
Submitted:
03 June 2024
Posted:
05 June 2024
You are already at the latest version
Abstract
Abstract: Background/Objectives: There are indications that the microbial composition of the maternal mucosal surfaces is associated with adverse events during pregnancy. The aim of this review is to investigate the link between vaginal microbiome alterations and gestational com-plications risk. Methods: This comprehensive literature review was performed using Medline and Scopus databases. The following search algorithm was used "Pregnancy Complications"[Mesh] AND (Vagin*) and after literature screening 44 studies were included in the final review. Results: The studies that were included investigated the association between vaginal microbial composi-tion and preterm birth, miscarriage, preeclampsia, ectopic pregnancy, gestational diabetes mellitus, chorioamnionitis and preterm premature rupture of membranes. In most of the studies, it was well established that increased microbial diversity is associated with these conditions. Also, the depletion of Lactobacillus species is linked to most of the gestational complication, while the increased relative abundance and especially L. crispatus may exerts a protective effect in favor of the pregnant woman. Several pathogenic taxa including Gardnerella, Prevotella, Sneathia, Bac-terial Vaginosis-Associated Bacteria-2, Atopobium and Megasphera seem to be correlated to higher maternal morbidity. Conclusions: Vaginal microbiome aberrations seem to have an as-sociation with pregnancy related adverse events, but more high-quality homogenous studies are necessary to reliably verify this link.
Keywords:
Keywords: Vaginal Microbiome
; Pregnancy Complications
; Preterm birth
; Miscarriage
; Gestational Diabetes Mellitus
; Preeclampsia
; Chorioamnionitis
; Ectopic pregnancy
; Preterm Premature Rapture of Membranes
1. Introduction
Maternal morbidity constitutes a public health issue of great importance because not only undermines both the mother’s and infant’s well-being, but also significantly increases the pregnancy-related incremental costs [1]. In fact, severe morbidity rates can complicate up to 8% of pregnancies taking place in hospital, while its rates have increased in the US from 74 per 100,000 delivery hospitalizations in 1998-99 to 163 in 2010-11 [2,3]. Pregnancy related complications include maternal hemorrhage, maternal sepsis and other pregnancy related infections, hypertensive disorders (HD), stillbirth, miscarriage, ectopic pregnancy, gestational diabetes mellitus (GDM) and spontaneous preterm birth (sPTB). Among them maternal hemorrhage is the most fatal accounting for up to 20% of maternal mortality, while some of the most common complications are GDM and HD with a prevalence of 10.9% and 5.2-8.2% respectively [4,5].
Given that every pregnancy can be prone to unpredictable and sudden onset complications, prompt access to appropriate care facilities is of paramount importance [6]. However, the ability to predict a woman’s predisposition to a certain complication based on demographic, genetic and social data would be an essential step in pregnancy related morbidity reduction [7]. Towards this direction points the analysis of either the mother’s or the newborn’s microorganism collections that inhabit the skin and multiple mucosal surfaces, named microbiota, to assess their association with several pregnancy-related complications [8]. The female gut, vagina, oral cavity, respiratory system, and uterus constitute major parts of the mucosal surfaces colonized by microorganisms [9]. For instance, recent advances in reproductive health indicate that preeclampsia is associated with abnormal gut microbiome, while women with gestational hypertension have more periodontal pathogens in their oral cavity compared to normotensive control study participants [10,11].
The human vagina harbors microorganisms that protect the host from several urogenital diseases, such as urinary tract infections, sexually transmitted diseases, and bacterial vaginosis. This disease preventing ability is attributed to the low vaginal pH due to lactic acid production by Lactobacillus sp., the production of bacteriostatic substances and the competitive exclusion [12,13,14]. Factors that can result in aberrations in the vaginal bacterial composition include obesity, hormonal changes, socioeconomic status, race, ethnicity, antibiotic administration, and urogenital infections [15,16].
During a non-complicated pregnancy, the vaginal microbiota remains stable with a predominance of Lactobacillus species [16]. Other bacteria that can be present in the vaginal microbiome include Gardnerella vaginalis, Atopobium vaginae, Prevotella sp., Sneathia amnii and Candidatus Lachnocurva vaginae [17]. To investigate more efficiently the highly diverse landscape of microbial taxa found in the vagina, a taxonomic classification into community state types (CST) based on the dominant bacterial species has been widely adopted. This classification includes CST-I (L. crispatus), CST-II (L. gasseri), CST-III (L. iners), CST-V (L. jensenii) and CST-IV dominated by several anaerobic species [18].
Since oral cavity and gastrointestinal tract microbiome abnormalities have been associated with certain pregnancy complications combined with evidence from single arm studies where in miscarriage cases there is diminished abundance of Lactobacillus sp. and increased isolation ratio of anaerobes ignites many questions about the association between vaginal microbiota composition and pregnancy related complications [10,11,19]. Thus, the aim of this literature review is to share reliable insights on the possible role of vaginal microbiome in gestational complications and further elucidate their association.
2. Materials and Methods
A literature search was performed using the MEDLINE and Scopus databases. The following terms were used in the search text fields (“Microbiota”[Mesh]) AND “Pregnancy Complications”[Mesh] AND (Vagin*). The search algorithm was adjusted for each database while maintaining a common overall architecture. Published observational and interventional studies investigating the association between vaginal microbiome and pregnancy complications published up to 22 April 2024 were included while reviews, letters, and commentaries were excluded.
Retrieved records underwent semi-automatic deduplication by Rayyan [20]. Unique records were imported in Rayyan. Two independent authors screened them for relevance based on titles and abstracts only. Disagreements were resolved through consensus or by discussion with a third author. Articles deemed as irrelevant were excluded and the full text copies of the remaining articles were assessed for eligibility as per the PICOS criteria by two blinded reviewers. Inconsistencies were, once again, resolved by consensus or by a third reviewer. The references of the full text copies were accessed to prevent potential loss of eligible studies that were missed by the database search (snowball procedure). The following data items were extracted from the eligible studies; Year of publication, study design, country, center and time period during which the study was conducted, number of participants, age, BMI, time point of vaginal swab collection, alpha diversity, beta diversity, dominant CST through the study arms, events of preterm birth (defined as live delivery of one or more infants at less than 37 completed weeks of gestation), miscarriages (defined as the loss of pregnancy before 20 weeks of gestation), preeclampsia and gestational diabetes mellitus.
3. Results
3.1. The selection Process of Included Studies
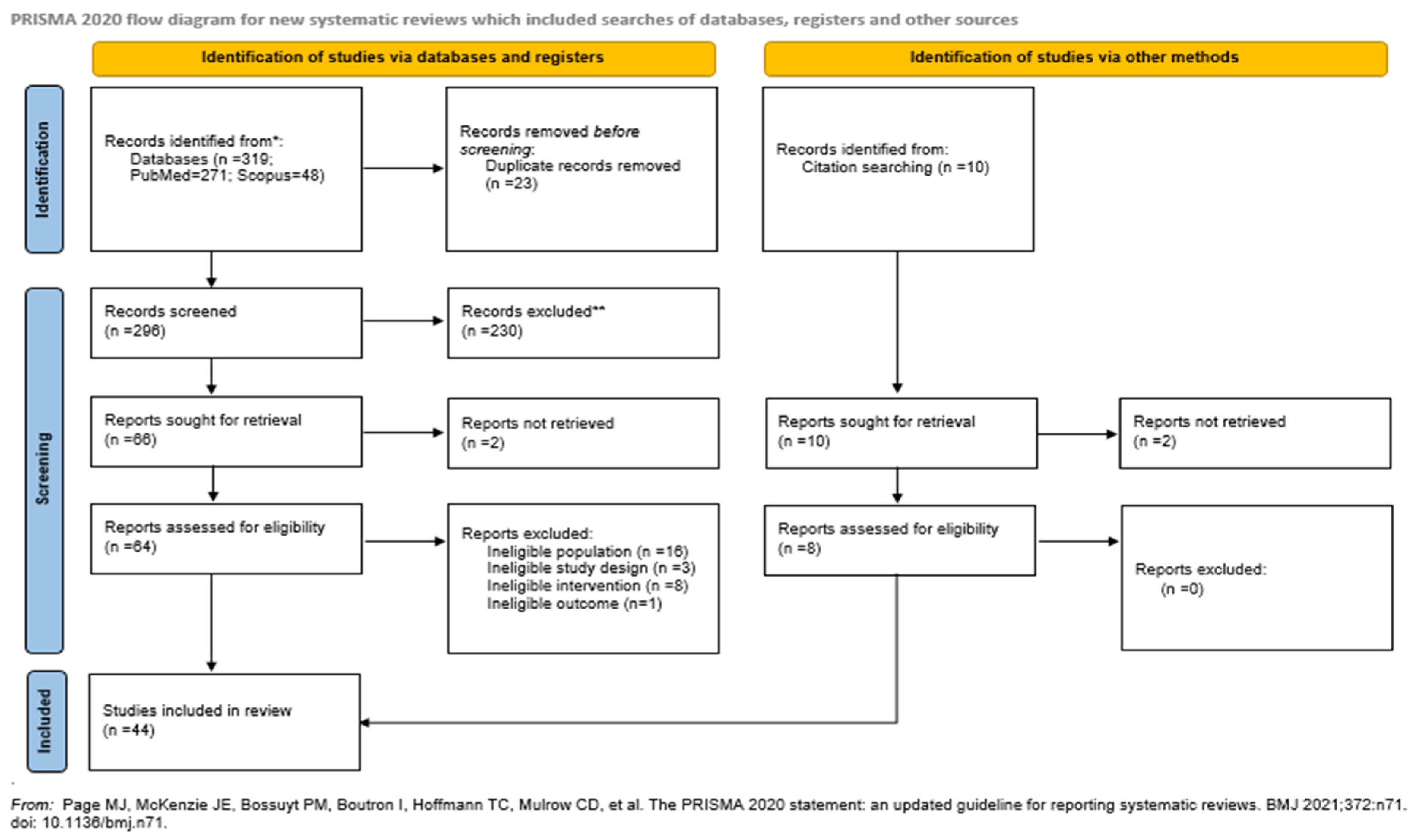
A flow diagram of the selection process is presented in Figure 1. 319 papers were initially identified and after duplicate removal, 296 were considered eligible for title-abstract screening. Subsequently, 66 articles were selected for full text screening, 20 of them were excluded for reasons presented in Figure 1, while 36 met the inclusion criteria and were included in this review. Furthermore, the references of the included studies and references from other relevant studies from high impact journal were hand-searched and 8 papers that were lost from the initial literature search were included as well. Thus, the 44 studies that were included in total investigated the association between vaginal microbial composition and preterm birth (PTB), miscarriage, preeclampsia (PE), ectopic pregnancy, gestational diabetes mellitus (GDM), chorioamnionitis (CAT) and preterm premature rupture of membranes (PPROM).
3.1.1. Preterm Birth
Preterm birth is one of the leading causes of neonatal morbidity and mortality accounting for virtually 15 million births annually worldwide [21]. Many conditions can provoke PTB, including preterm premature rapture of membranes or infections [22]. Lactobacilli, which are usually the predominant species in the vaginal microbiome, antagonize against dysbiosis-causing microorganisms and restrain the proliferation of anaerobes commonly correlated with bacterial vaginosis [23]. Thus, theoretically disruption in the vaginal microbial composition could undermine the protective mechanisms against vaginal dysbiosis and consequently against PTB.
Multiple studies have inquired into the probable association between vaginal microbial composition and PTB. The largest case-control study with 449 participants (94 in the PTB arm and 355 in the healthy control arm) suggested that L. gasseri/L. johnsonii, L. crispatus, L. acidophilus, L. iners, R. solanacearum and B. longum/B. breve might be associated with decreased risk of early but not late PTB. Furthermore, no correlation was found between CST assignment and PTB risk [24]. No correlation was either found between specific CSTs and PTB in a retrospective study, where Lactobacillus dominated communities had an inverse association with PTB in women swabbed before 12 weeks and a direct association with women swabbed at or after 12 weeks [25]. Similar results arise from a small retrospective study, a prospective study and two case-control studies where the abundance or depletion of Lactobacillus in the vaginal microbiome and the CST assignment were not significantly associated with PTB [26,27,28]. Additionally, results from another case-control study indicate that, although in the term group the richness and diversity of the microbiome remained stable through the course of pregnancy while in the PTB group both richness and diversity were significantly decreasing until labor, after taxonomic composition analysis, no association between any of the detected taxa and PTB reached statistical significance [29].
Given that the first indicator of important differences in microbial composition is the elevated alpha-diversity many studies have employed it as an initial step of their analysis. Starting from the alpha diversity analysis, which revealed statistically significant differences regarding richness and diversity between the groups (p<0.01), higher bacterial loads and higher rates of mollicutes, meaning Mycoplasma and Ureaplasma, were spotted in the PTB compared to the control group (p= 0.049 and p=0.012 respectively) [30]. In a prospective study, where the alpha-diversity assessed through Shannon indices and number of observed ASVs were higher in the PTB group (p= 0.0009 and 0.0003 respectively), increased rates of Atopobium, Gardnerella and Prevotella were observed in PTB cases. In addition, after hierarchical clustering analysis it was concluded that the PTB group was significantly more frequently assigned to CST-IV (p=0.004), while at the taxa level increased relative abundance of Lactobacillus was positively associated with term birth (p=0.007) [31]. Comparable outcomes emerge from a prospective cohort study where vaginal microbiotas of healthy uncomplicated cohort were characterized by low diversity and Lactobacillus dominance, while women who experienced PTB had intermediate or low profiles of Lactobacillus and high diversity (p=0.0011) even after the analysis was adjusted for potential confounders, such as ethnicity, BMI, smoking status, and medical interventions [32]. Except for Lactobacillus spp. depletion, increased abundance of Gardnerella is associated with PTB (p=0.0070). In fact, using an arbitrary threshold of 70% Lactobacillus spp. abundance and 0.1% of Gardnerella abundance the researchers calculated an OR for PTB of 5.81 (95% CI 1.12-33.7) and 5.12 (95% CI 1.05-31.1) respectively [33].
Besides diversity and richness of the vaginal microbiome, differences seem to be present in CST assignment as well [34]. More specifically, in a prospective study, women delivering at term were mostly assigned to CST-IV, while women from the PTB arm were mostly assigned to CST-III [35]. Contradicting results emerge from a cross-sectional study where both CST-III (L. iners dominated) and CST-IV were associated with spontaneous PTB with an OR as high as 4.1 (95% CI 1.1-infinity) and 7.7 (95% CI 2.2-infinity), respectively. In addition, in marginal analysis the relative abundance of Gardnerella vaginalis (p=0.011), non-iners Lactobacillus (p=0.016), and Mobiluncus curtisii (p=0.035) and the presence of Atopobium vaginae (p=0.049), Bacterial Vaginosis-Associated Bacteria (BVAB)-2 (p=0.024), Dialister microaerophilis (p=0.011), and Prevotella amnii (p=0.044) were associated with spontaneous PTB [36]. Moreover, in a prospective study, a higher frequency of CST-I assignment was observed in the term group compared to the preterm both at 20-22 weeks gestation and 26-28 weeks gestation (40.32 vs. 16.66% p=0.0002 and 20.69 vs. 16.66% p=0.03 respectively). Also, a diminished proportion of CST-V dominant microbiota was present in the term compared to the preterm group at both time points (9.68 and 22.22% p=0.0002 and 10.34 and 25% p=0.03 respectively). In addition, at 26-28 weeks gestation CST-II was assigned to 28% of term patients and to no patients in the preterm group (p<0.0001) [37].
Assigning vaginal microbiomes in CSTs constitutes the basis for a more systematic analysis but taxonomic composition analysis can provide further information. For example, a retrospective study concluded that the absence of elevated numbers of OTUs of L. iners and L. jensenii might be the main difference between women delivering at term compared to PTB and might serve as a biomarker for PTB prediction [38]. Accordingly, to the aforementioned results, a cross-sectional study indicated that in vaginal specimens obtained at 16 weeks gestation the dominance of L. iners was significantly higher in early PTB cases compared to term pregnancies (p=0.003), while L. Crispatus dominant microbiomes more frequently derived from the control group (p=0.009) [39]. The association of L. iners with PTB was identified by a prospective study too (p<0.001), which also added that the combination of two or more Lactobacillus species had a beneficial impact on pregnancy duration [40]. The relative abundance of various Lactobacillus species was investigated by a prospective study which used a PCR array specific for 15 bacteria and no significant association was observed with PTB, whereas high numbers of L. crispatus, L gasseri, or L jensenii was negatively associated with sPTB (p=0.05) while at the same time the detection of L. gasseri was predictive of term birth (p=0.017) [41].
Besides Lactobacillus spp. relative abundance evaluation microbiome analysis aims, also, at detecting differences in other microbial taxa. For instance, diversity in microbial composition was higher in the PTB group and the relative abundance of several taxa including BVAB1, Prevotella cluster 2 and Seanthia amnii were also higher in the PTG arm reaching statistical significance (p<0.05). In swabs collected early in pregnancy (6-24 weeks gestation), the presence of Megasphera type 1 and TMP-H1 were associated with PTB. In fact, the incorporation of S. amnii, Prevotella cluster-2, TMP-H1 and BVAB1 in an early prediction model for PTB in swabs collected at 24 weeks gestation or earlier exhibited a sensitivity of 77.4% and specificity of 76.3% [42].
From the analysis of the longitudinal samples from a prospective study it was concluded that women in the PTB group experience an increase in P. buccalis (p< 0.0001) while women delivering at term have a significant increase in L. crispatus and Finegolidia (p=0.0131 and p<0.0001 respectively) [43]. Several other taxa, including Gardnerella spp., Mobiluncus curtsii/mulieris and Sneathia sanguinegens, with M. curtsii/mulieris, genus Atopobium, and genus Megasphaera exert a significant association with PTB [44,45,46]. Interestingly, a case-control study expanding the analysis beyond bacterial to viral composition of the vaginal environment concluded that Among Papillomaviridae, Polyomaviridae, Herpesviridae, Poxviridae, Adenoviridae, and Anelloviridae, no virus or viral group was found to be associated with PTB. However, increased viral richness was a predictor of PTB risk (p=0.0005) and conversely low viral richness was correlated with term birth (p=0.03). Also, having both bacterial and viral diversity during the first trimester was a significant predictor of PTB (RR 3.12 95% CI 1.00-9.83 p=0.04) [47].
3.1.2. Miscarriage
Miscarriage is a common issue in obstetrics complicating about 25% of pregnancies worldwide and leading to an estimated average of 44 pregnancy losses per minute [48]. Miscarriages can be divided into two categories based on the time point of occurrence: early miscarriages happening before 12 weeks gestation and late miscarriages happening between 12- and 22-weeks’ gestation. Also, recurrent miscarriage defined as three or more consecutive miscarriages constitutes another variation of the condition [49,50].
Many studies attempted to elucidate the association between vaginal microbial composition and miscarriage. First, women experiencing miscarriage seem to have a vaginal microbiome characterized by increased diversity (p= 2.33*10⁻⁸) and richness (p=0.0005) compared to women with healthy ongoing pregnancies. This diversity might derive from differences in abundance rates of Bacteroides plebeius, Bifidobacterium breve, Gardnerella vaginalis, Mycoplasma girerdii and L. iners, Gardnerella and Prevotella, respectively, but no analysis for statistical significance regarding taxonomy was conducted in these studies [51,52]. Moreover, a relatively large prospective study observed that miscarriage in both first and second trimester is associated with Lactobacillus species depletion (p=0.0053), while the predominance of CST-IV was significantly different in the miscarriage group compared to the control group (p= 0.031). Interestingly, the Lactobacillus species depletion and the high bacterial diversity precede the diagnosis of miscarriage [53].
Furthermore, based on the results of the alpha and beta diversity analysis which revealed an increased microbial diversity in miscarriage cases (Shannon index 5.48 vs 5.18 p=0.02), a prospective study attempted to discover the origins of this intergroup diversity. Although Lactobacillus was the predominant species in both arms, the relative abundance was lower in the case than in the reference group (16.51% vs 23.00% p<0.05) and simultaneously a depleted abundance of L. jensenii and L. gasseri was observed in the miscarriage group (p = 0.00078 and p = 0.00069 respectively). Additionally, differences in the microbial diversity were attributed to greater abundance of Mycoplasma genitalium and Ureaplasmas (13.09% vs 10.38% and 9.18% vs 6.59% respectively, both p< 0.05) in the reference group [54]. Increased Ureaplasma species rate, specifically U. parvum, has been, also, reported in another study along with more frequent assignment to CST-III [34]. Contrary, a prospective study concluded that elevated BVAB-3 log concentration in women experiencing miscarriage was the only significant dissimilarity between the two groups (4.27 vs 3.71 p=0.012). More specifically, in women aged <21 years one unit increase in the BVAB-3 log concentration wound elevate the risk of miscarriage by 67.8% [55]. The same authors conducted the largest prospective relative study and concluded that the outcome of interest, meaning the second trimester pregnancy loss, was significantly associated with diminished loads of Lactobacilli early in pregnancy even after adjusting for confounding factors (HR 1.32 95% CI 1.10-1.64) [56].
Recurrent miscarriage (RM) has been investigated by several studies as a different outcome in non-pregnant women. The first study to ever inquire into this question employed the ACE, Simpson, and Chao diversity indices to assess alpha diversity and no statistically significant differences occurred. Only bacterial richness was higher in the RM group, meaning that more Operational Taxonomic Units (OTUs) were detected in the case group. After taxonomic composition analysis it was indicated that Atopobium, Prevotella and Streptococcus taxa were significantly more abundant in miscarriage group, whereas Lactobacillus and Gardnerella were more commonly found in the control group [57]. In agreement with these results comes a case-control study which revealed that Lactobacillus iners was significantly decreased while Ruminococcaceae_UCG-005 and Anaerococcus hydrogenalis were significantly more abundant in the RM group (p<0.05). Two taxa had significantly higher relative abundance in the control group including Lactobacillus and Gardnerella [58]. In partial contrast with the two studies above comes a cross-sectional study reporting decreased rates of Lactobacillus species in the RM group but increased relative abundance of Garnerella vaginalis, Prevotella bivia and Porphyromonas spp. (p<0.05) [59]. Elevated G. vaginalis rates have been again reported in the RM arm compared to a healthy control arm of a cross-sectional study (8.7% vs 5.7% p=0.001) [60]. Using women with medically induced abortion as controls, another cross-sectional study showed that alpha diversity is increased in RM women (p<0.05) and at the genus level the expressive abundance of Pseudomonas, Roseburia, Collinsella aerofaciens and Arthrobacter is higher [61]. Only one study concluded that although there was a significant difference in beta diversity (p=0.036) neither the alpha diversity not the taxonomic composition analysis of the vaginal microbiome revealed significant dissimilarities between the groups [62].
3.1.3. Gestational Diabetes Mellitus (GDM)
Gestational diabetes Mellitus (GDM) constitutes a major burden not only on the pregnant woman’s wellbeing but also on the health care system with a prevalence of approximately 5-20% [63]. The pathophysiology of GDM is partially known with the alterations in the hormonal and metabolic profile during pregnancy leading to a decrease in insulin sensitivity, which can sometimes result in the development of glucose intolerance and GDM [64].
Given that microbial composition of the vagina undergoes significant changes during pregnancy some studies tried to identify a connection between GDM development and vaginal microbiome. Cortez et al. reported that the vaginal specimens exhibited significant difference regarding alpha diversity with the control group having lower richness and diversity indices (p<0.01) with the GDM arm presenting a significantly higher abundance of the genera Bacteroides, Veillonella, Klebsiella, Escherichia-Shigella, Enterococcus, and Enterobacter (p<0.01) [65]. In respect of fungal microbiome, increased richness, and evenness indices in the GDM group pointed out that women with GDM have a more diverse fungal flora than healthy controls. This diversity was derived by the fact that in GDM-positive women except for Candida and Saccharomyces, which were the most predominant fungal species, the fungal load attributed to uncultured fungi was significantly lower than in the control arm (p<0.01) [66]. However, one study revealed no variations of the vaginal microbiome between women with GDM and healthy comparators [67].
3.1.4. Preeclampsia
Preeclampsia is a major complication of pregnancy associated with fetal prematurity and long term maternal cardiovascular morbidity [67]. It is estimated that affects approximately 4.6% of the pregnancies globally [68].
Only one study assessed the effect of vaginal microbiome aberrations on severe preeclampsia (SPE) development. In the context of alpha diversity, the two groups had similar richness but since Shannon and Gini-Simpson indices were higher in cases with SPE (p=0.001 and p<0.001 respectively) it was concluded that they had greater diversity than the controls. In the phylum level, the relative abundance of Bacteroides was significantly elevated in the SPE group (3.13% vs 0.18% p=0.015). After multivariable logistic regression in the genus and species level, Prevotella genus and more specifically P. bivia were significantly associated with SPE [69].
3.1.5. Chorioamnionitis (CAT)
Chorioamnionitis can present in 4% of full-term births but histological CAT can be detected in up to 94% of deliveries occurring between 21- and 24-weeks’ gestation [70]. Since CAT is caused by ascending infection originating from the cervical and vaginal area alterations in the vaginal microbiome can possibly be associated with CAT development [71].
Only one study investigated the association between CAT and vaginal microbiome composition. In this prospective observational study, the alpha diversity analysis indicated that the clinical chorioamnionitis group was characterized by a richer and diverse vaginal microbiome compared to the healthy control group. Also, the relative abundance of L. crispatus was significantly higher in the control group and conversely after logistic regression analysis it was found that depletion of L. crispatus was associated with higher risk of clinical CAT [72].
3.1.6. Ectopic Pregnancy
Ectopic pregnancy rupture accounts for 5-10% of all pregnancy related deaths and tubal pregnancy, meaning a gestational sac that implants in the salpinx, constitutes the most common type of ectopic pregnancy [73]. Women with confirmed ectopic pregnancy exhibited a greater alpha diversity as it is indicated by the Shannon diversity index (1.43 vs 0.99 p=0.03). Additionally, setting a relative abundance threshold of 85% for Lactobacillus the researchers classified the vaginal microbiomes as Lactobacillus dominated and Non-Lactobacillus-Dominated Vaginal Microbiota (NLDM). After adjusting for confounding factors, they concluded that there is a positive association between NLDM and ectopic pregnancy (OR 4.42 95% CI 1.33-14.71 p=0.02) [74].
3.1.7. Preterm Premature Rapture of Membranes (PPROM)
The prevalence of PPROM varies globally from 1-4% and its association with PTB has been well established contributing to virtually 30-40% of premature births [75,76]. One study attempted to provide insight in the relationship between characteristics of the vaginal microbiome and PPROM. The bacterial composition in the context of alpha diversity analysis differed significantly in richness, evenness, and diversity between the two groups. Also, increased relative abundance of Lactobacillus iners, Gardnerella vaginalis, Prevotella bivia, Ochrobactrum sp, Prevotella timonensis, and Ureaplasma parvum and decreased relative abundance of L. gasseri were correlated with PPROM [76].
4. Discussion
Vaginal microbiome constitutes a complex ecosystem constructed by epithelial cells, immune system cells and mediators and microorganisms interacting in complex ways [77]. Given that disruption of Lactobacillus dominance and high diversity can facilitate the colonization by pathogenic microbial taxa, for a pregnancy to progress uncomplicated to full term it is of highly importance that the microbiome retains its composition [78]. The specific mechanism linking alterations in the vaginal microbiome with different pregnancy complications is yet unclear but innovative laboratory techniques and study design have offered some relevant indications. First, depletion of beneficial bacteria loads, such as Lactobacilli, and increase in pathogens results in the formation of a dysbiotic environment which leads to inflammation of the cervix and surrounding tissues. This inflammation can undermine the integrity of cervical tissues and contribute to cervical insufficiency development [79]. In addition, changes in the bacterial composition have been associated with conditions such as bacterial vaginosis (BV) which might be linked to miscarriage [80].
In respect of PTB it has been clear that increased diversity of the vaginal microbiome and reduced relative abundance of Lactobacillus species were associated with high risk of PTB [31,32,33]. Also, the increased prevalence of Gardnerella, Prevotella, Megasphera, Sneathia, Atopobium and BVABs seem to be associated with shorter duration of pregnancy, while L. crispatus might have a protective role regarding pregnancy. Since the first are components of CST-IV, while the latter is the dominant species in CST-I, it is not surprising that CST-IV and CST-I are associated with preterm and term birth respectively [36,39]. Since Prevotella and Gardnerella trigger the immune response by promoting the proinflammatory cytokine production and are correlated with PPROM, it can be expected that they can affect the risk of PTB [76,81]. Lowered Lactobacillus loads and increased alpha diversity and relative abundance of non-lactobacilli bacteria are not only observed in PTB, but also in miscarriage, GDM, ectopic pregnancy, CAT and preeclampsia. This is attributed to virulence factors produced by these non-lactobacilli bacteria, such as sialidase, hyaluronidase or IgA protease, that activate inflammatory pathways that probably constitute the starting point of these complications [82,83].
Despite the mechanism through which the vaginal microbiome exerts its impact on pregnancy outcome is not totally understood, since among the factors that can modify it antibiotics pre and probiotics are included, an intervention strategy could be designed [84]. It has been already proven that probiotic lactobacilli reduce BV recurrence and can increase lactobacilli abundance [85]. However, the use of Lactobacillus and Bifidobacterium as probiotics in pregnant women seemed to have no effect on gestational age [86]. Moreover, the reliable establishment of certain taxa or CSTs associated with pregnancy complications can contribute to the formation of a microbial signature in high-risk pregnancies possibly utilized to promptly intervene [87].
This study has several limitations that merit careful considerations. First, relatively small sample sizes included in several of the studies jeopardize the reliability and the external validity of the results. In addition, most of the evidence included comprised of cross-sectional and retrospective cohort studies, meaning two study types both associated with selection bias. Furthermore, there were several factors regarding microbial analysis in the included studies that could have influenced the results, such as sample collection, extraction methods and hypervariable region used for sequencing. Among the included studies there was great heterogeneity regarding the vaginal sampling with some studies not even defining the sampling time point. Finally, the racioethnic composition of the cohorts is a factor that could have altered the effect size of vaginal microbiome alterations on pregnancy complication incidence.
5. Conclusions
To sum up, pregnancy complications pose a major burden on maternal and fetal health and well-being. New approaches are being applied in order not only to trace women at high risk of developing these disorders but also to develop appropriate prevention strategies. One of these approaches is facilitated through the analysis and the potential of the maternal reproductive tract microbiome. Given that high diversity and richness, depleted predominance of Lactobacillus species and increased abundance of specific taxa, such as Gardnerella and Prevotella, have been associated with several pregnancy complications indicates that vaginal microbiome constitutes a possible target. However, it is important that more high-quality studies with greater racioethnic diversity and more homogenous design in respect of sampling time point and sequencing techniques are conducted.
Author Contributions
Conceptualization, A.G. and E.D.; methodology, A.G., and E.V.; investigation, A.G, K.N., E.V., C.M.S. M.G., P.F., M.S. and S.S.; data curation, E.V.; writing—original draft preparation, A.G., K.N., E.V., C.M.S., M.G., P.F., M.S. and S.S.; writing—review and editing, A.G., S.P., K.D., N.N. and E.D.; visualization, E.V.; supervision, S.P., K.D., N.N and E.D.; project administration, E.D.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Law A, McCoy M, Lynen R, Curkendall SM, Gatwood J, Juneau PL, et al. The prevalence of complications and healthcare costs during pregnancy. J Med Econ [Internet]. 2015;18:533–41. [CrossRef]
- Creanga AA, Berg CJ, Ko JY, Farr SL, Tong VT, Bruce FC, et al. Maternal mortality and morbidity in the United States: where are we now? J Womens Health (Larchmt) [Internet]. 2014;23:3–9. Available from: https://pubmed.ncbi.nlm.nih.gov/24383493. [CrossRef]
- Say L, Pattinson RC, Gülmezoglu AM. WHO systematic review of maternal morbidity and mortality: the prevalence of severe acute maternal morbidity (near miss). Reprod Health [Internet]. 2004;1:3. Available from: https://pubmed.ncbi.nlm.nih.gov/15357863. [CrossRef]
- Paulo MS, Abdo NM, Bettencourt-Silva R, Al-Rifai RH. Gestational Diabetes Mellitus in Europe: A Systematic Review and Meta-Analysis of Prevalence Studies. Front Endocrinol (Lausanne) [Internet]. 2021;12:691033. Available from: https://pubmed.ncbi.nlm.nih.gov/34956073. [CrossRef]
- Umesawa M, Kobashi G. Epidemiology of hypertensive disorders in pregnancy: prevalence, risk factors, predictors and prognosis. Hypertension Research [Internet]. 2016;40:213–20. [CrossRef]
- Otolorin E, Gomez P, Currie S, Thapa K, Dao B. Essential basic and emergency obstetric and newborn care: From education and training to service delivery and quality of care. International Journal of Gynecology & Obstetrics [Internet]. 2015;130. [CrossRef]
- Poon LC, Nicolaides KH. Early prediction of preeclampsia. Obstet Gynecol Int [Internet]. 2014/07/17. 2014;2014:297397. Available from: https://pubmed.ncbi.nlm.nih.gov/25136369. [CrossRef]
- Stupak A, Kwaśniewski W. Evaluating Current Molecular Techniques and Evidence in Assessing Microbiome in Placenta-Related Health and Disorders in Pregnancy. Biomolecules [Internet]. 2023;13:911. Available from: https://pubmed.ncbi.nlm.nih.gov/37371491. [CrossRef]
- Taddei CR, Cortez R V, Mattar R, Torloni MR, Daher S. Microbiome in normal and pathological pregnancies: A literature overview. American Journal of Reproductive Immunology [Internet]. 2018;80. [CrossRef]
- Lv L-J, Li S-H, Li S-C, Zhong Z-C, Duan H-L, Tian C, et al. Early-Onset Preeclampsia Is Associated With Gut Microbial Alterations in Antepartum and Postpartum Women. Front Cell Infect Microbiol [Internet]. 2019;9:224. Available from: https://pubmed.ncbi.nlm.nih.gov/31297341. [CrossRef]
- Swati P, Thomas B, Vahab SA, Kapaettu S, Kushtagi P. Simultaneous detection of periodontal pathogens in subgingival plaque and placenta of women with hypertension in pregnancy. Arch Gynecol Obstet [Internet]. 2011;285:613–9. [CrossRef]
- Sobel JD. Is there a protective role for vaginal flora? Curr Infect Dis Rep [Internet]. 1999;1:379–83. [CrossRef]
- Boskey ER, Cone RA, Whaley KJ, Moench TR. Origins of vaginal acidity: high d/l lactate ratio is consistent with bacteria being the primary source. Human Reproduction [Internet]. 2001;16:1809–13. [CrossRef]
- Kaewsrichan J, Peeyananjarassri K, Kongprasertkit J. Selection and identification of anaerobic lactobacilli producing inhibitory compounds against vaginal pathogens. FEMS Immunology & Medical Microbiology [Internet]. 2006;48:75–83. [CrossRef]
- Fortenberry JD. The uses of race and ethnicity in human microbiome research. Trends Microbiol [Internet]. 2013;21:165–6. [CrossRef]
- Aagaard K, Riehle K, Ma J, Segata N, Mistretta T-A, Coarfa C, et al. A metagenomic approach to characterization of the vaginal microbiome signature in pregnancy. PLoS One [Internet]. 2012/06/13. 2012;7:e36466–e36466. Available from: https://pubmed.ncbi.nlm.nih.gov/22719832. [CrossRef]
- Serrano MG, Parikh HI, Brooks JP, Edwards DJ, Arodz TJ, Edupuganti L, et al. Racioethnic diversity in the dynamics of the vaginal microbiome during pregnancy. Nat Med [Internet]. 2019/05/29. 2019;25:1001–11. Available from: https://pubmed.ncbi.nlm.nih.gov/31142850. [CrossRef]
- Ravel J, Gajer P, Abdo Z, Schneider GM, Koenig SSK, McCulle SL, et al. Vaginal microbiome of reproductive-age women. Proc Natl Acad Sci U S A [Internet]. 2010/06/03. 2011;108 Suppl 1:4680–7. Available from: https://pubmed.ncbi.nlm.nih.gov/20534435. [CrossRef]
- Abdulla SR, Kareem SR, Hasan AH. Vaginal Microbiota Profile in first-trimester miscarriages cases. Cell Mol Biol [Internet]. 2023;69:9–17. [CrossRef]
- Ouzzani M, Hammady H, Fedorowicz Z, Elmagarmid A. Rayyan-a web and mobile app for systematic reviews. Syst Rev [Internet]. 2016;5:210. Available from: https://pubmed.ncbi.nlm.nih.gov/27919275. [CrossRef]
- Blencowe H, Cousens S, Oestergaard MZ, Chou D, Moller A-B, Narwal R, et al. National, regional, and worldwide estimates of preterm birth rates in the year 2010 with time trends since 1990 for selected countries: a systematic analysis and implications. The Lancet [Internet]. 2012;379:2162–72. [CrossRef]
- Baldwin EA, Walther-Antonio M, MacLean AM, Gohl DM, Beckman KB, Chen J, et al. Persistent microbial dysbiosis in preterm premature rupture of membranes from onset until delivery. PeerJ [Internet]. 2015;3:e1398–e1398. Available from: https://pubmed.ncbi.nlm.nih.gov/26644969. [CrossRef]
- Aldunate M, Srbinovski D, Hearps AC, Latham CF, Ramsland PA, Gugasyan R, et al. Antimicrobial and immune modulatory effects of lactic acid and short chain fatty acids produced by vaginal microbiota associated with eubiosis and bacterial vaginosis. Front Physiol [Internet]. 2015 [cited 2024 Mar 15];6:164. Available from: /pmc/articles/PMC4451362/. [CrossRef]
- Tabatabaei N, Eren AM, Barreiro LB, Yotova V, Dumaine A, Allard C, et al. Vaginal microbiome in early pregnancy and subsequent risk of spontaneous preterm birth: a case–control study. BJOG: An International Journal of Obstetrics & Gynaecology [Internet]. 2018;126:349–58. [CrossRef]
- Blostein F, Gelaye B, Sanchez SE, Williams MA, Foxman B. Vaginal microbiome diversity and preterm birth: results of a nested case-control study in Peru. Ann Epidemiol [Internet]. 2019/12/05. 2020;41:28–34. Available from: https://pubmed.ncbi.nlm.nih.gov/31883841. [CrossRef]
- Ng S, Chen M, Kundu S, Wang X, Zhou Z, Zheng Z, et al. Large-scale characterisation of the pregnancy vaginal microbiome and sialidase activity in a low-risk Chinese population. NPJ Biofilms Microbiomes [Internet]. 2021;7:89. Available from: https://pubmed.ncbi.nlm.nih.gov/34930922. [CrossRef]
- Hyman RW, Fukushima M, Jiang H, Fung E, Rand L, Johnson B, et al. Diversity of the vaginal microbiome correlates with preterm birth. Reprod Sci [Internet]. 2013/05/28. 2014;21:32–40. Available from: https://pubmed.ncbi.nlm.nih.gov/23715799. [CrossRef]
- Gulavi E, Mwendwa F, Atandi DO, Okiro PO, Hall M, Beiko RG, et al. Vaginal microbiota in women with spontaneous preterm labor versus those with term labor in Kenya: a case control study. BMC Microbiol [Internet]. 2022;22:270. Available from: https://pubmed.ncbi.nlm.nih.gov/36357861. [CrossRef]
- Stout MJ, Zhou Y, Wylie KM, Tarr PI, Macones GA, Tuuli MG. Early pregnancy vaginal microbiome trends and preterm birth. Am J Obstet Gynecol [Internet]. 2017/05/23. 2017;217:356.e1-356.e18. Available from: https://pubmed.ncbi.nlm.nih.gov/28549981. [CrossRef]
- Freitas AC, Bocking A, Hill JE, Money DM, Group VR. Increased richness and diversity of the vaginal microbiota and spontaneous preterm birth. Microbiome [Internet]. 2018;6:117. Available from: https://pubmed.ncbi.nlm.nih.gov/29954448. [CrossRef]
- Odogwu NM, Chen J, Onebunne CA, Jeraldo P, Yang L, Johnson S, et al. Predominance of Atopobium vaginae at Midtrimester: a Potential Indicator of Preterm Birth Risk in a Nigerian Cohort. mSphere [Internet]. 2021;6:e01261-20. Available from: https://pubmed.ncbi.nlm.nih.gov/33504666. [CrossRef]
- Brown RG, Marchesi JR, Lee YS, Smith A, Lehne B, Kindinger LM, et al. Vaginal dysbiosis increases risk of preterm fetal membrane rupture, neonatal sepsis and is exacerbated by erythromycin. BMC Med [Internet]. 2018 [cited 2024 Mar 15];16. Available from: /pmc/articles/PMC5782380/. [CrossRef]
- Callahan BJ, DiGiulio DB, Goltsman DSA, Sun CL, Costello EK, Jeganathan P, et al. Replication and refinement of a vaginal microbial signature of preterm birth in two racially distinct cohorts of US women. Proc Natl Acad Sci U S A [Internet]. 2017/08/28. 2017;114:9966–71. Available from: https://pubmed.ncbi.nlm.nih.gov/28847941. [CrossRef]
- Chang D-H, Shin J, Rhee M-S, Park K-R, Cho B-K, Lee S-K, et al. Vaginal Microbiota Profiles of Native Korean Women and Associations with High-Risk Pregnancy. J Microbiol Biotechnol [Internet]. 2020;30:248–58. Available from: https://pubmed.ncbi.nlm.nih.gov/31838792. [CrossRef]
- Shin H, Wu J, Nelson D, Dominguez-Bello M. The Gestational Vaginal Microbiome and Spontaneous Preterm Birth among Nulliparous African American Women. Am J Perinatol [Internet]. 2016;33:887–93. [CrossRef]
- Dunlop AL, Satten GA, Hu Y-J, Knight AK, Hill CC, Wright ML, et al. Vaginal Microbiome Composition in Early Pregnancy and Risk of Spontaneous Preterm and Early Term Birth Among African American Women. Front Cell Infect Microbiol [Internet]. 2021;11:641005. Available from: https://pubmed.ncbi.nlm.nih.gov/33996627. [CrossRef]
- Stafford GP, Parker JL, Amabebe E, Kistler J, Reynolds S, Stern V, et al. Spontaneous Preterm Birth Is Associated with Differential Expression of Vaginal Metabolites by Lactobacilli-Dominated Microflora. Front Physiol [Internet]. 2017;8:615. Available from: https://pubmed.ncbi.nlm.nih.gov/28878691. [CrossRef]
- de Freitas AS, Dobbler PCT, Mai V, Procianoy RS, Silveira RC, Corso AL, et al. Defining microbial biomarkers for risk of preterm labor. Braz J Microbiol [Internet]. 2019/07/22. 2020;51:151–9. Available from: https://pubmed.ncbi.nlm.nih.gov/31332740. [CrossRef]
- Kindinger LM, Bennett PR, Lee YS, Marchesi JR, Smith A, Cacciatore S, et al. The interaction between vaginal microbiota, cervical length, and vaginal progesterone treatment for preterm birth risk. Microbiome [Internet]. 2017;5:6. Available from: https://pubmed.ncbi.nlm.nih.gov/28103952. [CrossRef]
- Petricevic L, Domig KJ, Nierscher FJ, Sandhofer MJ, Fidesser M, Krondorfer I, et al. Characterisation of the vaginal Lactobacillus microbiota associated with preterm delivery. Sci Rep [Internet]. 2014;4:5136. Available from: https://pubmed.ncbi.nlm.nih.gov/24875844. [CrossRef]
- Payne MS, Newnham JP, Doherty DA, Furfaro LL, Pendal NL, Loh DE, et al. A specific bacterial DNA signature in the vagina of Australian women in midpregnancy predicts high risk of spontaneous preterm birth (the Predict1000 study). Am J Obstet Gynecol [Internet]. 2021;224:206.e1-206.e23. [CrossRef]
- Fettweis JM, Serrano MG, Sheth NU, Mayer CM, Glascock AL, Brooks JP, et al. Species-level classification of the vaginal microbiome. BMC Genomics [Internet]. 2012 [cited 2024 Mar 15];13:S17. Available from: /pmc/articles/PMC3535711/. [CrossRef]
- Kumar M, Murugesan S, Singh P, Saadaoui M, Elhag DA, Terranegra A, et al. Vaginal Microbiota and Cytokine Levels Predict Preterm Delivery in Asian Women. Front Cell Infect Microbiol [Internet]. 2021;11:639665. Available from: https://pubmed.ncbi.nlm.nih.gov/33747983. [CrossRef]
- Menard J, Fenollar F, Henry M, Bretelle F, Raoult D. Molecular Quantification ofGardnerella vaginalisandAtopobium vaginaeLoads to Predict Bacterial Vaginosis. Clinical Infectious Diseases [Internet]. 2008;47:33–43. [CrossRef]
- Elovitz MA, Gajer P, Riis V, Brown AG, Humphrys MS, Holm JB, et al. Cervicovaginal microbiota and local immune response modulate the risk of spontaneous preterm delivery. Nat Commun [Internet]. 2019;10:1305. Available from: https://pubmed.ncbi.nlm.nih.gov/30899005. [CrossRef]
- Liao J, Shenhav L, Urban JA, Serrano M, Zhu B, Buck GA, et al. Microdiversity of the vaginal microbiome is associated with preterm birth. Nat Commun [Internet]. 2023;14:4997. Available from: https://pubmed.ncbi.nlm.nih.gov/37591872. [CrossRef]
- Wylie KM, Wylie TN, Cahill AG, Macones GA, Tuuli MG, Stout MJ. The vaginal eukaryotic DNA virome and preterm birth. Am J Obstet Gynecol [Internet]. 2018/05/05. 2018;219:189.e1-189.e12. Available from: https://pubmed.ncbi.nlm.nih.gov/29738749. [CrossRef]
- Quenby S, Gallos ID, Dhillon-Smith RK, Podesek M, Stephenson MD, Fisher J, et al. Miscarriage matters: the epidemiological, physical, psychological, and economic costs of early pregnancy loss. The Lancet [Internet]. 2021;397:1658–67. [CrossRef]
- Larsen EC, Christiansen OB, Kolte AM, Macklon N. New insights into mechanisms behind miscarriage. BMC Med [Internet]. 2013;11:154. Available from: https://pubmed.ncbi.nlm.nih.gov/23803387. [CrossRef]
- Devall AJ, Coomarasamy A. Sporadic pregnancy loss and recurrent miscarriage. Best Practice & Research Clinical Obstetrics & Gynaecology [Internet]. 2020;69:30–9. [CrossRef]
- Gryaznova M, Kozarenko O, Smirnova Y, Burakova I, Syromyatnikov M, Maslov A, et al. Cervical and Vaginal Microbiomes in Early Miscarriages and Ongoing Pregnancy with and without Dydrogesterone Usage. Int J Mol Sci [Internet]. 2023;24:13836. Available from: https://pubmed.ncbi.nlm.nih.gov/37762139. [CrossRef]
- Shahid M, Quinlivan JA, Peek M, Castaño-Rodríguez N, Mendz GL. Is there an association between the vaginal microbiome and first trimester miscarriage? A prospective observational study. Journal of Obstetrics and Gynaecology Research [Internet]. 2021;48:119–28. [CrossRef]
- Al-Memar M, Bobdiwala S, Fourie H, Mannino R, Lee YS, Smith A, et al. The association between vaginal bacterial composition and miscarriage: a nested case-control study. BJOG [Internet]. 2019/10/31. 2020;127:264–74. Available from: https://pubmed.ncbi.nlm.nih.gov/31573753. [CrossRef]
- Sun D, Zhao X, Pan Q, Li F, Gao B, Zhang A, et al. The association between vaginal microbiota disorders and early missed abortion: A prospective study. Acta Obstet Gynecol Scand [Internet]. 2022/07/24. 2022;101:960–71. Available from: https://pubmed.ncbi.nlm.nih.gov/35871770. [CrossRef]
- Nelson DB, Hanlon AL, Wu G, Liu C, Fredricks DN. First Trimester Levels of BV-Associated Bacteria and Risk of Miscarriage Among Women Early in Pregnancy. Matern Child Health J [Internet]. 2015;19:2682–7. [CrossRef]
- Nelson DB, Bellamy S, Nachamkin I, Ness RB, Macones GA, Allen-Taylor L. First trimester bacterial vaginosis, individual microorganism levels, and risk of second trimester pregnancy loss among urban women. Fertil Steril [Internet]. 2007/04/16. 2007;88:1396–403. Available from: https://pubmed.ncbi.nlm.nih.gov/17434499. [CrossRef]
- Zhang F, Zhang T, Ma Y, Huang Z, He Y, Pan H, et al. Alteration of vaginal microbiota in patients with unexplained recurrent miscarriage. Exp Ther Med [Internet]. 2019/03/04. 2019;17:3307–16. Available from: https://pubmed.ncbi.nlm.nih.gov/30988706. [CrossRef]
- Jiao X, Zhang L, Du D, Wang L, Song Q, Liu S. Alteration of vaginal microbiota in patients with recurrent miscarriage. J Obstet Gynaecol (Lahore) [Internet]. 2021;42:248–55. [CrossRef]
- Soyer Caliskan C, Yurtcu N, Celik S, Sezer O, Kilic SS, Cetin A. Derangements of vaginal and cervical canal microbiota determined with real-time PCR in women with recurrent miscarriages. J Obstet Gynaecol (Lahore) [Internet]. 2022;42:2105–14. [CrossRef]
- Peuranpää P, Holster T, Saqib S, Kalliala I, Tiitinen A, Salonen A, et al. Female reproductive tract microbiota and recurrent pregnancy loss: a nested case-control study. Reprod Biomed Online [Internet]. 2022;45:1021–31. [CrossRef]
- Fan T, Zhong X-M, Wei X-C, Miao Z-L, Luo S-Y, Cheng H, et al. The alteration and potential relationship of vaginal microbiota and chemokines for unexplained recurrent spontaneous abortion. Medicine [Internet]. 2020;99:e23558–e23558. Available from: https://pubmed.ncbi.nlm.nih.gov/33371084. [CrossRef]
- Liu F-T, Yang S, Yang Z, Zhou P, Peng T, Yin J, et al. An Altered Microbiota in the Lower and Upper Female Reproductive Tract of Women with Recurrent Spontaneous Abortion. Microbiol Spectr [Internet]. 2022/05/23. 2022;10:e0046222–e0046222. Available from: https://pubmed.ncbi.nlm.nih.gov/35604131. [CrossRef]
- Ponzo V, Fedele D, Goitre I, Leone F, Lezo A, Monzeglio C, et al. Diet-Gut Microbiota Interactions and Gestational Diabetes Mellitus (GDM). Nutrients [Internet]. 2019;11:330. Available from: https://pubmed.ncbi.nlm.nih.gov/30717458. [CrossRef]
- Hasain Z, Mokhtar NM, Kamaruddin NA, Mohamed Ismail NA, Razalli NH, Gnanou JV, et al. Gut Microbiota and Gestational Diabetes Mellitus: A Review of Host-Gut Microbiota Interactions and Their Therapeutic Potential. Front Cell Infect Microbiol [Internet]. 2020;10:188. Available from: https://pubmed.ncbi.nlm.nih.gov/32500037. [CrossRef]
- Cortez R V, Taddei CR, Sparvoli LG, Ângelo AGS, Padilha M, Mattar R, et al. Microbiome and its relation to gestational diabetes. Endocrine [Internet]. 2018;64:254–64. [CrossRef]
- Zheng N-N, Guo X-C, Lv W, Chen X-X, Feng G-F. Characterization of the vaginal fungal flora in pregnant diabetic women by 18S rRNA sequencing. European Journal of Clinical Microbiology & Infectious Diseases [Internet]. 2013;32:1031–40. [CrossRef]
- Kuklina E V, Ayala C, Callaghan WM. Hypertensive Disorders and Severe Obstetric Morbidity in the United States. Obstetrics & Gynecology [Internet]. 2009;113:1299–306. [CrossRef]
- Abalos E, Cuesta C, Grosso AL, Chou D, Say L. Global and regional estimates of preeclampsia and eclampsia: a systematic review. European Journal of Obstetrics & Gynecology and Reproductive Biology [Internet]. 2013;170:1–7. [CrossRef]
- Lin C-Y, Lin C-Y, Yeh Y-M, Yang L-Y, Lee Y-S, Chao A, et al. Severe preeclampsia is associated with a higher relative abundance of Prevotella bivia in the vaginal microbiota. Sci Rep [Internet]. 2020;10:18249. Available from: https://pubmed.ncbi.nlm.nih.gov/33106556. [CrossRef]
- Suzuki S. Association between clinical chorioamnionitis and histological funisitis at term. J Neonatal Perinatal Med [Internet]. 2019;12:37–40. [CrossRef]
- Kim CJ, Romero R, Chaemsaithong P, Chaiyasit N, Yoon BH, Kim YM. Acute chorioamnionitis and funisitis: definition, pathologic features, and clinical significance. Am J Obstet Gynecol [Internet]. 2015;213:S29–52. Available from: https://pubmed.ncbi.nlm.nih.gov/26428501. [CrossRef]
- Guo X, Hong X, Qian H, Qiao D, Wang B, Yu H. Relationship between vaginal microbiota and chorioamnionitis: A prospective cohort study. Microb Pathog [Internet]. 2024;186:106458. [CrossRef]
- Houser M, Kandalaft N, Khati NJ. Ectopic pregnancy: a resident’s guide to imaging findings and diagnostic pitfalls. Emerg Radiol [Internet]. 2021;29:161–72. [CrossRef]
- Ruan X-F, Zhang Y-X, Chen S, Liu X-R, Zhu F-F, Huang Y-X, et al. Non-Lactobacillus-Dominated Vaginal Microbiota Is Associated With a Tubal Pregnancy in Symptomatic Chinese Women in the Early Stage of Pregnancy: A Nested Case-Control Study. Front Cell Infect Microbiol [Internet]. 2021;11:659505. Available from: https://pubmed.ncbi.nlm.nih.gov/34307190. [CrossRef]
- Nunes V, Cross J, Speich JE, Morgan DR, Strauss 3rd JF, Ramus RM. Fetal membrane imaging and the prediction of preterm birth: a systematic review, current issues, and future directions. BMC Pregnancy Childbirth [Internet]. 2016;16:387. Available from: https://pubmed.ncbi.nlm.nih.gov/27938341. [CrossRef]
- Yan C, Hong F, Xin G, Duan S, Deng X, Xu Y. Alterations in the vaginal microbiota of patients with preterm premature rupture of membranes. Front Cell Infect Microbiol [Internet]. 2022;12. [CrossRef]
- Gajer P, Brotman RM, Bai G, Sakamoto J, Schütte UME, Zhong X, et al. Temporal Dynamics of the Human Vaginal Microbiota. Sci Transl Med [Internet]. 2012;4. [CrossRef]
- Amabebe E, Anumba DOC. The Vaginal Microenvironment: The Physiologic Role of Lactobacilli. Front Med (Lausanne) [Internet]. 2018;5:181. Available from: https://pubmed.ncbi.nlm.nih.gov/29951482. [CrossRef]
- Ma Y, Li Y, Liu Y, Cao L, Han X, Gao S, et al. Vaginal Microbiome Dysbiosis is Associated with the Different Cervical Disease Status. Journal of Microbiology [Internet]. 2023;61:423–32. [CrossRef]
- Lewis FMT, Bernstein KT, Aral SO. Vaginal Microbiome and Its Relationship to Behavior, Sexual Health, and Sexually Transmitted Diseases. Obstetrics & Gynecology [Internet]. 2017;129:643–54. [CrossRef]
- Tsonis O, Gkrozou F, Harrison E, Stefanidis K, Vrachnis N, Paschopoulos M. Female genital tract microbiota affecting the risk of preterm birth: What do we know so far? A review. European Journal of Obstetrics & Gynecology and Reproductive Biology [Internet]. 2020;245:168–73. [CrossRef]
- Vornhagen J, Quach P, Boldenow E, Merillat S, Whidbey C, Ngo LY, et al. Bacterial Hyaluronidase Promotes Ascending GBS Infection and Preterm Birth. mBio [Internet]. 2016;7:e00781-16. Available from: https://pubmed.ncbi.nlm.nih.gov/27353757. [CrossRef]
- Cauci S, Hitti J, Noonan C, Agnew K, Quadrifoglio F, Hillier SL, et al. Vaginal hydrolytic enzymes, immunoglobulin A against Gardnerella vaginalis toxin, and risk of early preterm birth among women in preterm labor with bacterial vaginosis or intermediate flora. Am J Obstet Gynecol [Internet]. 2002;187:877–81. [CrossRef]
- Macklaim JM, Clemente JC, Knight R, Gloor GB, Reid G. Changes in vaginal microbiota following antimicrobial and probiotic therapy. Microb Ecol Health Dis [Internet]. 2015;26:27799. Available from: https://pubmed.ncbi.nlm.nih.gov/26282697. [CrossRef]
- Homayouni A, Bastani P, Ziyadi S, Mohammad-Alizadeh-Charandabi S, Ghalibaf M, Mortazavian AM, et al. Effects of Probiotics on the Recurrence of Bacterial Vaginosis. J Low Genit Tract Dis [Internet]. 2014;18:79–86. [CrossRef]
- Dugoua J-J, Machado M, Zhu X, Chen X, Koren G, Einarson TR. Probiotic Safety in Pregnancy: A Systematic Review and Meta-analysis of Randomized Controlled Trials of Lactobacillus, Bifidobacterium, and Saccharomyces spp. Journal of Obstetrics and Gynaecology Canada [Internet]. 2009;31:542–52. Http://dx.doi.org/. [CrossRef]
- Zhu B, Tao Z, Edupuganti L, Serrano MG, Buck GA. Roles of the Microbiota of the Female Reproductive Tract in Gynecological and Reproductive Health. Microbiol Mol Biol Rev [Internet]. 2022/10/12. 2022;86:e0018121–e0018121. Available from: https://pubmed.ncbi.nlm.nih.gov/36222685. [CrossRef]
Figure 1.
Flow diagram of the selection process of the included studies.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



