of bioactive peptides: Current opportunities. International Journal of Molecular Sciences, 2019, 20, 5978. [CrossRef]
- Qi, L. , Du, J., Sun, Y., Xiong, Y., Zhao, X., Pan, D., Zhi, Y., Dang, Y., Gao, X. 2023, Umami-MRNN: Deep learning-based prediction of umami peptide using RNN and MLP. Food Cheimstry, 405, A, 134935.
Figure 1.
Distribution of oil, hydrolysate, solids and bones generated using different enzyme combinations applied to Spanish mesopelagic trawls consisting of the species M. muelleri. Error bars represent SD (n = 3). Different letters indicate where significant difference exists between samples at 95% confidence.
Figure 1.
Distribution of oil, hydrolysate, solids and bones generated using different enzyme combinations applied to Spanish mesopelagic trawls consisting of the species M. muelleri. Error bars represent SD (n = 3). Different letters indicate where significant difference exists between samples at 95% confidence.
Figure 2.
Molecular weight distribution of hydrolysates generated using different proteolytic enzymes applied to mesopelagic trawls composed primarily of M. muelleri.
Figure 2.
Molecular weight distribution of hydrolysates generated using different proteolytic enzymes applied to mesopelagic trawls composed primarily of M. muelleri.
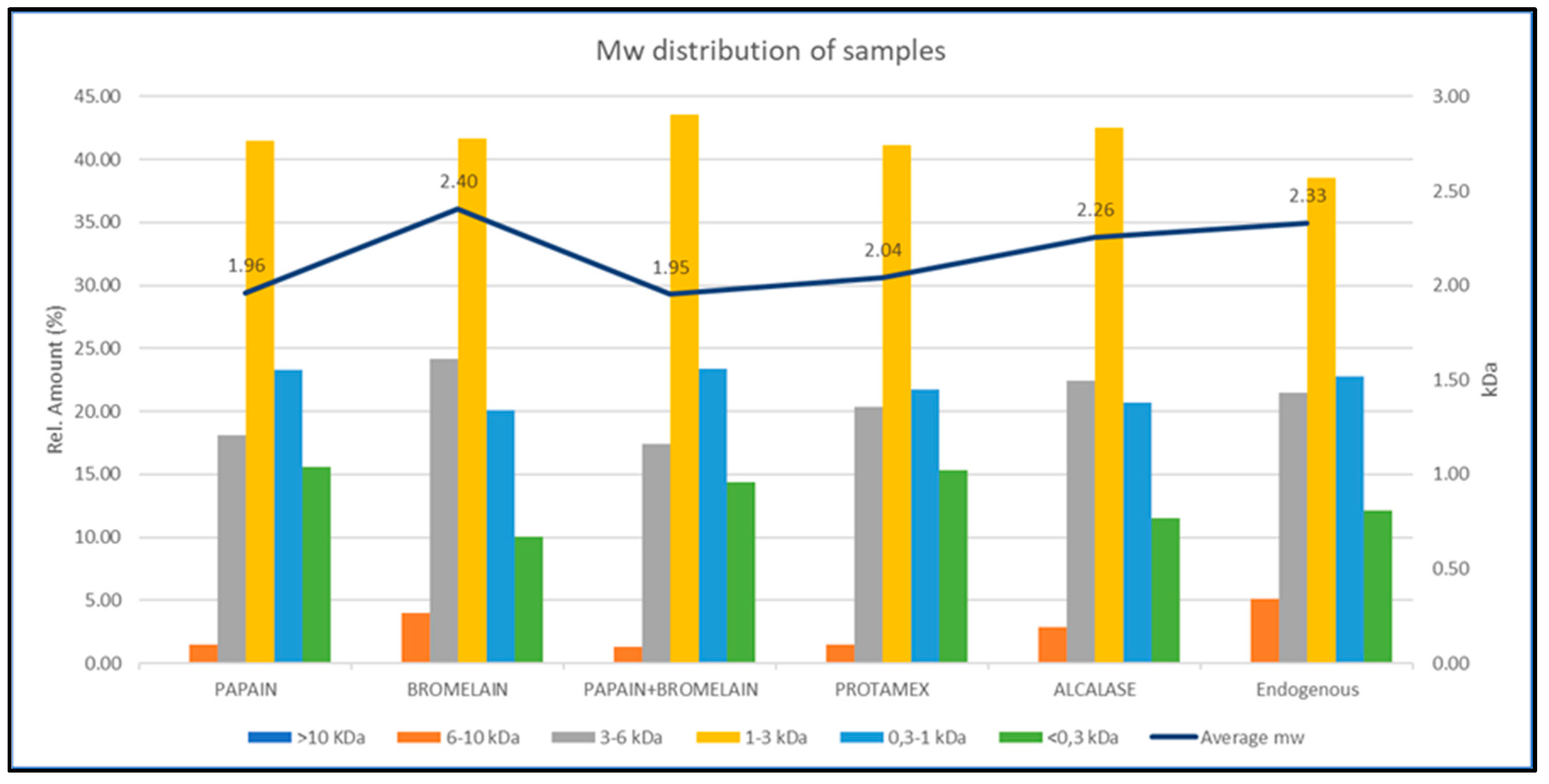
Figure 3.
COX-1 inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed at a concentration of 1 mg/mL and compared to a commercial control. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
Figure 3.
COX-1 inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed at a concentration of 1 mg/mL and compared to a commercial control. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
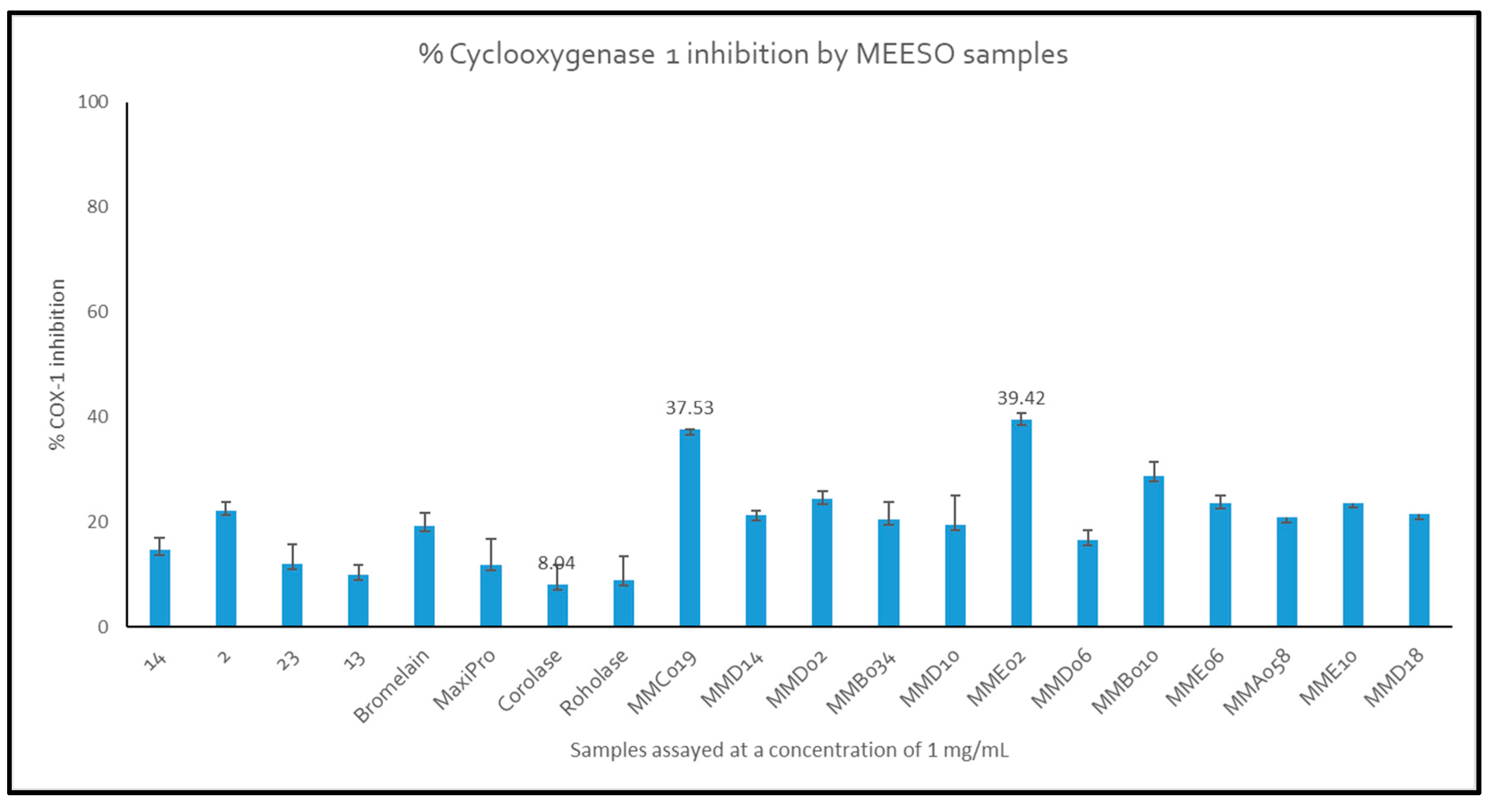
Figure 4.
COX-2 inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed at a concentration of 1 mg/mL and compared to a commercial control, Resveratrol. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
Figure 4.
COX-2 inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed at a concentration of 1 mg/mL and compared to a commercial control, Resveratrol. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
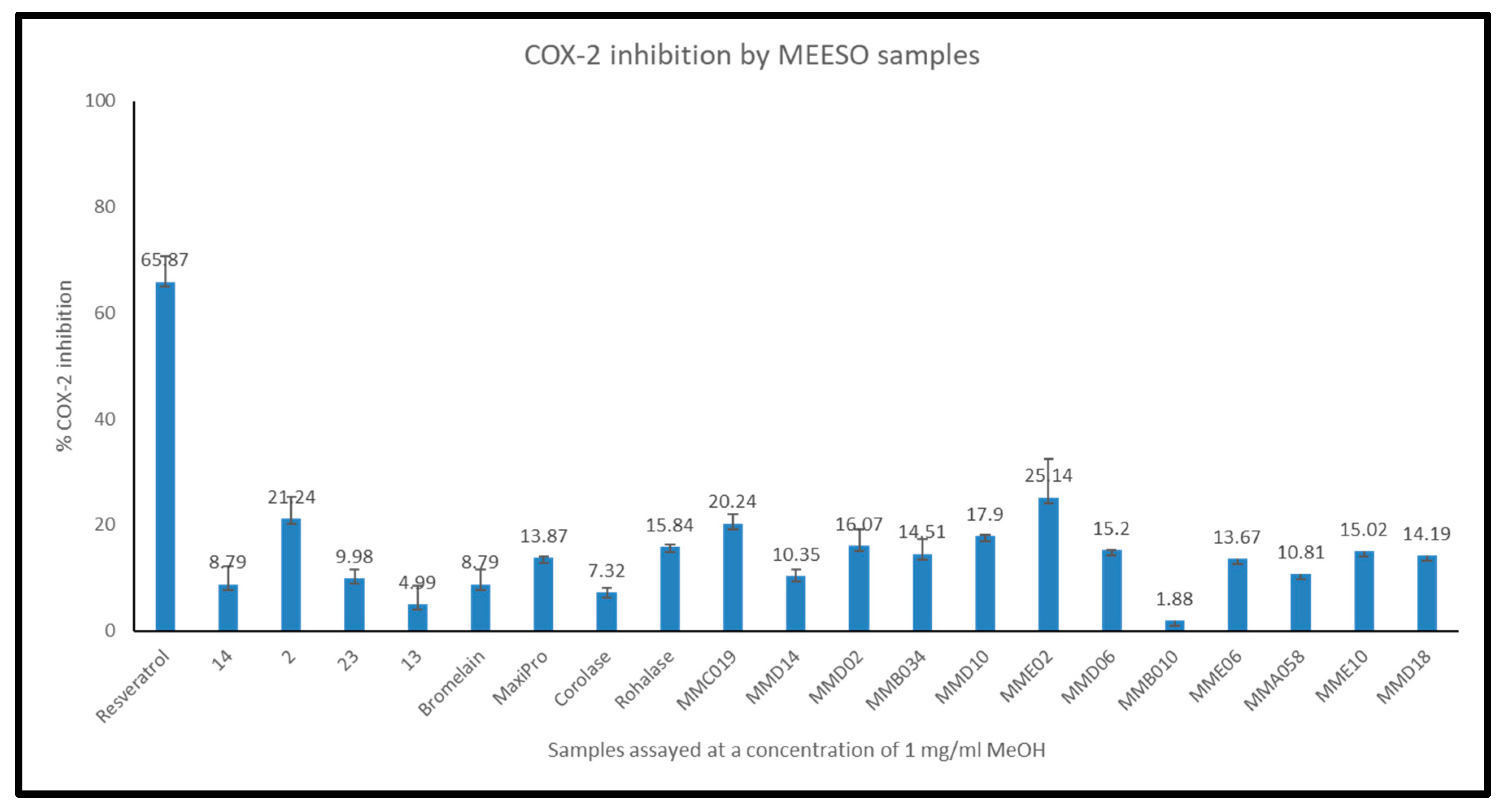
Figure 5.
Antioxidant activity of whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed using the ABTS antioxidant assay at a concentration of 1 mg/mL and compared to a commercial control Resveratrol. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
Figure 5.
Antioxidant activity of whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed using the ABTS antioxidant assay at a concentration of 1 mg/mL and compared to a commercial control Resveratrol. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
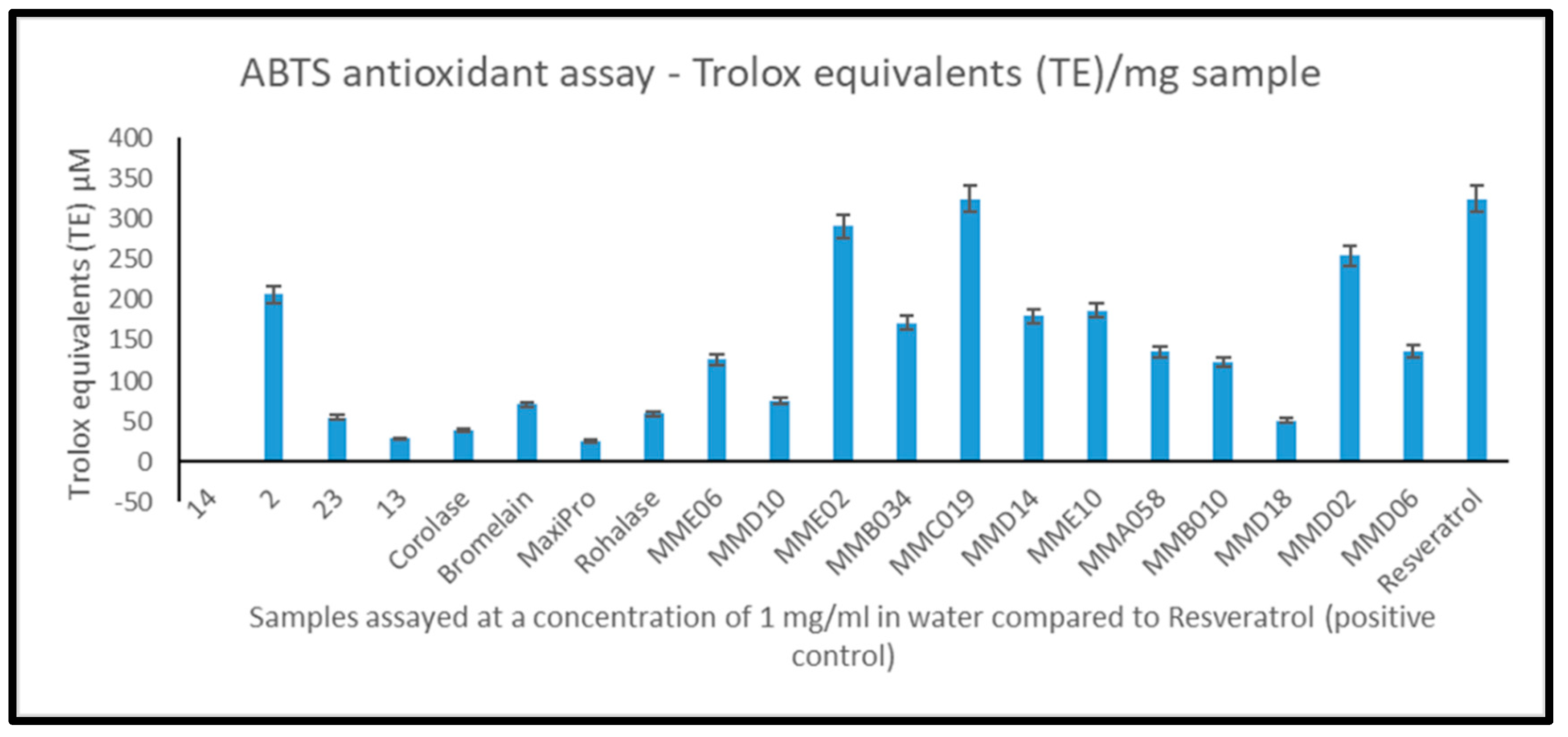
Figure 6.
Renin inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed using the renin inhibition assay method described in the materials and methods section at a concentration of 1 mg/mL and compared to commercial and internal controls SB, SP and Captopril. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
Figure 6.
Renin inhibition by whole, mesopelagic hydrolysates generated with different proteolytic enzymes. Samples were assayed using the renin inhibition assay method described in the materials and methods section at a concentration of 1 mg/mL and compared to commercial and internal controls SB, SP and Captopril. Assays were performed in triplicate (n=9). Irish hydrolysates: 14 corresponds to CE21004 Haul4 (Code 14) Benthosema glaciale biomass hydrolysed with Alcalase®, 2 corresponds to CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri hydrolysed with Alcalase®, 23 corresponds to CE21009 Haul23 Maurolicus muelleri biomass hydrolysed with Alcalase®, 13 corresponds to CE21004 Haul13 (Code 13) Maurolicus muelleri mixed biomass hydrolysed with Alcalase®. Norwegian hydrolysates: Bromelain corresponds to M. muelleri biomass hydrolysed with Bromelain, MaxiPro corresponds to M. muelleri biomass hydrolysed with MaxiPro enzymes, Corolase corresponds to M. muelleri biomass hydrolysed with Corolase and Roholase corresponds to M. muelleri hydrolysed with Roholase enzyme. Spanish hydrolysates: MMD02 and MME02, correspond to M. muelleri hydrolysed with Alcalase 2.4 LG, MMD06 and MME06, correspond to M. muelleri hydrolysed with Papain, MMD10 and MME10 correspond to M. muelleri hydrolysed with Bromelain, MMB010 and MMD18 correspond to M. muelleri hydrolysed with Protamex, MMA058 and MMD14 correspond to M. muelleri hydrolysed with Papain and Bromelain and MMB034 and MMC019 correspond to hydrolysates generated from M. muelleri using endogenous enzymes.
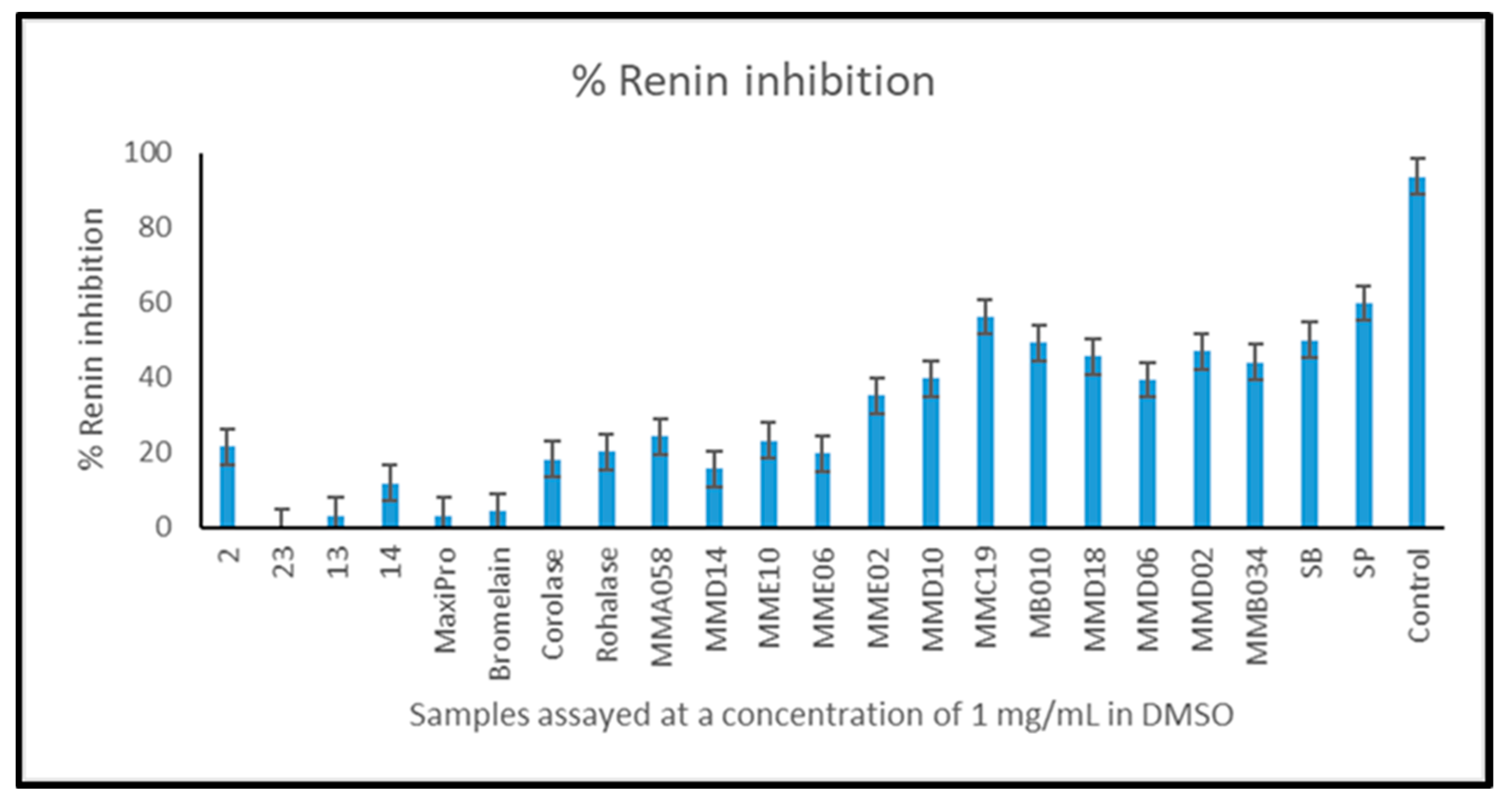

Table 1.
Peptide sequences identified from mesopelagic fish species hydrolysates generated using different enzymes including Alcalase, and in silico analysis predicted bioactivity results for selected peptides identified using mass spectrometry using different programmes including: PeptideRanker http://bioware.ucd.ie/~compass/biowareweb/, accessed on 30 September 2023; PreAIP http://kurata14.bio.kyutech.ac.jp/PreAIP/, accessed on 11th December 2023; BIO-PEP-UWM https://biochemia.uwm.edu.pl/biopep-uwm/, accessed on 6th January 2024; AntiDMPpred available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl, accessed in January 2024 and Umami-MRNN https://umami-mrnn.herokuapp.com/, accessed in January 2024.
Table 1.
Peptide sequences identified from mesopelagic fish species hydrolysates generated using different enzymes including Alcalase, and in silico analysis predicted bioactivity results for selected peptides identified using mass spectrometry using different programmes including: PeptideRanker http://bioware.ucd.ie/~compass/biowareweb/, accessed on 30 September 2023; PreAIP http://kurata14.bio.kyutech.ac.jp/PreAIP/, accessed on 11th December 2023; BIO-PEP-UWM https://biochemia.uwm.edu.pl/biopep-uwm/, accessed on 6th January 2024; AntiDMPpred available at http://i.uestc.edu.cn/AntiDMPpred/cgi-bin/AntiDMPpred.pl, accessed in January 2024 and Umami-MRNN https://umami-mrnn.herokuapp.com/, accessed in January 2024.
| Hydrolysate | Peptide Sequence | Peptide Ranker Value | PreAIP RF combined values | Anti-diabetic prediction (AntiDMPpred) | BIOPEP* | Umami |
|---|---|---|---|---|---|---|
| CE21009 Haul 23 Maurolicus muelleri (Code 23) Alcalase hydrolysate. Irish sample. Freeze-dried. High Confidence AIP (0.482). | KTLRKMGKWCCHCFPCCRGSGKSNVGAW | 0.999 | High confidence AIP (0.731) | Low probability | Novel | umami, predicted threshold: 25.470665mmol/L |
| DGINVLGLIVFCLVLGIVIGRKWEKGQIL | 0.996 | High confidence AIP (0.560) | Low probability | Novel | umami, predicted threshold: 12.649109mmol/L | |
| Origin Thin-lipped mullet | FDAFLPM | 0.955 | Medium confidence AIP (0.392) | Low probability | Novel | Non-umami |
| GLGGMLF | 0.939 | Low confidence AIP (0.370) | Low probability | Novel | umami, predicted threshold: 35.458916mmol/L | |
| Origin Salmo salar – Atlantic salmon | QCPLHRPWAL | 0.932 | High confidence AIP (0.499) | Low probability | Novel | Non-umami |
| LACNCNLHARRCRFNM | 0.908 | High confidence AIP (0.629) | Low probability | Novel | umami, predicted threshold: 37.657642 mmol/L | |
| TFSWGFDDFSCC | 0.889 | High confidence AIP (0.496) | Low probability | Novel | umami, predicted threshold: 14.819101 mmol/L | |
| GINVLGLIVFCLVLGI | 0.888 | High confidence AIP (0.620) | Low probability | Novel | umami, predicted threshold: 35.082146mmol/L | |
| LLSSELQSLLIATTCLRELISCC | 0.873 | High confidence (0.614) | Low probability | Novel | umami, predicted threshold: 13.255066 mmol/L | |
| Origin - Makaira nigricans - Atlantic Blue Marlin | NVGEVVCIFLTAALGLPEALI | 0.868 | High confidence AIP (0.612) | Likely to be anti-diabetic (probability of 0.8) | Novel | umami, predicted threshold: 19.895546 mmol/L |
| Maurolicus muelleri (MMC019) Endogenous enzyme autolysis (Spanish sample), spray dried. High confidence AIP (0.514) | SFVPNGASLEDCHCNLPCLA | 0.874 | High confidence AIP (0.506) | Low probability | Novel | umami, predicted threshold: 30.64746 mmol/L |
| GFSAVNMRKFG | 0.797 | High confidence AIP (0.527) | Low probability | Novel | umami, predicted threshold: 30.85351 mmol/L | |
| Origin - Cypinus carpio - Common carp | IAGFEIFDFNSLEQLC | 0.734 | High confidence AIP (0.540) | Low probability | Novel | umami, predicted threshold: 36.002125 mmol/L |
| NLFKDCNF | 0.693 | Medium confidence AIP (0.467) | Low probability | Novel | umami, predicted threshold: 18.799038 mmol/L | |
| PFGAADQDPF | 0.677 | Low confidence AIP (0.370) | Low probability | Novel | Non-umami | |
| NSGAGILPSPSTPRFP | 0.621 | Medium confidence AIP (0.453) | Low probability | Novel | umami, predicted threshold: 25.949366 mmol/L | |
| DVEFLPPQLPSDKFKDDPVG | 0.601 | Medium confidence AIP (0.433) | Low probability | Novel | umami, predicted threshold: 20.447205 mmol/L | |
| Origin - Takifuga rubipes - Japanese puffer fish | GFAGDDAPR | 0.598 | Negative AIP (0.284) | Low probability | Novel | umami, predicted threshold: 1.6841054 mmol/L |
| FSPFGAAD | 0.58 | Low confidence AIP (0.346) | Low probability | Novel | umami, predicted threshold: 17.39304 mmol/L | |
| PSRILYG | 0.574 | Medium confidence AIP (0.412) | Low probability | Novel | Non-umami | |
| Maurolicus muelleri (MME02 - Spanish haul) (Medium confidence AIP - 0.446) | VFIPFNPL | 0.871 | Low confidence AIP (0.382) | Low probability | Novel | Non-umami |
| NDLPWEF | 0.861 | Low confidence AIP (0.349) | Low probability | Novel | Non-umami | |
| VLLFFYAPWCGQ | 0.846 | High confidence AIP (0.524) | Low probability | Novel | Non-umami | |
| CGRASCPVLCSG | 0.845 | High confidence AIP (0.480) | Low probability | Novel | umami, predicted threshold: 23.602087mmol/L | |
| Origin - Makaira nigricans - Atlantic Blue Marlin | GFNPPDLDIM | 0.828 | Low confidence AIP (0.382) | Low probability | Novel | non-umami |
| Origin - Cypinus carpio - Common carp | SDNAYQFMLT | 0.72 | Medium confidence AIP (0.412) | Low probability | Novel | umami, predicted threshold: 34.065384mmol/L |
| CLGSPNPLDII | 0.687 | Medium confidence AIP (0.408) | Low probability | Novel | umami, predicted threshold: 36.979355mmol/L | |
| RCPEALF | 0.672 | High confidence AIP (0.556) | Low probability | Novel | non-umami | |
| ADDEDADGESSGEPPGAPKQEEAI | 0.667 | High confidence AIP (0.469) | Low probability | Novel | umami, predicted threshold: 8.319466mmol/L | |
| DSFGRLT | 0.662 | Low confidence AIP (0.387) | Low probability | Novel | umami, predicted threshold: 12.873926mmol/L |
Table 2.
Details of the Irish WESPAS survey. Haul dates, positions, depths and haul species compositions are shown.
Table 2.
Details of the Irish WESPAS survey. Haul dates, positions, depths and haul species compositions are shown.
| Haul code and species composition | Date | Latitude start | Longitude start | Bottom depth (m) | Target depth (m) |
|---|---|---|---|---|---|
| CE21004 Haul2 (Code 2) Notocopelus elongtus kroyeri |
23/03/2021 | 52°32.31 | 14°42.21 | 418 | 400 |
| CE21004 Haul4 (Code 14) Benthosema glaciale |
24/03/2021 | 53°29.98 | 14°18.52 | 860 | 500 |
| CE21004 Haul13 (Code 13) Maurolicus muelleri |
02/04/2021 | 59°49.64 | 13°18.39 | 1092 | 220-280 |
| CE21009 Haul23 Maurolicus muelleri (Code 23) |
24/06/2021 | 50°40.48 | 11°18.72 | 1029 | 175 |
Table 3.
Spanish haul information of Maurolicus muelleri samples.
| Haul code | Date | Latitude start | Longitude start | Bottom depth (m) | Target depth (m) |
|---|---|---|---|---|---|
| 9007 | 07/09/2019 | 43°51.24 | 5°58.23 | 400 | 200 |
| 9009 | 08/09/2019 | 43°51.50 | 5°26.42 | 320 | 300 |
| 9014 | 13/09/2019 | 43°47.81 | 6°28.82 | 400 | 206 |
| 9010 | 07/09/2020 | 43°54.09 | 5°47.62 | 500 | 192 |
| 9020 | 10/09/2020 | 43°39.98 | 4°45.74 | 189 | 138 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



