
Submitted:
10 June 2024
Posted:
11 June 2024
You are already at the latest version
Abstract
Hormone changes across women's menstrual cycle may lead to changes in perception of chemical signals and hormonal responses to these cues. The aim of the present study was to determine the role of menstrual cycle phase on the effects of extracts of male axillary secretions (EMAS) in women. We tested healthy reproductive age and premenopausal women (n=29). EMAS/control solution was applied once in two hours either in the follicular or luteal phase, while collecting saliva samples for LH and cortisol monitoring. LH and cortisol concentrations were analyzed using EIA technique. PANAS and VAS were used to assess the mood. We observed a significant increase in the number of LH peaks (p=0.0447) and their amplitude (p=0.0469) when EMAS was applied in the follicular phase. The same application in the luteal phase lowered the amplitude of LH peaks (p=0.0382). EMAS application increases salivary cortisol level in reproductive age women relative to premenopausal women (p=0.0032). PANAS revealed changes in positive and negative affect after EMAS application. Our data indicate the significance of the menstrual cycle phase for EMAS effects on LH secretion, mood, but not on cortisol secretion in women.
Keywords:
olfactory communication
; axillary secretions
; chemical signal
; human
; menstrual cycle
; emotional state
; LH
; cortisol
1. Introduction
Body odors are natural components of the environment in a person’s daily life. The past three decades have generated a suite of discoveries supporting on the one hand the idea that human olfaction has been underestimated during practically whole time of its investigation [1] and, on the other hand, concerning the potential involvement of body odors in various aspects of interpersonal communication, such as familial bonding, mate selection and emotional state in human [2,3]. Besides, it has been shown that body odor may be processed differently in the brain than other odors [4]. The mechanisms underlying such a communication are poorly documented to date.
Human body odors may change hormonal status of women in general, or regulate women’s menstrual cycles in particular [5,6,7]. Apocrine glands in the axillae are currently considered to be the most likely source of human chemical signals, or at least axillary secretions contribute most to the appearance of the so-called “body odor” [8]. Preti and colleagues have shown that extracts of male axillary secretions may directly affect LH-pulsing and mood of women in the follicular phase of menstrual cycle [6]. It has been shown that extracts of male axillary secretions affect the length of the menstrual cycle in women with a history of irregular cycle length [5,9]. In premenopausal women effects of application of extracts of male axillary secretions are even more profound than in younger women. It was shown that idiopathic repeated pregnancy loss was associated with altered olfactory responses to body odor [7]. At the same time, hormone changes across the women’s menstrual cycle may lead to changes in perception of chemical signals or other odors and hormonal responses to these cues [10,11].
2. Materials and Methods
2.1. Participants
29 healthy heterosexual women with known medical history, not taking hormonal pills were enrolled in the protocol. Informed consent was obtained from all participants.
Women were divided into two groups: 18-45 y.o. later referred to as “reproductive age women” with mean age of 24.7 ± 5.9 years and with regular menstrual cycle 26-30 days long, and over 45 y.o., later referred to as “premenopausal women” with mean age of 46.4 ± 3.5 years. For each woman, we had information on the length and regularity of her menstrual cycles for 6 months preceding the experiment. Length of menstrual cycles was self-recorded by participants of the study; the beginning of menses was considered the first day of the menstrual cycle. Women in the follicular phase were tested on the 9th or 10th day of the menstrual cycle. Women in the luteal phase were tested 5.3 ± 3.3 days prior menses, which start was also documented.
2.2. Extract of Male Axillary Secretions (EMAS)
Axillary secretions were collected during four hours on sterile cotton pads (0.2 × 11 × 9 cm) from 6 healthy heterosexual men of 32.2 ± 4.6 years old using the standard technique [14,15]. For seven days prior to collection all odor donors refrained from using deodorant/antiperspirants and washed/bathed using only plain soap without fragrance. After collecting the secretions, the pads from all the donors were combined and soaked with 70% ethanol (15 ml per pad) (ethanol 96.3% purity, serial # 386710, Ferejn, Russia) for 1 hour; about 85% of ethanol extract was recovered. Finally, a masking fragrance (0.005% solution of 2-phenylethanol, CAS Number: 60-12-8, Sigma-Aldrich® Cat. # 77861, ≥99.0% purity) was added to the ethanol extract [15]. Clear 70% ethanol solution scented with the same fragrance was used as an odor-matched control sample. All solutions were stored at -40 °C in aliquots and brought to room temperature before testing. Female participants were unable to distinguish between scented ethanol extract of male axillary secretions (EMAS) and control solution by smell. Male donors had no contact and were not familiar with the female participants.
2.3. Evaluation of Emotional State of Test Subjects
We evaluated the current emotional state of test subjects using two questionnaires: Positive and Negative Affect Schedule (PANAS) [16] adapted for the Russian population [17] and Visual Analogue Scales (VAS) [18,19]. The questionnaires were printed to fill in by hand.
PANAS questionnaire consists of a list of 20 adjectives that describe different feelings and emotions. Subjects were asked to note on a five-point Likert scale how they feel with regard to each statement - from “ very slightly or not at all” (score 1) to “very much” (score 5). To evaluate total positive or negative affect we summed up the scores of 10 positive (interested, excited, strong, enthusiastic, proud, alert, inspired, determined, attentive, active) or, respectively, 10 negative (distressed, upset, guilty, scared, hostile, irritable, ashamed, nervous, jittery, afraid) adjectives.
VAS was presented for 8 adjectives describing the mood. They were: attuned to communication, sincere, relaxed, sad, focused, experiencing sexual arousal, energetic, nervous. Participants were instructed to place a mark on a 100 mm scale with anchors “not at all” and “extremely” at the ends in accordance with their current mood.
2.4. Procedure
A within-subject, double-blind, repeated-measures design was used in such a way that each participant underwent testing both with control solution and EMAS. EMAS or control solution (0.5 ml) were applied to a cotton pad and placed within 15 cm of the woman’s nose once every 2 hours either in the follicular or luteal phase of her menstrual cycle. Procedure continued for 8 hours (from 10 a.m. to 6 p.m.). Neither experimenter, nor participant were informed about the order in which solutions were applied. None of the women correctly guessed the true nature of the stimuli that were being delivered to their nasal region. Saliva samples were taken every 10 min for reproductive age women (total of 48 samples per woman) using Sali-Caps® sets (IBL, Germany).
Women completed VAS and PANAS four times during the experiment: 15 minutes after the first application of the control solution at 10 a.m., 15 minutes before and after first application of EMAS at 2 p.m., at the end of the experiment.
Changes in salivary cortisol concentration after EMAS application in a group of reproductive age women were evaluated relative to a group of premenopausal women. For all groups testing continued for 4 hours from 12 a.m. to 4 p.m. and saliva samples were taken every 20 minutes (total of 12 samples per woman). EMAS or control solution (0.5 ml) were applied to a cotton pad and placed within 15 cm of the woman’s nose once every two hours.
2.5. Hormone’s Measurements and Further Calculations
Saliva samples were stored at -32 °C, thawed at room temperature and centrifuged before testing. Concentration of luteinizing hormone (LH) was assessed using EIA technique (abx576540, Abbexa, UK) in all saliva samples. After completion of the immunoassay, the absorbance in the wells were measured at λ = 450 nm with SpectraMax 340PC 384 spectrophotometer (Molecular Devices, United States). Data analysis was performed with software Spectra-MaxSoftware.
LH baseline was calculated individually for each woman and separately for control and experimental periods [20,21,22]. To calculate the basal LH level, the mean value of the LH concentration for each woman was added to 1.5 of standard deviation (SD). Values that exceeded the result were pro tempore excluded, and the value of the mean + 1.5 SD was calculated for remaining values again. This procedure was repeated until none of the remaining values in the sample size exceeded the mean +1.5 SD. The received remaining value was considered as the basal hormone concentration for a particular woman.
We counted the number of LH peaks (values exceeding the basal level) and determined their mean amplitude separately during control and experimental periods. We also took into account individual differences in the form of LH peak for each woman and the fact that the number of salivary LH peaks cannot exceed previously reported number of LH pulses in blood (on average one peak per hour in follicular phase and one peak per 2-4 hours in luteal phase) [6,23] (Figure 1).
Cortisol was monitored using an EIA technique (EIA Can-C-290, DBC, Canada). As cortisol secretion is cyclical in nature, we calculated the average value for two hours during control period and for two hours during EMAS application to catch the peaks that occur about every 90 min [24], we used samples that were collected from 12:20 p.m. to 3:40 p.m. from reproductive age women and premenopausal women. After completion of the immunoassay, the absorbance in the wells were obtained at λ = 450 nm with SpectraMax 340PC 384 spectrophotometer (Molecular Devices, United States). Data analysis was performed with software SpectraMaxSoftware.
2.6. Statistical Analyses
Data analysis was carried out with STATISTICA 8.0 software (StatSoft, Inc.) and with R (version 4.3.0).
All data are reported as mean ± SD. All data were checked to fit in normal distribution (Shapiro-Wilk test, p > 0.05) and to be homoscedastic (Levene’s test, p > 0.05) before performing Repeated Measures ANOVA or One-way ANOVA, otherwise we used data transformation prior analysis, non-parametric Wilcoxon Signed-Ranks test or permutation test with Monte-Carlo approximation in R (package lmPerm (https://cran.r-project.org/web/packages/lmPerm/lmPerm.pdf) function aovp(); and package rcompanion (https://cran.r-project.org/web/packages/rcompanion/rcompanion.pdf) function pairwisePermutationTest()). The alpha level for all analysis was set a priori to 0.05.
Analysis of variance for repeated measurements with the within-subject factor treatment (control solution vs. EMAS) and the between-subject factor phase (follicular vs. luteal) was used for comparison of the number of LH peaks.
LH amplitude data did not fit in normal distribution, that is why we counted percent of change between mean amplitude during control period and mean amplitude after beginning of EMAS application for each test subject, then we performed a One-way ANOVA followed by Fisher HSD test.
Cortisol data also did not fit in normal distribution, that is why we counted percent of change between mean cortisol level during control period and mean cortisol level during EMAS application for each test subject, then we performed permutation test with Monte-Carlo approximation in R.
3. Results
3.1. The Parameters of LH Peaks
We found that the frequency of LH peaks was affected by the phase of the menstrual cycle of test subjects (RM ANOVA, F(1, 17) = 45.37, p = 0.0000, n = 19). In particular, post-hoc analysis showed that the number of LH peaks depended on the phase of menstrual cycle after application of both control and EMAS solutions (p = 0.0002 and p = 0.0000, respectively, n = 19, post-hoc Fisher LSD test) (Figure 2).
Besides, we observed significant increase in the number of salivary LH peaks when EMAS was applied to women in follicular phase of menstrual cycle compared to the control (p = 0.0447, n = 10, post-hoc Fisher LSD test) (Figure 3), which indicates a reduction in the time interval between adjacent peaks, i.e., an increase in the pulsation of LH. While in a group of women in the luteal phase of their menstrual cycle the frequency of LH peaks did not change significantly after EMAS application (p = 0.8379, n = 9, post-hoc Fisher LSD test).
For the best of our knowledge this study is the first to show the rise of the average amplitude of LH peaks in a group of women in the follicular phase of the menstrual cycle (p = 0.0469, n = 10, Wilcoxon Signed-Ranks test). We are also the first to show the opposite effect of EMAS application on the average amplitude of LH peaks for the group of women in the luteal phase (p = 0.0382, n = 9, Wilcoxon Signed-Ranks test) (Figure 4).
3.2. Salivary cortisol
The change of salivary cortisol in reproductive age women didn’t depend on the phase of the menstrual cycle (RM ANOVA F(1, 17) = 0.92, p = 0.3516, n = 19). However, when comparing groups of reproductive age women and premenopausal women, we found that the change of salivary cortisol after application to EMAS depended on age group of test subjects (permutation test with Monte-Carlo approximation, p = 0.0032, n = 29) (Table 1).
We found that salivary cortisol increased by an average of 15.1% in a group of reproductive age women, and decreased by an average of 25% in a group of premenopausal women after application of EMAS (Figure 5). It should be noted that salivary cortisol values were within the normal range during the entire experiment, which suggests a stimulating effect of EMAS rather than a stressful one.
The number of test subjects reacted with an increase in the level of salivary cortisol after application of EMAS was significantly higher than it could be expected according the natural diurnal changes (follicular phase: p = 0.0269, n = 10, χ2; luteal phase: p = 0.0196, n = 9, χ2). In the group of premenopausal women, the number of test subjects who responded with an increase of salivary cortisol after EMAS application was within the expected level according to diurnal rhythm (p = 0.3428, n = 10, χ2) [24], in general, we observed a decrease in the salivary cortisol in this group.
3.3. Emotional State
Repeated measures ANOVA didn’t reveal significant changes in positive affect 15 min after the beginning of EMAS application neither in follicular nor in luteal phase (RM ANOVA, between-subject factor phase: F(1; 17) = 0.1236, p = 0.7294; within-subject factor treatment: F(1; 17) = 0.9082, p = 0.3540). At the same time, we observed a decrease in positive affect for the entire time (4 hours) of EMAS application (RM ANOVA, between-subject factor phase: F(1; 17) = 0.8690, p = 0.3643; within-subject factor treatment: F(1; 17) = 6.2821, p = 0.0227), which may also represent general fatigue of test subjects by the end of 8 hours testing.
We didn’t observe any changes in negative affect 15 min after the beginning of application to EMAS compared to control in women neither in follicular nor in luteal phase (follicular phase: p = 0.4652, n = 10, Wilcoxon Signed-Ranks test; luteal phase: p = 0.4227, n = 9, Wilcoxon Signed-Ranks test). We observe an increase in negative affect for the entire time (4 hours) of EMAS presentation only in women in the follicular phase (p = 0.0464, n = 10, Wilcoxon Signed-Ranks test).
VAS showed significant increase of a parameter “experiencing sexual arousal” 15 min after beginning of EMAS application in women in follicular phase of the menstrual cycle (p = 0.0431, n = 10, Wilcoxon Signed-Ranks test) and a tendency for increase in luteal phase of menstrual cycle (p = 0.0587, n = 9, Wilcoxon Signed-Ranks test) (Figure 6).
We also observed a short-term increase of a parameter “focused” 15 min after beginning of EMAS application in group of women in follicular phase of the menstrual cycle (p = 0.0284, n = 10, Wilcoxon Signed-Ranks test), but not in luteal phase (p = 0.9528, n = 9, Wilcoxon Signed-Ranks test).
4. Discussion
Presentation of extracts of male axillary secretions (EMAS) to healthy heterosexual women could cause a number of different responses as body odor may provide gender specific information even if it doesn’t register on a conscious level [4]. These responses may include changes in hormone secretion in both HPA and HPG axes as well as mood changes and other adaptive reactions in accordance with the context of the signal [3]. With this in mind we consider the following results of our study as its key findings: 1). The menstrual cycle phase of the recipient determines the hormonal EMAS effects: increase in number of salivary LH peaks and average LH peak amplitude for application in follicular phase and vice versa a decrease in LH amplitude for application in luteal phase. 2). EMAS application increases salivary cortisol level in reproductive age women relative to premenopausal women 3). EMAS may affect the emotional state of women, with changes of positive and negative affect by the end of testing as well as parameters “experiencing sexual arousal” and “focused” obtained with VAS.
To the best of our knowledge this is the first report of the influence of body odor on LH secretion during the luteal phase of the recipient. We observed a considerable decrease in the average amplitude of LH peaks after EMAS application during the luteal phase, while the number of salivary LH peaks remained the same. Our findings of increase in the number of LH peaks in saliva in follicular phase are in good agreement with findings of Preti et al. in blood [6] - extracts of male axillary secretions advance the onset of the next LH peak in women. Moreover, for the first time we found that the average amplitude of salivary LH peaks also increased. Apparently, the observed increase in LH secretion during follicular phase could be regarded as a possible mechanism for shortening of abnormally long menstrual cycles, shown in our previous research [9], since the increase in pulsation of LH may contribute to the acceleration of ovulation. Possible acceleration of the ovulation onset due to EMAS application in women with normal cycle length suggests a mechanism of cycle lengthening during the luteal phase. Otherwise, it is difficult to explain the constant cycle length [9]. In the present study we observed a decrease in the average amplitude of LH peaks (a decrease in hormone secretion) after EMAS application during the luteal phase of the menstrual cycle. We assume that the basis for these results lies in a completely different role that LH performs in the luteal phase compared to the follicular one [23]. At the same time, the onset of a new menstrual cycle is manifested by primarily sharp increase of LH pulse frequency, not change in their amplitude [25].
We have designed our experiments with the assumption that the frequency of LH peaks was affected by the phase of the menstrual cycle of test subjects. In particular, during a normal menstrual cycle one can expect a single LH pulse in blood every 60–90 min during the follicular phase and one pulse every 3–4 h during luteal phase [23]. Our current data obtained for saliva have a similar pattern. Besides, we obtained the values of LH concentrations in saliva similar to those obtained in other works [26,27].
Since the profile of the LH concentration curve in blood has a specific pulse shape with a pronounced maximum, the software used for detection of LH pulses in blood, for example MATLAB module Cluster Analysis, developed by Urban as described in [28] is based on capturing the sudden change in sign of the derivative of the curve from positive to negative. Meanwhile in saliva, LH peaks have smoother shape and as a result, they are not detected by the specific software described above as pulses. Due to the lack of a distinct pulsatile pattern, we have used the baseline method for peak detection of LH [20,21,22].
One can assume contribution of diurnal changes of LH secretions on the observed effects. Several works are devoted to diurnal changes of LH secretion [29,30,31,32], but their results are inconsistent. One of them showed absence of circadian rhythms of gonadotropin secretion in women in the early follicular phase [30]. Another showed presence of some diurnal changes of LH secretion only in the follicular phase [32]. One more study didn’t report menstrual cycle phase, and no circadian rhythms for LH secretion was found [31]. So it is difficult to conclude whether diurnal changes of LH secretion had an impact in our research. At the same time, according to Rossmanith [29] days of the cycle in which we have chosen to perform our experiment correspond to late follicular and mid-luteal phases of test subjects. For the late follicular phase there was no increase of LH pulsation in blood during 2 p.m. – 6 p.m. when we observed an increase in LH amplitude. For the luteal phase they observed an increase of LH pulsation from 1 p.m. till 9 p.m., whereas we found a decrease of LH amplitude for the period from 2 p.m. to 6 p.m.
In this work we also evaluated the effect of EMAS application on secretion of the main human glucocorticoid – cortisol in women of different age groups. We found that salivary cortisol level in reproductive age women did not depend on the phase of their menstrual cycle. However, comparing reproductive age and premenopausal women, we found that relative change of salivary cortisol after EMAS application depended on age group of test subjects. There are diurnal changes in salivary cortisol with the peak during the first hour after awakening - so called cortisol awakening response (CAR), which stay in consistent ranges for a single woman during the whole menstrual cycle [33]. During the experiment (12.00-16.00) for every 2 hours salivary cortisol doesn’t change significantly or have a tendency to decline [24,34]. On the contrary, EMAS caused an increase in the level of salivary cortisol in reproductive age women by an average of 15.1%. At the same time cortisol concentrations were within the physiological range both in the experiment and the control and lower than CAR [33], indicating a possible stimulating effect of EMAS rather than a stressful one.
To the best of our knowledge, this is the first report on the effect of intact male axillary secretions on the level of cortisol in women. However, for a single component of the axillary secretions – androstadienone, it was shown that its sniffing led to an increase in the level of salivary cortisol in reproductive age women with mean age of 20-23 y.o. [35].
In premenopausal women, application of EMAS, on the contrary, lowered the level of cortisol in saliva compared to the control period by 25%, which corresponded to diurnal changes of human cortisol secretion [24]. Such a different reaction is probably associated with a different hormonal status of women of different age groups.
We evaluated the influence of EMAS on the emotional state of women, depending on the phase of the menstrual cycle using Positive and Negative Affect Schedule (PANAS) and Visual Analogue Scales (VAS) [17,18]. We did not observe any short-term changes in positive or negative affect, but for the entire time (4 hours) of EMAS application we observed a decrease in positive affect for women in both phases as well as an increase of negative affect in women only in the follicular phase. We suppose that such result may be not only due to EMAS application, but rather represents general fatigue of test subjects by the end of 8 hours testing. Meanwhile, VAS showed significant short-term increase of a parameter “experiencing sexual arousal” during EMAS application in women in the follicular phase of menstrual cycle and a tendency to increase in the luteal phase of menstrual cycle. At the first glance, this is not very consistent with an increase in salivary cortisol level in the same subjects, since sexual arousal has been shown to be associated with a decrease in cortisol levels [36]. However, we evaluated a subjective sensation, not the physiological response, and as it was mentioned before, the increase was significant, but lower than the average value of CAR [33]. Stimulating effect of an increase in salivary cortisol level is also consistent with an increase of the parameter “focused” in the group of reproductive age women in the follicular phase of the menstrual cycle.
Various modulatory effects were shown for male axillary secretions or their components, usually using Visual Analogue Scales (VAS). For example, male axillary secretions made women more relaxed and less tense [6], while low concentrations of androstadienone made subjects more focused [37]. Activation of the cortical area of the brain, responsible for attention, was also shown when sniffing androstadienone at a sufficiently high concentration exceeding naturally occurring in the axillae [38]. Male axillary secretions can affect the mood of women through the modulation of the serotonin (5-HT) system, namely, by a rapid increase in the affinity of the 5-HT transporter without quantitative changes [39]. Although women in this study were tested only during the follicular phase of the menstrual cycle, besides, extract of male axillary secretions was applied directly to the area under the nose, in this case extract can be absorbed through the skin and enter the bloodstream.
It is possible that more than one compound in axillary secretions can induce endocrine and modulatory changes in human recipients, as far as chemical signals in mammals can be a multicomponent combination of molecules exhibiting a synergistic effect in contrast to an action of separate one [40]. The candidates for the role of chemical signal from axillary secretions could be among: organic unsaturated fatty acids 3-methyl-2-hexenoic acid (3M2H) and 3-hydroxy-3-methyl-hexanoic acid (HMHA), sulfur-containing 3-sulfanyl-3-methyl-hexan1-ol, and volatile steroids, such as 5α-androst-16-en-3α-ol (androstenol) and 4,16-androstadien-3-one (androstadienone), although their concentration is much lower than that of other compounds [41,42].
Up to day, most works on the study of components from axillary secretions as chemical signals were devoted to volatile steroids, whereas works on such components as fatty acids are sparse. In a preliminary study by Ferdenzi [41], the perception of the main compound of human sweat odor more characteristic of men, 3-hydroxy-3-methylhexanoic acid (HMHA), was studied in representatives of the European and Madagascar populations. A change in sensitivity to this acid was found to depend on the phase of the menstrual cycle: women in the fertile phase of the cycle (the period of 6 days before ovulation) perceived 3-hydroxy-3-methylhexanoic acid as more intense than women in the non-fertile phase, which made it a possible candidate for being a chemical signal, though further study is required. The changes in LH secretion after EMAS application shown in the current study can be used in future research as a bioassay to test activity of the mentioned components from axillary secretions. Candidate/s for the role of a human chemical signal should be necessary and sufficient to recreate the original activity of axillary secretions with the original bioassay at naturally occurring concentrations [40].
There are two hypotheses on how individuals assess potential mating partners via telereceptive senses such as vision, olfaction, and hearing. According to the backup signals hypothesis [43,44,45], multimodal cues provide redundant information, whereas the multiple messages hypothesis [43] suggests that different modalities provide independent and distinct information about an individual’s mating-related quality. Our results of the significant effect of EMAS applications on hormone secretion fall in better with multiple messages hypothesis. They are also in good agreement with meta-analysis [46] that showed that body odor may provide distinct and non-redundant information about an individual’s mating-related qualities as opposed to that accessible through either facial or vocal cues.
Author Contributions
Conceptualization, V.V.; methodology, T.L., V.V. and I.K.; validation, T.L., V.V. and I.K.; formal analysis, T.K. and I.K.; investigation, T.K., V.V. and I.K.; resources, V.V.; data curation, T.L. and I.K.; writing—original draft preparation, T.L.; writing—review and editing, I.K. and V.V.; visualization, T.L.; supervision, V.V.; project administration, V.V.; funding acquisition, V.V. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by RUSSIAN SCIENCE FOUNDATION, project number 16-15-10312 to VV “The Influence of Olfactory Environment on Physiological Status and Cognition in Humans”.
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Bioethics Committee at the A.N. SEVERTSOV INSTITUTE OF ECOLOGY & EVOLUTION OF RUSSIAN ACADEMY OF SCIENCES (no.#01.19.06.2017).
Informed Consent Statement
Informed consent was obtained from all participants involved in the study.
Data Availability Statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- McGann, J.P. Poor Human Olfaction Is a 19th-Century Myth. Science 2017, 356, eaam7263. [Google Scholar] [CrossRef] [PubMed]
- Mahmut, M.K.; Croy, I. The Role of Body Odors and Olfactory Ability in the Initiation, Maintenance and Breakdown of Romantic Relationships—A Review. Physiology & Behavior 2019, 207, 179–184. [Google Scholar] [CrossRef]
- Loos, H.M.; Schaal, B.; Pause, B.M.; Smeets, M.A.M.; Ferdenzi, C.; Roberts, S.C.; De Groot, J.; Lübke, K.T.; Croy, I.; Freiherr, J.; et al. Past, Present, and Future of Human Chemical Communication Research. Perspect Psychol Sci 2023, 17456916231188148. [Google Scholar] [CrossRef] [PubMed]
- Lundström, J.N.; Olsson, M.J. Functional Neuronal Processing of Human Body Odors. In Vitamins & Hormones; Elsevier, 2010; Vol. 83, pp. 1–23. ISBN 9780123815163. [Google Scholar] [CrossRef]
- Cutler, W.B.; Preti, G.; Krieger, A.; Huggins, G.R.; Garcia, C.R.; Lawley, H.J. Human Axillary Secretions Influence Women’s Menstrual Cycles: The Role of Donor Extract from Men. Hormones and Behavior 1986, 20, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Preti, G.; Wysocki, C.J.; Barnhart, K.T.; Sondheimer, S.J.; Leyden, J.J. Male Axillary Extracts Contain Pheromones That Affect Pulsatile Secretion of Luteinizing Hormone and Mood in Women Recipients1. Biology of Reproduction 2003, 68, 2107–2113. [Google Scholar] [CrossRef] [PubMed]
- Rozenkrantz, L.; Weissgross, R.; Weiss, T.; Ravreby, I.; Frumin, I.; Shushan, S.; Gorodisky, L.; Reshef, N.; Holzman, Y.; Pinchover, L.; et al. Unexplained Repeated Pregnancy Loss Is Associated with Altered Perceptual and Brain Responses to Men’s Body-Odor. eLife 2020, 9, e55305. [Google Scholar] [CrossRef]
- Prokop-Prigge, K.A.; Greene, K.; Varallo, L.; Wysocki, C.J.; Preti, G. The Effect of Ethnicity on Human Axillary Odorant Production. J Chem Ecol 2016, 42, 33–39. [Google Scholar] [CrossRef]
- Voznessenskaya, V.V.; Laktionova, T.K. Influence of the Male Axillary Extracts on Regulation of Menstrual Cycles in Women. Dokl Biol Sci 2018, 478, 19–21. [Google Scholar] [CrossRef]
- Derntl, B.; Schopf, V.; Kollndorfer, K.; Lanzenberger, R. Menstrual Cycle Phase and Duration of Oral Contraception Intake Affect Olfactory Perception. Chemical Senses 2013, 38, 67–75. [Google Scholar] [CrossRef]
- Yao, F.; Chen, K.; Zhuang, Y.; Shen, X.; Wang, X. Mid-Luteal Olfactory Abilities Reveal Healthy Women’s Emotional and Cognitive Functions. Front. Neurosci. 2022, 16, 826547. [Google Scholar] [CrossRef]
- Roberts, S.C.; Třebická Fialová, J.; Sorokowska, A.; Langford, B.; Sorokowski, P.; Třebický, V.; Havlíček, J. Emotional Expression in Human Odour. Evolut. Hum. Sci. 2022, 4, e44. [Google Scholar] [CrossRef] [PubMed]
- De Groot, J.H.B.; Haertl, T.; Loos, H.M.; Bachmann, C.; Kontouli, A.; Smeets, M.A.M. Unraveling the Universality of Chemical Fear Communication: Evidence from Behavioral, Genetic, and Chemical Analyses. Chemical Senses 2023, 48, bjad046. [Google Scholar] [CrossRef] [PubMed]
- Preti, G.; Cutler, W.B.; Garcia, C.R.; Huggins, G.R.; Lawley, H.J. Human Axillary Secretions Influence Women’s Menstrual Cycles: The Role of Donor Extract of Females. Hormones and Behavior 1986, 20, 474–482. [Google Scholar] [CrossRef]
- Zeng, X.-N.; Leyden, J.J.; Lawley, H.J.; Sawano, K.; Nohara, I.; Preti, G. Analysis of Characteristic Odors from Human Male Axillae. J Chem Ecol 1991, 17, 1469–1492. [Google Scholar] [CrossRef] [PubMed]
- Watson, D.; Clark, L.A.; Tellegen, A. Development and Validation of Brief Measures of Positive and Negative Affect: The PANAS Scales. Journal of Personality and Social Psychology 1988, 54, 1063–1070. [Google Scholar] [CrossRef]
- Osin, E.N. Measuring Positive and Negative Affect: Development of a Russian-language Analogue of PANAS: Psychology. Journal of Higher School of Economics 2012, 9, 91–110. (In Russian) [Google Scholar]
- Aitken, R.C. Measurement of Feelings Using Visual Analogue Scales. Proc R Soc Med 1969, 62, 989–993. [Google Scholar]
- Seubert, J.; Gregory, K.M.; Chamberland, J.; Dessirier, J.-M.; Lundström, J.N. Odor Valence Linearly Modulates Attractiveness, but Not Age Assessment, of Invariant Facial Features in a Memory-Based Rating Task. PLoS ONE 2014, 9, e98347. [Google Scholar] [CrossRef]
- Brown, J.L.; Schmitt, D.L.; Bellem, A.; Graham, L.H.; Lehnhardt, J. Hormone Secretion in the Asian Elephant (Elephas Maximus): Characterization of Ovulatory and Anovulatory Luteinizing Hormone Surges. Biology of Reproduction 1999, 61, 1294–1299. [Google Scholar] [CrossRef]
- Bruin, P.R.D.; Medger, K.; Bennett, N.C.; Ganswindt, A. Assessment of Reproductive Function in Southern African Spiny Mice ( Acomys Spinosissimus ) Using Faeces as Hormone Matrix. African Zoology 2014, 49, 44–53. [Google Scholar] [CrossRef]
- Kvasha, I.G.; Laktionova, T.K.; Voznessenskaya, V.V. The Presentation Rate of Chemical Signals of the Domestic Cat Felis Catus Affects the Reproductive Status of the House Mouse. Biol Bull Russ Acad Sci 2018, 45, 278–283. [Google Scholar] [CrossRef]
- Gingold, J.A.; Jain, M.; Jalai, C. Hypothalamic-Pituitary-Ovarian Axis and Control of the Menstrual Cycle. In Clinical Reproductive Medicine and Surgery; Falcone, T., Hurd, W.W., Eds.; Springer International Publishing: Cham, 2022; pp. 1–22. ISBN 9783030995959 9783030995966. [Google Scholar] [CrossRef]
- Mezzullo, M.; Fanelli, F.; Fazzini, A.; Gambineri, A.; Vicennati, V.; Di Dalmazi, G.; Pelusi, C.; Mazza, R.; Pagotto, U.; Pasquali, R. Validation of an LC–MS/MS Salivary Assay for Glucocorticoid Status Assessment: Evaluation of the Diurnal Fluctuation of Cortisol and Cortisone and of Their Association within and between Serum and Saliva. The Journal of Steroid Biochemistry and Molecular Biology 2016, 163, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Hall, J.E.; Schoenfeld, D.A.; Martin, K.A.; Crowley, W.F. Hypothalamic Gonadotropin-Releasing Hormone Secretion and Follicle-Stimulating Hormone Dynamics during the Luteal-Follicular Transition. The Journal of Clinical Endocrinology & Metabolism 1992, 74, 600–607. [Google Scholar] [CrossRef] [PubMed]
- Loewit, K.K.; Kraft, H.-G.; Ortlieb, A. Measurement of LH in Saliva; a New Approach to Ovulation Detection. In Future Aspects in Contraception; Runnebaum, B., Rabe, T., Kiesel, L., Eds.; Springer: Dordrecht, Netherlands, 1985; pp. 31–47. ISBN 9789401086783 9789400949164. [Google Scholar] [CrossRef]
- Saibaba, G.; Srinivasan, M.; Priya Aarthy, A.; Silambarasan, V.; Archunan, G. Ultrastructural and Physico-Chemical Characterization of Saliva during Menstrual Cycle in Perspective of Ovulation in Human. DD&T 2017, 11, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Urban, R.J.; Johnson, M.L.; Veldhuis, J.D. In Vivo Biological Validation and Biophysical Modeling of the Sensitivity and Positive Accuracy of Endocrine Peak Detection. I. The LH Pulse Signal*. Endocrinology 1989, 124, 2541–2547. [Google Scholar] [CrossRef] [PubMed]
- Rossmanith, W.G.; Lauritzen, C. The Luteinizing Hormone Pulsatile Secretion: Diurnal Excursions in Normally Cycling and Postmenopausal Women. Gynecological Endocrinology 1991, 5, 249–265. [Google Scholar] [CrossRef] [PubMed]
- Klingman, K.M.; Marsh, E.E.; Klerman, E.B.; Anderson, E.J.; Hall, J.E. Absence of Circadian Rhythms of Gonadotropin Secretion in Women. The Journal of Clinical Endocrinology & Metabolism 2011, 96, 1456–1461. [Google Scholar] [CrossRef] [PubMed]
- Van Kerkhof, L.W.M.; Van Dycke, K.C.G.; Jansen, E.H.J.M.; Beekhof, P.K.; Van Oostrom, C.T.M.; Ruskovska, T.; Velickova, N.; Kamcev, N.; Pennings, J.L.A.; Van Steeg, H.; et al. Diurnal Variation of Hormonal and Lipid Biomarkers in a Molecular Epidemiology-Like Setting. PLoS ONE 2015, 10, e0135652. [Google Scholar] [CrossRef]
- Rahman, S.A.; Grant, L.K.; Gooley, J.J.; Rajaratnam, S.M.W.; Czeisler, C.A.; Lockley, S.W. Endogenous Circadian Regulation of Female Reproductive Hormones. The Journal of Clinical Endocrinology & Metabolism 2019, 104, 6049–6059. [Google Scholar] [CrossRef]
- Ozgocer, T.; Ucar, C.; Yildiz, S. Daily Cortisol Awakening Response and Menstrual Symptoms in Young Females. Stress and Health 2022, 38, 57–68. [Google Scholar] [CrossRef]
- Vining, R.F.; McGinley, R.A.; Maksvytis, J.J.; Ho, K.Y. Salivary Cortisol: A Better Measure of Adrenal Cortical Function than Serum Cortisol. Ann Clin Biochem 1983, 20, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Wyart, C.; Webster, W.W.; Chen, J.H.; Wilson, S.R.; McClary, A.; Khan, R.M.; Sobel, N. Smelling a Single Component of Male Sweat Alters Levels of Cortisol in Women. J. Neurosci. 2007, 27, 1261–1265. [Google Scholar] [CrossRef]
- Hamilton, L.D.; Rellini, A.H.; Meston, C.M. Cortisol, Sexual Arousal, and Affect in Response to Sexual Stimuli. The Journal of Sexual Medicine 2008, 5, 2111–2118. [Google Scholar] [CrossRef] [PubMed]
- Lundström, J. Psychological Effects of Subthreshold Exposure to the Putative Human Pheromone 4,16-Androstadien-3-One. Hormones and Behavior 2003, 44, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Gulyás, B.; Kéri, S.; O’Sullivan, B.T.; Decety, J.; Roland, P.E. The Putative Pheromone Androstadienone Activates Cortical Fields in the Human Brain Related to Social Cognition. Neurochemistry International 2004, 44, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Masala, I.; Baroni, S.; Polini, M.; Massimetti, G.; Giannaccini, G.; Betti, L.; Italiani, P.; Fabbrini, L.; Caglieresi, C.; et al. Male Axillary Extracts Modify the Affinity of the Platelet Serotonin Transporter and Impulsiveness in Women. Physiology & Behavior 2010, 100, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, T.D. Pheromones and Animal Behavior: Chemical Signals and Signatures, 2nd edition; Cambridge University Press: Cambridge, 2014; ISBN 9780521112901 9780521130196. [Google Scholar]
- Ferdenzi, C.; Razafindrazaka, H.; Baldovini, N.; Poupon, D.; Pierron, D.; Bensafi, M. Influence of Gender and Culture on the Perception of Acidic Compounds of Human Body Odor. Physiology & Behavior 2019, 210, 112561. [Google Scholar] [CrossRef] [PubMed]
- Natsch, A.; Emter, R. The Specific Biochemistry of Human Axilla Odour Formation Viewed in an Evolutionary Context. Philosophical Transactions of the Royal Society B: Biological Sciences, 2020; 375, 20190269. [Google Scholar] [CrossRef]
- Moller, A.P.; Pomiankowski, A. Why Have Birds Got Multiple Sexual Ornaments? Behav Ecol Sociobiol 1993, 32. [Google Scholar] [CrossRef]
- Thornhill, R. The Scent of Symmetry: A Human Sex Pheromone That Signals Fitness? Evolution and Human Behavior 1999, 20, 175–201. [Google Scholar] [CrossRef]
- Grammer, K.; Fink, B.; Juette, A.; Ronzal, G.; Thornhill, R. Female Faces and Bodies: N-Dimensional Feature Space and Attractiveness. In Facial attractiveness: Evolutionary, cognitive, and social perspectives; Advances in visual cognition, vol. 1.; Ablex Publishing: Westport, CT, US, 2002; pp. 91–125. ISBN 9781567506365 9781567506372. [Google Scholar]
- Třebický, V.; Delplanque, S.; Ferdenzi, C.; Fink, B.; Jelínková, L.; Pátková, Ž.; Roberts, S.C.; Röder, S.; Saxton, T.K.; Schwambergová, D.; et al. Cross-Modal Associations of Human Body Odour Attractiveness with Facial and Vocal Attractiveness Provide Little Support for the Backup Signals Hypothesis: A Systematic Review and Meta-Analysis. Evolution and Human Behavior 2023, 44, 19–29. [Google Scholar] [CrossRef]
Figure 1.
Example of salivary LH pattern of woman tested in follicular phase of menstrual cycle. Arrow indicates the beginning of EMAS application (14.00).
Figure 1.
Example of salivary LH pattern of woman tested in follicular phase of menstrual cycle. Arrow indicates the beginning of EMAS application (14.00).
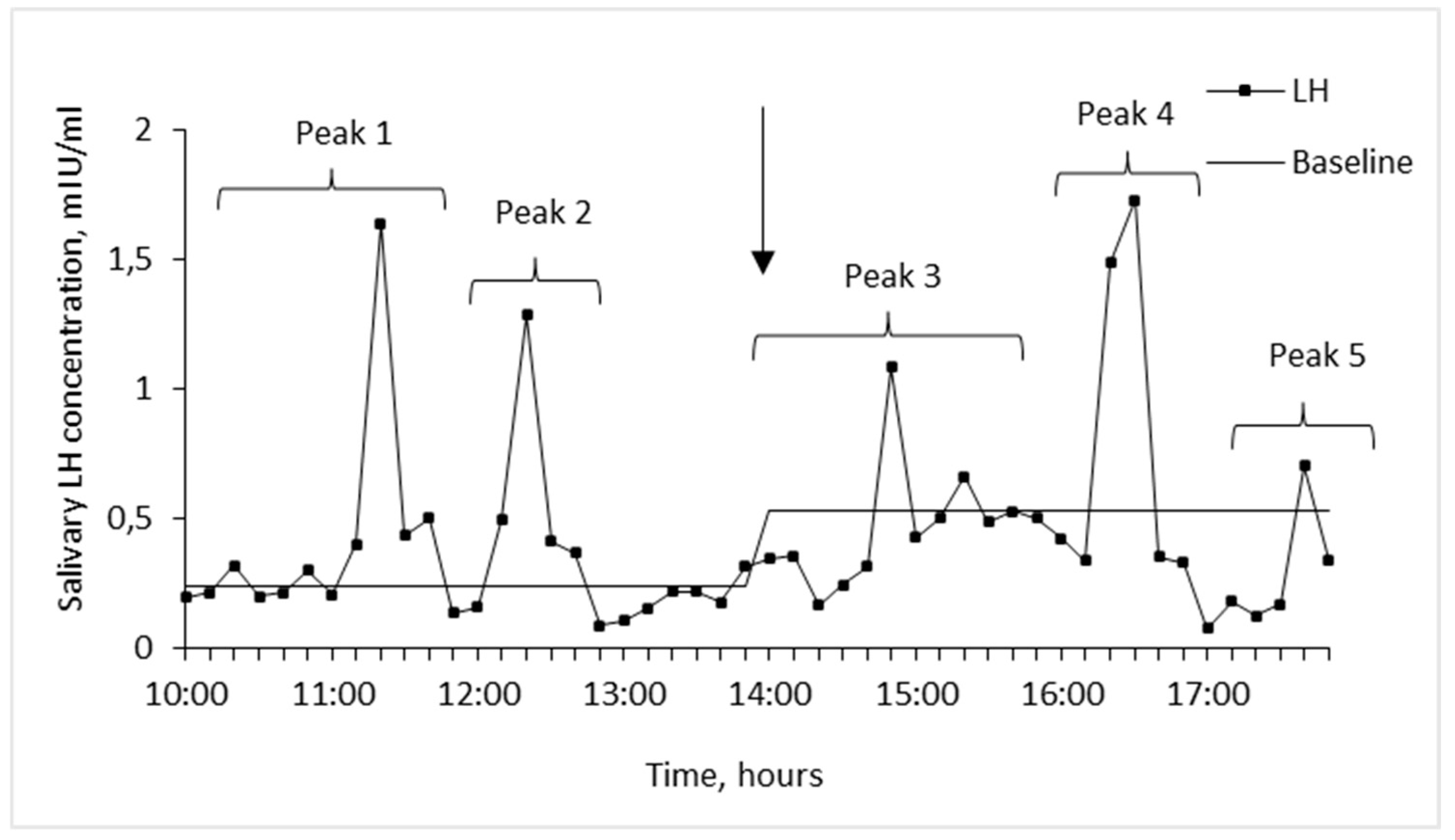
Figure 2.
The influence of the menstrual cycle phase on the number of LH peaks (Control: p = 0.0002; EMAS: p = 0.0000, n = 19, post-hoc Fisher LSD test). The data are presented as: M ± SD.
Figure 2.
The influence of the menstrual cycle phase on the number of LH peaks (Control: p = 0.0002; EMAS: p = 0.0000, n = 19, post-hoc Fisher LSD test). The data are presented as: M ± SD.
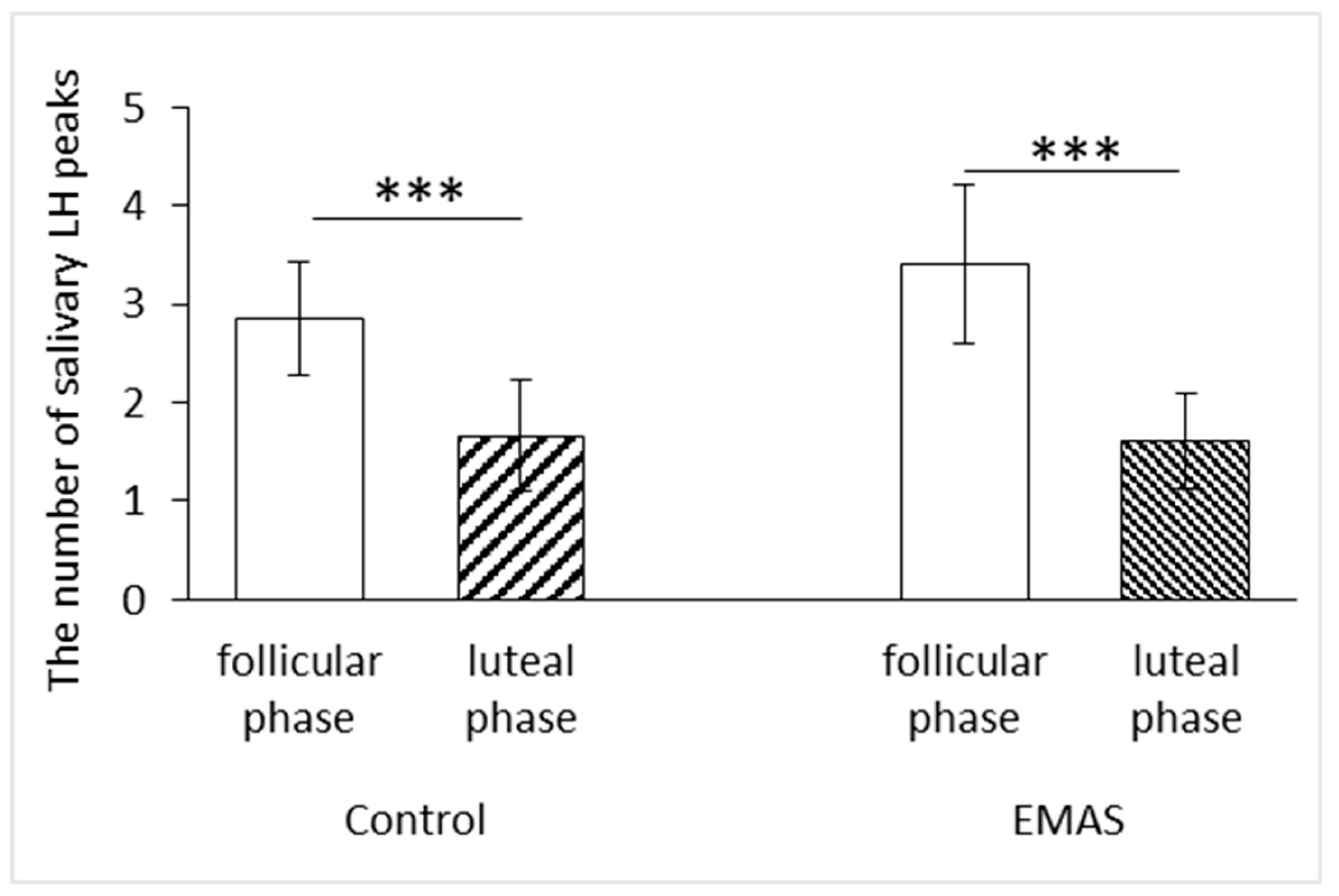
Figure 3.
The influence of EMAS application on the number of salivary LH peaks in a group of women in follicular (p = 0.0447, n = 10, post-hoc Fisher LSD test). The data are presented as: M ± SD.
Figure 3.
The influence of EMAS application on the number of salivary LH peaks in a group of women in follicular (p = 0.0447, n = 10, post-hoc Fisher LSD test). The data are presented as: M ± SD.
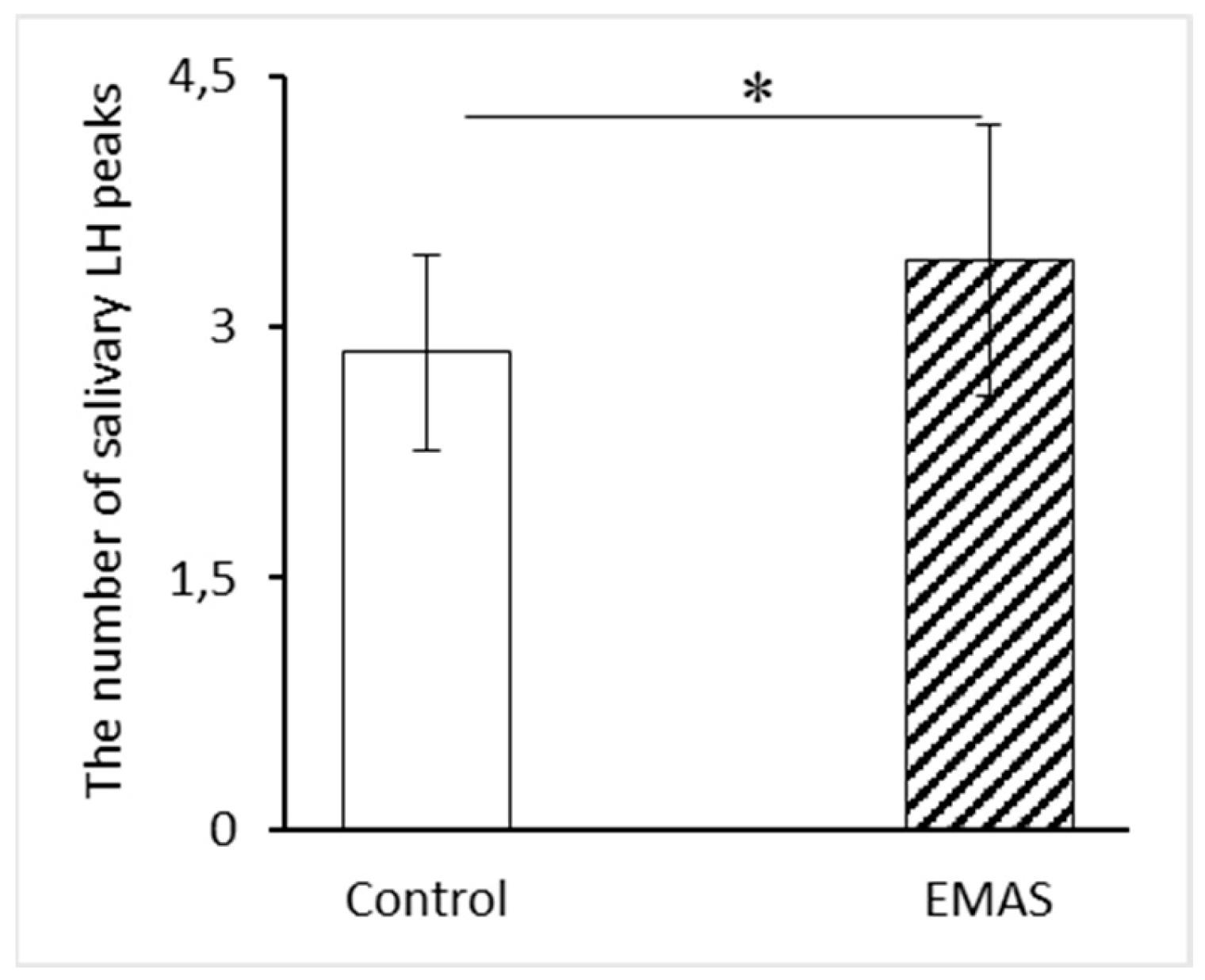
Figure 4.
The influence of EMAS application on the average amplitude of salivary LH peaks depending on the phase of menstrual cycle of test subjects (follicular phase: p = 0.0469, n = 10; luteal phase: p = 0.0382, n = 9, Wilcoxon Signed-Ranks test). The data are presented as: M ± SD.
Figure 4.
The influence of EMAS application on the average amplitude of salivary LH peaks depending on the phase of menstrual cycle of test subjects (follicular phase: p = 0.0469, n = 10; luteal phase: p = 0.0382, n = 9, Wilcoxon Signed-Ranks test). The data are presented as: M ± SD.
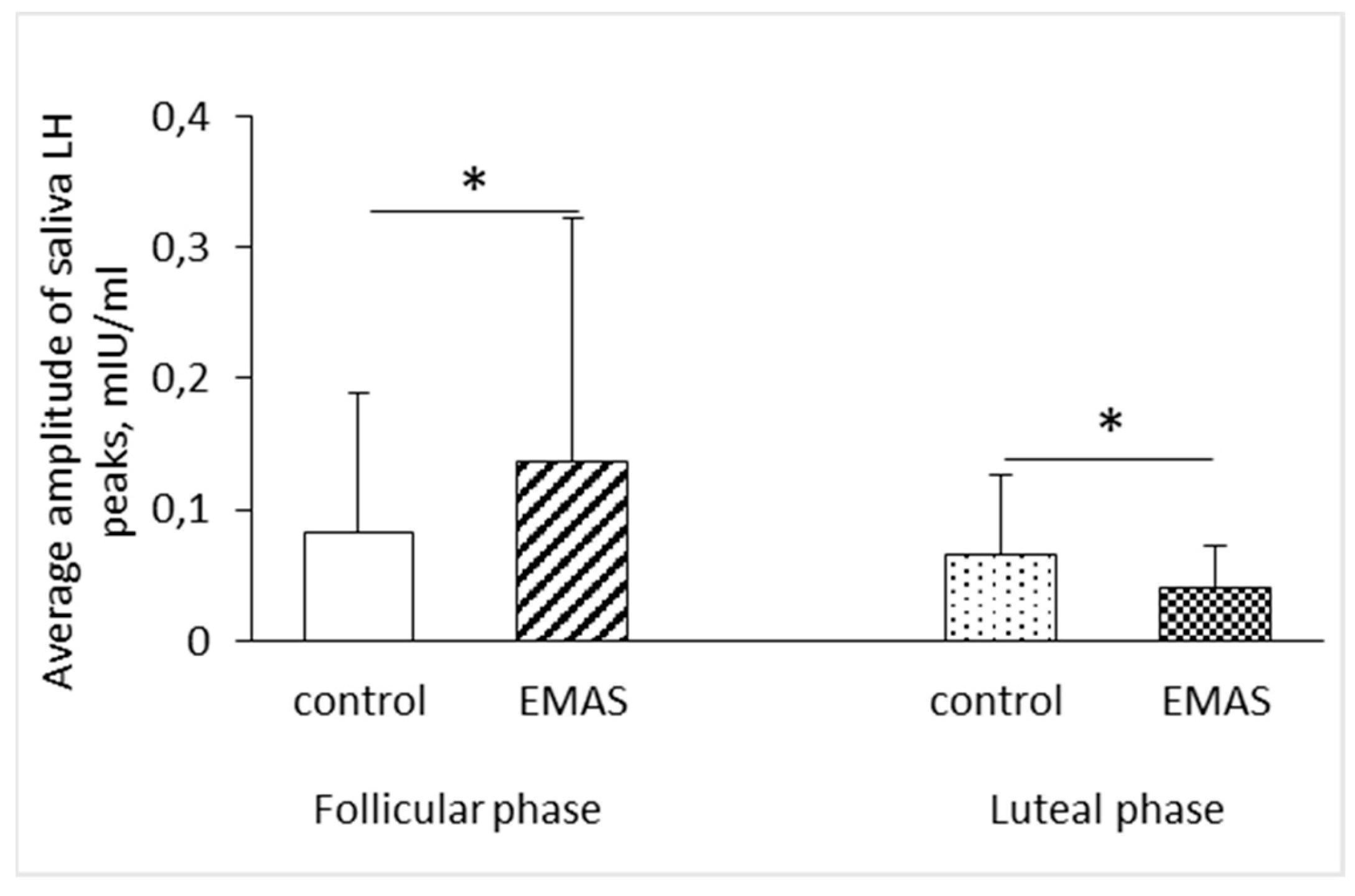
Figure 5.
Relative salivary cortisol level change (in %) after application of EMAS in reproductive age women and in premenopausal women. The data are presented as: M ± SD; ** p ˂ 0.01, n = 29, permutation test with Monte-Carlo approximation.
Figure 5.
Relative salivary cortisol level change (in %) after application of EMAS in reproductive age women and in premenopausal women. The data are presented as: M ± SD; ** p ˂ 0.01, n = 29, permutation test with Monte-Carlo approximation.
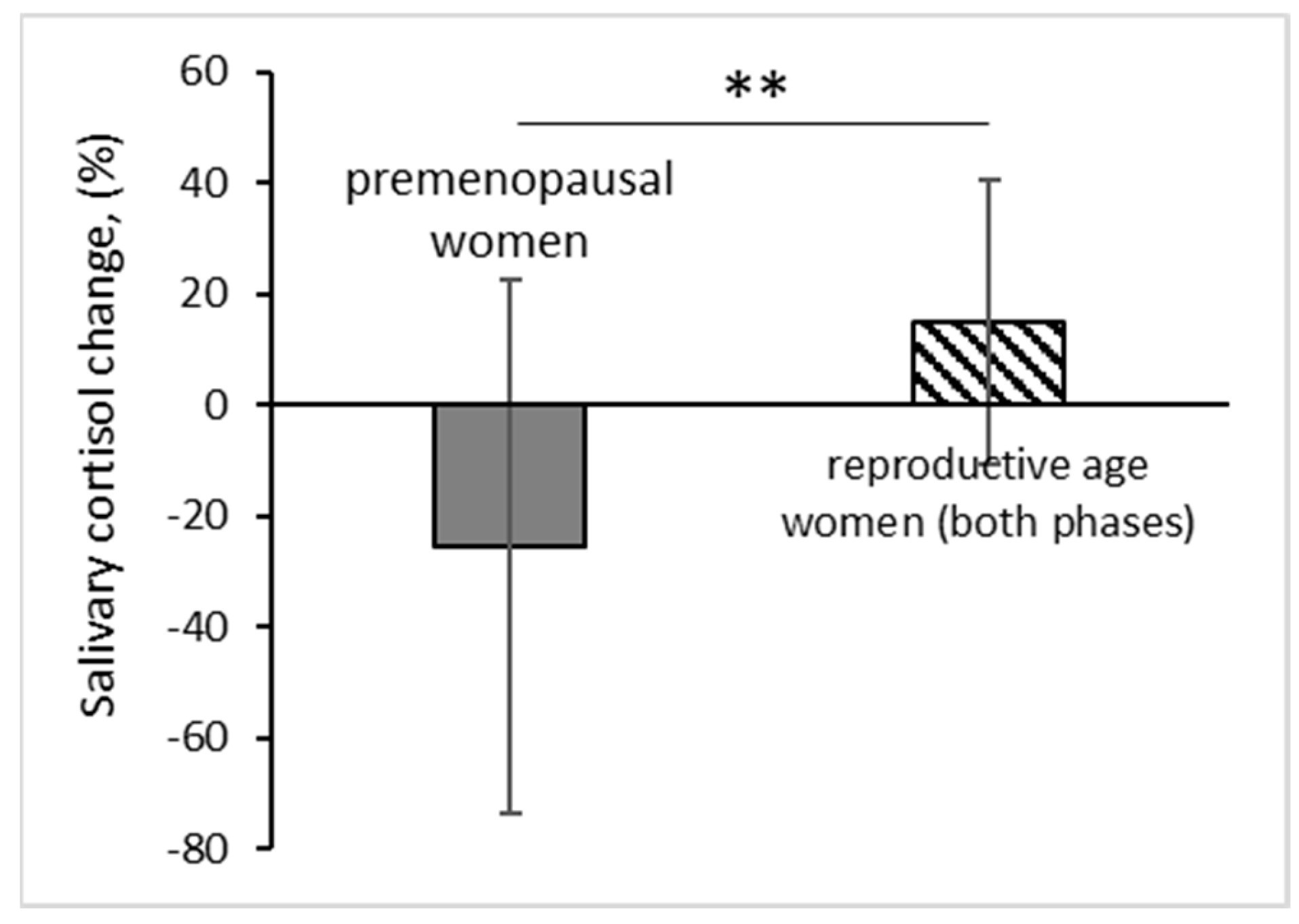
Figure 6.
The influence of EMAS application on parameter “experiencing sexual arousal” in a group of women in follicular (p = 0.0431, n = 10, Wilcoxon Signed-Ranks test) and luteal (p = 0.0587, n = 9, Wilcoxon Signed-Ranks test) phase of the menstrual cycle.
Figure 6.
The influence of EMAS application on parameter “experiencing sexual arousal” in a group of women in follicular (p = 0.0431, n = 10, Wilcoxon Signed-Ranks test) and luteal (p = 0.0587, n = 9, Wilcoxon Signed-Ranks test) phase of the menstrual cycle.
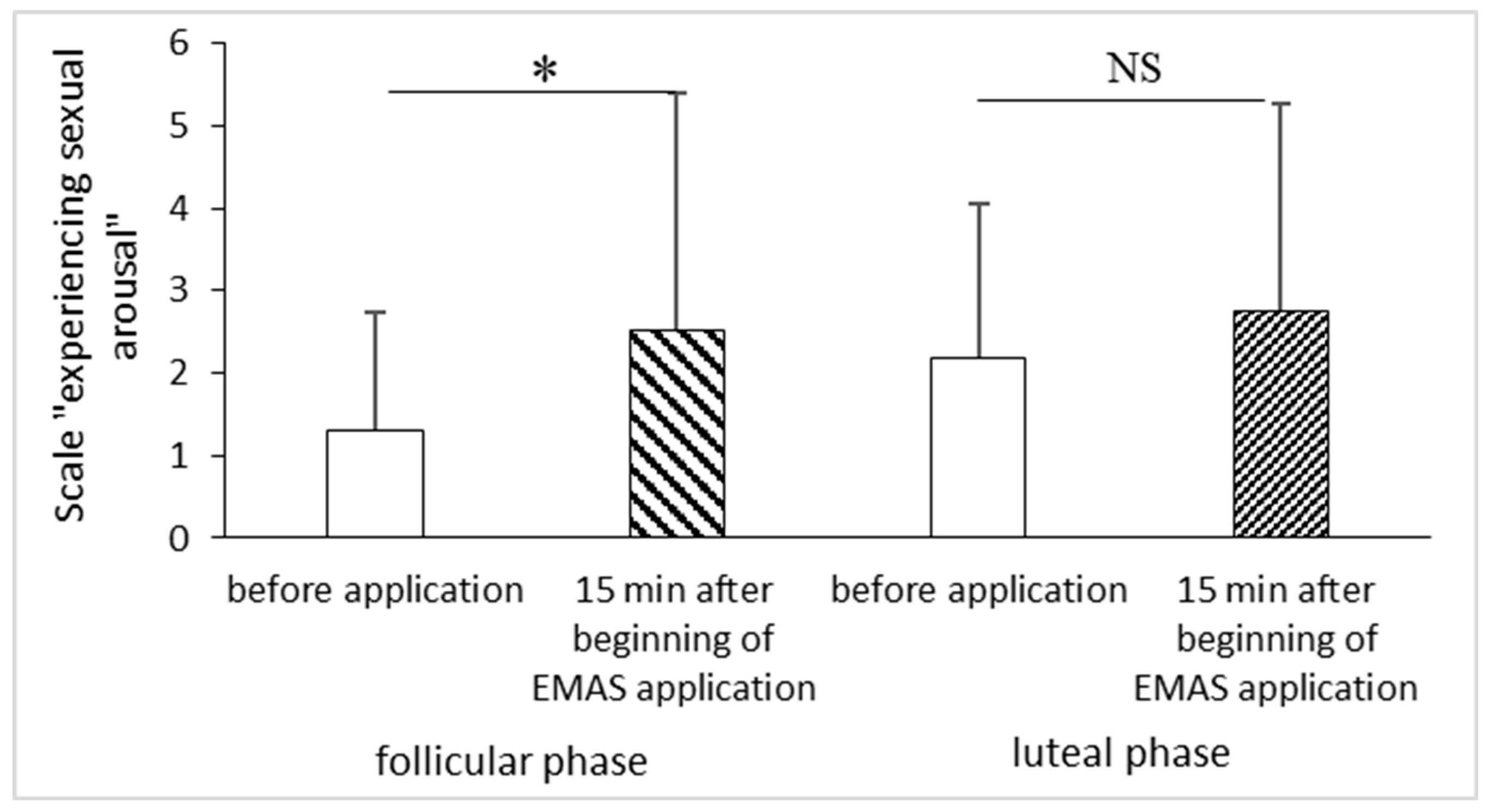

Table 1.
Results of permutation test with Monte-Carlo approximation.
| Component 1: | |||||
|---|---|---|---|---|---|
| Df | R Sum Sq | R Mean Sq | Iter | Pr(Prob) | |
| age group | 1 | 10697 | 10696.9 | 5000 | 0.0032 ** |
| Residuals | 27 | 32726 | 1212.1 |
Signif. codes: 0 ‘***’ 0.001 ‘**’ 0.01 ‘*’ 0.05 ‘.’ 0.1 ‘ ‘ 1.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



