
Submitted:
07 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Carpels are a reproductive feature restricted to angiosperms, therefore they are a focus of many botanical studies. However, there are controversies over the nature of carpels. A reason underly-ing these controversies is mixing implications given by conflicting interpretations on fossil carpels in early fossil angiosperms from the Cretaceous. These controversies hinder a clear understanding of angiosperm evolution and systematics. A key to these questions is older fossil fruits bearing concerned information. Here we report a new fossil fruit, Xenofructus dabuensis gen. et sp. nov, from the Middle Jurassic of Liaoning, China. Unlike previously reported fruits of early angio-sperms that were interpreted as bearing seeds either on adaxial or abaxial margin by various au-thors, our fossil demonstrates clearly that the seeds in Xenofructus are neither borne on the adaxial nor abaxial margin of the fruit, instead the seeds of Xenofructus are borne on an axis positioned between two margins. This new feature implies that a placenta in carpels is an ovule/seed-bearing axis, a carpel is a composite organ comprising an enclosing leaf (fruit wall) and an axis (placenta). The adaxial or abaxial position of ovules/seeds frequently seen in fossil and extant angiosperms is a consequence derived through long time evolution (coalescence of placenta with either margin of fruits). Carpels can be taken as foliar structures enclosing their associated ovulate branches.
Keywords:
Xenofructus
; fruits
; carpels
; evolution
; angiosperms
; Jurassic
; China
1. Introduction
Angiosperms are the most diversified plant group that is closely related with the well-being of the humans. Carpels are a feature idiosyncratic to angiosperms, and thus a focus of studies and debates. Formerly, a carpel was taken as an equivalent of a leaf [1]. However, this ancient saying dated back to the Goethe age has never been honored by any fossil evidence [2] despite palaeobotanists have been painstakingly searching for related evidence. A recently raised hypothesis stated that a carpel is a composite organ derived from an ovule-bearing axis and a subtending and enclosing leaf [3,4,5]. The latest progresses include 1) recognizing the branch-terminal position of seeds in gymnosperms [6,7], 2) treating ovule and carpel (wall) separately, which become tightly synorganized, in extant angiosperms [8]. Although these moves are in the right direction, the controversy over nature of carpels continues, partially due to on-going debates on ovule/seed position in fruits of early angiosperms, including famous Archaefructus [9,10,11,12] and Archaeanthus [13,14] (Figure 3). In these debates, the seeds were interpreted as borne either on the adaxial or the abaxial margin (two opposite positions) of the fruits, and neither of the confronting schools could convince the other. How to reconcile these conflicting parties? The stalemate between these two conflicting parties cannot be broken until independent informative fossil occurs. Here we report a new fruit, Xenofructus dabuensis gen. et sp. nov, from the Middle Jurassic of Liaoning, China. It appears that this new fossil provides a new insight on nature of carpels.
2. Materials and Methods
The specimen was collected from an outcrop of the upper member of the Dabu Formation in the Tianshifu Coal Mine (N41˚14′58″, E124˚23′03″) in 1978 by Dr. Shaolin Zheng and Dr. Wu Zhang. The specimen bearing the fossil plants is dark-colored siltstone, lacking good lamination. There are two similar fruits preserved on the surface of the same block. Previously reported fossil plants from the upper member of the Dabu Formation include Equisetum beani, E. laterale, Neocalamites hoerensis, Todites williamsoni, Dicksonia concinna, Eboracia lobifolia, Clathropteris pekingensis, Cladophlebis asiatica, C. sp., Coniopteris simplex, Pterophyllum propinquum, P. angustum, P. sp., Ctenis kaneharai, Williamsoniella buracovae, W. minima, W. dabuensis, Nilssonia tenuinervis, Ginkgo concinna, G. sp., Czekanowskia sp., Baiera gracilis, B. furcata, Sphenobaiera gyron, Phoenicopsis augustifolia, Sphenarion latifolia, “Elatidies” chinensis, Storgaardia mentoukouensis, Lindleycladus lanceolatus, Carpolithus sp., Raticites sp.. Other than these taxa, the fossil plants from the Dabu Formation include Lycopodites falcatus, Neocalamites carrerei, Annulariopsis sp., A. sp., Marattiopsis hoerensis, M. muensteri, Coniopteris hymenophylloides, C. kuandianensis, C. karatiubensis, Kuandiania crassicaulis, Clathropteris meniscioides, Hausmannia cf. dentata, Todites denticulata, Dicksonia kendalli, Cladophlebis raciborskii, C. argutula, C. shansiensis, C. fukiensis, C. haiburnensis, C. nebbensis, Pterophyllum issykkulense, Nilsonia mosserai, N. aff. acuminata, N. cf. compta, N. orientalis, N. mosserayi, Anomozamites cf. major, A. inconstans, A. spp., Pterophyllum aequale, P. ctenoites, P. nathorsti, Ctenis lingyuanensis, Pityophyllum nordenskioldi, P. longifolium, Tyrmia grandifolia, Baiera asadai, B. cf. lindleyana, Ginkgo beijingensis, G. obrutschewi, G. manchurica, G. sibirica, Sphenobaiera huangi, Czekanowskia rigida, Phoenicopsis aff. speciosa, Elatidies ovalis, Elatocladus (Cephalotaxopsis?) angustifolius, E. lindongensis, Ferganiella cf. podozamioides, Solenites vimineus, Leptostrobus laxifolia, Isostrobus heeri [15,16]. The fossil flora is comparable to the Coniopteris-Phoenicopsis flora, and the fossiliferous strata are thought belonging to the early Middle Jurassic as it shares 17 species with the typical Middle Jurassic Yorkshire Flora [15,16].
The whole specimen was photographed using a Huawei digital camera, and details of the fossil were photographed using a Nikon SMZ1500 stereomicroscope at the Nanjing Institute of Geology and Palaeontology, Nanjing, China. The specimen was investigated on a Phoenix v|tome|x m scanner using a voltage of 190 Kv and a current of 140 μA at the State Key Laboratory of Continental Dynamics (Northwest University), Xi’an, Shaanxi Province, China. The specimen was mounted on the stage using self-adhesive tape. In total, 2000 projections were obtained. The dataset had a resolution of 74.56 μm. Three-dimensional reconstructions were generated using VG Studio 3.2. All images were saved in TIFF or JPEG format. The brightness and contrast of images were adjusted in whole. All figures were organized for publishing using a Photoshop 7.0.
3. Results
Angiosperms
Xenofructusgen. nov
Synonym: Williamsoniella Zheng and Zhang 1990
Type species: Xenofructus dabuensis gen. et sp. nov
Etymology: Xeno– for unexpected morphology and occurrence of the fossil, –fructus Latin word for fruit.
Generic diagnosis: Fruit with a short stalk, straight on abaxial, convex on adaxial. Abaxial ridge robust. Placenta positioned between abaxial and adaxial margins of fruit. Seeds enclosed in fruit, in cluster, sickle-shaped.
Remarks:
The specimens studied here were previously identified as Williamsoniella dabuensis Zheng and Zhang in 1990 [15], which was placed in Bennettitales. Indeed, the general morphology did resemble what shown in Text-Figure 3 of Harris [17]. However, the features of our specimen appear to fall out of the morphological scope of Bennettitales, which are expected to have conic receptacle bearing numerous seeds and interseminal scales or pollen sacs on the adaxial surface of microsporophylls [17,18]. Forms and variations of seeds in our specimen suggest that they are not pollen sacs, as suggested by Zheng and Zhang (1990), which are expected to be more uniform in morphology. Therefore, we re-studied the specimen and revealed some new features.
The fruit of Xenofructus dabuensis gen. et sp. nov is unique in its placenta position (internal in the fruit, positioned between the adaxial and abaxial margins of the fruit) and sickle-shaped seeds borne in cluster on a branch. The combination of both features has never been seen in previously reported in any fossil taxa, thus justifying a new genus.
Xenofructus dabuensis gen. et sp. nov
Synonym: Williamsoniella dabuensis Zheng and Zhang 1990, Plate V, Figure 7A, page 220 [15]
Diagnosis: The same as that of the genus.
Description: The specimens studied here include two isolated fruit parallel oriented on surface of the same block (Figure 1, Figure 2 and Figure S1). Both fruits are preserved as coalified compressions (Figure 1a,b and Figure S1a). The general morphology of these two fruits agrees each other, although there is slight difference in dimension (Figure 1a,b,g). One of the fruit (Fruit A) measures 19 mm long and 8.4 mm wide (Figure 1a), while the other (Fruit B) measures 21 mm long and 6.9 mm wide (Figure 1b). Fruit A has a stalk 4.7 mm long and 2.5 mm wide, gradually transitional to the base of the fruit (Figure 1a and Figure 2a). Fruit B has a stalk 5.6 mm long and 1.8 mm wide, gradually transitional to the base of the fruit (Figure 1b andFigure 2b). Due to difference in breaking, different features are shown separately in these two fruits (Figure 1a–f, Figure 2a,b and Figure S1a–d). The foreground part of Fruit A is almost fully removed, exposing the internal cluster of seeds and robust abaxial ridge (Figure 1a,g,h, Figure 2a and Figure S1a–c), while the foreground part of Fruit B is partially removed, with the clustered seeds (placenta) are partially covered by the fruit wall (Figure 1b,c,e, Figure 2b and Figure S1a,c). The internal position of placenta (cluster of seeds) within the fruit is suggested by the presence of the stubs of the fruit wall (Figure 1b,c,e). The placenta is a central axis bearing several seeds (Figure 1a–c, Figure 2a,b and Figure S1a,c). The seeds are sessile, have variable orientations and morphologies, depending on their positions on the placenta (Figure 1a–c,f, Figure 2a,b and Figure S1a,c,d). The basal seeds tend to be corpulent and round in shape, up to 1.9 mm long and 0.95 mm thick (Figure 1c and Figure S1c,d), while the distal ones tend to more slender and sickle-shaped, up to 2.5 mm long and 1.1 mm thick (Figure 1c,f). There appears to be some trace of micropyle at the distal of a seed, suggestive of orthotropous ovules (Figure 1c,f).
Holotype specimen: KP10-18A.
Etymology: dabuensis–, species epithet conserved from Williamsoniella dabuensis Zheng and Zhang, former name of the specimen.
Type locality: Tianshifu Coal Mine, Liaoning, China.
Stratigraphic horizon: The Dabu Formation (the Middle Jurassic, >164 Ma).
Depository: The Shenyang Center of Geological Survey and Institute of Geology and Mineral Resources, Ministry of National Land and Resources, Shenyang, China.
Figure 1.
Xenofructus dabuensis gen. et sp. nov and its details. Specimen number KP10-18A. a. Holotype, showing profile of Fruit A and its seeds exposed. Scale bar = 1 mm. b. Fruit B and its internal seeds partially exposed and partially covered by the broken fruit wall. Scale bar = 1 mm. c. Detailed view of the black rectangle in Figure 1b, showing three seeds (s) of variable shapes and fruit wall (upper right) covering other seeds. Scale bar = 1 mm. d. Detailed view of the rectangle in Figure 1a, showing robust abaxial margin (between the triangles) of the fruit. Scale bar = 1 mm. e. Broken fruit wall (upper right) and exposed internal of the fruit (lower left), and the boundary in between (triangles), enlarged from the white rectangle in Figure 1b. Scale bar = 0.5 mm. f. A sickle-shaped seed with a distal invagination (possible micropyle, triangle), enlarged from the rectangle in Figure 1c. Scale bar = 1 mm. g. A Micro-CT rendering of the specimen showing two fruits parallel deployed on the surface of the specimen. h-j. Serial sections of Fruit B, showing the seed-bearing placenta (triangles) independent from both margins of the fruit. Scale bar = 2 mm.
Figure 1.
Xenofructus dabuensis gen. et sp. nov and its details. Specimen number KP10-18A. a. Holotype, showing profile of Fruit A and its seeds exposed. Scale bar = 1 mm. b. Fruit B and its internal seeds partially exposed and partially covered by the broken fruit wall. Scale bar = 1 mm. c. Detailed view of the black rectangle in Figure 1b, showing three seeds (s) of variable shapes and fruit wall (upper right) covering other seeds. Scale bar = 1 mm. d. Detailed view of the rectangle in Figure 1a, showing robust abaxial margin (between the triangles) of the fruit. Scale bar = 1 mm. e. Broken fruit wall (upper right) and exposed internal of the fruit (lower left), and the boundary in between (triangles), enlarged from the white rectangle in Figure 1b. Scale bar = 0.5 mm. f. A sickle-shaped seed with a distal invagination (possible micropyle, triangle), enlarged from the rectangle in Figure 1c. Scale bar = 1 mm. g. A Micro-CT rendering of the specimen showing two fruits parallel deployed on the surface of the specimen. h-j. Serial sections of Fruit B, showing the seed-bearing placenta (triangles) independent from both margins of the fruit. Scale bar = 2 mm.
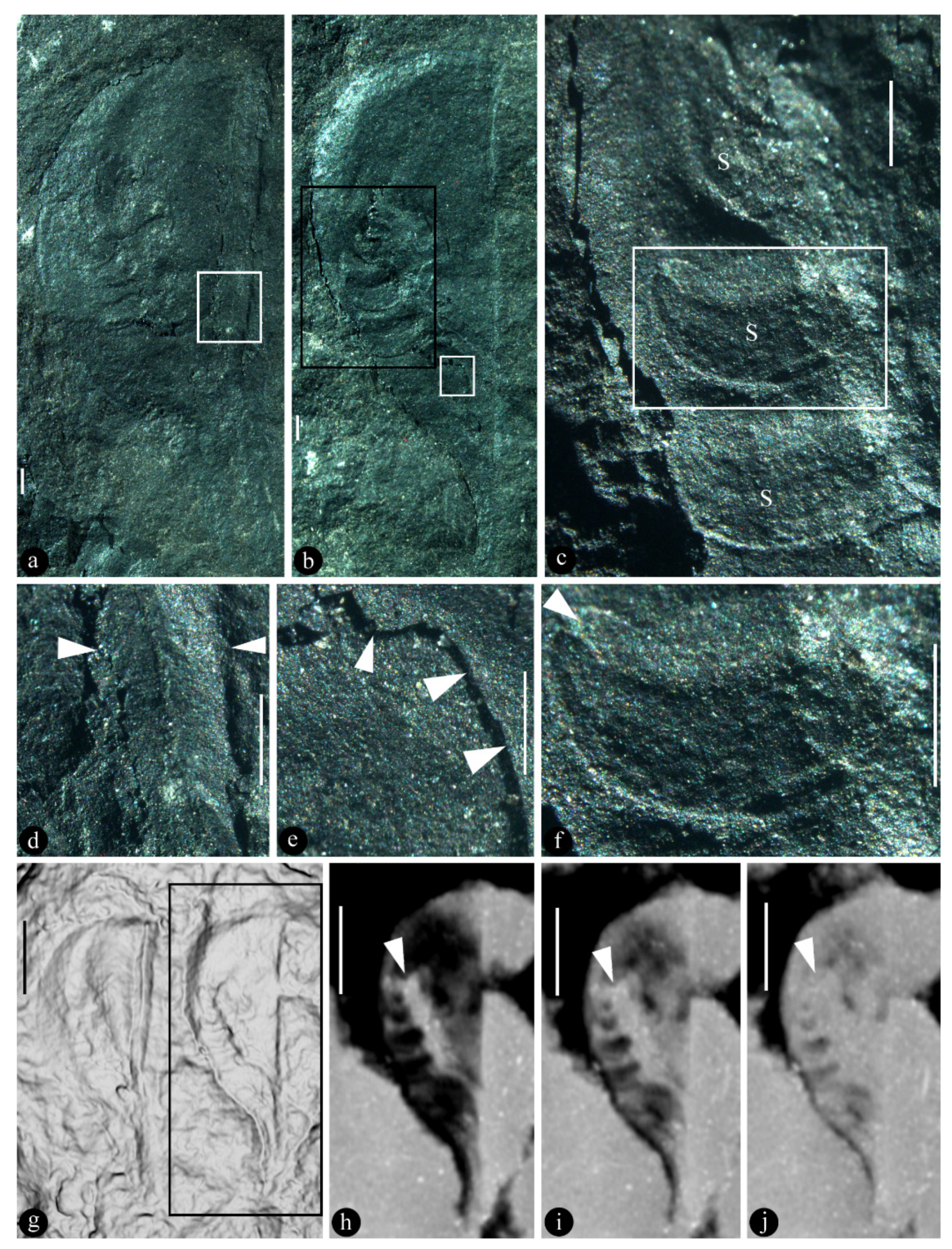
Figure 2.
Sketches of two Xenofructus fruits. Fruit outline is in black, while seed outline is in gray. a. The fruit in Figure 1a,b. The fruit in Figure 1b.
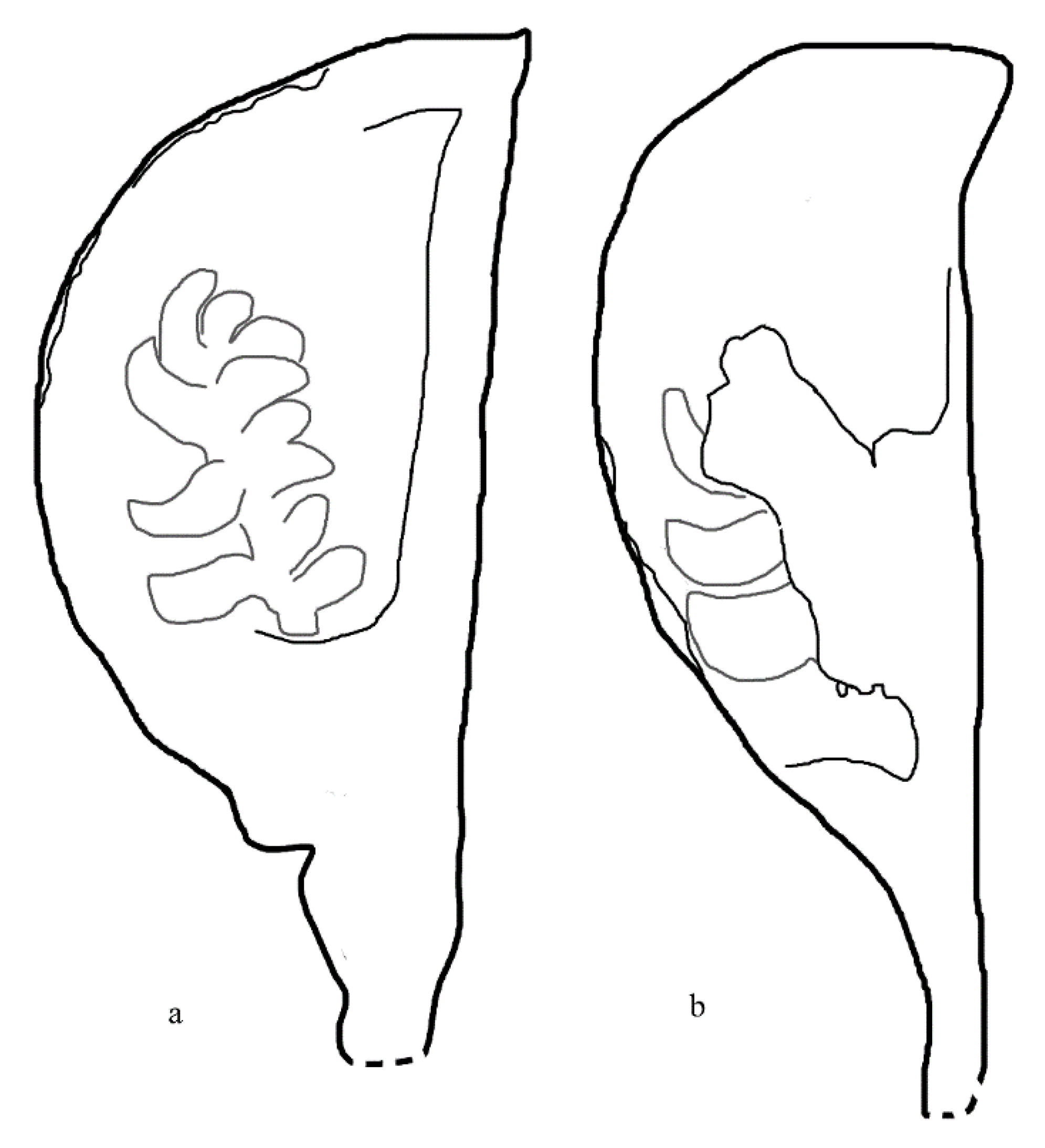
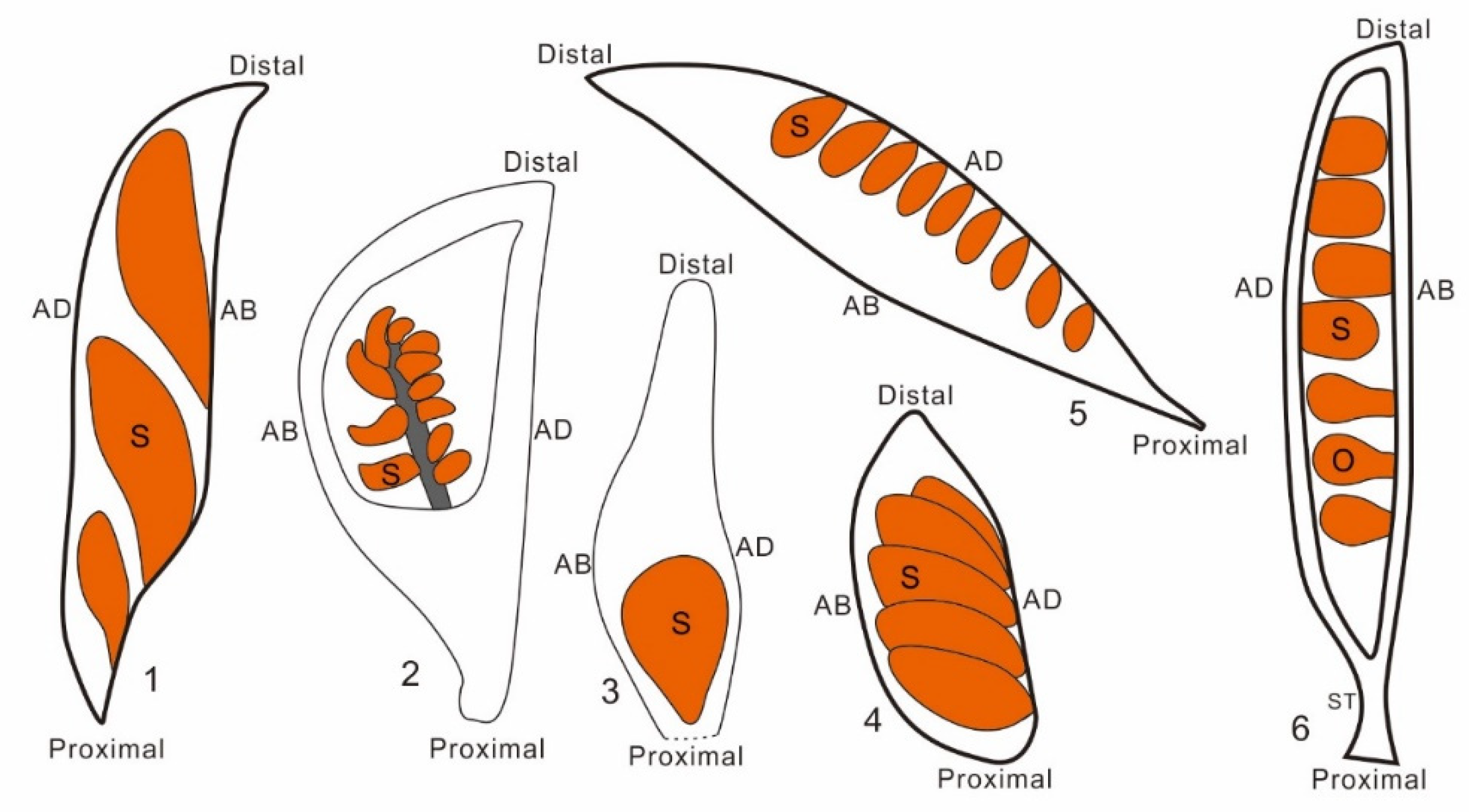
Figure 3.
Several representatives of early angiosperms, showing variable seed arrangement in the fruit. Seed (S)/ovule(O) are inserted either along the adaxial (AD), abaxial (AB), or both margins of a fruit, or is basally fixed in a fruit. 1. Archaefructus, according to Wang and Zheng [12]; 2. Xenofructus; 3. Qingganninginfructus, according to Han et.al. [19]; 4.Gansufructus,according to Du et al. [20]; 5. Sinocarpus, according to Leng and Friis [21]; 6. Archaeanthus,according to Wang [13].
Figure 3.
Several representatives of early angiosperms, showing variable seed arrangement in the fruit. Seed (S)/ovule(O) are inserted either along the adaxial (AD), abaxial (AB), or both margins of a fruit, or is basally fixed in a fruit. 1. Archaefructus, according to Wang and Zheng [12]; 2. Xenofructus; 3. Qingganninginfructus, according to Han et.al. [19]; 4.Gansufructus,according to Du et al. [20]; 5. Sinocarpus, according to Leng and Friis [21]; 6. Archaeanthus,according to Wang [13].
4. Discussion
The term “angiosperms” was coined by German botanist Paul Hermann (1646-1695) in 1690. He combined two Greek words, angeion (vessel) and spermos (seed), into a new word “angiosperm” designating “plant with seeds contained in a protective vessel” (www.etymonline.com). This definition has been accepted widely by many leading botanists [22]. For example, the so-called “First Flower” Archafructus liaoningensis was recognized following this definition. Wang [3,23,24,25,26,27,28,29,30,31,32,33,34,35] has been applying this criterion identifying various early angiosperms. Thus here we adopt this criterion for angiosperms to identify our fossil plant, Xenofructus gen. nov.
As seeds are supposed to be enclosed in angiosperm fruits, so angiosperm seeds are not to be visible to naked eyes unless the enclosing fruit wall is broken. This makes proving enclosed seeds in a fossil plant a great challenge for palaeobotanists, especially when there is only one specimen of fossil fruit available for observation. We are lucky in that there are two fruits preserved side by side in the same specimen, AND these two fruits are preserved in slightly different statuses (Figure 1a,b and Figure S1a). Fruit A in Figure 1a is a broken fruit, namely, the foreground fruit wall is almost fully removed (Figure 1a,g,h and Figure S1c), exposing the seeds formerly enclosed in the fruit, and allows us to view all clustered seeds. These seeds are clustered along an axis, forming an equivalent of placenta. The counterpart of this placenta is only partially seen in Fruit B (Figure 1b,c), as the placenta in this fruit is partially covered and eclipsed by the fruit wall (Figure 1b,c). Comparing these two fruits suggests that the placentae (including the seeds on them) of Xenofructus are sandwiched between to fruit walls and thus fully enclosed in the formerly integral fruits, satisfying the criterion of angiosperms.
The presence of fruit wall in Xenofructus is suggested by the partially preserved placenta in Fruit B. Comparing Figure 1a–c, it is easy to see that there are less seeds in Figure 1b,c than in Figure 1a, and that the deployment of seeds in both fruits differ. These differences are due to the partial removal and partial preservation of fruit wall in Figure 1b,c. The presence of fruit wall is further suggested by Figure 1e, which shows a detailed view of the basal portion of Fruit B in Figure 1b, in which the fallen off fruit wall left a clear stub. Both Figure 1e and Figure 1b,c indicate that the fruit wall covers both basal and upper parts of the fruit. Actually, the presence of fruit wall is obvious on the adaxial and abaxial margins if a careful observation is paid: in Figure 1b,c, the dark shadow of the fruit wall, although thin, on the left is clearly shown for Fruit B. Similar residue of fruit wall or epidermis is also visible in Fruit A in Figure S1b: a robust straight ridge (abaxial margin) of Fruit A (Figure 1a,d,g,h and Figure S1a,b) is covered by an additional thin layer. It is not a pure imagination that the fruit wall was integral and covered the whole fruit, although the current preservation cannot demonstrate this.
Seeds of Xenofructus vary in their morphology, even in the same fruit (Figure 1a–c,f andFigure S1a–d). Such a variation is rather expected, as ovules borne on the same placenta may be in their different developmental stages, as seen in extant Nymphaea [36][M2] . For example, three adjacent seeds of Fruit B are of different shapes (Figure 1c). The lowermost one is more corpulent and round in shape, the uppermost one is apparently more slender and smaller, sickle-shaped, while the middle one is intermediate between these two. Such variation suggests an acropetal development of the ovules in Xenofructus. It is noteworthy that all seeds are consistently sickle-shaped. Not alone, a similar variation of ovules can be seen in Fruit A (Figure 1a and Figure S1a,c,d). It is important that there seems to be an invagination at the tip of seed/ovule (Figure 1f). This conjures to micropyle, a feature frequently seen at the tip of an ovule, rejecting former interpretation of these seeds as pollen sacs. We attribute the above differences in seed morphology to the various developmental stages of the ovules in Xenofructus.
The most intriguing feature of Xenofructus may be its placenta positioning. Unlike previously reported fossil plants including Archaefructus [9,10,11,12], Sinocarpus [21,37], Archaeanthus [13,14], in which the seeds/ovules were interpreted as borne either on adaxial or abaxial margin of fruits, the seeds in Xenofructus are clustered around an axis independent of both adaxial and abaxial margins. This phenomenon is so far never seen in extant angiosperms. Therefore its occurrence in a Jurassic pioneer angiosperm Xenofructus is of significance, as it potentially represents an ancestral status of placenta. The placenta intermediately positioned between the adaxial and abaxial margins in fruit of Xenofructus indicates that previously seen adaxial or abaxial position of seeds in a (fossil or extant) fruit may represent a derived feature of carpels, and placenta originally is an ovule-bearing branch that is independent of either carpel margin. This conclusion is in agreement with result of studies on Michelia [5] and Illicium [4], and with the generalization given by Wang [3]. Most importantly, studies on gene function have long given a similar implications [38,39,40,41], which, unfortunately, have been largely ignored by many botanists. Such a consensus among palaeobotany, plant morphology, and molecular studies seems to indicate that we are approaching the truth.
There are increasing independent molecular datings suggesting a Jurassic or even Triassic origin of angiosperms [42,43,44,45,46,47]. A recent study using numerous genes of most taxa in angiosperms and 62 fossil calibrations [46,48] suggested that there is a “Jurassic Gap” for angiosperms. This conclusion is further strengthened by the latest phylogenomic study on 353 gene sets in almost 800 genera of angiosperms carried out by 279 scientists from 138 institutions in 27 countries [47]. Now our Xenofructus represents a Middle Jurassic (>164 Ma) angiosperm, filling the so-called “Jurassic Gap”. It appears that the impression of “Jurassic Gap” is either due to our lack of knowledge on Jurassic angiosperms, or due to intentional turning back to Jurassic angiosperms, just as Herendeen et al. [49] did. We cannot know the truth until we calm down and study fossils carefully. Actually, there have been enough studies suggestive of pre-Cretaceous angiosperms [3,19,26,27,28,33,34,50,51,52,53,54,55,56,57].
We still do not know anything about the whole plant of Xenofructus yet. The occurrence of two Xenofructus fruits on the surface of the same specimen seems to suggest that Xenofructus was not rare or proliferated in the flora. These fruits are parallel in orientation, suggesting either that they belonged to the same plant and were connected each other by an unpreserved part, or that they represented two of many fruits in the fossil flora, implying many unseen fruits awaiting further digging in the fossil flora. We expect future studies will shed more light on Xenofructus and other angiosperms that lived in the Jurassic of Liaoning, China.
Previous studies on megafossils and palynology of the Dabu Formation showed that the fossil flora bearing Xenofrucuts was dominated by Cycadales (28.57%), ferns (27.27%) and Ginkgoales (24.68%), followed by Coniferales (11.69%), Equisetales (6.49%), and Lycopodiales (1.3%) [16]. These plants constitute a forest dominated by trees of Ginkgoales, Czekanowskiales, and Coniferales, with flourished bushes of Cycadales and Bennettitales, and under store of ferns [16]. We hope this information can be helpful for future studies on the ecological background of angiosperm evolution. It is rather expected that some other primitive angiosperms may still survive in a more or less similar extant environment.
5. Conclusions
Xenofructus represents a new taxon of Jurassic angiosperms. Its early age and unique morphology not only confirm the truthful occurrence of angiosperms in the pre-Cretaceous age, but also shed a new light on the homology of carpels and on the origin of angiosperms.
Author Contributions
S. Z. collected the specimen; X.W. and Q.F. carried out the observation and photography; J.S. carried out the Micro-CT observation and rendering. X.W. and Q.F. drafted the manuscript. Q.F., J.S., S.Z. and X.W. modified and finalized the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research is supported by the National Natural Science Foundation of China (42288201).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data generated or analyzed during this study are included in this published article.
Acknowledgments
Dr. Wu Zhang is graciously thanked for collecting this invaluable specimen.
Conflicts of Interest
Q.F. and X.W. contributed to the study conception and design. S.Z. collected the material for study. J.S. carried out the Micro-CT observations and figure processing. All authors analyzed the data. The first draft of the manuscript was written by X. W. and all authors commented on versions of the manuscript. All authors read and approved the final manuscript.
References
- Cronquist, A. The evolution and classification of flowering plants., 2nd ed.; New York Botanical Garden: Bronx, 1988; p. 555. [Google Scholar]
- Wang, X.; Liu, Z.-J.; Liu, W.; Liao, W.; Zhang, X.; Liu, Z.; Hu, G.; Guo, X.; Wang, Y. Stepping out of the shadow of Goethe: for a more scientific plant systematics. Chinese Bulletin of Botany 2020, 55(4), 505–512. [Google Scholar]
- Wang, X. , The Dawn Angiosperms., 2nd ed.; Springer: Cham, Switzerland, 2018; p. 407. [Google Scholar]
- Zhang, X. , Floral ontogeny of Illicium lanceolatum (Schisandraceae) and its implications on carpel homology. Phytotaxa 2019, 416(3), 200–210. [Google Scholar] [CrossRef]
- Zhang, X.; Liu, W.; Wang, X. , How the ovules get enclosed in magnoliaceous carpels. PLOS ONE 2017, 12(4), e0174955. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Leslie, A. B.; Herendeen, P. S.; Herrera, F.; Ichinnorov, N.; Takahashi, M.; Knopf, P.; Crane, P. R. , Early Cretaceous Umkomasia from Mongolia: implications for homology of corystosperm cupules. New Phytologist 2016, 210(4), 1418–1429. [Google Scholar] [CrossRef] [PubMed]
- Crane, P. R.; Herendeen, P. S.; Herrera, F.; Shi, G., Diversity and homologies of corystosperm seed-bearing structures from the Early Cretaceous of Mongolia and China. In 10th European Palaeobotany and Palynology Conference, McElwain, J., Ed. Trinity College Dublin: Dublin, 2018; p 88.
- Endress, P. K. , The morphological relationship between carpels and ovules in angiosperms: pitfalls of morphological interpretation. Botanical Journal of the Linnean Society 2019, 189(3), 201–227. [Google Scholar] [CrossRef]
- Sun, G.; Dilcher, D. L.; Zheng, S.; Zhou, Z. , In search of the first flower: a Jurassic angiosperm, Archaefructus, from Northeast China. Science 1998, 282, 1692–1695. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.; Ji, Q.; Dilcher, D. L.; Zheng, S.; Nixon, K. C.; Wang, X. , Archaefructaceae, a new basal angiosperm family. Science 2002, 296, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Ji, Q.; Li, H.; Bowe, M.; Liu, Y.; Taylor, D. W., Early Cretaceous Archaefructus eoflora sp. nov. with bisexual flowers from Beipiao, Western Liaoning, China. Acta Geologica Sinica (English edition) 2004, 78 (4), 883-896.
- Wang, X.; Zheng, X.-T., Reconsiderations on two characters of early angiosperm Archaefructus. Palaeoworld 2012, 21 (3–4), 193-201.
- Wang, X. , New observation on seed/ovule position in the fruit of Archaeanthus and its systematic implications. China Geology 2021, 4, 752–755. [Google Scholar] [CrossRef]
- Dilcher, D. L.; Crane, P. R. , Archaeanthus: An early angiosperm from the Cenomanian of the Western Interior of North America. Annals of the Missouri Botanical Garden 1984, 71(2), 351–383. [Google Scholar] [CrossRef]
- Zheng, S.-L.; Zhang, W. , Early and Middle Jurassic fossil flora from Tianshifu Liaoning. Liaoning Geology 1990, 3, 212–237. [Google Scholar]
- Zheng, S.-L.; Zhang, W. , Basic characteristics of Tianshifu Flora. Liaoning Geology 1990, 4, 322–334. [Google Scholar]
- Harris, T. M., III. On Williamsoniella, a new type of bennettitalean flower. Philosophical Transactions of the Royal Society of London. Series B 1916, 207, 335–347. [Google Scholar]
- Harris, T. M. , Williamsoniella lignieri: Its pollen and the compression of spherical pollen grains. Palaeontology 1974, 17(1), 125–148. [Google Scholar]
- Han, L.; Zhao, Y.; Zhao, M.; Sun, J.; Sun, B.; Wang, X. , New fossil evidence suggests that angiosperms flourished in the Middle Jurassic. Life 2023, 13(3), 819. [Google Scholar] [CrossRef] [PubMed]
- Du, B.; Zhang, M.; Sun, B.; Li, A.; Zhang, J.; Yan, D.; Xie, S.; Wu, J. , An exceptionally well-preserved herbaceous eudicot from the Early Cretaceous (late Aptian-early Albian) of Northwest China. National Science Review 2021, 8(12), nwab084. [Google Scholar] [CrossRef] [PubMed]
- Leng, Q.; Friis, E. M., Sinocarpus decussatus gen. et sp. nov., a new angiosperm with basally syncarpous fruits from the Yixian Formation of Northeast China. Plant Systematics and Evolution 2003, 241, 77-88.
- Dilcher, D. L.; Cronquist, A.; Stevenson, D. W.; Zimmermann, M. H.; Berry, P. E.; Stevens, P. Angiosperm. https://www.britannica.com/plant/angiosperm (accessed 8 July).
- Wang, X. , Criterion is a touchstone in study of early angiosperms. Open Journal of Plant Science 2021, 6(1), 91–93. [Google Scholar]
- Wang, X. , A novel Early Cretaceous flower and its implications on flower derivation. Biology 2022, 11(7), 1036. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Shih, C.; Liu, Z.-J.; Lin, L.; Singh, K. J. , Reconstructing the Callianthus plant–An early aquatic angiosperm from the Lower Cretaceous of China. Cretaceous Research 2021, 128, 104983. [Google Scholar] [CrossRef]
- Fu, Q.; Diez, J. B.; Pole, M.; Garcia-Avila, M.; Liu, Z.-J.; Chu, H.; Hou, Y.; Yin, P.; Zhang, G.-Q.; Du, K.; Wang, X. , An unexpected noncarpellate epigynous flower from the Jurassic of China. eLife 2018, 7, e38827. [Google Scholar] [CrossRef] [PubMed]
- Fu, Q.; Diez, J. B.; Pole, M.; García-Ávila, M.; Wang, X. , Nanjinganthus is an angiosperm, isn’t it? China Geology 2020, 3(3), 359–361. [Google Scholar] [CrossRef]
- Fu, Q.; Hou, Y.; Yin, P.; Diez, J. B.; Pole, M.; García-Ávila, M.; Wang, X. Micro-CT results exhibit ovules enclosed in the ovaries of Nanjinganthus. Scientific Reports 2023, 13, 426. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Fu, X.; Liu, Z.-J.; Wang, X., A new angiosperm genus from the Lower Cretaceous Yixian Formation, Western Liaoning, China. Acta Geologica Sinica (English edition) 2013, 87 (4), 916-925.
- Han, G.; Liu, Z.; Wang, X., A Dichocarpum-like angiosperm from the Early Cretaceous of China. Acta Geologica Sinica (English edition) 2017, 90 (1), 1-8.
- Han, G.; Wang, X., A new infructescence of angiosperms from the Early Cretaceous of China. Acta Geologica Sinica (English edition) 2020, 94 (5), 1711-1713.
- Liu, X.; Ma, L.; Liu, B.; Liu, Z.-J.; Wang, X. , A novel angiosperm including various parts from the Early Cretaceous sheds new light on flower evolution. Historical Biology 2021, 33(11), 2706–2714. [Google Scholar] [CrossRef]
- Liu, Z.-J.; Wang, X. , A perfect flower from the Jurassic of China. Historical Biology 2016, 28(5), 707–719. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-J.; Wang, X. , Yuhania: a unique angiosperm from the Middle Jurassic of Inner Mongolia, China. Historical Biology 2017, 29(4), 431–441. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.-J.; Wang, X., A novel angiosperm from the Early Cretaceous and its implications on carpel-deriving. Acta Geologica Sinica (English edition) 2018, 92 (4), 1293-1298.
- Endress, P. K. , Multicarpellate gynoecia in angiosperms: occurrence, development, organization and architectural constraints. BOTANICAL JOURNAL OF THE LINNEAN SOCIETY 2014, 174(1), 1–43. [Google Scholar] [CrossRef]
- Leng, Q.; Friis, E. M., Angiosperm leaves associated with Sinocarpus infructescences from the Yixian Formation (Mid-Early Cretaceous) of NE China. Plant Systematics and Evolution 2006, 262 (3-4), 173-187.
- Roe, J. L.; Nemhauser, J. L.; Zambryski, P. C. , TOUSLED participates in apical tissue formation during gynoecium development in Arabidopsis. Plant Cell 1997, 9(3), 335–353. [Google Scholar] [PubMed]
- Rounsley, S. D.; Ditta, G. S.; Yanofsky, M. F. , Diverse roles for MADS box genes in Arabidopsis development. Plant Cell 1995 7, 1259–1269.
- Skinner, D. J.; Hill, T. A.; Gasser, C. S., Regulation of ovule development. Plant Cell 2004, 16 (suppl_1), S32-S45.
- Mathews, S.; Kramer, E. M. , The evolution of reproductive structures in seed plants: A re-examination based on insights from developmental genetics. New Phytologist 2012, 194(4), 910–923. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.; Gierl, A.; Saedler, H. , Molecular evidence for pre-Cretaceous angiosperm origins. Nature 1989, 339, 46–48. [Google Scholar] [CrossRef]
- Martin, W.; Gierl, A.; Saedler, H. , Angiosperm origins. Nature 1989, 342, 132. [Google Scholar] [CrossRef]
- Soltis, D. E.; Bell, C. D.; Kim, S.; Soltis, P. S. , Origin and early evolution of angiosperms. Annals of the New York Academy of Sciences 2008, 1133, 3–25. [Google Scholar] [CrossRef] [PubMed]
- Soltis, P. S.; Soltis, D. E. , The origin and diversification of angiosperms. Am. J. Bot. 2004, 91(10), 1614–1626. [Google Scholar] [CrossRef] [PubMed]
- Li, H.-T.; Yi, T.-S.; Gao, L.-M.; Ma, P.-F.; Zhang, T.; Yang, J.-B.; Gitzendanner, M. A.; Fritsch, P. W.; Cai, J.; Luo, Y.; Wang, H.; van der Bank, M.; Zhang, S.-D.; Wang, Q.-F.; Wang, J.; Zhang, Z.-R.; Fu, C.-N.; Yang, J.; Hollingsworth, P. M.; Chase, M. W.; Soltis, D. E.; Soltis, P. S.; Li, D.-Z. , Origin of angiosperms and the puzzle of the Jurassic gap. Nature Plants 2019, 5(5), 461–470. [Google Scholar] [CrossRef] [PubMed]
- Zuntini, A. R.; Carruthers, T.; Maurin, O.; Bailey, P. C.; Leempoel, K.; Brewer, G. E.; Epitawalage, N.; Françoso, E.; Gallego-Paramo, B.; McGinnie, C.; Negrão, R.; Roy, S. R.; Simpson, L.; Toledo Romero, E.; Barber, V. M. A.; Botigué, L.; Clarkson, J. J.; Cowan, R. S.; Dodsworth, S.; Johnson, M. G.; Kim, J. T.; Pokorny, L.; Wickett, N. J.; Antar, G. M.; DeBolt, L.; Gutierrez, K.; Hendriks, K. P.; Hoewener, A.; Hu, A.-Q.; Joyce, E. M.; Kikuchi, I. A. B. S.; Larridon, I.; Larson, D. A.; de Lírio, E. J.; Liu, J.-X.; Malakasi, P.; Przelomska, N. A. S.; Shah, T.; Viruel, J.; Allnutt, T. R.; Ameka, G. K.; Andrew, R. L.; Appelhans, M. S.; Arista, M.; Ariza, M. J.; Arroyo, J.; Arthan, W.; Bachelier, J. B.; Bailey, C. D.; Barnes, H. F.; Barrett, M. D.; Barrett, R. L.; Bayer, R. J.; Bayly, M. J.; Biffin, E.; Biggs, N.; Birch, J. L.; Bogarín, D.; Borosova, R.; Bowles, A. M. C.; Boyce, P. C.; Bramley, G. L. C.; Briggs, M.; Broadhurst, L.; Brown, G. K.; Bruhl, J. J.; Bruneau, A.; Buerki, S.; Burns, E.; Byrne, M.; Cable, S.; Calladine, A.; Callmander, M. W.; Cano, Á.; Cantrill, D. J.; Cardinal-McTeague, W. M.; Carlsen, M. M.; Carruthers, A. J. A.; de Castro Mateo, A.; Chase, M. W.; Chatrou, L. W.; Cheek, M.; Chen, S.; Christenhusz, M. J. M.; Christin, P.-A.; Clements, M. A.; Coffey, S. C.; Conran, J. G.; Cornejo, X.; Couvreur, T. L. P.; Cowie, I. D.; Csiba, L.; Darbyshire, I.; Davidse, G.; Davies, N. M. J.; Davis, A. P.; van Dijk, K.-j.; Downie, S. R.; Duretto, M. F.; Duvall, M. R.; Edwards, S. L.; Eggli, U.; Erkens, R. H. J.; Escudero, M.; de la Estrella, M.; Fabriani, F.; Fay, M. F.; Ferreira, P. d. L.; Ficinski, S. Z.; Fowler, R. M.; Frisby, S.; Fu, L.; Fulcher, T.; Galbany-Casals, M.; Gardner, E. M.; German, D. A.; Giaretta, A.; Gibernau, M.; Gillespie, L. J.; González, C. C.; Goyder, D. J.; Graham, S. W.; Grall, A.; Green, L.; Gunn, B. F.; Gutiérrez, D. G.; Hackel, J.; Haevermans, T.; Haigh, A.; Hall, J. C.; Hall, T.; Harrison, M. J.; Hatt, S. A.; Hidalgo, O.; Hodkinson, T. R.; Holmes, G. D.; Hopkins, H. C. F.; Jackson, C. J.; James, S. A.; Jobson, R. W.; Kadereit, G.; Kahandawala, I. M.; Kainulainen, K.; Kato, M.; Kellogg, E. A.; King, G. J.; Klejevskaja, B.; Klitgaard, B. B.; Klopper, R. R.; Knapp, S.; Koch, M. A.; Leebens-Mack, J. H.; Lens, F.; Leon, C. J.; Léveillé-Bourret, É.; Lewis, G. P.; Li, D.-Z.; Li, L.; Liede-Schumann, S.; Livshultz, T.; Lorence, D.; Lu, M.; Lu-Irving, P.; Luber, J.; Lucas, E. J.; Luján, M.; Lum, M.; Macfarlane, T. D.; Magdalena, C.; Mansano, V. F.; Masters, L. E.; Mayo, S. J.; McColl, K.; McDonnell, A. J.; McDougall, A. E.; McLay, T. G. B.; McPherson, H.; Meneses, R. I.; Merckx, V. S. F. T.; Michelangeli, F. A.; Mitchell, J. D.; Monro, A. K.; Moore, M. J.; Mueller, T. L.; Mummenhoff, K.; Munzinger, J.; Muriel, P.; Murphy, D. J.; Nargar, K.; Nauheimer, L.; Nge, F. J.; Nyffeler, R.; Orejuela, A.; Ortiz, E. M.; Palazzesi, L.; Peixoto, A. L.; Pell, S. K.; Pellicer, J.; Penneys, D. S.; Perez-Escobar, O. A.; Persson, C.; Pignal, M.; Pillon, Y.; Pirani, J. R.; Plunkett, G. M.; Powell, R. F.; Prance, G. T.; Puglisi, C.; Qin, M.; Rabeler, R. K.; Rees, P. E. J.; Renner, M.; Roalson, E. H.; Rodda, M.; Rogers, Z. S.; Rokni, S.; Rutishauser, R.; de Salas, M. F.; Schaefer, H.; Schley, R. J.; Schmidt-Lebuhn, A.; Shapcott, A.; Al-Shehbaz, I.; Shepherd, K. A.; Simmons, M. P.; Simões, A. O.; Simões, A. R. G.; Siros, M.; Smidt, E. C.; Smith, J. F.; Snow, N.; Soltis, D. E.; Soltis, P. S.; Soreng, R. J.; Sothers, C. A.; Starr, J. R.; Stevens, P. F.; Straub, S. C. K.; Struwe, L.; Taylor, J. M.; Telford, I. R. H.; Thornhill, A. H.; Tooth, I.; Trias-Blasi, A.; Udovicic, F.; Utteridge, T. M. A.; Del Valle, J. C.; Verboom, G. A.; Vonow, H. P.; Vorontsova, M. S.; de Vos, J. M.; Al-Wattar, N.; Waycott, M.; Welker, C. A. D.; White, A. J.; Wieringa, J. J.; Williamson, L. T.; Wilson, T. C.; Wong, S. Y.; Woods, L. A.; Woods, R.; Worboys, S.; Xanthos, M.; Yang, Y.; Zhang, Y.-X.; Zhou, M.-Y.; Zmarzty, S.; Zuloaga, F. O.; Antonelli, A.; Bellot, S.; Crayn, D. M.; Grace, O. M.; Kersey, P. J.; Leitch, I. J.; Sauquet, H.; Smith, S. A.; Eiserhardt, W. L.; Forest, F.; Baker, W. J. , Phylogenomics and the rise of the angiosperms. Nature 2024. [Google Scholar] [CrossRef] [PubMed]
- Mandel, J. R. , A Jurassic leap for flowering plants. Nature Plants 2019, 5, 455–456. [Google Scholar] [CrossRef] [PubMed]
- Herendeen, P. S.; Friis, E. M.; Pedersen, K. R.; Crane, P. R. , Palaeobotanical redux: revisiting the age of the angiosperms. Nature Plants 2017, 3, 17015. [Google Scholar] [CrossRef] [PubMed]
- Hochuli, P. A.; Feist-Burkhardt, S. , A boreal early cradle of angiosperms? angiosperm-like pollen from the Middle Triassic of the Barents Sea (Norway). Journal of Micropalaeontology 2004, 23, 97–104. [Google Scholar] [CrossRef]
- Hochuli, P. A.; Feist-Burkhardt, S. , Angiosperm-like pollen and Afropollis from the Middle Triassic (Anisian) of the Germanic Basin (Northern Switzerland). Frontiers in Plant Science 2013, 4, 344. [Google Scholar] [CrossRef]
- Prasad, V.; Strömberg, C. A. E.; Leaché, A. D.; Samant, B.; Patnaik, R.; Tang, L.; Mohabey, D. M.; Ge, S.; Sahni, A. , Late Cretaceous origin of the rice tribe provides evidence for early diversification in Poaceae. Nature Communication 2011, 2, 480. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; You, H.-L.; Li, X.-Q. , Dinosaur-associated Poaceae epidermis and phytoliths from the Early Cretaceous of China. National Science Review 2018, 5, 721–727. [Google Scholar] [CrossRef]
- Wu, Z.-Y.; Lu, A.-M.; Tang, Y.-C.; Chen, Z.-D.; Li, D.-Z. , Synopsis of a new “polyphyletic-polychronic-polytopic” system of the angiosperms. Acta Phytotaxonomica Sinica 2002, 40(4), 289–322. [Google Scholar]
- Han, G.; Liu, Z.-J.; Liu, X.; Mao, L.; Jacques, F. M. B.; Wang, X., A whole plant herbaceous angiosperm from the Middle Jurassic of China. Acta Geologica Sinica (English edition) 2016, 90 (1), 19-29.
- Cui, D.-F.; Hou, Y.; Yin, P.; Wang, X., A Jurassic flower bud from China. Geological Society, London, Special Publications 2022, 521, 81-93.
- Wang, X.; Fu, Q. , Taiyuanostachya: An abominable angiosperm from the Early Permian of China. Journal of Biotechnology and Biomedicine 2023, 6(3), 371–379. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



