
Submitted:
12 June 2024
Posted:
12 June 2024
You are already at the latest version
Abstract
(1) Background:The objective of this study was to explorethe dynamic changes of gut microbiota in Simmental calves before weaning, and compared microbial composition and functionality differences between diarrhea and health calves; (2) Methods: Fourteen neonatal Simmental calves were divided into a health group (n = 8) and diarrhea group (n = 6). The rectal stool samples were collected from each calf on days 1, 3, 5, 7, 9, 12, 15, 18, 22, 26, 30, 35, and 40. The high-throughput sequencing analysis of the 16S rRNA gene V1–V9 region was conducted to explore the changes in gut microbiota over time in both groups and assess the influence of diarrhea on microbiota structure and function; (3) Results: E.coli, B.fragilis, and B.vulgatus were the top three bacterial species of pre-weaning Simmental calves.Meanwhile, the major functions of fecalmicrobiota included the “Metabolic pathways”,“Biosynthesis of secondary metabolites”,“Biosynthesis of antibiotics”, “Microbial metabolism in diverse environments”, and “Biosynthesis of amino acids”. For calves in health group, PCoA revealed that the bacterial profiles generated from days 1, 3, 5, 7, and 9 separated from days 15, 18, 22, 26, 30, 35, and 40. While the microbial profiles of calves on day 12 crossed with both clusters, suggesting that the dynamic changes of microbial structures with the increasing of ages. When compared the difference of the relative abundance of bacteria between calves in health and diarrhea groups, the results showed that the relative abundance of the beneficial L.johnsonii, F.prausnitzii, and Limosilactobacillus in the health group were significantly higher than those in the diarrhea group (p
Keywords:
Calf diarrhea
; Intestinal flora
; High throughput 16S rRNA
1. Introduction
Simmental cattle, a famous dairy-beef dual-purpose cattle breed originating from Switzerland, are characterized for their outstanding meat and milk production performance. Therefore, Simmental cattle were mainly used to cross-breed dairy and meat cattle after being introduced into China[1] Inner Mongolia is a main area for beef cattle breeding in China, with Tong Liao serving as the core area.In Tongliao, the beef cattle population exceeds three million, with Simmental cattle as the main breed. Calf health is the most critical aspect of beef cattle breeding[2]. Currently, calf diarrhea lead to the highest morbidity and mortality in beef cattle breeding, which causes important economic losses to beef cattle breeding[3].Therefore, it is of significant important to pay attention to the intestinal health of pre-weaning calves during early life[3].
The early colonization of the gut microbiota could significantly affects the gut health of calves [4]. For neonatal calves, microbial colonization process is a complex and dynamic process, which could be influenced by host-microbial interactions, along with external factors [5].Such factors include maternal microbiota, birth process, diet, antibiotics, and weaning and etc. Studies have shown that the intestinal flora of ruminants presented since the fetal period, and when the cattle reaches 5 months during pregnancy, the microflora can be detected in the rumen, cecum, meconium and amniotic fluid of calves [6,7]. After birth,and the intestinal microbiota of neonatal calvesundergos rapid changes. Within approximately 8 h after birth, Escherichia coli and Streptococcus colonize in all gastrointestinal regions (, followed by the detection of lactic acid bacteria and Clostridium perfringens[8]. From the second day to one week after birth, lactic acid bacteria are predominantin both cecum and stoolsamples [9]. For 3 week old calves, the dominant bacterial genus were Bacteroides, Prevotella, Coccus-Useriella and Faebacillus [10] While Lactococcus flavus and cellulolytic bacteria appearede in the 5th week after birth [9]. Within 12 weeks after birth, Prevotella, Bacteroides, Clostridium, and Eubacterium were the main intestinal microbiota of calves [11]. The above studies on microbial dynamic changes were mainly carried out in Holstein calves, however, limited studies have been performed on Simmental calves to explore the microbial colonization process and microbial functions.
Calf diarrhea is the main cause of calf death before weaning. While 39% of calf death in the United States were caused by diarrhea, and 53.4% of calf death were in South Korea, as well as 9.6%-60% mortality rates were reported in Xinjiang, Gansu, Henan, Beijing in China [12]. The factors that cause neonatal calves’ diarrhea are complex, which mainly include nutritional factors, intestinal inflammation, stress, pathogen infection and etc.Antibiotics, such as β-lactams and sulfonamides, have been commonly applied for the treatments of calf diarrhea. The overuse of antibiotics could disrupt the gut microbiota of calves, leading to intestinal disorders. Specifically, antibiotics such as methylene salicylic acid and bacitracin, used in calf diarrhea treatment, may enhance the colonization ofpotential pathogens such as E. coli, Enterococcus, and Shigella in calf intestines, therefore affecting the intestinal microbial balance [12]. Meanwhile, the application of antibiotics also affected the metabolism pattern of intestinal microbiota and the absorption of nutrients in the intestine, hence the long-term use of antibiotics could promote drug-resistant bacteria colonization and increase infection risks of preweaning calves[13]. Additionally,antibiotics residues can have adverse effects on the environment, therefore, it is also important to find alternatives to antibiotics to prevent and treat calves’ diarrhea.Probiotics, including Bifidobacterium pseudocatenulatum, Lactobacillus acidophilus [14], and Bacillus subtilis [15,16], serve as sustainable alternatives to antibiotics for the prevention and treatment of early calf diarrhea. They could resist the pathogen adhesion and enhance the intestinal barrier function, thereby reducing the intestinal damage. Studies have shown that Bifidobacterium pseudomidobacterium could produce lactic acid and short chain fatty acids (SCFAs) to facilitate the prevention and treatment of calf diarrhea [17]. Moreover, the proliferation of probiotics in the intestine could lower the undigested carbohydrate levels, thus reducing the risk of diarrhea resulting from osmotic gradient disruption [18]. The above research results were mainly based on Holstein calves, and there is a dearth of reports on the probiotic applications and diarrhea treatments in Simmental calves.
This study investigated the changes of gut microbiota of healthy Simmental calves and those who experienced diarrhea before weaning over time, as well aspredicting intestinal microbialfunctions. Additionally, we compared the difference in microbial composition and functions of calves in health and diarrhea group before, during, and after diarrhea period
2. Materials and Methods
2.1. Animal Breeding
The experiment was conducted at a commercial ranch from February to April 2023, in Tong Liao City, Inner Mongolia, following the protocols proved by the Laboratory Animal Welfare and Ethics Committee of the College of Animal Science and Technology, Inner Mongolia Minzu University (protocol code: 2022058). In total, 14 neonatal Simmental calves (39 ± 5 kg) were enrolled in our study immediately after birth, and they were managed according to the standard feeding protocols of the ranch. All the calves were immediately separated from maternal cows after birth, and they were raised in individual hutches after umbilical cord disinfection. The calves were received 4 L of colostrum within 1 h, followed by an additional 2 L of colostrum within 8–10 h after birth. Then they were fed 5 L of whole milk/day onwards. Calves had ad libitum access to calf starter from the 3rd day after birth.
2.2. Daily Health Monitoring
Calves were monitored and scored daily before morning and evening feeding. Nasal and eye discharges, coughing, umbilical cord inflammation, rectal temperature, dehydration, and overall condition were recorded and the specific scoring criteria refer to the previously published article [19]. When fecal samples were collected, fecal scores were recorded based on fecal fluidity: 1 = normal, 2 = soft, 3 = runny, or4 = watery. Calves that had a fecal score of 3 or 4 were considered diarrheic[20]. The scoring criteria are provided in (Supplementary File 1).
2.3. Grouping
The calves were categorized into two groups according to the occurrence of diarrhea during the study period: diarrheal group (A)(n=6) and healthy group (B)(n=8). All the calves in diarrheal group experienced diarrhea on day 5 and they were recovered on day 9 after treated with an oral electrolyte solution. Calves in the healthy group remained healthy throughout the whole experimental period. The stool samples were collected from all the calves at 8:00 am, on days 1, 3, 5, 7, 9, 12, 15, 18, 22, 26, 30, 35, and 40 after birth. The fecal samples were temporarily stored in liquid nitrogen, and then transferred to the -80℃ refrigerator for long-term storage.
2.4. High-throughput 16S Ribosomal RNA Gene Sequencing
The total genomic DNA was extracted from the stool samples using the TGuide S96 Magnetic Stool DNA Kit (Tiangen Biotech (Beijing) Co., Ltd.) following the manufacturer’s instructions. The quality and quantity of the extracted DNA were assessed via electrophoresis on a 1.8% agarose gel, and the DNA concentration and purity were determined using a NanoDrop 2000 UV-Vis spectrophotometer (Thermo Scientific, Wilmington, USA). The full-length 16S rRNA gene was amplified using the primer pair 27F: AGRGTTTGATYNTGGCTCAG and 1492R: TASGGHTACCTTGTTASGACTT. Both the forward and reverse 16S primers were tagged with sample-specific PacBio barcode sequences to enable the multiplexed sequencing. Barcoded primers were selected to minimize the chimera formation compared to an alternative protocol involving a second PCR reaction. The PCR amplification was performed using the KOD One PCR Master Mix (Toyobo Life Science) for 25 cycles, comprising an initial denaturation at 95 °C for 2 min, followed by 10-second denaturation at 98°C, annealing at 55°C for 30 s, extension at 72°C for 1 min 30 s per cycle, and a final step at 72°C for 2 min. The total PCR amplicons were purified using VAHTSTM DNA Clean Beads (Vazyme, Nanjing, China) and quantified with the Qubit dsDNA HS Assay Kit and Qubit 3.0 Fluorometer (Invitrogen, Thermo Fisher Scientific, Oregon, USA). After the individual quantification, the amplicons were pooled in equal proportions. The SMRTbell libraries were then generated from the amplified DNA using the SMRTbell Express Template Prep Kit 2.0 according to the manufacturer’s instructions (Pacific Biosciences). The purified SMRTbell libraries derived from the pooled and barcoded samples were subsequently sequenced on a PacBio Sequel II platform (Beijing Biomarker Technologies Co., Ltd., Beijing, China) using the Sequel II binding kit 2.0.
2.5. Bioinformatic Analysis
The qualified sequences exceeding a 97% similarity threshold were assigned to operational taxonomic units (OTUs) using USEARCH (version 10.0). Taxonomic annotation of the OTUs was conducted using the Naive Bayes classifier in QIIME2[21], utilizing the SILVA database[22] (release 138.1) with a confidence threshold of 70%. The α-diversity test was performed to determine the complexity of species diversity in each sample using the QIIME2 software. The beta diversity was analyzed using the principal coordinate analysis (PCoA) to evaluate the species diversity across the samples. The bacterial abundance and diversity were compared using one-way analysis of variance . The gut microbiome was considered significant if its relative abundance exceeded 0.1% and was present in over half of the animals within at least one age group. The FAPROTAX software was employed to predict the function of intestinal fecal microorganisms in calves, focusing on metabolic pathways with a CPM > 5 observed in at least 50% of the animals within each treatment group.
2.6. Statistical Analysis
The data were analyzed using Microsoft Excel 2019 and SPSS 25.0. Non-parametric Wilcoxon test was performed to assess the difference of α-diversity index and the relative abundance of microbiotabetween the control and diarrhea groups at the same sampling day. The top five bacterial species were compared between adjacent time points in the two groups separately,with P<0.05 defines statistical significance, and 0.05<p<0.10 represents the tendency of significance.Meanwhile, the similarity analysis between two groups were performed with Anosim. When p<0.05 and r<0.5, the intestinal microbiota of the two age groups was similar; when p<0.05 and r>0.75, it indicated that the intestinal microbiota of the two age groups were completely different, different when 0.5<r<0.75, and tended to be different when 0.3<r<0.5. There was no difference at r<0.3. Wilcoxon test was applied to compare intergroup differences between the control and diarrhea groups on the same sampling day for gut microbiota function. p<0.05 indicated statistical significance.
2.7. Nucleotide Sequence Accession Numbers
All sequences were depositedat NCBI Sequence Read Archive and are publicly accessible under the accessionnumber PRJNA1072024. These files could be accessed in the following link: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1072024.
3. Results
3.1. Data Collection and Diversity of Gut Microbiota
In total, amplicon sequencing of the pre-weaning Simmental calves’ intestinal microbiota generated 2,185,694 high-quality sequences that were assigned to a total of 1,122 OTUs based on 97% nucleotide sequence similarity (Supplementary File2). Both the sparsity curve and the rank abundance curve showed a saturation trend, indicating that the depth and uniformity of sequencing could meet the requirements for subsequent analysis (Supplementary File3).The calves in healthy group exhibited 34 shared OTUs at various time points with additional 12,15,28,11,8,7,9,16,27,13,20,16,123 OTUs specific to day 1, 3, 5, 7, 9, 12, 15, 18, 22, 26, 30, 35, and 40 (Figure 1a). For the calves in diarrheal group also exhibited 34 shared OTUs at different time points with additional 24, 15,22,12,12,4,8,10,16,27,20,29,108 OTUs specific to day 1, 3, 5, 7, 9, 12, 15, 18, 22, 26, 30, 35, and 40 (Figure 1b). Additionally, the α-diversity indices, including ACE, Chao1, PD-whole-tree, Shannon, Simpson were compared at adjacent time points for the calves in both healthy and diarrheal Groups, revealing no significant differences for all the calves during pre-weaning period (Supplementary File3).
The PCoA results indicated that the microbial profiles of the calves in the healthy group exhibited similar structures on days 1, 3, 5, 7, and 9, as well as on days 15, 18, 22, 26, 30, 35, and 40. However, microbial profile of calves on day 12 intersected with the two distinct clusters (Figure 2a). For calves in diarrheal group, the microbial profiles of the calves was similar on days 1, 3, and 5, as well as on days 15, 18, 22, 26, 30, 35, and 40. However, microbial profiles of the calves on days 7, 9, and 12 intersected with the above two time clusters (Figure 2b).
3.2. Taxonomic Composition of Healthy Simmental Calves during Pre-Weaning Period
For healthy Simmental calves, regardless of ages, 20 bacterial phyla were identified in the stool samples of calves throughout the entire experiment. The top 10 bacterial phyla were Firmicutes (39.586±1.773%), Bacteroidota (29.488±1.821%), and Proteobacteria (25.959±2.487%) Fusobacteriota (2.538±0.539%), Verrucomicrobiota (0.772±0.204%), Actinobacteriota (0.454±0.098%), Desulfobacterota (0.453±0.072%), Cyanobacteria (0.400±0.124%), Campylobacterota (0.267±0.105%), and Elusimicrobiota (0.062±0.023%) (Figure 3a). Additionally, 328 bacterial genera were identified at the genus level. The 10 predominant bacterial genus were Escherichia_Shigella (20.628±2.483%), Bacteroides (19.208±1.504%), Faecalibacterium (4.374±0.627%), Alloprevotella (3.330±0.559%), Clostridium_sensu_stricto_1 (2.367±0.639%), Streptococcus (2.269±0.520%), Fusobacterium (2.538±0.539%), Butyricicoccus (2.433±0.429%), Lachnoclostridium (2.338±0.253%), and Parabacteroides (2.081±0.300%) (Figure 3b).Moreover, 503 bacterial species were identified, with E.coli (20.628±2.483%), B.fragilis (8.023±1.309%), B.vulgatus (6.589±0.830%), F.prausnitzii (3.837±0.602%), S.pasteurianus (2.103±0.520%), B.pullicaecorum (2.308±0.432%), C.perfringens (1.717±0.468%), F.mortiferum (1.939±0.501%), R.gnavus_CC55_001C (1.769±0.355%), and C.kerstersii (1.447±0.433%) being the 10 predominant bacterial species (Figure 3c). Furthermore, no significant difference in the relative abundance of the top five bacterial species between adjacent time points were detected (Figure 3d).
3.3. Taxonomic Composition of Simmental Calves Infected with Diarrhea during Pre-Weaning Period
For the calves in the diarrheal group, 17 phyla were detected throughout the experiment regardless of ages. The top 10 bacterial phyla included Firmicutes (41.492±2.317%), Bacteroidota (29.384±2.341%), Proteobacteria (21.615±3.043%), Fusobacteriota (3.815±1.009%), Verrucomicrobiota (2.170±0.564%), Actinobacteriota (0.722±0.188%), Cyanobacteria (0.480±0.133%), Desulfobacterota (0.292±0.066%), Elusimicrobiota (0.119±0.0019%), and Campylobacterota (0.020±0.016%) (Figure 4A). Additionally, 135 bacterial genera were identified, with Bacteroides (20.756±1.895%) being the most dominant genera, followed by Bacteroides (20.756±1.895%), Escherichia_Shigella (17.129±3.057%), Faecalibacterium (4.433±0.689%), Fusobacterium (3.813±1.099%), Lactobacillus (2.971±1.265%), Lachnoclostridium (2.383±0.989%), Clostridium_sensu_stricto_1 (2.654±0.342%), Akkermansia (2.164±0.564%), [Ruminococcus]_gnavus_group (1.646±0.374%), and Tyzzerella(1.751±0.276%) (Figure 4b). At the species level, 458 bacterial species were identified. The top 10 annotated species were E.coli (17.129±3.057%), B.vulgatus (7.695±1.051%), B.fragilis (5.308±1.339%), F.prausnitzii (3.449±0.536%), F.mortiferum (2.598±0.977%), L.amylovorus (2.638±1.186%), A.muciniphila (2.164±0.564%), B.uniformis (1.909±0.351%), R.bacterium (1.713±0.432%), and R.gnavus_CC55_001C (1.451±0.345%) (Figure 4c).
Moreover, the significant changes in the relative abundances of the top five bacterial species were compared between adjacent time points. The results showed significant differences in the relative abundance of B.fragilis between day 5 (13.000±12.460%) and day 7 (6.868±6.322%) (p=0.047), and F.prausnitzii exhibited significant differences between day 12 (4.573 ± 1.396%) and day 15 (6.377 ± 2.011%) (p=0.046) (Figure 4d). No significant differences were observed in the relative abundance of other bacteria at the species level.
3.4. Similarity Analysis of Gut Microbiota of Simmental Calves during Pre-Weaning Period
In our study, Anosim analysis was applied to conduct pairwise comparisons of the gut microbial structures of pre-weaning calves between the healthy and diarrheal groups at 13 time points. For calves in healthy group, the gut microbiota exhibited similarity from days 1- 9 (r<0.5), and days 15- 40 (r<0.5). However, microbial structure of calves on day 12 differs with these two age groups (Table 1). In the diarrheal group, similar gut microbial patterns were observed in two age groups, days 1-15 (r<0.5) and days 18-35 (r<0.5) (Table 2).
3.5. Differential Analysis of Intestinal Microbiota of Simmental Calves between Healthy and Diarrheal Groups
In this study, we conducted a comparative analysis to identify potential difference in bacterial taxonomy between the calves in the healthy group and those in the diarrheal group from birth through the stage of diarrheal resolution (1 d, 3 d, 5 d, 7 d, and 9 d). Notably, on day 1 after birth, significant differences were observed in the relative abundances of three bacterial species between calves in healthy and diarrheal groups. Specifically, L.mucosae (Diarrheal group:0, Healthy group:0.035±0.021%, p=0.046) and P.mirabilis (Diarrheal group:0.073±0.071%, Healthy group:0.043±0.01%, p=0.032) exhibited higher relative abundances in diarrheal group than healthy group. Conversely, L.johnsonii showed lower relative abundance in diarrheal group than healthy group (Diarrheal group : 0.012±0.010%, Healthy group : 0.030±0.019%, p=0.012) (Table 3).
On day 3 following birth, significant differences were observed in the relative abundances of four bacterial species between both groups. Specifically, the relative abundance of F.prausnitzii was higher in calves from diarrheal group than healthy group (Diarrheal group:1.619±1.461%, Healthy group:0.029±0.023%, p=0.014), While three other bacterial species displayed significant lower relative abundance in darrheal group compared to healthy group, P.russellii (Diarrheal group:0.023±0.018%, Healthy group : 1.330±0.720%,p=0.008), E.ramosum (Diarrheal group:0.778±0.416%, Healthy group:2.283±0.611%, p=0.023), and L.johnsonii (Diarrheal group : 0, Healthy group:1.208±1.113%, p=0.023) (Table 3).
On day 5 after birth, when diarrheal occurred, the relative abundance of three bacterial species showed significantly lower relative abundances in diarrheal group compared with those in healthy group, F.umbilicata (Diarrheal group:0, Healthy group:0.269±0.159%, p=0.010), G.genomosp. 3 (Diarrheal group:0.057±0.037%, Healthy group:2.764±1.556%, p=0.019), and C.pharyngocola (Diarrheal group:0.309±0.309%, Healthy group:1.376±0.609%, p=0.040) (Table 3).
On day 7 after birth,the relative abundance of five bacterial species exhibited markedly higher relative abundances in diarrheal group compared to those in healthy group. Namely, S.mitis (Diarrheal group:0.380±0.380%, Healthy group : 0.005±0.008%, p=0.013), E.ramosum (Diarrheal group : 0.638±0.252%, Healthy group : 0.087±0.053%, p=0.019), P.mirabilis (Diarrheal group : 0.180±0.099%, Healthy group : 0.026±0.017%, p=0.019), A.muciniphila (Diarrheal group : 2.967±2.760%, Healthy group : 0.004±0.003%, p=0.040), and L.amylovorus (Diarrheal group : 0.011±0.004%, Healthy group : 0.002±0.001%, p=0.040) (Table 3).
After the recovery phase of calf diarrheal, specifically on day 9 after birth, significant differences were noted in the relative abundance of ten bacterial species between both groups. Five bacterial species exhibited higher relative abundances in diarrheal group compared to those in healthy group, including L.murinus (Diarrheal group : 8.123±7.134%, Healthy group : 0.128±0.127%, p=0.010), F.necrophorum (Diarrheal group : 0.071±0.042%, Healthy group : 0.001±0.001%, p=0.012), A.muciniphila (Diarrheal group : 0.159±0.096%, Healthy group : 0.008±0.006%, p=0.020), B.vulgatus (Diarrheal group : 16.951±7.283%, Healthy group : 4.473±2.922%, p=0.020), and K.pneumoniae (Diarrheal group : 1.384±1.285%, Healthy group : 0.003±0.002%, p=0.028). While the relative abundances of five other bacterial speceis in diarrheal group were significantly lower than those in healthy group: P.russellii (Diarrheal group : 0.032±0.026%, Healthy group : 6.153±5.149%, p=0.004) S.mitis (Diarrheal group : 0.012±0.018%, Healthy group : 8.893±0.003%, p=0.012), P.dorei (Diarrheal group : 0.003±0.003%, Healthy group : 0.005±0.005%, p=0.020), E.coli (Diarrheal group : 27.465±11.439%, Healthy group : 52.753±7.410%, p=0.039), and L.johnsonii (Diarrheal group : 0.262±0.253%, Healthy group : 2.409±1.656%, p=0.020) (Table 3).
3.6. Prediction of Intestinal Microbial Functions of Simmental Calves
In total, 364 KEGG pathways (the metabolic pathways with CPM > 5, in at least 50% of the animals within each treatment group) were predicted with fecal samples of healthy neonatal calves. However, 25 pathways were removed as exogenous pathways. Subsequently, 181 pathways were utilized for further analyses (Supplementary File2). The 181 identified KEGG pathways belonged to 4 first level KEGG functions, which were “Cellular Processes” (2.900±0.029%), “Environmental Information Processing” (7.001%±0.104%), “Genetic Information Processing” (7.941%±0.083%), and “Metabolism” (78.293%±0.093%) (Figure 5a). Additionally, 27 secondary level of KEGG functions were identified, with “Global and overview maps” (41.855%±0.084%), “Carbohydrate metabolism” (10.202%±0.056%), “Amino acid metabolism” (6.479%±0.033%), “Membrane transport” (4.313%±0.073%), “Metabolism of cofactors and vitamins” (4.136%±0.017%), “Energy metabolism” (3.999%±0.015%), “Nucleotide metabolism” (3.752%±0.025%), “Translation” (3.309%±0.044%), “Replication and repair” (2.971%±0.029%), and “Signal transduction” (2.649%±0.039%) being the top 10 functions (Figure 5b). Furthermore, the top 10 KEGG pathways were: “Metabolic pathways” (17.483±0.031%), “Biosynthesis of secondary metabolites” (7.743±0.021%), “Biosynthesis of antibiotics” (5.684±0.023%), “Microbial metabolism in diverse environments” (4.422±0.032%), “Biosynthesis of amino acids” (3.942±0.031%), “ABC transporters” (3.230±0.035%), “Carbon metabolism” (2.719±0.005), “Two-component system” (2.312±0.033%), “Ribosome” (2.274±0.024%),and “Purine metabolism” (2.162±0.011%)(Figure 5c).
3.7. Differences in Microbial Functions of Calves between Healthy and Diarrheal Groups
The second level KEGG functions were compared between calves in healthy and diarrheal groups on day 5 d, 9d, 15 d, 30 d, 35 d, and 40 d. Specifically, on day 5 after birth, the three functions demonstrated significant disparities between both groups. Among these, two functions exhibited significantly higher relative content in calves in diarrheal group than calves in healthy group, namely “Environmental adaptation” (Diarrheal group:0.156±0.010%, Healthy group:0.142±0.009%,p=0.046) and “Cellular community-prokaryotes” (Diarrheal group:1.588±0.162%, Healthy group:1.361±0.124%, p=0.047).Conversely, the relative content of the “Folding, sorting and degradation” function was notably lower in diarrheal group calves compared to healthy group (Diarrheal group:1.401±0.072%, Healthy group:1.505±0.069%,p=0.048). On day 9 after birth, following diarrheal recovery, a notable difference was observed between the two groups in one function, with the relative content significantly higher in diarrheal group compared to healthy group, “Endocrine system” (Diarrheal group:0.527±0.045%, Healthy group:0.460±0.057%, p=0.044). Similarly, on day 15, a significant difference was noted in one function between both groups, namely “Cell motility” (Diarrheal group:0.369±0.153%, Healthy group:0.587±0.177%, p=0.043). Likewise, on day 30, a single function showed a significant disparity between the groups, namely “Metabolism of terpenoids and polyketides” (Diarrheal group:1.034±0.045%, Healthy group:1.104±0.030%, p=0.016). Additionally, on day 35, one function exhibited a significant difference between the two groups: “Folding, sorting and degradation” (Diarrheal group:1.587±0.036%, Healthy group:1.527±0.028%, p=0.012). On day 40, significant differences were observed in seven functions between the two groups. The relative content of four functions was notably higher in diarrheal group calves than healthy group: “Transcription” (Diarrheal group:0.173±0.011%, Healthy group:0.153±0.009%, p=0.012), “Replication and repair” (Diarrheal group:3.173±0.107%, Healthy group:2.975±0.160%, p=0.025), “Nucleotide metabolism” (Diarrheal group:3.871±0.146%, Healthy group:3.676±0.152%, p=0.046), and “Translation” (Diarrheal group:3.628±0.119%, Healthy group:3.377±0.239%, p=0.036). Conversely, the relative content of three functions was significantly lower in diarrheal group calves than healthy group: “Energy metabolism” (Diarrheal group:3.927±0.107%, Healthy group:4.104±0.078%, p=0.013), “Amino acid metabolism” (Diarrheal group :6.500±0.150%, Healthy group:6.780±0.203%, p=0.031), and “Digestive system” (Diarrheal group:0.041±0.016%, Healthy group:0.063±0.013%, p<0.05) (Figure 6).
4. Discussion
For neonatal ruminants, nutrient digestion primarily occurs in the intestine because of underdeveloped rumen. The gut microbiota is crucial for host nutrition, absorption, metabolism, immune regulation, and gut health[23]. In this study, 14 pre-weaning Simmental calves were selected, and the stool samples were subjected to high-throughput sequencing of the V1-V9 region of the 16S rRNA gene. The microbial profiling revealed a diverse and dense microbial colonization of neonatal calves. Throughout the experimental period, the α-diversity indices, including Chao1, Simpson, and ACE showed no significant difference with ages in both healthy and diarrheal groups, which may be related to the individual variation or the limited number of pre-weaning calves. Inconsistent with our studies, significant difference was found on Chao1 and Shannon indices among different age groups of Holstein calves[24]. Such discrepancy may stem from breed differences between Holstein and Simmental calves or different sampling points.
For healthy Simmental calves, when considering about the changes of intestinal microbiota with ages during pre-weaning period through PCoA analysis, the structure of the gut microbe has changed. The intestinal microbial structure of calves was similar from days 1 to 9, and the microbial structure of calves showed transition on day 12, which is probably linked to the intestinal maturation and the changes in feed intake. However, the feed intake was not measured in this study and further investigations should be taken to explore the correlations between feed intake and intestinal microbial colonization. Meanwhile, the microbial structure of calves was also similar from days 15-40, which may indicate that the microorganisms tend to mature and stabilize during this period. Moreover, our findings were similar to those of previous studies on Holstein calves, which also exhibited the age-related changes, with the microbial structures on day 21 and 42 resembling each other but differing from that on day 7 of pre-weaning calves[24]. Both studies showed a gradual maturation process of intestinal microbiota, but the transition time to maturity and stability may be closely related to the breed of calves and the sampling time point.
Similar to the results of previous studies on calf fecal intestinal microbiota targeting the V1–V4 hypervariable regions of the 16S rRNA gene through 454 pyrosequencing[25], Firmicutes, Bacteroidetes, and Proteobacteria were the dominant phyla of pre-weaning Simmental calves, constituting over 90% of the total bacterial composition. Moreover, E.coli, B.fragilis, and B.vulgatus emerged as the dominant species, which was consistent with the studies on the intestinal microbial dynamic changes of pre-weaning Holstein calves[26]. Escherichia, the facultative anaerobes, could help to create a conducive environment for anaerobic bacteria colonization in the early stage of calf birth[27]. Therefore, the large abundance of Escherichia is closely related to intestinal oxygen consumption. Additionally, nutrients could be metabolized by Bacteroides to produce acetic acid, which could be served as a substrate for Butyricoccus and Megamonas to produce butyric and propionic acids[28]. Butyrate was the crucial energy sources for intestinal epithelial cells[29], as well as inhibiting pro-inflammatory cytokine signaling pathways[30]. E.coli, B.fragilis, and B.vulgatus all play important roles in the gut, hence the large abundance of their colonization during early life may contribute to the maturation and health of intestinal function.
During days 5–7, coinciding with the onset of diarrhea, the relative abundance of B.vulgatus notably decreased for the calves in diarrheal group. B.vulgatus could ferment carbohydrates to produce butyrate, which positively affected the gut health and energy acquisition of intestinal cells[31]. Therefore, the reduction of the relative abundance of B.vulgatus may influence the growth and health of pre-weaning calves infected with diarrhea. It was suggested that oral administration of F.prausnitzii significantly reduced the incidence of severe diarrhea and calf mortality in lactating Holstein calves, as well as promoting calves’ growth and intestinal health[32]. In our study, the relative abundance of Faecalibacterium prausnitzii in the gut microbiota of calves in diarrheal group was higher on day 15 compared with day 12.This indicated that the levels of Faecalibacterium prausnitzii were elevated after the calves were recovered from diarrhea, which is in consistence with previously publication that Faecalibacterium prausnitzii is closely related to intestinal health of neonatal calves. The comparison was conducted to assess the significant differences in the relative abundance of intestinal microbiota between calves in healthy and diarrheal groups at the same sample collection points. It has demonstrated that milk supplementated with Lactobacillus reuteri L81 and Lactobacillus johnsonii L29 enhanced the growth performance, immunity, and antioxidant capacity of pre-weaning Holstein calves, as well as reducing calves’ diarrhea[33]. Interestingly, higher relative abundance of L.johnsonii in the intestine of Simmental calves in healthy group on days 1, 3, and 9 was detected The probiotic effect of L.johnsonii may be closely related to the absence of diarrhea for the Simmental calves in Healthy group. Oral administration of F.prausnitzii is closely associated with the reduced rate of calf diarrhea[34]. In our study, the relative abundance of F.prausnitzii was significantly higher in the intestine of calves in healthy group at day 3 after birth, which may be related to the absence of diarrhea in this group. Meanwhile, Limosilactobacillus supplementation can alleviate the symptoms of ETEC K88-induced diarrhea in piglets by modulating macrophage phenotypes[35], therefore, the higher relative abudannce of Limosilactobacillus on day 3 of neonatal Simmental calves could be a contributor for the preventation of calf diarrhea. Therefore, L.johnsonii, F.prausnitzii, and Limosilactobacillus may play a certain role in the prevention of Simmental calves’ diarrhea and could be developed as a potential probiotic bacteria. E.coli is a potential pathogenic bacteria of causing neonatal calves’ diarrhea[36]. On day 9, when the calves has just recovered from diarrhea, the relative abundance of E. coli in diarrheal group significantly exceeded that in the healthy group, which may suggested that calves in diarrheal group may have a greater risk of recurrent diarrhea.
Inconsistent with the predicted main intestinal functions of Holstein calves[24], the core functions of pre-weaning Simmental calves not only included functions related to nutrients metabolism, but also included “Biosynthesis of antibiotics”, which is likely attributable to breed specific productive properties. It may also suggest that gut microbes of pre-weaning Simmental calves may tend to protect intestinal health by producing antibiotics. Meanwhile, the difference in intestinal microbial functions between calves in healthy and diarrheal groups were compared. Significantly higher relative abundance of "Environmental adaptation" function were detected on day 5 of calves in healthy group, which may indicate that intestinal microbiota of healthy calves had stronger adaptability to intestinal environment and the homeostasis is not easily destroyed[37]. When the calves were on day 40, the relative abundance of “energy metabolism” and “amino acid metabolism” functions in the healthy group exceeded that of the diarrheal group. These two functions are important functions in the hindgut of pre-weaning calves, which could provide energy for the host[38]. Therefore, our results may indicate that the gut microbiome of calves in healthy group may provide more energy and nutrients for the host compared to the calves in diarrheal group.
5. Conclusions
This study investigated the dynamic changes of intestinal microbiota of pre-weaning Simmental calves and predicted the functions of their gut microbiota. It suggested that Escherichia coli, Bacteroides fragilis, and Bacteroides vulgatus were the dominant bacterial species.“Metabolic pathways” “Biosynthesis of secondary metabolites” “Biosynthesis of antibiotics” “Microbial metabolism in diverse environments” and “Biosynthesis of amino acids” were the major intestinal functions. Moreover, pronounced dynamic changes of intestinal microbiota of the calves in both healthy and diarrheal groups with increasing of ages were detected. Furthermore, significantly higher relative abundance of Lactobacillus johnsonii, Faecalibacterium prausnitzii, and Limosilactobacillus were detected in the gut of the calves in healthy group, which may be closely related to absence of calves’ diarrhea. Our study could offer valuable insights into the prevention of diarrhea and the development of probiotics for Simmental calves.
5. Conclusions
This study investigated the dynamic changes of intestinal microbiota of pre-weaning Simmental calves and predicted the functions of their gut microbiota. It suggested that Escherichia coli, Bacteroides fragilis, and Bacteroides vulgatus were the dominant bacterial species. “Biosynthesis of secondary metabolites” “Biosynthesis of antibiotics” “Microbial metabolism in diverse environments” and “Biosynthesis of amino acids” were the major intestinal functions. Moreover, pronounced dynamic changes of intestinal microbiota of the calves in both healthy and diarrheal groups with increasing of ages were detected. Furthermore, significantly higher relative abundance of L.johnsonii, F.prausnitzii, and Limosilactobacillus were detected in the gut of the calves in healthy group, which may be closely related to absence of calves’ diarrhea. Our study could offer valuable insights into the prevention of diarrhea and the development of probiotics for Simmental calves.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary File1,.Supplementary File2, Supplementary File3.
Author Contributions
J.Y.,Y.S. and Q.W. designed this experiment; Q.W., M.N., S.J.,M.S.,S.P and S.G. performed the animal experiment; Q.W. and W.D. participated in results and statistics; Q.W. and Y.S. were responsible for revising the article; J.Y.and Y.S. supervised all processes from performing the experiment to writing the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Program for Program for National Thermal Science Foundation(M2142002; Inner Mongolia, China),Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (NJYT22053; Inner Mongolia, China),Science Innovation Project for Master Students of Inner Mongolia Autonomous Region (S20231212Z; Inner Mongolia, China).
Institutional Review Board Statement
The animal study protocol was approved by the Experimental Animal Welfare and Ethics Committee of the College of Animal Science and Technology of Inner Mongolia Minzu University (protocol code: No. 2022058).
Informed Consent Statement
Not applicable.
Data Availability Statement
The data that support the findings of this study are available from the corresponding authors upon reasonable request.
Acknowledgments
We would like to thank Chi Feng for polishing the manuscript, and thank Di Sun for her contribution to this article. All the above involved personnel have confirmed and agreed to the content.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Wang, Y.; Wang, Z.; Hu, R.; Peng, Q.; Xue, B.; Wang, L. Comparison of carcass characteristics and meat quality between Simmental crossbred cattle, cattle-yaks and Xuanhan yellow cattle. J Sci Food Agric. 2021, 101, 3927–3932. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Shen, Y.; You, L; Zhang, Y; Su, Z; Peng, G; Deng, J; Zuo, Z; Zhong, Z; Ren, Z; Yu, S; Zong, X; Zhu, Y; Cao, S. Pueraria lobata polysaccharides alleviate neonatal calf diarrhea by modulating gut microbiota and metabolites. Front Vet Sci. 2023, 9, 1024392. [Google Scholar] [CrossRef] [PubMed]
- Tamrat, H.; Mekonnen, N.; Ferede, Y.; Cassini, R.; Belayneh, N. Epidemiological study on calf diarrhea and coccidiosis in dairy farms in Bahir Dar, North West Ethiopia. Ir Vet J. 2020, 73, 14. [Google Scholar] [CrossRef] [PubMed]
- Kim, HS.; Whon, TW.; Sung, H.; Jeong, YS.; Jung, ES.; Shin, N.R.; Hyun, D.W.; Kim, P.S.; Lee, J. Y, Lee, C.H, Bae, J.W. Longitudinal evaluation of fecal microbiota transplantation for ameliorating calf diarrhea and improving growth performance. Nat Commun. 2021, 12, 161. [Google Scholar] [CrossRef] [PubMed]
- Du, Y.; Gao, Y.; Hu, M.; Hou, J.; Yang, L.; Wang, X.; Du, W.; Liu, J.; Xu, Q. Colonization and development of the gut microbiome in calves. J Anim Sci Biotechnol. 2023, 14, 46. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Tu, Y.; Zhang, N.; Wang, S.; Zhang, F.; Suen, G.; Shao, D.; Li, S.; Diao, Q. Multiomics analysis reveals the presence of a microbiome in the gut of fetal lambs. Gut. 2021, 70, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Guzman, CE.; Wood, J.L.; Egidi, E.; White-Monsant, A.C.; Semenec, L.; Grommen, S.V.H.; Hill-Yardin, E.L.; De Groef, B.; Franks, A.E. A pioneer calf foetus microbiome. Sci Rep. 2020, 10, 17712. [Google Scholar] [CrossRef]
- Schaedler,R. W.; Dubos,R.; Costello, R. The development of the bacterial flora in the gastrointestinal tract of mice. J Exp Med. 1965, 122, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Uyeno, Y.; Sekiguchi, Y.; Kamagata, Y. rRNA-based analysis to monitor succession of faecal bacterial communities in Holstein calves. Lett Appl Microbiol. 2010, 51, 570–577. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Griebel, P.J.; Guan, le.L. Taxonomic identification of commensal bacteria associated with the mucosa and digesta throughout the gastrointestinal tracts of preweaned calves. Appl Environ Microbiol. 2014, 80, 2021–2028. [Google Scholar] [CrossRef]
- Vlková, E.; Trojanová, I.; Rada, V. Distribution of bifidobacteria in the gastrointestinal tract of calves. Folia Microbiol (Praha). 2006, 51, 325–328. [Google Scholar] [CrossRef] [PubMed]
- Guo,S.H.H.Study on intestinal flora diversity and fecal metabolome of calves with diarrhea.2021. Ningxia university, MA thesis.
- Penati, M.; Sala, G.; Biscarini, F.; Boccardo, A.; Bronzo, V.; Castiglioni, B.; Cremonesi, P.; Moroni, P.; Pravettoni, D.; Addis, M.F. Feeding Pre-weaned Calves With Waste Milk Containing Antibiotic Residues Is Related to a Higher Incidence of Diarrhea and Alterations in the Fecal Microbiota. Front Vet Sci. 2021, 8, 650150. [Google Scholar] [CrossRef]
- Lépine, A.F.P.; de Wit, N.; Oosterink, E.; Wichers, H.; Mes, J.; de Vos, P. Lactobacillus acidophilus Attenuates Salmonella-Induced Stress of Epithelial Cells by Modulating Tight-Junction Genes and Cytokine Responses. Front Microbiol. 2018, 9, 1439. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Mustapha, A.; Lin, M.; Zheng, G. Biocontrol of the internalization of Salmonella enterica and Enterohaemorrhagic Escherichia coli in mung bean sprouts with an endophytic Bacillus subtilis. Int J Food Microbiol. 2017, 250, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Memon, F.U.; Yang, Y.; Leghari, I.H.; Lv, F.; Soliman, A.M.; Zhang, W.; Si, H. Transcriptome Analysis Revealed Ameliorative Effects of Bacillus Based Probiotic on Immunity, Gut Barrier System, and Metabolism of Chicken under an Experimentally Induced Eimeria tenella Infection. Genes. 2021, 12, 536. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Nie, C.; Luo, R.; Qi, F.; Bai, X.; Chen, H.; Niu, J.; Chen, C.; Zhang, W. Effects of Multispecies Probiotic on Intestinal Microbiota and Mucosal Barrier Function of Neonatal Calves Infected With E. coli K99. Front Microbiol. 2022, 12, 813245. [Google Scholar] [CrossRef] [PubMed]
- Rivière, A.; Selak, M.; Lantin, D.; Leroy, F.; De Vuyst, L. Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front Microbiol. 2016, 7, 979. [Google Scholar] [CrossRef] [PubMed]
- Stöber,M.“Kennzeichen, Anamnese, Grundregeln der Untersuchungstechnik, Allgemeine Untersuchung,” in Die klinische Untersuchung des Rindes, eds G. Dirksen, H. -D. Grunder, M. Stöber, Stuttgart (Erlangen: Enke Verlag), 2012, 75–141.
- Lesmeister,KE. ; Heinrichs,A.J.; Effects of corn processing on growth characteristics, rumen development, and rumen parameters in neonatal dairy calves. J Dairy Sci. 2004, 87, 3439–3450. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, MR.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat Biotechnol. Erratum in: Nat Biotechnol. 2019 Sep;37(9):1091. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–6. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, P.; Dingkao, R.; Du, M.; Ahmad, A.A.; Liang, Z.; Zheng, J.; Shen, J.; Yan, P.; Ding, X. Fecal Microbiota Dynamics Reveal the Feasibility of Early Weaning of Yak Calves under Conventional Grazing System. Biology. 2021, 11, 31. [Google Scholar] [CrossRef]
- Song, Y.; Malmuthuge, N.; Steele, M.A.; Guan, L.L. Shift of hindgut microbiota and microbial short chain fatty acids profiles in dairy calves from birth to pre-weaning. FEMS Microbiol Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Klein-Jöbstl, D.; Schornsteiner, E.; Mann, E.; Wagner, M.; Drillich, M.; Schmitz-Esser, S. Pyrosequencing reveals diverse fecal microbiota in Simmental calves during early development. Front Microbiol. 2014, 5, 622. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, Y.; Huang, K.; Yang, B.; Zhang, Y.; Yu, Z.; Wang, J. Fecal microbiota dynamics and its relationship to diarrhea and health in dairy calves. J Anim Sci Biotechnol. 2022, 13, 132. [Google Scholar] [CrossRef] [PubMed]
- Mayer, M.; Abenthum, A.; Matthes, JM.; Kleeberger, D.; Ege, M.J.; Hölzel, C.; Bauer, J.; Schwaiger, K. Development and genetic influence of the rectal bacterial flora of newborn calves. Vet Microbiol. 2012, 161, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; Collins, MD. 16S rRNA gene similarities indicate that Hallella seregens (Moore and Moore) and Mitsuokella dentalis (Haapsalo et al.) are genealogically highly related and are members of the genus Prevotella: emended description of the genus Prevotella (Shah and Collins) and description of Prevotella dentalis comb. nov. Int J Syst Bacteriol. 1995, 45, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Trachsel, J.; Humphrey, S.; Allen, H.K. Butyricicoccus porcorum sp. nov., a butyrate-producing bacterium from swine intestinal tract. Int J Syst Evol Microbiol. 2018, 68, 1737–1742. [Google Scholar] [CrossRef] [PubMed]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; LeLeiko, N.; Snapper, S.B.; Bousvaros, A.; Korzenik, J.; Sands, B.E.; Xavier, R. J, Huttenhower, C. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Nishida, A.; Fujimoto, T.; Fujii, M.; Shioya, M.; Imaeda, H.; Inatomi, O.; Bamba, S.; Sugimoto, M.; Andoh, A. Reduced Abundance of Butyrate-Producing Bacteria Species in the Fecal Microbial Community in Crohn's Disease. Digestion. 2016, 93, 59–65. [Google Scholar] [CrossRef]
- Foditsch,C. ;Pereira,R.V.;Ganda,E.K.;Gomez,M.S.;Marques,E.C.;Santin,T.;Bicalho,R.C.Oral Administration of Faecalibacterium prausnitzii Decreased the Incidence of Severe Diarrhea and Related Mortality Rate and Increased Weight Gain in Preweaned Dairy Heifers. PLoS One. 2015, 10, e0145485. [Google Scholar]
- Li, Y.; Li, X.; Nie, C.; Wu, Y.; Luo, R.; Chen, C.; Niu, J.; Zhang, W. Effects of two strains of Lactobacillus isolated from the feces of calves after fecal microbiota transplantation on growth performance, immune capacity, and intestinal barrier function of weaned calves. Front Microbiol. 2023, 14, 1249628. [Google Scholar] [CrossRef] [PubMed]
- Foditsch, C.; Santos, T.M.; Teixeira, A.G.; Pereira, R.V.; Dias, J.M.; Gaeta, N.; Bicalho, R.C. Isolation and characterization of Faecalibacterium prausnitzii from calves and piglets. PLoS One. 2014, 9, e116465. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Feng, S.; Wang, Z.; He, J.; Zhang, Z.; Zou, H.; Wu, Z.; Liu, X.; Wei, H.; Tao, S. Limosilactobacillus mucosae-derived extracellular vesicles modulates macrophage phenotype and orchestrates gut homeostasis in a diarrheal piglet model. NPJ Biofilms Microbiomes. 2023, 9, 33. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, R.; Burdukiewicz, M.; Schierack, P. A systematic review and meta-analysis of the epidemiology of pathogenic Escherichia coli of calves and the role of calves as reservoirs for human pathogenic E. coli. Front Cell Infect Microbiol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, S.; Blanton, L.V.; Frese, S.A.; Charbonneau, M.; Mills, D.A.; Gordon, J.I. Cultivating healthy growth and nutrition through the gut microbiota. Cell. 2015, 161, 36–48. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- McNeil, NI. The contribution of the large intestine to energy supplies in man. Am J Clin Nutr. 1984, 39, 338–342. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Petalograms of intestinal microbial OTUs at different time points of pre-weaning Simmental (a) Petalograms of intestinal microbial OTUs at different time points of calves in healthy group (b) Petalograms of microbial OTUs at different time points of calves in diarrheal group.
Figure 1.
Petalograms of intestinal microbial OTUs at different time points of pre-weaning Simmental (a) Petalograms of intestinal microbial OTUs at different time points of calves in healthy group (b) Petalograms of microbial OTUs at different time points of calves in diarrheal group.
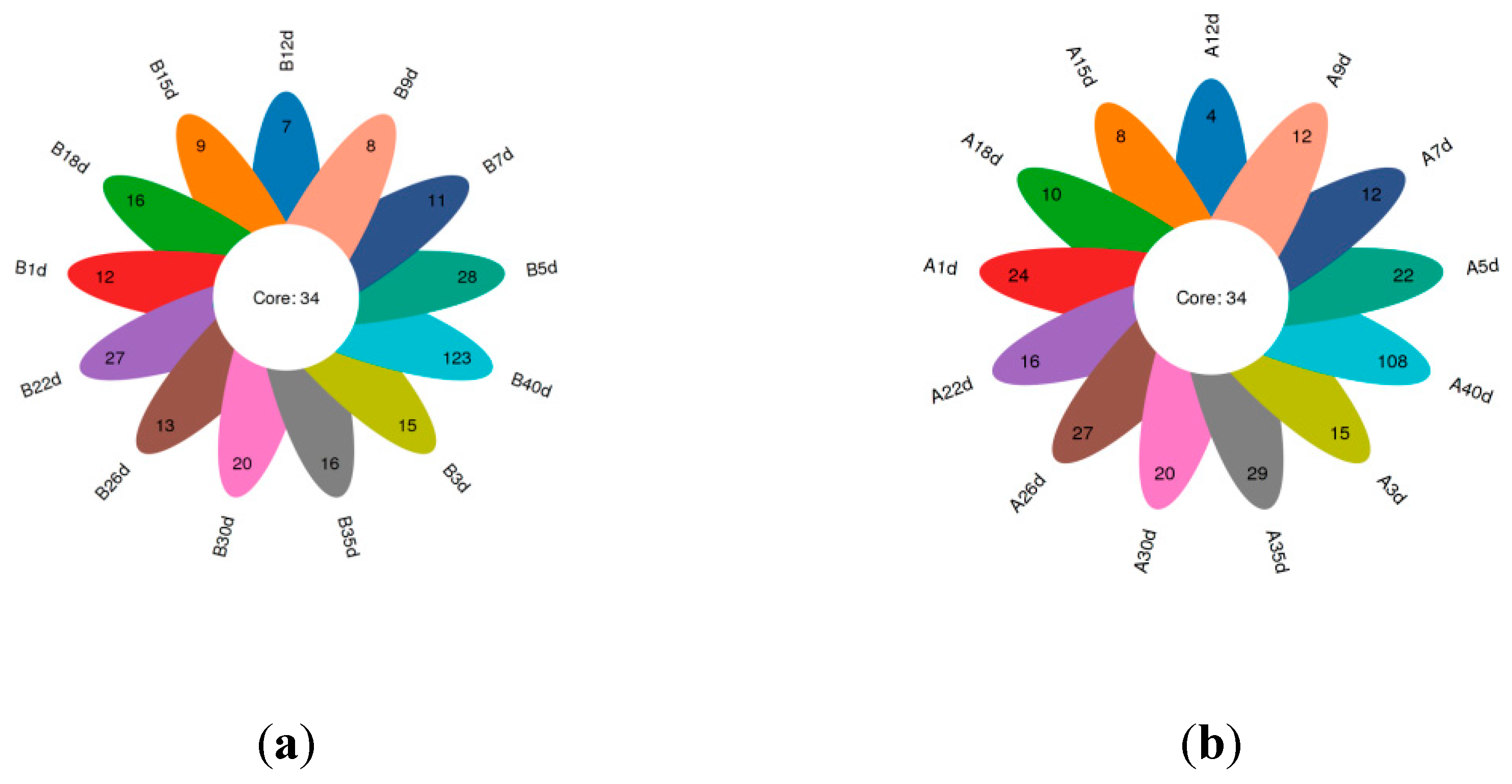
Figure 2.
Bacterial profiles of calves were compared across different sampling point using principal coordinate analysis (PCoA).(a)PCoA was generated using unweighted Jaccard for 13 different time points of calves in healthy groups. The two components explained 22.65% and 5.90% of the variance.(b)PCoA was generated using unweighted Jaccard for 13 different time points of calves in diarrheal groups. The two components explained 18.13% and 7.42% of the variance.
Figure 2.
Bacterial profiles of calves were compared across different sampling point using principal coordinate analysis (PCoA).(a)PCoA was generated using unweighted Jaccard for 13 different time points of calves in healthy groups. The two components explained 22.65% and 5.90% of the variance.(b)PCoA was generated using unweighted Jaccard for 13 different time points of calves in diarrheal groups. The two components explained 18.13% and 7.42% of the variance.
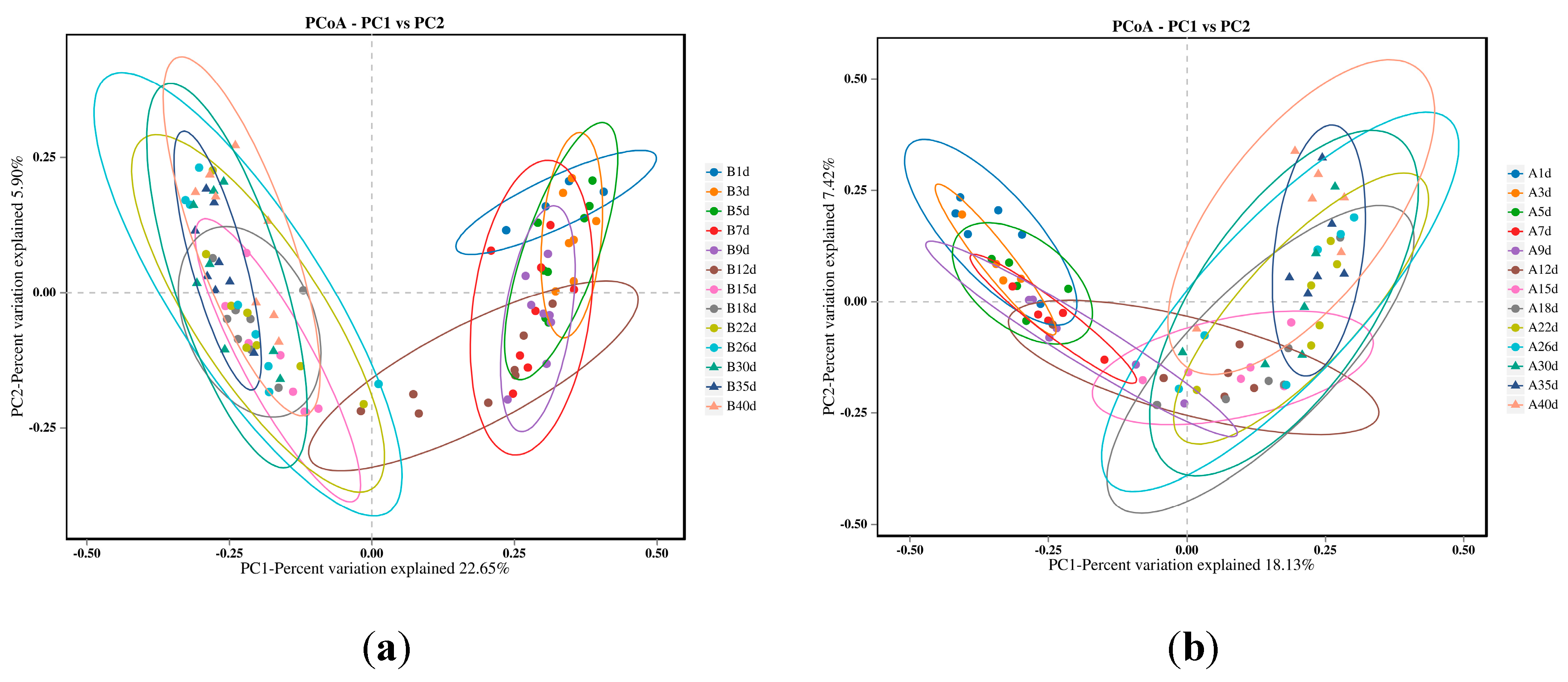
Figure 3.
Gut microbial composition of calves at different time points in healthy group. (a) The relative abundance of top 10 microbial composition at phylum level. (b) The relative abundance of top 10 microbial composition at genus level. (c) The relative abundance of top 10 microbial composition at species level. (d)Dynamic changes of the top 5 bacterial species of calves in healthy group with increasing of ages.
Figure 3.
Gut microbial composition of calves at different time points in healthy group. (a) The relative abundance of top 10 microbial composition at phylum level. (b) The relative abundance of top 10 microbial composition at genus level. (c) The relative abundance of top 10 microbial composition at species level. (d)Dynamic changes of the top 5 bacterial species of calves in healthy group with increasing of ages.
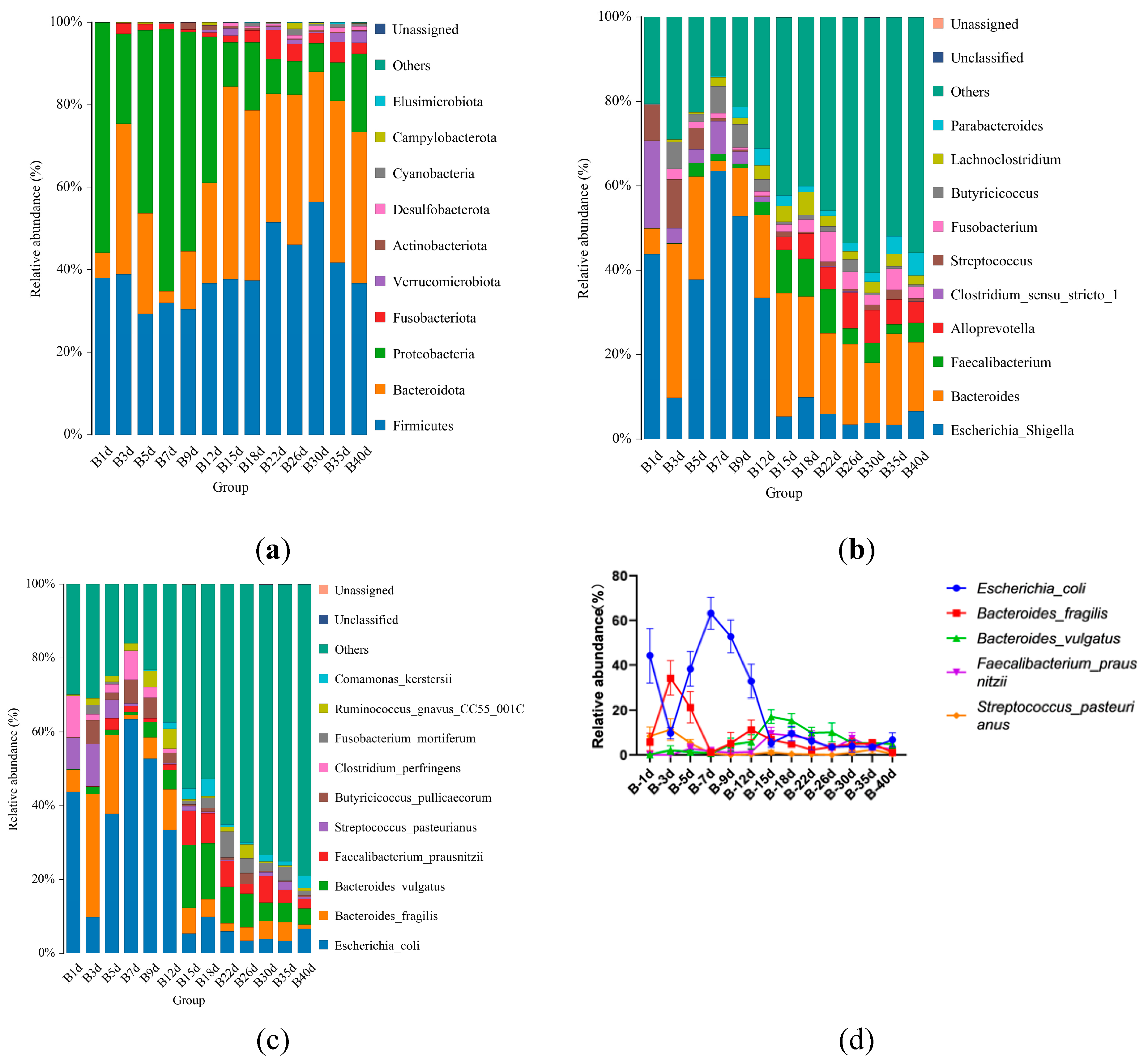
Figure 4.
Gut microbial composition of calves at different time points in diarrheal group. (a) The relative abundance of top 10 microbial composition at phylum level. (b) The relative abundance of top 10 microbial composition at genus level. (c) The relative abundance of top 10 microbial composition at species level. (d)Dynamic changes of the top 5 bacterial species of calves in diarrheal group with increasing of ages.
Figure 4.
Gut microbial composition of calves at different time points in diarrheal group. (a) The relative abundance of top 10 microbial composition at phylum level. (b) The relative abundance of top 10 microbial composition at genus level. (c) The relative abundance of top 10 microbial composition at species level. (d)Dynamic changes of the top 5 bacterial species of calves in diarrheal group with increasing of ages.
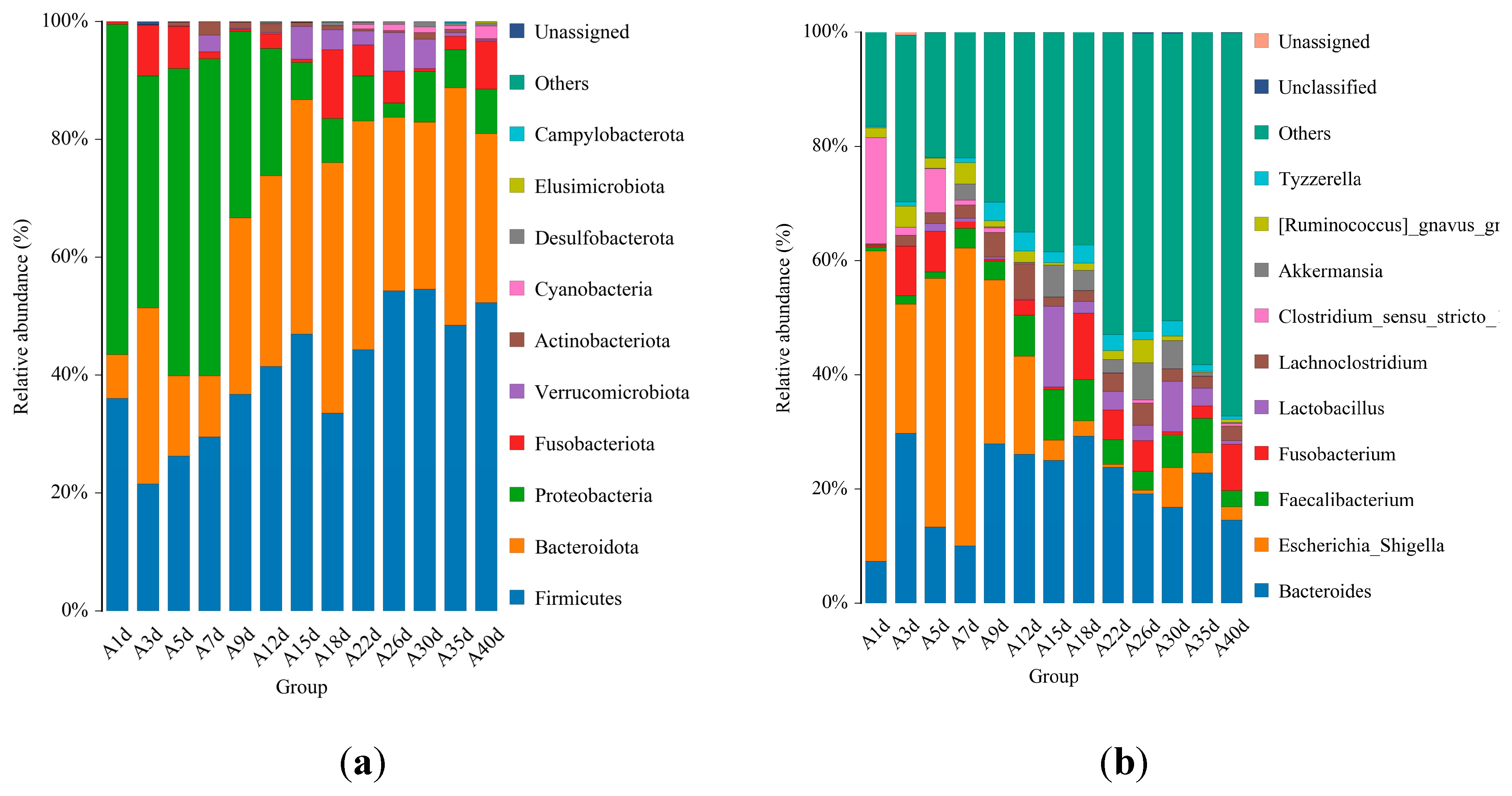
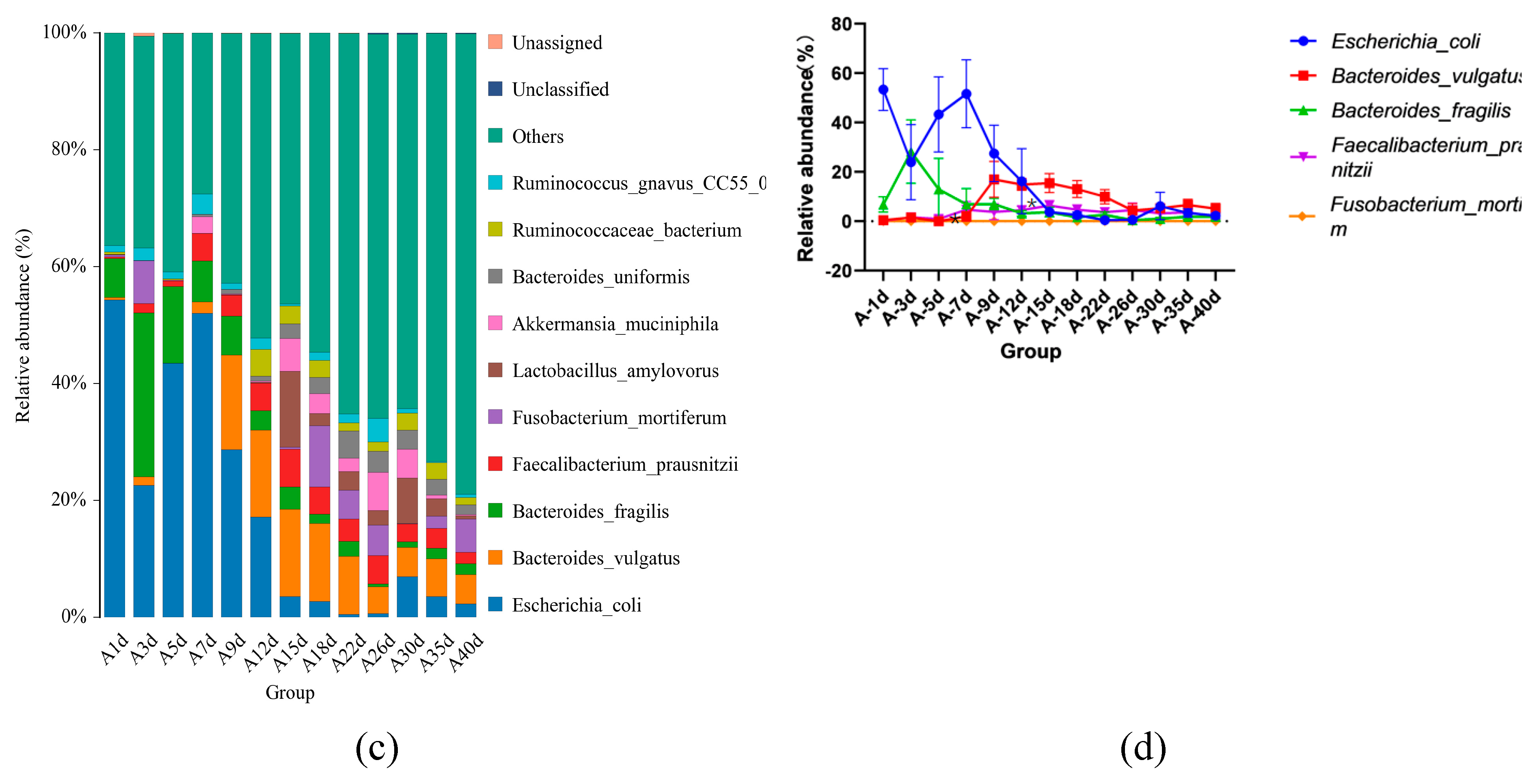
Figure 5.
Prediction of intestinal microbial functions of healthy pre-weaning Simmental calves. (A) The top 5 first level KEGG pathways.(B) The top 5 second level KEGG pathways. (C) The top 10 KEGG pathways.
Figure 5.
Prediction of intestinal microbial functions of healthy pre-weaning Simmental calves. (A) The top 5 first level KEGG pathways.(B) The top 5 second level KEGG pathways. (C) The top 10 KEGG pathways.
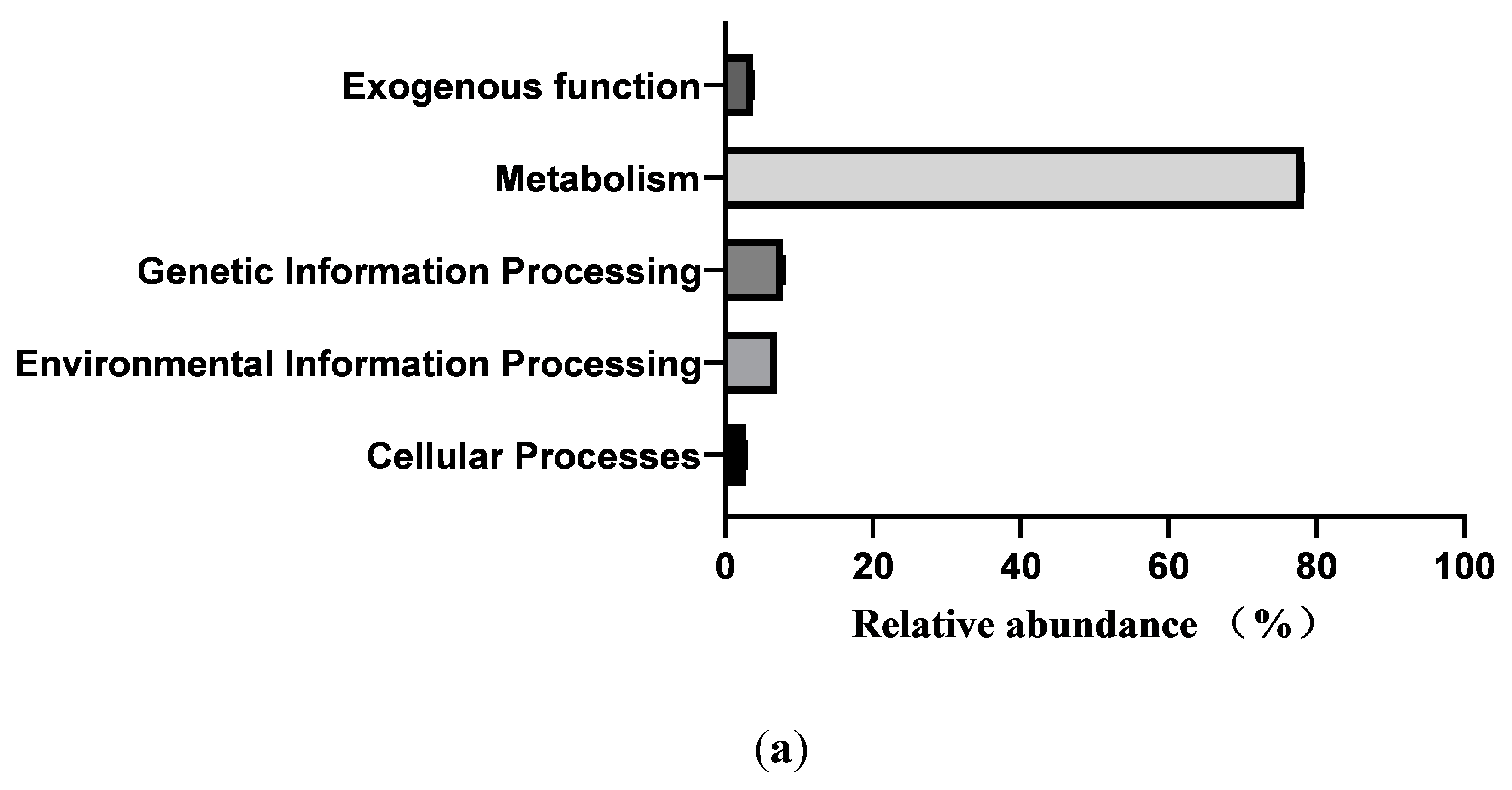
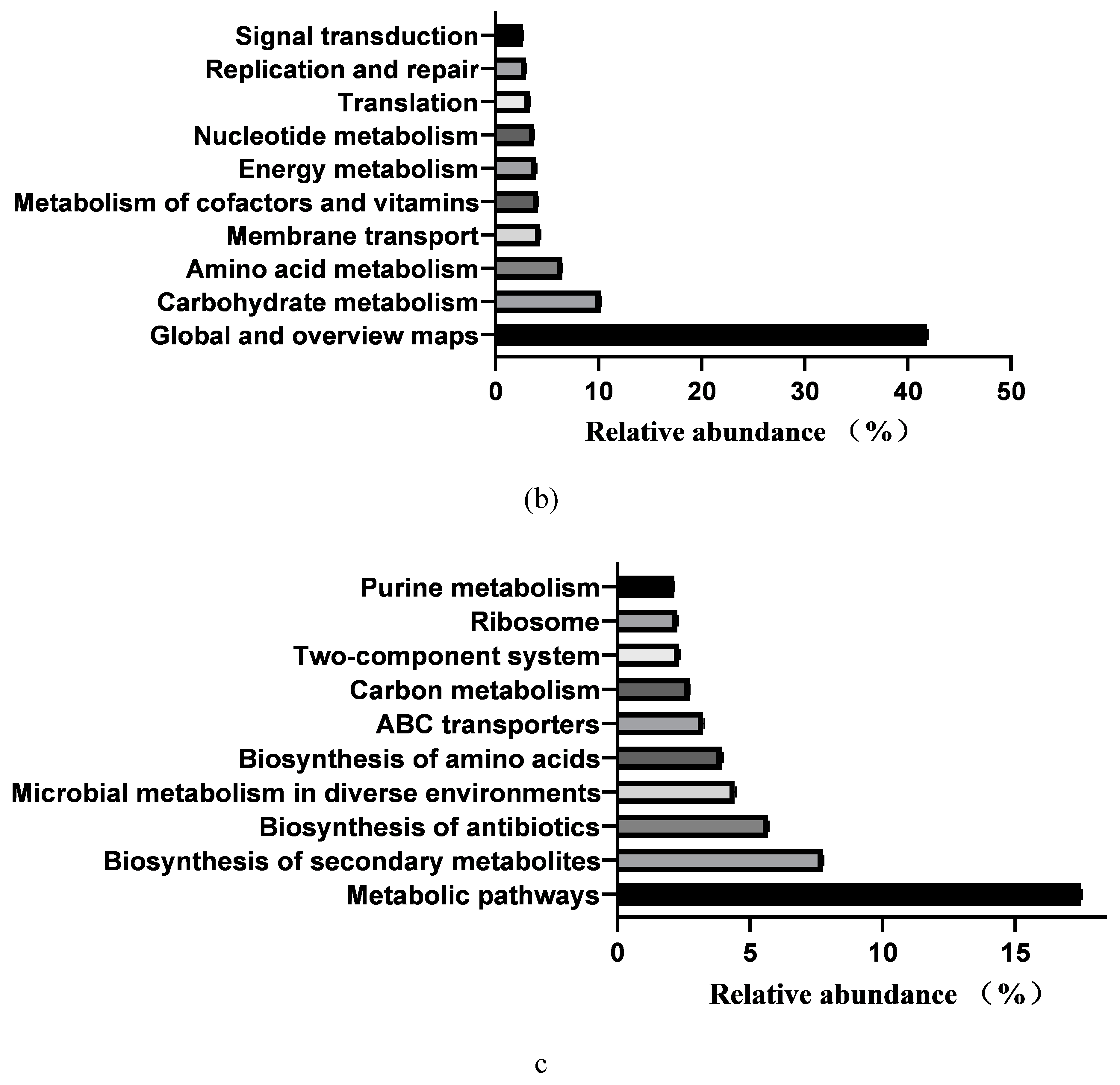
Figure 6.
Differential analysis of intestinal microbial functions of Simmental calves in healthy and diarrheal groups before weaning.
Figure 6.
Differential analysis of intestinal microbial functions of Simmental calves in healthy and diarrheal groups before weaning.
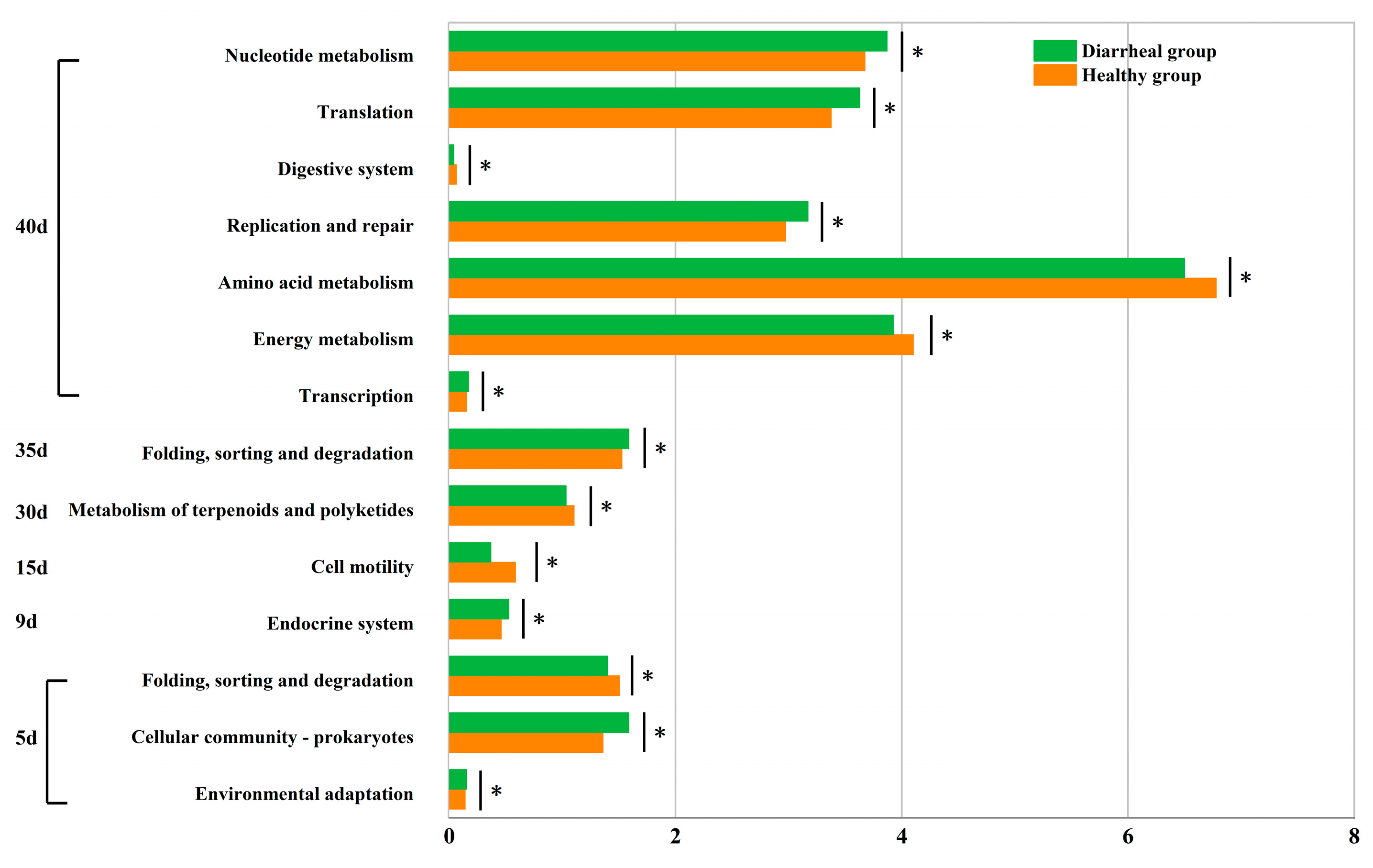

Table 1.
A matrix of R-values of pair-wise comparison of the fecal microbiota of calves in healthy group n=8 using ANOSIM.
Table 1.
A matrix of R-values of pair-wise comparison of the fecal microbiota of calves in healthy group n=8 using ANOSIM.
| Healthy group | 1d | 3d | 5d | 7d | 9d | 12d | 15d | 18d | 22d | 26d | 30d | 35d | 40d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1d | 0 | ||||||||||||
| 3d | r=0.398* | 0 | |||||||||||
| 5d | r=0.402* | r=-0.001 | 0 | ||||||||||
| 7d | r=0.340* | r=0.363* | r=0.328* | 0 | |||||||||
| 9d | r=0.718* | r=0.544* | r=0.491* | r=-0.021 | 0 | ||||||||
| 12d | r=0.861* | r=0.789* | r=0.667* | r=0.199* | r=0.137* | 0 | |||||||
| 15d | r=0.889* | r=1* | r=0.988* | r=0.871* | r=0.954* | r=0.646* | 0 | ||||||
| 18d | r=1* | r=1* | r=0.996* | r=0.947* | r=0.992* | r=0.845* | r=0.027 | 0 | |||||
| 22d | r=1* | r=0.985* | r=0.978* | r=0.925* | r=0.951* | r=0.801* | r=0.388* | r=0.122 | 0 | ||||
| 26d | r=0.985* | r=0.961* | r=0.962* | r=0.900* | r=0.934* | r=0.735* | r=0.314* | r=0.068 | r=-0.109 | 0 | |||
| 30d | r=1* | r=1* | r=0.996* | r=0.965* | r=0.996* | r=0.907* | r=0.525* | r=0.213* | r=-0.034 | r=-0.634 | 0 | ||
| 35d | r=1* | r=1* | r=0.999* | r=0.996* | r=1* | r=0.969* | r=0.824* | r=0.422* | r=0.181* | r=0.007 | r=-0.047 | 0 | |
| 40d | r=1* | r=0.998* | r=0.991* | r=0.980* | r=0.996* | r=0.906* | r=0.611* | r=0.413* | r=0.252* | r=0.014 | r=0.091 | r=0.064 | 0 |
*represents the pair-wise comparison with p-value<0.05. Those with r-value<0.5are blod With p-value<0.05, the fecal microbiotas of two ages were considered completely different at r-value>0.75; different at 0.5<r-value<0.75; tended to be different data 0.3<r-value<0.5; not different at r-value <0.3.
Table 2.
A matrix of R-values of pair-wise comparison of the fecal microbiota of calves in diarrheal group (n=6) using ANOSIM.
Table 2.
A matrix of R-values of pair-wise comparison of the fecal microbiota of calves in diarrheal group (n=6) using ANOSIM.
| Diarrhea group | 1d | 3d | 5d | 7d | 9d | 12d | 15d | 18d | 22d | 26d | 30d | 35d | 40d |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1d | 0 | ||||||||||||
| 3d | r=0.017 | 0 | |||||||||||
| 5d | r=0.362* | r=0.125 | 0 | ||||||||||
| 7d | r=0.533* | r=0.409* | r=0.022 | 0 | |||||||||
| 9d | r=0.635* | r=0.619* | r=0.024 | r=-0.192 | 0 | ||||||||
| 12d | r=0.861* | r=0.833* | r=0.709* | r=0.347* | r=0.128 | 0 | |||||||
| 15d | r=0.938* | r=1* | r=0.924* | r=0.728* | r=0.425* | r=-0.099 | 0 | ||||||
| 18d | r=0.942* | r=0.976* | r=0.887* | r=0.728* | r=0.474* | r=0.039 | r=-0.110 | 0 | |||||
| 22d | r=0.983* | r=0.996* | r=0.979* | r=0.867* | r=0.648* | r=0.109 | r=0.051 | r=-0.011 | 0 | ||||
| 26d | r=0.935* | r=0.976* | r=0.864* | r=0.715* | r=0.611* | r=0.094 | r=0.132 | r=-0.057 | r=-0.185 | 0 | |||
| 30d | r=0.972* | r=0.968* | r=0.949* | r=0.888* | r=0.709* | r=0.240* | r=0.174 | r=-0.002 | r=-0.067 | r=-0.204 | 0 | ||
| 35d | r=0.998* | r=1* | r=0.997* | r=0.992* | r=0.898* | r=0.583* | r=0.462* | r=0.302* | r=-0.019 | r=-0.093 | r=-0.004 | 0 | |
| 40d | r=0.959* | r=0.968* | r=0.947* | r=0.912* | r=0.863* | r=0.561* | r=0.581* | r=0.435* | r=0.146 | r=0.056 | r=0.105 | r=-0.098 | 0 |
Represents the pair-wise comparison with p-value<0.05. Those with r-value<0.5are blod With p-value<0.05, the fecal microbiotas of two ages were considered completely different at r-value>0.75; different at 0.5<r-value<0.75; tended to be different data 0.3<r-value<0.5; not different at r-value <0.3.
Table 3.
Comparison of the relative abundance of intestinal microbiota between calves in healthy and diarrheal groups at different time points.
Table 3.
Comparison of the relative abundance of intestinal microbiota between calves in healthy and diarrheal groups at different time points.
| Day | Bacteria | Group | P-value | |
| A | B | |||
| 1d | L.johnsonii | 0.012±0.010% | 0.030±0.019% | 0.088 |
| Limosilactobacillus | 0 | 0.035±0.021% | 0.088 | |
| P.mirabilis | 0.073±0.071% | 0.043±0.017% | 0.088 | |
| 3d | P.russellii | 0.023±0.018% | 1.330±0.720% | 0.008 |
| F.prausnitzii | 0.029±0.023% | 1.619±1.461% | 0.014 | |
| E.ramosum | 0.778±0.416% | 2.283±0.611% | 0.023 | |
| L.johnsonii | 0.002±0.002% | 1.208±1.113% | 0.023 | |
| 5d | F.umbilicata | 0 | 0.269±0.159% | 0.010 |
| G.genomosp._3 | 0.057±0.037% | 2.764±1.556% | 0.019 | |
| C.pharyngocola | 0.309±0.309% | 1.376±0.609% | 0.040 | |
| 7d | S.mitis | 0.380±0.380% | 0.005±0.008% | 0.013 |
| E.ramosum | 0.638±0.252% | 0.087±0.053% | 0.019 | |
| P.mirabilis | 0.180±0.099% | 0.026±0.017% | 0.019 | |
| A.muciniphila | 2.967±2.760% | 0.004±0.003% | 0.040 | |
| L.amylovorus | 0.011±0.004% | 0.002±0.001% | 0.040 | |
| 9d | P.russellii | 0.032±0.026% | 6.153±5.149% | 0.004 |
| L.murinus | 8.123±7.134% | 0.128±0.127% | 0.010 | |
| F.necrophorum | 0.071±0.042% | 0.001±0.001% | 0.012 | |
| S.mitis | 0.012±0.018% | 8.893±0.003% | 0.012 | |
| A.muciniphila | 0.159±0.096% | 0.008±0.006% | 0.020 | |
| B.vulgatus | 16.951±7.283% | 4.473±2.922% | 0.020 | |
| L.johnsonii | 0.262±0.253% | 2.409±1.656% | 0.020 | |
| P.dorei | 0.003±0.003% | 0.005±0.005% | 0.020 | |
| K.pneumoniae | 1.384±1.285% | 0.003±0.002% | 0.028 | |
| E.coli | 27.465±11.439% | 52.753±7.410% | 0.039 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



