
Submitted:
07 June 2024
Posted:
07 June 2024
You are already at the latest version
Abstract
Abstract: To obtain bovine oocytes in high quality, effects of vitamin C (VC) on in vitro maturation (IVM) of bovine oocytes and early embryo development were investigated. The results showed that: 1) The IVM medium containing 50 µg/mL VC improved the oocyte maturation rate, while no effects on the development of parthenogenetic embryos; 2) The in vitro culture (IVC) medium containing 20 µg/mL VC increased the cleavage rate of the in vitro fertilized (IVF) embryos, enhanced the mRNA levels of pluripotency gene Oct4, Sox2, Cdx2 and Nanog in the blastocysts, but no effect on the blastocyst rate; 3) Combing supplemention of 50 µg/mL in IVM medium + 20 µg/ml VC in IVC medium (VC 50/20) increased the cleavage rate of IVF embryos and enhanced the mRNA levels of Oct4, Sox2, Cdx2 and Nanog in the blastocysts; 4) Combination of VC 0/20 and VC 50/20 enhanced the expression of anti-apoptotic gene Bcl-2 and VC 50/0 weakened the expression of pro-apoptotic gene Bax, while VC 0/40 and VC 0/60 increased Bax expression and decreased the Bcl-2/Bax ratio in blastocysts. Together, VC supplementation at tuned concentration as well its combination in IVM and IVC media improves the developmental rates of bovine oocytes and embryos.
Keywords:
cattle
; embryo
; in vitro maturation
; oocyte
; vitamin C (VC)
1. Introduction
In vitro maturation (IVM) of oocyte and the subsequent early embryo development are two key steps in assisted reproductive technology, no matter for the treatment of human infertility; or for high-yield, top-quality and disease-resistant livestock breeding. In order to increase oocyte IVM rate and embryo development rate, great efforts have been made, and multiple parameters have been found involving in these processes [1,2]. However, the development rates obtained with these techniques are still limited, particularly in large domestic animal like cattle.
Vitamin C (VC, also known as Ascorbic Acid/L-Ascorbic Acid) is an essential nutrient that must be obtained through the diet in adequate amounts to prevent hypovitaminosis C, deficiency and its consequences-including the potentially fatal deficiency disease scurvy. As a water-soluble molecule, VC is well known for its multiple functions such as promotion of wound healing, detoxification, alleviation of diabetes and neurogenic/synaptogenic disease, epigenetic regulation, serving as a cofactor for various enzymes, cytotoxicity against tumor/cancer cells, and antioxidant activity [3,4]. Among these functions, VC fulfils its roles in collagen synthesis, vasculogenesis, aging, cell proliferation, and differentiation by donating electrons in various enzymatic and non-enzymatic reactions [5]. Recently, it’s demonstrated that treatment with VC improved the development of mouse IVF embryo derived [6], and the supplements of VC as well other molecules seems to be also beneficial to the development of bovine embryos [7]. Later studies have shown that VC-treated bovine donor cells improved the development of cloned embryos by influencing embryonic transcription [8]. Based on its versatile function, VC improves the developmental competence of porcine oocytes [9] and enhances the survival of primary ovarian follicles cultured in vitro [10]. However, the appropriate concentration of VC still needs to be tuned for bovine oocyte IVM and the subsequent embryo development. Additionally, the effects of combined application of VC both in oocyte IVM and embryo culture of cattle are unclear.
To address the questions mentioned above, here we investigated the effects of VC and its combinations in IVM medium and IVC medium on bovine oocyte IVM and early embryonic development, aiming to improve the efficiency of bovine embryo IVC and procure high-quality embryos.
2. Materials and Methods
2.1. Bovine Ovaries and Chemicals
Bovine ovaries were collected from a local abattoir (Changchun Jixing meat industry Co., LTD, Changchun, China) and transported to the laboratory in 0.9% NaCl supplemented with 200 IU/mL penicillin at 36–37.5°C within 2 h. All experimental materials and procedures used here received endorsement from the Animal Welfare and Research Ethics Committee at Jilin University (SY201903002). Unless stated otherwise, chemicals and reagents were all procured from Sigma-Aldrich and the basic medium was acquired from Gibco (CA, USA).
2.2. In Vitro Maturation (IVM)
The follicles with a diameter of 2–8 mm were selected, and the oocytes were aspired using a 10 mL syringe with a 12-gauge needle. The cumulus-oocyte complexes (COCs) containing intact and compact cumulus cells were selected under a stereomicroscope. The COCs were washed three times with maturation solution [TCM-199 enriched with 25 mmoL/L NaHCO3, 0.38 mmoL/L sodium pyruvate, 10 μg follicle stimulating hormone (FSH)/mL, 10 μg luteinizing hormone (LH)/mL, 1 μg 17β-estradiol (E2)/mL and 10% (v/v) fetal bovine serum (FBS)]. Next, the COCs were transferred into 100 µL maturing solution microdroplets (10-15 pieces/droplet), and the IVM was conducted at 38.5℃, 5% CO2 and saturated humidity for 20–22 h. VC at concentrations of 0, 25, 50, 100 µg/mL was added into the maturing solution based on our preliminary experiments. The maturation rate was calculated and statistically analyzed after the oocytes being cultured for 23–24 h. After IVM, the oocyte with the first polar body, an homogeneous cytoplasm and at least three layers of cumulus cells, was considered to be a mature oocyte with high morphological quality.
2.3. Parthenogenetic Activation
The cumulus cells of the IVM oocytes were removed by the following steps. Firstly, they were digested for 2–3 min in the operating medium (15 mmol/L Hepes, 5 mmol/L NaHCO3, and 3 mg/mL of bovine serum albumin) containing 0.2% (w/v) hyaluronidase. Post-digestion, the cumulus cells were repeatedly and gently blown away with a 200 µL pipette, and the oocytes were washed three times with the operating medium. After collection of the mature oocytes, parthenogenesis is activated by a combination treatment of ionomycin and 6-dimethylaminopurine (6-DMAP). Briefly, the oocytes were firstly incubated with operating solution containing 5 µmol/L ionomycin for 5 min. Next, the eggs were transferred into the modified synthetic oviductal fluid (mSOF) droplets enriched with 2 mmol/L 6-DMAP, and cultured at 38.5 ℃, 5% CO2 and saturated humidity for 4 h. The mSOF consists of 108 mM NaCl, 7.2 mM KCl, 0.3 mM KH2PO4, 5 mM NaHCO3, 3.3 mM sodium lactate, 0.07 mM kanamycin monosulfate, 0.33 mM pyruvate, 1.7 mM CaCl2·2H2O, 0.3% (w/v) fatty acid-free bovine albumin, 1% (v/v) non-essential amino acid, and 2% (v/v) essential amino acid. The activated oocytes were subsequently transferred into new mSOF droplets covered with mineral oil and continuously cultivated under the conditions mentioned above. The culture medium containing VC (0, 25, 50, 100 µg/mL) was changed every 48 h. The cleavage and blastocyst rates were calculated at 2 d and 7 d respectively.
2.4. In Vitro Fertilization (IVF) and Embryo Culture
The matured oocytes obtained above were used for IVF by procedures as described [11]. Briefly, frozen bull semen was thawed in a water bath at 39.0℃ for 30–40 s. The semen was washed three times by centrifugation in pre-warmed Dulbecco's phosphate buffered saline (D-PBS). Subsequently, the sperm was washed in 5 mL TALP [Tyrode’s medium base, albumin, lactate and pyruvate, containing 0.67 mg NaCl/mL, 0.024 mg KCl/mL, 0.005 mg NaH2PO4/mL, 0.029 mg CaCl2·2H2O/mL, 0.01 mg MgCl2·6H2O/mL, 0.09 mg glucose/mL, 0.01 mg sodium pyruvate/mL, 0.19 μL sodium lactate (60%, v/v)/mL, 0.21 mg NaHCO3/mL, 0.3 mg BSA/mL], and incubated at 38.5 ℃, 5% CO2 and saturated humidity for 30 min. During sperm incubation, the COCs cultured for 23–24 h were washed in TALP for 3 times, transferred into 90 µL TALP droplets with 25–30 COCs per droplet, and continuously incubated at 38.5℃, 5% CO2 and saturated humidity.
Next, the sperm was washed two times by centrifugation at 1500 r/min for 5 min in TALP, and diluted with TALP to reach a final density of 2–4 ×106/mL. Subsequently, 10 µL sperm was added into 90 µL TALP droplets containing COCs, and the sperm-COCs were coincubated for 24 h.
After sperm-COCs coincubation, the putative zygotes were transferred into pre-warmed operating solution (500 µL). The remaining cumulus cells around the oocytes were removed by gentle pipetting repeatedly. The zygotes were washed 3 times with mSOF and transferred into new mSOF containing VC (0, 20, 40 and 60 µg/mL based on preliminary experiments). Generally, 25–30 zygotes were put into 100 µL mSOF with/without VC. The medium was replaced every 48 h. Similarly, the cleavage and blastocyst rates of each group were calculated at cultivation of 2 d and 7 d, respectively.
2.5. Blastocyst Cell Count
Four to five blastocysts from the IVF embryos cultured 7 d were selected for nuclear staining. Briefly, the embryos were washed 3 times with phosphate buffered saline (PBS)/PVA (0.01% polyvinyl alcohol in PBS). The zona pellucida of the blastocysts was removed with the acidic Tyrode’s solution (0.8 mg NaCl/mL, 0.02 mg KCl/mL, 0.024 mg CaCl2·2H2O/mL, 0.0047 mg MgCl2·6H2O/mL, 0.1 mg glucose/mL, and 0.4 mg polyvinylpyrrolidone (PVP)/mL). After washing 3 times with PBS/PVA, the embryos were stabilized using 3.7% paraformaldehyde at room temperature for 3 min, washed 3 times with PBS/PVA, and stained with Hoechst33342 (10 µg/mL) for 10 min in darkness. Finally, the blastocysts were washed 3 times with PBS/PVA, mounted between a cover slip and a glass slide supported by four columns of a mixture of petroleum jelly and paraffin (9:1) and immediately observed under a fluorescence microscope (Nikon, Tokyo, Japan). The number of blastomeres in each blastocyst was counted; the average cell numbers were quantified and statistically compared among groups.
2.6. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
The total RNAs of the blastocysts (approximately 25 embryos for each group) was extracted using the DNA/RNA Micro Kit (Qiagen, Germany). The RNA was reverse-transcribed into cDNA using TransScript® II One-Step gDNA Removal and cDNA Synthesis SuperMix AH311-03 Kit (Transgen Biotech, Beijing, China). The reverse transcription was performed in a 20 µL system, using following program: 95 ℃ for 30 s, 95 ℃ for 5 s, and 60 ℃ for 30 s, total 40 cycles, adhering to the protocols provided by the manufacturers. The primer sequences are detailed in Table 1 and GAPDH served as the reference gene to normalize the data. The 2−△△CT method was employed to analyze the relative abundance of mRNA transcripts.
2.7. Statistical Analysis
Data were presented as mean ± SEM and analyzed using IBM SPSS Statistics 19.0 software. P < 0.05 was considered as significant difference (*p < 0.05, **p < 0.01, ***p < 0.001). Replication of each experimental procedure was done three times to ensure reliability.
3. Results
3.1. Effects of VC on Oocyte IVM and Parthenogenetic Embryo Development
The morphology of bovine oocytes (immature, IVM 24 h, and mature) was shown in Figure 1 (A-C). The IVM rates of oocytes among four groups were compared, showing 50 µg/mL VC significantly improved the IVM rate of bovine oocytes (Figure 1D, p < 0.05). No statistical difference was observed for the cleavage and blastocyst rates of the parthenogenetic embryos among four groups (Figure 1E, F).
3.2. Effects of VC on IVF Embryo Development
Since IVF embryo might have different response to VC, its concentration gradient in IVC medium was tuned to 0, 20, 40 and 60 µg/mL according to our preliminary experiments. The results showed that VC at 20 µg/mL significantly increased the embryo cleavage rate (Figure 2A, P < 0.05). In terms of the blastocyst rate, there was no statistical difference among the groups (Figure 2B). Consistent with the cleavage rate, qRT-PCR analysis showed that the mRNA expressions of pluripotency-related gene Oct4, Sox2, Cdx2, and Nanog in blastocysts were remarkably enhanced by VC at 20 µg/mL (p < 0.05 or 0.001). Interestingly, supplementing VC at higher concentrations (40 and 60 µg/mL) didn’t show positive effects to the expressions of these genes except Nanog (Figure 2C).
3.3. Effects of VC Combination on IVF Embryo Development
Accordingly, IVM solution containing 50 µg/mL VC + IVC medium without/with 20 µg/mL VC (50/0 group and 50/20 group) was applied as a combined strategy to evaluate the effects of VC on the development of bovine IVF embryos, with none supplemented IVM solution + IVC medium (0/0 group) as the control. The results showed that the embryo cleavage rates were greatly increased both in 50/0 group and 50/20 group (Figure 3A). However, no significant difference of blastocyst formation and the cell numbers in blastocysts was observed among the groups (Figure 3B-D). Further qRT-PCR analysis showed that VC combination of 50/0 and 50/20 dramatically enhanced the transcriptional expression of Oct4, Cdx2, and Nanog, while only VC 50/20 boosted Sox2 expression, in bovine blastocysts (Figure 3E).
3.4. Effect of VC Combination on mRNA Expressions of Apoptotic Genes
Since apoptosis might occur during oocyte IVM and embryo development, we next investigated the expression of apoptosis-related gene Bcl-2 and Bax, and the changes of Bcl-2/Bax ratio, in IVF blastocysts derived by varied VC combinations in IVM solution + IVC medium (VC 0/0, VC 0/20, VC 0/40, VC 0/60, VC 50/0, VC 50/20, Figure 4). Compared with VC 0/0 group, VC 0/20 and VC 50/20 significantly elevated the expression levels of Bcl-2, while VC 0/40 and VC 0/60 dramatically increased Bax expression. However, Bax expression was greatly decreased in VC 50/0 group (Figure 4A). Furthermore, the ratios of Bcl-2/Bax were analyzed among these groups, showing increases in VC 0/20, VC 50/0 and VC 50/20 groups (all > VC 0/0 = 1, although not significantly), and markedly decreases in VC 0/40 and VC 0/60 groups (Figure 4B).
4. Discussion
Techniques of IVM, IVF, IVC and somatic cell cloned embryos have been successfully applied in research, production, animal breeding and medical fields. Oocyte IVM mainly includes nuclear maturation and cytoplasmic maturation. Many factors can affect oocyte IVM and subsequent embryonic development, such as female age and ovarian function, the temperature and time of ovarian transport, the method for COC aspiration, season, oocyte quality, and culture medium as well supplements, etc. Optimizing these factors is pivotal to procure high-quality oocytes and embryos for animal breeding. Since VC is an essential nutrient having multiple functions, here we evaluated the effects of VC at appropriate concentration as well its combination on bovine oocyte IVM and early embryo development.
Previously, in order to optimize embryo culture conditions, researchers applied varied supplements including VC to improve oocyte IVM, IVF rate and embryo development. It’s reported that VC improved the developmental ability of ovine and porcine oocytes and subsequent embryonic development [12,13]. However, its effects on bovine oocytes and early embryo development are somewhat controversial. Dalvit et al. reported that VC has no significant influence on the maturation of bovine oocytes [14], while Córdova et al. proved that after adding VC to IVM solution for 12 h in vitro, the subsequent development ability and blastocyst formation rate of bovine oocytes were significantly improved [15]. It’s also demonstrated that VC increases the cleavage rate of somatic cloned embryos, but has no positive effects on the blastocyst rate, and even negatively decreased the blastocyst rate and blastocyst quality [16]. We speculate that, for bovine oocyte IVM and embryo development, VC concentration needs be tuned appropriately. Based on preliminary experiments, we found that VC at 50 µg/mL (in gradients of 0, 25, 50, and 100 μg/mL) is significantly beneficial to bovine oocyte IVM, while VC at 20 µg/mL (in gradients of 0, 20, 40, and 60 μg/mL) is evidently helpful to improve the cleavage rate of bovine IVF embryos. Consistently, supplementing VC of 20 µg/mL in IVC medium markedly elevated the transcription levels of pluripotent gene Oct4, Sox2, Cdx2, and Nanog. Interestingly, adding VC at higher concentration into IVM solution (100 μg/mL) or IVC medium (40 and 60 μg/mL respectively) showed no positive effects on the development of both Parthenogenetic and IVF embryos in cattle. These results verify that our speculation is reasonable and practical. VC at tuned concentration might also alleviate cytotoxicity during oocyte IVM as reported in yak [17].
In order to achieve a better effect, IVM solution containing 50 μg/mL VC + IVC medium containing 20 μg/mL VC (VC 50/20 combination) was employed. Consistently, both VC 50/0 and VC 50/20 showed statistical positive role on the cleavage rate and slight increasing effect of the blastocyst rate. These combination effects were further confirmed by the up-regulated expressions of Oct4, Sox2, Cdx2, and Nanog in the IVF blastocysts. It’s well documented that these pluripotency-related genes are important factors in maintaining the proliferation and differentiation of bovine embryonic stem cells and induced pluripotent stem cells [18,19,20,21]. Thus, appropriate combination of VC might enhance the developmental capacity by regulating endogenous pluripotency factors.
In the process of embryo IVC, cell apoptosis might occur in its early development due to changes in the culture environment [22]. It’s reported that cell apoptosis mainly occurs at the stage of 9-16 cells and compacted morula [23]. Therefore, we next asked whether the effects of VC combinations are related to their anti-apoptosis role in bovine embryos. Our data indicate that VC 0/20 and VC 50/20 indeed boost anti-apoptotic gene Bcl-2 but inhibit pro-apoptotic gene Bax. The factors leading to embryo damage are variable, sometimes even including electromagnetic waves. Recently, it’s demonstrated that an in ovo VC injection reduces the harmful effects of electromagnetic waves emitted from mobile phones on chicken embryos [24]. Commonly, exposure to radiation and other detrimental factors increases the concentration of free radicals in embryos and causes oxidative stress damage. Our results and the above report suggest that VC and its combination prevent embryo damage and death possibly by anti-apoptosis and anti-oxidation activities.
5. Conclusions
Obtaining high quality oocytes is pivotal for animal breeding, particularly for large livestock like cattle. In the present study, the effects of VC on bovine oocyte IVM and early embryo development were evaluated. We found that supplementation of VC at tuned concentration into IVM solution and IVC medium as well as its combination application is beneficial to improve the development rates of bovine embryos. Additional experiments may be needed to reveal the underlying mechanism. Looking from a future perspective, the results of the present study may be of practical importance in the IVM and IVF of bovine oocytes as well the subsequent embryo IVC.
Author Contributions
Y.W. and A.W.: experimental design, data acquisition and analysis, manuscript writing. H.L.: data analysis. R.Y. and B.Z.: data acquisition. B.T. and Z.L.: fund raising. X.Z.: study design, analysis, manuscript writing and fund raising. All authors approved the final version of the manuscript.
Funding
This research was funded by National Natural Science Foundation of China (No. 32172803) and the Natural Science Foundation of Jilin Province, China (No. 20240101255JC).
Institutional Review Board Statement
The animal study protocol was approved by the Jilin University Institutional Animal Care and Use Committee for use of animals/tissues (SY201903002).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding author, upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Luciano, A.M.; Franciosi, F.; Barros, R.G.; Dieci, C.; Lodde, V. The variable success of in vitro maturation: can we do better? Anim Reprod 2018, 15, 727–736. [Google Scholar] [CrossRef] [PubMed]
- Strączyńska, P.; Papis, K.; Morawiec, E.; Czerwiński, M.; Gajewski, Z.; Olejek, A.; Bednarska-Czerwińska, A. Signaling mechanisms and their regulation during in vivo or in vitro maturation of mammalian oocytes. Reprod Biol Endocrinol 2022, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Abdollahifar, M.A.; Azad, N.; Sajadi, E.; Mofarahe, Z.S.; Zare, F.; Moradi, A.; Rezaee, F.; Gholamin, M.; Abdi, S. Vitamin C restores ovarian follicular reservation in a mouse model of aging. Anat Cell Biol 2019, 52, 196–203. [Google Scholar] [CrossRef] [PubMed]
- Lykkesfeldt, J.; Michels, A.J.; Frei, B. Vitamin C. Adv Nutr 2014, 5, 16–18. [Google Scholar] [CrossRef] [PubMed]
- Travica, N.; Ried, K.; Sali, A.; Scholey, A.; Hudson, I.; Pipingas, A. Vitamin C status and cognitive function: A systematic review. Nutrients 2017, 9, 960. [Google Scholar] [CrossRef]
- Navid, S.; Saadatian, Z.; Talebi, A. Assessment of developmental rate of mouse embryos yielded from in vitro fertilization of the oocyte with treatment of melatonin and vitamin C simultaneously. BMC Womens Health 2023, 23, 525. [Google Scholar] [CrossRef]
- Sovernigo, T.C.; Adona, P.R.; Monzani, P.S.; Guemra, S.; Barros, F.; Lopes, F.G.; Leal, C. Effects of supplementation of medium with different antioxidants during in vitro maturation of bovine oocytes on subsequent embryo production. Reprod Domest Anim 2017, 52, 561–569. [Google Scholar] [CrossRef]
- Zhang, L.; Zhang, Y.; Han, Z.; Fang, J.; Chen, H.; Guo, Z. Transcriptome analyses reveal effects of Vitamin C-treated donor cells on cloned bovine embryo development. Int J Mol Sci 2019, 20, 2628. [Google Scholar] [CrossRef]
- Kere, M.; Siriboon, C.; Lo, N.W.; Nguyen, N.T.; Ju, J.C. Ascorbic acid improves the developmental competence of porcine oocytes after parthenogenetic activation and somatic cell nuclear transplantation. J Reprod Dev 2013, 59, 78–84. [Google Scholar] [CrossRef]
- Tagler, D.; Makanji, Y.; Tu, T.; Bernabé, B.P.; Lee, R.; Zhu, J.; Kniazeva, E.; Hornick, J.E.; Woodruff, T.K.; Shea, L.D. Promoting extracellular matrix remodeling via ascorbic acid enhances the survival of primary ovarian follicles encapsulated in alginate hydrogels. Biotechnol Bioeng 2014, 111, 1417–1429. [Google Scholar] [CrossRef]
- Zhang, J.; Hao, L.; Wei, Q.; Zhang, S.; Cheng, H.; Zhai, Y.; Jiang, Y.; An, X.; Li, Z.; Zhang, X.; Tang, B. TET3 overexpression facilitates DNA reprogramming and early development of bovine SCNT embryos. Reproduction 2020, 160, 379–391. [Google Scholar] [CrossRef]
- Tatemoto, H.; Ootaki, K.; Shigeta, K.; Muto, N. Enhancement of developmental competence after in vitro fertilization of porcine oocytes by treatment with ascorbic acid 2-O-alpha-glucoside during in vitro maturation. Biol Reprod 2001, 65, 1800–1806. [Google Scholar] [CrossRef]
- Karimian, M.; Zandi, M.; Sanjabi, M.R.; Masoumian, M.; Ofoghi, H. Effects of grape seed extract, quercetin and vitamin C on ovine oocyte maturation and subsequent embryonic development. Cell Mol Biol 2018, 64, 98–102. [Google Scholar] [CrossRef]
- Dalvit, G.; Llanes, S.P.; Descalzo, A.; Insani, M.; Beconi, M.; Cetica, P. Effect of alpha-tocopherol and ascorbic acid on bovine oocyte in vitro maturation. Reprod Domest Anim 2005, 40, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Córdova, B.; Morató, R.; Izquierdo, D.; Paramio, T.; Mogas, T. Effect of the addition of insulin-transferrin-selenium and/or L-ascorbic acid to the in vitro maturation of prepubertal bovine oocytes on cytoplasmic maturation and embryo development. Theriogenology 2010, 74, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, Y.S.; Wang, L.J.; Zhang, H.; Li, R.Z.; Cui, C.C.; Li, W.Z.; Zhang, Y.; Jin, Y.P. Vitamin C supplementation enhances compact morulae formation but reduces the hatching blastocyst rate of bovine somatic cell nuclear transfer embryos. Cell Reprogram 2014, 16, 290–297. [Google Scholar] [CrossRef]
- Li, Q.; Zhao, T.; He, H.; Robert, N.; Ding, T.; Hu, X.; Zhang, T.; Pan, Y.; Cui, Y.; Yu, S. Ascorbic acid protects the toxic effects of aflatoxin B1 on yak oocyte maturation. Anim Sci J 2022, 93, e13702. [Google Scholar] [CrossRef] [PubMed]
- Bogliotti, Y.S.; Wu, J.; Vilarino, M.; Okamura, D.; Soto, D.A.; Zhong, C.; Sakurai, M.; Sampaio, R.V.; Suzuki, K.; Izpisua Belmonte, J.C.; Ross, P.J. Efficient derivation of stable primed pluripotent embryonic stem cells from bovine blastocysts. Proc Natl Acad Sci U S A 2018, 115, 2090–2095. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; An, X.L.; Yu, H.; Cai, N.N.; Zhai, Y.H.; Li, Q.; Cheng, H.; Zhang, S.; Tang, B.; Li, Z.Y.; Zhang, X.M. Transcriptome profile of bovine iPSCs derived from Sertoli cells. Theriogenology 2020, 146, 120–132. [Google Scholar] [CrossRef]
- Jiang, Y.; Cai, N.N.; An, X.L.; Zhu, W.Q.; Yang, R.; Tang, B.; Li, Z.Y.; Zhang, X.M. Naïve-like conversion of bovine induced pluripotent stem cells from Sertoli cells. Theriogenology 2023, 196, 68–78. [Google Scholar] [CrossRef]
- Zhao, L.; Gao, X.; Zheng, Y.; Wang, Z.; Zhao, G.; Ren, J.; Zhang, J.; Wu, J.; Wu, B.; Chen, Y.; Sun, W.; Li, Y.; Su, J.; Ding, Y.; Gao, Y.; Liu, M.; Bai, X.; Sun, L.; Cao, G.; Tang, F.; Bao, S.; Liu, P.; Li, X. Establishment of bovine expanded potential stem cells. Proc Natl Acad Sci U S A 2021, 118, e2018505118. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Ibeas, P.; Gimeno, I.; Cañón-Beltrán, K.; Gutiérrez-Adán, A.; Rizos, D.; Gómez, E. Senescence and apoptosis during in vitro embryo development in a bovine model. Front Cell Dev Biol 2020, 8, 619902. [Google Scholar] [CrossRef] [PubMed]
- Antunes, G.; Chaveiro, A.; Santos, P.; Marques, A.; Jin, H.S.; Moreira da Silva, F. Influence of apoptosis in bovine embryo's development. Reprod Domest Anim 2010, 45, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Yenilmez, F. Effect of in ovo vitamin C injection against mobile phone radiation on post-hatch performance of broiler chicks. Vet Sci 2022, 9, 613. [Google Scholar] [CrossRef]
Figure 1.
Effects of VC on bovine oocyte IVM and parthenogenetic embryo development. (A-C): Oocyte morphology at different stages (Immature, IVM 24 h, and Mature, respectively) was shown with arrows showing the first polar bodies; (D): IVM rates (Oocyte number = 89, 78, 94 and 81 for different groups, respectively); (E, F): Cleavage and blastocyst rates of the parthenogenetic embryos (Oocyte number = 102, 96, 111, and 85, respectively). *P < 0.05; Scale bar = 50 μm.
Figure 1.
Effects of VC on bovine oocyte IVM and parthenogenetic embryo development. (A-C): Oocyte morphology at different stages (Immature, IVM 24 h, and Mature, respectively) was shown with arrows showing the first polar bodies; (D): IVM rates (Oocyte number = 89, 78, 94 and 81 for different groups, respectively); (E, F): Cleavage and blastocyst rates of the parthenogenetic embryos (Oocyte number = 102, 96, 111, and 85, respectively). *P < 0.05; Scale bar = 50 μm.
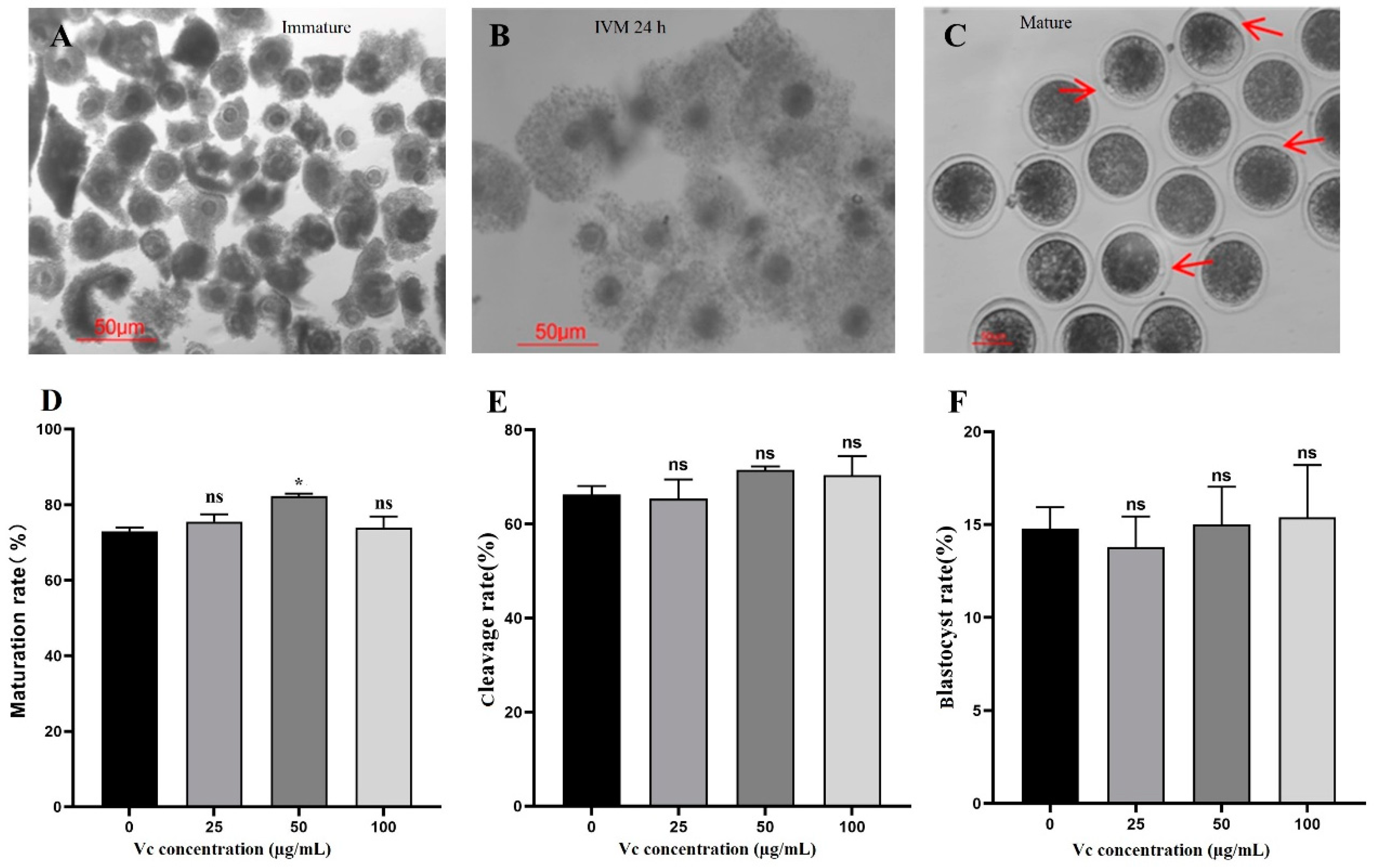
Figure 2.
Effects of VC on the development of bovine IVF embryos. (A, B): The cleavage and blastocyst rates of IVF embryos (Oocyte number = 96, 106, 93 and 100 for different groups, respectively); (C): Relative expressions of pluripotency-related gene Oct4, Sox2, Cdx2, and Nanog in IVF embryos. *p < 0.05, ***p < 0.001.
Figure 2.
Effects of VC on the development of bovine IVF embryos. (A, B): The cleavage and blastocyst rates of IVF embryos (Oocyte number = 96, 106, 93 and 100 for different groups, respectively); (C): Relative expressions of pluripotency-related gene Oct4, Sox2, Cdx2, and Nanog in IVF embryos. *p < 0.05, ***p < 0.001.
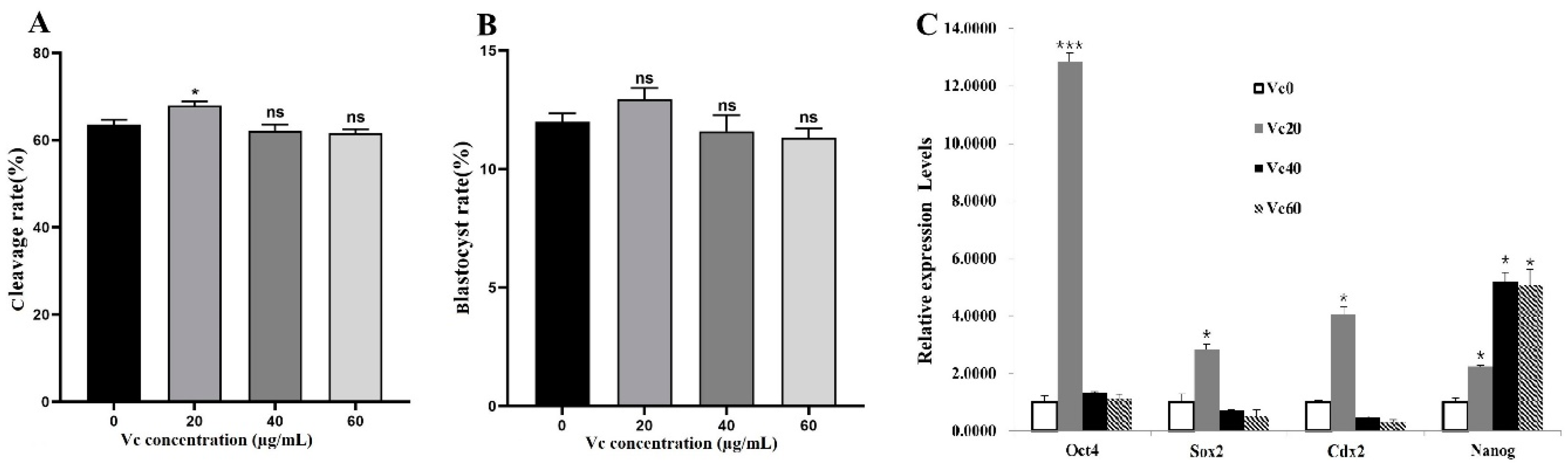
Figure 3.
Effects of VC combination on the development of bovine IVF embryo. (A-C): The cleavage and blastocyst rates as well as the blastomere numbers in IVF embryos (Oocyte number = 79, 88, and 95 for different groups, respectively); (D): Representative Hoechst33342 staining images of the IVF blastocysts (200×); (E): mRNA expression of Oct4, Sox2, Cdx2, and Nanog in IVF embryos. *p < 0.05, **p < 0.01, Scale bar = 50 μm.
Figure 3.
Effects of VC combination on the development of bovine IVF embryo. (A-C): The cleavage and blastocyst rates as well as the blastomere numbers in IVF embryos (Oocyte number = 79, 88, and 95 for different groups, respectively); (D): Representative Hoechst33342 staining images of the IVF blastocysts (200×); (E): mRNA expression of Oct4, Sox2, Cdx2, and Nanog in IVF embryos. *p < 0.05, **p < 0.01, Scale bar = 50 μm.
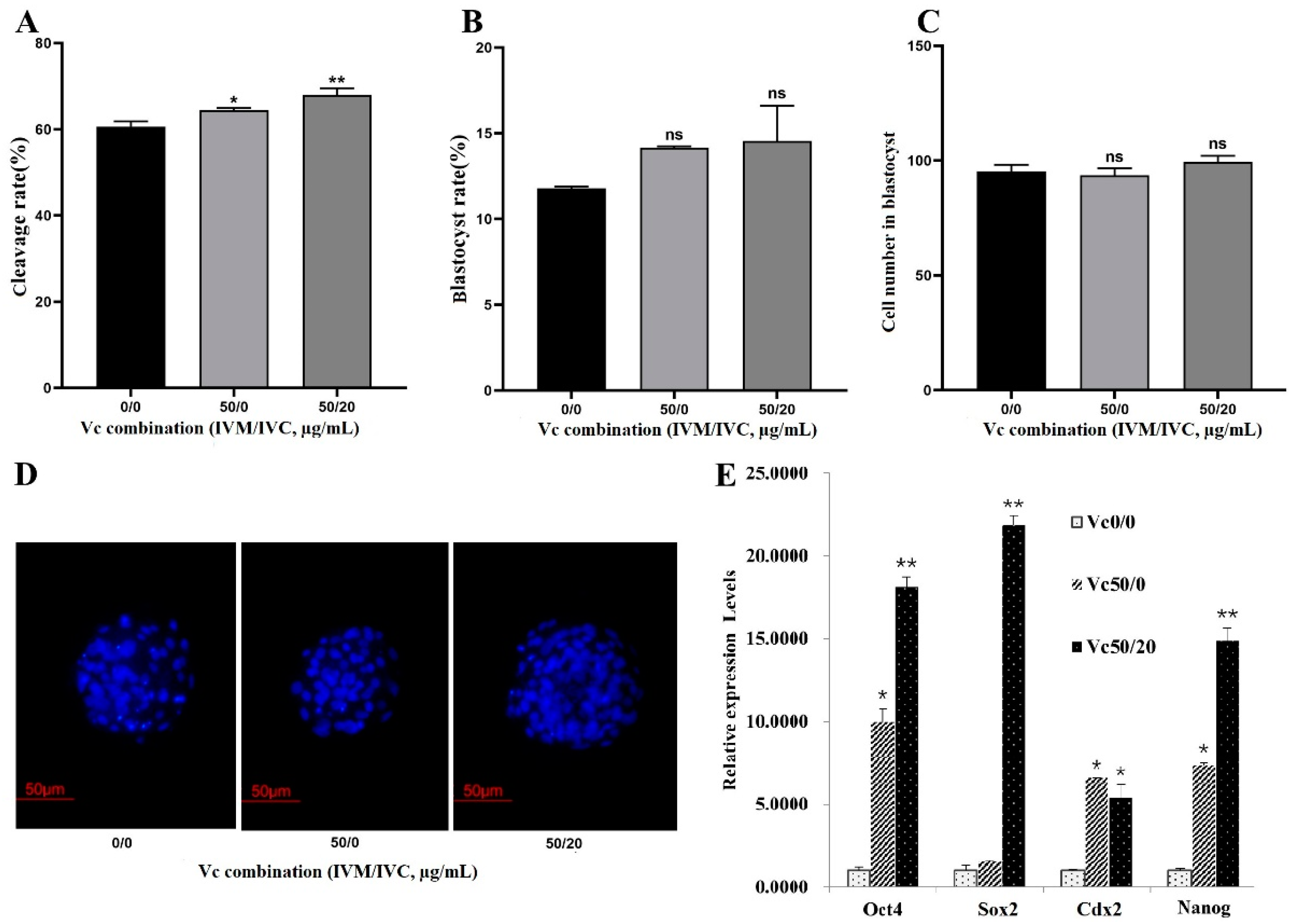
Figure 4.
Effect of Vc combinations on the mRNA expressions of apoptosis-related genes in IVF blastocysts. (A): mRNA levels of Bcl-2 and Bax in groups containing different Vc combinations in IVM solution + IVC medium (VC 0/0, VC 0/20, VC 0/40, VC 0/60, VC 50/0, VC 50/20). (B): mRNA expression ratios of Bcl-2/Bax. *P < 0.05.
Figure 4.
Effect of Vc combinations on the mRNA expressions of apoptosis-related genes in IVF blastocysts. (A): mRNA levels of Bcl-2 and Bax in groups containing different Vc combinations in IVM solution + IVC medium (VC 0/0, VC 0/20, VC 0/40, VC 0/60, VC 50/0, VC 50/20). (B): mRNA expression ratios of Bcl-2/Bax. *P < 0.05.
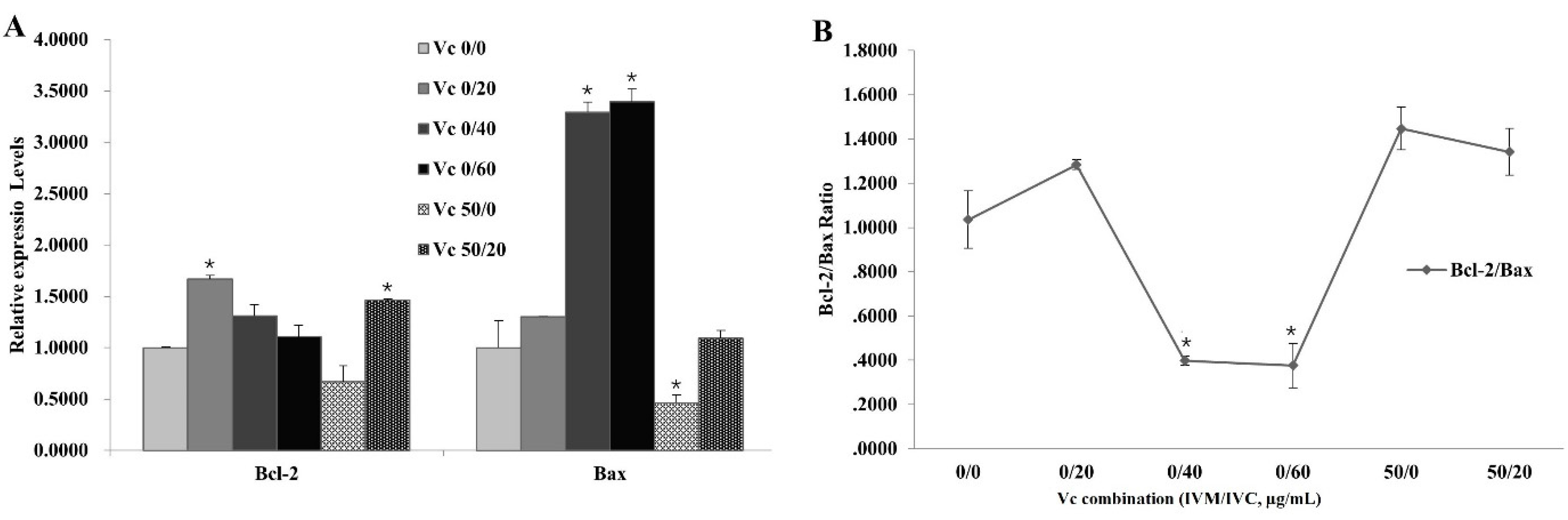

Table 1.
Table 1. Primers used for qRT-PCR.
| Gene | Forward Primer (5'-3') | Reverse Primer (5'-3') |
|---|---|---|
| GAPDH | CGGCACAGTCAAGGCAGAGAAC | CGGCACAGTCAAGGCAGAGAAC |
| Oct4 | GATTTGGATGAGTTTTTAAGGGTT | ACTCCAACTTCTCCTTATCCAACTT |
| Sox2 | CTATGACCAGCTCGCAGA | GGAAGAAGAGGTAACCACG |
| Cdx2 | CTTTCCTCCGGATGGTGATA | AGCCAAGTGAAAACCAGGAC |
| Nanog | AAACAACTGGCCGAAGGAATA | AGGAGTGGTTGVTCCAAGAC |
| Bcl-2 | GAGTCGGATCGCAACTTGGA | CTCTCGGCTGCTGCATTGT |
| Bax | GCGCATCGGAGATGAATTG | CCACAGCTGCGATCATCCT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



