
Submitted:
06 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Pygeum africanum bark has been shown to inhibit the production of pro-inflammatory prostaglandins in the prostate and reduce the production of leukotrienes and other 5-lipoxygenase (5-LO) metabolites. It has been suggested that inflammation plays an important role in the pathophysiology of benign prostatic hyperplasia (BPH). In clinical trials, P. africanum improved the symptoms and objective measures of BPH. This in vitro study aimed to assess the anti-inflammatory potential of a proprietary Pygeum bark standardized extract (Prunera®) on cytokine release by lipopolysaccharide-simulated human peripheral blood mononuclear cells (PBMCs). PBMCs were obtained from four donors and a bead-based assay (ProcartaPlex™ panel) was used for the detection and quantitation of cytokines. Pygeum africanum bark standardized extract (PABE) showed the following effects: IL-2 was lowered in all donors in absence of a clear dose–response relationship; IL-4, IL-5, IL-6, IL-9, and IL-13 levels were decreased in most donors; IL-22 levels seemed to be suppressed only for donor 4 at lower and medium concentrations; and IL-27 and levels of TNF-α decreased at all PABE concentrations in all donors. The anti-inflammatory effect of PABE supports the potential use of this natural compound in the management of BPH and other conditions in which pro-inflammatory cytokines are involved in their underlying pathophysiological mechanisms.
Keywords:
Pygeum africanum
; inflammation
; cytokine release
; lymphocites
; natural products
; phytotherapy
; benign prostatic hyperplasia
1. Introduction
Benign prostatic hyperplasia (BPH) is a histological diagnosis and refers to a nodular overgrowth of the epithelium and fibromuscular tissue within the prostate transition zone and periurethral areas. However, several definitions exist in the literature when describing BPH. These include bladder outlet obstruction, lower urinary tract symptoms (LUTS), and benign prostatic enlargement. BPH describes the histological changes, benign prostatic enlargement describes the increased size of the gland (usually secondary to BPH) and bladder outlet obstruction describes the obstruction to flow. BPH is the most common condition in ageing men, associated with LUTS. LUTS simply describe urinary symptoms shared by disorders affecting the bladder and prostate (when in reference to men). LUTS can be subdivided into storage and voiding symptoms, and these terms have largely replaced those historically termed “prostatism.”
The development of BPH is characterized by stromal and epithelial cell proliferation in the prostate transition zone (surrounding the urethra), this leads to compression of the urethra and development of bladder outflow obstruction which can result in clinical manifestations of LUTS, urinary retention or infections due to incomplete bladder emptying [1]. Multiple pleiotropic mechanisms have been involved in the prostatic tissue remodeling process, but the pathogenesis of BPH is still not fully understood. It has been shown that prostate tissue remodeling in the transition zone is characterized by i) hypertrophic basal cells; ii) altered secretions of luminal cells leading to calcification, clogged ducts, and inflammation; iii) lymphocytic infiltration with production of pro-inflammatory cytokines; and iv) increased production of basic fibroblast growth factor (bFGF) and transforming growth factor beta 1 (TGF-beta 1), leading to stromal proliferation, transdifferentiation, and extracellular matrix production [1]. An investigation of lymphocyte-derived growth factors on prostatic stromal cells showed that BPH-tissues expressed interferon gamma (IFN-γ) and interleukin (IL) IL-2 and IL-4 mRNA, concluding that chronic inflammation may induce an increased growth pattern of fibromuscular tissue in BPH [2]. Other studies further support the link between chronic inflammation and the progression of BPH [3,4]. BPH myxoid nodules are a constant finding in BPH surgical specimens [5], and BPH nodules frequently occur with interstitial distribution of chronic inflammatory infiltrates mainly composed of chronically activated T-cell lymphocytes and macrophages [6]. A chronic inflammatory state may lead to tissue damage, activate the release of cytokines, increase the concentration of growth factors, and cause a local vicious cycle. The role and the relationship of pro-inflammatory cytokines in chronic inflammation seem to be a critical component of BPH pathogenesis [7]. Accordingly, reducing inflammation has been postulated as an approach in the management of BPH [8].
Pygeum africanum (Prunus Africana) commonly known as African cherry is a member of the Rosaceae family of an evergreen species found across the entire continent of Africa at altitudes of 3,000 feet or higher. The leaves impart a characteristic almond smell when bruised. The tree bears creamy-white flowers and purplish-red cherries that turn black when ripe. Prunus africana is widespread in many African countries including South Africa, Zimbabwe, Kenya, Ethiopia, Democratic Republic of Congo, Cameroon and Madagascar. The bark is exploited for its medicinal properties.
Pygeum bark contains fat-soluble sterols and fatty acids as its major active components, including phytosterols like beta-sitosterol. These phytosterols possess anti-inflammatory properties that effectively inhibit the prostate gland's production of pro-inflammatory prostaglandins. Pygeum bark also contains pentacyclic triterpenes, ferulic acid, n-docosanol, and tetracosanol, which contribute to reducing prolactin levels and preventing cholesterol accumulation in the prostate. Extracts of Pygeum standardized to 13%–14% phytosterols have been extensively researched in animal studies as well as human clinical trials. [9]. Pygeum africanum bark extract (PABE) has potentially relevant biological properties, including inhibition of chemotactic activity of leukotrienes; antagonization of 5-lipoxygenase metabolite production; inhibition of growth factors (bFGF, insulin growth factor-1 [IGF-1], epidermal growth factor [EGF]) and fibroblast proliferation; improvement of prostatic histology; and restoration of prostatic and bulbourethral epithelium secretory activity [10,11,12,13,14].
In a study of the proliferation of rat prostatic stromal cells stimulated with different growth factors (epidermal growth factor [EGF], basic fibroblast growth factor [bFGF], and insulin-like growth factor 1 [IGF-I]), P. africanum inhibited both basal and stimulated growth with IC50 values of 4.5, 7.7 and 12.6 µg/ml. for EGF, IGF-I and bFGF, respectively, compared to 14.4 µg/ml. for untreated cells [11]. Also, P. africanum inhibited the proliferation induced by protein kinases (TPA or PDBu) in a concentration-dependent manner. These results suggest that the beneficial effect of P. africanum may be related to inhibition of growth factors responsible for the prostatic overgrowth in man. In another study of primary cultures of prostatic stromal cells obtained from histologically confirmed human BPH by enzymatic digestion, an extract of P. africanum inhibited the proliferation of prostatic myofibroblasts and fibroblasts [14].
Phytotherapy has become an increasingly popular option for the treatment of BPH, with various herbal extracts leading to subjective and objective symptom improvement beyond a placebo effect in men with BPH. PABE as one of the common phytotherapeutic agents used to ameliorate LUTS attributable to BPH [15,16]. In a review of the literature of the role of phytotherapy in the management of BPH, proposed mechanisms of action of P. africanum bark in LUTS include anti-androgen by inhibiting androgen and progesterone receptors, inhibition of prostate cell proliferation and apoptosis of stromal cells, anti-inflammatory through inhibition of 5-lipoxygenase, and histamine-mediated protective response to the detrusor muscle in the bladder [15]. A 2020 monograph of the European Scientific Cooperative on Phytotherapy (ESCOP) on Pygeum africanum bark summarizes the therapeutic indications of this compound for the symptomatic treatment of micturition disorders (dysuria, pollakiuria, nocturia, urine retention) in BPH (stages I, II, and III as defined by Alken and Vahlensieck) [17]. In a systematic review of 18 studies involving 1562 men with symptomatic BPH, PABE significantly improved urologic symptoms and flow measures [18]. In a multicenter study carried out in central Europe, PABE treatment for 2 months improved domains of the International Prostate Symptom Score (IPSS) questionnaire, quality of life (QoL), and urinary flow parameters [19]. In a cross-sectional study of men with LUTS associated with BPH treated in real-world practice with PABE for 6 months, significant symptom improvement and QoL as well as patient satisfaction and compliance with treatment was observed [20]. In a report on Prunus africana of the Committee on Herbal Medicinal Products of the European Medicines Agency, based on data of clinical reviews involving a total of 1310 patients and daily doses of 75-200 mg of lipophilic extracts and treatment periods from 15 to 120 days, it was concluded that Pygeum bark improved the symptoms and the objective measures of BPH and was well tolerated [21].
Cytokines are involved in both the inflammatory process and the epithelial/stromal prostatic cell interactions [22] and given the well-documented effects of PABE on inflammation and cell proliferation, it was considered of interest to assess the effects of its bark extract on cytokine production using an established in vitro model of lipopolysaccharide (LPS)-stimulated human peripheral blood mononuclear cells (PBMCs). Results of the study will contribute to define the mechanisms by which PABE improves symptoms of BPH as shown in clinical studies.
2. Results
2.1. Cytokine Release
The mean (standard deviation, SD) values of changes of the release of cytokines observed in the four donors are shown in Table 1.
Changes in cytokine levels compared to untreated controls are shown in Figure 1. PABE showed the following effects on the investigated cytokines: IL-1β levels remained unaffected by the compound in all four donors. IL-2 was lowered upon exposure to PABE in all donors in the absence of a clear dose–response relationship. IL-4 and IL-5 levels were lowered in donors 1–3, whereas donor 4 showed only marginal changes. IL-6 levels were strongly reduced in donors 1–3, while donor 4, showed only minor changes. IL-9 levels were reduced in donors 1, 2, and 4 while donor 3 featured no clear effects. IL-13 levels were suppressed in all four donors. IL-22 levels seemed to be suppressed only for donor 4, at lower and medium concentrations. Finally, IL-27 and TNF-α were lowered at all concentrations in all four donors.
The mean values of changes in the individual cytokines according to the doses of PABE are shown in Table S1 of the Supplementary material.
2.2. Viability Assays
Viability of the PBMCs was determined by CTG assay after the centrifugation step and withdrawal of the supernatant for cytokine analysis. As shown in Figure 2, PABE had no negative impact on viability of PBMCs, and all data of the cytokine release assay were valid.
3. Discussion
The present in vitro model provides evidence of the full anti-inflammatory potential of PABE with a trend towards suppression of pro-inflammatory cytokines; although a clear dose-response relationship was not found. However, it may be possible that concentrations of PABE might be still too high to make apparent a biological dose-response. Further studies with lower doses of the active compound would be needed to clarify this point.
Results of the study shed light on the mechanisms by which PABE ameliorates the symptoms of BPH, and are, at the very least, suggestive of the potential of this bark extract to be effective in the management of other clinical conditions, the etiology of which may involve a relevant role of pro-inflammatory cytokines. On the other hand, all cytokines selected for our study possess characteristics that contribute to inflammation, cell proliferation, and other pathophysiologic conditions which can be factors in the development of BPH (Table 2).
BPH and associated symptoms of urinary tract outlet obstruction are consequences of hyperproliferation, mainly within the stromal component of the prostate with increased tonicity of the prostatic smooth muscles. Nodular and fibromuscular hyperplasia are the primary cellular events involved in the pathogenesis of BPH. Studies in cultured prostatic stromal cells obtained from histologically confirmed BPH have shown the antiproliferative effects of Pygeum, with inhibition of prostatic myofibroblasts and fibroblasts and induction of apoptosis of stromal cells [23,24]. It has been suggested that the putative molecular basis of inhibition of prostate epithelial and stromal cells include cell cycle arrest, inhibition of cell signaling pathways at the level of protein kinase C, apoptosis, and binding ability to estrogen and androgen receptors [24].
Growth factors, particularly bFGF, which is found at elevated levels in BPH tissue, appear to play a role in the pathogenesis of BPH. In a study that examined the effects of PABE on basal cell proliferation and on the proliferation induced by bFGF, EGF and IGF-1, it was found that it had a much larger inhibitory effect on the proliferation of 3T3 fibroblasts when proliferation was induced by 0.5 µg/mL of bFGF, with the effect being significant at 1 µg/mL. PABE also inhibited EGF-induced cell proliferation but to a lesser extent [25], and inhibition of cell growth induced by certain growth factors may explain at least in part the therapeutic effect of PABE in BHP.
Although BPH is primarily characterized by prostatic cell proliferation, inflammation seems to play an important role in the initiation, development, and progression of BPH [22]. Human stromal prostate cells obtained from BPH tissue can actively contribute to the inflammatory process by secreting pro-inflammatory cytokines as well as chemokines to recruit lymphomononuclear cells [26]. Elevated expression of pro-inflammatory cytokines (IL-6, IL-8, and IL-17) may perpetuate chronic immune response in BPH and induce fibromuscular growth by an autocrine or paracrine loop or via induction of COX-2 expression [27,28]. In addition, a possible inhibitory effect of Pygeum on fibroblast growth factor (FGF-beta 1) and an anti-inflammatory role on neutrophil granulocytes has been reported [29]. A lipophilic extract of PABE inhibited the production of 5-lipoxygenase metabolites, such as chemotactic leukotrienes, in human polymorphonuclear cells stimulated by the calcium ionophore A23187 [30].
Results of the present study should be interpreted considering the small sample size, which limits its external validity. However, the large panel of cytokines analyzed is an interesting contribution of the study.
4. Materials and Methods
4.1. Human Peripheral Blood Mononuclear Cells
Institutional Review Board approval of the study protocol was provided by Quorum Review IRB (Seattle, WA, USA).
Human peripheral blood mononuclear cells (PBMCs) (ePBMC®- uncharacterized cryopreserved human PBMC, size < 10 x 106 cells per vial) were purchased from Cellular Technology Limited (CLT) (Bonn, Germany). Description of the product was as follows: human PBMC isolated from leukopacks and frozen in CTL-CryoABC™ serum-free freezing medium. These leukopacks were ethically collected from four healthy donors (Table 3) with no risk of breaching privacy. All samples tested negative for hepatitis B surface antigen (HBsAg), hepatitis B core antibody (HBcAb), hepatitis C virus (HCV), human T lymphotropic virus types 1 and 2 (HTLV I/II), and standard tests for syphilis (STS) by serology, as well as human immunodeficiency virus type 1 (HIV-1), HCV, and West Nile virus (WNV) by nucleic acid testing.
A 100 micromolar (100x) master stock concentration of P. africanum bark extract (Prunera®, Euromed, S.A., Mollet del Vallés, Barcelona, Spain) was made in ethanol at 50 mg/mL. The Cmax in the final assay was 500 µg/mL corresponding to 1% ethanol. All subsequent dilutions and the control were prepared using culture media supplemented with 1% ethanol. The stock solution had a brownish appearance with no signs of undissolved particles.
The test product, Prunera®, is a lipid sterolic extract derived from the dried bark of the stems and branches of Prunus africana, sustainably sourced and certified by the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), ensuring ethical and environmentally responsible practices. It is meticulously standardized to contain no less than 13% of beta-sitosterol, determined using a spectrophotometric UV method. This extract undergoes rigorous analysis by gas chromatography (GC), following the chromatographic method outlined in the United States Pharmacopoeia (USP) Pygeum extract monograph [31], to quantify the total sterols content, expressed as beta-sitosterol. The method entails the analysis of sterols including campesterol, stigmasterol, and beta-sitosterol in a Pygeum africanum extract (Figure 3). The internal standard, 5 alpha-cholestane, is employed for accurate measurements, and the result is expressed as beta-sitosterol. The analytical results consistently indicate that the total sterol content in the test product exceeds 10%.
4.2. Cell Viability Assay
For detection of viable cells, the CellTiter-Glo® Luminescent Cell Viability Assay (Promega Corporation, Madison, WI, USA) was used. This is a homogeneous method of determining the number of viable cells in culture based on quantitation of the ATP present, as an indicator of metabolically active cells. The procedure involves adding a single reagent (CellTiter-Glo® reagent) directly to cells cultured in serum-supplemented medium. The homogeneous “add-mix-measure” format results in cell lysis and generation of a luminescent signal proportional to the amount of ATP present. The assay system is characterized by a high sensitivity and excellent linearity.
For the dose-response relationship, absolute luminescence (background subtracted) was related to the negative (medium) control, and relative viability values were plotted against the test item concentration.
4.3. Cytokine Release Assay
Due to the small amount of cytokines released by PBMCs into the supernatant, a bead-based assay (ProcartaPlex™, Santa Clara, CA, USA) using the Luminex® instrument platform (Luminex Corporation, Austin, TX, USA) was used to quantitate 18 different cytokines in parallel within a 50 μL sample using appropriate calibration standards. Human cryopreserved PBMCs were thawed according to the manufacturer’s instructions. Cells were washed, resuspended in RPMI1640 containing 10% fetal bovine serum (FBS), plated in 96-well microtiter plates at 300,000 PBMCs/well, stimulated with lipopolysaccharide (LPS) at 10 µg/mL, and then exposed to different concentrations of PABE for 24 hours. Plates were centrifuged and cell-free supernatant was collected and forwarded to cytokine bead-array assay, which was performed according to the manufacturer’s instructions and read using a MagPix multiplex reader.
For the dose-response relationship, absolute concentrations were calculated by the MagPix software using two separate calibration series as provided by the manufacturer and summarized in table form.
5. Conclusions
The anti-inflammatory effect of P. africanum standardized bark extracts based on inhibition of cytokine release in the in vitro model of LPS-stimulated human PBMCs supports the potential use of this natural compound in the management of BPH and other conditions in which pro-inflammatory cytokines are involved in their underlying pathophysiological mechanisms.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org, Table S1.
Author Contributions
Conceptualization, A.V. and A.M..; methodology, A.V.; formal analysis, A.V.; investigation, A.V.; resources, A.Z.; data curation, A.V. and F.S.-F.; writing—original draft preparation, A.V.; writing—review and editing, A.V., A.M., F.S.-F. and A.Z.; visualization, A.V., F.S.-F. and A.Z.; supervision, A.V., F.S.-F. and A.Z.; project administration, A.V., F.S.-F. and A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study protocol was provided by Quorum Review IRB (Seattle, WA, USA).
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are available from the authors upon request.
Acknowledgments
The authors thank Marta Pulido, MD, for editing the manuscript and editorial assistance.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Untergasser, G.; Madersbacher, S.; Berger, P. Benign prostatic hyperplasia: age-related tissue-remodeling. Exp Gerontol 2005, 40, 121–128. [Google Scholar] [CrossRef]
- Kramer, G.; Steiner, G.E.; Handisurya, A.; Stix, U.; Haitel, A.; Knerer, B.; Gessl, A.; Lee, C.; Marberger, M. Increased expression of lymphocyte-derived cytokines in benign hyperplastic prostate tissue, identification of the producing cell types, and effect of differentially expressed cytokines on stromal cell proliferation. Prostate 2002, 52, 43–58. [Google Scholar] [CrossRef]
- Steiner, G.; Gessl, A.; Kramer, G.; Schöllhammer, A.; Förster, O.; Marberger, M. Phenotype and function of peripheral and prostatic lymphocytes in patients with benign prostatic hyperplasia. J Urol 1994, 151, 480–484. [Google Scholar] [CrossRef]
- Norström, M.M.; Rådestad, E.; Sundberg, B.; Mattsson, J.; Henningsohn, L.; Levitsky, V.; Uhlin, M. Progression of benign prostatic hyperplasia is associated with pro-inflammatory mediators and chronic activation of prostate-infiltrating lymphocytes. Oncotarget 2016, 7, 23581–23593. [Google Scholar] [CrossRef]
- Manzarbeitia, F.; Vela Navarrete, R.; Fernández-Aceñero, M.J. Early histopathological aspects of benign prostatic hyperplasia: myxoid-inflammatory nodules. Actas Urol Esp 2010, 34, 549–554. [Google Scholar] [CrossRef]
- Wang, L.; Yang, J.R.; Yang, L.Y.; Liu, Z.T. Chronic inflammation in benign prostatic hyperplasia: implications for therapy. Med Hypotheses 2008, 70, 1021–1023. [Google Scholar] [CrossRef]
- Naiyila, X.; Li, J.; Huang, Y.; Chen, B.; Zhu, M.; Li, J.; Chen, Z.; Yang, L.; Ai, J.; Wei, Q.; et al. A novel insight into the immune-related interaction of inflammatory cytokines in benign prostatic hyperplasia. J Clin Med 2023, 12, 1821. [Google Scholar] [CrossRef]
- Taille de la, A. Therapeutic approach: The importance of controlling prostatic inflammation. Eur J Urol Suppl 2013, 12, 116–122. [Google Scholar] [CrossRef]
- Anonymous. Pygeum africanum (Prunus africana) (African plum tree) Monograph. Altern Med Rev 2002, 7, 71–74. [Google Scholar]
- Andro, M.C.; Riffaud, J.P. Pigeum africanum extract for the treatment of patients with benign prostatic hyperplasia: review of 25 years of published experience. Curr Ther Res 1995, 56, 796–817. [Google Scholar] [CrossRef]
- Yablonsky, F.; Nicolas, V.; Riffaud, J.P.; Bellamy, F. Antiproliferative effect of Pygeum africanum extract on rat prostatic fibroblasts. J Urol 1997, 157, 2381–2387. [Google Scholar] [CrossRef]
- Paubert-Braquet, M.; Cave, A.; Hocquemiller, R.; Delacroix, D.; Dupont, C.; Hedef, N.; Borgeat, P. Effect of Pygeum africanum extract on A23187-stimulated production of lipoxygenase metabolites from human polymorphonuclear cells. J Lipid Mediat Cell Signal 1994, 9, 285–290. [Google Scholar]
- Lucchetta, G.; Weill, A.; Becker, N.; Clavert, A.; Bollack, C. Reactivation of the secretion from the prostatic gland in cases of reduced fertility. Biological study of seminal fluid modifications. Urol Int 1984, 39, 222–224. [Google Scholar] [CrossRef]
- Boulbès, D.; Soustelle, L.; Costa, P.; Haddoum, M.; Bali, J.P.; Hollande, F.; Magous, R. Pygeum africanum extract inhibits proliferation of human cultured prostatic fibroblasts and myofibroblasts. BJU Int 2006, 98, 1106–1113. [Google Scholar] [CrossRef]
- Antoniou, V.; Gauhar, V.; Modi, S.; Somani, B.K. Role of phytotherapy in the management of BPH: A summary of the literature. J Clin Med 2023, 12, 1899. [Google Scholar] [CrossRef]
- Jena, A.K.; Vasisht, K.; Sharma, N.; Kaur, R.; Dhingra, M.S.; Karan, M. Amelioration of testosterone induced benign prostatic hyperplasia by Prunus species. J Ethnopharmacol 2016, 22, 33–45. [Google Scholar] [CrossRef]
- ESCOP Monographs. The Scientific Foundation for Herbal Medicinal Products. Pruni africanae cortex. Pygeum africanum bark. 2020. Available online: https://www.sefit.es/corteza-pigeum-pruni-africanae-cortex-pygeum-africanum-bark-monografia-online-escop/ (accessed on 31 March 2023).
- Pagano, E.; Laudato, M.; Griffo, M.; Capasso, R. Phytotherapy of benign prostatic hyperplasia. A minireview. Phytother Res 2014, 28, 949–955. [Google Scholar] [CrossRef]
- Ishani, A.; MacDonald, R.; Nelson, D.; Rutks, I.; Wilt, T.J. Pygeum africanum for the treatment of patients with benign prostatic hyperplasia: a systematic review and quantitative meta-analysis. Am J Med 2000, 109, 654–664. [Google Scholar] [CrossRef]
- Breza, J.; Dzurny, O.; Borowka, A.; Hanus, T.; Petrik, R.; Blane, G.; Chadha-Boreham, H. Efficacy and acceptability of tadenan (Pygeum africanum extract) in the treatment of benign prostatic hyperplasia (BPH): A multicentre trial in central Europe. Curr Med Res Opin 1998, 143, 127–139. [Google Scholar] [CrossRef]
- Cambronero, J.; Osca-García, J.M.; Merino-Salas, S.; Miguel, J.M.; Borralleras, C.; López-Alcina, E. Effectiveness of treatment with Pygeum africanum in patients with lower urinary tract symptoms and benign prostatic hyperplasia: a cross-sectional study in the real-world clinical practice in Spain (The PROFIT Study). Arch Esp Urol 2022, 75, 219–227. [Google Scholar]
- European Medicines Agency. Committee on Herbal Medicinal Products (HMPC). 12 July 2016. EMA/HMPC/680624/2’13. Assessment report on Prunus africana (Hook f.) Kalkm., cortex. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-prunus-africana-hook-f-kalkm-cortex_en.pdf (accessed on 31 March 2023).
- Chughtai, B.; Lee, R.; Te, A.; Kaplan, S. Inflammation and benign prostatic hyperplasia: clinical implications. Curr Urol Rep 2011, 12, 274–277. [Google Scholar] [CrossRef]
- Boulbès, D.; Soustelle, L.; Costa, P.; Haddoum, M.; Bali, J.P.; Hollande, F.; Magous, R. Pygeum africanum extract inhibits proliferation of human cultured prostatic fibroblasts and myofibroblasts. BJU Int 2006, 98, 1106–1113. [Google Scholar] [CrossRef]
- Quiles, M.T.; Arbós, M.A.; Fraga, A.; de Torres, I.M.; Reventós, J.; Morote, J. Antiproliferative and apoptotic effects of the herbal agent Pygeum africanum on cultured prostate stromal cells from patients with benign prostatic hyperplasia (BPH). Prostate 2010, 70, 1044–1053. [Google Scholar] [CrossRef]
- Paubert-Braquet, M.; Monboisse, J.C.; Servent-Saez, N.; Serikoff, A.; Cavé, A.; Hocquemiller, R.; Dupont, C.; Forneau, C.; Borel, J.P. Inhibition of bFGF and EGF-induced proliferation of 3T3 fibroblasts by extract of Pygeum africanum (Tadenan®). Biomed Pharmacother 1994, 48 (Suppl. 1), 43s–47s. [Google Scholar]
- Penna, G.; Fibbi, B.; Amuchastegui, S.; Cossetti, C.; Aquilano, F.; Laverny, G.; Gacci, M.; Crescioli, C.; Maggi, M.; Adorini, L. Human benign prostatic hyperplasia stromal cells as inducers and targets of chronic immuno-mediated inflammation. J Immunol 2009, 182, 4056–4064. [Google Scholar] [CrossRef]
- Kramer, G.; Marberger, M. Could inflammation be a key component in the progression of benign prostatic hyperplasia? Curr Opin Urol 2006, 16, 25–29. [Google Scholar] [CrossRef]
- Szolnoki, E.; Reichart, E.; Marchal, S.; Szegedi, G. The effect of Pygeum africanum on fibroblast growth factor (FGF) and transforming growth factor beta (TGF beta 1/LAP) expression in animal model. Acta Microbiol Immunol Hung 2001, 48, 1–9. [Google Scholar] [CrossRef]
- Sidoti, C.; Hedef, N.; Delacroix, D.; Cavet, A.; Dupontt, C.H.; Borgeat, H. Inhibitory effect of Pygeum africanum extract (Tadenan®) on A23187-stimulated lipoxygenase metabolite production from human polymorphonuclear cells. The Pharmacologist 1993, 35, 196. [Google Scholar]
- United States Pharmacopeia. Dietary Supplement Monographs, Pygeum Extract. USP-NF. Rockville, MD: United States Pharmacopeia, 2023. [CrossRef]
Figure 1.
Changes in cytokines IL-1β, IL-2, IL-4, IL-5, IL-6, IL-9, IL-13, IL-22, IL-27, and TNF-α levels (pg/mL) after 24 hours exposure to PABE at different concentrations (500.0 to 2.0 µg/mL) (panels from left to right and from up to down). PABE, Pygeum africanum bark extract.
Figure 1.
Changes in cytokines IL-1β, IL-2, IL-4, IL-5, IL-6, IL-9, IL-13, IL-22, IL-27, and TNF-α levels (pg/mL) after 24 hours exposure to PABE at different concentrations (500.0 to 2.0 µg/mL) (panels from left to right and from up to down). PABE, Pygeum africanum bark extract.
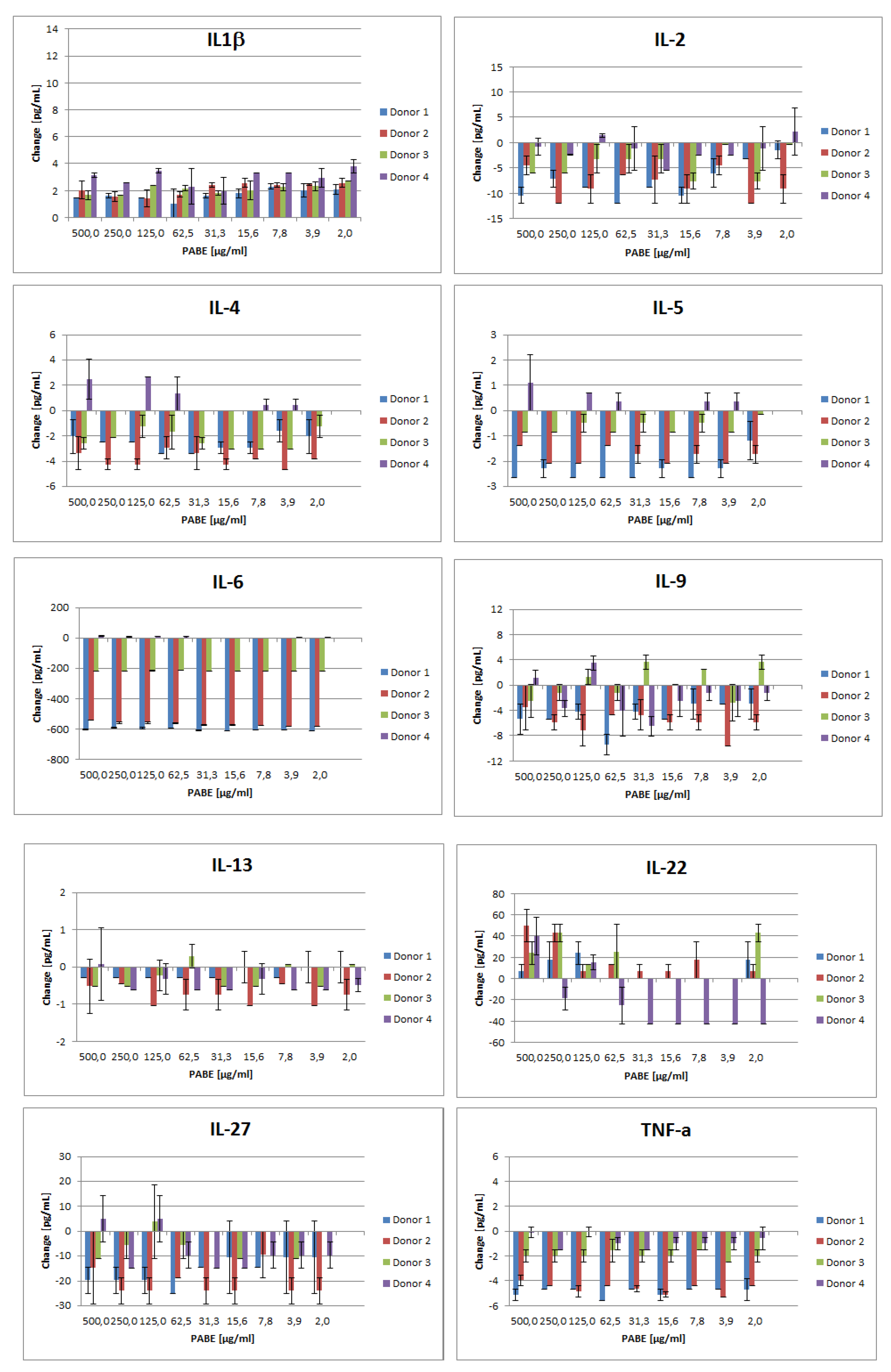
Figure 2.
Viability of PBMCs (% concentration) after 24 hours exposure to PABE (µg/mL) in each of the four donors.
Figure 2.
Viability of PBMCs (% concentration) after 24 hours exposure to PABE (µg/mL) in each of the four donors.
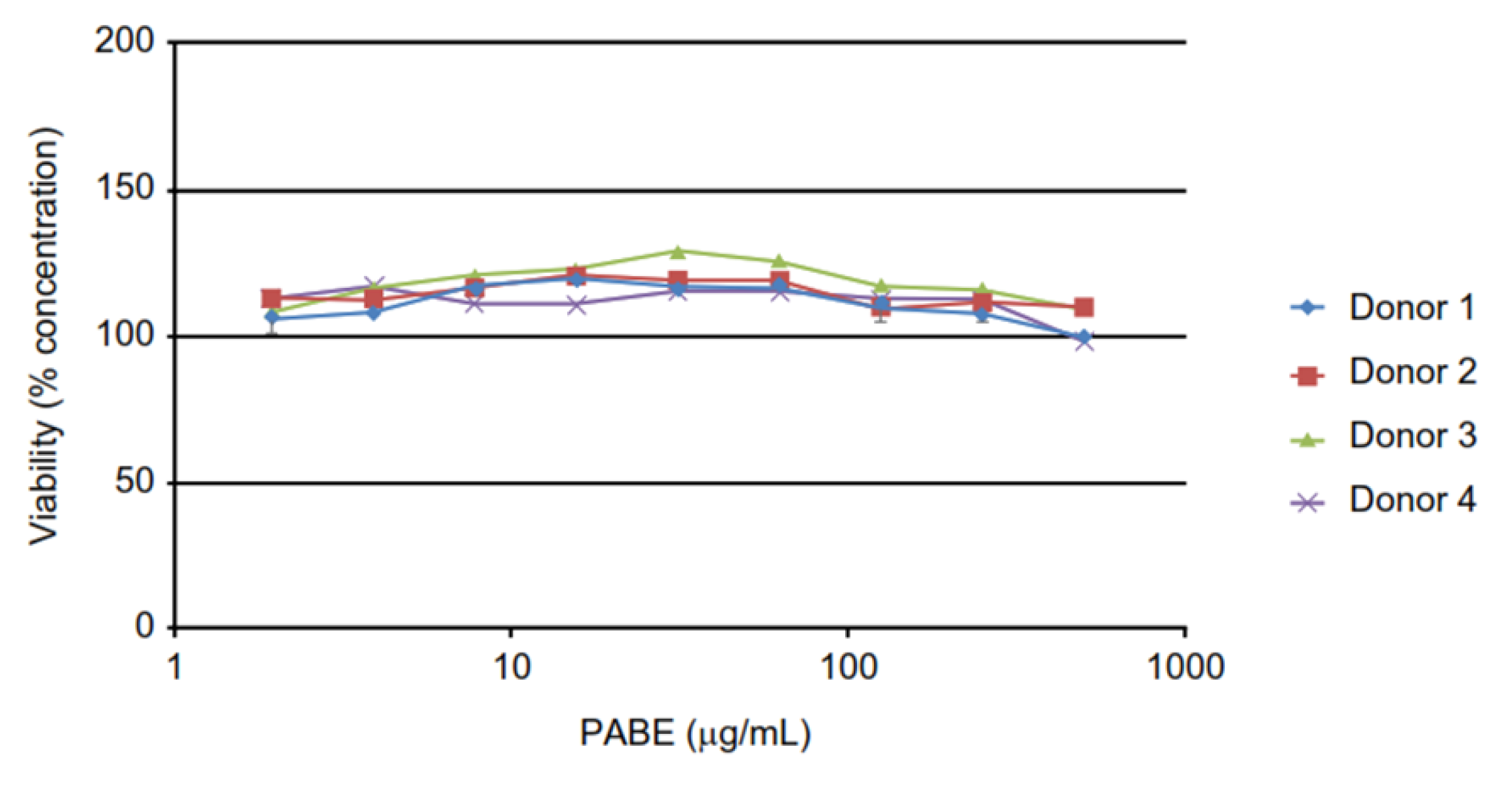
Figure 3.
Gas chromatography–flame ionization detector (GC–FID) chromatogram relative to Pygeum africanum extract Prunera®. Peaks in order of elution: Lauric Acid, Myristic Acid, Palmitic Acid, Linoleic Acid, Oleic Acid, Steric Acid, Eicosanol, Arachidic Acid, Docosanol, Behenic Acid, Tetracosanol, Lignoceric Acid, Octacosanol, Campesterol, Campestanol, Stigmasterol, beta-Sitosterol, Stigmastanol, Delta-7-Stigmastenol, Oleanolic Acid, Ursolic Acid, Docosyl Ferulate, beta-Sitosterol Glucoside.
Figure 3.
Gas chromatography–flame ionization detector (GC–FID) chromatogram relative to Pygeum africanum extract Prunera®. Peaks in order of elution: Lauric Acid, Myristic Acid, Palmitic Acid, Linoleic Acid, Oleic Acid, Steric Acid, Eicosanol, Arachidic Acid, Docosanol, Behenic Acid, Tetracosanol, Lignoceric Acid, Octacosanol, Campesterol, Campestanol, Stigmasterol, beta-Sitosterol, Stigmastanol, Delta-7-Stigmastenol, Oleanolic Acid, Ursolic Acid, Docosyl Ferulate, beta-Sitosterol Glucoside.
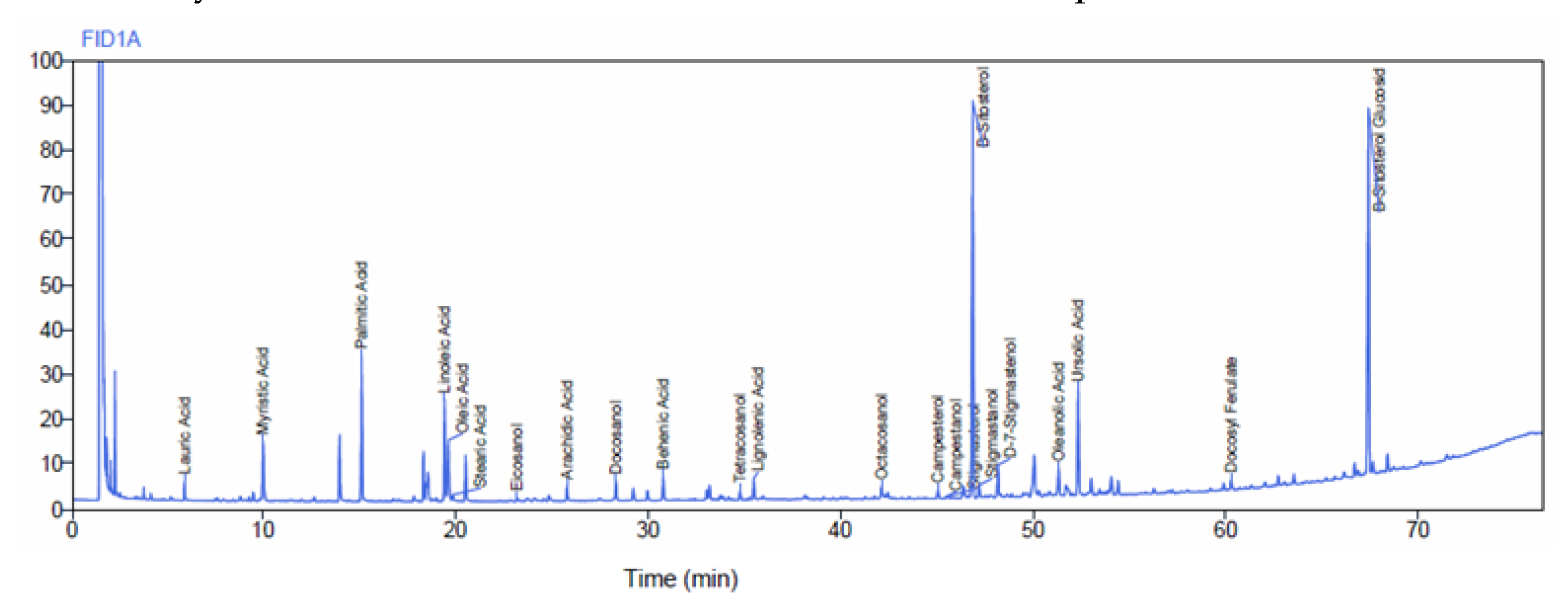

Table 1.
Mean values of cytokines in the four donors.
| Cytokines pg/mL |
Donor 1 | Donor 2 | Donor 3 | Donor 4 |
|---|---|---|---|---|
| IL-1β | 15.43 (2.81) | 19.08 (2.90) | 18.95 (1.98) | 26.79 (3.86) |
| IL-2 | -68.67 (9.34) | -74.08 (16.70) | -38.14 (11.54) | -12.53 (15.33) |
| IL-4 | -21.15 (3.13) | -30.86 (4.92) | -19.36 (3.13) | 7.41 (3.83) |
| IL-5 | -21.20 (1.80) | -16.20 (1.05) | -5.90 (1.05) | 2.85 (2.15) |
| IL-6 | -5439.97 (18.48) | -5109.79 (18.09) | -1972.39 (6.58) | 31.83 (13.65) |
| IL-9 | -42.65 (11.22) | -53.08 (13.19) | 3.24 (11.49) | -16.84 (16.54) |
| IL-13 | -1.76 (1.25) | -6.71 (1.98) | -2.36 (0.73) | -4.15 (1.98) |
| IL-22 | 65.52 (52.03) | 150.29 (67.43) | 148.46 (52.11) | -202.86 (53.73) |
| IL-27 | -144.31 (60.04) | -186.46 (56.07) | -40.19 (25.36) | -73.58 (40.03) |
| TNF-α | -43.92 (1.86) | -41.39 (1.38) | -17.48 (3.77) | -7.70 (3.65) |
Table 2.
Mechanism of action of cytokines selected for the study.
| Cytokine | Characteristics |
| IL-1β | Pro-inflammatory cytokine expressed by monocytes, macrophages, and dendritic cells, synthesized in response to inflammatory stimuli (e.g. pathogens). |
| IL-2 | Plays a central role in activation and proliferation of lymphocytes that have been primed by antigens; pivotal for expansion of most T-cells, natural killer [NK] cells, and B-cells during various phases of the immune response- |
| IL-4 | Anti-inflammatory cytokine, secreted by activated T-cells and NK cells. |
| IL-5 | Acts as hematopoietic growth factor, promoting proliferation, activation, and differentiation of certain bone marrow cells. |
| IL-6 | Pleiotropic cytokine produced mainly by monocytes; plays a central role in host defense mechanisms. |
| IL-9 | Pro-inflammatory cytokine having pleiotropic effects on Th2 lymphocytes and B-cells and mast cells; implicated in asthma and other allergies. |
| IL-13 | Pleiotropic cytokine expressed by activated T-helper, T-suppressor and NK cells; suppresses macrophage cytotoxic activity and inflammatory cytokine expression. |
| IL-22 | Regulates production of acute phase proteins of the immunological response; involved in the inflammatory immune response. |
| TNF-α | Pleiotropic cytokine that plays key roles in innate and adaptive immunity; most often associated with regulation of pro-inflammatory properties. |
Table 3.
Data of four donors from whom PBMCs were obtained.
| Sample identifier |
Gender | Aye, years | Ethnicity/race | ABO group/Rh | Antigen |
| HHU20130318 | Male | 38 | Asian | A/positive | SFC |
| HHU20130715 | Male | 41 | Hispanic | 0/positive | SFC |
| HHU20150826 | Male | 49 | Caucasian | Unknown | SFC |
| HHU20150910 | Female | 23 | African American | Unknown | SFC |
PBMCs: peripheral blood mononuclear cells; SFC: spot forming cells, PBMCs tested in ImmunoSpot® IFN-γ ELISPOT assay system 400,000 cells/well.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



