
Submitted:
03 June 2024
Posted:
04 June 2024
You are already at the latest version
Abstract
Helicobacter pylori infection affects over 50% of the world population and leads to chronic inflammation and gastric disorders, being the main pathogen correlated to gastric cancer development. The increasing on antibiotic resistance levels is a major global concern and alternative treatments are needed. Soybean peptides and other compounds might be an alternative in the treatment to avoid, eradicate and/or control symptoms of H. pylori infection. This study aimed to characterize a lunasin-enriched soybean extract (LSE) by proteomics tools and to evaluate its antioxidant, anti-inflammatory and antibacterial properties against H. pylori infection. By LC-MS/MS analysis, 124 proteins were identified, with the 2S albumin (lunasin and large chain subunits) being the fourth most abundant protein (8.9%). Lunasin consisted of 44 amino acid residues and an intramolecular disulfide bond. LSE at a low dose (0.0625 mg/mL) reduced ROS production in both H. pylori-infected and non-infected AGS gastric cells. This led to a significant reduction of 6.71% in the levels of pro-inflammatory interleukin (IL)-8. LSE also showed antibacterial activity against H. pylori which can be attributed to other soybean proteins and phenolic compounds. Our findings suggest that LSE might be a promising alternative in the management of H. pylori infection and associated symptoms.
Keywords:
Helicobacter pylori
; proteomics
; soybean
; lunasin
; oxidative stress
; inflammation
1. Introduction
Helicobacter pylori (H. pylori) is a gram-negative microaerophilic flagellated pathogen that infects the gastric epithelial lining, generating a host’s immune response. It is estimated that more than 50% of the world population is infected by H. pylori, although the global prevalence varies according to regions and demographic groups worldwide [1,2]. Colonization with this pathogen generally occurs in the early stages of human life and will not disappear unless diagnosed and treated [3]. The severity of the infection is conditioned by multiple variables, such as genetic factors, gastric physiology of the host and the characteristics of the pathogen itself [3,4]. The infection can be symptomatic or asymptomatic, and clinical manifestations range from mild dyspeptic symptoms to severe peptic ulcers and gastric cancer [5,6]. The findings in molecular and cellular research of H. pylori-induced physiopathological changes are helping to elucidate the mechanisms of this pathogen to manipulate signaling pathways and evade immune responses [7]. Furthermore, the dynamics between H. pylori and the gut microbiota can influence systemic health, causing problems not only at gastrointestinal level [8].
The immune response raised by H. pylori infection induces a chronic inflammatory response that is associated with gastric cancer, being the strongest risk factor for the development of this disease [9]. Classified by the International Agency for Research on Cancer as a class I carcinogen, the pathogen is correlated with 80% of gastric cancer cases [10,11]. For this reason, eradicative treatment has been indicated. The treatment is conducted with traditional antibiotic therapies combined with other medicines to assist and diminish side-effects. However, these adverse side-effects together the rising antibiotic resistance of pathogens highlights the urgency to search for alternative approaches [12,13,14].
Despite the known importance of changes in diet to lead a healthy life and specially in the context of diseases, diet therapy is often neglected. The diet influences directly in gastrointestinal health. Dietary choices can act favoring H. pylori proliferation and inducing clinical manifestations and ultimately gastric cancer [15]. Therefore, changes in diet could be beneficial in the treatment/management of H. pylori infection. Besides the diet itself, food-derived bioactive compounds can be allies against the progression of infectious diseases such as H. pylori infection. Thus, current studies are focused on demonstrating the biological properties of food-derived compounds as basis of their use as ingredients of functional foods or nutraceuticals [16].
Soybean is one of the main sources of bioactive compounds, such as phytochemicals, peptides, oligosaccharides, dietary fibers, isoflavones, saponins, phenolic and phytic acids. Those are related to several health benefits like cardiometabolic and bone health, cancer prevention, immunomodulation and antiaging effects. Antibacterial activity against H. pylori has been reported in some soybean extracts, both fermented and unfermented, with different criteria regarding the compounds involved in the antibacterial response [17,18]. In the last years, the study of soybean-derived bioactive peptides has gained importance because of their demonstrated human health benefits. Soybean-derived peptides have been reported to exert antimicrobial potential against gram-positive and gram-negative bacteria, although no data on their effects against H. pylori are still available [18]. One of the most studied soybean-derived peptides is lunasin, which multifunctionality has been associated to its anticarcinogenic, anti-inflammatory, antioxidant, and hypocholesterolemic properties [19]. Although no antimicrobial effects have been reported for lunasin, its anti-inflammatory and antioxidant properties demonstrated in cell and animal models [20] suggest a potential this peptide in the control/management of H. pylori infection-associated symptoms. Thus, the main objectives of the present work were to characterize the proteome of a lunasin-enriched soybean extract (LSE) and to evaluate its antimicrobial, antioxidant, and anti-inflammatory properties as basis of its potential against the H. pylori infection.
2. Materials and Methods
2.1. Materials
The albumin-enriched soybean extract (SE) was supplied by Reliv International Inc. (Chesterfield, MO, USA). 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazol bromide (MTT), dimethyl sulfoxide (DMSO), 2′7′-dichlorofluorescein diacetate (DCFH-DA), tert-butyl hydroperoxide (t-BOOH) were purchased from Sigma-Aldrich (St. Louis, MO, USA). All other reagents were of analytical grade.
2.2. Preparation of Lunasin-Enriched Soybean Extract (LSE)
500 mg of SE were dissolved in 50 mL of PBS, stirred at 4°C overnight, and centrifuged at 1000 × g for 5 min twice, collecting the supernatant by decantation and further lyophilization (LSE), and kept at −20°C until further assays. The total protein concentration was determined by Bradford assay in a flat-bottom 96-well microplate (Kasvi, São José dos Pinhais, PR, Brazil) using Bio-Rad reagent (Bio-Rad Laboratories, Inc., Hercules, CA, USA) according to the manufacture procedures. Bovine serum albumin (BSA) was used as standard at concentrations ranged from 50 to 1000 µg/mL. LSE samples were prepared in two different concentrations in deionized water. The assays were performed in triplicate. Absorbance measurements at 595 nm were carried out on a Spectramax M5 microplate reader (Molecular Devices, LLC., San Jose, CA, USA).
2.3. Proteomic Analysis of the Lunasin-Enriched Soybean Extract (LSE)
The strategy employed to perform proteomic analysis of LSE is schematized in Figure 1. The sample was prepared dissolving 4 mg of LSE in 250 µL of 0.1 M NH4HCO3 solution, pH 8.3, to a total protein concentration of 12 µg/µL.
2.3.1. In-Solution Digestion
The enzymatic hydrolysis of LSE was based in the methodology described by Rech et al. [21]. A volume of 100 µL of LSE (12 µg/µL) was collected for in-solution digestion. RapiGest SF surfactant (Waters Corporation, Milford, MA, USA) was added to a final concentration of 0.1% (p/v) and incubated for 15 min at 80°C under agitation in a thermo shaker K80-01 (Kasvi). Then, dithiothreitol (DTT) was added to a final concentration of 10 mM and incubated for 30 min at 60°C to reduce cysteine residues. Alkylation of the cysteines was performed adding iodoacetamide (IAA) to a final concentration of 10 mM and reaction was carried out for 30 min at room temperature in the dark. Trypsin digestion was conducted overnight by adding 2 µg of the enzyme. The RapiGest SF was precipitated by reacting the sample with 0.5% trifluoroacetic acid (TFA) at 37°C for 90 min. Afterwards, the sample was centrifugated at 17,530 x g for 30 min at 5°C and the supernatant was collected and dried to a final volume of approximately 100 µL in a vacuum centrifuge (Savant SPD111V SpeedVac Concentrator, Thermo Electron Corp., Milford, MA, USA).
2.3.2. Analysis of the In-Solution Digested Lunasin-Enriched Soybean Extract (LSE) Samples by Liquid Chromatography-Tandem Mass Spectrometry (LC–MS/MS)
LSE sample from the in-solution digestion was submitted to qualitative and quantitative analysis. For absolute quantification, alcohol dehydrogenase from Saccharomyces cerevisiae (Waters Corp.) was used as standard protein and added into the sample to a final concentration of 200 fmol. The tryptic peptides were desalted on-line using a Waters Symmetry C18 180 μm X 20 mm, 5 μm trap column, and a sample injection volume of 1 µL (or 6 µg) was applied in five replicates. Both quantitative and qualitative analyses was performed by LC using an ACQUITY UPLC M-CLASS HSS T3 C18 column, 100 μm x 100 mm, 1.8 μm (Waters Corp.) with a flow of 0.5 μL/min. The mobile phase A (0.1% (v/v) formic acid in water) and mobile phase B (0.1% (v/v) formic acid in acetonitrile) were combined in a step gradient as follows: 3 to 5% solvent B (0 to 3 min), and increased to 40% solvent B (3 to 95 min), 40 to 85% solvent B (95 to 96 min), held at 85% solvent B (96 to 106 min) and decreased to 3% solvent B (106 to 107 min) and, finally held at the gradient initial conditions until the end of the run (130 min).
The quantitative analyses were carried out on a Waters SynaptTM G1 HD/MS High-Definition Mass Spectrometer (Waters Corp.) set at MSE mode, following a label-free protein quantification methodology [22,23], and coupled to the Nano ACQUITY UPLC chromatography system. The ESI voltage was set at 3000 V, the source temperature was 80°C and the cone voltage was 40 V. The MassLynx data system (Version 4.1, Waters Corp.) was utilized to control the instrument and for data acquisition. The experiments were performed by scanning from a mass-to-charge ratio (m/z) of 50–2000 using a scan time of 0.8 s. MSE label-free quantification acquisitions were performed by alternating the collision energy between 6 V in low energy and ramped from 12-35 V in high energy and using argon as the collision gas at a pressure of 40 psi.
For the qualitative analysis, the mass spectral data were acquired on the same mass spectrometer as described above in Data Dependent Analysis (DDA) in positive ion detection mode. The ToF mass analyzer was calibrated with [Glu1]-Fibrinopeptide B fragment ions in the range of 50 to 2000 m/z and the intact double-charged ion at 785.8426 m/z was used as a reference (“lock mass”) for accurate correction of the mass measurement. A sample volume of 1 µL (or 6 µg) was injected in three replicates into the same LC-MS/MS system described above. The ESI voltage was set at 4000 V, the source temperature was 80°C and the cone voltage was 40 V. The MassLynx data system (Version 4.1, Waters Corp.) was also used to control the instrument and for data acquisition. The experiments were performed by scanning from a m/z of 50–2000 using a scan time of 1 s and an interscan time of 0.02 s. The three more intense peptide ion signals were selected to fragmentation and the collision energy was automatically adjusted according to the ion m/z range.
2.3.3. Bioinformatics Analysis
Data were processed using the Progenesis QI for proteomics version 4.2 software platform (Nonlinear Dynamics, Waters Corp.). Protein and peptide tolerance were set as 100 ppm and 50 ppm respectively. A false discovery rate was set to 1%. All quantified proteins with an ANOVA p-value lower than 0.05 were considered as a positive identification for raw abundance analysis and followed by a quantification using abundance of top three non-conflicting peptides of the external standard as a reference to estimate the amount (fmol) of the other injected proteins.
For quantitative analysis, raw data were searched against Glycine max Uniprot non-reviewed protein database, including possible contaminants, such as human keratin proteins and Sus scrofa trypsin. Trypsin enzyme was utilized, and one missed cleavage site was allowed. Cysteine carbamidomethylation was set as fixed modification, peptide N-terminal carbamidomethylation, methionine oxidation, glutamine to pyroglutamic acid at the N-terminal, and asparagine and glutamine deamidation were set as variable modification.
For the protein identification (qualitative analysis), raw mass spectral data were processed on a Protein Lynx Global Server (PLGS) version 3.0 licensed software (Waters Corp.) and the processed data were searched against the Uniprot Glycine max protein database using the MASCOT search engine (license version 2.8). SemiTrypsin enzyme was utilized, and one missed cleavage site was allowed. The precursor as well as fragment ion tolerances were 1.0 Da and 0.5 Da, respectively. False-positive discovery rate (FDR) was set at 1%, significance threshold value p < 0.05 and peptide ions score ≥ 25. Cysteine carbamidomethylation was set as fixed modification, and carbamidomethylation at N-terminus, deamidation of asparagine and glutamine, conversion of N-terminal glutamine or glutamate to pyroglutamic acid, and methionine oxidation were set as variable modifications. Biological process and molecular function information were retrieved from UniProtKB and Gene Ontology databases.
2.4. Analysis of the Intact Proteins (2S Albumin and Its Subunits) in the Lunasin-Enriched Soybean Extract (LSE)
2.4.1. Separation of Lunasin-Enriched Soybean Extract (LSE)-Fractions by Ultrafiltration
To better characterize whether lunasin was dissociated or associated to the large chain of the 2S albumin in the LSE, an ultrafiltration was performed with reduced and non-reduced LSE samples. Firstly, 0.1 g of LSE was added in two of each microcentrifuge tubes and dissolved in 1 mL of 0.1 M NH4HCO3 solution, pH 8.0. To reduce LSE sample, a final concentration of 10 mM DTT was added. LSE samples were submitted to sonication for 6 min at 30°C to better solubilization. Then, the LSE sample with 10 mM DTT was incubated for 35 min at 37°C to complete reduction. Both samples - with and without DTT - were then centrifuged (Centrifuge 5804R, Eppendorf AG, Hamburg, Germany) at 13,000 x g for 15 min at 4°C. This procedure was repeated three times until the supernatant was reasonably clear. The supernatants were transferred to Amicon® Ultra 0.5 mL centrifugal filter units (Merck Millipore Ltd., Ireland) with a 10 kDa cutoff, followed by centrifugation at 13,000 x g for 10 min at 4°C. This step was performed three times. The retentate and filtrated fractions of both reduced and non-reduced samples were collected, partially concentrated in a vacuum centrifuge (Concentrator plus, Eppendorf AG) and kept at -20°C.
2.4.2. Analysis of the Intact Proteins by LC–MS
The retentate and filtrated fractions of both reduced and non-reduced samples were diluted to approximately 2 µg/µL with 3% (v/v) acetonitrile and 0.1% (v/v) formic acid. The samples were centrifugated at 13,500 x g for 10 min at room temperature and then, the supernatant was transferred to injection glass vials. For the analyses of the filtrated and the retentate fractions, C18 and C4 reversed-phase capillary columns were used, respectively. The filtrated samples (1 µL) were injected into an ACQUITY UPLC M-Class HSS T3 Column, 100 Å, 1.8 µm, 75 µm X 150 mm (Waters Corp.), while the retentate fractions (1 µL) were injected into a nanoACQUITY BEH300 C4 Column, 300 Å, 1.7 µm, 75 µm x 100 mm (Waters Corp.). In both the analysis, the flow rate was at 0.4 μL/min. The mobile phases A and B were the same as described in the section 2.3.2. and the linear gradient was as follows: 3 to 30% solvent B (0 to 0.1 min), held at 30% solvent B (0.1 to 6.25 min) and increased to 60% solvent B (6.25 to 6.26 min), held at 60% solvent B (6.26 to 12.5 min) and increased to 85% solvent B (12.5 to 15.0 min), held at 85% solvent B (15.0 to 18.75 min) and decreased to 3% solvent B (18.75 to 22.5 min) and, finally held at the gradient initial conditions until the end of the run (30 min). Data were analyzed using MassLynx 4.0 (Waters Corp.) package. Each scan of the chromatogram (elution time) was manually checked to search for molecular mass corresponding to the intact 2S albumin and its subunits (lunasin and large chain) in both retentate and filtrated fractions.
2.5. Antioxidant and Anti-Inflammatory Activity of the Lunasin-Enriched Soybean Extract (LSE)
2.5.1. Cell Culture
Human cells derived from a stomach adenocarcinoma (AGS) (American Type Culture Collection, ATCC, Rockville, MD, USA), were grown in modified Dulbecco Eagle/F12 medium (DMEM/F12) (Lonza, Basel, Switzerland) and supplemented with 10% (v/v) fetal bovine serum (FBS) (Hyclone, GE Healthcare, Logan, UK) and 1% penicillin/streptomycin/amphotericin (v/v) (Lonza). Cells were grown at 37°C under constant conditions humidity, 5% CO2, and 95% air.
2.5.2. Effects on Cell Viability
The evaluation of the effect of LSE on cell viability was carried out using the MTT assay. Cells were seeded onto 96-well plates (Sarstedt AG & Co., Nümbrecht, Germany) at a density of 1×105 cells/well and incubated at 37°C for 24 h. After removing the culture medium, the samples of LSE (0.125 to 5 mg of protein/mL) were dissolved in DMEM/F12 without FBS and added to the wells, the plate was incubated for 2 and for 24 h at 37°C. DMEM without FBS was used as a negative control. After discarding the treatment, MTT solution (0.5 mg/mL) was added, and cells were incubated for 2 h at 37°C. The formazan crystals were dissolved in DMSO, and the absorbance was measured at 570 nm in the reader Synergy HT (BioTek Instruments Inc., Winooski, VT, USA). The results were expressed as a percentage of the control, which was considered as 100%.
2.5.3. Cell Infection by H. pylori
AGS cells were seeded in plates according to the assay to be conducted. After the pre-treatment with the samples of LSE, cells were infected with a bacterial inoculum (500 μL/well, 1 × 108 colony forming units CFU/mL) of the H. pylori strain Hp59. This strain was isolated from a gastric biopsy of a symptomatic patient at the Microbiology Department of the University Hospital La Princesa (Madrid, Spain). It is resistant to metronidazole, one of the most widely used antimicrobials in H. pylori therapy, and has pathogenicity attributes associated with high virulence and the most severe clinical conditions [24]. Strain was dissolved in DMEM/F12 without FBS or antibiotics. The infected cells were incubated at 37°C in microaerophilic atmosphere (VAIN). For ROS production, the incubation time was of 3 h. For cytokines production, the incubation time was 24 h.
2.5.4. Effects on Reactive Oxygen Species (ROS) Generation
The effects LSE on the intracellular ROS levels were determined using DCFH-DA as a fluorescent probe. Cells were seeded (1 × 105 cells/well) onto a 24-well plate (Sarstedt, Sevelen, Switzerland) and incubated at 37°C for 24 h. After discarding the medium, cells were pre-treated and incubated for 2 h at 37°C with 500 μL of LSE dissolved in DMEM/F12 without FBS, filtered through 0.22 µM filter units. The cells were washed with DPBS and 500 µL of DCFH-DA (20 µM, dissolved in DMEM/F12 without FBS) was added to each well and incubated for 30 minutes. The assayed conditions were AGS cells challenged with t-BOOH as a pro-oxidative agent and AGS cells infected with the Hp59 strain. For the t-BOOH challenged cells, 500 µL of t-BOOH (2.5 mM, dissolved in DMEM/F12 without FBS) was added to each well and incubated for 3 h at 37°C, with readings every 90 minutes. For the cells challenged by the infection, 500 µL of bacterial inoculum (previously described) was added to each well and incubated for 3 h at 37°C, with readings every 60 minutes. The fluorescence was measured at the excitation and emission wavelengths of 485 nm and 530 nm, respectively, in a Synergy HT plate reader (BioTek Instruments Inc.). The results were expressed as ROS levels (% compared to the controls, which was considered as 100%).
2.5.5. Effects on Cytokine Production
AGS cells were seeded (1 mL/well, cell density 2 x 105) in 48 well plates and infected with bacterial inoculum, as previously described. Before infection, the cells were treated (2 h) with LSE. After the infection, the supernatants were collected from the wells, centrifuged (10 minutes at 12,000 rpm) and then stored at -20°C. Pre-treated and not infected cells were used as a negative control and AGS cells infected without a pre-treatment were used as a positive control. The cytokines production (interleukin IL-10 and tumor necrosis factor TNF-α) was measured with Enzyme-linked Immunosorbent Assay (ELISA) using kits, according to the manufacturers’ instruction (eBioscienceTM, Thermo Fisher Scientific). The cytokine IL-8 secretion was measured using the Diaclone ELISA kit (Besançon, France), following the manufacturers’ instruction.
2.6. Antimicrobial Activity against H. pylori of Lunasin-Enriched Soybean Extract (LSE)
H. pylori strain Hp59 was suspended in Brucella Broth (Becton, Dickinson, & Co., Madrid, Spain) with 20% glycerol as a cryoprotective agent and stored in cryovials at -80°C. The reactivation of the strain was done by inoculation of 200 µL on Müeller-Hinton agar supplemented with 5% defibrinated sheep's blood (MHB) (Becton, Dickinson, & Co.), and then incubated in microaerophilic atmosphere using a variable atmosphere incubator (VAIN) (85% N2, 10% CO2, 5% O2) (MACS-VA500, Don Whitley Scientific, Bingley, UK) at 37°C for 72 h. For the experiments using liquid medium for bacterial growth, Brucella Broth (BB, Becton, Dickinson, & Co.), supplemented with 10% horse serum (HS) (Biowest, Barcelona, Spain) was used.
The antimicrobial activity of LSE against H. pylori was carried out as follows: 1 mL of LSE samples (final concentration 1 mg/mL) was dissolved in water and filtered through 0.22 μm filter units, mixed with 4 mL of BB supplemented with 10% HS in a conical flask. Bacterial biomass grown in plates was diluted in 2 mL of BB supplemented with 10% HS, under aseptic conditions and this bacterial inoculum was added (100 μL, ~1×108 CFU/mL). The conical flasks were incubated under agitation (150 rpm) in a microaerophilic atmosphere using a VAIN at 37°C for 24 h. Positive growth controls were prepared with 1 mL of water in 4 mL of BB supplemented with 10% HS and 100 μL of bacterial inoculum. A negative control was prepared with 5 mL of BB supplemented with 10% HS. After incubation, serial decimal dilutions were prepared in saline solution (0.9% NaCl) and 20 μL of each was seeded on plates of MHB agar and incubated in a microaerophilic atmosphere (VAIN) at 37°C for 120 h. All samples were analyzed in quadruplicate (n = 4). After 120 h of incubation, the number of CFU was evaluated and the results were expressed as the logarithm of CFU per mL (log CFU/mL).
2.7. Statistical Analysis
The obtained results were analyzed using a one-way ANOVA, followed by a turkey test or a t student test, according to the data set. The statistical significance was considered in a p ≤ 0,05. The analyses were conducted using the statistical analysis program GraphPad Prism 8.0 (GraphPad Software, San Diego, CA, USA).
3. Results and Discussion
3.1. Proteomic Characterization of the Lunasin-Enriched Soybean Extract (LSE)
Qualitative analysis of the proteins in the LSE was performed by LC-MS/MS. The Mascot server search identified a total of 124 proteins, including different isoforms (Table S1). Predominantly, these proteins were seed storage proteins belonging to the 11S, 7S, and 2S families, as detailed in Table 1. Furthermore, the LSE contained several protease inhibitors (e. g., Kunitz and Bowman-Birk trypsin inhibitors), which could protect lunasin from digestion in the gastrointestinal tract, as previously reported [25]. In addition, oleosins and lipoxygenases were also present in the LSE. By using UniProtKB and Gene
Ontology, we found that 112 identified proteins had molecular functions (Figure 2A) and that the predominant ones were nutrient reservoirs (17 proteins), oxidoreductases (16 proteins), metal ion binding proteins (15 proteins) and endopeptidase inhibitors (11 proteins), corresponding to 15%, 14%, 13% and 10% of total proteins, respectively. These proteins were involved in 20 biological processes (Figure 2B), such as response to stress (15 proteins), proteolysis (11 proteins), response to abscisic acid (11 proteins) and lipid metabolic process (7 proteins).
A quantification analysis of the proteins contained in the LSE was performed using the label-free MSE approach (Table S2). The 2S albumin (lunasin and large chain subunits) was the fourth most abundant protein (5939.51 fmol) among those detected in the LSE (Table 2). It corresponds to 8.9% of the total proteins quantified. LSE was constituted mainly by the globulins glycinins and β-conglycins subunits (Table 2), which are considered the major components of soybean seed storage proteins [26]. They corresponded to 31.98% and 20.36% of the total proteins quantified in LSE, respectively. These proteins, which together account for more than half of the total amount of LES proteins, have demonstrated antibacterial activity against gram-positive and gram-negative species [27,28].
3.2. Structural Analyses of the Lunasin in the Lunasin-Enriched Soybean Extract (LSE)
In our qualitative analysis, we identified peptides comprising the sequences of both the small (with a sequence coverage of 88.6%) and large (with a sequence coverage of 76.6%) chains (Figure 3). Notably, a peptide corresponding to the C-terminal of the lunasin, containing an asparagine residue, was also identified. This result indicated that lunasin present in the LSE had a length of 44 amino acid residues, consistent with findings in other studies that extracted lunasin form soybean [29,30]. The lunasin peptide corresponds to the small chain of the mature form of soybean 2S albumin. The precursor 2S albumin sequence is constituted of a signal peptide, a small chain, a propeptide, and a large chain (Figure 3) [31]. This protein is processed by proteolytical cleavage, and its mature form is constituted of the small (lunasin) and large chains connected by disulfide bonds. In the study of Serber et al., the mature form of the 2S albumin (14 kDa) in soybean extract was identified, emphasizing the importance of including a reducing step in the purification of lunasin from soybean extract to recover this peptide separately [30].
To further evaluate whether lunasin was dissociated or associated to the large chain of the 2S albumin in the LSE, we conducted a fractionation step through ultrafiltration (10 kDa cut-off) with reduced and non-reduced LSE samples. Since 2S albumin has a 14 kDa molecular weight, while lunasin and the large chain are both < 10 kDa, we expected to find lunasin in the filtrated fraction only if it was dissociated from the other chain. MS analyses of the intact proteins were performed with the retentate and filtrated fractions of the reduced and non-reduced LSE samples. The filtrated fraction (< 10 kDa) of the reduced and non-reduced samples mass spectra showed the presence of ions corresponding to both lunasin and the large chain of the 2S albumin individually (Figure 4 and Figure 5). These results indicate that lunasin was dissociated from the large chain in the LSE. Both lunasin and the large chain monoisotopic masses indicated that the cysteine residues in these proteins were forming intramolecular disulfide bonds (Figure 4C and Figure 5C). Lunasin intact mass detected in the non-reduced LSE sample (5,137.3 Da) corresponds to the theoretical monoisotopic mass of its form with 44 residues (containing a C-terminal asparagine) and an intramolecular disulfide bond. It was not identified any peak corresponding to the mass of the 2S albumin in any of the retentate samples.
3.3. Antioxidant and Anti-Inflammatory Activity of LSE
After AGS cell treatment with LSE, cell viability decreased at all concentrations tested (0.125 to 5 mg of protein/mL) in a dose-dependent manner (data not shown). Thus, the evaluation of the potential antioxidant and anti-inflammatory activities was conducted with a LSE concentration of 0.0625 mg protein/mL, consistent with a cell viability higher than 80%. Despite the low concentration used, LSE reduced ROS production by 22.7% in comparison to the negative control in basal cells, indicating an antioxidant effect (Figure 6). However, in t-BOOH-challenged AGS cells, LSE did not show any effect. This could be due to the high stability of t-BOOH in aqueous solution, generating a persistent oxidative effect in the cells that could not be reverted by LSE at the assayed conditions [32]. This result differed from some previous studies conducted with different soybean preparations and derived compounds, under different conditions. Thus, a simulated gastrointestinal hydrolyzate from soybean flour reduced ROS production in H2O2-challenged AGS cells, attributing the effects to medium and short peptides [33]. Our previous studies have demonstrated the ability of synthetic lunasin to inhibit ROS production in different cell lines. Thus, this peptide, at concentrations ranged from 0.5 to 10 µM reduced ROS levels from 190% to 122% in t-BOOH-induced HepG2 cells [34]. In Caco-2 cells, the antioxidant effect of this peptide was also demonstrated, reducing ROS levels in a dose-dependent manner, with a reduction of 62.7% at 25 µM [35]. Recently, we demonstrated that a soybean protein isolate reduced ROS generation in lipopolysaccharide (LPS)-challenged macrophages RAW264.7, being medium and low-molecular weight peptides the main responsible for the observed effects [36]. The discrepancies between these studies and our current results could be due to different factors such as the type of cell line used, the concentration of lunasin or antioxidant peptides present in LSE or the presence of other compounds in our extract that could antagonize the effect of lunasin and other antioxidant compounds [19,37].
In contrast to what is observed in chemically induced AGS cells, LSE significantly reduced the generation of ROS in Hp59 infected-cells by 10.4% (Figure 7A). In H. pylori infection, ROS production plays a major role, having a paradoxical effect. The overproduction of this biomarker by cells aimed at eliminating the pathogen can damage cells of the host, and also foster a propitious environment for the bacteria to colonize and to develop multidrug resistance [38,39]. Thus, control ROS levels is of paramount importance. The results obtained indicated that LSE might exert a protective effect against oxidative stress in H. pylori infected cells that could be attributed to protein/peptides or other bioactive compounds contained in the soybean extract. The antioxidant effects of lunasin and other soybean peptides have been previously reported in different cell and animal models, but not in H. pylori-infected cells [40,41]. Furthermore, other compounds present in LSE and not determined in this work could contribute to the reduction of ROS production by the cells. Thus, a reduction of ROS levels in AGS cells infected with H. pylori was observed in a study conducted with anthocyanins from black soybean at concentrations ranged from 12.5 to 50 µM [42].
The reduction in ROS production in AGS cells caused by LSE was consistent with the decrease in IL-8 production, the main pro-inflammatory cytokine associated to H. pylori infection and the development of gastric cancer [43]. It provoked a significant reduction of 6.71% in the levels of IL-8 produced by AGS cells (Figure 7B). The anti-inflammatory activity of the peptide lunasin is associated with a suppression of NF-κB pathway by its reduction on the release of cytokines such as IL-1, IL-6 and IL-8 and also the ability of inhibiting integrins [41], another important mechanism in H. pylori infection. Although there are no studies conducted in H. pylori-infected cells with the protein fraction of soybeans, bioactive compounds from sprouted soybeans digest reduced in 19.5% the release of IL-8 in Caco-2 cells [44]. Other compounds such as the anthocyanins fraction from black soybean have also been reported to decrease IL-8 levels by 45.8% in the same infected cell model used in our study [42]. LSE did not provoke significant changes in the release of the cytokines TNF-α and IL-10 in Hp59 infected cells (data not shown). These results suggest that, together with lunasin, other compounds present in the extract may be involved in the modulation of the inflammatory response provoked by LSE and should be studied in future research with the purpose to enhance the anti-inflammatory capacity of this type of extracts.
3.4. Antimicrobial Activity of LSE against H. pylori
Figure 8 shows the antibacterial effect of LSE at 1 mg protein/mL against H. pylori. The extract caused a small but significant decrease of 0.40 log CFU in H. pylori growth. It has been described that peptides with greater antibacterial activity are usually of small size (10-50 amino acids) and highly cationic, with a tendency to adopt amphipathic structures, which increases their affinity for bacterial membranes [45]. The lunasin peptide consists in a sequence of 43-44 amino acids, with many hydrophilic and charged residues, and a theoretical pl of 4.43 [46,47]. To date, the antibacterial activity of this peptide has not been reported because of its length and structural properties that do not seem consistent with previosly identified antibacterial peptides. However, other proteins or peptides contained in LSE could be responsible for the observed effects against H. pylori. Thus, glycinins and β-conglycinins, predominant proteins in LSE (Table 1) could contribute on this activity since hydrolyzates from these proteins have been demonstrated to exert potent antimicrobial effects against E. coli [48]. The Kunitz-type trypsin and Bowman-Birk serine protease inhibitors, also identified in LSE, have been associated to antibacterial activity [49]. Other soybean-derived peptides such as PGTAVFK and IKAFKEATKKVDKVVVVLWTA have also shown antibacterial activity against other gram-negative bacteria such as Pseudomonas aeruginosa [50]. In addition, the phenolic fraction could also contribute since phenolic compounds have been related with the inhibition of H. pylori growth exerted by fermented soybean sprouts [17].
4. Conclusions
The proteome of LSE revealed the presence of 124 proteins with 2S albumin (lunasin and large chain subunits) being the fourth most abundant protein. Lunasin in LSE consisted of 44 residues (containing a C-terminal asparagine) and an intramolecular disulfide bond. LSE at 0.0625 mg/mL had an antioxidant effect by reducing ROS production in both H. pylori-infected and uninfected AGS gastric cells. Moreover, the production of the pro-inflammatory cytokine IL-8 was significantly reduced after the pre-treatment of infected cells by LSE. The antimicrobial effect of LSE at 1 mg/mL could be attributed to several soybean proteins contained in the extract although the contribution of other compounds such as phenolic compounds could not be discarded. Our research suggests that LSE may potentially be a useful tool in the treatment of H. pylori gastric infection diminishing the oxidative stress and inflammation associated to this disease. Further studies are required to explore the role of other components of the extract in the observed behavior.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Table S1: Data from Qualitative Protein Analysis in LES; Table S2: Data from Quantitative Protein Analysis in LES.
Author Contributions
G.F.-O., S.M.P. and S.M.A.-S.: formal analysis and investigation; G.F.-O and S.M.A.-S.: writing—original draft preparation; D.E.K.: methodology, investigation, writing—review and editing, and formal analysis; T.L.F.S.: methodology, investigation, writing—review and editing, supervision, and funding acquisition; A.J.M.-R.: conceptualization, supervision, and writing—review and editing, and funding acquisition; B.H.-L.: conceptualization, supervision, writing—review and editing, and funding acquisition. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Projects AGL2017-89566-R (HELIFOOD) (funded by MCIN/AEI/10.13039/501100011033/ and by “ERDF A Way of Making Europe”) and by Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ), grant number E-26/210.767/2021. G. Franca-Oliveira acknowledges the Spanish Ministry of Science, Innovation, and Universities for the predoctoral fellowship FPU20/06555.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Hooi: J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global prevalence of Helicobacter pylori infection: systematic review and meta-analysis. Gastroenterology 2017, 153, 420–429. [CrossRef]
- Díaz, P.; Valenzuela Valderrama, M.; Bravo, J.; Quest, A.F.G. Helicobacter pylori and gastric cancer: adaptive cellular mechanisms involved in disease progression. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Ferwana, M.; Abdulmajeed, I.; Alhajiahmed, A.; Madani, W.; Firwana, B.; Hasan, R.; Altayar, O.; Limburg, P.J.; Murad, M.H.; Knawy, B. Accuracy of urea breath test in Helicobacter pylori infection: meta-Analysis. World J. Gastroenterol. 2015, 21, 1305–1314. [Google Scholar] [CrossRef]
- Amieva, M.; Peek, R.M. Pathobiology of Helicobacter pylori-induced gastric cancer. Gastroenterology 2016, 150, 64–78. [Google Scholar] [CrossRef]
- O’Connor, A.; Liou, J.-M.; Gisbert, J.P.; O’Morain, C. Review: treatment of Helicobacter pylori infection 2019. Helicobacter 2019, 24, e12640. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, M.A.; Canales, J.; Corvalán, A.H.; Quest, A.F.G. Helicobacter pylori-induced inflammation and epigenetic changes during gastric carcinogenesis. World J. Gastroenterol. 2015, 21, 12742–12756. [Google Scholar] [CrossRef]
- Backert, S.; Neddermann, M.; Maubach, G.; Naumann, M. Pathogenesis of Helicobacter pylori infection. Helicobacter 2016, 21 Suppl 1, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Liou, J.-M.; Malfertheiner, P.; Lee, Y.-C.; Sheu, B.-S.; Sugano, K.; Cheng, H.-C.; Yeoh, K.-G.; Hsu, P.-I.; Goh, K.-L.; Mahachai, V.; et al. Screening and eradication of Helicobacter pylori for gastric cancer prevention: The Taipei global consensus. Gut 2020, 69, 2093–2112. [Google Scholar] [CrossRef] [PubMed]
- Polk, D.B.; Peek, R.M. Helicobacter pylori: Gastric cancer and beyond. Nat. Rev. Cancer 2010, 10, 403–414. [Google Scholar] [CrossRef]
- Schistosomes, liver flukes and Helicobacter pylori. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 61, 1–241.
- Iwu, C.D.; Iwu-Jaja, C.J. Gastric cancer epidemiology: current trend and future direction. Hygiene 2023, 3, 256–268. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Camargo, M.C.; El-Omar, E.; Liou, J.-M.; Peek, R.; Schulz, C.; Smith, S.I.; Suerbaum, S. Helicobacter pylori infection. Nat. Rev. Dis. Primers 2023, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Guo, Q.; Yuan, Y.; Gong, Y. The antibiotic resistance of Helicobacter pylori to five antibiotics and influencing factors in an area of China with a high risk of gastric cancer. BMC Microbiol. 2019, 19, 152. [Google Scholar] [CrossRef] [PubMed]
- Ho, J.J.C.; Navarro, M.; Sawyer, K.; Elfanagely, Y.; Moss, S.F. Helicobacter pylori antibiotic resistance in the United States between 2011 and 2021: A systematic review and meta-analysis. Am. J. Gastroenterol. 2022, 117, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Cover, T.L.; Peek, R.M. Diet, microbial virulence, and Helicobacter pylori-induced gastric cancer. Gut Microbes 2013, 4, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Samtiya, M.; Aluko, R.E.; Dhewa, T.; Moreno-Rojas, J.M. Potential health benefits of plant food-derived bioactive components: an overview. Foods 2021, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Apostolidis, E.; Kwon, Y.; Shinde, R.; Ghaedian, R.; Shetty, K. Inhibition of Helicobacter pylori by fermented milk and soymilk using select lactic acid bacteria and link to enrichment of lactic acid and phenolic content. Food Biotechnol. 2011, 25, 58–76. [Google Scholar] [CrossRef]
- Pina-Pérez, M.C.; Ferrús Pérez, M.A. Antimicrobial potential of legume extracts against foodborne pathogens: A review. Trends Food Sci. Tech. 2018, 72, 114–124. [Google Scholar] [CrossRef]
- Kim, I.-S.; Yang, W.-S.; Kim, C.-H. Beneficial effects of soybean-derived bioactive peptides. Int. J. Mol. Sci. 2021, 22, 8570. [Google Scholar] [CrossRef]
- Fernández-Tomé, S.; Indiano-Romacho, P.; Mora-Gutiérrez, I.; Pérez-Rodríguez, L.; Ortega Moreno, L.; Marin, A.C.; Baldán-Martín, M.; Moreno-Monteagudo, J.A.; Santander, C.; Chaparro, M.; et al. Lunasin peptide is a modulator of the immune response in the human gastrointestinal tract. Mol. Nutr. Food Res. 2021, 65, e2001034. [Google Scholar] [CrossRef]
- Murad, A.; Souza, G.; Garcia, J.; Rech, E. Characterisation and quantitation expression analysis of recombinant proteins in plant complex mixtures using nanoUPLC mass spectrometry. Protocol Exchange 2011, 1. [Google Scholar] [CrossRef]
- Silva, J.C.; Denny, R.; Dorschel, C.; Gorenstein, M.V.; Li, G.-Z.; Richardson, K.; Wall, D.; Geromanos, S.J. Simultaneous qualitative and quantitative analysis of the Escherichia coli proteome: a sweet tale. Mol. Cell Proteomics 2006, 5, 589–607. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.C.; Gorenstein, M.V.; Li, G.-Z.; Vissers, J.P.C.; Geromanos, S.J. Absolute quantification of proteins by LCMSE: A virtue of parallel MS acquisition. Mol. Cell Proteomics 2006, 5, 144–156. [Google Scholar] [CrossRef]
- Silvan, J.M.; Gutierrez-Docio, A.; Guerrero-Hurtado, E.; Domingo-Serrano, L.; Blanco-Suarez, A.; Prodanov, M.; Alarcon-Cavero, T.; Martinez-Rodriguez, A.J. Pre-treatment with grape seed extract reduces inflammatory response and oxidative stress induced by Helicobacter pylori infection in human gastric epithelial cells. Antioxidants 2021, 10, 943. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Huerta, E.; Fernández-Tomé, S.; Arques, M.C.; Amigo, L.; Recio, I.; Clemente, A.; Hernández-Ledesma, B. The protective role of the Bowman-Birk protease inhibitor in soybean lunasin digestion: the effect of released peptides on colon cancer growth. Food Funct. 2015, 6, 2626–2635. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, H.B. Biochemistry and molecular biology of soybean seed storage proteins. J. New Seeds 2000, 2, 1–25. [Google Scholar] [CrossRef]
- Sitohy, M.Z.; Mahgoub, S.A.; Osman, A.O. In vitro and in situ antimicrobial action and mechanism of glycinin and its basic subunit. Int. J. Food Microbiol. 2012, 154, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.-P.; Li, Y.-Q.; Sun, G.-J.; Mo, H.-Z. Antibacterial actions of glycinin basic peptide against Escherichia coli. J. Agric. Food Chem. 2017, 65, 5173–5180. [Google Scholar] [CrossRef] [PubMed]
- Seber, L.E.; Barnett, B.W.; McConnell, E.J.; Hume, S.D.; Cai, J.; Boles, K.; Davis, K.R. Scalable purification and characterization of the anticancer lunasin peptide from soybean. Plos One 2012, 7, e35409. [Google Scholar] [CrossRef]
- Serra, A.; Gallart-Palau, X.; See-Toh, R.S.-E.; Hemu, X.; Tam, J.P.; Sze, S.K. Commercial processed soy-based food product contains glycated and glycoxidated lunasin proteoforms. Sci. Rep. 2016, 6, 26106. [Google Scholar] [CrossRef]
- Galvez, A.F.; de Lumen, B.O. A soybean cDNA encoding a chromatin-binding peptide inhibits mitosis of mammalian cells. Nat. Biotechnol. 1999, 17, 495–500. [Google Scholar] [CrossRef] [PubMed]
- Alía, M.; Ramos, S.; Mateos, R.; Bravo, L.; Goya, L. Response of the antioxidant defense system to tert-butyl hydroperoxide and hydrogen peroxide in a human hepatoma cell line (HepG2). J. Biochem. Mol. Toxicol. 2005, 19, 119–128. [Google Scholar] [CrossRef]
- Cavaliere, C.; Montone, A.M.I.; Aita, S.E.; Capparelli, R.; Cerrato, A.; Cuomo, P.; Laganà, A.; Montone, C.M.; Piovesana, S.; Capriotti, A.L. Production and characterization of medium-sized and short antioxidant peptides from soy flour-simulated gastrointestinal hydrolysate. Antioxidants 2021, 10, 734. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Tomé, S.; Ramos, S.; Cordero-Herrera, I.; Recio, I.; Goya, L.; Hernández-Ledesma, B. In vitro chemo-protective Effect of bioactive peptide lunasin against oxidative stress in human HepG2 cells. Food Res. Int. 2014, 62, 793–800. [Google Scholar] [CrossRef]
- García-Nebot, M.J.; Recio, I.; Hernández-Ledesma, B. Antioxidant activity and protective effects of peptide lunasin against oxidative stress in intestinal Caco-2 cells. Food Chem. Toxicol. 2014, 65, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Franca-Oliveira, G.; Martinez-Rodriguez, A.J.; Morato, E.; Hernández-Ledesma, B. Contribution of proteins and peptides to the impact of a soy protein isolate on oxidative stress and inflammation-associated biomarkers in an innate immune cell model. Plants 2023, 12, 2011. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, C.; Gleddie, S.; Xiao, C.-W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [PubMed]
- Butcher, L.D.; den Hartog, G.; Ernst, P.B.; Crowe, S.E. Oxidative stress resulting from Helicobacter pylori infection contributes to gastric carcinogenesis. Cell Mol. Gastroenterol. Hepatol. 2017, 3, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Cai, Y.; Chen, Z.; Li, H.; Xu, Z.; Li, W.; Jia, J.; Sun, Y. SpoT-mediated NapA upregulation promotes oxidative stress-induced Helicobacter pylori biofilm formation and confers multidrug resistance. Antimicrob. Agents Chemother. 2021, 65, e00152–21. [Google Scholar] [CrossRef]
- Franca-Oliveira, G.; Hernández-Ledesma, B.; Martinez-Rodriguez, A. Bioactive peptides as alternative treatment for Helicobacter pylori infection. Bioact. Compd. Health Dis. 2024, 7, 245–264. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, G.; Diao, J.; Wang, C. Recent advances in exploring and exploiting soybean functional peptides—a review. Front Nutr 2023, 10. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-M.; Kim, K.-M.; Park, E.-H.; Seo, J.-H.; Song, J.-Y.; Shin, S.-C.; Kang, H.-L.; Lee, W.-K.; Cho, M.-J.; Rhee, K.-H.; et al. Anthocyanins from black soybean inhibit Helicobacter pylori-induced inflammation in human gastric epithelial AGS Cells. Microbiol. Immunol. 2013, 57, 366–373. [Google Scholar] [CrossRef]
- Lee, K.E.; Khoi, P.N.; Xia, Y.; Park, J.S.; Joo, Y.E.; Kim, K.K.; Choi, S.Y.; Jung, Y.D. Helicobacter pylori and interleukin-8 in gastric cancer. World J. Gastroenterol. 2013, 19, 8192–8202. [Google Scholar] [CrossRef] [PubMed]
- Nolasco, E.; Krassovskaya, I.; Hong, K.; Hansen, K.; Alvarez, S.; Obata, T.; Majumder, K. Sprouting alters metabolite and peptide contents in the gastrointestinal digest of soybean and enhances in-vitro anti-inflammatory activity. J. Funct. Foods 2023, 109, 105780. [Google Scholar] [CrossRef]
- Parreira, P.; Duarte, M.F.; Reis, C.A.; Martins, M.C.L. Helicobacter pylori infection: a brief overview on alternative natural treatments to conventional therapy. Crit. Rev. Microbiol. 2016, 42, 94–105. [Google Scholar] [CrossRef] [PubMed]
- Odani, S.; Koide, T.; Ono, T. Amino acid sequence of a soybean (Glycine max) seed polypeptide having a poly(L-Aspartic acid) structure. J. Biol. Chem. 1987, 262, 10502–10505. [Google Scholar] [CrossRef]
- de Souza, S.M.A.; Hernández-Ledesma, B.; de Souza, T.L.F. Lunasin as a promising plant-derived peptide for cancer therapy. Int. J. Mol. Sci. 2022, 23, 9548. [Google Scholar] [CrossRef]
- Vasconcellos, F.; Woiciechowski, A.; Thomaz-Soccol, V.; Mantovani, D.; Soccol, C. Antimicrobial and antioxidant properties of β-conglycinin and glycinin from soy protein isolate. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 144–157. [Google Scholar]
- Dhayakaran, R.; Neethirajan, S.; Weng, X. Investigation of the antimicrobial activity of soy peptides by developing a high throughput drug screening assay. Biochem. Biophys. Rep. 2016, 6, 149–157. [Google Scholar] [CrossRef]
- Nascimento, A.M. de S.; Segundo, V.H. de O.; Aguiar, A.J. F. C.; Piuvezam, G.; Passos, T. S.; Damasceno, K.S.F. da S. da S. C.; Morais, A.H.A. Antibacterial action mechanisms and mode of trypsin inhibitors: a systematic review. J. Enzyme Inhib. Med. Chem. 2022, 37, 749–759. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the methodology employed for lunasin-enriched soybean extract (LSE) characterization using proteomic analysis.
Figure 1.
Schematic representation of the methodology employed for lunasin-enriched soybean extract (LSE) characterization using proteomic analysis.
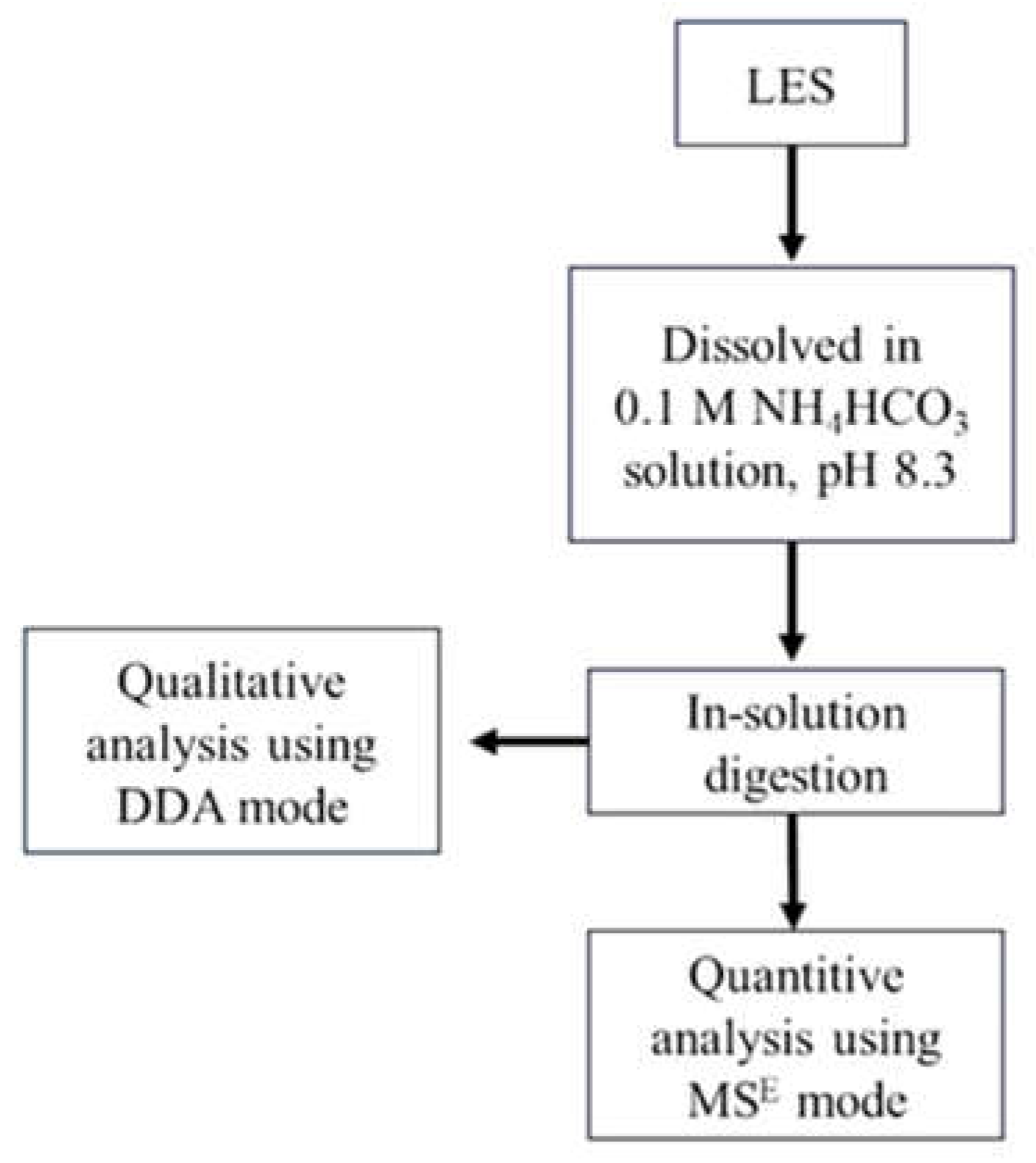
Figure 2.
(A) Molecular functions and (B) biological processes of the proteins identified in the lunasin-enriched soybean extract (LSE). Information was obtained from the UniProtKB and Gene Ontology. All the molecular functions and biological processes identified are presented.
Figure 2.
(A) Molecular functions and (B) biological processes of the proteins identified in the lunasin-enriched soybean extract (LSE). Information was obtained from the UniProtKB and Gene Ontology. All the molecular functions and biological processes identified are presented.
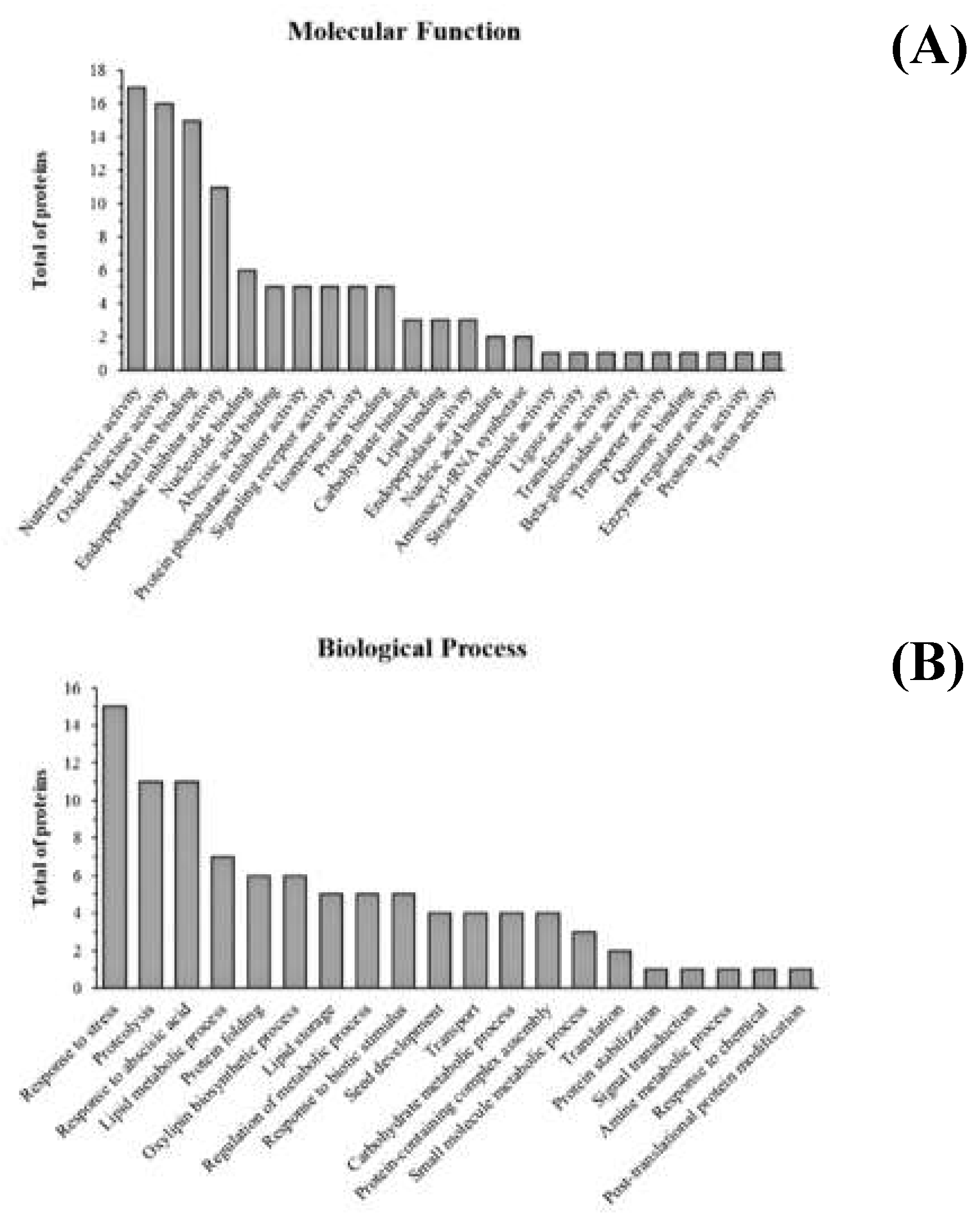
Figure 3.
2S albumin sequence coverage by LC-MS/MS analysis. Each part of the sequence is identified and highlighted in corresponding colours. The sequence parts identified by MS analysis are underlined.
Figure 3.
2S albumin sequence coverage by LC-MS/MS analysis. Each part of the sequence is identified and highlighted in corresponding colours. The sequence parts identified by MS analysis are underlined.

Figure 4.
Analysis of the lunasin in the filtrated non-reduced and reduced lunasin-enriched soybean extract (LSE) samples by LC-MS. (A) Base peak ion chromatograms of the filtrated non-reduced and reduced LSE samples. Arrows indicate the time of elution of lunasin. (B) Mass spectra of lunasin in both non-reduced and reduced LSE samples from chromatograms at the times 8.26 and 8.11 min, respectively. (C) Comparison of the mass spectra of lunasin in both non-reduced and reduced LSE samples for 5+ charge species.
Figure 4.
Analysis of the lunasin in the filtrated non-reduced and reduced lunasin-enriched soybean extract (LSE) samples by LC-MS. (A) Base peak ion chromatograms of the filtrated non-reduced and reduced LSE samples. Arrows indicate the time of elution of lunasin. (B) Mass spectra of lunasin in both non-reduced and reduced LSE samples from chromatograms at the times 8.26 and 8.11 min, respectively. (C) Comparison of the mass spectra of lunasin in both non-reduced and reduced LSE samples for 5+ charge species.
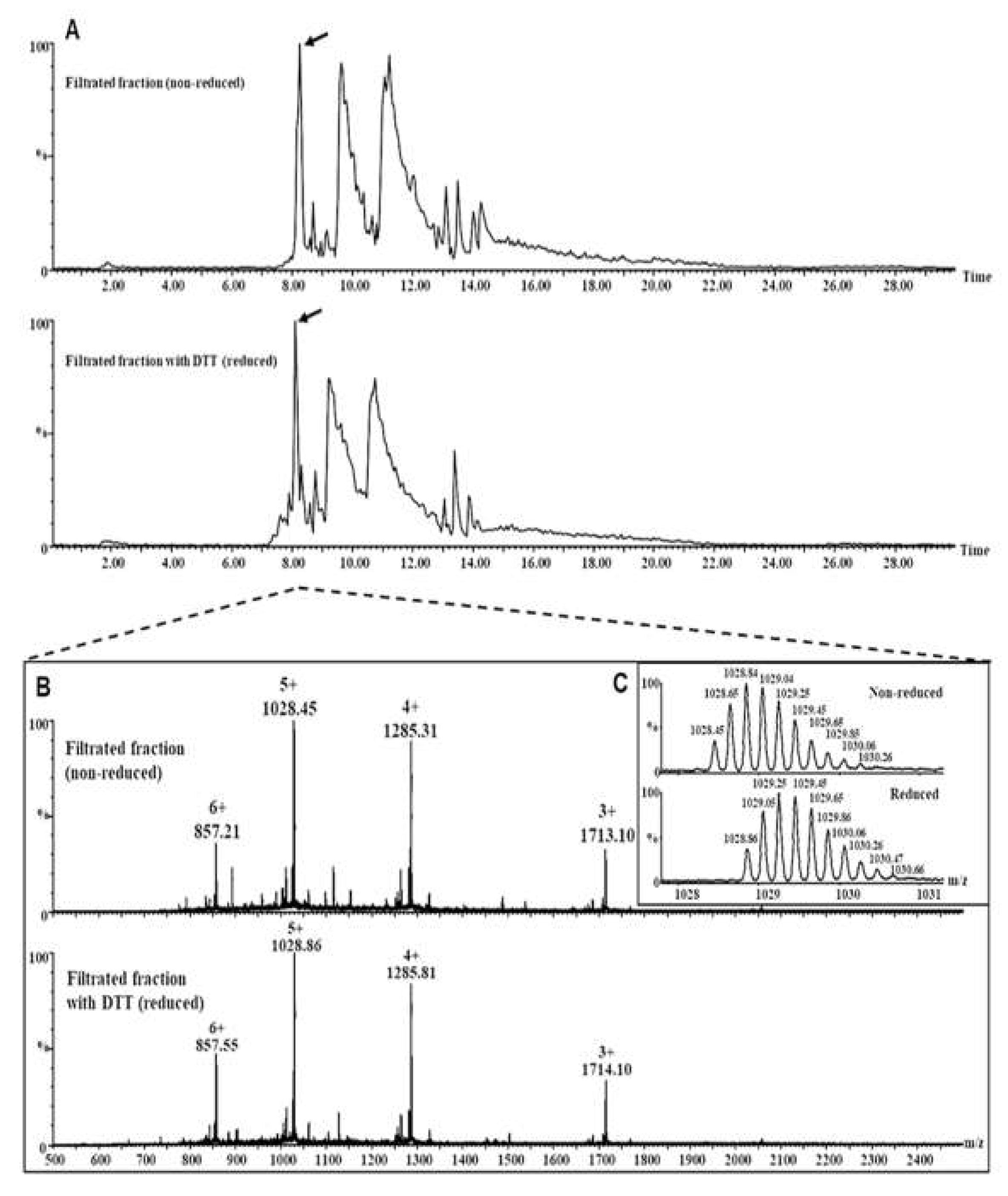
Figure 5.
Analysis of the 2S albumin large chain in the filtrated non-reduced and reduced lunasin-enriched soybean extract (LSE) samples by LC-MS. (A) Base peak ion chromatograms of the filtrated non-reduced and reduced LSE samples. Arrows indicate the time of elution of the 2S albumin large chain. (B) Mass spectra of 2S albumin large chain in both non-reduced and reduced LSE samples obtained from chromatograms at the times 10.05 and 10.99 min, respectively. (C) Comparison of the mass spectra of 2S albumin large chain in both non-reduced and reduced LSE samples for 9+ charge species.
Figure 5.
Analysis of the 2S albumin large chain in the filtrated non-reduced and reduced lunasin-enriched soybean extract (LSE) samples by LC-MS. (A) Base peak ion chromatograms of the filtrated non-reduced and reduced LSE samples. Arrows indicate the time of elution of the 2S albumin large chain. (B) Mass spectra of 2S albumin large chain in both non-reduced and reduced LSE samples obtained from chromatograms at the times 10.05 and 10.99 min, respectively. (C) Comparison of the mass spectra of 2S albumin large chain in both non-reduced and reduced LSE samples for 9+ charge species.
Figure 6.
Antioxidant effects of lunasin-enriched soybean extract (LSE) in AGS cells. Effects of LSE (0.065 mg protein/mL) on reactive oxygen species (ROS) production (expressed as %) in AGS cells under basal conditions and chemically-challenged with tert-butyl hydroperoxide (t-BOOH, 2.5 mM). Negative control indicates 100% ROS production. * p ˂ 0.0001.
Figure 6.
Antioxidant effects of lunasin-enriched soybean extract (LSE) in AGS cells. Effects of LSE (0.065 mg protein/mL) on reactive oxygen species (ROS) production (expressed as %) in AGS cells under basal conditions and chemically-challenged with tert-butyl hydroperoxide (t-BOOH, 2.5 mM). Negative control indicates 100% ROS production. * p ˂ 0.0001.
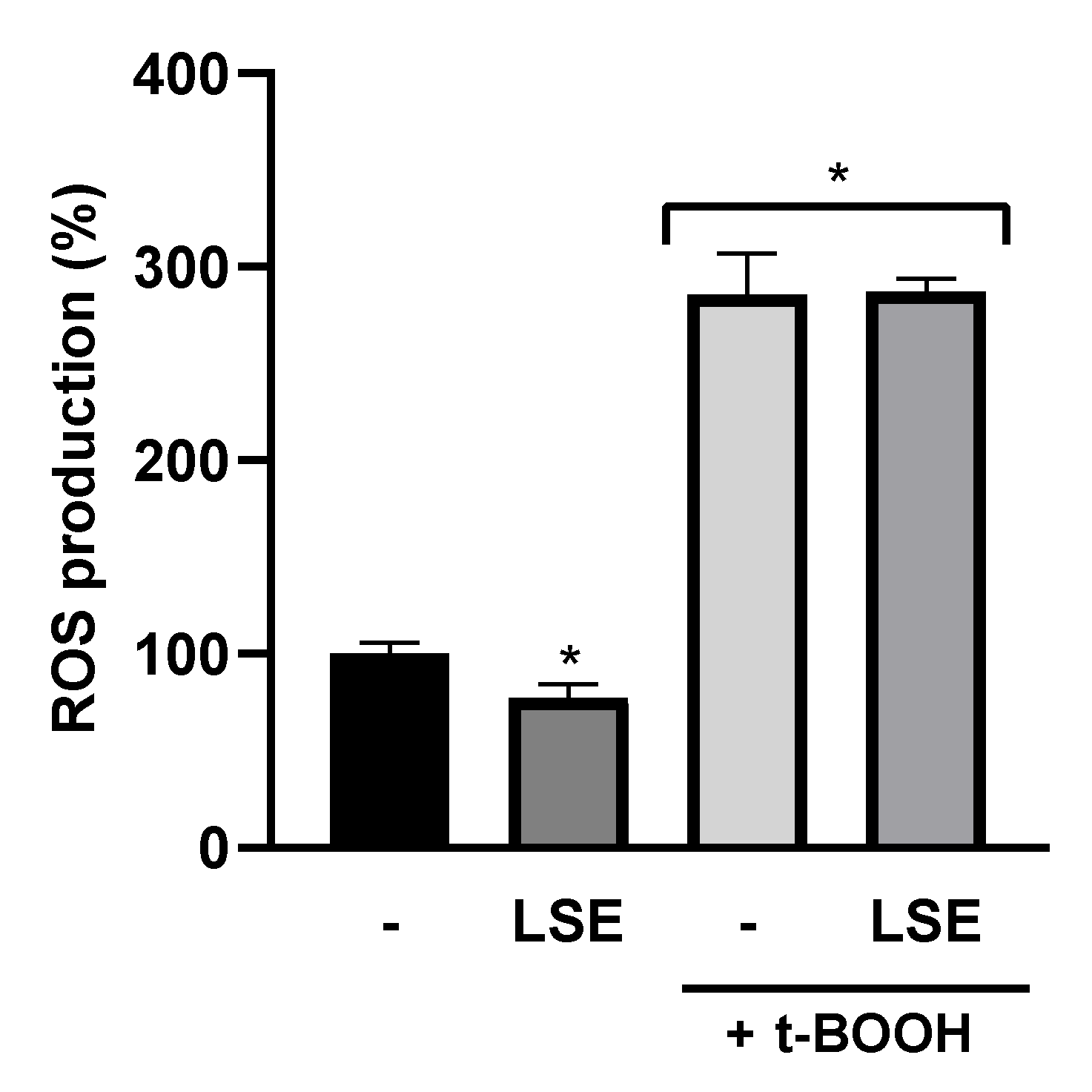
Figure 7.
Effects of lunasin-enriched soybean extract (LSE, 0.065 mg protein/mL) on the (A) reactive oxygen species (ROS) production (expressed as % of ROS in comparison with non-treated infected cells, considered as 100%), and (B) interleukin (IL)-8 (expressed as pg/mL) in Helicobacter pylori infected-AGS cells. * p = 0.0222; ** p = 0.0087.
Figure 7.
Effects of lunasin-enriched soybean extract (LSE, 0.065 mg protein/mL) on the (A) reactive oxygen species (ROS) production (expressed as % of ROS in comparison with non-treated infected cells, considered as 100%), and (B) interleukin (IL)-8 (expressed as pg/mL) in Helicobacter pylori infected-AGS cells. * p = 0.0222; ** p = 0.0087.
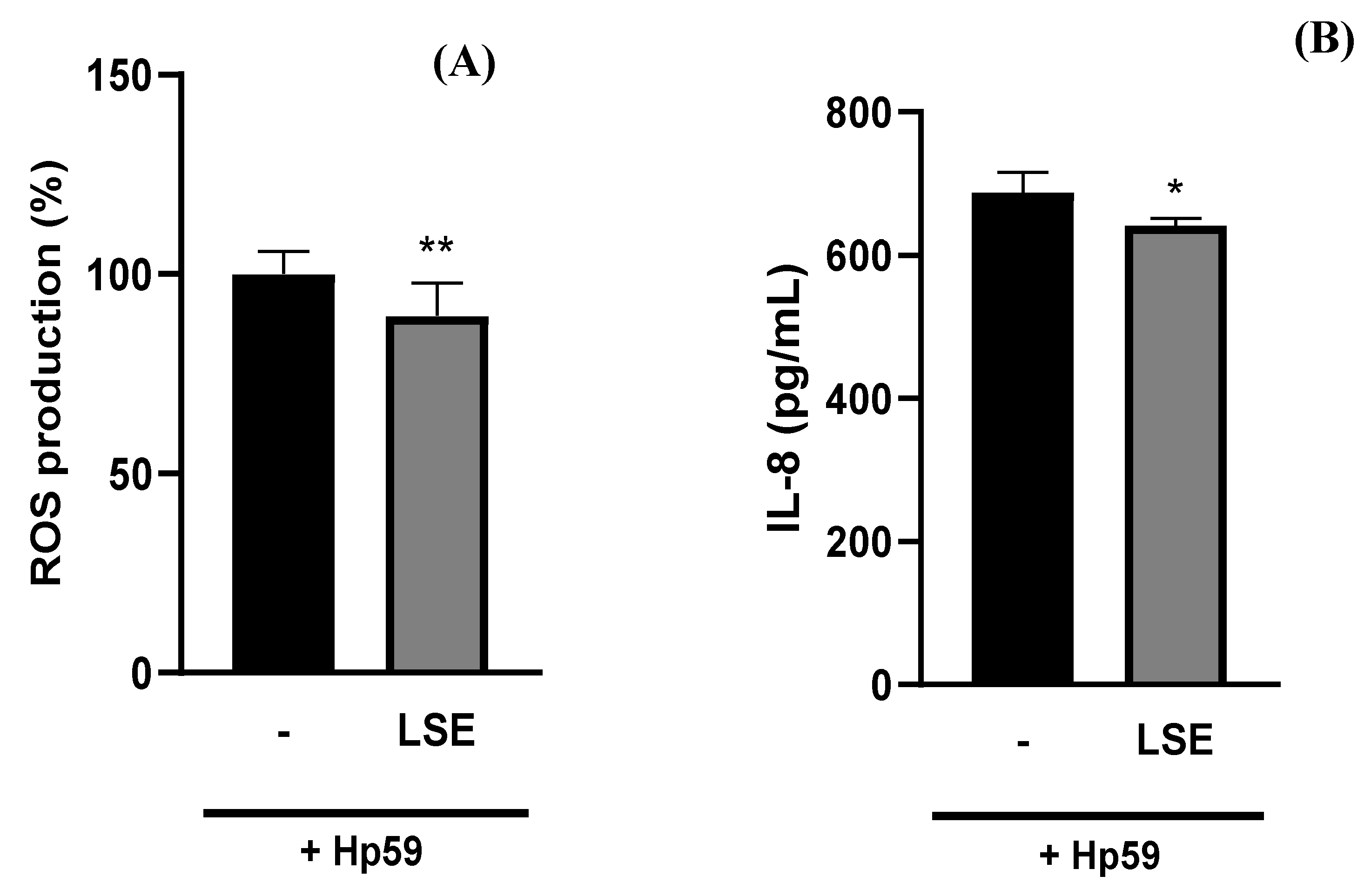
Figure 8.
Antimicrobial effect of the lunasin-enriched soybean extract (LSE, 1 mg protein/mL) against Helicobacter pylori. The results were expressed as log UFC/mL (mean ± SD) (n = 4). * p = 0.0034.
Figure 8.
Antimicrobial effect of the lunasin-enriched soybean extract (LSE, 1 mg protein/mL) against Helicobacter pylori. The results were expressed as log UFC/mL (mean ± SD) (n = 4). * p = 0.0034.

Table 1.
Summary of the proteins identified in the lunasin-enriched soybean extract (LSE) by mass spectrometry (MS) analysis.
Table 1.
Summary of the proteins identified in the lunasin-enriched soybean extract (LSE) by mass spectrometry (MS) analysis.
| Protein Family | Proteins | Accession Number |
|---|---|---|
| 11S seed storage protein (globulins) | Glycinins | P04405, P04776, P02858, P11828, A0A0R0GMV1, A0A0R0KK84, A0A0R0KRW0, P02858-2, P04776-2, P04347 |
| 7S seed storage protein | β-conglycinins | P0DO15, P11827, F7J077, P25974, A0A0R0HYM3, I1NGH2, K7N3H7, A0A0R0I6G3 |
| 2S seed storage albumins | 2S albumin and Napin-type 2S albumin 1 | P19594 and Q9ZNZ4 |
| Protease inhibitor I3 (leguminous Kunitz-type inhibitor) | Kunitz-type trypsin inhibitors | P01070, C6SWW4, P01071, P25273, A0A0R0IWE9, I1KYW9 |
| Bowman-Birk serine protease inhibitor | Bowman-Birk serine protease inhibitors | I1MQD2, P01063, A0A0R0IF61, I1MAC4 |
| Oleosin | Oleosins | P29531, I1N747, K7KTR9, C3VHQ8, C6SZ13 |
| Lipoxygenase | Lipoxygenases | B3TDK6, P09186, I1KH67, I1M597, I1M596, P09439 |
| BetVI | Bet v I | C6T588, C6TFW4, K7L3M0, I1KMV1 |
| Protein disulfide isomerase | Protein disulfide-isomerases | I7FST9, I1JZ42, I1KAB7, A0A0R0JFL1 |
| Glyceraldehyde-3-phosphate dehydrogenase | Glyceraldehyde-3-phosphate dehydrogenases | I1KC68, I1JXG9, A0A0R0KA84 |
| Small hydrophilic plant seed protein | Proteins SLE1, SLE2, and SLE3 | I1JLC8, I1N2Z5, C6T0L2, A0A0R0J847 |
| Seed biotin-containing protein SBP65 | Seed biotin-containing proteins SBP65 | I1M3M9, K7LW58, A0A0R0JWF4 |
| Small heat shock protein (HSP20) | Seed maturation protein PM31 and SHSP domain-containing protein | Q9XET1 and C6T1V2 |
| LEA type SMP | SMP domain-containing proteins | C6T1V2, I1NGG4, I1LE41, I1L849 |
| LEA type 1 | 18 kDa seed maturation proteins | Q01417, I1L2M6 |
| LEA type 2 | Seed maturation protein PM22 | Q9XER5 |
| LEA type 4 | Late embryogenesis abundant protein | Q39871 |
| Peptidase A1 | Basic 7S globulin and Basic 7S globulin 2 |
P13917 and Q8RVH5 |
| Peptidase C1 | P34 probable thiol protease | P22895 |
| Zinc-containing alcohol dehydrogenase | Alcohol dehydrogenase 1 | A0A0R4J4U4 and I1MA91 |
| D-isomer specific 2-hydroxyacid dehydrogenase | Formate dehydrogenase, mitochondrial | I1M005 |
| Plant LTP |
Non-specific lipid-transfer protein and Hydrophobic seed protein domain-containing protein |
C6TFC1 and I1MG41 |
| Leguminous lectin | Lectin | P05046 |
| Plant dehydrin | Dehydrin | O23957 |
| Cystatin | Cysteine proteinase inhibitor | I1M0K3 |
| Calcineurin regulatory subunit | Calcineurin B-like protein | K7KRW8 |
| Universal ribosomal protein uS7 | Ribosomal protein S7 domain-containing protein | I1JFW6 |
| Class-II aminoacyl-tRNA synthetase | Glycine—tRNA ligase | I1KZJ9 |
| Mitochondrial carrier (TC 2.A.29) | Mitochondrial carrier protein | A0A0R0KAF4 |
| Copper/topaquinone oxidase | Amine oxidase | I1MRA7 |
| Glycosyl hydrolase 1 | Sugar phosphate transporter domain-containing protein | A0A0R0JIT8 |
| OBAP | Uncharacterized protein | A0A0R0GZT5 |
| Caleosin | EF-hand domain-containing protein | A0A0R0HVK7 |
Table 2.
Quantification of proteins in the lunasin-enriched soybean extract (LSE) by mass spectrometry (MS) using MSE method.
Table 2.
Quantification of proteins in the lunasin-enriched soybean extract (LSE) by mass spectrometry (MS) using MSE method.
| Accession. | Protein | Peptide Count | Unique Peptides | Confidence Score | Theoretical Mass (Da) | Total Experimental Amount (fmol) | Percentage in Total Protein (%) |
|---|---|---|---|---|---|---|---|
| P04776 | Glycinin G1 | 83 | 48 | 801.50 | 56333.71 | 8290.88 | 12.39 |
| P0DO15; I1NGH2; K7KGR6; K7N005; P0DO16 | Beta-conglycinin alpha subunit 2 | 75 |
54 |
813.70 |
70591.40 |
7155.67 |
10.69 |
| P02858 | Glycinin G4 | 80 | 66 | 782.68 | 64253.61 | 5943.14 | 8.88 |
| P19594 | 2S seed storage albumin protein | 32 | 32 | 235.11 | 19030.31 | 5939.51 | 8.88 |
| P11827 | Beta-conglycinin alpha’ subunit | 95 | 77 | 692.06 | 72513.18 | 3939.93 | 5.89 |
| A0A0R0GMV1 | Glycinin G1 (Fragment) | 48 | 10 | 567.38 | 55600.23 | 3816.23 | 5.70 |
| P01070; A0A0R0IWE9; I1KYW9; Q39898 | Trypsin inhibitor A |
49 |
36 |
293.94 |
24290.46 |
3659.55 |
5.47 |
| K7LEQ5 | Dehydrin | 36 | 29 | 339.19 | 26687.07 | 3482.17 | 5.20 |
| Q9ZNZ4 | Napin-type 2S albumin 1 | 33 | 32 | 255.07 | 18404.98 | 1858.82 | 2.78 |
| P04405 | Glycinin G2 | 67 | 23 | 612.42 | 54961.11 | 1593.32 | 2.38 |
| P25974; A0A0R0I6G3 | Beta-conglycinin beta subunit 1 | 70 | 29 | 505.48 | 50532.97 | 1215.24 | 1.82 |
| I1NGG4 | Late embryogenesis abundant protein D-34-like | 29 | 24 | 252.52 | 26172.16 | 1027.04 | 1.53 |
| P11828 | Glycinin G3 | 48 | 18 | 537.14 | 54869.09 | 1026.31 | 1.53 |
| F7J077 | Beta-conglycinin beta subunit 2 | 38 | 2 | 497.00 | 50498.95 | 1021.06 | 1.53 |
| P22895; O64458 | P34 probable thiol protease | 6 | 5 | 52.93 | 43136.04 | 837.89 | 1.25 |
| P01064; I1MQD2 | Bowman-Birk type proteinase inhibitor D-II | 12 | 12 | 123.67 | 10323.16 | 769.52 | 1.15 |
| P29531 | P24 oleosin isoform B | 13 | 7 | 140.98 | 23392.69 | 736.24 | 1.10 |
| P04347 | Glycinin G5 | 51 | 12 | 460.16 | 58412.39 | 730.70 | 1.09 |
| P05046 | Lectin | 8 | 8 | 62.18 | 30928.02 | 702.78 | 1.05 |
| I1L957 | Late embryogenesis abundant protein LEA | 42 | 28 | 375.05 | 48795.62 | 651.76 | 0.97 |
Only the 20 most abundant proteins are presented in the table. Theoretical mass corresponds to the precursor proteins. Total protein amount corresponds to the sum of all detected ions.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



