Submitted:
13 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
Microcystis-dominated cyanobacterial blooms (MCBs) frequently occur in freshwaters worldwide due to massive Microcystis colonies formation and severely threaten human and ecosystems health. Quorum sensing (QS) is a direct cause of Microcystis colonies formation, possibly regulating MCBs occur and Microcystis population behaviors [1, 2]. Many novel findings in fundamental knowledge of Microcystis QS phenomenon and the signaling molecules have been documented. However, little effort has devoted to comprehensively summarize and discuss research progresses and exploration direction on QS signaling molecules-mediated QS system in Microcystis. This article summarizes action process of N-acyl homoserine lactones (AHLs) as major signaling molecules in Microcystis and discussed the detailed roles of AHLs-mediated QS system in cells aggregation, colonies formation, ecological adaptability and competitive advantage. And the progress was summarized in research on the QS mechanism in Microcystis. Compared to other QS system, the LuxI/LuxR-type QS system is more likely to found in Microcystis. Also, we introduced Quorum quenching (QQ) — QS blocking process in Microcystis, with emphasizing on the potential of QS inhibitors in MCBs control. Finally, in response to the research deficiencies and gaps in Microcystis QS and QQ, we proposed several future research directions in these fields. This review may deepen the understanding of Microcystis QS knowledge and provide theoretical guidance for the development of strategies to monitor, control and harness MCBs.
Keywords:
Microcystis
; Quorum sensing
; Quorum quenching
1. Introduction
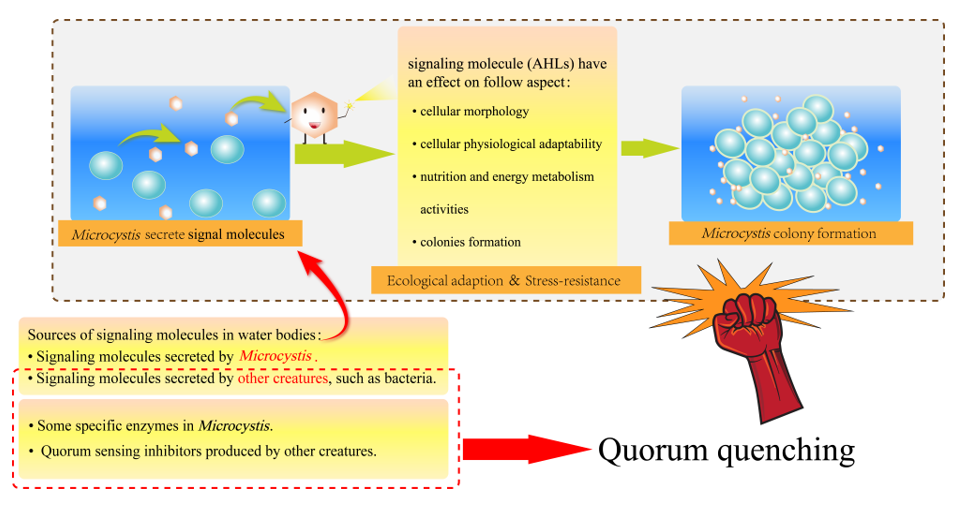
Cyanobacterium, an ancient unicellular photosynthetic prokaryote, is crucial in emergence of high-level aerobic organisms [1]. With global warming and rising eutrophication of water bodies, cyanobacterial blooms frequently occur worldwide via massive cell proliferation. Microcystis is the most common genus that dominates cyanobacterial blooms in freshwaters, and consists of two ecotypes: microcystin-producing (MC+) and non-microcystin-producing (MC-) Microcystis [2,3], where microcystins are widespread cyanotoxins endangering eco-safety [4]. Over past decades, outbreaks of MC+ Microcystis-dominated cyanobacterial blooms (MCBs) in freshwaters have endangered ecological and human health and aroused public concern [5,6,7]. Development and depletion of MCBs involve following processes: Microcystis cells aggregation, colonies formation and colonies disaggregation [8]. Microcystis cells aggregation into colonies enhances their buoyancy to allow rapid flotation nearby freshwater surface, so their colonies formation contributes to MCBs outbreak/maintenance and Microcystis dominance/prevalence in cyanobacterial blooms [9,10,11]. From colonies formation view, many researchers explored the cause of MCBs outbreaks [12,13,14,15,16], while the regulatory mechanisms of Microcystis colonies formation deserve further attention. Although most studies suggested that the production and secretion of MCs and extracellular polymers (EPSs), as well as other cellular traits, of Microcystis lead to Microcystis cells aggregation and colonies formation, yet it is noteworthy that QS is directly responsible for Microcystis colonies formation. Thus, QS as direct driving force for Microcystis colonies formation are non-negligible.
QS is a density-dependent phenomenon that regulates intercellular communication between and within species. QS system is prevalent in bacterial kingdom, with the majority reported for Gram-negative (G-) bacteria. The bacteria with QS system can secrete signaling molecules called autoinducer (AI), where the secretion increases with rising cell density to regulate bacterial population characteristics and behaviors [17]. When diffusible signaling molecules accumulate to required threshold in extracellular environment, bacterial population exhibits obvious changes of phenotypic and behavioral traits, involving cell aggregation, swarming motility, bioluminescence, biofilm formation, colonization, sporulation, cytotoxins/antibiotics syntheses and virulence factor production [18,19,20,21]. Generally, QS exerts effect via three stages: signaling molecule-production/secretion by cells in a population, signaling molecule-sensing by other cells within or outside the population, and signaling molecule-binding by receptor protein. QS phenomenon has recently been discovered in Microcystis population. The regulatory roles and processes of QS in Microcystis population behaviors and MCBs outbreak/maintenance are becoming research hotspot and well documented nowadays. However, little effort has dedicated to comprehensively summarize and discuss research progresses and exploration direction on Microcystis QS.
This paper provided an updated comprehensive summary and review for current research progress of Microcystis QS, aiming to facilitate an integrative understanding on QS-regulated MCBs outbreak mechanisms. Specifically, the discovery processes of QS phenomenon and QS signaling molecules in Microcystis were firstly described. Detail roles of QS signaling molecules secreted by Microcystis itself or exogenously originated in cells aggregation and colonies formation were reviewed and discussed from several aspects of cellular growth and morphology, physiological adaptability (i.e., MCs and EPSs production/release), and nutrition/energy metabolisms. To grasp QS mechanisms, genetic information and action process including signals syntheses, sensing and binding of Microcystis were summarized. Various exogenous signal analogs as QS inhibitors and QS-blocking process in Microcystis through degrading QS signals or disrupting QS signals syntheses and receptor binding were introduced, with emphasizing on the potential of QS inhibitors in MCBs control. Ultimately, we proposed several future research directions in field of Microcystis QS, with the purpose for deepening understanding on knowledge in this field and developing strategies to monitor, control and harness MCBs in context of climate change.
2. QS Discovery and Signaling Molecules Recognition in Microcystis
2.1. Cell Density-Dependent Regulatory Behaviors in Microcystis
QS regulates bacterial population characteristics via signaling molecules-mediated intercellular communication, where individual cell can perceive cell density by sensing signaling molecules concentration diffused in population. When cell density reaches the responding threshold, the cells adjust functional genes expression to alter phenotypic traits. Thus, QS eventually regulates population characteristics by such a manner that cannot be achieved by a single cell [22,23]. Microcystis, as G- bacterium-like prokaryote, has a similar cellular structure with G- bacteria. To explore fundamental mechanisms of MCBs outbreak for better MCBs control/management, many studies aimed to discover QS phenomenon in Microcystis.
Using semi-continuous culture experiment, Pereira et al. observed that different cell densities led to obviously different metabolites in Microcystis, and proposed the presence of QS phenomenon that causes different metabolites production in Microcystis [24]. Wood et al. found a significant positive correlation between MCs amount and Microcystis cell density, where MCs amount produced by each cell increases with rising cell density, suggesting that Microcystis cell density could drive some physiological processes [25,26]. Xie et al. and Wang et al. experimentally confirmed that the expression level of MCs-synthesizing genes and enhanced MCs concentration were positively proportional to Microcystis cell density, which further indicated that Microcystis cells could adjust their physiological metabolism and cell aggregation (mediated by MCs) by perceiving cell density [27,28]. Yet, above studies could not give a definite answer to the question that whether QS exists in Microcystis until the signaling molecules and its regulatory manner were identified in Microcystis.
2.2. Discovery and Recognition of Signaling Molecules in Microcystis
Bacterial QS signaling molecules are classified into four major categories: N-acyl homoserine lactones (AHLs), autoinducer peptides (AIPs), autoinducer-2 (AI-2) and autoinducer-3 (AI-3) [29,30,31]. Typically, G- bacteria mainly possess AHL-mediated QS system that secretes AHLs as signaling molecules for intraspecific communication. AHLs-mediated QS system has multiple signal-receptor gene homologs including LuxI/LuxR, LasI/LasR, RhlI/RhlR, AfeI/AfeR, BtaI/BtaR, and TofI/TofR [32]. AIPs are major QS signal molecules in Gram-positive (G+) bacteria for their intraspecific communication, while AI-2 (encoded by luxS gene) is an interspecific communication signal secreted by both G- and G+ bacteria [33]. AI-3 is a metabolite of previously unknown structure involving pathogenesis of Escherichia coli, which drove bacteria/host inter-kingdom communication [34,35].
Microcystis possesses similar cellular structure with G- bacteria, thus most studies speculated that the QS system of Microcystis might be alike to that of G- bacteria, where AHLs acted as signaling molecules to regulate Microcystis population characteristics. The molecular structure of AHLs consists of two parts, a homoserine lactone (HSL) ring and a variable amide side chain, so the diversity of AHLs structures is caused by differences in side chain length, substituent group and substituent position [36,37,38]. AHLs comprises short-chain (C4-HSL~C8-HSL) and long-chain (C10-HSL~C18-HSL) variants according to side chain length, while OH- and O- groups, as common substituent groups, often replace the H atom at C3 position of AHLs (Figure 1). Early in 2008, Sharif et al. demonstrated that cyanobacterium Gloeothece produces C8-AHL in axenic culture [39]. Over recent years, a growing number of studies have successfully extracted AHLs from axenic culture of Microcystis, and confirmed that AHLs could be synthesized and secreted by Microcystis. Using liquid chromatography-mass spectrometry (LC-MS) and bioreporter assay, Zhai et al. for the first time to evidence that Microcystis can produce AHLs in axenic culture [40]. Meanwhile, researchers revealed that AHLs concentration changed with dependence on Microcystis cell density, where AHLs concentration peaked as Microcystis cell density reached the required threshold [28,40]. It was also found that an obvious accumulation of AHLs occurred only when cell density threshold reached, which accorded with the regulation manner of many QS systems [39]. Above studies proved that Microcystis could depend on cell density to adjust AHLs production and secretion. Namely, Microcystis can produce AHLs as signaling molecules based on cell density, which has a potential to induce a series of physiological/behavioral changes at the concentration threshold of AHLs. The AHLs analogs produced by different Microcystis strains were summarized in Table 1. Table 1 presents information about previous studies that successfully extracted the molecules of AHLs from Microcystis, including media, molecular formula as well as Microcystis species.
3. AHLs' Roles in Regulating Physiology and Colonies Formation of Microcystis
3.1. Regulatory Effects of Endogenous AHLs
The roles of endogenous AHLs produced by Microcystis have been widely explored. Most studies extracted AHLs from Microcystis cells, and added such endogenous AHLs into Microcystis culture at growth stage to observe physiological and population changes of Microcystis. Endogenous AHLs were found to affect Microcystis in many aspects, including cellular morphology, physiological adaptation, nutrition/energy metabolism activities, and triggering cells aggregation for colonies formation.
Firstly, in aspect of cellular morphology, the secreted AHLs can deform cell wall, and promote the forming of more gas vesicles in Microcystis with increasing cell density. These columnar-shaped gas vesicles regulate cell buoyancy, which enables Microcystis to occupy the surface layer of water body that favors its competitive advantages maintenance in aquatic ecosystem [44]. For instance, Xu et al. identified novel long-chain AHLs from Microcystis, and demonstrated a significant correlation between AHLs concentration and cell density. By adding the AHLs into culture, the authors also confirmed that the AHLs enhanced cell buoyancy and the expression of vesicle-related genes [43]. This result evidenced that Microcystis could secrete AHLs to enhance cell buoyancy via vesicle formation, which allows Microcystis to occupy water surface layer to become advantageous competitor [28].
Secondly, in aspect of cellular physiological adaptability, endogenous AHLs can regulate and modify the production of MCs and EPSs. As widely reported, MCs may strengthen ecological fitness of Microcystis by resisting biochemical stressors (e.g., hydrogen peroxide, metal ions, predators) [45,46], raising the adaptability to high-radiation and oxidation conditions [46,47], enhancing competitiveness over its MC- counterparts [48], promoting large-size colonies formation [49,50,51], and helping Microcystis overwintering and recovery from cold environment [52,53]. EPSs also improve Microcystis resistance to stresses caused by many factors, such as grazing pressure by protozoa, allelochemical pressure by anti-cyanobacterial allelochemicals [54,55,56]. Thus, Microcystis can improve ecological adaption and stress-resistance by adjusting MCs and EPSs contents, which could be driven by AHLs. Wang et al. observed that the concentration of MC-LR (a common homolog of MCs) increased with Microcystis cell density in growth culture, but remained stable when cell density is kept constant by adding medium into culture, indicating a close correlation between MC-production and cell density of Microcystis. Meanwhile, the authors detected a similar trend in AHLs concentration, with AHLs being detectable even at low cell density where MCs were undetectable [27]. This phenomenon reflected that the MCs increase is caused by AHLs. Additionally, Xu et al. observed the promotive effects of AHLs on MC-LR secretion and MC-synthesis genes expression by using qPCR analysis [43].
Thirdly, in aspect of nutrition and energy metabolism activities, some AHLs homologs can activate relative enzymes to improve carbon and nitrogen metabolism efficacy in Microcystis, and can also adjust the synthesis and secretion of various metabolites by affecting genes expression and enzymes activity of multiple pathways. For instance, Yan et al. identified 3OH-C4-HSL (a AHL homolog) as QS signaling molecule of Microcystis, and verified that the addition of 3OH-C4-HSL could up-regulate QS-related genes Dpp and Sec, and the expression of genes related to NADH dehydrogenase, succinate dehydrogenase, cytochrome coxidase were also up-regulated to promote all ATP-synthesizing genes expression. Consequently, carbon and energy metabolisms were promoted in Microcystis [41]. This suggested that some AHLs homologs acted as the trigger to initiate a series of downstream metabolisms. Xu et al. found up-regulated expression of photosynthesis-related genes (e.g., apcABF、petE、psaBFK、psbUV), promoted nitrogen metabolism and ribosomal metabolism and increased content of chlorophyll by the action of AHLs extract [57].
Fourthly, in aspect of colonies formation, AHLs influence Microcystis cell aggregation to promote colonies formation, which involve EPSs concentration and composition controlled by AHLs [58]. As reviewed above, AHLs also affect cellular morphology, physiological adaptation and metabolism activities of nutrition and energy to expand the advantage of Microcystis in waters, thereby jointly affecting its cell growth and colonies formation. Zhai et al. found the existence of a special AHL molecule named (E)-7-hydroxy-5-oxo-N-(2oxotetrahydrofuran-3-yl) oct-2-enamide in pure culture of Microcystis aeruginosa PCC-7820, and such AHL promoted cell aggregation [40]. The sticky EPSs possess various functional groups (e.g., -OH, C-O) that strongly bind with Ca2+ and Mg2+ in waters, which assist Microcystis cells bind together to form bio-aggregates alike to flocculent sludge, bioparticle and biofilm [59,60]. Zhai et al. revealed that AHLs extracted from Microcystis promoted biofilm formation to induce cell aggregation [40]. Xu et al. discovered a decreased aggregating ability of Microcystis after cellular EPSs were extracted, and such decrease of Microcystis aggregation was more obvious in field sample than in lab-culture [61]. Noteworthy, most previous studies ascribed Microcystis cells aggregation to the stickiness of extracellular polysaccharides (ex-poly) in EPSs. However, more recent studies proposed the key function of extracellular proteins (ex-pro) of EPSs in promoting cells aggregation and colonies formation. For instances, Xu et al. found that AHLs extracted from Microcystis did not obviously promote EPSs syntheses and secretion, but increased ex-pro content in EPSs to enhance Zeta potential and hydrophobicity, thus promoting Microcystis cell aggregation [43]. Using bioinformatics and comparative genomics analysis, Qiu et al. found the existence of genes encoding PEP-CTERM domain proteins, and suggested that these genetically controlled proteins secreted on cell surface may form complex polymers with ex-poly through glycosylation process to induce cell aggregation and colonies formation [17].
Schematic diagram of AHLs-mediated QS for Microcystis colonies formation is shown in Figure 1. Concretely, AHLs concentration can increase with rising cell density. When cell density reaches a specific threshold, AHLs concentration culminates in aqueous phase to promote Microcystis cell aggregation and survival/competition advantages by improving cellular morphological/physiological adaptability and nutrition/energy metabolism, and thus promote colonies formation for MCBs occurrence (Figure 2).
3.2. Regulatory Effects of Exogenous AHLs
Besides endogenous AHLs produced by Microcystis, researchers also conducted extensive studies to explore the effects of exogenous AHLs on Microcystis. The premise of exploring exogenous AHLs function is that these AHLs produced by other organisms can affect Microcystis. Yan et al. found that the addition of 3-OH-C4-HSL extracted from bacterial sludge could promote microalgal growth, alike to the function of endogenous AHLs [41], whereas Xue et al. found that exogenous AHLs addition decreased Microcystis growth rate [62]. Based on numerous studies, exogenous AHLs affect Microcystis in several aspects alike to endogenous AHLs. This means that both endogenous and exogenous AHLs influence the same four aspects of Microcystis, including cellular morphology, physiological adaptation, nutrition/energy metabolism activities, and cells aggregation for colonies formation (Figure 2).
By adding exogenous AHLs into Microcystis culture, Xie et al. showed that exogenous AHLs could change cell wall shape, and the sizes of gas vesicles increased under the action of C4-HSL and C8-HSL [28]. Xie et al. also found that some exogenous AHLs raised the expression level of genes related to biological processes, cellular component synthesis and molecular function to promote Microcystis cells growth. Among them, the promoting effect of 3-OH-C4-HSL is most obvious [28]. However, exogenous C8-HSL had no promoting effect on Microcystis cells growth and even decreased the expression level of genes related to ATP enzymatic activity and hydrolyase activity. In aspect of physiological adaptation, EPSs and MCs content of Microcystis can be regulated and controlled by exogenous AHLs. Exogenous AHLs were observed to promote EPSs secretion of Microcystis, and the exogenous AHLs with growth-promoting effect could decrease MCs secretion but increase MCs syntheses, while the ones without growth-promoting effect caused high MCs content. This suggested that even the same exogenous AHLs analogs may pose opposite effects on EPSs and MCs secretion [28]. In aspect of nutrition/energy metabolism activities, a part of exogenous AHLs can change relative genes expression and enzymes activities to regulate metabolic processes such as carbon/nitrogen metabolism in Microcystis [28,62,63]. C6-HSL can increase carbon sequestration efficiency to promote Microcystis growth [62]. Under joint action of above three aspects, Microcystis cells growth and aggregation for colonies formation also changed. For instance, N-octanoyl-L-homoserine lactone (C8-HSL), N-(3-oxooctanoyl)-L-homoserine lactone (3-oxo-C8-HSL), and N-butyryl-DL-homoserine lactone (C4-HSL) can promote the formation of biofilm-like membrane on Microcystis and thus significantly strengthen colonies formation [40,64].
Notably, there is a close correlation between MCs syntheses/secrete and EPSs composition of Microcystis [50]. MCs can up-regulate ex-poly syntheses-related genes such as capD, csaB, tagH and epsL to significantly increase ex-poly content in EPSs [65,66]. As major component of EPSs, the tightly bound ex-poly can facilitate capsule formation around Microcystis cells to increase cell surface viscosity and promote colonies formation [17,67,68]. This provides further explanation for the positive correlation between MCs content and colony size. Based on above, AHLs not only act on MCs and EPSs secretion alone, but also regulate the linkage of MCs and EPSs.
Diverse AHLs variants with different molecular structures can exert vastly distinct functions in QS regulation pathway of G- bacteria [36,37,38]. Likewise, different AHLs variants also exert distinct effects on Microcystis [24,26,28,64]. As revealed by Xie et al., the AHLs with phenyl groups, oxo groups, ether groups and bromide substituents decrease MCs content in aqueous phase [42], and the AHLs differing in hydrophilicity and side chain length seemed to posed different influences on MCs syntheses and secretion by Microcystis [28]. Most existing studies focused on how cells growth and morphology, cells aggregation/colonies formation, chlorophyll content and photosynthesis of Microcystis are affected by adding exogenous AHLs variants. The effects of different exogenous AHLs variants addition on cell growth and physiological aspects of Microcystis are summarized in Table 2. Exogenous AHLs addition could mimic a naturally algal-bacterial co-existent circumstance where Microcystis is affected by AHLs secreted by other algae and/or bacteria. Owing to different AHLs variants used for addition experiments, the observed effects are always divergent (e.g, promotive or inhibitory effect) among experiments. Applying exogenous AHLs with inhibitory effect on Microcystis growth provides a new option for controlling and restricting MCBs outbreak.
4. Genetic Information of AHLs-Mediated QS in Microcystis
The AHLs-mediated QS system widely prevalent in G- bacteria is termed as LuxI/LuxR-type QS system, which includes AHLs synthase (termed 'LuxI protein') and AHLs receptor (termed 'LuxR protein'), with AHLs as signaling molecule [37,71]. In such system, luxI gene encodes AHLs synthetase, while luxR gene encodes AHLs receptor to sense AHLs and become transcription activators for other genes (e.g., luxI, luxA, luxB, luxC, luxD, luxE), among which luxA-E participate in regulating specific behaviors caused by QS [30,72,73,74,75,76,77]. AHLs synthesized in cells can be diffused to surroundings via simple diffusion and/or transport by specific transporter [22,78]. With rising cell density, AHLs gradually accumulate in surroundings. Only when AHLs concentration accumulates up to specific threshold, AHLs can bind with LuxR protein to form signal-receptor complex dimers/multimers to activate downstream genes expression and induce corresponding behavioral changes of G- bacteria [71,79]. The AHLs concentration threshold required for activating specific gene expression is highly specific. AHLs act as a trigger to activate differential functional genes in sequence when each specific threshold reaches [78,80].
Because Microcystis shares a similar cellular structure and similar QS signaling molecule to G- bacteria, so it is questioned that ‘does any similarity present in genetic mechanisms of QS between Microcystis and G- bacteria?’ Xie et al. conducted genetic-level research for QS in Microcystis, and proposed that AHLs initiated QS by affecting expression of QS-related genes, such as BH695_RS06140, but the authors did not identify gene function [28]. Chen et al. identified slr2100 and slr1259 gene that homologous to LuxI and AiiA gene (AHLs-lytic enzyme gene), respectively, in cyanobacterium Synechocystis sp. PCC6803, and found that slr2100 and slr1259 gene expression was detected in Microcystis at different growth stages. This implies that Microcystis can express homologous proteins to LuxI and AiiA for AHLs syntheses and degradation, respectively, which causes self-regulation of AHLs level. Chen et al. proposed that the QS system in Microcystis is likely to the AHLs-mediated QS system of G- bacteria (i.e., LasI/LasR-type QS system in Pseudomonas aeruginosa) [63]. Interestingly, Gisella et al. found luxS gene expression in Microcystis aeruginosa PCC7806 [79]. LuxS gene encodes the crucial enzyme for synthesizing AI-2, which is the QS signaling molecule for another QS system [22,29,30]. Despite this finding, AI-2 as QS signal is not yet identified in Microcystis until now. Hence, it is unclear for whether there is AI-2-mediated QS system in Microcystis and whether luxS gene in Microcystis functioned as same as in bacteria to encode AI-2 synthetase.
At present, the enzymes and signaling molecules of these two possible QS systems have not been detected simultaneously. However, the LuxI / LuxR-type QS system is more likely the QS system in Microcystis, based on the evidence of slr2100 and slr1259 gene expression in Microcystis found [63]. This system model suggested that i) when Microcystis cell density is low, AHLs concentration remained at low level, so AHLs cannot be sensed and bound by AHLs receptor, thus failing to activate various genes expression; ii) when Microcystis cell density is high, AHLs concentration increases up to high level, so AHLs can bind with AHLs receptor to form signal-receptor complex, which in turn activates AHLs-syntheses and induces other specific genes expression to produce functional proteins (Figure 3). However, substantial conclusive evidences for LasI/LasR-type or LuxI/LuxR-type QS system presence in Microcystis are still insufficient, and genetic information for AHLs-mediated QS in Microcystis remains largely lacking, which deserve urgent research.
5. Quorum Quenching (QQ) of Microcystis and Application Implication
QQ is a phenomenon that negatively disrupt microbial colonies formation and population behavior by interfering with any step of QS process, such as inhibiting AHLs syntheses, promoting AHLs degradation and disrupting AHLs binding to receptor [28]. In G- bacteria, Several substances have inhibitory effect on AHLs syntheses, such as the structural analogs of intermediates during AHLs syntheses process, purine nucleotide, the derivative of HSL, and the homologs and analogs of some antibiotics [81]. Researchers have found some of these substances disrupt AHLs syntheses process by competing with intermediates [82]. For examples, as AHLs synthase, LuxI can catalyze the formation of an amide bond between S-adenosylmethionine (SAM, an essential intermediate) and acyl-acyl carrier protein for AHLs syntheses. The structural analogs of SAM (e.g.,S-adenosylhomocysteine, S-adenosylcysteine and sinefungin ) have inhibitory effect on AHLs syntheses to cause QQ. Parsek et al. Identified that the inhibiting function may be caused by the competition between the SAM analog and SAM [6].
Besides, AHLs can be degraded in vivo through biochemical metabolisms and specific enzymes including AHL-acylase, AHL-lactonase and oxidoreductases, and synthetic AHLs structural analogues compete with corresponding AHLs signals for the binding sites of AHLs receptor, thus AHLs binding to receptor can be prevented [37,71,81]. The expression of sll1392 gene that homologous to LuxR gene was suppressed but slr1259 gene was stimulated after adding exogenous AHLs structural analog (i.e., ɑ-amino-γ-butyrolactone hydrobromide) into Synechocystis culture. Such response of gene expression to AHLs structural analog is alike to corresponding response manner of AHLs-mediated QS system in G- bacteria. These suggested that cyanobacterial QS could be quenched by promoting AHLs degradation and disrupting AHLs binding to receptor, and QQ mechanism in cyanobacteria might be similar to that in G- bacteria [63]. Romero et al. indicated that an acylases named AiiC in cyanobacterium Anabaena sp. PCC 7120 might be responsible for its self-regulation of AHLs level, and recombinant AiiC could degrade a series of AHLs molecules, thus causing QQ against QS signals [69]. Although the homologous gene to AiiC was not identified in Microcystis, yet AiiA homologous gene (slr1259) encoding AHLs-lytic enzyme can be expressed in Microcystis, implying that AHLs degradation was mediated by different genes to cause QQ in different cyanobacteria. Based on above, a schematic diagram for QQ mechanisms in Microcystis was summarized in Figure 4.
Consequently, QS of Microcystis is not only regulated by AHLs signals that secreted by its own cells, but also negatively impacted by those secreted by other organisms such as bacteria. The principle of QQ provides a new strategy to develop eco-benign algicidal method to control toxigenic MCBs caused by excessive proliferation and colonies formation of Microcystis. Exogenous AHLs produced by other algae and bacteria may impact Microcystis growth in bacteria-algae co-existence environments. For instances, Zhou et al. reported that some bacteria secreted α-amino-γ-butyrolactone hydrobromate (acyl-HSLs) to impact Microcystis growth [56], and Manuel et al. found that chlorophyta greatly disrupted cyanobacterial QS through QQ effect in their co-existence system [69]. Based on this, researchers could control MCBs by introducing QQ signals- secreting microorganisms. To achieve QQ, many studies also applied exogenous AHLs and/or AHLs structural analogs as QS inhibitors against Microcystis cell proliferation [62]. For instances, Microcystis aeruginosa and Synechocystis sp. growth were inhibited by acyl-HSLs addition [83]. The addition of C4-HSL and C8-HSL caused obvious down-regulation of major genes related to QS in Microcystis to decelerate QS process [28].
Researchers also revealed that a variety of plant-originated and synthetic organic compounds could serve as QS inhibitors, such as vanillin extracted from vanilla bean and a heterocyclic oxygenic compound named furan [28,41,69]. Zhai et al. found that one of the furanone, dihydro-3-amino-2-(3H)-furanone (FN) and Microcystis's QS signaling molecule could bind to the same site on receptor [40]. Thus, FN can compete with QS signal of Microcystis for receptor, indicating that FN acts as QS inhibitor by blocking receptor-binding. Further research reported that FN could bind with such two residues as Asn164 (A) and His167 (A) of AHLs receptor LuxR to form hydrogen bonds. The hydrophobic interactions between FN and binding pockets formed by residues Gln137(A), Val140-(A), Ala163(A), Asn164(A) and His167(A) also enables FN to have more competitive advantage in receptor-binding than QS signal [41]. Such advantage effectively inhibited carbon assimilation and energy metabolism of Microcystis to control its cell density.
6. Future Perspectives
Compared to G- bacteria, research on QS and QQ of Microcystis is relatively insufficient, especially in aspects of molecular mechanisms, so genetic information on QS- and QQ-related genes and associated function should be further screened and probed in Microcystis. Thus, bioinformatics (e.g., sequence blast and alignment) could be used to clarify whether Microcystis possesses QS- and QQ-related genes homologous to those in Anabaena and other G- bacteria. By screening new target genes, we can unveil gene function and track the action pathways of QS and QQ in Microcystis using gene knockout and isotopic tracer techniques for better understanding on QS and QQ mechanisms of Microcystis, which will lay a solid theoretical basis for toxigenic MCBs control.
Owing to climate change and eutrophication, MCBs intensity and frequency may continuously increase [84], so the interaction of Microcystis’s AHLs and other microorganisms should be elucidated to facilitate understanding on complex roles of Microcystis’s AHLs in aquatic ecosystem. It is also essential to develop an efficient and specific AHLs-detection technology as technical support for harnessing MCBs by promptly gaining QS signals level and AHLs species distribution. Meanwhile, how environmental factors (e.g., light, temperature, nutrient salts level) affect aqueous AHLs species distribution and AHLs' effect should be concerned, which is helpful in predicting AHLs-mediated outbreak of MCBs as environment factors change in context of climate change. Based on this review, we suggested following research directions that crucial for advancing knowledge understanding in this field and developing strategies to monitor, control and harness MCBs, which deserve further attention and comprehensive research:
i) Elucidating the molecular mechanisms for QS and QQ of Microcystis, especially clarifying related gene functions and action pathway, using modern molecular biological technologies and bioinformatics;
ii) Further identifying AHLs species produced by Microcystis, and exploring their effects on growth and physiology of many other bloom-forming microalgae;
iii) Exploring other QS signaling molecules besides AHLs in Microcystis, and whether some AHLs species produced by Microcystis can inhibit Microcystis’s QS, and comparing the molecular structure difference between QS-inducing and QS-inhibiting AHLs for Microcystis;
iv) Surveying the effect of diverse environmental factors on aqueous AHLs species distribution and AHLs' effect.
7. Conclusive Remarks
Microcystis can secrete AHLs as QS signaling molecules to regulate its population characteristics and behaviors. As previously proposed, the diffused concentration of endogenous AHLs increases with rising cell density until a specific threshold reaches for binding with AHLs receptor to form signal-receptor complex, which can further activate functional gene expression. Consequently, AHLs-mediated QS system regulates physiological/behavioral changes of Microcystis population. The QS system promotes morphological/physiological adaptability of Microcystis through enhancing cell density, cell buoyancy via gas vesicle formation, EPSs and MCs secretion, nutrition and energy metabolism, which ultimately cause cell aggregation and colonies formation of Microcystis to strengthen its survival/competition advantages. Notably, differences in molecule structures of exogenous AHLs and/or exogenous AHLs structural analogs secreted by other bacteria/algae exert distinct regulatory effects on Microcystis physiology and behavior, especially those secreted by some specific bacteria may pose inhibitory or quenching effect on Microcystis QS. This suggested that applying QS inhibitor to interfere QS signaling pathway could be a promising strategy for MCBs control.
However, several deficiencies in field of Microcystis QS and QQ still exist. To further explore genetic mechanisms at molecular level, as well as species distribution and complex roles of QS signals under influence of environmental factors, is imperatively desirable. Overall, by encompassing research progress, this review provided an updated comprehensive review on Microcystis QS, and proposed new insights on future research direction to advance understanding on underlying mechanisms.
Acknowledgments
This work was funded by National Natural Scientific Foundation of China (No. 32271707, 31872694).
References
- Carroll, S.B. Chance and necessity: The evolution of morphological complexity and diversity. Nature 2001, 409, 1102–1109. [Google Scholar] [CrossRef]
- Gagala, I.; Izydorczyk, K.; Jurczak, T.; Pawelczyk, J.; Dziadek, J.; Wojtal-Frankiewicz, A.; Jozwik, A.; Jaskulska, A.; Mankiewicz-Boczek, J. Role of environmental factors and toxic genotypes in the regulation of microcystins-producing cyanobacterial blooms. Microb. Ecol. 2014, 67, 465–479. [Google Scholar] [CrossRef]
- Zuo, J.; Chen, L.T.; Shan, K.; Hu, L.L.; Song, L.R.; Gan, N.Q. Assessment of different mcy genes for detecting the toxic to non-toxic Microcystis ratio in the field by multiplex qPCR. J. Oceanol. Limnol. 2018, 36, 1132–1144. [Google Scholar] [CrossRef]
- Song, W.J.; Wang, W.X.; Qiu, D.H.; Zheng, W.Z.; Li, X. Evaluating the effectiveness of various biochemical and molecular techniques to assess microcystin risk during the onset process of Microcystis blooms (delay-development stages). J. Cleaner Prod. 2022, 369, 133335. [Google Scholar] [CrossRef]
- Guo, Z.H.; Li, J.M.; Luo, D.; Zhang, M.X. Novel ecological implications of non-toxic Microcystis towards toxic ecotype in population-promoting toxic ecotype dominance at various n levels and cooperative defense against luteolin-stress. FEMS Microbiol. Ecol. 2023, 99, 11–17. [Google Scholar] [CrossRef]
- Li, J.M.; Li, R.H.; Li, J. Current research scenario for microcystins biodegradation - a review on fundamental knowledge, application prospects and challenges. Sci. Total Environ. 2017, 595, 615–632. [Google Scholar] [CrossRef]
- Li, J.M.; Shimizu, K.; Zhou, Y.L.; Utsumi, M.; Sakharkar, M.K.; Zhang, Z.Y.; Sun, H.W.; Sugiura, N. Biodegradation of microcystins by bacterial communities co-existing with the flagellate monas guttula and concurrent succession of community structures. J Water Supply Res T 2011, 60, 352–363. [Google Scholar] [CrossRef]
- Shen, H.; Niu, Y.; Xie, P.; Tao, M.; Yang, X. Morphological and physiological changes in Microcystis aeruginosa as a result of interactions with heterotrophic bacteria. Freshwater Biol. 2011, 56, 1065–1080. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Shiah, F.-K.; Chen, Y.-L. Importance of large colony formation in bloom-forming cyanobacteria to dominate in eutrophic ponds. Ann Limnol-int J Lim 2011, 47, 167–173. [Google Scholar] [CrossRef]
- Visser, P.M.; Ibelings, B.W.; Mur, L.R.; Walsby, A.E. The ecophysiology of the harmful cyanobacterium Microcystis. In Harmful cyanobacteria, Huisman, J., Matthijs, H.C.P., Visser, P.M., Eds.; Springer Netherlands: Dordrecht, 2006; Volume 3, pp. 109–142. [Google Scholar]
- Bi, X.D.; Dai, W.; Zhang, S.L.; Xing, K.Z.; Guo, Y.J. Research progress on the competitive advantages and formation mechanism of Microcystis colony. Environ. Sci. Technol. 2014, 37, 41–44,65. [Google Scholar]
- Liu, Y.; Xu, Y.; Wang, Z.J.; Xiao, P.; Yu, G.L.; Wang, G.X.; Li, R.H. Dominance and succession of Microcystis genotypes and morphotypes in lake taihu, a large and shallow freshwater lake in china. Environ. Pollut. 2016, 219, 399–408. [Google Scholar] [CrossRef]
- Lehman, P.W.; Kurobe, T.; Teh, S.J. Impact of extreme wet and dry years on the persistence of Microcystis harmful algal blooms in san francisco estuary. Quat. Int. 2022, 621, 16–25. [Google Scholar] [CrossRef]
- Zhou, J.; Yang, G.J.; Qin, B.Q.; Zhang, G.S.; Wang, L.J.; Gao, Y.H.; Li, H.Y.; Chen, M.Y.; Zhong, C.N. Effect of the ingestion of metazooplankton on the formation of Microcystis blooms in summer in lake taihu. Sci. Limnol. Sin. 2013, 25, 398–405. [Google Scholar]
- Van Le, V.; Srivastava, A.; Ko, S.-R.; Ahn, C.-Y.; Oh, H.-M. Microcystis colony formation: Extracellular polymeric substance, associated microorganisms, and its application. Bioresour. Technol. 2022, 360, 127610. [Google Scholar] [CrossRef]
- Qiu, D.R. Biosynthesis pathway of extracellular polymeric substances and colonial formation of cyanobacteria underlying water blooms of Microcystis. Acta Hydrobiol. Sin. 2020, 44, 1008–1013. [Google Scholar] [CrossRef]
- Wu, Q.Q.; Wu, K.; Ye, Y.W.; Dong, X.H.; Zhang, J.M. Quorum sensing and its roles in pathogenesis among animal-associated pathogens-a eview. Acta Microbiol. Sin. 2009, 49, 853–858. [Google Scholar]
- Sanchez, S.; Ng, W.-L. Motility control as a possible link between quorum sensing to surface attachment in vibrio species. Adv. Exp. Med. Biol. 2023, 1404, 65–75. [Google Scholar] [CrossRef]
- Eickhoff, M.J.; Bassler, B.L. Snapshot: Bacterial quorum sensing. Cell 2018, 174, 1328. [Google Scholar] [CrossRef]
- Mangwani, N.; Dash, H.R.; Chauhan, A.; Das, S. Bacterial quorum sensing: Functional features and potential applications in biotechnology. J. Mol. Microbiol. Biotechnol. 2012, 22, 215–227. [Google Scholar] [CrossRef]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal-response systems in gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef]
- Waters, C.M.; Bossier, B.L. Quorum sensing: Cell-to-cell communication in bacteria. In Annu. Rev. Cell dev. Biol.; Annual review of cell and developmental biology; 2005; Volume 21, pp. 319–346.
- Pereira, D.A.; Giani, A. Cell density-dependent oligopeptide production in cyanobacterial strains. FEMS Microbiol. Ecol. 2014, 88, 175–183. [Google Scholar] [CrossRef]
- Wood, S.A.; Rueckert, A.; Hamilton, D.P.; Cary, S.C.; Dietrich, D.R. Switching toxin production on and off: Intermittent microcystin synthesis in a Microcystis bloom. Environ. Microbiol. Rep. 2011, 3, 118–124. [Google Scholar] [CrossRef]
- Wood, S.A.; Dietrich, D.R.; Cary, S.C.; Hamilton, D.P. Increasing Microcystis cell density enhances microcystin synthesis: A mesocosm study. Inland Waters 2012, 2, 17–22. [Google Scholar] [CrossRef]
- Wang, S.L.; Ding, P.; Lu, S.Y.; Wu, P.; Wei, X.Q.; Huang, R.X.; Kai, T.H. Cell density-dependent regulation of microcystin synthetase genes (mcy) expression and microcystin-lr production in Microcystis aeruginosa that mimics quorum sensing. Ecotoxicol. Environ. Saf. 2021, 220, 112330. [Google Scholar] [CrossRef]
- Xie, Y.T. Preliminary study on signaling molecules regulating the synthesis and secretion of microcystins in Microcystis aeruginosa[D]. Changchun: Northeast Normal University, 2021.
- Dai, C.X.; Qu, Y.Y.; Wu, W.Z.; Li, S.Z.; Chen, Z.; Lian, S.Y.; Jing, J.W. Qsp: An open sequence database for quorum sensing related gene analysis with an automatic annotation pipeline. Water Res. 2023, 235, 119814. [Google Scholar] [CrossRef]
- Mi, J.Q.; Yu, Z.Y.; Yu, H.; Zhou, W.B. Quorum sensing systems in foodborne salmonella spp. And corresponding control strategies using quorum sensing inhibitors for food storage. Trends Food Sci. Technol. 2024, 144, 104320. [Google Scholar] [CrossRef]
- Yi, L.; Dong, X.; Grenier, D.; Wang, K.C.; Wang, Y. Research progress of bacterial quorum sensing receptors: Classification, structure, function and characteristics. Science of the Total Environment 2021, 763, 143031. [Google Scholar] [CrossRef]
- Liu, L.L.; Zeng, X.Y.; Zheng, J.; Zou, Y.M.; Qiu, S.Y.; Dai, Y.F. Ahl-mediated quorum sensing to regulate bacterial substance and energy metabolism: A review. Microbiol. Res. 2022, 262, 127102. [Google Scholar] [CrossRef]
- Bassler, B.L. How bacteria talk to each other: Regulation of gene expression by quorum sensing. Curr. Opin. Microbiol. 1999, 2, 582–587. [Google Scholar] [CrossRef]
- Markus, V.; Paul, A.A.; Terali, K.; Ozer, N.; Marks, R.S.; Golberg, K.; Kushmaro, A. Conversations in the gut: The role of quorum sensing in normobiosis. Int. J. Mol. Sci. 2023, 24, 3722. [Google Scholar] [CrossRef]
- Kim, C.S.; Gatsios, A.; Cuesta, S.; Lam, Y.C.; Wei, Z.; Chen, H.W.; Russell, R.M.; Shine, E.; Wang, R.R.; Wyche, T.P.; et al. Characterization of autoinducer-3 structure and biosynthesis in e. Coli. ACS Cent. Sci. 2020, 6, 197–206. [Google Scholar] [CrossRef]
- Dow, L. How do quorum-sensing signals mediate algae-bacteria interactions? Microorganisms 2021, 9, 1391. [Google Scholar] [CrossRef]
- Huang, J.H.; Shi, Y.H.; Zeng, G.M.; Gu, Y.L.; Chen, G.Q.; Shi, L.X.; Hu, Y.; Tang, S.; Zhou, J.X. Acyl-homoserine lactone-based quorum sensing and quorum quenching hold promise to determine the performance of biological wastewater treatments: An overview. Chemosphere 2016, 157, 137–151. [Google Scholar] [CrossRef]
- Bhedi, C.D.; Prevatte, C.W.; Lookadoo, M.S.; Waikel, P.A.; Gillevet, P.M.; Sikaroodi, M.; Campagna, S.R.; Richardson, L.L. Elevated temperature enhances short-to medium-chain acyl homoserine lactone production by black band disease-associated vibrios. FEMS Microbiol. Ecol. 2017, 93, fix005. [Google Scholar] [CrossRef]
- Sharif, D.I.; Gallon, J.; Smith, C.J.; Dudley, E. Quorum sensing in cyanobacteria: N -octanoyl-homoserine lactone release and response, by the epilithic colonial cyanobacterium gloeothece pcc6909. ISME J. 2008, 2, 1171–1182. [Google Scholar] [CrossRef]
- Zhai, C.M.; Zhang, P.; Shen, F.; Zhou, C.X.; Liu, C.H. Does Microcystis aeruginosa have quorum sensing? FEMS Microbiol. Lett. 2012, 336, 38–44. [Google Scholar] [CrossRef]
- Yan, G.; Fu, L.; Ming, H.; Chen, C.L.; Zhou, D.D. Exploring an efficient and eco-friendly signaling molecule and its quorum quenching ability for controlling Microcystis blooms. Environ. Sci. Technol. 2023, 57, 16929–16939. [Google Scholar] [CrossRef]
- Lamas-Samanamud, G.; Reeves, T.; Tidwell, M.; Bohmann, J.; Lange, K.; Shipley, H. Changes in chemical structure of n-acyl nomoserine lactones and their effects on microcystin expression from Microcystis aeruginosa pcc7806. Environ. Eng. Sci. 2022, 39, 29–38. [Google Scholar] [CrossRef]
- Xu, C.; Ni, L.; Li, S.; Du, C.; Sang, W.; Jiang, Z. Quorum sensing regulation in Microcystis aeruginosa: Insights into ahl-mediated physiological processes and mc-lr production. Science of the Total Environment 2024, 919, 170867. [Google Scholar] [CrossRef]
- Xu, R.; Long, H.; Wang, Y.H.; Huang, K.Y. A new method for isolating gas vesicles from Microcystis for ultrasound contrast. Shengwu Gongcheng Xuebao 2022, 38, 1589–1601. [Google Scholar] [CrossRef]
- Utkilen, H.; Gjolme, N. Iron-stimulated toxin production in Microcystis-aeruginosa. Appl. Environ. Microbiol. 1995, 61, 797–800. [Google Scholar] [CrossRef]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Boerner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of Microcystis under oxidative stress conditions. Plos One 2011, 6, e17615. [Google Scholar] [CrossRef]
- Meissner, S.; Steinhauser, D.; Dittmann, E. Metabolomic analysis indicates a pivotal role of the hepatotoxin microcystin in high light adaptation of Microcystis. Environ. Microbiol. 2015, 17, 1497–1509. [Google Scholar] [CrossRef]
- Yang, J.; Deng, X.R.; Xian, Q.M.; Qian, X.; Li, A.M. Allelopathic effect of Microcystis aeruginosa on Microcystis wesenbergii : microcystin-lr as a potential allelochemical. Hydrobiologia 2014, 727, 65–73. [Google Scholar] [CrossRef]
- Sedmak, B.; Elersek, T. Microcystins induce morphological and physiological changes in selected representative phytoplanktons. Microb. Ecol. 2006, 51, 508–515. [Google Scholar] [CrossRef]
- Gan, N.Q.; Xiao, Y.; Zhu, L.; Wu, Z.X.; Liu, J.; Hu, C.L.; Song, L.R. The role of microcystins in maintaining colonies of bloom-forming Microcystis spp. Environ. Microbiol. 2012, 14, 730–742. [Google Scholar] [CrossRef]
- Harke, M.J.; Steffen, M.M.; Gobler, C.J.; Otten, T.G.; Wilhelm, S.W.; Wood, S.A.; Paerl, H.W. A review of the global ecology, genomics, and biogeography of the toxic cyanobacterium, Microcystis spp. Harmful Algae 2016, 54, 4–20. [Google Scholar] [CrossRef]
- Ihle, T.; Jähnichen, S.; Benndorf, J. Wax and wane of Microcystis (cyanophyceae) and microcystins in lake sediments:: A case study in quitzdorf reservoir (germany). J. Phycol. 2005, 41, 479–488. [Google Scholar] [CrossRef]
- Schatz, D.; Keren, Y.; Vardi, A.; Sukenik, A.; Carmeli, S.; Boerner, T.; Dittmann, E.; Kaplan, A. Towards clarification of the biological role of microcystins, a family of cyanobacterial toxins. Environ. Microbiol. 2007, 9, 965–970. [Google Scholar] [CrossRef]
- Yin, L.; Xu, Y.; Kong, D.S.; Wang, J.; Shi, K.P.; Zhang, Y.; He, H.; Yang, S.G.; Ni, L.X.; Li, S.Y. Role of extracellular polymeric substances in resistance to allelochemical stress on Microcystis aeruginsosa and its mechanism. J. Oceanol. Limnol. 2023, 41, 2219–2231. [Google Scholar] [CrossRef]
- U, B.; P, H.; S, D.; P, B. Effects of the mixotrophic flagellate ochromonas sp. On colony formation in Microcystis aeruginosa. Aquat. Ecol. 2001, 35, 11–17. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Shi, X.L.; Zhang, M.; Xing, P.; Cao, H.S. Changes in the morphology and polysaccharide content of Microcystis aeruginosa (cyanobacteria) during flagellate grazing. J. Phycol. 2008, 44, 716–720. [Google Scholar] [CrossRef]
- Xu, C.; Ni, L.X.; Du, C.H.; Shi, J.H.; Ma, Y.S.; Li, S.Y.; Li, Y.P. Decoding Microcystis aeruginosa quorum sensing through ahl-mediated transcriptomic molecular regulation mechanisms. Sci. Total Environ. 2024, 926, 172101. [Google Scholar] [CrossRef]
- Zeng, X.Y.; Zou, Y.M.; Zheng, J.; Qiu, S.Y.; Liu, L.L.; Wei, C.Y. Quorum sensing-mediated microbial interactions: Mechanisms, applications, challenges and perspectives. Microbiol. Res. 2023, 273, 127414. [Google Scholar] [CrossRef]
- Xu, H.C.; Lv, H.; Liu, X.; Wang, P.F.; Jiang, H.L. Electrolyte cations binding with extracellular polymeric substances enhanced Microcystis aggregation: Implication for Microcystis bloom formation in eutrophic freshwater lakes. Environ. Sci. Technol. 2016, 50, 9034–9043. [Google Scholar] [CrossRef]
- Xiao, R.; Zheng, Y. Overview of microalgal extracellular polymeric substances (eps) and their applications. Biotechnol. Adv. 2016, 34, 1225–1244. [Google Scholar] [CrossRef]
- Xu, H.C.; Jiang, H.L.; Yu, G.H.; Yang, L.Y. Towards understanding the role of extracellular polymeric substances in cyanobacterial Microcystis aggregation and mucilaginous bloom formation. Chemosphere 2014, 117, 815–822. [Google Scholar] [CrossRef]
- Liao, L.; Chen, B.; Deng, K.K.; He, Q.; Lin, G.J.; Guo, J.S.; Yan, P. Effect of the n-hexanoyl-l-homoserine lactone on the carbon fixation capacity of the algae–bacteria system. Int. J. Environ. Res. Public Health 2023, 20, 5047. [Google Scholar] [CrossRef]
- Chen, X.L. Inhibition of ahls analog on the growth of bloom-forming cyanobacteria and its mathematic model[D]. Kunming: Yunnan University, 2008.
- Herrera, N.; Echeverri, F. Evidence of quorum sensing in cyanobacteria by homoserine lactones: The origin of blooms. Water 2021, 13, 1831. [Google Scholar] [CrossRef]
- Rzymski, P.; Klimaszyk, P.; Jurczak, T.; Poniedzialek, B. Oxidative stress, programmed cell death and microcystin release in Microcystis aeruginosa in response to daphnia grazers. Front Microbiol 2020, 11, 1201. [Google Scholar] [CrossRef]
- Gan, N.; Xiao, Y.; Zhu, L.; Wu, Z.; Liu, J.; Hu, C.; Song, L. The role of microcystins in maintaining colonies of bloom-forming Microcystis spp. Environ Microbiol 2012, 14, 730–742. [Google Scholar] [CrossRef]
- De Philippis, R.; Vincenzini, M. Exocellular polysaccharides from cyanobacteria and their possible applications. FEMS Microbiol. Rev. 1998, 22, 151–175. [Google Scholar] [CrossRef]
- Thornton, D. Diatom aggregation in the sea: Mechanisms and ecological implications. Eur. J. Phycol. 2002, 37, 149–161. [Google Scholar] [CrossRef]
- Romero, M.; Diggle, S.P.; Heeb, S.; Camara, M.; Otero, A. Quorum quenching activity in anabaena sp pcc 7120:: Identification of aiic, a novel ahl-acylase. FEMS Microbiol. Lett. 2008, 280, 73–80. [Google Scholar] [CrossRef]
- Xue, H.F.; Zhou, Y.; Luo, K.; Chen, X.L. The influence of ahls and its analog on the biosynthesis of uv-protectants in cyanobacteria. J. Yunnan Natl. Univ. 2009, 18, 135–138. [Google Scholar]
- Lade, H.; Paul, D.; Kweon, J.H. N-acyl homoserine lactone-mediated quorum sensing with special reference to use of quorum quenching bacteria in membrane biofouling control. BioMed Res. Int. 2014, 2014, 162584. [Google Scholar] [CrossRef]
- Engebrecht, J.; Silverman, M. Identification of genes and gene-products necessary for bacterial bioluminescence. PNAS 1984, 81, 4154–4158. [Google Scholar] [CrossRef]
- Piper, K.R.; Vonbodman, S.B.; Farrand, S.K. Conjugation factor of agrobacterium-tumefaciens regulates ti-plasmid transfer by autoinduction. Nature 1993, 362, 448–450. [Google Scholar] [CrossRef]
- Passador, L.; Cook, J.M.; Gambello, M.J.; Rust, L.; Iglewski, B.H. Expression of pseudomonas-aeruginosa virulence genes requires cell-to-cell communication. Science 1993, 260, 1127–1130. [Google Scholar] [CrossRef]
- Bainton, N.J.; Bycroft, B.W.; Chhabra, S.R.; Stead, P.; Gledhill, L.; Hill, P.J.; Rees, C.E.D.; Winson, M.K.; Salmond, G.P.C.; Stewart, G.; et al. A general role for the lux autoinducer in bacterial-cell signaling - control of antibiotic biosynthesis in erwinia. Gene 1992, 116, 87–91. [Google Scholar] [CrossRef]
- Shadel, G.S.; Devine, J.H.; Baldvvin, T.O. Control of the lux regulon of vibrio fischeri. J. Biolumin. Chemilumin. 1990, 5, 99–106. [Google Scholar] [CrossRef]
- Egland, K.A.; Greenberg, E.P. Quorum sensing in vibrio fischeri :: Elements of the luxl promoter. Mol. Microbiol. 1999, 31, 1197–1204. [Google Scholar] [CrossRef]
- Rasmussen, T.B.; Givskov, M. Quorum sensing inhibitors: A bargain of effects. Microbiol-Sgm 2006, 152, 895–904. [Google Scholar] [CrossRef]
- Lamas-Samanamud, G.; Montante Iii, A.; Mertins, A.; Phan, D.; Loures, C.; Naves, F.; Reeves, T.; Shipley, H.J. The role of quorum sensing in the development of Microcystis aeruginosa blooms: Gene expression. Microorganisms 2023, 11, 383. [Google Scholar] [CrossRef]
- Schuster, M.; Lostroh, C.P.; Ogi, T.; Greenberg, E.P. Identification, timing, and signal specificity of pseudomonas aeruginosa quorum-controlled genes:: A transcriptome analysis. J. Bacteriol. 2003, 185, 2066–2079. [Google Scholar] [CrossRef]
- Cui, T.Q.; Bai, F.L.; Li, J.R. Advance on quorum-sensing regulation and quenching mechanism of gram-negative bacteria mediated by ahls. Journal of Chinese Institute of Food Science and Technology 2020, 20, 308–320. [Google Scholar] [CrossRef]
- Parsek, M.R.; Val, D.L.; Hanzelka, B.L.; Cronan, J.E.; Greenberg, E.P. Acyl homoserine-lactone quorum-sensing signal generation. PNAS 1999, 96, 4360–4365. [Google Scholar] [CrossRef]
- Zhou, L.J.; Chen, X.L.; Wang, B.; Chen, S.N. The effects of exogenous ahls on the growth and metabolism of blue-green algae. J. Yunnan Univ., Nat. Sci. Ed. 2007, 29, 303–307. [Google Scholar]
- Hutchins, D.A.; Jansson, J.K.; Remais, J.V.; Rich, V.I.; Singh, B.K.; Trivedi, P. Climate change microbiology - problems and perspectives. Nat Rev Microbiol 2019, 17, 391–396. [Google Scholar] [CrossRef]
Figure 1.
The process of Microcystis colonies formation mediated by AHLs. The AHLs secreted by Microcystis increased with the Microcystis cell density rising. In the process of cell proliferation, single Microcystis cell form the Microcystis colony under the influence of AHLs.
Figure 1.
The process of Microcystis colonies formation mediated by AHLs. The AHLs secreted by Microcystis increased with the Microcystis cell density rising. In the process of cell proliferation, single Microcystis cell form the Microcystis colony under the influence of AHLs.
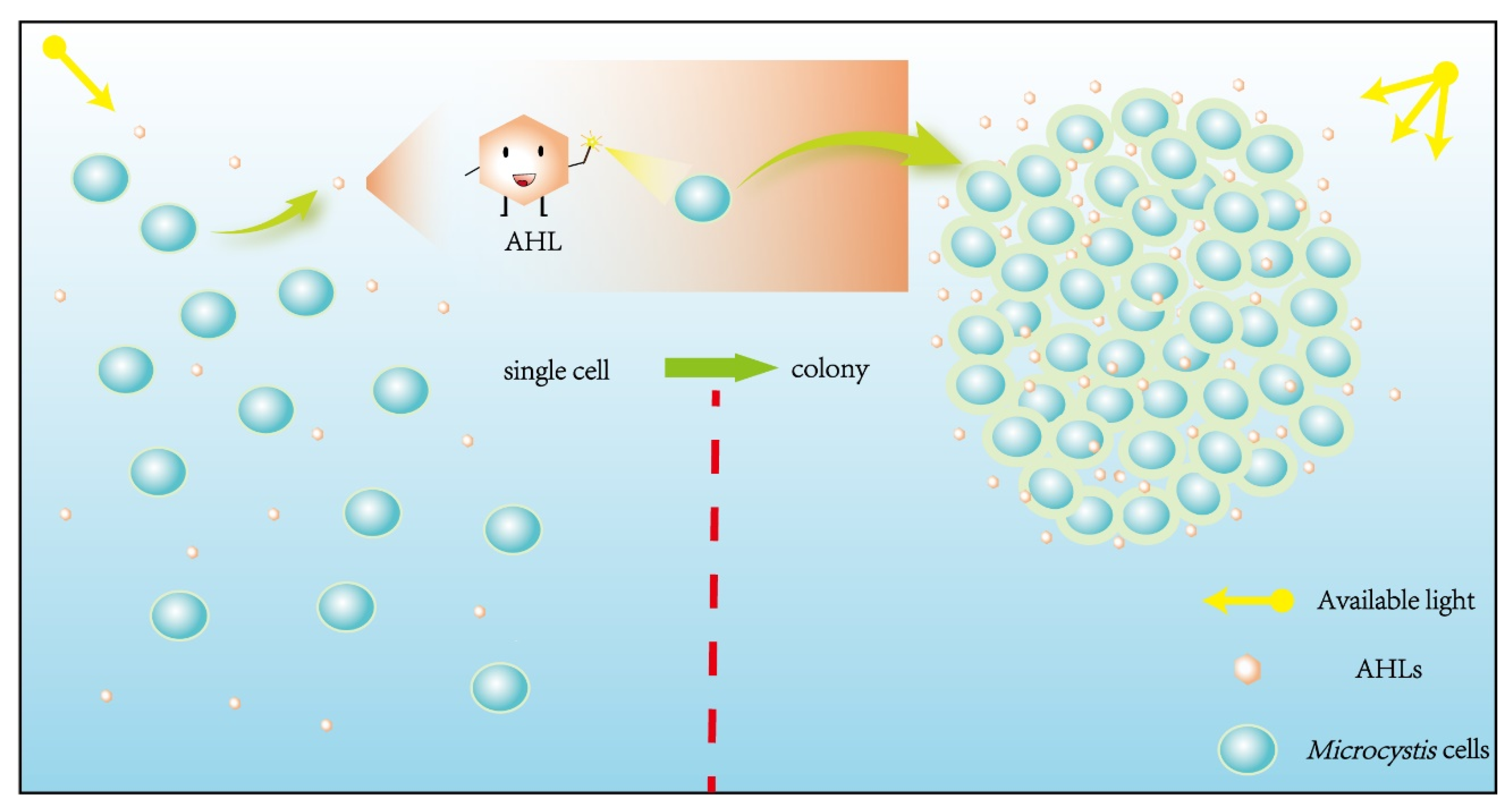
Figure 2.
The effects of AHLs are reflected in four aspects in Microcystis. These aspects include cellular morphology, cellular physiological adaptability, nutrition and energy metabolism activities, colonies formation.
Figure 2.
The effects of AHLs are reflected in four aspects in Microcystis. These aspects include cellular morphology, cellular physiological adaptability, nutrition and energy metabolism activities, colonies formation.
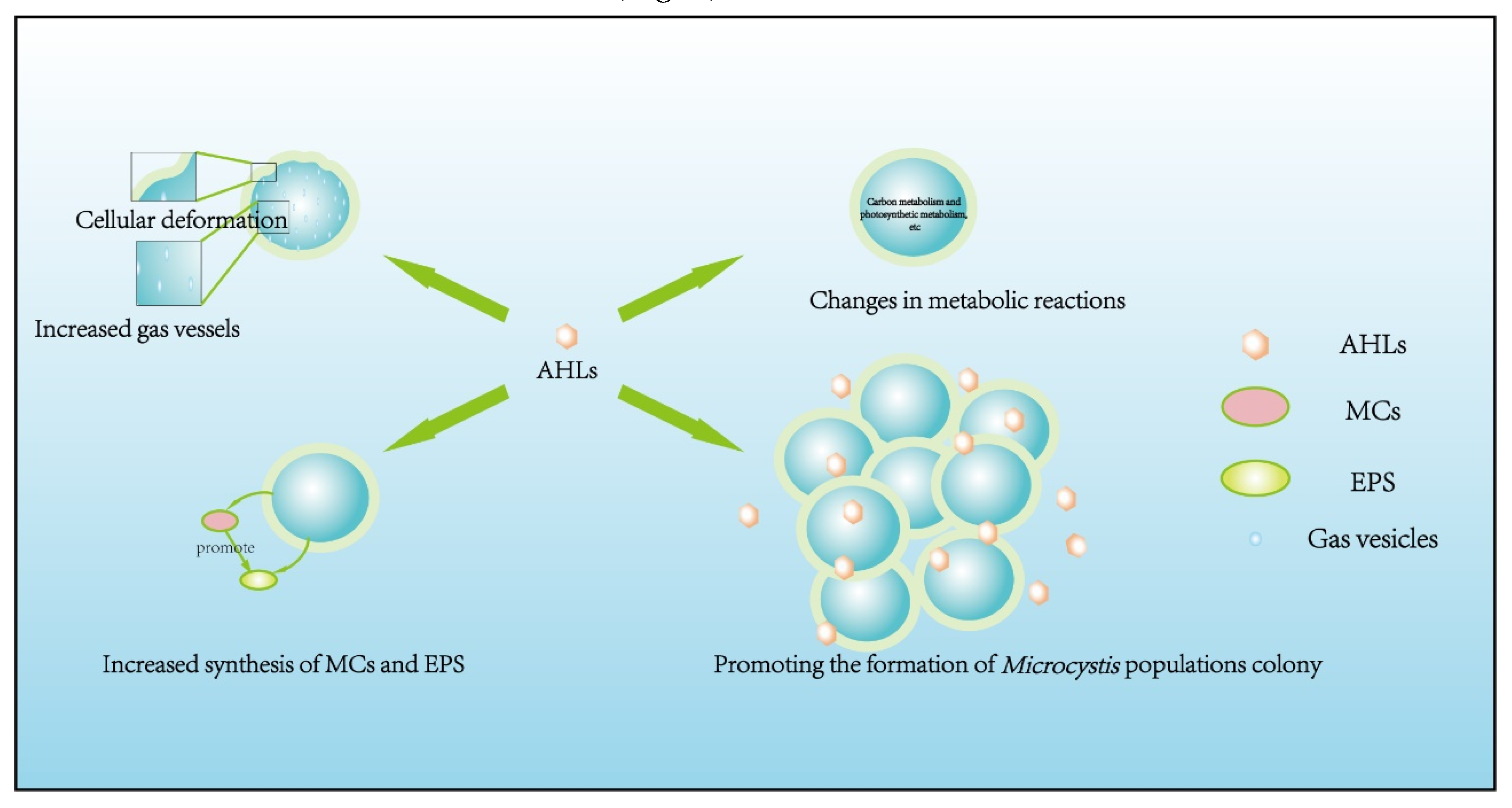
Figure 3.
The LuxI / LuxR-type QS system mediated by AHLs which may be present in Microcystis. LuxI controls the synthesis of AHLs; LuxR is a receptor for AHLs; When the concentration of AHLs reach a specific level, AHLs can bind to receptors and form the complex of LuxR and AHLs to activate the expression of related genes.
Figure 3.
The LuxI / LuxR-type QS system mediated by AHLs which may be present in Microcystis. LuxI controls the synthesis of AHLs; LuxR is a receptor for AHLs; When the concentration of AHLs reach a specific level, AHLs can bind to receptors and form the complex of LuxR and AHLs to activate the expression of related genes.
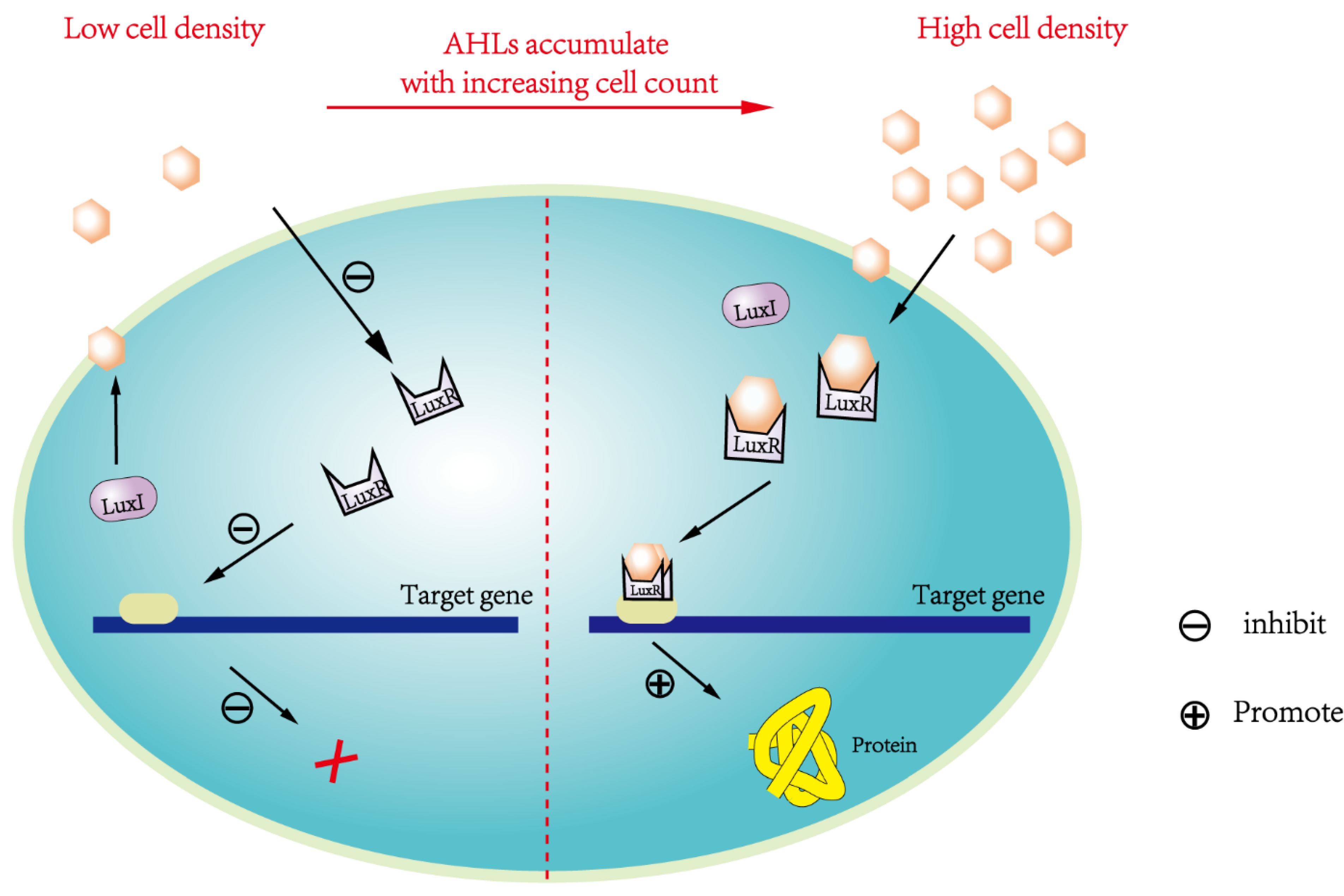
Figure 4.
The QQ process corresponding to the LuxI / LuxR-type QS system in Microcystis. The QQ effect can be achieved by the way of disrupting the signal pathways including the production of AHLs, the accumulation of signaling molecules and the binding of AHLs and receptor.
Figure 4.
The QQ process corresponding to the LuxI / LuxR-type QS system in Microcystis. The QQ effect can be achieved by the way of disrupting the signal pathways including the production of AHLs, the accumulation of signaling molecules and the binding of AHLs and receptor.
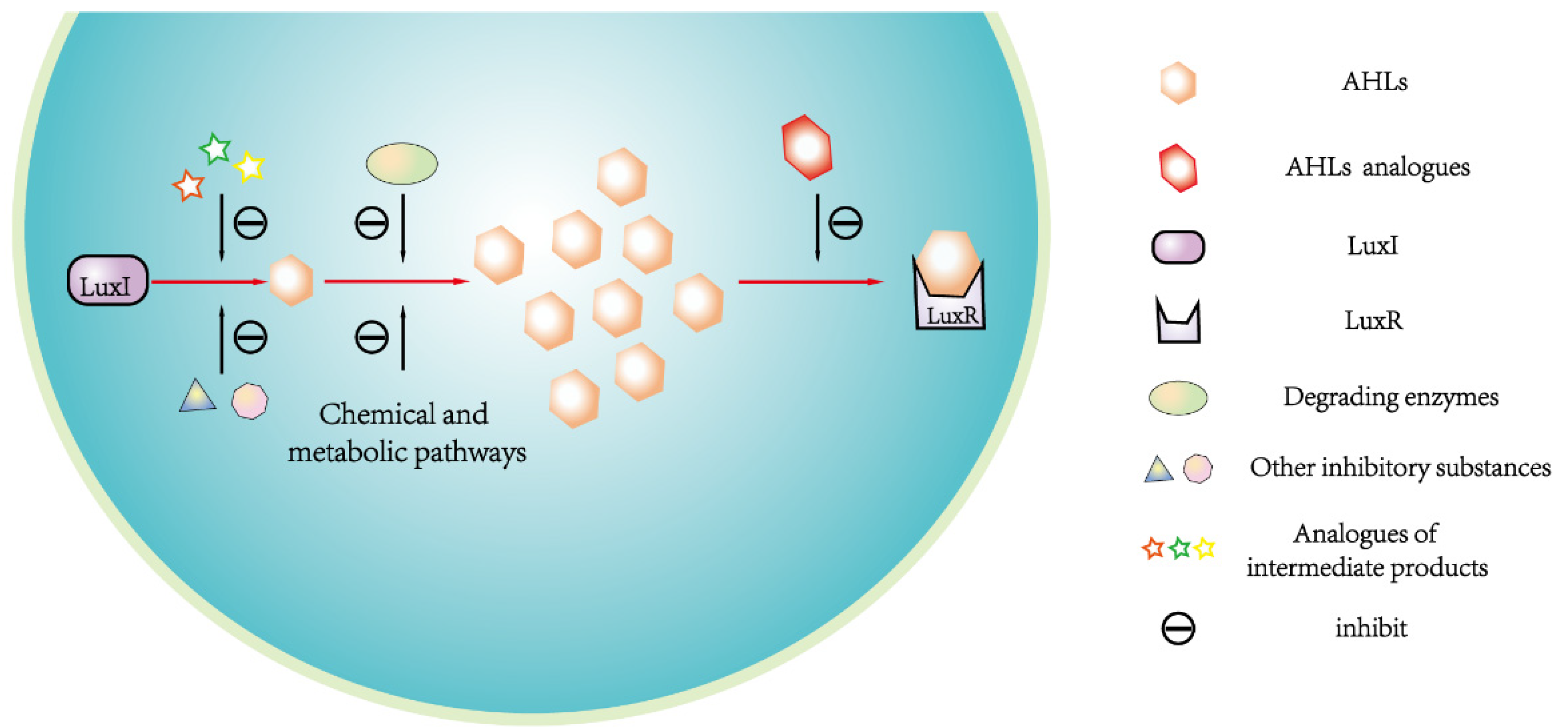

Table 1.
Signaling molecules extracted from Microcystis.
| Article | Microcystis strain | AHLs homologs | Culture medium |
|---|---|---|---|
| [24] | M. aeruginosa FACHB-905 | AHLs(C12H17O5N) | BG-11 |
| [28] | M. aeruginosa PCC7820 | C8 HSLs | Allen-BG11 |
| [40] | M. aeruginosa PCC-7820 | AHLs(C12H17O5N) | BG-11 |
| [41] | M. aeruginosa HB 836 | 3-OXO-C5-HSL | BG-11 |
| C6-HSL | BG-11 | ||
| 3-OXO-C7-HSL | BG-11 | ||
| 3-OH-C4-HSL | BG-11 | ||
| [42] | M. aeruginosa PCC7806 | C3-HSL | BG-11 |
| C4-HSL | BG-11 | ||
| [43] | M. aeruginosa FACHB 905 | AHLs(C13H19O8N) | BG-11 |
Table 2.
The effect of exogenous signaling molecules on Microcystis.
| Article | Microcystis | Concentration | AHLs | Cell growth | Cell morphology | Colony formation | Chlorophyll a | Photosynthesis | Rubisco activity | Superoxide free radical content | Phycocyanin | Intracellular polysaccharide | MCs | EPS |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| [28] | M. aeruginosa HB909 | 0.2 μmol/L | 3-OH-C4-HSL | √ | √ | √ | × | |||||||
| C4-HSL | √ | √ | √ | × | √ | |||||||||
| C8-HSL | × | √ | - | √ | √ | |||||||||
| [40] | M. aeruginosa PCC-7820 | 0.02 μmol/L | AHLs(Unknown type) | √ | √ | |||||||||
| [43] | M. aeruginosa FACHB905 | √ | √ | √ | √ | √ | ||||||||
| [62] | M. aeruginosa FACHB905 | 5 ng/L | C6-HSL | × | √ | |||||||||
| 10 ng/L | √ | √ | ||||||||||||
| 50 ng/L | × | × | ||||||||||||
| 500 ng/L | × | √ | ||||||||||||
| 1000 ng/L | × | √ | ||||||||||||
| [63] | M. aeruginosa FACHB905 | 5 μmol/L | C4H7NO2·HBr | - | ||||||||||
| 10 μmol/L | × | |||||||||||||
| 20 μmol/L | × | |||||||||||||
| 40 μmol/L | × | |||||||||||||
| 50 μmol/L | × | × | √ | × | ||||||||||
| 60 μmol/L | × | |||||||||||||
| 80 μmol/L | × | |||||||||||||
| 100 μmol/L | × | |||||||||||||
| [64] | M. aeruginosa | 0.004 μmol/L | 3-oxo-C10 HSL | × | - | |||||||||
| C6-HSL | √ | √ | ||||||||||||
| C10-HSL | ||||||||||||||
| C7-HSL | √ | √ | ||||||||||||
| C12-HSL | √ | √ | ||||||||||||
| C8-HSL | × | √ | ||||||||||||
| 3-oxo-C8-HSL | √ | √ | ||||||||||||
| C4-HSL | × | √ | ||||||||||||
| [69] | M. aeruginosa FACHB905 | 10 μmol/L | C4H7NO2·HBr | × | √ | × | √ | |||||||
| 20 μmol/L | × | √ | × | √ | ||||||||||
| 40 μmol/L | × | √ | × | √ | ||||||||||
| 60 μmol/L | × | √ | × | √ | ||||||||||
| 80 μmol/L | × | √ | × | √ | ||||||||||
| [70] | M. aeruginosa FACHB905 | 50 μmol/L | C4H7NO2·HBr | √ |
√:Positive effect was observed; ✕: Negative effect was observed; -: no valid effect was observed.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



