
Submitted:
12 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
Lindernia dubia (L.) Pennell is a species with invasive behavior outside of its native range of distribution (North America), linked mainly to aquatic habitats. This annual species has been acknowledged as a weed in rice paddies in Europe and Asia. Due to the impacts of this invasive plant, some authors have even listed this species as a global invader. The present work focused on spontaneous plant species occurring in cattail seedlings grown in central Spain for the establishment of constructed wetlands. Weed inventory revealed the presence of L. dubia as a dominant spontaneous species in this crop environment. A suite of mesocosm experiments were designed to study the distribution of L.dubia versus the distribution of the other dominant plant species, and to determine traits associated with its weedy potential. Results showed that L.dubia presents competitive attributes such as morphological variability, early flowering, long seeding time, short growth cycle, small and light seeds and high seed production and germination rate (25 ⁰C), meaning a high reproductive capacity in a cycle of about three months, in non-limiting conditions for plant growth. Data from this work provide a basis for understanding the weedy potential of L.dubia, and for management decisions of a potentially invasive species, little investigated in Europe
Keywords:
Lindernia dubia
; invasive species
; plant traits
; cattail seedlings
; constructed wetlands
1. Introduction
Lindernia dubia (L.) Pennell (yellowseed false-pimpernel) is an annual herb of small size, native to North America, where it grows in borders of ponds, river banks and other moist to wet habitats. According to Lewis [1], this species shows great morphological plasticity, especially in vegetative characters; stems can be erect, ascending or prostrate, and they can root at the lower nodes; leaves are simple, opposite and sessile, and their shape is variable, from lanceolate to ovate or suborbicular. Flowers are gamopetalous, zygomorphic, chasmogamous or cleistogamous [2]; the fruit is a septicidal capsule with ellipsoid or globose shape, containing numerous seeds, ellipsoid or rectangular [1].
Nowadays, the distribution of this species has expanded from America to various other regions of the World, as distant as Europe, Korea or Taiwan [3,4]. The Global Biodiversity Information database [4] mentions that there is evidence of impact by this species in Italy, Romania and Japan. Specifically, in Europe L. dubia (hereafter referred as LIDDU) has been appointed as an invasive species in natural aquatic habitats of the Netherlands [5], Romania [6], and Italy [7].
In crop environments, LIDDU has been acknowledged as a weed in rice paddies, where its occurrence seems to be expanding [8,9,10]. Despite the fact that rice is a staple food crop, there is little information on the traits possessed by LIDDU that promote its spread and on its performance as a weed, probably because other weeds of rice, such as Echinochloa spp. and Cyperus spp., are by far much more harmful [11]. In the case of Europe, the cultivation of rice is concentrated in two countries, Italy (60.3 %) and Spain (15.5 %) (rice cropped in Russian Federation not included) [12], and in both countries LIDDU has been reported as an alien weed of rice [13]. Additionally, LIDDU may also occur in other productive moist habitats, such as meadows, plant nurseries or treatment wetlands.
Treatment wetlands are constructed aquatic habitats intended for the restoration of ecosystems or the management of wastewater; they are regarded as nature-based solutions in various contexts [14]; among the most commonly plants used in constructed wetlands (CW) are Typha spp. [15]. Typha spp. can be established in constructed wetlands from rhizomes, seeds or seedlings; while the supply of rhizomes requires much resources (rhizome collection, logistics expenses), seeds are easy to collect and use. However, direct sowing is less successful than the establishment from seedlings, due to seed drift by water movement. This fact, along with the need for large number of plants (5 to 50 individuals·m-2 depending on the type of wetland) [16], has placed the focus on nursery-propagated plants for CW establishment [17]. In this respect, proper sanitation is essential to produce marketable plants and to prevent the spread of pests; some authors highlight that weed growth in container-grown nursery stock is a serious problem not only in terms of economic losses but also in terms of the risk of spread of troublesome plant species, such as alien or invasive plant species [18]. In this regard, we hypothesized that helophyte nurseries may represent an environment prone to weed infestation, due to high availability of growth resources such as water and nutrients. To the best of our knowledge, the occurrence of weeds in helophyte nurseries has not been addressed in the published literature.
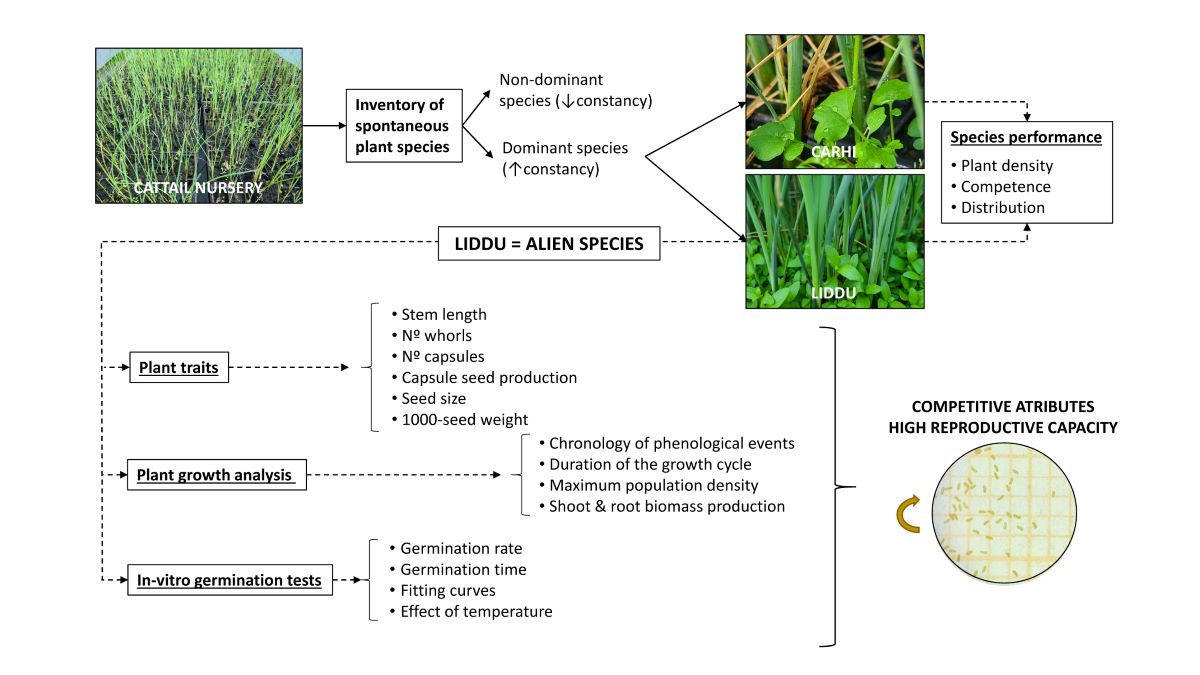
In order to shed some light on spontaneous flora occurring in helophyte nurseries and the interspecific competition between dominant species, we set up a mesocosm experiment with the helophyte Typha domingensis Pers. (commonly known as cattail) intentionally grown from seed in a nursery of Spain for constructed wetlands. The primary objectives were to develop an inventory of potential weeds in cattail nurseries and to assess the occurrence of the prevailing taxa and their introduction pathways in order to understand the source and species that should be controlled. In the course of species identification works, the alien species Lindernia dubia (L.) Pennel was recorded; this finding encouraged the research on the performance and characterization of this species. Thus, in this paper, i) a case study of weeds in cattail seedlings is presented, ii) a new record of the alien species L.dubia is reported, iii) the distribution of dominant weeds is assessed, and iv) traits of L.dubia, a dominant alien species, are determined in order to build a dataset for the estimation of its weedy potential.
2. Results
2.1. Weed inventory and assessment of dominant species
Plant species identified as weeds of cattail seedlings and their constancy values are shown in Table 1. In all, there were five families: Linderniaceae, Brassicaceae, Asteraceae, Cyperaceae, Onagraceae and Poaceae, represented in this particular environment; per life cycle, five annual species (in four families) and four perennials (in three families). The Poaceae family presented taxa in both categories; the annuals Echinochloa crus-galli (L.) P.Beauv. (ECHCG; common name: barnyardgrass) and Digitaria sanguinalis (L.) Scop. (DIGSA; hairy crabgrass) and the perennial Agrostis stolonifera L. (AGSST; creeping bentgrass). Per native range, three alien species were recorded: Lindernia dubia (L.) Pennel (LIDDU; yellowseed false pimpernel), Cyperus eragrostis Lam. (CYPER; tall flatsedge) and Cyperus rotundus L. (CYPRO; nutgrass), according to the list of alien species in Europe provided by EASIN [19] (CYPRO is considered partly native to Europe). The occurrence of LIDDU represented a new record of this alien species for Madrid (Figure 1) [20].
LIDDU was the most frequent species in flooded seedling trays, showing 0.9828 constancy (Table 1), while CARHI was second in the rank (0.9425 constancy). LIDDU and CARHI surpassed by far the occurrence of other species, which presented <0.34 constancy. The existence of dominance was also confirmed by the estimate of the Simpson index (D=0.73), while the estimate of the Shannon index (H=1.53) indicated low diversity.
Population density by dominant species and tray position are shown in Figure 2. In all, the population density ranged from a minimum of 606 to a maximum of 1543 individuals·m-2, showing a mean value of 1024+263 individuals·m-2. Per plant species, LIDDU presented 465+251 individuals·m-2 on average, while CARHI, 560+223 individuals·m-2. Per tray position mean values (LIDDU+CARHI) were 1128+240 and 929+256 individuals·m-2, for the border and inner trays, respectively. Noteworthily, the variability of density was higher in the inner than in the border trays. No significant differences were found between species nor tray position.
2.2. The growth cycle of LIDDU
Along the duration of the experiment, no plant species was observed in the control treatment, meaning that the substrate was free of plant propagules. Concerning the sown pots, the chronology of phenological events is shown in Table 2. Seeds germinated within a very short of time, soon after, cotyledons appeared visible to the naked eye. Flowering started a little before a month. As plants developed leaf whorls, new flower buds arose in the leaf axils; thus, the stages of flowering and fruiting were almost contemporary of the stage of seed dehiscence, which was first noted on DAS 57. According to these records, the length of LIDDU reproductive cycle -i.e. from sowing to the start of seed dispersal- was nearly two months. Plants dried up from DAS 83 and were considered dead on DAS 100, that is, the duration of the growth cycle was estimated at 100 days in these experimental conditions. (See supplementary material S1).
Once the cycle was over, the whole plants (shoots + roots) were removed from the pots for growth measurements. Plant counts showed that stem density was equivalent to 6833 individuals·m-2 on average (7.1 % coefficient of variation, cv), which would be equivalent to 14-times the population recorded for LIDDU in the nursery. The dry matter content of the above ground biomass was 26.5 % (cv 1.4%); its dry weight was equivalent to 368.8 g·m-2 (12.7 % cv). The ratio shoots to roots was 6.3 (dry weight basis). Mean growth rate -from sowing to senescence- was equivalent to 19.4 g·week-1·m-2.
2.3. Germination of LIDDU seeds
The start of the germination (germination time) was observed on Day 2 for the naturally-dehisced seeds (NDS) in the 25 ⁰C treatment whereas in the 20 ⁰C treatment the germination started on Day 3. First observations of open cotyledons were made on Day 4 (19% of the germinated seeds) and Day 6 (1%), respectively. In the 25 ⁰C treatment, the first hypocotyl fibrous root started to grow on Day 10, whereas this observation was recorded on Day 17 for the other treatment (see supplementary material S1). The germination period (number of days from the first observed germination to the last one) was 2.3 times higher in the 20 ⁰C treatment than in the 25 ⁰C one.
The time-course of germination in the two temperature treatments is shown in Figure 3. Great differences were observed between the results of both treatments, especially in final cumulative rate; 97.8 +1.4 % at 25 ⁰C versus 22.8 +7.7 % at 20 ⁰C. In addition, the variability within replicates was usually higher in the 20 ⁰C treatment. Consistent with this, the fit of data to the Gompertz model gave better coefficient of correlation for the 25 ⁰C treatment (r = 0.9948) than for the 20 ⁰C one (r = 0.8435).
As regards the lot of whole fruit seeds (WFS), the germination at 25 ⁰C resulted in 71.8+6.6 % final germination rate on average. The asymptote in the fitted germination curve took the value of 69.8 %, while the coefficient of correlation was 0.9656.
2.4. LIDDU traits
The stem length of LIDDU plants that accompanied cattails ranged from 3.6 to 24.7 cm (10.5 cm on average, 41 % coefficient of variation) (Figure 4-a) and exhibited from 5 to 21 leaf whorls (Figure 4-b), which meant about 1 leaf whorl per cm. The number of capsules per stem also varied greatly, from 3 to 43, in line with the variability found for stem length; on average there were 17.6 capsules per stem (43 % c.v.) (Figure 4-c). Nevertheless, there was a close relationship between number of fruits and stem length (Figure 5), suggesting a regular pattern of flowering and fruiting linked to leaf whorls. Seed production also varied greatly among capsules; it ranged from 34 to 292 seeds, with a mean value of 149.4 seeds·capsule-1 (30 % c.v.) (Figure 4-d).
The variability found for seed measurements was low, showing that the seeds were rather uniform in size and mass, and tiny. Mean seed length and seed width were 0.35 mm (9 % c.v.) (Figure 6-e) and 0.14 mm (13 % c.v.) (Figure 6-f), respectively. Seed ratio length to width ranged from 1.60 to 4.22, with a mean value of 2.6; this value reflected the rectangular seed shape (Figure 6-g). The weight of a thousand seeds was from 2.86 to 3.27 mg; the NDS lot was slightly heavier (3.09+0.17 mg) than the WHS lot (3.00+0.12 mg), but the difference was not statistically significant. The mean value for 1000-seed weight was 3.04 mg (Figure 6-h).
The potential seed production of a plant, or plant seed number, was estimated at 2629 seeds·plant-1 from the mean values found for the number of capsules per stem and the number of seeds per capsule; this amount would mean that the population seed number in the seedling trays might achieve over 1.2·106 seeds·m-2 at the end of the growth cycle. In order to assess the reproductive capacity of a plant in absence of competence, it was assumed 97.1% optimum germination rate (value of the asymptote on the germination fitted curve of NDS at 25 ⁰C) and 72.7 % seed dispersal rate (from the asymptote values of NDS and WFS germination fitted curves). With these assumptions, the maximum reproductive capacity (sexual propagation) would be estimated at 1857 individuals in a cycle of about three months, as long as temperature, substrate and water were not limiting factors.
3. Discussion
Nowadays, the spread of alien species is a matter of concern, especially when they present invasive traits. In an agricultural or horticultural environment, the record of an alien plant species may represent a potential weed infestation and/or a risk for nearby ecosystems. One of the main goals of this work was to gain insight into the spontaneous flora occurring in cattail nurseries. Certainly, the use of potted cattail seedlings infested with weeds for constructed wetlands may result in the spread of undesirable plant species, including potentially invaders.
In the present work, the inventory of species accompanying cattail seedlings -grown in a substrate free of plant propagules- revealed low diversity; it also showed that such species have preference for moist habitats [21]. Thus, CYPER was reported as the most frequent alien species on river banks of the Cantabric watershed of Spain [22]. AGSST is widely distributed in Europe and Spain, where it appears in humid meadows [21]. CYPRO and ECHCG have been acknowledged as major weeds of rice fields globally [11], and DIGSA has been reported as a weed in rice fields of India and USA [11]. In Spain, CYPRO, ECHCG and SONOL were reported as main weeds in sprinkler irrigated rice [23]. In a study of the success of alien weeds in irrigated Mediterranean orchards, CYPRO and ECHCG were categorized as ‘high successful’ invaders [24]. Notwithstanding the above, the constancy values of the above-mentioned species were low in our case-study; the dominant species were others, LIDDU (first in the rank) and CARHI.
To the best of our knowledge, the occurrence of the alien species LIDDU in our area of study (Madrid, Spain) has not been reported so far [20,25,26]. According to the Information System of the Plants of Spain [20] the closest record of the occurrence of LIDDU was located in the proximities of the Rosarito reservoir, i.e. 138 km from our location. It is worth noting that this species was cited as a global plant invader by Laginhas & Bradley [27], and that it was reported as an invasive plant species in some regions of Europe [5]. In eastern Spain (provinces of Tarragona and Valencia) it was reported within the list of emergent rice field flora (flooded rice crops), although with little presence [28]. Results of seed weight in this work and literature data [29,30] showed that LIDDU seeds are lightweight so, the accidental introduction of this alien species into our region might have occurred by a wide range of pathways. It could have even occurred by waterfowl, a pathway suggested by Lovas-Kiss et al. [31] for LIDDU dispersal in Hungary, and by Soons et al. [32] for small seeds in general. In this respect, it is worth noting that our facilities are about 500 m from the Manzanares river (closest river).
CARHI (hairy bittercress) is a well-known weed species, widely studied in the literature. Vaughn et al. [33] stated that CARHI was a major pest in nurseries in USA and highlighted that CARHI occurred ‘in patches in nursery crops and can solidly fill nursery pots if left uncontrolled’ (sic). Warnings about CARHI as a weed of plant nurseries have also been launched in Spain at least since 1997 [34]. These authors highlighted as competitive attributes of CARHI, large seed production (up to 5000 seeds per plant), quick generation times (30-60 days per generation, in favorable climates), and enhanced mechanisms of seed dispersal (ballistic seed dispersal).
The occurrence of LIDDU and CARHI across seedling trays seemed to be influenced by the proximity to the corridors, although no statistically significant differences were found due to high variability in the counts; e.g. LIDDU presented 63 % coefficient of variation for plant counts in the inner trays. Despite variability, it was noticed that LIDDU performed a little worse in cells all around surrounded by cattails (i.e. in the inner trays). This observation is considered in line with [5], who inferred from their results that LIDDU would not easily out-compete fast-growing erect plant species; however, these authors suggested that the impact of LIDDU on small or minute plant communities could be adverse. Competition could explain the differences observed between tray positions in the present study. Another explanation could be the unintentionally human contribution to seed dispersal (workers walked through the corridors) and wind dispersal; open corridors may result in preferential air flow paths, entailing higher seed dispersal towards the closest trays.
In-vitro germination tests showed that the seeds of LIDDU have the potential to germinate very quickly at 25 ⁰C, and that the germination rate is good, especially for naturally-dehisced seeds. Interestingly, growth of hypocotyl roots was observed at an early plantlet stage (see supplementary material S1), an observation that has not been previously reported in literature. The only datum on LIDDU germination found in relevant literature was the 63 % germination rate reported for the control treatment in the study of waterfowl seed dispersal by Lovas-Kiss et al. [31]; their test was conducted at room temperature (20-25 ⁰C), and 16 h light/8 h dark photoperiod. Consistent with those temperature conditions, the value reported was intermediate between the rates we found at 25 ⁰C and 20 ⁰C in this work. Furthermore, the trends observed in our work seem to be consistent with previous studies of freshwater wetlands and other aquatic habitats, that highlighted the capacity of LIDDU to germinate over other species present in seed banks [35] [36]. In our tests the germination rate sharply decreased at 20 ⁰C, a fact that pointed to thermophilic preferences of this species. Although our findings refer to seed lots from an only location, the response of seed germination to temperature obtained in this work suggests that infestation by LIDDU is not likely to happen in the cold season of temperate climates, at least in a short period of time since the introduction of this species. Nevertheless, Simons & Jansen [5] categorized LIDDU as a naturalized invasive in the Netherlands (temperate Atlantic north-western European climate), but estimated that the transition from being a casual alien plant to that status could have taken about ten years. Additionally, there is evidence of the environmental impact by LIDDU in warmer countries of Europe such as Italy and Romania [4].
Chronology of phenological events and plant traits data presented in this work suggest that LIDDU possess competitive attributes such as morphological variability, early flowering, long seeding time, short growth cycle, small and light seeds, and high seed production. Peralta and Royuela [37] cited that LIDDU had a summer phenology in North Spain, flowering from July to September, like we observed in central Spain. Data accompanying the botanical description of LIDDU in North America by [38] reads ‘flowering year-round’; Baker [30] observed that LIDDU flowering ended in autumn (September) in California. As regards plant size, values in this work were in line with the botanical description of LIDDU by Lewis [1]. Results of seed length and width were consistent with the taxonomical study of Linderniaceae conducted in Korea by Bazarragchaa et al [39], and also with data provided in the study of endozoochorous seed dispersal in Hungary [31]. Bazarragchaa et al.[39] described the inflorescence of LIDDU as in axillary flower pairs, solitary axillary or branching in subtending leaf. Despite the fact that the number of capsules in a single stem was not reported by those authors, it can be inferred that the finding of nearly 2 fruits (capsules) per leaf whorl in our work is in line with their inflorescence description. Data on capsule seed number or plant seed number were not found in the consulted literature. Some data on 1000-seed weight were found but appeared controversial to a certain extend. Thus, the Seed Information Database [29] reports 0.01 g as mean value for 1000-seed weight, whereas the Baker Seed Collection database, 0.003 g for the same seed amount [30]. However, a note on the former value says that minor covering structures could have been included in the weights. Therefore, our results of 1000-seed weight are consistent with Baker’s, and support the hypothesis that minor covering structures were included in the weights reported for LIDDU by the Seed Information Database [29].
In the literature there has been much discussion on what plant traits can be associated with invasiveness, as well as on the dependence of plant traits on the environmental context; however, it is widely accepted that traits related to fecundity and species dispersal are essential to assess the potential success of an invader [40]. Concerning weed fecundity, Norris [41] underlined the risks of extrapolating data from controlled conditions to field conditions, but at the same time acknowledged that many papers report experiments carried out under artificial conditions; besides, this author encouraged for data on more weed species. In this sense, our estimates of plant seed production and maximum plant reproductive capacity should be taken with caution, however they provide a basis along with other results from this work for management decisions of LIDDU, a potentially invasive species little investigated in Europe.
4. Materials and Methods
4.1. Site description
This study was developed at the nursery of aquatic plants of the Technical University of Madrid (UPM, Spain) (40 26’ 36” N, 3º 44’ 18” W, 650 m a.s.l.). The climate at the site is temperate with dry and hot summers; normal values of temperature are: 14.6 ⁰C annual mean temperature, -7.4 ⁰C absolute minimum temperature (January) and 40 ⁰C absolute maximum temperature (August) [42].
The aquatic plant facilities included experimental wetlands, a greenhouse for plant propagation and laboratories for plant characterization. These facilities were intended to produce cattail plants for the establishment of external constructed wetlands, as well as for research on phytodepuration. The greenhouse was endowed with an automated system for ventilation (20 ⁰C set point); in addition, doors remained open during summertime. In the interior of the greenhouse, benches were conventionally arranged; two adjacent rows of benches in the middle (central rows) plus two separate rows at each side of the central rows with two aisle spaces of 1.10 m width in between. Benches were watertight by means of a polyethylene (PE) pond liner. They were periodically filled up with tap water to simulate shadow ponds. Every year in the period from 2017 to 2023, cattails were raised from seed in 96-cell plastic trays (52.5 x 33 x 8 cm) placed in the simulated water ponds; before seeding, trays were filled with a commercial substrate (Pindstrup Mosebrug S.A.E. code 51212). After emergence, seedlings were thinned to one per cell and let to grow until planting in constructed wetlands. The alien LIDDU was noticed for the first time in the summer of 2022.
4.2. Weed inventory and assessment of the dominant species
The occurrence of spontaneous plant species in cattail seedlings was studied in autumn 2022. At that time, cattails were about 50-65 cm tall and exhibited 5-7 green leaves. Identification of spontaneous plant species growing in the trays was carried out through botanical keys and monographs [21] [43]. Species constancy was assessed as the ratio of the number of seedling trays with a determined species to the total number of seedling trays (n=174). The indexes of diversity (H) [44] and dominance (D) [45] were calculated from tray counts.
The occurrence of LIDDU was assessed in terms of population density and distribution across the nursery, as compared to CARHI, the other dominant weed species in this study. A randomized block design with four blocks and two replications per block was used. To this end, two categories: border (trays placed next to the aisle) and inner (trays placed on the inner area of the bench table) were made. Population density (individuals·m-2) was calculated from the number of individuals counted in each seedling tray, considering the tray surface area (1536 cm2·tray-1).
4.3. Growth chamber experiments
Two types of experiments were developed in a plant growth chamber (300 L capacity, forced air circulation, 6 x F30W/Gro-Lux T8) to gain insight into LIDDU growth cycle and seed germination.
The first one was a pot experiment, where the pot dimensions were: top diameter, 10.5 cm; bottom diameter, 8.5 cm; and height, 9.5 cm. Pots (n=4) were filled up with the same substrate as the one used in the cattail nursery (Pindstrup Mosebrug S.A.E. code 51212). A control (no sown treatment) was used to check the absence of plant propagules in the substrate. Mature fruits (light-brown capsules, starting to break) were taken from senescent LIDDU plants in the seedling trays; seeds were spread on the pot surface, using 50 capsules·pot-1; capsules were left on the ground. Then, the just-sown pots were placed in a water bath inside the growth chamber at 25 ⁰C constant temperature and 14 h day/10 h dark cycle. Water level in the bath was kept at pot surface level (daily water replenishment). Dates of the start of the phenological stages: visible cotyledons, flowering, fruiting, seed dehiscence and plant senescence (supplementary material S1), were recorded and expressed as DAS (Days After Sowing). Once the seed dehiscence was observed, naturally-dehisced seeds from the potted plants were collected on a daily basis; after seed cleaning (debris separation under stereomicroscope) they were gathered in a seed lot named NDS (=Naturally-Dehisced Seeds). When plants were fully dry (brown-colored shoots), they were uprooted in order to determine the following parameters: population density (individuals·m-2), shoots dry matter content (%), average growth rate (dry mass accumulated per week and ground area, g·week-1·m-2) and ratio shoots/roots (dry weight basis). Dry weights were determined by oven-drying at 105 ⁰C.
The other type of experiments were in-vitro seed germination tests. Germination of NDS was performed in covered Petri-dishes filled with filter paper moistened with distilled water; 2 thermal regimes (20 and 25 ⁰C constant temperature) were tested using three replicates of about 100 seeds each (n = 3 x 100). The photoperiodic regime was 14 h light/10 h dark. Observations were made on a daily basis under a Leica EZ4 stereomicroscope 35 x (transmitted illumination). A seed was considered to have germinated when the radicle emerged (see supplementary material S1). The germination rate was calculated as the number of germinated seeds divided by the total number of seeds in the Petri dish. Dates of the first observation of visible cotyledons and lateral root growth were annotated as well.
Another seed lot was prepared and tested for in-vitro germination. All seeds (naturally-dehisced plus not-dehisced seeds) inside 50 mature LIDDU capsules were extracted by shaking and by hand with the help of a thin brush and a spatula; after seed cleaning under stereomicroscope, seeds were gathered in a seed lot named WFS (Whole Fruit Seeds). WFS germination was tested at 25 ⁰C constant thermal regime, following the same procedure as for NDS tests. Final germination rate of WFS was compared to that of NDS in order to assess the seed dispersal rate of LIDDU capsules.
4.4. LIDDU traits
Plant traits studied were the following: stem length (SL), number of whorls per stem (NW), number of capsules per stem (NC), seed production per capsule (NS), seed size, 1000-seed weight, and maximum reproductive capacity in absence of competence. One hundred shoots at senescence stage (brown shoots) were randomly sampled from cattail seedling trays in the nursery for SL, NW and NC; from these plants, fifty capsules -dry capsules starting to open- were taken at random for NS. Seed extraction and seed cleaning (separation from debris) were performed by hand with the help of laboratory equipment (thin brushes, tweezers, spatulas, stereomicroscope); afterwards, seeds from each capsule were counted under a stereomicroscope to determine NS. Seed size measurements included seed length (L) and width (W) and ratio L/W [39]; the sample size for seed measurements was one hundred seeds. Thousand-seed weight (air-dry seeds) was determined in triplicate using a five decimal precision balance. This trait was studied for the two lots of seeds, NDS and WFS (n=3 x 2 x 1000).
The maximum reproductive capacity (individuals per ground area, pl·m-2) was calculated as the product of mean NS, mean NC, seed dispersal rate and optimum germination rate. It was assumed that the optimum germination rate was the highest asymptote value found for the germination curves in the germination tests.
4.5. Statistical analysis
Descriptive statistics (mean, standard deviation, minimum, maximum, coefficients of variation) were calculated. Data of LIDDU traits were graphically presented in terms of frequency (percentage in a category). Population densities of dominant species were analyzed by one-way ANOVA with the location of plants in the nursery as between-subject factor and two weed dominant species (factor with two levels) as a within-subject factor. For the study of seed germination, data were fitted to the S-shaped Gompertz function: Y=a*exp(-b*exp(-c*T), using the cumulative germination rate as dependent variable and the germination time (days) as independent variable; parameters a, b and c in the Gompertz function were calculated by successive iterations. Data analyses were conducted using the statistics software Statgraphics 19®.
Supplementary Materials
The following supporting information can be downloaded at the website of this paper posted on Preprints.org. Supplementary Material S1: Photographic report.
Author Contributions
Conceptualization, investigation, methodology, formal analysis, writing—original draft preparation, MDC; data curation, investigation, formal analysis, GS; resources, visualization, validation, PLA; writing—review and editing, validation, supervision, ISM. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Data Availability Statement
Data are contained within the article.
Acknowledgments
Technical support for the study of LIDDU germination was provided by Eng. J.G. Apodaca. Support from Projects FPA220000PA1402 and Life16 CCM/GR/000044 is also gratefully acknowledged.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Lewis, D.Q. A Revision of the New World Species of Lindernia (Scrophulariaceae). Castanea 2000, 65, 93–122. [Google Scholar]
- Ikeda, E.; Miura, R. A Note on the Proportion of Cleistogamous Flowers in Lindernia Dubia (L.) Pennel (Scrophulariaceae) in a Paddy Field*. Weed Research, Japan 1994, 39, 177–179. [Google Scholar] [CrossRef]
- Seebens, H.; Blackburn, T.M.; Hulme, P.E.; van Kleunen, M.; Liebhold, A.M.; Orlova-Bienkowskaja, M.; Pyšek, P.; Schindler, S.; Essl, F. Around the World in 500 Years: Inter-Regional Spread of Alien Species over Recent Centuries. Global Ecology and Biogeography 2021, 30, 1621–1632. [Google Scholar] [CrossRef]
- Global Biodiversity Information GBIF.Org (16 April 2024). Available online: https://www.gbif.org/en/ (accessed on 20 April 2024).
- Simons, E.L.A.N.; Jansen, M.G.M. Ecology of Naturalized Invasive Species Lindernia Dubia (L.) Pennell in the Netherlands. Gorteria: Tijdschrift voor Onderzoek aan de Wilde Flora 2018, 40, 1–10. [Google Scholar]
- Neacșu, A.; Imbrea, I.; Lațo, A.; Arsene, G.G. Lindernia Dubia (L.) Pennell: A New Invasive in the Romanian Banat. AgroLife Scientific Journal 2021, 10, 129–135. [Google Scholar] [CrossRef]
- Galasso, G.; Domina, G.; Ardenghi, N.M.G.; Aristarchi, C.; Bacchetta, G.; Bartolucci, F.; Bonari, G.; Bouvet, D.; Brundu, G.; Buono, S.; et al. Notulae to the Italian Alien Vascular Flora: 7. Italian Botanist 2019, 7, 157–182. [Google Scholar] [CrossRef]
- Webb, D.A.; Philcox, D. Lindernia All. In Flora Europaea, volume 3; 1972; pp. 203–204. [Google Scholar]
- Kumar, V.; Mahajan, G.; Sheng, Q.; Chauhan, B.S. Weed Management in Wet Direct-Seeded Rice (Oryza Sativa L.): Issues and Opportunities. In Advances in Agronomy; Academic Press Inc., 2023; Vol. 179, pp. 91–133. ISBN 9780443192623. [Google Scholar]
- Yoshino, N.; Wang, G.X.; Ito, M.; Auld, B.; Kohara, H.; Enomoto, T. Naturalization and Dissemination of Two Subspecies of Lindernia Dubia (Scrophulariaceae) in Japan. Weed Biol Manag 2006, 6, 174–176. [Google Scholar] [CrossRef]
- Kraehmer, H.; Jabran, K.; Mennan, H.; Chauhan, B.S. Global Distribution of Rice Weeds - A Review. Crop Protection 2016, 80, 73–86. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations FAOSTAT. Available online: https://www.fao.org/faostat/en/#data (accessed on 30 May 2024).
- Carretero, J.L. La Vegetación Emergente de Los Arrozales Europeos. An Biol 1988, 15, 135–141. [Google Scholar]
- Sonneveld, B.G.J.S.; Amani, A.; Merbis, M.D. Nature-Based Solutions for Agricultural Water Management. 2022; Vol. 1, ISBN 9783030424626. [Google Scholar]
- Vymazal, J. Plants in Constructed, Restored and Created Wetlands. Ecol Eng 2013, 61, 501–504. [Google Scholar] [CrossRef]
- Kataki, S.; Chatterjee, S.; Vairale, M.G.; Dwivedi, S.K.; Gupta, D.K. Constructed Wetland, an Eco-Technology for Wastewater Treatment: A Review on Types of Wastewater Treated and Components of the Technology (Macrophyte, Biolfilm and Substrate). J Environ Manage 2021, 283. [Google Scholar] [CrossRef]
- Rogers, S.M.D. Tissue Culture and Wetland Establishment of the Freshwater Monocots Carex, Juncus, Scirpus, and Typha. In Vitro Cellular and Developmental Biology - Plant 2003, 39, 1–5. [Google Scholar] [CrossRef]
- Case, L.T.; Mathers, H.M.; Senesac, A.F. A Review of Weed Control Practices in Container Nurseries. Horttechnology 2005, 15, 535–545. [Google Scholar] [CrossRef]
- European Commission EASIN - European Alien Species Information Network Available online: https://easin.jrc.ec.europa.eu/easin.
- Anthos. Information System of the Plants of Spain. Real Jardín Botánico, CSIC - Fundación Biodiversidad. Available online: www.anthos.es.
- Castroviejo (Coord.), S. Flora Iberica (22 Vol.); Real Jardín Botánico, CSIC: Madrid, 1986; ISBN 84-00-06221-3. [Google Scholar]
- Liendo, D.; Biurrun, I.; Campos, J.A.; Herrera, M.; Loidi, J.; García-Mijangos, I. Invasion Patterns in Riparian Habitats: The Role of Anthropogenic Pressure in Temperate Streams. Plant Biosyst 2015, 149, 289–297. [Google Scholar] [CrossRef]
- Cavero, J.; Zaragoza, C.; Cirujeda, A.; Anzalone, A.; Faci, J.M.; Blanco, O. Selectivity and Weed Control Efficacy of Some Herbicides Applied to Sprinkler Irrigated Rice (Oryza Sativa L.). Spanish Journal of Agricultural Research 2011, 9, 597. [Google Scholar] [CrossRef]
- Juárez-Escario, A.; Valls, J.; Solé-Senan, X.O.; Conesa, J.A. A Plant-Traits Approach to Assessing the Success of Alien Weed Species in Irrigated Mediterranean Orchards. Annals of Applied Biology 2013, 162, 200–213. [Google Scholar] [CrossRef]
- Madrid Council Flora Urbana de Madrid Available online: https://datos.madrid.es/portal/site/egob.
- Casas, I.; Gavilán, R.; Echevarría, J. Catálogo de La Flora Vascular de La Ciudad Universitaria de Madrid (España). Bot Complut 1993, 18, 175–201. [Google Scholar]
- Laginhas, B.B.; Bradley, B.A. Global Plant Invaders a Compendium of Invasive Plant Taxa Documented by the Peer-reviewed.Pdf. Ecology 2022, 103, e03569. [Google Scholar] [CrossRef]
- Carretero, J.L. Rice Field Flora and Vegetation in the Provinces of Valencia and Tarragona. Collectanea Botanica (Barcelona) 1987, 17, 113–124. [Google Scholar] [CrossRef]
- Society for Ecological Restoration; International Network for Seed Based Restoration; Royal Botanic Gardens Kew Seed Information Database (SID) Available online: https://ser-sid.org/.
- Baker, H.G. Baker Seed Collection Database Https://Ucjeps.Berkeley.Edu/Baker/. Available online: https://ucjeps.berkeley.edu/baker/ (accessed on 11 June 2024).
- Lovas-Kiss, Á.; Navarro-Ramos, M.J.; Vincze, O.; Löki, V.; Urgyán, R.; Pallér-Kapusi, F.; van Leeuwen, C.H.A.; Green, A.J.; Lukács, B.A. Traits for Transport: Alien Wetland Plants Gain an Advantage during Endozoochorous Seed Dispersal by Waterfowl. Freshw Biol 2023, 68, 1703–1715. [Google Scholar] [CrossRef]
- Soons, M.B.; Van Der Vlugt, C.; Van Lith, B.; Heil, G.W.; Klaassen, M. Small Seed Size Increases the Potential for Dispersal of Wetland Plants by Ducks. Journal of Ecology 2008, 96, 619–627. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Bowling, A.J.; Ruel, K.J. The Mechanism for Explosive Seed Dispersal in Cardamine Hirsuta (Brassicaceae). Am J Bot 2011, 98, 1276–1285. [Google Scholar] [CrossRef]
- Bay, T.; Gracia, I.; Cavero, J.; Zaragoza, C. Cardamine Hirsuta Infestante de Los Viveros...¿Importada Con La Turba? In Proceedings of the Actas del Congreso 1997 de la Sociedad Española de Malherbología (SEMH); SEMH, Ed.; SEMH: Valencia, Spain, 1997; pp. 177–180.
- Abernethy, V.J.; Willby, N.J. Changes along a Disturbance Gradient in the Density and Composition of Propagule Banks in Floodplain Aquatic Habitats. Plant Ecol 1999, 140, 177–190. [Google Scholar] [CrossRef]
- Neff, K.P.; Rusello, K.; Baldwin, A.H. Rapid Seed Bank Development in Restored Tidal Freshwater Wetlands. Restor Ecol 2009, 17, 539–548. [Google Scholar] [CrossRef]
- Peralta, J.; Royuela, M. Herbario-Departamento de Ciencias de La Universidad Pública de Navarra. Available online: https://www.unavarra.es/herbario/htm/Lind_dubi.htm (accessed on 28 May 2024).
- Lewis, D.Q. Lindernia Dubia. Available online: http://floranorthamerica.org/Lindernia_dubia (accessed on 22 May 2024).
- Bazarragchaa, B.; Yang, S.; Kim, H.S.; Lee, S.J.; Lee, J. An Updated Taxonomy of the Family Linderniaceae in Korea. Korean Journal of Agricultural Science 2019, 46, 1007–1018. [Google Scholar] [CrossRef]
- Lau, J.A.; Funk, J.L. How Ecological and Evolutionary Theory Expanded the ‘Ideal Weed’ Concept. Oecologia 2023, 203, 251–266. [Google Scholar] [CrossRef]
- Norris, R.F. Weed Fecundity: Current Status and Future Needs. Crop Protection 2007, 26, 182–188. [Google Scholar] [CrossRef]
- Agencia Estatal de Meteorología; Instituto de Meteorología de Portugal Iberian Climate Atlas: Air Temperature and Precipitation (1971-2000); Agencia Nacional de Meteorología de España (AEMET), Instituto de Meteorología de Portugal (IM), Eds.; 2011; ISBN 9788478370795.
- Recasens, J.; Conesa, J.A. Malas Hierbas En Plántula. Guía de Identificación; Editions de la Universitat de Lleida, Bayer CropScience, 2009; ISBN 978-84-8409-290-4.
- Shannon, C.; Wever, W. The Mathematical Theory of Communication; University of Illinois Press, Urbana, IL, USA, 1949; ISBN 0-252-72546-8.
- Simpson, E.H. Measurement of Diversity. Nature 1949, 163, 688–688. [Google Scholar] [CrossRef]
Figure 1.
Confirmed records of Linaria dubia (L.) Pennel in the Iberian Peninsula (red dots), according to the Information System of the plants of Spain [20]. (Simplified map). Yellow dot = Madrid.
Figure 1.
Confirmed records of Linaria dubia (L.) Pennel in the Iberian Peninsula (red dots), according to the Information System of the plants of Spain [20]. (Simplified map). Yellow dot = Madrid.
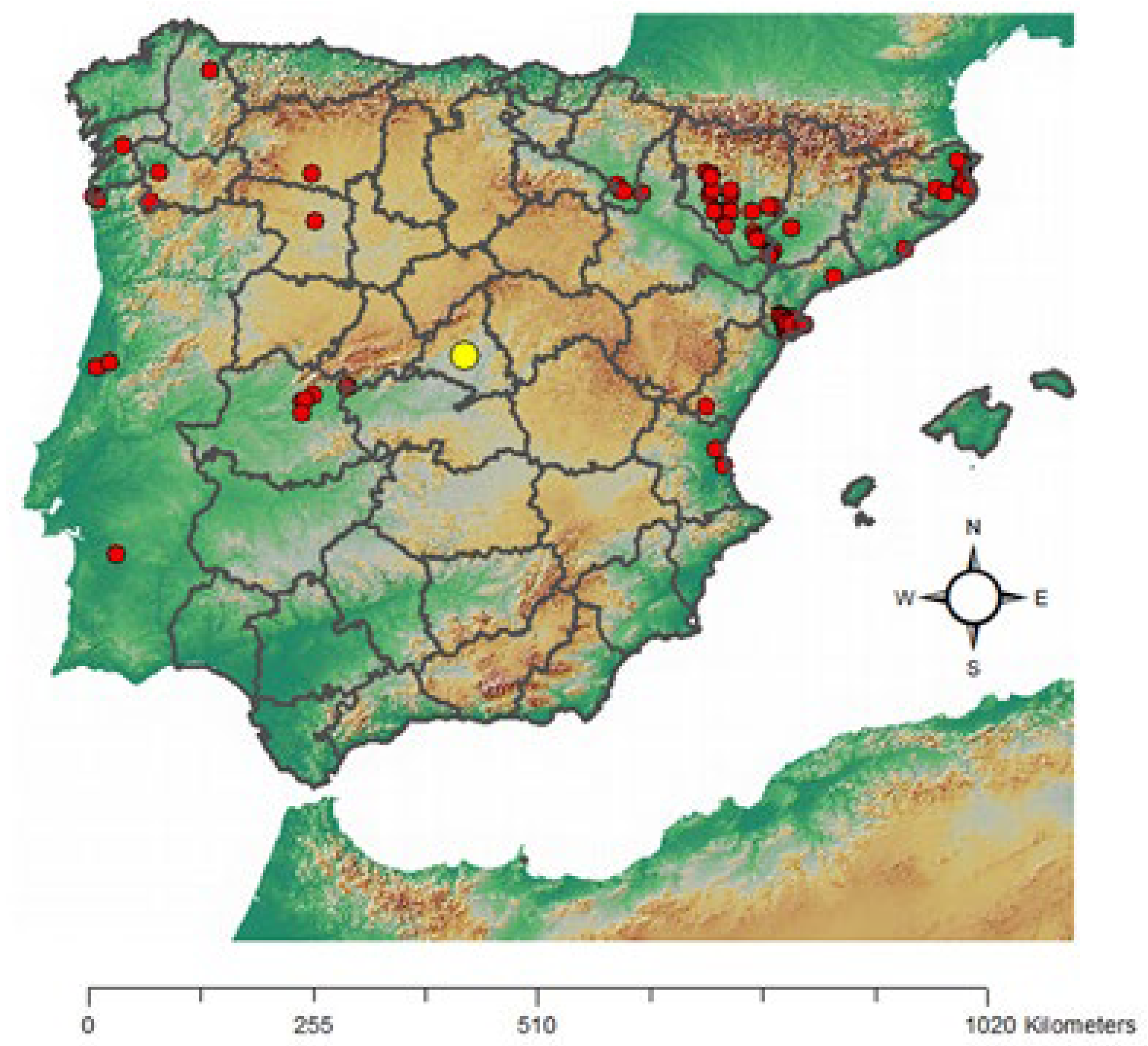
Figure 2.
Population density of dominant plant species. CARHI = Cardamine hirsuta L.; LIDDU = Lindernia dubia (L.) Pennel; B= border position; I = inner position. The bars show + standard error.
Figure 2.
Population density of dominant plant species. CARHI = Cardamine hirsuta L.; LIDDU = Lindernia dubia (L.) Pennel; B= border position; I = inner position. The bars show + standard error.
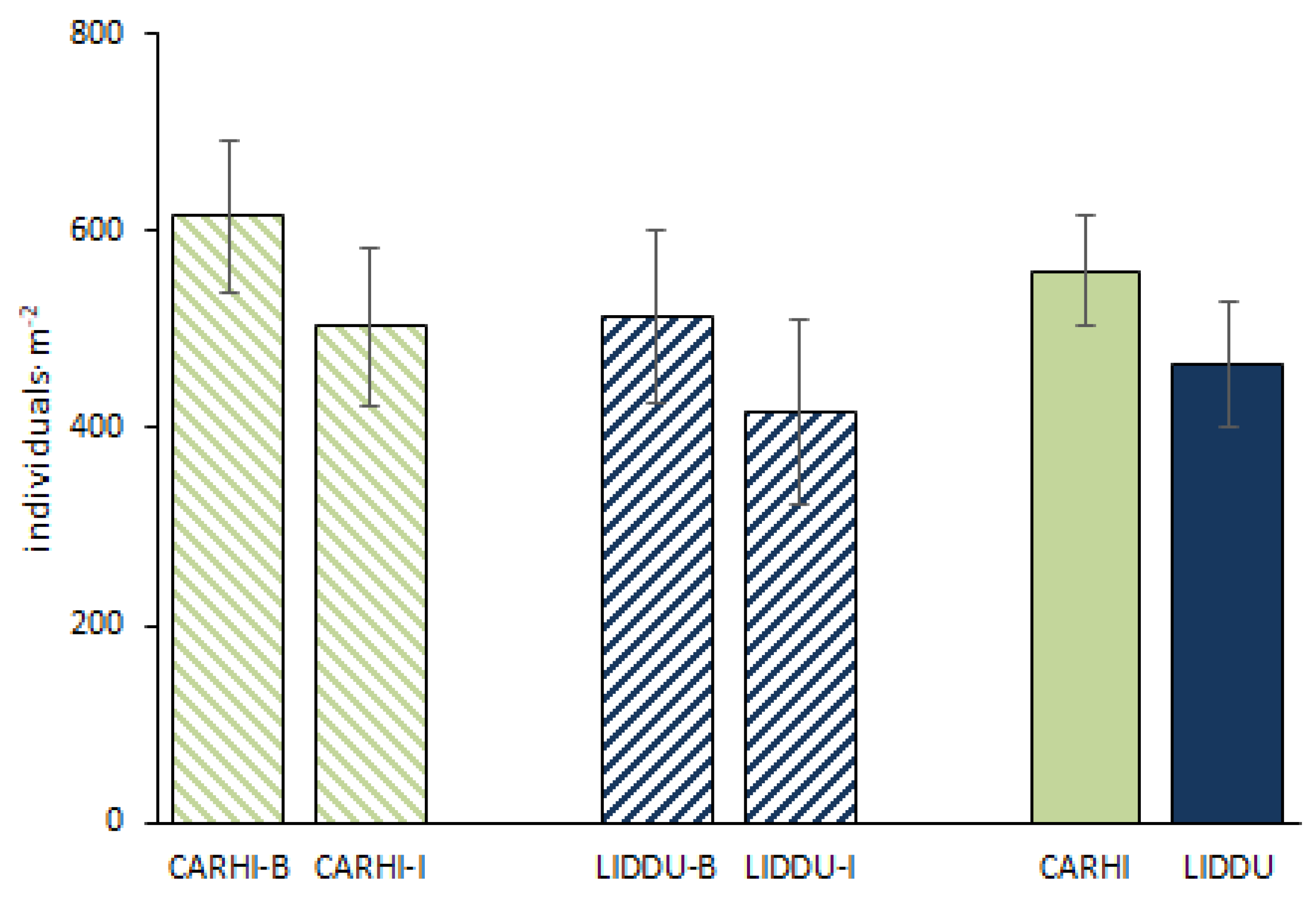
Figure 3.
Fitted curves for the germination of naturally-dehisced seeds of Lindernia dubia. The bars show +standard deviation values.
Figure 3.
Fitted curves for the germination of naturally-dehisced seeds of Lindernia dubia. The bars show +standard deviation values.
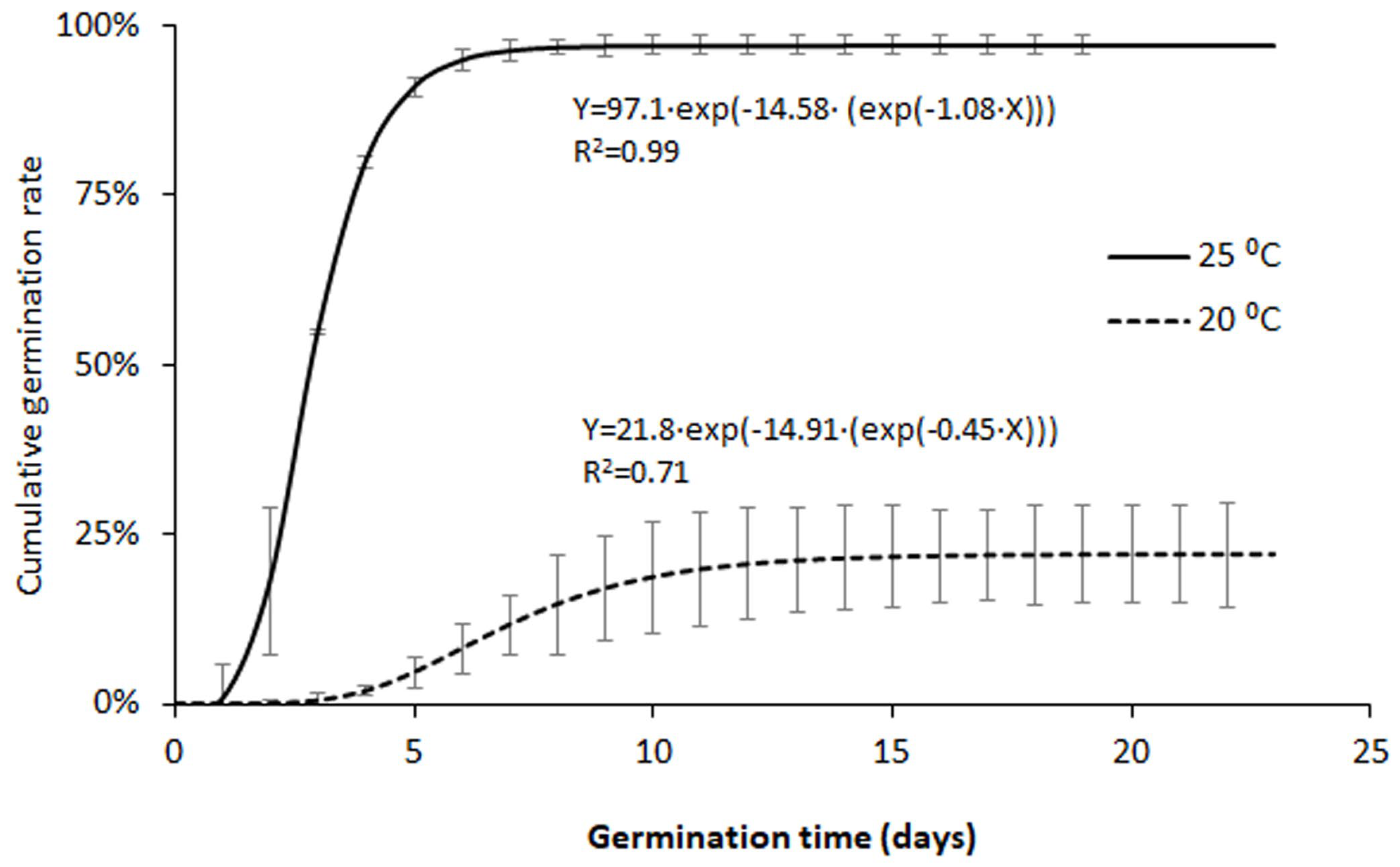
Figure 4.
Frequency diagrams for data on traits of Lindernia dubia: (a) Stem length; (b) Number of leaf whorls per stem; (c) Number of capsules per stem; (d) Number of seeds per capsule.
Figure 4.
Frequency diagrams for data on traits of Lindernia dubia: (a) Stem length; (b) Number of leaf whorls per stem; (c) Number of capsules per stem; (d) Number of seeds per capsule.

Figure 5.
Relationship between the number of capsules and the length of the stem of Lindernia dubia.
Figure 5.
Relationship between the number of capsules and the length of the stem of Lindernia dubia.
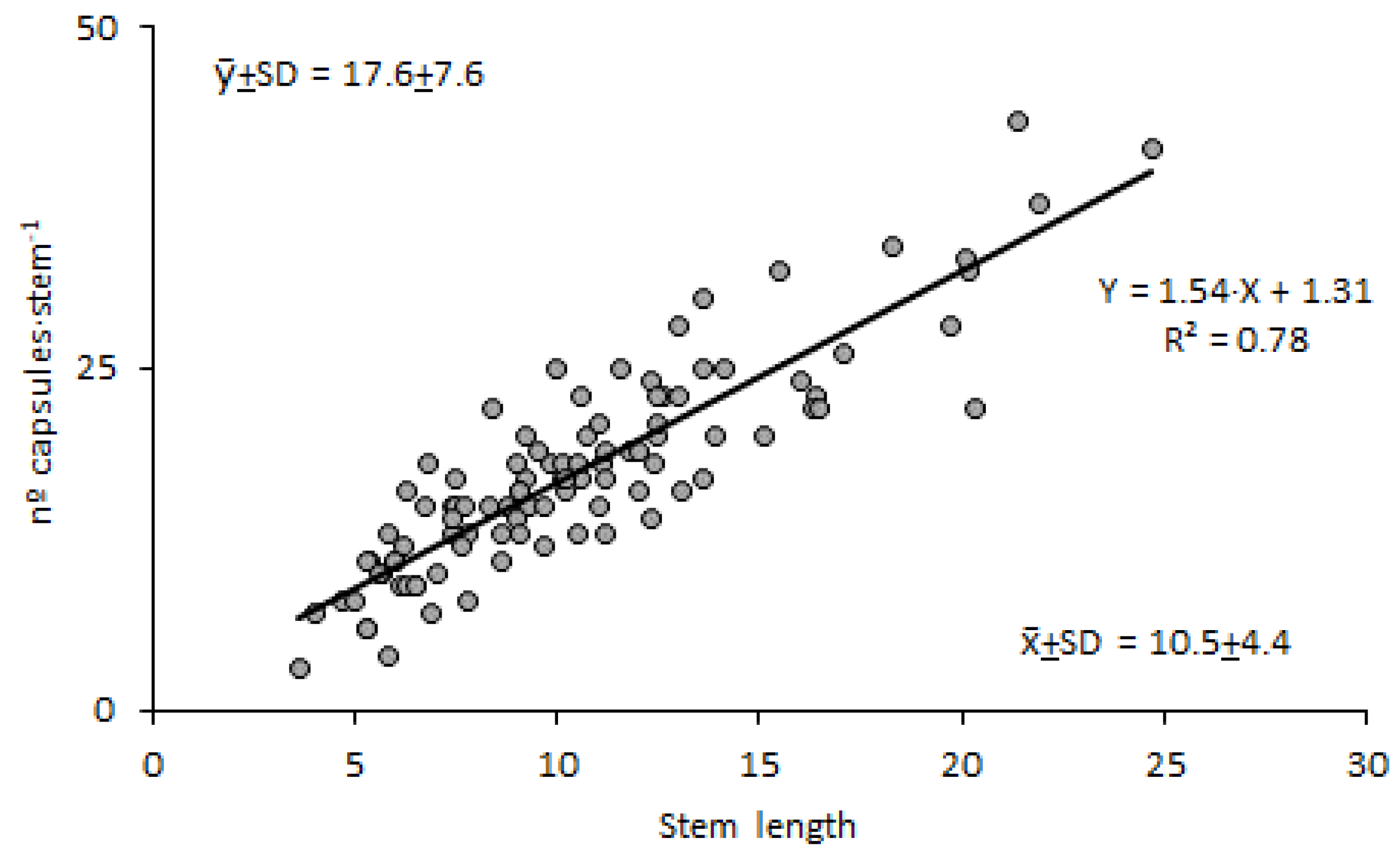
Figure 6.
Frequency diagrams for data on traits of Lindernia dubia: (e) Seed length; (f) Seed width; (g) Seed ratio length to width; (h) 1000-seed weight. NDS = Naturally-Dehisced Seeds; WFS = Whole Fruit Seeds. The bars show + standard deviation.
Figure 6.
Frequency diagrams for data on traits of Lindernia dubia: (e) Seed length; (f) Seed width; (g) Seed ratio length to width; (h) 1000-seed weight. NDS = Naturally-Dehisced Seeds; WFS = Whole Fruit Seeds. The bars show + standard deviation.
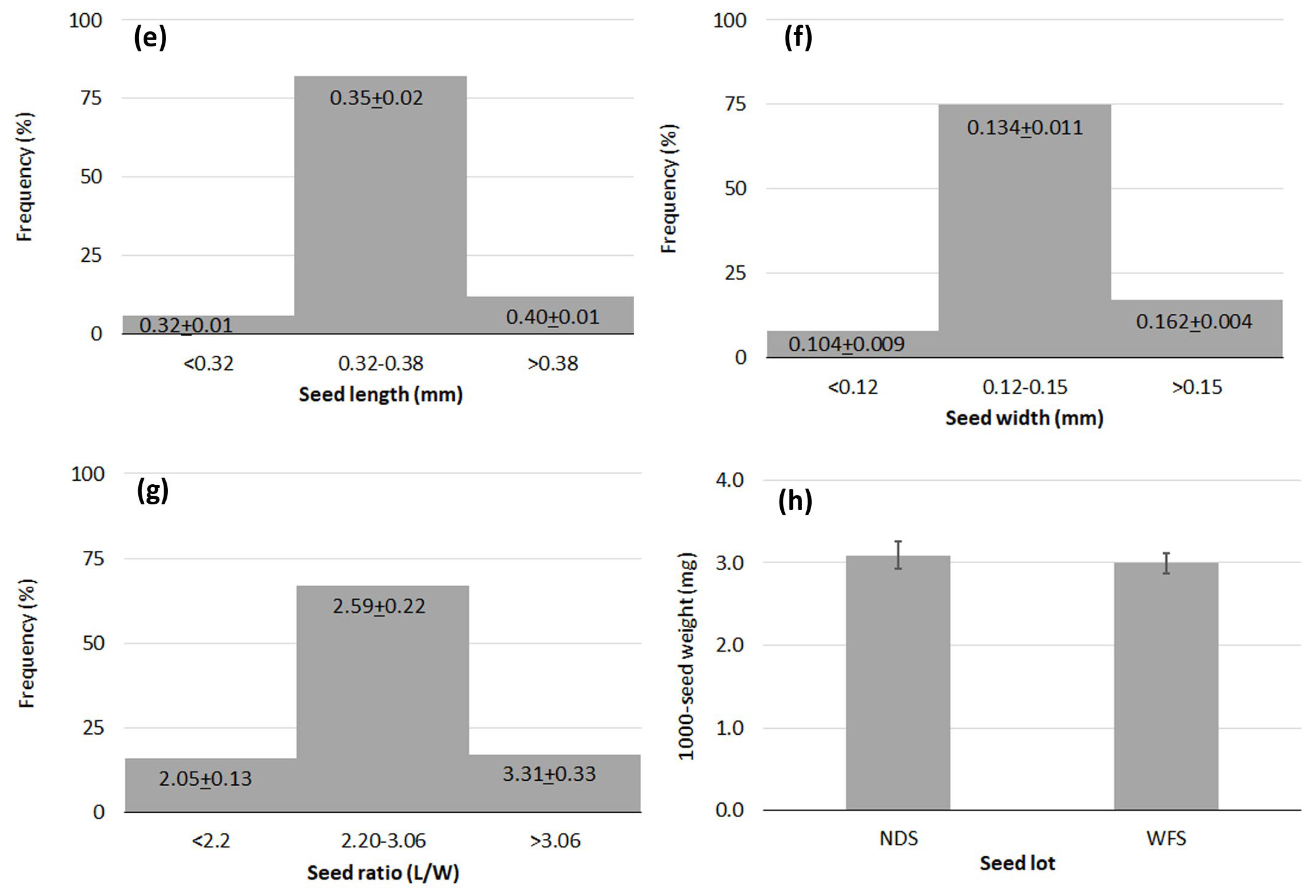

Table 1.
Inventory of plant species present in cattail seedling trays and constancy values.
| Taxon | EPPO code1 | Family | Life cycle | Origin | Constancy |
|---|---|---|---|---|---|
| Lindernia dubia (L.) Pennel | LIDDU | Linderniaceae | Annual | North America | 0.9828 |
| Cardamine hirsuta L. | CARHI | Brassicaceae | Winter annual | Europe | 0.9425 |
| Cyperus eragrostis Lam. | CYPER | Cyperaceae | Perennial | Tropical America | 0.3391 |
| Epilobium roseum Schreb. | EPIRO | Onagraceae | Perennial | Europe to China | 0.2471 |
| Agrostis stolonifera L. | AGSST | Poaceae | Perennial | Eurasia, North Africa | 0.0862 |
| Digitaria sanguinalis (L.) Scop | DIGSA | Poaceae | Summer annual | Mediterranean region to Central Asia and Malaysia | 0.0632 |
| Cyperus rotundus L. | CYPRO | Cyperaceae | Perennial | Indian sub-continent | 0.0287 |
| Sonchus oleraceus L. | SONOL | Asteraceae | Summer annual (biennial) | Mediterranean region, Europe | 0.0230 |
| Echinochloa crus-galli (L.) P.Beauv. | ECHCG | Poaceae | Summer annual | Europe | 0.0172 |
1 1 EPPO = European and Mediterranean Plant Protection Organization https://gd.eppo.int.
Table 2.
Chronology of phenological observations. Lindernia dubia grown from seeds at 25 ⁰C. DAS = Days After Sowing.
Table 2.
Chronology of phenological observations. Lindernia dubia grown from seeds at 25 ⁰C. DAS = Days After Sowing.
| DAS | Start of phenological stage |
|---|---|
| 6 | Visible cotyledons |
| 15 | Leaf development |
| 30 | Flowering |
| 42 | Fruit development |
| 57 | Seed dehiscence |
| 83 | Senescence |
| 100 | Dead plants |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



