
Submitted:
05 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Trichinellosis is a major worldwide health concern caused by T. spiralis infection without highly efficient and safe therapeutic method so far. To solve the problem of uncomplete drug treatment schemes and the rise of anthelmintic resistance, a potential protective effect of probiotic strains against zoonotic T. spiralis infection was investigated in the framework of a new therapeutic strategy aimed at using probiotics to control parasitic zoonoses. The study was focused on L. reuteri protected against T. spiralis infection in intestinal phase throughout the detection of adult worm number, inflammatory factors mRNA expression, antioxidant enzyme expression and Nrf2 signal pathway protein expression. Mice were divided randomly into 3 groups: 1) Control group: no treatment; 2) Ts group: T. spiralis infection; 3)LAB group: T. spiralis infected after orally administered L. reuteri for 7 days. Probiotics bacteria were administered daily in a dose of 109 CFU/mL, and mice were infected with 400 T. spiralis larvae in the experimental groups. The number of adult worms was also significantly increased after administration of probiotics. Meanwhile, L. reuteri alleviate the inflammatory response caused by T. spiralis infection in intestinal has been confirmed. In addition, the promotion of antioxidant enzyme expression can be inferred that L. reuteri regulate Nrf2 signal pathways against T. spiralis infection. Our study demonstrates that L. reuteri provide protection against T. spiralis infection in intestinal. Therapeutic approaches with probiotic strains admission could help to reduce the risks of trichinellosis or complement classical anti-parasite treatments.
Keywords:
Trichinella spiralis
; Lactobacillus reuteri
; intestinal
; mice
1. Introduction
Trichinellosis is a widespread food-borne parasitic zoonosis resulted from Trichinella spiralis (T. spiralis) infection, which the pathological mechanism is associated with initial inflammatory response during the intestinal phase [1] Numerous studies demonstrate that T. spiralis is an intestinal nematode that can modulate the host immune system [2]. The efficacy of classic therapy with antiparasitic drugs limited due to weak activity against T. spiralis larvae and the emergence of antiparasitic resistance [3,4]. The development of alternative therapeutic approaches is essential, therefore, the utilisation of the beneficial-probiotic bacteria has been proposed [5].
Positive effects of probiotic bacteria reducing the parasite burden and pathological changes in experimental trichinellosis were described previously [6,7,8,9,10]. Probiotic bacteria can provide an indirect protection to the host, probably by modulation the effect on newborn and muscle T. spiralis larvae [11]. The main mechanisms of probiotic actions include enhancement of the gut epithelial barrier, increase of adhesion to the intestinal mucosa and simultaneous inhibition of pathogen adhesion, competitive elimination of pathogens, production of anti-microbial molecules, and modulation of the immune system.
It is noteworthy that increasing evidence has revealed that intestinal helminth infections are closely correlated with the gastrointestinal microbiota [12,13,14,15]. Lactobacillus (LAB), a large group of autochthonous intestinal microbiota in humans and animals, are especially distinguished for their probiotic properties [16]. Probiotics contribute to regulate Nrf2 signal pathway by balancing intestinal immunological barrier,relieveing inflammatory response [17].
T. spiralis infection is determined not only by the dose of infective larvae, but also by the immune status and inflammatory responses of the host to the parasite antigens [18].The host elicits an immune response to the parasite invasion by activation of immune mechanisms at the intestinal level, the site where the interaction between the parasite and gut microflora modify each other and the host immune system. This study aimed to explore the change in intestinal flora in the intestinal during T. spiralis infection, as well as the potential effects of probiotic on parasite survival or its underlying pathogenesis.
2. Materials and Methods
2.1. Animals and Ethics Statement
Specific pathogen free (SPF) KM mice aged 6-8 weeks were purchased from the Harbin Medical University (Harbin, China), which were housed in cages with a 12 h light and 12 h dark cycle with free access to a standard diet. All animal care procedures were in accordance with the Animal Management Committee of Northeast Agricultural University (SYXK [Hei] 2016-007). Moreover, animal suffering was minimized during the experimental procedure.
2.2. Parasite and Probiotic Strain
T. spiralis (ISS533 strain) was stored in our lab and maintained in KM mice. In addition, the muscle larvae (ML) were released through pepsin-acid digestion as previously described [19], and kept in saline solution until inoculation of experimental mice.
Probiotic strains were separated, screened, identified, characterized and evaluated according to the previous studied [5]. Lactobacillus reuteri (L. reuteri)50319 possessed extraordinary probiotic properties in vivo was prepared for experiment.
2.3. Experimental Design
Mice were divided randomly into 4 groups: Control group -no treatment; Ts (n = 15)-T. spiralis infection without the administration of bacterial strains; LAB (n = 15)- L. reuteri +T. spiralis. Probiotic strains were administered per os daily at a dose of 109 CFU/mL in a total volume of 200 μL. Mice were infected per os with 400 T. spiralis larvae/mouse.
Five mice from each experimental group were euthanized on day 1, 4, 7 after infection, so as to collect the intestinal samples. During sampling procedure (Figure 1), make sure that samples shall not be contaminated. Additionally, Control group was euthanized on day 1, which the samples collected as the uninfected controls. All samples were preserved at -80 C for later use.
2.4. Intestinal Worm Burdens
The intestinal phase of infection was investigated on day 4, and 7. The small intestine was cut into pieces and incubated in 37 °C NaCl (0.9 % saline) overnight. The worms in the sediment were counted after incubation.
2.5. Oxidative Stress Assay
To investigate the oxidative stress of intestinal tissues, the levels of serum superoxide dismutase (SOD) and malondial dehyde (MDA) in the homogenates of intestinal tissues were measured by commercially kits on day 1, 4, and 7, according to the manufacturer's instruction. The level of SOD was measured as an indicator of cellular defense against reactive oxygen species. Also, the level of MDA was measured as an index of lipid peroxides.
2.6. Measurement of Inflammatory Cytokines
Total RNA from each tissue sample was extracted as described [20]. The RNA quantity and purity were assessed. Extracted RNA was diluted to the same concentration and the synthesis of cDNA was performed as the manufacturer’s instructions. The cDNA samples were diluted used for qRT-PCR amplification. Each sample was analyzed in triplicate. For the relative quantification of the gene expression levels, the logarithmic-scaled threshold cycle(Ct) values were used in the 2−∆∆Ct method before calculating the mean and standard error of the mean (SEM) for the references and individual targets.
The encoded gene expression levels of inflammatory response, including interleukin 4 (IL-4), interleukin 10 (IL-10), interleukin 17 (IL-17), and interferon-γ (IFN-γ) expression in the intestinal samples were investigated on day 1, 4, and 7. All oligonucleotide sequences of the forward and reverse primers used in this experiment are obtained as described [21].
2.7. Western Blot Analysis
Proteins were extracted from the intestinal tissues using protein extraction reagent on day 7.Protein concentration was determined by BCA protein assay kit. Proteins were separated by SDS PAGE and blotted onto PVDF membranes .After blockade for 2h at room temperature, the membranes were probed with primary antibodies (Nrf2, HO-1, and β-actin) at 4°C overnight. Followed by incubation with secondary antibody(peroxidase conjugated goat anti-rabbit IgG)with incubation for 2h at room temperature, proteins were visualized with ECL-chemiluminescent kit.
2.8. Statistical Analysis
Statistical differences were assessed using one-way ANOVA, followed by post hoc Tukey’s test (a value of P<0.05 was considered significant), which allowed comparison between each two groups at each time point.
3. Results
3.1. Adult Worm Reduction
As shown in Table 1, a significant reduction of adult worm in the intestine occurred in LAB group (18.24%) and treated with L. reuteri 50319 on day 4. The highest numbers of adult worm were detected in Ts group on days 7. The probiotic treatment resulted in a significant adult worm count reduction in LAB group (44.49%). Experiment results confirmed that L. reuteri promote AD excretion and reduce colonization to against T. spirali infection.
3.2. Oxidative Stress Marker Levels
As shown in Figure 2, after T. spiralis infection day 1, the level of MDA in Ts group was significantly increased than that of the Control group (P<0.01), as well as day 4 and day 7. While, the level of MDA in LAB group was significantly increased than that of the Control group (P<0.01) on day 1 and day 4 (P<0.05).
The level of SOD in Ts group and LAB group were significantly increased than that of the Control group (P<0.01), as well as day 4. While, on day 7 the level of MDA in Ts group and LAB group were significantly increased than that of the Control group (P<0.05).
3.3. Inflammatory Response
Statistical significances were found between Control group and experimental groups (Ts group and LAB group) throughout the entire process, as shown in Figure 3. After infection with T. spiralis, the IL-4 expression levels in Control group had statistically significant values for Ts group and LAB group, and LAB group were higher than Ts group. In the same instant, significantly increasing was obverved in the IL-10 expression levels between Ts group and LAB group,compared with Control group. IL-17 expression results showed a similar tendency at experimental period. On the contrary, compared with Control group, IFN-γ expression significantly increased in Ts group and LAB group. The LAB group gradually recovered to the IFN-γ expression level without no treatment, while Ts group higher than in LAB group consistently .The experiment results confirmed that L. reuteri could inhibit the inflammatory reaction caused by invading intestinal in mice.
3.4. Nrf2 and HO-1 Expression
The expression of Nrf2 and HO-1 in intestinal tissues was detected by western blot analysis, as shown in Figure 4. Compared with the control group, the mice Nrf2 and HO-1 expression level in Ts group and LAB group increases in intestinal tissue.
4. Discussion
T. spiralis is a kind of the typical tissue-dwelling pathogen, which develops in a life cycle including muscle larva (ML), adult worm (Ad) and the newborn larva (NBL) [22]. NBL can be released into the mucosa at day 4 after infection [23,24], which represents the intestine phase of T. spiralis infection [25]. Significantly, T. spiralis is a kind of intestinal parasite that modulate the host immune system strongly.The crucial drugs currently used to control helminth infections including benzimidazoles, imidazothiazoles and macrocyclic lactones families, however, evidences have revealed increasing anthelmintic resistance in animals which a potential health problem. To deal with limitation of anthelmintic drugs, alternative treatments based on probiotics have been developed. Probiotics consumed in adequate amounts, confer benefit to the host [26]. It has been proven that probiotic microbes have the ability to exert an immunomodulatory effect in vivo [27]. As is known to all, the oral administration of probiotics contributes to controlling T. spiralis infections since it has been demonstrated that probiotic could stimulate intestinal immune response, among other effects on the host immune system. The present paper aimed to explore the protection of probiotics in intestinal phase during T. spiralis infection, as well as its potential effects on parasite survival or its underlying pathogenesis. To investigate the protection of probiotics against T. spiralis infection on intestinal, mice were infected with T. spiralis infectious larvae or probiotic by oral gavage. Previous studies demonstrate that T. spiralis produce diverse immunomodulatory effects at different stages. Therefore, samples were also collected at different time points of infection, so as to examine the effects of intestinal stages of T. spiralis infection.
As previously describe, probiotic reduced the parasite burden in experimental trichinellosis.It has been reported that the reduced parasite burden is associated with decreased fecundity induced by probiotic strains. In present paper, L. reuteri reduced adult worms after T. spiralis infection, in agreement with prior observation that LAB administered to induce resistance against T. spiralis infection in mice [28].
Host immune dependent damage to helminth parasites is mediated by the generation of oxygen-derived free radicals via nonspecific defense reaction [29]. Therefore, administered L. reuteri after T. spiralis induction, significant increases in the levels of the antioxidant enzyme SOD starting from day 1, lasting through day 7 were observed. However, significant decreases in the level of antioxidant enzyme MDA. This may be caused by LAB-mediated antioxidant protective effects. The antioxidant enzyme levels in our experimental model are consistent with the results of previous study [30].
In general terms, the immune response in helminth infections is a type 2 immune response which is characterized by the production of interleukin including IL-4, IL-5, IL-9, IL-10, and IL-13 et al [31]. Previous study has been reported that IFN-γ activated macrophages which rapidly processed T. spiralis antigens improving the acquired immune response against the parasite. These macrophages also produced nitric oxide (NO) and probably promoted inflammatory response in the intestine [32]. IL-17 cytokine plays an important role in the pathogenesis of various autoimmune inflammatory diseases [33]. Inflammatory response is a protective mechanism to protect against injury, infection, trauma, and noxious stimuli [34]. The purpose of this process is to eliminate injurious agents or intruder [35]. Mounting evidence suggests that LAB ameliorates inflammation during intestinal injury [36,37]. Therefore, our observation in agreement with the previous literature that LAB alleviate the inflammatory response caused by T. spiralis infection in intestinal.
Nrf2 is essential for the regulation of detoxifying and antioxidant genes [38]. It can activate the Nrf2 signaling pathway to regulate the expression of antioxidant enzymes, alleviate oxidative stress, and alleviate inflammatory reactions. To investigate the anti-oxidative effects of L. reuteri , Nrf2 signaling pathway was measured. In this study, our results showed that L. reuteri dependently up regulated the expression of Nrf2 and HO-1. These results suggested that L. reuteri inhibited T. spiralis induced oxidative stress by activating Nrf2 signaling pathway.
The use of probiotics as a treatment for helminth infection is an incipient research line with promising perspectives. Nonetheless, several limitations should be noted in our study. The sample size is small relatively, which may affect the statistical power. A larger sample size should be expanded in prospective studies.
5. Conclusions
Therapeutic approaches with probiotic strains benefit to reduce the risks of trichinellosis or complement classical anti-parasite treatments. Present study demonstrates that probiotic bacteria can provide strain-specific protection against T. spiralis infection throughout reduced parasite burder, regulated intestinal flora and alleviated inflammatory response. Several additional mechanisms involved in the anti-parasite defense should be further studied and elucidated to justify the therapeutic use of probiotics. We look forward to more findings as a wide range of possibilities to be explored that may deliver groundbreaking treatment strategies for helminth diseases. In addition, to the best of our knowledge, this is the first experimental report on the antiparasitic effects of L. reuteri used as a novel treatment option against nematode T. spiralis infection.
Author Contributions
Conceptualization and methodology S.X. and Y.X.L.; Conduction of the experimental work and data curation, S.X.; writing—review and editing, S.X. and Y.X.L. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The animal study protocol was approved by the Animal Management Committee of Northeast Agricultural University (Animal Ethics Committee approval number SYXK [Hei] 2016-007).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Hasby Saad, M.; Safwat, O.; El-Guindy, D.; Raafat, R.; Elgendy, D.; Hasby, E. Biomolecular Changes and Cortical Neurodegenerative Lesions in Trichinella Spiralis Infected BALB/c Mice: A Preliminary Study Elucidating a Potential Relationship Between Systemic Helminthic Infections and Idiopathic Parkinson's. Helminthologia 2018, 55, 261–274. [Google Scholar] [CrossRef]
- Liu, S.; Pan, J.; Meng, X.; Zhu, J.; Zhou, J.; Zhu, X. Trichinella spiralis infection decreases the diversity of the intestinal flora in the infected mouse. J Microbiol Immunol Infect 2021, 54, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, E.; Sjölund, M.; Wallgren, T.; Lind, E.O.; Höglund, J.; Wallgren, P. Management practices related to the control of gastrointestinal parasites on Swedish pig farms. Porcine Health Manag 2021, 7, 12. [Google Scholar] [CrossRef]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Raisová Stuchlíková, L.; Matoušková, P. Anthelmintics in the future: current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov Today 2020, 25, 430–437. [Google Scholar] [CrossRef]
- Vargová, M.; Hurníková, Z.; Revajová, V.; Lauková, A.; Dvorožňáková, E. Probiotic Bacteria can Modulate Murine Macrophage's Superoxide Production in Trichinella Spiralis Infection. Helminthologia 2020, 57, 226–234. [Google Scholar] [CrossRef]
- Bautista-Garfias, C.R.; Ixta, O.; Orduña, M.; Martínez, F.; Aguilar, B.; Cortés, A. Enhancement of resistance in mice treated with Lactobacillus casei: effect on Trichinella spiralis infection. Vet Parasitol 1999, 80, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Gómez, F.; Fuentes-Castro, B.E.; Bautista-Garfias, C.R. The intraperitoneal inoculation of Lactobacillus casei in mice induces total protection against Trichinella spiralis infection at low challenge doses. Parasitol Res 2011, 109, 1609–1617. [Google Scholar] [CrossRef] [PubMed]
- El Temsahy, M.M.; Ibrahim, I.R.; Mossallam, S.F.; Mahrous, H.; Abdel Bary, A.; Abdel Salam, S.A. Evaluation of newly isolated probiotics in the protection against experimental intestinal trichinellosis. Vet Parasitol 2015, 214, 303–314. [Google Scholar] [CrossRef]
- Dvorožňáková, E.; Bucková, B.; Hurníková, Z.; Revajová, V.; Lauková, A. Effect of probiotic bacteria on phagocytosis and respiratory burst activity of blood polymorphonuclear leukocytes (PMNL) in mice infected with Trichinella spiralis. Vet Parasitol 2016, 231, 69–76. [Google Scholar] [CrossRef]
- Bucková, B.; Hurníková, Z.; Lauková, A.; Revajová, V.; Dvorožňáková, E. The Anti-parasitic Effect of Probiotic Bacteria via Limiting the Fecundity of Trichinella Spiralis Female Adults. Helminthologia 2018, 55, 102–111. [Google Scholar] [CrossRef]
- Travers, M.A.; Florent, I.; Kohl, L.; Grellier, P. Probiotics for the control of parasites: an overview. J Parasitol Res 2011, 2011, 610769. [Google Scholar] [CrossRef] [PubMed]
- Harris, N.L. Intimate gut interactions: helminths and the microbiota. Cell Res 2016, 26, 861–862. [Google Scholar] [CrossRef] [PubMed]
- Gause, W.C.; Maizels, R.M. Macrobiota - helminths as active participants and partners of the microbiota in host intestinal homeostasis. Curr Opin Microbiol 2016, 32, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Peachey, L.E.; Jenkins, T.P.; Cantacessi, C. This Gut Ain't Big Enough for Both of Us. Or Is It? Helminth-Microbiota Interactions in Veterinary Species. Trends Parasitol 2017, 33, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Su, L.; Li, Y.; Long, S.R.; Chang, J.; Zhang, W.; Walker, W.A.; Xavier, R.J.; Cherayil, B.J.; Shi, H.N. Helminth-induced alterations of the gut microbiota exacerbate bacterial colitis. Mucosal Immunol 2018, 11, 144–157. [Google Scholar] [CrossRef] [PubMed]
- Walter, J. Ecological role of lactobacilli in the gastrointestinal tract: implications for fundamental and biomedical research. Appl Environ Microbiol 2008, 74, 4985–4996. [Google Scholar] [CrossRef] [PubMed]
- Rheinallt, M.J.; Chirayu, D.; Trevor, M.D.; Liping, L.; Alexandra, A.W.; Christopher, D.S.; Courtney, S. A; April. R.R; Erin. S.K.; Andrew, S.N. Lactobacilli Modulate Epithelial Cytoprotection through the Nrf2 Pathway[J]. Cell Reports, 2015; 12, 1217–1225. [Google Scholar] [CrossRef]
- Bolas-Fernández, F. Biological variation in Trichinella species and genotypes. J Helminthol 2003, 77, 111–118. [Google Scholar] [CrossRef]
- Gamble, H.R.; Bessonov, A.S.; Cuperlovic, K.; Gajadhar, A.A.; van Knapen, F.; Noeckler, K.; Schenone, H.; Zhu, X. International Commission on Trichinellosis: recommendations on methods for the control of Trichinella in domestic and wild animals intended for human consumption. Vet Parasitol 2000, 93, 393–408. [Google Scholar] [CrossRef] [PubMed]
- Park, I.; Lee, Y.; Goo, D.; Zimmerman, N.P.; Smith, A.H.; Rehberger, T.; Lillehoj, H.S. The effects of dietary Bacillus subtilis supplementation, as an alternative to antibiotics, on growth performance, intestinal immunity, and epithelial barrier integrity in broiler chickens infected with Eimeria maxima. Poult Sci 2020, 99, 725–733. [Google Scholar] [CrossRef]
- Xie, Z.; Zhang, G.; Liu, R.; Wang, Y.; Tsapieva, A.N.; Zhang, L.; Han, J. Heat-Killed Lacticaseibacillus paracasei Repairs Lipopolysaccharide-Induced Intestinal Epithelial Barrier Damage via MLCK/MLC Pathway Activation. Nutrients 2023, 15. [Google Scholar] [CrossRef]
- Pozio, E. Searching for Trichinella: not all pigs are created equal. Trends Parasitol 2014, 30, 4–11. [Google Scholar] [CrossRef]
- Song, Y.Y.; Zhang, Y.; Ren, H.N.; Sun, G.G.; Qi, X.; Yang, F.; Jiang, P.; Zhang, X.; Cui, J.; Wang, Z.Q. Characterization of a serine protease inhibitor from Trichinella spiralis and its participation in larval invasion of host's intestinal epithelial cells. Parasit Vectors 2018, 11, 499. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.Y.; Zhu, X.P.; Xu, K.C.; Lu, Q.; Boireau, P. Biological and genetic characteristics of two Trichinella isolates in China; comparison with European species. Parasite 2001, 8, S34–38. [Google Scholar] [CrossRef]
- Wakelin, D.; Goyal, P.K. Trichinella isolates: parasite variability and host responses. Int J Parasitol 1996, 26, 471–481. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. , et al. Expert consensus document. The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat Rev Gastroenterol Hepatol 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Saracino, M.P.; Vila, C.C.; Baldi, P.C.; González Maglio, D.H. Searching for the one(s): Using Probiotics as Anthelmintic Treatments. Front Pharmacol 2021, 12, 714198. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.T.; Cheng, P.C.; Pan, T.M. The immunomodulatory effects of lactic acid bacteria for improving immune functions and benefits. Appl Microbiol Biotechnol 2012, 96, 853–862. [Google Scholar] [CrossRef] [PubMed]
- Mido, S.; Fath, E.M.; Farid, A.S.; Nonaka, N.; Oku, Y.; Horii, Y. Trichinella spiralis : Infection changes serum paraoxonase -1 levels, lipid profile, and oxidative status in rats[J]. Experimental Parasitology 2012, 131, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Farid, A.S.; Mido, S.; Linh, B. K.; Hayashi, T.; Horii, Y. An atherogenic lipid profile with low serum paraoxonase-1 activity during nematode infection in rats[J]. European Journal of Clinical Investigation, 2010; 40, 984–993. [Google Scholar] [CrossRef]
- Allen, J.E.; Maizels, R.M. Diversity and dialogue in immunity to helminths. Nat Rev Immunol 2011, 11, 375–388. [Google Scholar] [CrossRef]
- Bautista-Garfias, C.R.; Ixta-Rodríguez, O.; Martínez-Gómez, F.; López, M.G.; Aguilar-Figueroa, B.R. Effect of viable or dead Lactobacillus casei organisms administered orally to mice on resistance against Trichinella spiralis infection. Parasite 2001, 8, S226–S228. [Google Scholar] [CrossRef]
- Rudner, X.L.; Happel, K.I.; Young, E.A.; Shellito, J.E. Interleukin-23 (IL-23)-IL-17 cytokine axis in murine Pneumocystis carinii infection. Infect Immun 2007, 75, 3055–3061. [Google Scholar] [CrossRef] [PubMed]
- Choy, C.S.; Hu, C.M.; Chiu, W.T.; Lam, C.S.; Ting, Y.; Tsai, S.H.; Wang, T.C. Suppression of lipopolysaccharide-induced of inducible nitric oxide synthase and cyclooxygenase-2 by Sanguis Draconis, a dragon's blood resin, in RAW 264.7 cells. J Ethnopharmacol 2008, 115, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Kanmani, P.; Kim, H. Protective Effects of Lactic Acid Bacteria Against TLR4 Induced Inflammatory Response in Hepatoma HepG2 Cells Through Modulation of Toll-Like Receptor Negative Regulators of Mitogen-Activated Protein Kinase and NF-κB Signaling. Front Immunol 2018, 9, 1537. [Google Scholar] [CrossRef] [PubMed]
- Roselli, M.; Finamore, A.; Britti, M.S.; Mengheri, E. Probiotic bacteria Bifidobacterium animalis MB5 and Lactobacillus rhamnosus GG protect intestinal Caco-2 cells from the inflammation-associated response induced by enterotoxigenic Escherichia coli K88. Br J Nutr 2006, 95, 1177–1184. [Google Scholar] [CrossRef]
- Wang, Y.; Kirpich, I.; Liu, Y.; Ma, Z.; Barve, S.; McClain, C.J.; Feng, W. Lactobacillus rhamnosus GG treatment potentiates intestinal hypoxia-inducible factor, promotes intestinal integrity and ameliorates alcohol-induced liver injury. Am J Pathol 2011, 179, 2866–2875. [Google Scholar] [CrossRef]
- Fan, L.; Yunyi, Y.; Hui, H.; Hua, H.; Mingzhong, Y. Xanthohumol attenuates cisplatin-induced nephrotoxicity through inhibiting NF-κB and activating Nrf2 signaling pathways[J]. International Immunopharmacology, 2018; 61, 277–282. [Google Scholar] [CrossRef]
Figure 1.
Schematic outline of the experimental design.
Figure 2.
Oxidative stress markers detection results. *P<0.05 and **P<0.01 mean statistically significant differences from the Control group.
Figure 2.
Oxidative stress markers detection results. *P<0.05 and **P<0.01 mean statistically significant differences from the Control group.
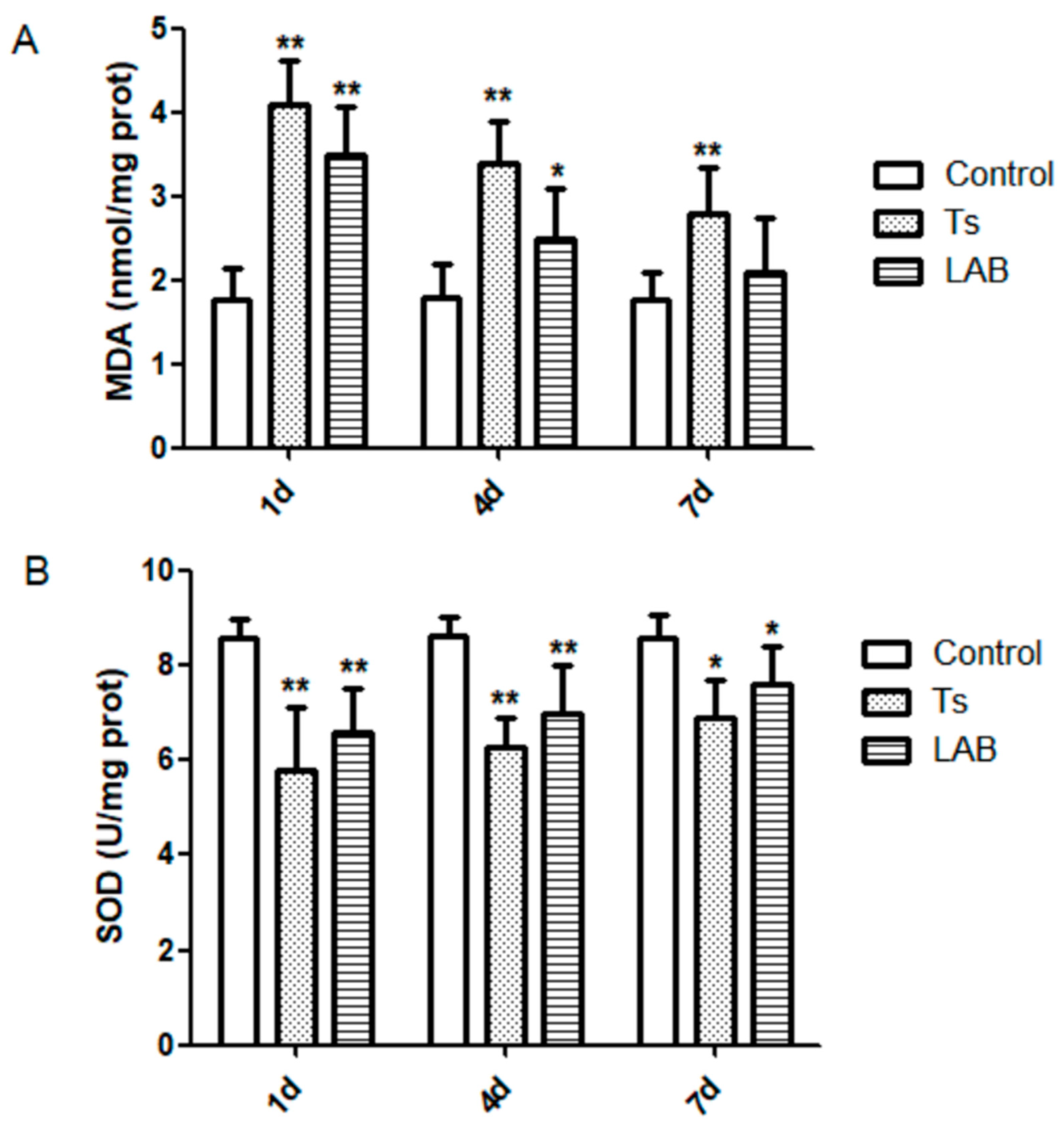
Figure 3.
Inflammatory factor expression results. *P<0.05 and **P<0.01 mean statistically significant differences from the Control group.
Figure 3.
Inflammatory factor expression results. *P<0.05 and **P<0.01 mean statistically significant differences from the Control group.
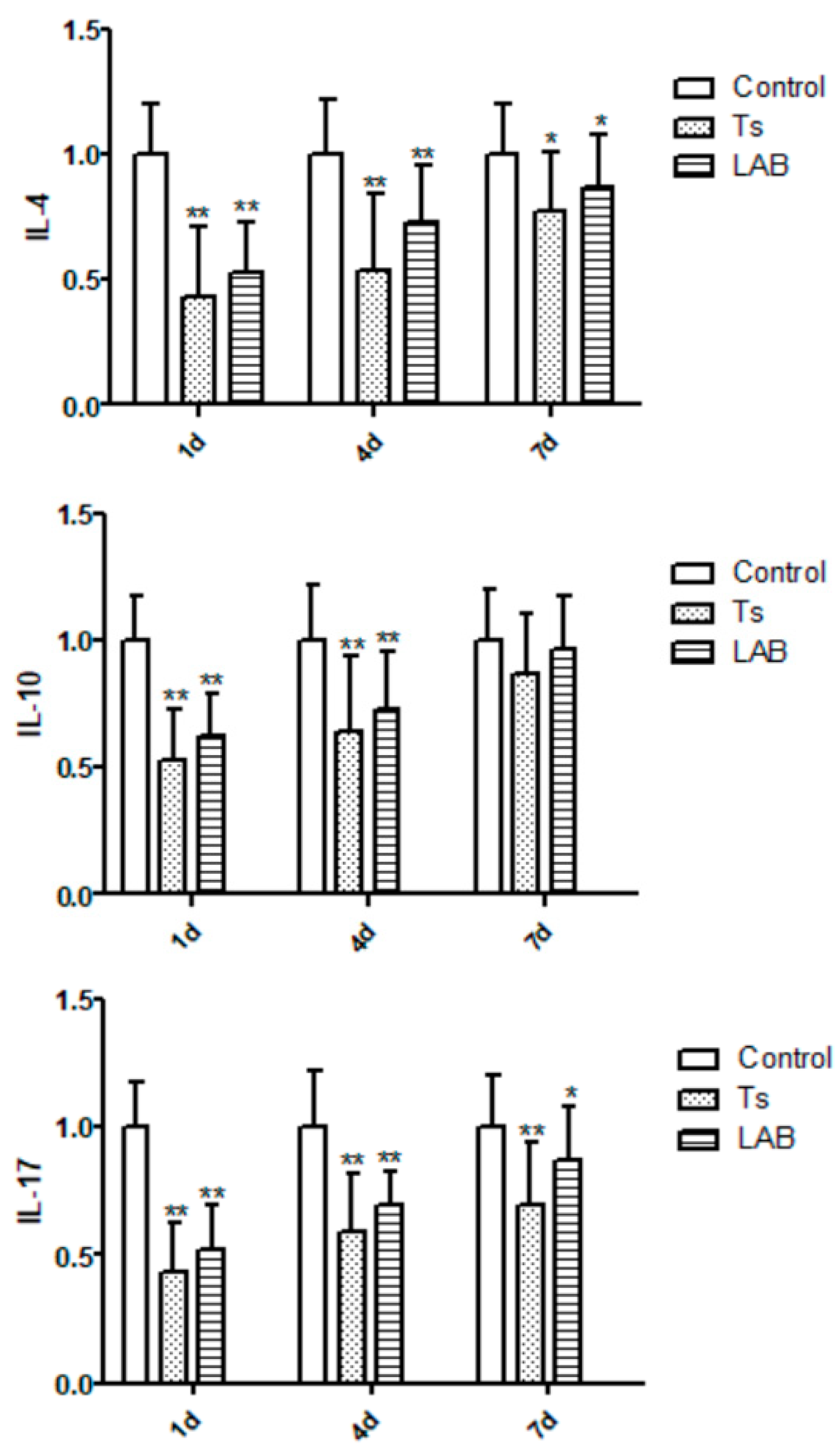
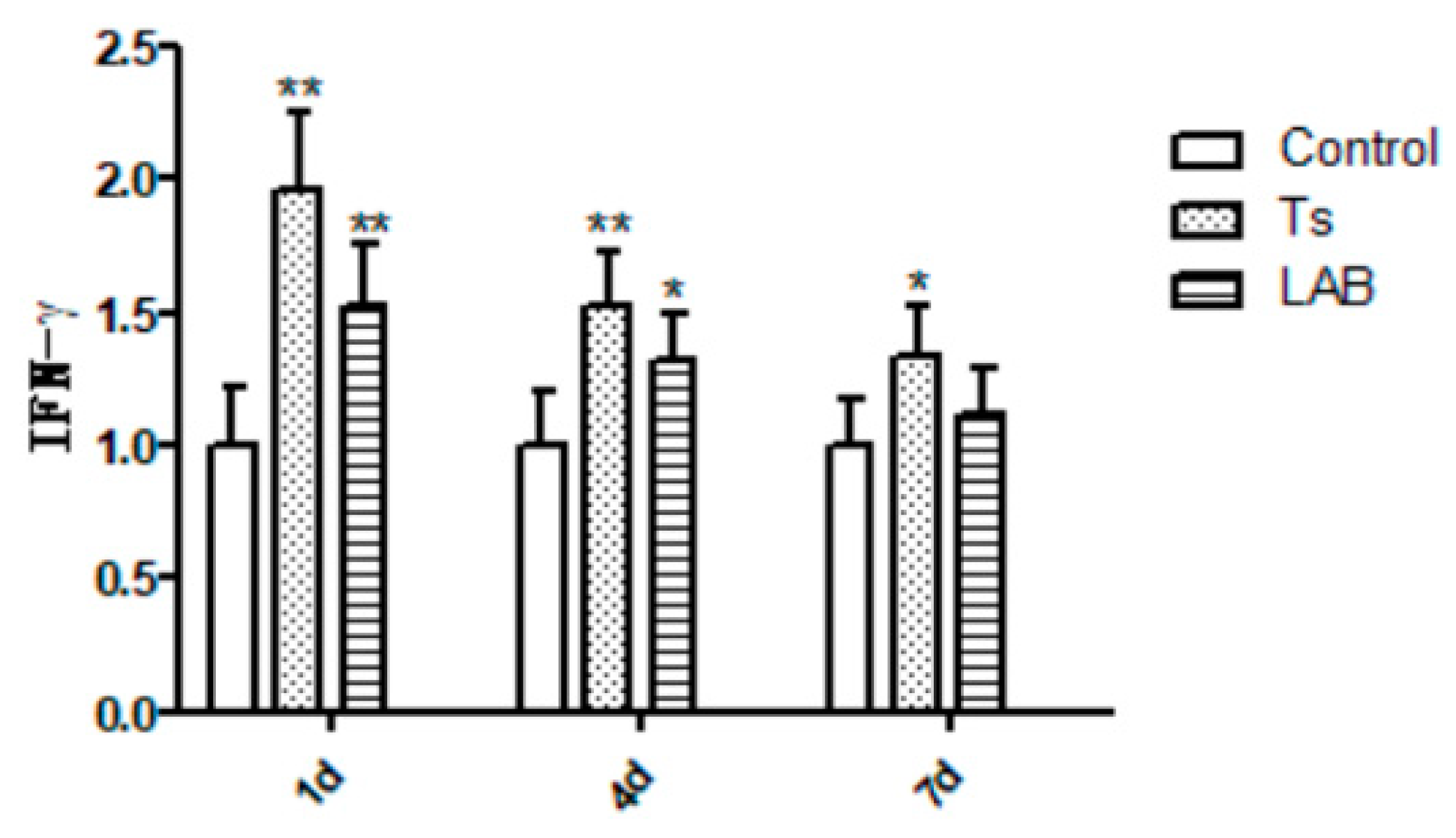
Figure 4.
Nrf2 and HO-1 protein expression results. *P<0.05 means statistically significant differences from the Control group.
Figure 4.
Nrf2 and HO-1 protein expression results. *P<0.05 means statistically significant differences from the Control group.
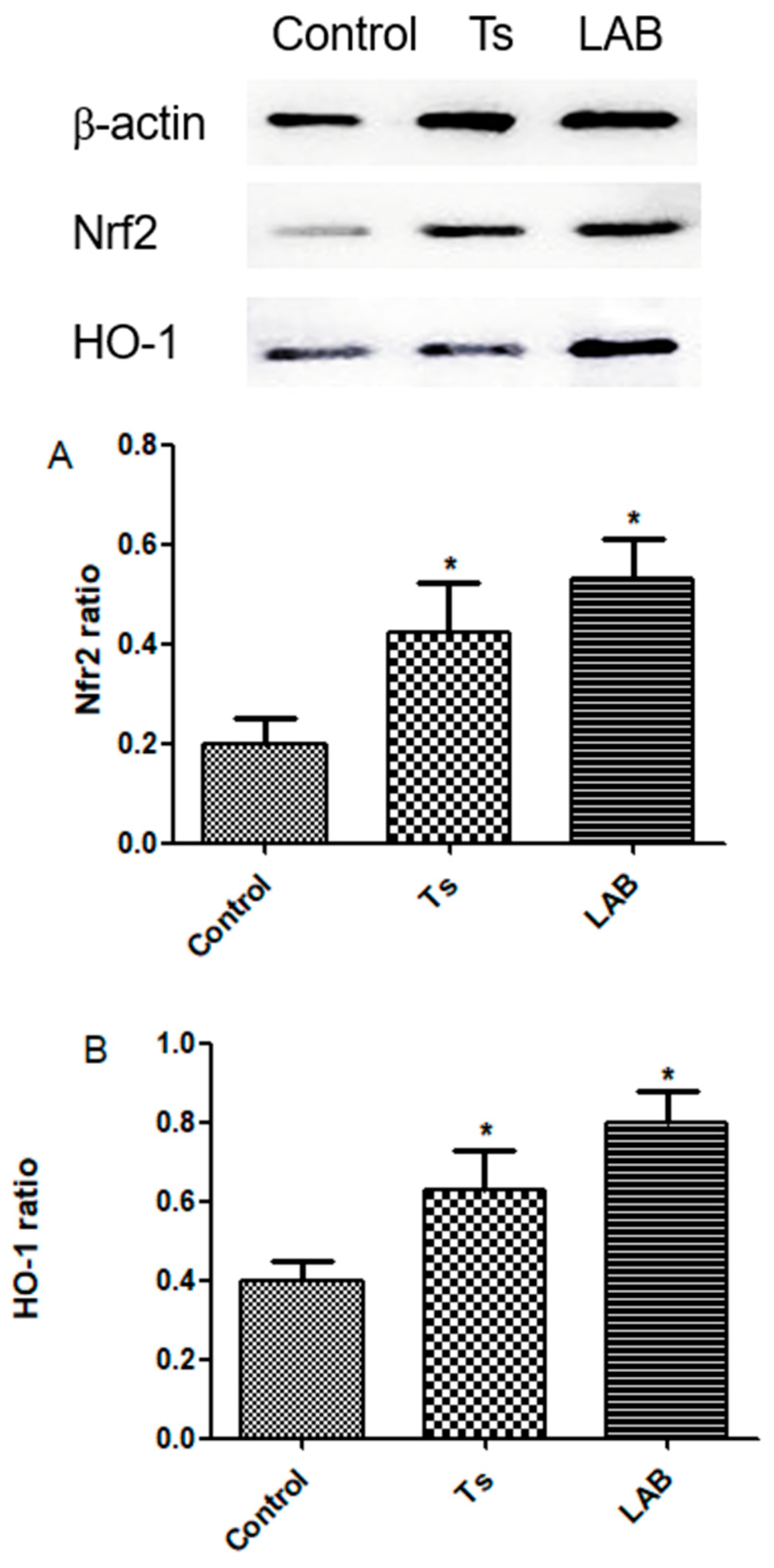

Table 1.
Parasite burden in mice treated with probiotic bacteria and infected with T. spiralis.
| Day 4 | Day 7 | |||
|---|---|---|---|---|
| mean±SD | reduction | mean±SD | reduction | |
| Ts | 296±31 | —— | 227±15 | —— |
| LAB | 242±26 | 18.24% | 126±23* | 44.49% |
*P<0.05 means statistically significant differences from T. spiralis infected group.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



