
Submitted:
13 June 2024
Posted:
13 June 2024
You are already at the latest version
Abstract
Nowadays, there has been a significant rise in the consumption of kefir, a functional beverage touted for its perceived health benefits. To offer a high-quality beverage to consumers, it is imper-ative to scrutinize and fine-tune the fermentation process. This study seeks to investigate the im-pact of fermentation time and the number of subcultures on the physicochemical, microbiological, and volatile composition, as well as the visual appearance of kefir beverages obtained from four consecutive 24- or 48-h batch subcultures. All fermented beverages exhibited low lactose, ethanol and acids levels, with counts of viable probiotic lactic acid bacteria and yeast exceeding 106 colony forming units/mL. The four kefir beverages from the 48-h batch subcultures notably showed the lowest total concentrations of volatile compounds likely due to overfermentation and overacidifi-cation of the beverages. This caused separation of the whey and curd, along with the formation of large gas bubbles, negatively affecting the visual appearance of the products. These findings em-phasize the importance of fine-tuning the fermentation process to ensure the production of high-quality kefir beverages that align with consumer preferences. The four beverages from the 24-h batch subcultures exhibited high microbiological and physicochemical stability during stor-age at 4 °C for 28 days.

Keywords:
batch fermentation
; kefir grains
; probiotic culture
; whole milk
; volatile compounds
; visual ap-pearance
1. Introduction
Kefir, a fermented beverage primarily produced in the Caucasian mountains by fermentation of sheep's milk with kefir grains, is becoming increasingly popular in European countries, Japan, and the United States [1]. Currently, other dairy substrates (e.g. whole, semi-skimmed or skimmed milk cow's milk, goat, sheep, buffalo, camel, and donkey milks) have been used for kefir production [2,3].
The fermented beverage has a high nutritional value since it contains mainly proteins, amino acids, kefiran (a bioactive exopolysaccharide composed of repeated glucose and galactose units), vitamins, minerals, bacteriocins and low levels of ethanol, lactic acid (responsible of its acidic taste), and fat [4].
Additionally, this functional beverage is widely known as an excellent source of probiotics with a high content in viable lactic acid bacteria (LAB) and yeasts, both originate from the kefir grains, that is composed of, in dry mass basis, kefiran (57 %), proteins (33 %), fat (4 %), and ash (6 %) [4]. The complex microbiota present in kefir grains, which is responsible for the aroma and flavour profile of kefir, includes genera such as Lactobacillus, Lactococcus, Streptococcus, Pediococcus, Leuconostoc, Acetobacter, Gluconobacter, Saccharomyces, Candida, Pichia, Kluyveromyces, Zygosaccharomyces, among others [2,3,4]. The microbiota and the compounds (e.g. lactose, proteins, lactic acid, etc.) present in the fermented beverage can produce beneficial (nutritional and therapeutic) effects for the health of humans and animals [2,3,4,5].
Thus, consumption of kefir has shown a considerable potential to i) positively modulate the microbiome, immune and digestive systems, and cholesterol metabolism, ii) alleviate or delay the development of different types of cancer, iii) reduce the high blood pressure by inhibiting the activity of the angiotensin-converting enzyme, iv) heal wounds faster and reduce wound size, v) reduce the incidence of allergic disease and asthma, vi) improve humoral immune response, vii) be consumed by people with lactose intolerance due to its lower lactose content compared with milk [3,5]. In addition to these health benefits, kefir is considered a food with greater nutritional and dietary value than milk, since the slow digestion of proteins during fermentation makes kefir more easily digestible for humans [6]. Furthermore, the presence of lactic acid in fermented milk can improve calcium absorption, as the acidic pH of the beverage ionizes calcium, facilitating its intestinal absorption [7].
Previous studies have shown that the chemical, microbiological, and volatile composition of kefir can be affected by the origin of kefir grains [8], as well as the type and composition of the fermentation substrate and culture conditions [9]. Therefore, in this study, the chemical, microbiological, and volatile composition, as well as the visual appearance of beverages obtained from four consecutive batch subcultures of 24 or 48 h, were statistically compared to determine differences and similarities between them. To the best of our knowledge, this is the first study to address the effect of incubation time and the number of subcultures on milk kefir production. The chemical and microbiological stabilities of the kefir beverages obtained at the optimal fermentation time were studied during their refrigerated storage at 4 °C for 28 days.
2. Materials and Methods
2.1. Kefir Grains Preparation
The milk kefir grains, submerged in milk, were obtained from Kefiralia (a commercial brand of the company Burumart Commerce S.L., Arrasate, Guipúzcoa, Spain) in a closed plastic zipper bag. The grains were kept at 4 °C in the fridge until they were subsequently activated.
To activate them, the kefir grains were separated from the milk by filtration using a sterile plastic strainer in a biosafety cabinet. Subsequently, the grains were washed with mineral water (Cabreiroá, Ourense, España) and transferred into a clean and sterilized 3.1-L glass jar. Then, 1 L of fresh ultra-high temperature (UHT) whole milk (Central Lechera Asturiana, Asturias, Spain) were poured over the grains. The whole milk was chosen to produce a thick and creamy kefir. The jar mouth was covered with a sterile and breathable cloth secured by a rubber band and subsequently, the jar was then placed at room temperature during 24 h, avoiding exposure to direct sunlight during incubation. This activation protocol was repeated for six times before utilizing the kefir grains as the inoculum in the production of kefir (beverage) in the four consecutive 24- or 48-h batch cultures [10].
2.2. Repeated Batch Fermentations of 24 or 48 h
The UHT whole milk (Central Lechera Asturiana, Asturias, Spain), purchased from a local market in Ourense (Spain), was utilized as the fermentation substrate in this study. According to the producer, the mean composition of the UHT whole milk was 46.0 g sugar/L, 32.0 g proteins/L, 36.0 g fats/L, 24.0 g saturated fats/L, 1.0 g salt/L, 1.2 g calcium/L, with an initial pH of 6.70.
Each batch involved four transfers of the kefir grains into fresh UHT whole milk every 24 h (Fermentation I) or 48 h (Fermentation II), using clean and sterile 3.1-L glass jars, each containing 1 L of UHT whole milk. Repeated batch fermentations of 24 h (I-24 h, I-48 h, I-72 h, and I-96 h) or 48 h (II-48 h, II-96 h, II-144 h, and II-192 h) were conducted in triplicate under static conditions.
Before starting the first subculture of each fermentation, 30.6 ± 0.94 g (drained wet weight) of activated kefir grains were rinsed with mineral water (Cabreiroá, Ourense, España), air-dried for 10 min in a tissue paper under aseptic conditions in a biosafety cabinet, and used to inoculate the UHT whole milk.
Upon completion of the first subculture, the three glass jars containing the fermented milk were used to perform analytical, microbiological, and volatile compounds determinations. First, the kefir grains were separated from the fermented milk by filtration using a sterile plastic strainer. Subsequently, the grains were washed with mineral water (Cabreiroá, Ourense, España), air-dried for 10 min on tissue paper under aseptic conditions in a biosafety cabinet and weighed to estimate the drained wet weight [9,10]. These kefir grains were then utilized to inoculate another 1 L of fresh UHT whole milk to perform the second subculture of 24 or 48 h in triplicate [10].
The fermented milks from the three glass jars were analyzed independently. Each fermented beverage (1 L) was divided into four aliquots (A, B, C, and D) of 30 mL (A), 210 mL (B), 210 mL (C), and 400 mL (D).
Aliquot A was further divided into two equivolume subaliquots, each of which was used to count viable lactic acid bacteria (LAB), acetic acid bacteria (AAB), Enterobacteriaceae, Pseudomonas, and yeasts. Aliquot B was centrifuged (5000 rpm/5 min at 4 °C), and the supernatant was divided into two equivolume subaliquots. Each subaliquot was utilized to measure the culture pH and the concentrations of sugars, organic acids, alcohols, and volatile compounds present in the fermented beverages.
Aliquot C was adjusted to pH 3.5, heated for 3 min in a boiling water bath, centrifuged (5000 rpm/5 min at 4 °C), and then the supernatant was divided into two equivolume subaliquots. Each supernatant sample was used to quantify the antibacterial activity produced by the microbiota of the milk kefir grains [10].
Aliquot D from the four subcultures of the 24-h fermentation was stored in the refrigerator at 4 °C for 28 days. Samples (50 mL) were collected on days 7, 14, 21 and 28 to analyze, in duplicate, the evolution of beverage pH, viable cell counts (LAB, AAB, and yeasts), lactose, organic acids, and alcohols during storage period.
The first, second, third, and fourth subcultures in the 24- and 48-h fermentations were conducted and analyzed under the same conditions.
2.3. Microbiological and Chemical Characterization of the Fermented Beverages
Enumeration of bacteria (LAB, AAB, Enterobacteriaceae, and Pseudomonas) and yeasts was conducted by plating serial decimal dilutions of appropriate volumes from UHT whole milk and aliquot A on the corresponding culture medium. The different microbial groups were counted using specific growth media: MRS agar containing 0.1 g amphotericin B/L for LAB, Carr agar for AAB, double-layered violet red bile glucose agar for Enterobacteriaceae, Pseudomonas agar base containing 10 g cetrimide fucidin/L for Pseudomonas, and yeast extract-glucose agar containing 0.1 g chloramphenicol/L for yeasts [10,11].
Analytical determinations of lactose (L), lactic acid (LA), acetic acid (AA), ethanol (EtOH) and glycerol (GOH) in UHT whole milk and fermented milks were performed using HPLC [10].
Bacterial and yeast counts, expressed as log CFU (colony-forming units) per mL, along with culture pH values, and concentrations (in g/L) of lactose and fermentation products, were reported as means ± standard deviations from three experiments, each with two analytical replications.
2.4. Volatile Characterization
The separation, identification, and quantification of volatile organic compounds (VOCs) present in the UHT whole milk and fermented samples (aliquot B) were conducted using a gas chromatograph GC 7820 A (Agilent Technologies, Santa Clara, CA, USA) coupled to a mass spectrometry detector Agilent MSD series 5975. The GC-MS system was equipped with a capillary column ZB (Phenomenex; 60 m × 0.25 mm × 0.25 μm film thickness).
For the quantification of VOCs in milk fermented samples, the headspace solid-phase microextraction gas chromatography mass spectrometry (HS-SPME GC-MS) method was used [12]. Thus, 20 μL of an internal standard solution of 3-octanol (48 mg/L in absolute ethanol) and 5 mL of sample (UHT whole milk or fermented beverages) were added to a 20 mL vial containing a 1.0 g of NaCl (to enhance the activity and concentration of VOCs in the headspace) along with a magnetic stir bar. The vial was sealed with a screw cap and silicone septum, and then immersed in a water bath at 60 °C for 3 min, at 500 rpm.
For the microextraction of VOCs present in the unfermented and fermented milk samples, a 2 cm, 50/30 µm, DVB/Carboxen/PDMS Stableflex fiber (Agilent Technologies Ltd, Ireland) was used. This type of fiber was used considering its suitability for the extraction of VOCs in dairy foods [12]. The fiber was inserted into the vial and remained in the headspace for 60 min at 60 °C and 500 rpm. The elution of the volatile compounds retained in the fiber was performed by inserting it into the injection port of the chromatograph in splitless mode for 5 min at 250 °C.
For the separation of VOCs, the column temperature was held at 30 °C for 5 min, then increased at a rate of 2 °C/min up to 100 °C, and finally a ramp of 3 °C/min was set up to 150 °C and held constant for 10 min. The flow rate of the column was maintained constant at 1.2 mL/min, using hydrogen as the carrier gas. Mass spectra were scanned at 70 eV in a mass range of m/z 10 to 1000.
All VOCs were identified by comparing their mass spectra with those existing in the Wiley spectral library collection. When the pure volatile organic compound was available, its identification was confirmed by comparing the corresponding retention times [10].
The quantification of VOCs was performed using the internal standard quantification method, utilizing 3-octanol. The concentration of VOCs in the samples was determined based on the concentration of 3-octanol and the ratio between the area of the volatile compound and that of the internal standard.
2.5. Odor Activity Value (OAV)
The OAV, which is a measure of the perception intensity of a specific compound in a food matrix, was calculated as follows [10]:
Where [VOC] is the concentration of the volatile organic compound (mg/L) and OPT is its corresponding odor perception threshold (mg/L).
Compounds with an OAV ≥ 1.0 will make a high contribution to the aroma of beverages, while those with OAV < 1.0 may have either a positive or a negative impact on the aroma through synergistic or antagonistic effects, respectively, with the other volatile compounds present in the beverages [10].
In this study, the OAVs were calculated for those compounds for which information is available in the literature regarding their perception thresholds and aromatic descriptors.
2.6. Aromatic Profile of the Fermented Milks
To obtain the aromatic profile of the beverages, Odor Activity Values (OAVs) were grouped based on volatile compounds exhibiting the same olfactory sensation, as described in their odor profile description obtained from various references. Thus, the volatile compounds were classified into seven aromatic series: microbiological, fruity, floral, herbaceous, solvent, fatty, and sweet [13,14,15].
The olfactory sensations of the UHT whole milk and fermented beverages were compared based on these seven aromatic series using spider maps to detect differences or similarities between them.
2.7. Antibacterial Activity Assay
The total antibacterial activity (TAA) contained in the different fermented beverages was measured using the methodology described by Costas et al. [16], and was quantified as activity units (AU) per milliliter cell-free supernatant against Carnobacterium piscicola CECT 4020 (indicator strain).
2.8. Statistical Analyses
The data regarding beverages characterization, including the culture pH, concentrations (g/L) of lactose, organic acids, alcohols, and volatile compounds (µg/L), bacterial and yeast counts (as log CFU/mL), and antibacterial activity (AU/mL), were obtained from triplicate samples and duplicate analytical determinations after each subculture of 24 or 48 h. Therefore, six observations (3 samples × 2 analytical replicates) for each corresponding variable were obtained in each subculture. The data regarding the drained kefir grains weight were obtained from triplicate samples and one analytical determination after each subculture of 24 or 48 h.
Subsequently, the corresponding mean levels of the variables in each subculture of fermentation I and II were statistically compared using the paired-samples Student's t-test at a significance level of 5% utilizing the software package IBM® SPSS® Statistics for Windows (version 21.0, IBM SPSS Inc., Armonk, NY, 2012) [10]. The comparison between the mean concentration of a volatile compound and its corresponding olfactory perception threshold was performed using the one-sample Student's t-test (significance level of 5%) of the same software package.
The relationships between chemical and volatile compositions of the UHT whole milk and the different kefir beverages were statistically analyzed by applying principal component analysis (PCA) using the software package IBM® SPSS® Statistics for Windows (version 21.0, IBM SPSS Inc., Armonk, NY, 2012). Before conducting the PCA, the suitability of the data for this analysis was confirmed using Bartlett’s sphericity test.
3. Results and Discussion
3.1. Static Fermentation of Whole Milk in Consecutive 24-h Batch Subcultures
The chemical composition of different kefir beverages obtained through fermentation of UHT whole milk with kefir grains in consecutive 24-h batch subcultures is depicted in Figure 1. The results showed that the initial pH in each subculture (6.69 ± 0.01) decreased to 5.17 ± 0.02, 4.99 ± 0.09, 4.70 ± 0.09, and 4.70 ± 0.06 at the end of the first (0 to 24 h), second (24 to 48 h), third (48 to 72 h), and fourth (72 to 96 h) subcultures, respectively. Consequently, the resulting pH drops (ΔpH) were 1.52 ± 0.01, 1.70 ± 0.08, 1.99 ± 0.08, and 1.99 ± 0.07 in the first, second, third, and fourth subcultures, respectively. This indicates an increase in the acidification capacity of the microbial population of the kefir grains with the progression of subcultures.
The consumption of nutrients such as lactose (8.47 ± 1.76, 9.81 ± 2.40, 14.87 ± 2.88, and 9.02 ± 2.10 g/L), total nitrogen (0.96 ± 0.09, 1.11 ± 0.08, 1.68 ± 0.14, and 1.02 ± 0.22 g/L), and proteins (5.99 ± 1.02, 6.94 ± 1.77, 10.51 ± 0.98, and 6.38 ± 1.66 g/L) exhibited a slight increase from the first to the third subculture, but decreased in the fourth one.
In the different subcultures, higher LA concentrations (5.59 ± 0.19, 7.11 ± 0.21, 7.38 ± 0.19, and 4.49 ± 0.11 g/L) and EtOH (3.34 ± 0.11, 3.90 ± 0.15, 3.72 ± 0.12, and 2.55 ± 0.11 g/L) were synthesized compared to the productions of AA (0.78 ± 0.10, 0.80 ± 0.08, 1.97 ± 0.21, and 1.28 ± 0.09 g/L) and GOH (1.38 ± 0.12, 0.35 ± 0.10, 0.37 ± 0.01, and 0.24 ± 0.02 g/L). Lactic acid is likely synthesized by LAB [16,17,18] or yeasts [19], while ethanol and glycerol are predominantly synthesized by yeasts [19,20,21]. Conversely, acetic acid is mainly produced by AAB [22].
The TAA titers produced in milk (Figure 1) increased exponentially from the first to the third subculture (5.31 ± 0.25, 5.85 ± 0.41, and 8.19 ± 0.51 AU/mL) and exhibited a slight decrease (6.97 ± 0.40 AU/mL) in the fourth passage. TAA is the result of the sum of the antibacterial activity of the metabolic products synthesized by the microorganisms of the milk kefir grains (bacteriocins, lactic acid, acetic acid, ethanol and glycerol) [10]. Thus, the increase in the TAA values from the first to the third subculture could be related to the exponential increase in acetic acid concentration and the logistic increase in the production of lactic acid and ethanol (Figure 1). In the latter case, it could be noted that its concentration in the second and third subculture were not statistical different (P > 0.05).
In contrast, the increase in TAA could not be related to the production of glycerol since the concentration of this metabolite showed an exponential decrease during the whole fermentation period (Figure 1).
Considering that milk kefir grains contain bacteriocin-producing LAB strains (e.g. Lactococcus and Lactobacillus species) [10,18,23,24], it could be logical to suppose that the increase in the TAA levels could be also related to the production of bacteriocins by these strains.
The decline in TAA in the four subculture (Figure 1) could be linked to a reduction in the metabolic activity of the microorganisms present in the kefir grains [10]. This is evidenced by a decrease in the consumption of nutrients (L, TN, and Pr) [18], as well as a decrease in the production of lactic acid, acetic acid, ethanol, and glycerol, and possibly in bacteriocin synthesis.
The drained kefir grains weight (KGW) increased (although not significantly, P > 0.05) from 12.24 ± 0.44 to 12.44 ± 0.90, 12.81 ± 0.91, 13.24 ± 0.83, and 13.50 ± 1.04 g in the first, second, third, and fourth subculture (Figure 1). The growth of the kefir grains is commonly associated to an increase in the microbial population (bacteria and yeasts) anchored to the grains and the production of kefiran, an exopolysaccharide produced by Lactobacillus kefiranofaciens, Lb. kefirgranum, Lb. parakefir, Lb. kefir, and Lb. delbrueckii subsp. bulgaricus [10,25].
Also noteworthy are the high final counts of LAB (8.66 ± 0.06, 8.96 ± 0.08, 8.84 ± 0.02, and 9.14 ± 0.08 log CFU/mL), AAB (8.89 ± 0.02, 8.64 ± 0.05, 8.95 ± 0.03, and 9.13 ± 0.06 log CFU/mL), and yeasts (6.30 ± 0.09, 6.21 ± 0.03, 7.30 ± 0.05, and 6.29 ± 0.04 log CFU/mL) in each subculture. These counts significantly exceeded the threshold level of 6.0 log CFU of probiotic cells per mL or gram needed to observe beneficial effects on the host [26,27]. The counts of Enterobacteriaceae and Pseudomonas were below the minimum detection limit (counts were performed only for plates containing 30–300 colonies), indicating that the four beverages have good hygienic conditions. This is an important consideration in the production of fermented beverages, as contamination with undesirable bacteria could affect consumer health.
The statistically higher (P < 0.05) LAB and AAB counts, compared to those of yeasts, could be related to the initial pH of the milk (6.69 ± 0.01), which was probably more favorable for the growth of the two bacterial populations [28,29] than for the yeasts population [30,31].
Considering that the UHT whole milk is an ultra-pasteurized product, packaged in sterilized container, the initial concentration of microorganisms is considerably low in this substrate. Therefore, the final concentrations of LAB, AAB, and yeasts in the fermented beverages at the end of each subculture solely comprise the microbiota of the milk kefir grains. This indicates that a probiotic-enriched beverage can be obtained by fermenting UHT whole milk with milk kefir grains.
3.2. Static Fermentation of Whole Milk in Consecutive 48-h Batch Subcultures
To determine the impact of fermentation time on the chemical, microbiological and volatile composition of milk kefir beverages, a series of consecutive 48 h-batch fermentations of UHT whole milk with milk kefir grains was conducted under static conditions (fermentation II). This approach allows for a discussion of the similarities and differences between fermentations I and II.
In fermentation II, the final pH values in the first (4.38 ± 0.03), second (4.02 ± 0.07), and third (4.12 ± 0.05) subcultures (Figure 2) were statistically different (P < 0.05). However, the final pH in the fourth subculture (4.09 ± 0.03) did not show a statistically difference (P > 0.05) compared to those of the second and third subcultures.
The consumptions of lactose (21.13 ± 1.75, 23.92 ± 2.165, 17.77 ± 2.62, and 14.59 ± 1.49 g/L) and proteins (13.91 ± 1.89, 15.75 ± 1.35, 11.69 ± 0.83, and 9.60 ± 1.90 g/L) were only not statistically different (P > 0.05) in the third and fourth 48-h subcultures (Figure 2). In contrast, the consumption of total nitrogen in the four subcultures (2.23 ± 0.13, 2.52 ± 0.08, 1.87 ± 0.09, and 1.54 ± 0.22 g/L, respectively) showed statistically differences (P < 0.05).
Regarding the synthesis of fermentation products, it can be observed that LA production exhibited a significant (P < 0.05) small linear increase (9.46 ± 0.16, 10.58 ± 0.18, 10.56 ± 0.21, and 11.50 ± 0.14 g/L) at an average rate of 0.61 g/L/subculture. Acetic acid production increased from the first (1.08 ± 0.16 g/L) to the second subculture (3.80 ± 0.10 g/L), but then decreased in the third (2.74 ± 0.16 g/L) and fourth (2.01 ± 0.11 g/L) subculture. In contrast, the concentrations of EtOH (5.17 ± 0.09, 2.84 ± 0.11, 1.22 ± 0.16, and 0.80 ± 0.19 g/L) and GOH (0.44 ± 0.08, 0.27 ± 0.01, 0.08 ± 0.01, and 0.05 ± 0.00 g/L) exhibited decreasing profiles (Figure 2).
Providing an accurate discussion on the levels of the metabolic products (LA, AA, EtOH and GOH) synthesized in the different subcultures is very difficult. This is probably due to the existence of a complex microbial population in the milk kefir grains, comprising LAB, AAB, and yeasts [2,3,4], which establish complex metabolic relationships among themselves. Thus, it has been reported that certain LAB, such as Lactobacillus buchneri, Lb. parabuchneri [32], Lb. plantarum [33], and L. lactis [34], can use lactic acid as a carbon source. Furthermore, some non-lactose-consuming yeasts, such as strains of S. cerevisiae [35,36], S. exiguous, S. carlsbergensis, and Debaryomyces hansenii, can assimilate lactic acid [36], Candida guilliermondii can metabolize acetic acid [37], and S. cerevisiae can use both lactic and acetic acid [38] as carbon sources.
Regarding the EtOH production, it has been recognized that species of AAB such as Acetobacter and Gluconobacter can oxidize this metabolite to produce acetic acid [39]. Additionally, assimilation of GOH by some strains of LAB, such as Lb. collinoides, Lb. diolivorans, Lb. reuteri [40], or Lb. buchneri [41], has also been reported.
Therefore, it seems logical to assume that the final concentration of the different metabolic products in the different subcultures is the result of the difference between their production and consumption by the corresponding producing and consuming strains present in the milk kefir grains, respectively.
TAA production increased logistically (P < 0.05) during the fermentation (Figure 2), contrasting with the exponentially decreasing profiles exhibited by ethanol and glycerol production (from the first to the fourth subculture), and acetic acid synthesis (from the second to the fourth subculture). Thus, although lactic acid production showed a slightly linear increase, the rise in TAA levels appeared to be associated with the increase in bacteriocin production by the LAB strains present in the grains [10,18,23,24].
The drained KGW exhibited again a no significant increase (P > 0.05) throughout the fermentation process (Figure 2), consistent with observations from the 24-h subcultures (Figure 1).
The viability of microbial populations increased to 7.40 ± 0.09, 9.03 ± 0.03, 8.48 ± 0.05, and 9.12 ± 0.06 log CFU/mL (in the case of LAB), 8.48 ± 0.06, 9.32 ± 0.09, 8.57 ± 0.06, and 8.97 ± 0.07 log CFU/mL (in the case of AAB), and 7.42 ± 0.05, 6.69 ± 0.06, 6.18 ± 0.07, and 5.60 ± 0.04 log CFU/mL (in the case of yeasts) in the first, second, third, and fourth subcultures, respectively. As observed, only the yeasts count corresponding to the fourth subculture did not surpass the threshold level of 6.0 log CFU of probiotic cells per mL needed to observed beneficial effects on the host [26,27].
As occurred in the previous 24-h subcultures, the counts of Enterobacteriaceae and Pseudomonas were considerably lower than the minimum detection limit of 30 colonies indicating once again that these beverages are in good hygienic condition.
Comparing the fermentations I and II (Figure 1 and Figure 2), it is evident that the 48-h batch subcultures provided higher (P < 0.05) ΔpH values (2.31 ± 0.04, 2.67 ± 0.08, 2.57 ± 0.06, and 2.60 ± 0.04) and showed greater (P < 0.05) consumption of nutrients (L: 21.13 ± 1.75, 23.92 ± 2.16, 17.77 ± 2.62, and 14.59 ± 1.49 g/L, TN: 2.23 ± 0.13, 2.52 ± 0.08, 1.87 ± 0.09, and 1.54 ± 0.22 g/L, and Pr: 13.91 ± 1.89, 15.75 ± 1.35, 11.69 ± 0.83, and 9.60 ± 1.90 g/L). However, the increase in nutrient consumption did not lead to an increase in product synthesis (LA, AA, EtOH, and GOH) as observed in Figure 2. These results suggest that extending the duration of each subculture from 24 to 48 h, the microbiota of the kefir grains consumed higher amounts of nutrients to support the growth and maintenance of cells rather than for product synthesis.
In terms of microbial content, the four beverages obtained from fermentation I had counts of LAB (except in the first subculture) and AAB (except in the second subculture) that were almost similar to those from fermentation II. Meanwhile, yeast counts at the end of the subcultures from fermentation II decreased as the number of subcultures increased, contrasting with their behavior in the 24-h subcultures.
Moreover, the 48-h kefir beverages exhibited slightly higher acidic, lower alcohol content (except in the first subculture), and increased antibacterial activity (P < 0.05) compared to the 24-h kefir beverages (Figure 1 and Figure 2). However, the 24-h kefir beverages, particularly those obtained from the first two subcultures, showed statistically significant higher final lactose concentrations (P < 0.05) compared to those from fermentation II.
3.3. Volatile Composition of Beverages Obtained through Static Discontinuous Fermentations I and II
As shown in Table 1, fermenting UHT whole milk (containing 20 volatile compounds) with kefir grains in 24-h subcultures resulted in a considerable increase in both the number (more than doubling) and the total concentrations (although not always significantly) of volatile compounds in the four fermented beverages.
In the 48-h subcultures, the number of volatile compounds found in the four fermented beverages was also higher than in unfermented milk. However, surprisingly, the total concentration of these compounds in the four 48-h beverages decreased (P < 0.05) compared to that in unfermented milk (Table 1).
The volatile compounds detected in the unfermented milk and fermented samples were grouped into seven families for study: organic acids, alcohols, aldehydes, ketones, esters, hydrocarbons, and a miscellaneous group (other compounds). These compounds exhibited different profiles, with some detected only in the unfermented milk, some only in all fermented beverages, some specifically in the 24-h subcultures, others in the 48-h subcultures, and some in both unfermented and fermented milk.
The olfactory perception thresholds, odor descriptors and aroma series for the volatile compounds found in the literature are shown in Table 2.
The presence of different compound families in unfermented whole milk and in kefir beverages obtained in fermentations I and II is described below.
3.3.1. Organic Acids
Carboxylic acids in fermented milk, like kefir, generally originate from lipolysis, proteolysis, or lactose fermentation. These compounds are responsible for the acidity and contribute to the flavor and aroma of the kefir beverages [45,69], and act as precursors for a variety of aromatic compounds including alcohols, esters, methyl ketones, and others [45].
Both hexanoic acid and 2-ethyl hexanoic acid were found in all fermented samples, although at levels lower than their corresponding olfactory perception threshold (OPT) values (3000 and 27000 μg/L, respectively). Hexanoic acid was the organic acid detected at the highest concentrations, ranging between 7.63 and 23.45 μg/L in the 24-h subcultures, and between 3.85 and 7.82 μg/L in the subcultures of fermentation II. The highest concentration of 2-ethyl hexanoic acid (2.61 ± 0.63 μg/L) was found in the third subculture of fermentation II (Table 1).
Acetic acid was not detected in the unfermented whole milk or in the fermented beverage from the first 24-h subculture. Considering each fermentation independently, it can be noted that the highest concentrations of this compound were found in the fermented beverages obtained from the third subcultures of fermentations I and II, with values of 2.69 ± 0.83 μg/L and 3.38 ± 0.23 μg/L, respectively (Table 1). However, these levels are considerably lower than the OPT for this acid (99000 μg/L) [42] and those (between 7.89 to 24.65 μg/L) detected by Dan et al. [84,85] in milks fermented with different proportional combinations of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. This suggests that the fermented milk beverages obtained in this study will not have unpleasant notes of vinegar or sourness [43,44].
Concentrations of butanoic acid considerably lower than its OPT (1400 μg/L) [42] were obtained in the fermented samples from the first and third subcultures of fermentation I, as well as in all beverages from fermentation II. This indicates that this compound alone will not influence the aroma of the fermented beverages.
Four organic acids were exclusively detected in the 24-h subcultures: pentanoic, octanoid, ethyl boronic, and 8-methyl-6-nonenoic acid. Pentanoic acid and octanoic acid were only found in fermentation I, although at concentrations lower than the OPT (3000 μg/L) for these compounds [42]. Pentanoic acid was present solely in the third (0.76 μg/L) and fourth (0.87 μg/L) subcultures, while octanoic acid was detected in the fourth subculture (1.69 μg/L). Similarly, ethyl boronic acid (0.59 ± 0.14 μg/L) and 8-methyl-6-nonenoic acid (0.71 μg/L) were only identified in the third subculture of fermentation I.
In contrast, two organic acids were only detected in the repeated batch fermentations of 48 h: 2-methyl-propanoic acid in the last three subcultures at concentrations between 0.64 and 2.02 μg/L (< OPT: 6550.5 μg/L) [42], and trans-2-undecenoic acid (1.54 μg/L) in the second subculture (Table 1).
Heptanoic acid was detected in the second subculture of fermentation I at a mean concentration of 0.92 μg/L (second subculture) and gradually decreased to 0.78 μg/L at 96 h (Table 1). This organic acid was also detected in the first, second and third subcultures of fermentation II at concentrations of 0.20, 0.23, and 0.31 μg/L, respectively, which were lower than those found in the 24-h subcultures. All these concentrations are below its OPT (640 μg/L) [42].
Nonanoic acid was detected in the unfermented substrate (4.89 μg/L < OPT: 8800 μg/L [42]), but its concentration decreased in the second and third subcultures of the repeated batch fermentations of 24 h, and in the second, third, and fourth subcultures of the repeated batch fermentations of 48 h.
Cyclopentane-undecanoic acid was only detected in unfermented milk (7.55 ± 0.19 μg/L), while cis-11-eicosenoic acid was the acid found at the highest concentration in unfermented milk (28.38 ± 1.43 μg/L) and was also detected in all fermented samples. However, its concentration decreased during fermentation (Table 1).
Acetic, hexanoic, octanoic, nonanoic acids was also found in kefir samples obtained by fermentation of full-fat pasteurized milk with three types of kefir grains [52] or in other fermented milks [85,86].
According to the results obtained in this study, acetic, butanoic, pentanoic, hexanoic, 2-ethyl hexanoic, 2-methyl-propanoic, heptanoic, octanoic, and nonanoic acids will not have a direct contribution to the sensory profile of the fermented beverages, although these compounds could indirectly influence it through synergistic effects with other identified volatile compounds. Production of volatile compounds in milk kefir beverages at concentrations lower than their corresponding olfactory perception thresholds were also obtained in commercial kefir beverages samples [87].
3.3.2. Alcohols
The formation of alcohols in milk fermented with kefir grains is mainly due to the metabolism of yeasts [19,20,21,88]. Like ketones, alcohols that are generated by the reduction of the corresponding aldehydes [89] are important contributors to the flavor of dairy products [84].
In this study, the fermentation of milk in repeated 24-h subcultures increased both the number and total concentration of alcohols compared to the unfermented milk (Table 1). Two alcohols were detected in the unfermented milk, whereas 9, 10, 10 and 6 alcohols were found in the fermented samples from the first, second, third, and fourth 24-h subcultures, respectively. The total concentration of alcohols increased in the first two subcultures (82.71 ± 5.84 μg/L and 108.89 ± 16.56 μg/L) but decreased in the last two (89.47 ± 13.96 μg/L and 31.63 ± 8.45 μg/L), although it did not reach the lower concentration (15.13 ± 0.00 μg/L) found in unfermented milk (Table 1). These results are consistent with the statement by Fang et al. [90], who describe kefir as a fermented dairy product with a wide variety of volatile alcoholic compounds.
The increase in the number and concentrations of these compounds during fermentation is the result of increasing biochemical reactions [91]. However, the concentration of some alcohols can also decrease during fermentation due to their reaction with organic acids to form esters [45].
The only two alcohols detected in unfermented milk were 2-phenylethanol and 2-ethyl hexanol, indicating that the other alcohols found in the kefir beverages originated during repeated 24- or 48-h subcultures. In the 24-h subcultures, the concentration of 2-phenylethanol increased from 8.86 ± 0.00 μg/L (unfermented milk) to 12.36 ± 5.54, 23.09 ± 9.51, 22.90 ± 3.73, and 11.41 ± 1.74 μg/L in the first, second, third, and fourth subcultures, respectively. The detection of relatively high concentrations of 2-phenylethanol was also described by Oliveira et al. [51] in whey samples mixed with beet juice and fermented Kluyveromyces lactis. The presence of this alcohol in the milk kefir samples is related to the metabolism of amino acids [52].
In the repeated 48-h fermentation, the concentration of 2-phenylethanol increase to 10.70 ± 4.84 μg/L in the first subculture, but decreased gradually in the second (9.06 ± 0.39 μg/L), third (5.34 ± 1.00 μg/L), and fourth (3.20 ± 0.87 μg/L) passages (Table 1). As observed, the concentrations of this compound in the second, third, and fourth subcultures of fermentation II were significantly lower (P < 0.05) than those detected in the corresponding subcultures of fermentation I. However, the concentrations of 2-phenylethanol in the first subcultures of fermentations I and II were not statistically different (P > 0.05).
2-Ethyl-1-hexanol exhibited the same behavior as 2-phenylethanol in the 24-h subcultures, as it was detected in both unfermented milk (6.27 ± 0.00 μg/L) and in the four fermented beverages (12.00 ± 0.59, 19.23 ± 3.27, 17.59 ± 0.83, and 8.03 ± 7.34 μg/L). In contrast, the concentration of this compound decreased in the beverages from fermentation II, reaching only mean concentrations between 2.43 and 3.37 μg/L (Table 1).
The alcohol detected at the highest concentration during fermentations I and II was 1-pentanol, although in both cases, its concentration decreased gradually from the first to the fourth subculture. However, as observed for 2-phenylethanol and 2-ethyl-1-hexanol, the concentrations of 1-pentanol in the 24-h subcultures were higher than those found in the 48-h subcultures (Table 1). This volatile compound was also identified in dairy beverages fermented with kefir grains [52], and in yogurts produced with cocultures of traditional LAB strains, although at relatively low concentrations [53].
Methyl alcohols can be generated from the reduction of the corresponding methyl ketones, a reaction catalyzed by enzymes with reductase activity [84]. In this study, nine methyl alcohols were detected in the fermented samples from the 24-h subcultures: 2-methyl-1-propanol, 3-methyl-1-pentanol, 2,10-dimethyl-9-undecen-1-ol, 2,4-dimethyl-3-pentanol, 2,7-dimethyl-1-octanol, 2-methyl-5-(1-methylethenyl)-cyclohexanol, 3,6-dimethyl-3-heptanol, 3-methyl-1,5-pentanediol, and 5,9-dimethyl-1-decanol. However, these compounds were not detected in all subcultures. A similar situation occurred with the alcohols 2-butyl-1-octanol, 2-butyl-2,7-octadien-1-ol, 1,3-octanediol, 1,5-hexanediol, 1-heptanol, 5-nonadecen-1-ol, and Z-10-pentadecen-1-ol (Table 1).
2-Heptanol was detected in the milk kefir samples at low mean concentrations (0.64 and 0.80 μg/L in the first and second passages, respectively) in the 24-h subcultures, consistent with the levels obtained in milk fermented with different proportional combinations of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus [84,85].
In fermentation II, the only methyl alcohols found were 2,4-dimethyl-3-pentanol (in the first and third passages) and 4-methyl-2-pentanol (in the fourth subculture) but at relatively low concentrations (Table 1). Additionally, the following alcohols were identified in these subcultures: 1,5-hexanediol and 2-heptanol (both in the first three subcultures), 2-butyl-1-octanol (in the first, third, and fourth subcultures), and 2-butyl-2,7-octadien-1-ol (in all subcultures).
With the increase in the incubation period to 48 h, eight additional alcohols were found. One of them, butane-1,3-diol, was detected in the first subculture. Three alcohols, including (E)-2-tridecen-1-ol, 1-eicosanol, and 2-nonen-1-ol, were found in the second subculture. In the fourth subculture, three alcohols were identified: 1,3,5-pentanetriol, 2-furanmethanol, and 2-hexyl-1-octanol. Additionally, 3-tetradecin-1-ol was detected in the first and second subcultures.
Butane-1,3-diol was also detected in the volatile composition of cheese samples produced with a mixed inoculum of Kluyveromyces lactis and Torulaspora delbrueckii (yeasts present in kefir grains), and matured for 21 days [45]. 2-Furanmethanol was identified in creams from yogurt and milks with different percentages of fat [63]. (E)-2-tridecen-1-ol was found in tomato juice fermented con Lactobacillus plantarum and Lb. casei, two LAB present in the kefir grains [92].
3.3.3. Aldehydes
Aldehydes are also an important group of volatile compounds responsible for the formation of the characteristic aromatic profile of dairy products by Dimitrellou et al. [67]. These researchers observed that the aldehydes developed during milk fermentation with encapsulated strains of Lb. casei, a bacterium present in kefir grains, improved the aroma of the final product. In this regard, the researchers [67] highlighted the productions of 2-nonenal (8.4 μg/kg) and nonanal (16.8 μg/kg), two compounds also detected in the fermented samples of the second and third subcultures of the repeated 24-h fermentation, respectively. However, the production of both aldehydes was no detected in the repeated 48-h fermentation (Table 1).
In this study, no aldehydes were detected in milk (Table 1). However, the total number of aldehydes increased from the first to the third subculture and then decreased in the fourth subculture of fermentations I and II. The concentrations of aldehydes increased from the first to the third subculture (although not always significantly) and exhibited a significant decreased (P < 0.05) in the fourth subculture of fermentation I. In fermentation II, the concentration of aldehydes increased significantly (P < 0.05) from the first to the third subculture, but the slight decrease in the fourth subculture compared to the third was not significant (P > 0.05).
A comparison between the fermentations I and II shows that the total number of aldehydes in the first three subcultures of fermentation I was higher than in the corresponding subcultures of fermentation II. However, in the fourth subculture of fermentations I and II, two and three aldehydes were detected, respectively. Additionally, the concentrations of aldehydes in all subcultures of fermentation I were always significantly higher (P < 0.05) than in fermentation II (Table 1).
In addition to 2-nonenal and nonanal, eight other aldehydes were identified in the fermented samples from fermentation I (Table 1). These include (Z)-14-methyl-8-hexadecenal (24-48 h, 48-72 h, and 72-96 h subcultures), 10-undecenal (0-24 h subculture), 2,6-dimethylbenzaldehyde (0-24 h subculture), 2-ethyl hexanal (24-48 h and 48-72 h subcultures), 2-methyl-undecanal (24-48 h, and 48-72 h subcultures), benzaldehyde (0-24 h, 24-48 h, and 48-72 h subcultures), dodecanal (48-72 h and 72-96 h subcultures), and phenylacetaldehyde (24-48 h and 48-72 h subcultures).
In this study, only two aldehydes were detected at concentrations higher than their corresponding OPT: (E)-2-nonenal and nonanal. The first aldehyde was detected in the second subculture at a mean concentration of 0.84 μg/L (Table 1), above its perception threshold in water (0.40 μg/L) [42], suggesting that this compound would contribute directly to the aroma of the beverage. (E)-2-nonenal, described as fatty and green [59], was also detected (3.93 μg/L) in a fermented milk fermented with strains of Lb. delbrueckii subsp. bulgaricus and S. thermophiles (at a ratio 1:1) [85]. Nonanal was detected in the third subculture at a concentration of 1.61 ± 0.81 μg/L (Table 1). Since this concentration is higher than its corresponding OPT in water (1.10 μg/L) [42], it can be concluded that this compound would contribute directly to the aroma of the beverage imparting green, fresh [47], citrus-like, soapy [51], and fruity notes [67] (Table 2).
In the fermented samples from the 48-h subcultures, only four aldehydes were detected. 2-Ethyl hexanal and phenylacetaldehyde were present in all subcultures, while 2-methyl-undecanal and (Z)-14-methyl-8-hexadecenal were found in two subcultures (Table 1). The concentrations of these four compounds were always lower than those in fermentation I (Table 1).
3.3.4. Ketones
Ketones are well-known for their impact on the aroma of most dairy products due to their low perception thresholds [69]. The formation of these compounds is mainly due to the breakdown of fatty acids, sugars, and amino acids, and not solely due to microbial activity [93].
In this study, three ketones (2,6-dimethyl-4-heptanone, 2-heptanone, and 2-nonanone) were identified in UHT whole milk. In the fermented samples from fermentation I, four, two, two and four ketones were identified in the first, second, third, and fourth subcultures, respectively. During the repeated 48-h fermentation, three volatile compounds were found in the first subculture, while in the subsequent subcultures, only two ketones were detected.
Among the ketones detected, 2,6-dimethyl-4-heptanone was the only compound found in both UHT whole milk and all fermented samples. The mean concentration of this ketone increased from 2.18 μg/L in milk to 4.98, 2.23, 2.52, and 3.19 μg/L in the fermented samples obtained after 24, 48, 72, and 96 h of incubation, respectively (Table 1). However, in the 48-h subcultures, relatively low concentrations (between 0.06 and 0.42 μg/L) of this compound were obtained (Table 1). In any case, the concentrations of 2,6-dimethyl-4-heptanone did not exceed its OPT of 110 μg/L [68] (Table 2).
The mean concentration of 2-heptanone in UHT whole milk was 28.96 μg/L but it decreased during fermentation to mean levels of 5.52, 4.05, and 2.39 μg/L in the first, second, and fourth subcultures from fermentation I. The fermented beverages from the repeated 48-h subcultures had lower concentrations of 2-heptanone, ranging from 0.40 to 2.14 μg/L. However, only the unfermented substrate and the fermented beverage from the first subculture of fermentation I had concentrations of 2-heptanone higher than its OPT determined in milk (5 μg/L) [69], thus directly contributing to the aroma of both samples (Table 2).
2-Heptanone was also detected in milk fermented with encapsulated Lb. casei cells [67], with Lb. delbrueckii subsp. bulgaricus and S. thermophilus [85], and in traditional yogurt [53]. This ketone was found in the volatile composition of commercial UHT whole and skimmed milks [89], and in milks fermented with 1:1, 1:10, 1:50, 1:100, 1:1000, and 1:10000 mixtures of Lb. delbrueckii subsp. bulgaricus and S. thermophilus strains [85]. The highest concentration of 2-heptanone (27.48 μg/L) was found in milk fermented with the 1:10000 mixture of LAB.
Regarding 2-nonanone, its maximum concentration was found in the substrate (UHT whole milk) at a level of 14.26 μg/L, which is higher than its OPT determined in milk (5 μg/L) [69], making it one of the compounds contributing to the sensory profile of milk (Table 2).
2-Heptanone and 2-nonanone were found in yak milk fermented in five eco-regions of the Qinghai-Tibetan plateau [90]. When present at appropriate concentrations, these compounds are considered responsible for the natural and pleasant flavor of milk [93]. These observations indicate that 2-heptanone and 2-nonanone are commonly present in both milk and fermented dairy products [85].
However, fermentation of milk in repeated 24- or 48-h subcultures resulted in a drastic reduction in the concentrations of both 2-heptanone and 2-nonanone (Table 1).
2-Methyl-4-octanone was found in the first (1.06 μg/L), third (1.30 μg/L), and fourth (1.92 μg/L) subcultures, and 8-hydroxy-2-octanone was only detected in the fourth passage (2.06 μg/L) of the repeated 24-h fermentation.
In contrast, 5-methyl-3-heptanone was only detected in the first subculture of the repeated 48-h fermentation (Table 1), at a concentration of 0.16 μg/L, considerably lower than its OPT of 41 μg/L [42]. According to Dan et al. [85], 5-methyl-3-heptanone was found (1.31 μg/L) in milk fermented with a 1:1 mixture of Lb. delbrueckii subsp. bulgaricus and S. thermophilus strains. Surprisingly, this ketone was not detected when the milk was fermented with other combinations of both strains (1:10, 1:50, 1:100, 1:1000, and 1:10000) [85].
3.3.5. Esters
According to Liu et al. [71], esters in dairy products are commonly synthesized via two enzyme-catalyzed reactions: esterification or alcoholysis. The first reaction occurs between alcohols and carboxylic acids, while the second can occur in two ways: between alcohols and acylglycerols, or between alcohols and fatty acyl-CoAs synthesized during the metabolism of fatty acids, amino acids, and/or carbohydrates [71]. Other researchers have indicated that esters are produced during microbial degradation of lactose or amino acid [94], or due to the lipid metabolism of yeasts [95]. These compounds are desirable in beverages due to their high volatility and fruity aromatic characteristics, which can help mask undesirable aromas of other compounds, including acids, aldehydes, and amines [45].
It has been reported that esters, even at low levels, can contribute to the formation of fruity and floral aromas and flavors in fermented and non-fermented foods, including fermented dairy products, because most of these compounds have a low taste threshold [67,85,95].
In the present study, esters comprised the majority of the volatile compounds identified in the unfermented and fermented milk (Table 1), consistent with previous findings in a beverage obtained by fermenting whey with Kluyveromyces lactis, a yeast present in milk kefir grains and known for its efficiency in producing these compounds [51].
Five esters (2-phenylethyl acetate, allyl pentadecyl oxalate, ethyl octanoate, ethyl decanoate, and methyl hippurate) were identified in unfermented milk (Table 1). Fermentation of this substrate with kefir grains led to an increase in the number of esters. Thus, in the fermented samples from the 24-h subcultures, the number of esters increased to 14 (24 h), 11 (48 h), 13 (72 h), and 15 (96 h). Regarding the concentrations of these volatile compounds, it can be noted that repeated 24-h fermentation improved the concentration of esters compared to the unfermented substrate (P < 0.05), suggesting an increase in fruity aromatic notes in the four fermented beverages. A similar increase in the number and concentrations of esters with fermentation was observed when soy whey was fermented with water kefir grains for 48 h [96].
In the repeated 48-h subcultures, the number of esters were also higher than in whole milk. Thus, 13, 16, 11, and 9 volatile compounds were found in the first, second, third, and fourth subcultures, respectively (Table 1).
The fermentation of milk in repeated 48-h subcultures did not always result in an increase in the total concentration of these compounds. Specifically, the first two subcultures provided total esters concentrations higher (P < 0.05) than in unfermented milk. The total concentrations of esters in the third and fourth subcultures were either similar (P < 0.05) or lower (P < 0.05) compared to that found in milk.
2-Phenylethyl acetate, ethyl octanoate, and ethyl decanoate were the only esters compounds identified in both unfermented milk and all subcultures (Table 1). Of these esters, only ethyl octanoate was found at mean concentrations higher than its corresponding OPT of 19.30 μg/L [42] (Table 1 and Table 2). This was observed in the beverages obtained from the first two subcultures (30.70 and 30.12 μg/L) of the repeated 24-h fermentation. Additionally, in the beverage from the fourth subculture, the mean concentration of this compound (18.92 μg/L) was very near to 19.30 μg/L. Therefore, this compound is one of the key compounds in the aromatic profile of these beverages.
Isoamyl acetate was detected in the fermented beverages obtained in three subcultures (0-24, 48-72, and 72-96 h) from fermentation I, at mean concentrations above its OPT of 2 μg/L [42]. Therefore, this compound will have a direct contribution to the aroma of these beverages, with aromatic notes of banana, fruity, and sweet [47,73]. In contrast, the mean concentrations of isoamyl acetate found in the beverages from the 48-h subcultures were lower than its OPT (Table 1 and Table 2).
Ethyl hexanoate was the ester detected at the highest concentration during fermentation. The mean concentrations found in the four samples of the repeated 24-h fermentation (36.47, 34.07, 19.74, and 64.05 μg/L) were considerably higher than its OPT (1.00 μg/L) [42], thus contributing to the different beverages a characteristic flavor and aroma with banana and apple notes (Table 2). In the repeated 48-h fermentation, the mean concentrations of this compound were considerably lower (P < 0.05) than in repeated 24-h fermentation, but the first three subcultures provided beverages containing ethyl hexanoate levels higher than 1.00 μg/L. This compound was also detected by Walsh et al. [52] in milk kefir samples.
The other esters compounds were detected at concentrations lower than their corresponding OPT (Table 1 and Table 2). Some were detected in all fermented samples, some only in all subcultures of the repeated 24-h fermentation, and some only in certain subcultures of the first and/or second repeated fermentations.
Ethyl acetate was detected in all subcultures, with its mean concentration always being higher (P < 0.05) in the subcultures from fermentation I compared to the corresponding subcultures from fermentation II. This compound was also detected in skim milk kefir fermented with a mixture of concentrated cultures of strains from kefir grains: Lb. acidophilus LA5, Lb. delbrueckii subsp. bulgaricus LB12, S. thermophilus TH4, and K. marxianus subsp. marxianus LAF4 [88].
The rest of the esters detected during fermentation showed low concentrations, but their presence and possible synergistic effects could impart characteristic fruity aromas to the beverages [71].
3.3.6. Aromatic Hydrocarbons
Aromatic hydrocarbons have considerably high OPT values (Table 2), so their presence in fermented milk does not significantly affect the flavor of the beverage. However, at certain concentrations, aromatic hydrocarbons can serve as supplementary flavoring compounds, enhancing the flavor of the fermented product and giving it a fuller taste [69].
In this study, five aromatic hydrocarbons were identified in UHT whole milk, while 4, 5, 6, and 7 compounds were found in the four fermented beverages from the 24-h subcultures. The longer subcultures (48 h) provided a lower number of hydrocarbons (2, 2, 2, and 3). Additionally, the concentrations of aromatic hydrocarbons decreased significantly (P < 0.05) during milk fermentation, particularly in the four 48-h subcultures (Table 1).
3.3.7. Other Compounds
Within this group are included minor compounds whose chemical structure does not match any of the previous families. In fact, both the number and concentrations of these volatile compounds in unfermented and fermented milks were considerably low (Table 1).
3.4. Analysis of the Effect of Incubation Time on the Volatile Composition of the Fermented Beverages
In general, the results obtained in this study indicate that performing more than two 48-h subcultures led to a decrease in the number of volatile compounds compared to the repeated 24-h fermentation. Additionally, increasing the incubation time from 24 to 48 h provoked a drastic reduction in the total concentrations of the detected volatile compounds (see last two rows of Table 1).
The following two hypothesis could explain this phenomenon:
1. The metabolism of some microorganisms in kefir grains [97,98,99] and low pH values [98] could lead to the decrease in the number and/or concentration of these volatile compounds, resulting in the production of other volatile metabolites [97,98,99]. For instance, the reduction in the total concentration of hydrocarbons during the spontaneous or inoculated fermentation (with Lb. curvatus BIOTEC) of nham was attributed to the possible microbial oxidation of these compounds for the production of methyl ketones. Additionally, the reduction in the total concentration of aldehydes and ketones was attributed to the microbial conversion of these compounds for the synthesis of alcohols [97].
A decrease in the concentration of most esters, aldehydes, alcohols, and some organic acids was observed with the increase in incubation time during natural and inoculated (with a W-4 LAB culture) fermentation of minced pipper (Capsicum annuum L.) cv. “yanhong” [98]. The decrease in esters was related to their hydrolysis at low pH values, while the reduction in aldehydes was attributed to their conversion into acids and alcohols by microorganisms. Additionally, the decrease in the concentrations of alcohols was explained by their oxidation to organic acids due to microbial metabolism at low pH values [98], their assimilation by acetic acid bacteria, or microbial transformation or esterification in the production of kombucha [99]. The decline in organic acid levels was assumed to be caused by esterification reactions [99].
2. A more pronounced acidification of the milk in the 48-h subcultures resulted in final pH values (4.38, 4.02, 4.12, and 4.09) in the fermented samples, which were lower than those (5.17, 4.99, 4.70, and 4.70) in the beverages obtained from fermentation I. This significant decrease in the culture pH in the 48-h subcultures provoked a greater precipitation of proteins [100], which could facilitate the release of volatile compounds from the fermented substrate into the air, leading to a partial or total reduction of their concentration in the beverage.
In any case, the hypotheses proposed by Rotsatchakul et al. [97], Chen et al. [98], and Meng et al. [99] could not explain the observed reduction in the total concentrations of volatile compounds in the beverages from repeated 48-h fermentation, compared to those of the unfermented substrate and the beverages from fermentation I. This is mainly because the total concentrations of the seven families of volatile compounds (organic acids, alcohols, aldehydes, ketones, esters, hydrocarbons, and other compounds) decreased when the incubation time of milk with kefir grains increase from 24 to 48 h (Table 1).
Thus, the second hypothesis could be a more plausible reason to explain the reduction in the total concentrations of volatile compounds in the beverages from fermentation II. For this reason, the physical structure of the beverages obtained after different incubation times (24, 26, 28, 30, and 48 h) was examined (Figure 3).
As can be observed, the kefir beverage obtained at 24 h presents a creamy consistency very similar to that of yogurt with some clumps on the surface (Figure 3A). The gradual formation of gas bubbles becomes apparent from 26 h (Figure 3B), increasing in number and size at 28 (Figure 3C) and 30 (Figure 3D). Noticeable separation of whey and the formation of large gas bubbles was observed in the fermented milk obtained after 48 h of fermentation (Figure 3E), resulting in a beverage with an unappealing appearance. In addition, we observed that these gas bubbles progressively escaped from the liquid, probably dragging along the volatile components of the drink.
The results obtained in this study indicate that 24 h of fermentation appears to be sufficient to produce a milk kefir beverage with desirable odor and flavor notes and a better visual appearance compared to the beverages from the 48-h subcultures.
3.5. Analysis of the Aromatic Profile of the Fermented Milks
In UHT whole milk, the volatile compounds that had an odor activity value (OAV) ˃ 1.0 were 2-heptanone (OAV = 5.79) and 2-nonanone (OAV = 2.85), so they will contribute a greater impact to the odor of this substrate.
Regarding fermentations I and II, it could be noted that first fermentation provided the fermented beverages with the highest OAV values due to the aforementioned decrease in the concentrations of volatile compounds in fermentation II.
Ethyl hexanoate was the volatile compound with the highest OAV in the fermented beverages (except the fourth beverage from fermentation II). Thus, the OAV calculated for this compound in the four fermented beverages from the first, second, third, and fourth subcultures of fermentation I were 36.47, 34.07, 19.74, and 64.05, respectively. The OAV calculated for the four fermented beverages from the first, second, and third subcultures of fermentation II were 8.10, 2.36, and 2.52, respectively.
The volatile compounds that presented an OAV ˃ 1.0 in different subcultures were isoamyl acetate, in beverages from the first (1.29), third (1.01), and fourth (1.89) subcultures, and ethyl octanoate in beverages from the first (1.59) and second (1.56) passages in fermentation I. In contrast, other compounds only exhibited OAV ˃ 1.0 in one subculture: 2-heptanone (OAV = 1.10) in the first subculture, E-2-nonenal (OAV = 2.01) in the second subculture, and nonanal (OAV = 1.46) in the third subculture of fermentation I. Another compound with an OAV ˃ 1.0 was ethyl isobutanoate (OAV = 2.60), which was detected in the second subculture of fermentation II (Table 1 and Table 2).
Figure 4 and Figure 5 show the OAVs calculated for whole milk and the four subcultures of fermentations I and II (separated for better analysis) based on their classification into seven aromatic series (microbiological, fruity, floral, herbaceous, solvent, animal fatty, and sweet).
The aromatic series with the greatest contribution for whole milk were the fruity, floral and microbiological series. In the first subculture of fermentation I, the fruity was the most intense series (with an intensity 4.7 times greater than in whole milk), followed by the microbiological and floral series, but both with a lower magnitude than in whole milk (upper left part of Figure 4). In the second, third and fourth beverage from fermentation I, the fruity series predominated with an intensity 4.2, 2.6, and 7.6 times greater, respectively, than in whole milk. In the second and third subcultures, the fruity series was followed by the microbiological and herbaceous series, but in both cases, with less intensity than the fruity series. In the beverage from the fourth subculture, only the fruity series predominated (lower right part of Figure 4).
The results obtained for fermentation II (Figure 5) indicate that the fruity series predominated in the beverage from the first subculture with an intensity slightly higher than in whole milk (upper left part of Figure 5), while in the subsequent subcultures (48-96, 96-144, and 144-192 h), this series progressively lost intensity with the increase in the number of passages.
However, in the second subculture (96 h), the herbaceous series was also important, although with a smaller contribution than the fruity series. In contrast, in the last subculture, the intensity of the seven aromatic series was considerably lower than in unfermented milk, as the OAV of all volatile compounds in this subculture was below 1.0. These observations suggest, once again, that fermentation time significantly influences the sensory profile of kefir beverages.
3.6. Comparison of the Chemical Composition of Unfermented Milk and the Beverages Obtained in the Repeated 24- and 48-h Subcultures
Considering the different chemical composition of the nine milk beverages (Figure 1 and Figure 2), it is very difficult to visualize, a priori, the similarities and differences between them. Therefore, principal component analysis (PCA) was used to reduce the initial number of variables (pH, L, Pr, TN, LA, AA, EtOH, GOH, and TAA), transforming them into factors that explain the variability observed in the experimental data.
This variability can be conveniently explained by two factors (1 and 2) with eigenvalues greater than 1.0, which accounted for 67.75% and 18.86% of the total variance of the data, respectively. Together, these factors explained 86.61% of the total variance (Figure 6).
UHT whole milk was located in the lower right quadrant as the most different beverage. This is attributed to its high pH value (6.70), high nutrients (L, Pr, TN) contents, and low product (LA, AA, EtOH, GOH, and TAA) concentrations, compared to the fermented beverages (Figure 1 and Figure 2).
The fermented beverages from the first and second subculture of fermentation I (I−24 h and I−48 h) were located in the upper right quadrant because both beverages had higher final pH values, higher residual nutrient (L, Pr, TN), EtOH, and GOH concentrations, and lower LA, AA, and TAA levels compared to the other fermented beverages (Figure 1 and Figure 2).
The fermented beverage from the third subculture of fermentation I (I−72 h) was located in the upper left quadrant, as this beverage contained the lowest LA concentration (Figure 1 and Figure 2). The I-96 h beverage was located in the lower right quadrant, although far from the unfermented milk (Figure 6), since this fermented beverage contained the highest lactose concentration and the lowest LA level compared to the other fermented beverages (Figure 1 and Figure 2).
Regarding the fermentation II, it can be observed that the first fermented beverage (I−48 h) was located in the upper left quadrant since this sample contained the highest ethanol concentration (Figure 1 and Figure 2). The other three fermented samples (II−96 h, II−148 h and II−192 h) were located in the lower left quadrant (Figure 6) because these three beverages were the most acidic, have the lowest pH values and the highest levels of organic acids (LA and AA). Additionally, the three beverages contained the lowest concentrations of alcohols (EtOH and GOH) and the highest TAA titers (Figure 1 and Figure 2).
3.7. Comparison of the Volatile Composition of Unfermented Milk and the Beverages Obtained in the Repeated 24- and 48-h Subcultures
The high number (107) of volatile compounds detected in the nine milk beverages (Table 1) makes difficult to determine the relationship between the volatile compositions of the different beverages. Therefore, PCA was also applied to these data to easily interpret the relationships between the different beverages considering the total mean concentrations of each family of volatile compounds (organic acids, alcohols, aldehydes, ketones, esters, hydrocarbons, and other compounds).
In this analysis, factors 1 and 2 explained 46.01% and 37.89% of the total variation in the volatile composition of the beverages, respectively, accounting for a combined total of 83.90% of the variance (Figure 7). Factor 1 increased with higher concentrations of organic acids, hydrocarbons, ketones, and other compounds, while factor 2 increases with increasing concentrations of aldehydes, alcohols, and esters.
Once again, UHT whole milk was separated as an independent sample from the eight fermented samples and was located in the lower right quadrant, as it had the highest concentrations of ketones, hydrocarbons, organic acids, and other compounds and the lowest content in aldehydes. The four fermented samples from fermentation I were grouped together in the upper right quadrant due to their high content of alcohols, aldehydes, and esters compared to UHT milk and the four kefir beverages obtained from fermentation II. In contrast, the four fermented samples from fermentation II were grouped together in the lower left quadrant, as these beverages had the lowest concentrations of organic acids, ketones, hydrocarbons, and other compounds (Figure 7, Table 1). These results support the observation that increasing the fermentation time from 24 to 48 h leads to a decrease in the concentrations of volatile compounds in milk beverages fermented with kefir grains.
3.8. Microbiological Stability of the Beverages Obtained in the Repeated 24-h Subcultures
Considering that the four beverages from the repeated 24-h fermentation had the best visual appearance, their microbiological and chemical stability under refrigerated storage at 4 °C was analyzed over 28 days. Regarding the microbiological analysis, it can be noted that the LAB counts in the four subcultures decreased slightly at mean rates of 0.050, 0.016, 0.020, and 0.090 log CFU/mL/day reaching final levels of 7.28, 8.48, 8.49, and 8.42 log CFU/mL, respectively, in the four subcultures by day 28 (Figure 8). The AAB counts also decreased slightly, with mean rates of 0.016, 0.021, 0.013, and 0.015 log CFU/mL/day during storage, resulting in final counts of 8.42, 8.40, 8.69, and 8.34 log CFU/mL, respectively, by day 28 (Figure 8). In the case of yeasts, the counts decreased at mean rates of 0.003, 0.007, 0.025, and 0.007 log CFU/mL/day, with final counts of 6.13, 6.08, 6.57, and 6.08 log CFU/mL, respectively, on the 28th day in the four subcultures (Figure 8).
These decreases could be associated to the slight decrease in pH values (Figure 9) and the slight increase in the productions of LA, AA, and EtOH (Figure 10), which probably affected the viability of the LAB, AAB and yeasts [101]. This increase in product formation paralleled the consumption of lactose and proteins (Figure 9), which were consumed during storage at 4 °C at rates considerably lower than those during the corresponding 24-h subcultures (Figure 1 and Figure 9).
A similar trend in LAB viability, pH decrease, and protein consumption was observed previously in kefir [101] or yogurt [102] samples stored under refrigeration at 4 °C for 28 days. Irigoyen et al. [103] also observed a decreased in the lactobacilli and lactococci counts during the first 14 days of storage at 5 ± 1 °C, which then stabilized until the end of the storage period (28 days). In contrast, the AAB and yeast counts, as well as the pH and lactose content, did not show significant variation during storage.
However, the bacterial counts of the three microbial groups by day 28 always surpassed the minimal number of probiotic cells needed to observe a probiotic effect on the host [101]. Additionally, it is expected that a high number of these cells can reach the intestine and colonize it, as it has been suggested that the food matrix (milk) can protect these cells against gastrointestinal conditions [101,104].
5. Conclusions
The main contribution of this work is the description of the fermentation of UHT whole milk with milk kefir grains in repeated fermentations of 24 or 48-h, comparing the microbiological, physicochemical, and volatile compositions, as well as the visual appearance of the different kefir beverages.
Fermenting milk for 48 h with kefir grains produced a beverage containing high counts of LAB (> 106 UFC/mL in the four subcultures) and yeasts (> 106 UFC/mL, except in the four subculture). However, the concentration of volatile compounds decreased considerably compared to the kefir beverages obtained from the 24-h subcultures. Additionally, the increase in the fermentation time from 24 to 48 h negatively affected the visual appearance of the beverage due to the separation of whey and curd. In contrast, fermenting milk for 24 h was an appropriate strategy to produce kefir beverages with counts of probiotic cells (LAB and yeasts) higher than 106 UFC/mL, with relatively high concentrations of aromatic volatile compounds, and low concentrations of lactose, organic acids, and alcohols. Furthermore, a fermentation time of 24 h is sufficient to obtain a probiotic beverage with a good visual appearance compared to the beverages from the repeated 48-h subcultures.
Storage at 4 °C for 28 days of fermented milks from the four 24-h subcultures resulted in a slight reduction in the concentrations of lactose and proteins, as well as in the counts of LAB, AAB, and yeasts. However, in all cases, the counts of these microbial groups remained higher than 106 UFC/mL. This indicates that these beverages can be appropriately preserved by refrigerated storage during at least four weeks.
Author Contributions
Conceptualization, N.P.G.; methodology, D.L.B.T. and P.G.R.; software, N.P.G.; validation, D.L.B.T., S.C.D., and P.G.R.; formal analysis, N.P.G. and J.M.D.; investigation, N.P.G. and D.L.B.T.; resources, N.P.G. and J.M.D.; data curation, N.P.G., S.C.D., and D.L.B.T.; writing—original draft preparation, N.P.G. and D.L.B.T.; writing—review and editing, N.P.G. and J.M.D.; visualization, N.P.G. and D.L.B.T.; supervision, N.P.G.; project administration, N.P.G. and J.M.D.; funding acquisition, N.P.G. and J.M.D. All authors have read and agreed to the published version of the manuscript.
Funding
Delicia L. Bazán thanks the National University of Jaén (Perú) and the National Fellowship Program (PRONABEC) for their research grant (President of the Republic 2017-I-PRONABEC—Perú). Pablo G. Del-Río would like to express gratitude to Consellería de Cultura, Educación, Formación Profesional e Universidades of Xunta de Galicia for the financial support (ED481B-2022-020).
Data Availability Statement
Data is contained within the article.
Acknowledgments
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Teijeiro, M.; Pérez, P.F.; De Antoni, G.L.; Golowczyc, M.A. Suitability of Kefir Powder Production using Spray Drying. Food Res. Int. 2018, 112, 169–174. [Google Scholar] [CrossRef]
- Farag, M.A., Jomaa, S.A.; El-Wahed, A.A.; El-seedi, H.R. The Many Faces of Kefir Fermented Dairy Products: Quality Characteristics, Flavor Chemistry, Nutritional Value, Health Benefits, and Safety. Nutrients, 2020, 12, 1–21. [CrossRef]
- Guzel-Seydim, Z.B.; Çağlar, G.; Greene, A.K. A Comparison of Milk Kefir and Water Kefir: Physical, Chemical, Microbiological and Functional Properties. Trends Food Sci. Technol. 2021, 113, 42–53. [Google Scholar] [CrossRef]
- Ganatsios, V.; Nigam, P.; Plessas, S.; Terpou, A. Kefir as a Functional Beverage Gaining Momentum towards Its Health Promoting Attributes. Beverages 2021, 7, 48. [Google Scholar] [CrossRef]
- Bourrie, B.C.T.; Willing, B.P.; Cotter, P.D. The Microbiota and Health Promoting Characteristics of the Fermented Beverage Kefir. Frontiers Microbiol. 2016, 7, 647. [Google Scholar] [CrossRef]
- Pawlos, M.; Znamirowska, A.; Szajnar, K.; Kalicka, D. The Influence of the Dose of Calcium Bisglycinate on Physicochemical Properties, Sensory Analysis, and Texture Profile of Kefirs during 21 Days of Cold Storage. Acta Sci. Pol. Technol. Aliment. 2016, 15, 37–45. [Google Scholar] [CrossRef]
- Singh, G.; Muthukumarappan, K. Influence of Calcium Fortification on Sensory, Physical, and Rheological Characteristics of Fruit Yogurt. LWT - Food Sci. Technol. 2008, 41, 1145–1152. [Google Scholar] [CrossRef]
- Dertli, E.; Çon, A.H. Microbial Diversity of Traditional Kefir Grains and Their Role in Kefir Aroma. LWT - Food Sci. Technol. 2017, 85, 151–157. [Google Scholar] [CrossRef]
- Tzavaras, D.; Papadelli, M.; Ntaikou, I. From Milk Kefir to Water Kefir: Assessment of Fermentation Processes, Microbial Changes and Evaluation of the Produced Beverages. Fermentation 2022, 8, 135. [Google Scholar] [CrossRef]
- Bazán, D.L.; G. del Río, P.; Domínguez, J.M.; Cortés-Diéguez, S.; Mejuto, J.C.; Pérez-Guerra, N. The Chemical, Microbiological and Volatile Composition of Kefir-like Beverages Produced from Red Table Grape Juice in Repeated 24-h Fed-Batch Subcultures. Foods 2022, 11, 3117. [Google Scholar] [CrossRef]
- Randazzo, W.; Corona, O.; Guarcello, R.; Francesca, N.; Germanà, M.A.; Erten, H.; Moschetti, G.; Settanni, L. Development of New Non-Dairy Beverages from Mediterranean Fruit Juices fermented with Water Kefir Microorganisms. Food Microbiol. 2016, 54, 40–51. [Google Scholar] [CrossRef]
- Clarke, H.J.; Mannion, D.T.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Development of a Headspace Solid-Phase Microextraction Gas Chromatography Mass Spectrometry Method for the Quantification of Volatiles Associated with Lipid Oxidation in Whole Milk Powder using Response Surface Methodology. Food Chem. 2019, 292, 75–80. [Google Scholar] [CrossRef]
- García, M.; Esteve-Zarzoso, B.; Crespo, J.; Cabellos, J.M.; Arroyo, T. Influence of Native Saccharomyces cerevisiae Strains from D.O. “Vinos de Madrid” in the Volatile Profile of White Wines. Fermentation 2019, 5, 94. [Google Scholar] [CrossRef]
- Thomsen, M.; Martin, C.; Mercier, F.; Tournayre, P.; Berdagué, J.L.; Thomas-Danguin, T.; Guichard, E. Investigating Semi-Hard Cheese Aroma: Relationship between Sensory Profiles and Gas Chromatography-Olfactometry Data. Int. Dairy J. 2012, 26, 41–49. [Google Scholar] [CrossRef]
- Sánchez-Palomo, E.; Trujillo, M.; García Ruiz, A.; González Viñas, M.A. Aroma Profile of Malbec Red Wines from La Mancha Region: Chemical and Sensory Characterization. Food Res. Int. 2017, 100, 201–208. [Google Scholar] [CrossRef]
- Costas, M.; Alonso, E.; Guerra, N.P. Nisin Production in Realkalized Fed-Batch Cultures in Whey with Feeding with Lactose- or Glucose-Containing Substrates. Appl. Microbiol. Biotechnol. 2016, 100, 7899–7908. [Google Scholar] [CrossRef]
- Costas, M.; Alonso, E.; Bazán, D.L.; Bendaña, R.J.; Guerra, N.P. Batch and Fed-Batch Production of Probiotic Biomass and Nisin in Nutrient-Supplemented Whey Media. Braz. J. Microbiol. 2019, 50, 915–925. [Google Scholar]
- Costas, M.; Alonso, E.; Bendaña, R.J.; Guerra, N.P. The Joint Effect of pH Gradient and Glucose Feeding on the Growth Kinetics of Lactococcus lactis CECT 539 in Glucose-Limited Fed-Batch Cultures. Polish J. Microbiol. 2019, 68, 269–280. [Google Scholar]
- Valli, M.; Sauer, M.; Branduardi, P.; Borth, N.; Porro, D.; Mattanovich, D. Improvement of Lactic Acid Production in Saccharomyces cerevisiae by Cell Sorting for High Intracellular pH. Appl. Environ. Microbiol. 2006, 72, 5492–549. [Google Scholar] [CrossRef]
- Yalçin, S.K.; Özbaş, Z.Y. Determination of Growth and Glycerol Production Kinetics of a Wine Yeast Strain Saccharomyces cerevisiae Kalecik 1 in Different Substrate Media. World J. Microbiol. Biotechnol. 2005, 21, 1303–1310. [Google Scholar] [CrossRef]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in Sustainable Bioethanol Production: A Review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar]
- Ribéreau-Gayon, P.; Dubourdieu, D.; Donèche, B. B.; Lonvaud, AA.; Darriet, P.; Towey, J. Handbook of Enology. 3rd ed.; John Wiley & Sons Ltd: Chichester, UK, 2021; Volume 1: The Microbiology of Wine and Vinifications; pp. 229–241.
- Neysens, P.; Messens, W.; Gevers, D.; Swings, J.; De Vuyst, L. Biphasic Kinetics of Growth and Bacteriocin Production with Lactobacillus amylovorus DCE 471 Occur Under Stress Conditions. Microbiol. 2003, 149, 1073–1082. [Google Scholar] [CrossRef]
- Yang, R.; Ray, B. Factors Influencing Production of Bacteriocins by Lactic Acid Bacteria. Food Microbiol. 1994, 11, 281–291. [Google Scholar] [CrossRef]
- Moradi, Z.; Kalanpour, N. Kefiran, a Branched Polysaccharide: Preparation, Properties, and Applications: A Review. Carbohydr. Polym. 2019, 223, 115100. [Google Scholar]
- Sayes, C.; Leyton, Y.; Riquelme, C. Probiotic Bacteria as a Healthy Alternative for Fish Aquaculture. In Antibiotic Use in Animals, 1st ed.; Savic, S., Ed.; InTechOpen Limited, London, UK, 2018; Chapter 7, pp. 115–132.
- Gallina, D.A.; Barbosa, P.P.M.; Ormenese, R.C.S.C.; Garcia, A.O. Development and Characterization of Probiotic Fermented Smoothie Beverage. Rev. Ciênc. Agron. 2019, 50, 378–386. [Google Scholar]
- Poolman, B.; Konings, W.N. Relation of Growth of Streptococcus lactis and Streptococcus cremoris to Amino Acid Transport. J. Bacteriol. 1988, 170, 700–707. [Google Scholar] [CrossRef]
- Fajardo, P.; Rodríguez, I.; Pastrana, L.; Guerra, N.P. Production of a Potentially Probiotic Culture of Lactobacillus casei subsp. casei CECT 4043 in Whey. Int. Dairy J. 2008, 18, 1057–1065. [Google Scholar]
- Membré, J.M.; Kubaczka, M.; Chéné, C. Combined Effects of pH and Sugar on Growth Rate of Zygosaccharomyces rouxii, a Bakery Product Spoilage Yeast. Appl. Environ. Microbiol. 1999, 65, 4921–4925. [Google Scholar] [CrossRef]
- Arroyo, F.N.; Orlić, S.; Querol, A.; Barrio, E. Effects of Temperature, pH, and Sugar Concentration on the Growth Parameters of Saccharomyces cerevisiae, S. kudriavzevii and Their Interspecific Hybrid. Int. J. Food Microbiol. 2009, 131, 120–127. [Google Scholar] [CrossRef]
- Oude, S.J.W.H.; Krooneman, J.; Gottschal, J.C.; Spoelstra, S.F.; Faber, F.; Driehuis, F. Anaerobic Conversion of Lactic Acid to Acetic Acid and 1,2-Propanediol by Lactobacillus buchneri. Appl. Environ. Microbiol. 2001, 67, 125–132. [Google Scholar] [CrossRef]
- Outeiriño, D.; Costa-Trigo, I.; Ochogavias, A.; Pinheiro de Souza Oliveira, R.; Guerra, N.P.; Salgado, J.M.; Domínguez, J.M. Biorefinery of Brewery Spent Grain to obtain Bioproducts with High Value-added in the Market. New Biotechnol. 2024, 79, 111–119. [Google Scholar] [CrossRef]
- Guerra, N.P.; Pastrana, L. Enhancement of Nisin Production by Lactococcus lactis in Periodically Re-Alkalized Cultures. Proc. Biochem. 2002, 37, 1005–1015. [Google Scholar] [CrossRef]
- Tada, S.; Katakura, Y.; Ninomiya, K.; Shioya, S. Fed-Batch Coculture of Lactobacillus kefiranofaciens with Saccharomyces cerevisiae for Effective Production of Kefiran. J. Biosci. Bioeng. 2007, 103, 557–562. [Google Scholar] [CrossRef]
- Cheirsilp, B.; Radchabut, S. Use of Whey Lactose from Dairy Industry for Economical Kefiran Production by Lactobacillus kefiranofaciens in Mixed Cultures with Yeasts. New Biotechnol. 2011, 28, 574–580. [Google Scholar] [CrossRef]
- Felipe, M.G.; Vieira, D.C.; Vitolo, M.; Silva, S.S.; Roberto, I.C.; Manchilha, I.M. Effect of Acetic Acid on Xylose Fermentation to Xylitol by Candida guilliermondii. J. Basic Microbiol. 1995, 35, 171–177. [Google Scholar] [CrossRef]
- Kirtadze, E.; Nutsubidze, N. Metabolic Potential of Alcoholic Fermentation Yeasts. Bull. Georgian Natl. Acad. Sci. 2009, 3, 110–116. [Google Scholar]
- Gomes, R.J.; Borges, M.F.; Rosa, M.F.; Castro-Gómez, R.J.H.; Spinosa, W.A. Acetic Acid Bacteria in the Food Industry: Systematics, Characteristics and Applications. Food Technol. Biotechnol. 2018, 56, 139–151. [Google Scholar] [CrossRef]
- Garai-Ibabe, G.; Ibarburu, I.; Berregi, I.; Claisse, O.; Lonvaud-Funel, A.; Irastorza, A.; Dueñas, M.T. Glycerol Metabolism and Bitterness Producing Lactic Acid Bacteria in Cidermaking. Int. J. Food Microbiol. 2008, 121, 253–261. [Google Scholar] [CrossRef]
- Mamlouk, D.; Gullo, M. Acetic Acid Bacteria: Physiology and Carbon Sources Oxidation. Indian J. Microbiol., 2013, 53, 377–384. [Google Scholar] [CrossRef]
- van Gemert, L.J. Compilations of Odour Threshold Values in Air, Water and other Media. 2nd ed.; Oliemans Punter & Partners BV, The Netherlands, 2011.
- Escudero, A.; Gogorza, B.; Melús, M.A.; Ortín, N.; Cacho, J.A.; Ferreira, V. Characterization of the Aroma of a Wine from Maccabeo. Key Role played by Compounds with Low Odor Activity Values. J. Agric. Food Chem. 2004, 52, 3516–3524. [Google Scholar] [CrossRef]
- Siebert, T.E.; Smyth, H.E.; Capone, D.L.; Neuwöhner, C.; Pardon, K.H.; Skouroumounis, G.K.; Herderich, M.J.; Sefton, M.A.; Pollnitz, A.P. Stable Isotope Dilution Analysis of Wine Fermentation Products by HS-SPME-GC-MS. Anal. Bioanal. Chem. 2005, 381, 937–947. [Google Scholar] [CrossRef]
- Pereira, R.; Resende, D.; Alencar, A.C.; de Abreu, L.R.; Ferreira, W. Survival of Kluyveromyces lactis and Torulaspora delbrueckii to Simulated Gastrointestinal Conditions and their Use as Single and Mixed Inoculum for Cheese Production. Food Res. Int. 2019, 125, 108620. [Google Scholar]
- Czerny, M.; Schieberle, P. Important Aroma Compounds in Freshly Ground Wholemeal and White Wheat Flour Identification and Quantitative Changes during Sourdough Fermentation. J. Agric. Food Chem. 2002, 50, 6835–6840. [Google Scholar] [CrossRef]
- Li, B.; Gao, X.; Li, N.; Mei, J. Fermentation Process of Mulberry Juice-Whey based Tibetan Kefir Beverage Production. Czech J. Food Sci. 2018, 36, 494–501. [Google Scholar] [CrossRef]
- Meilgaard, M.C. Flavor Chemistry of Beer: Part II: Flavor and Threshold of 239 Aroma Volatiles. Tech. Q. Master Brew. Assoc. Am. 1975, 12, 151–168. [Google Scholar]
- Dertli, E.; Çon, A.H. Microbial Diversity of Traditional Kefir Grains and their Role on Kefir Aroma. LWT-Food Sci. Technol. 2017, 85, 151–157. [Google Scholar] [CrossRef]
- Vinh, T.; Schwartz, G.; Moll, M. Identification of 2-Ethyl-Hexanoic (2-Ethyl-Caproic) Acid in Beer. J. Am. Soc. Brew. Chem. 1981, 39, 2–5. [Google Scholar] [CrossRef]
- Oliveira, D.R.; Lopes, A.C.A.; Pereira, R.A.; Cardoso, P.G.; Duarte, W.F. Selection of Potentially Probiotic Kluyveromyces lactis for the Fermentation of Cheese Whey–based Beverage. Ann. Microbiol. 2019, 69, 1361–1372. [Google Scholar] [CrossRef]
- Walsh, A.M.; Crispie, F.; Kilcawley, K.; O’Sullivan, O.; O’Sullivan, M.G.; Claesson, M.J.; Cotter, P.D. Microbial Succession and Flavor Production in the Fermented Dairy Beverage Kefir. MSystems 2016, 1, e00052–16. [Google Scholar] [CrossRef]
- Tian, H.; Shi, Y.; Zhang, Y.; Yu, H.; Mu, H.; Chen, C. Screening of Aroma–Producing Lactic Acid Bacteria and their Application in improving the Aromatic Profile of Yogurt. J. Food Biochem. 2019, 43, e12837. [Google Scholar] [CrossRef]
- Clarke, H.J.; Mannion, D.T.; O’Sullivan, M.G.; Kerry, J.P.; Kilcawley, K.N. Development of a Headspace Solid-Phase Microextraction Gas Chromatography Mass Spectrometry Method for the Quantification of Volatiles associated with Lipid Oxidation in Whole Milk Powder using Response Surface Methodology. Food Chem. 2019, 292, 75–80. [Google Scholar] [CrossRef]
- Hayaloglu, A.A.; Brechany, E.Y.; Deegan, K.C.; McSweeney, P.L.H. Characterization of the Chemistry, Biochemistry and Volatile Profile of Kuflu Cheese, a Mould-ripened Variety. LWT-Food Sci. Technol. 2008, 41, 1323–1334. [Google Scholar] [CrossRef]
- Zheng, X.; Li, K.; Shi, X.; Ni, Y.; Li, B.; Zhuge, B. Potential Characterization of Yeasts isolated from Kazak Artisanal Cheese to produce Flavoring Compounds. MicrobiologyOpen. 2018, 7, e00533. [Google Scholar] [CrossRef]
- Salgado, J.M.; González, C.; Rodríguez, R.; Simal, J.; Domínguez, J.M.; Cortés, S. Study of the Volatile Compounds produced by Debaryomyces hansenii NRRL Y-7426 during the Fermentation of Detoxified Concentrated Distilled Grape Marc Hemicellulosic Hydrolysates. World J. Microbiol. Biotechnol. 2012, 28, 3123–3134. [Google Scholar] [CrossRef]
- Pico, J.; Martínez, M.M.; Bernal, J.; Gómez, M. Evolution of Volatile Compounds in Gluten-free Bread: From Dough to Crumb. Food Chem. 2017, 227, 179–186. [Google Scholar] [CrossRef]
- Czerny, M.; Christlbauer, M.; Christlbauer, M.; Fischer, A.; Granvogl, M.; Hammer, M.; Hartl, C.; Hernandez, N.M.; Schieberle, P. Re-investigation on Odour Thresholds of Key Food Aroma Compounds and Development of an Aroma Language based on Odour Qualities of defined Aqueous Odorant Solutions. Eur. Food Res. Technol. 2008, 228, 265–273. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, W.; Yu, W.; Zhao, L.; Song, S.; Xu, W.; Zhang, C.; Ma, C.; Wang, L.; Wang, S. Study on the Volatile Composition of Table Grapes of Three Aroma Types. LWT-Food Sci. Technol. 2019, 115, 108450. [Google Scholar] [CrossRef]
- Chastrette, M. , Cretin, D., El Aïdi, C. El. Structure-Odor Relationships: Using Neural Networks in the Estimation of Camphoraceous or Fruity Odors and Olfactory Thresholds of Aliphatic Alcohols. J. Chem. Inf. Comput. Sci. 1996, 36, 108–113. [Google Scholar] [CrossRef]
- Komes, D.; Ulrich, D.; Lovric, T. Characterization of Odor-Active Compounds in Croatian Rhine Riesling Wine, Subregion Zagorje. Eur. Food Res. Technol. 2006, 222, 1–7. [Google Scholar] [CrossRef]
- Pionnier, E.; Hugelshofer, D. Characterisation of Key Odorant Compounds in Creams from Different Origins with Distinct Flavours. Dev. Food Sci. 2006, 43, 233–236. [Google Scholar]
- Sarhir, S.T.; Amanpour, A.; Bouseta, A.; Selli, S. Key Odorants of a Moroccan fermented Milk Product “Lben” using Aroma Extract Dilution Analysis. J. Food Sci. Technol. 2019, 56, 3836–3845. [Google Scholar] [CrossRef]
- Verma, D.K.; Srivastav, P.P. A Paradigm of Volatile Aroma Compounds in Rice and their Product with Extraction and Identification Methods: A Comprehensive Review. Food Res. Int. 2020, 130, 108924. [Google Scholar] [CrossRef]
- Welke, J.E.; Zanus, M.; Lazzarotto, M.; Alcaraz, C. Quantitative Analysis of Headspace Volatile Compounds using Comprehensive Two-dimensional Gas Chromatography and their Contribution to the Aroma of Chardonnay Wine. Food Res. Int. 2014, 59, 85–99. [Google Scholar] [CrossRef]
- Dimitrellou, D.; Kandylis, P.; Lević, S.; Petrović, T.; Ivanović, S.; Nedović, V.; Kourkoutas, Y. Encapsulation of Lactobacillus casei ATCC 393 in Alginate Capsules for Probiotic fermented Milk Production. LWT-Food Sci. Technol. 2019, 116, 108501. [Google Scholar] [CrossRef]
- Bingham, E.; Cohrssen, B.; Powell, C.H. Patty's Toxicology. Volume 6, 5th ed.; John Wiley & Sons. Inc. New York, USA, 2001; pp. 318.
- Dan, T.; Chen, H.; Li, T.; Tian, J.; Ren, W.; Zhang, H.; Sun, T. Influence of Lactobacillus plantarum P-8 on Fermented Milk Flavor and Storage Stability. Front. Microbiol. 2019, 9, 3133. [Google Scholar] [CrossRef]
- Montgomery, J.H. Groundwater Chemicals, 4th ed.; Taylor & Francis Group. Boca Raton, Florida, USA, 2007; pp. 718.
- Liu, S.Q.; Holland, R.; Crow, V.L. Esters and their Biosynthesis in Fermented Dairy Products: A Review. Int. Dairy J. 2004, 14, 923–945. [Google Scholar] [CrossRef]
- Gil, M.; Cabellos, J.M.; Arroyo, T.; Prodanov, M. Characterization of the Volatile Fraction of Young Wines from the Denomination of Origin “Vinos de Madrid” (Spain). Anal. Chim. Acta. 2006, 563, 145–153. [Google Scholar] [CrossRef]
- Peinado, R.A.; Moreno, J.; Bueno, J.E.; Moreno, J.A.; Mauricio, J.C. Comparative Study of Aromatic Compounds in Two Young White Wines subjected to Prefermentative Cryomaceration. Food Chem. 2004, 84, 585–590. [Google Scholar] [CrossRef]
- Ferreira, V.; Lopez, R.; Cacho, J.F. Quantitative Determination of the Odorants of Young Red Wines from Different Grape Varieties. J. Sci. Food Agric. 2000, 80, 1659–1667. [Google Scholar] [CrossRef]
- Burdock, G.A. Fenaroli´s Handbook of Flavour Ingredients, 6th ed.; CRC Press. Taylor & Francis Group LLC, Boca Raton, Florida, USA, 2010.
- Nikolaou, A.; Tsakiris, A.; Kanellaki, M.; Bezirtzoglou, E.; Akrida-Demertzi, K.; Kourkoutas, Y. Wine Production using Free and Immobilized Kefir Culture on Natural Supports. Food Chem. 2019, 272, 39–48. [Google Scholar] [CrossRef]
- Cheng, H. Volatile Flavor Compounds in Yogurt: A Review. Crit. Rev. Food Sci. Nutr. 2010, 50, 938–950. [Google Scholar] [CrossRef]
- Curioni, P.M.G.; Bosset, J.O. Key Odorants in Various Cheese Types as determined by Gas Chromatography-Olfactometry. Int. Dairy J. 2002, 12, 959–984. [Google Scholar] [CrossRef]
- Bugaud, C.; Alter, P. Volatile and Non-Volatile Compounds as Odour and Aroma Predictors in Dessert Banana (Musa spp.). Postharvest Biol. Technol. 2016, 112, 14–23. [Google Scholar] [CrossRef]
- López, M.L.; Villatoro, C.; Fuentes, T.; Graell, J.; Lara, I.; Echeverría, G. Volatile Compounds, Quality Parameters and Consumer Acceptance of ‘Pink Lady®’ Apples stored in Different Conditions. Postharvest Biol. Technol. 2007, 43, 55–66. [Google Scholar] [CrossRef]
- Zepka, L.Q.; Garruti, D.S.; Sampaio, K.L.; Mercadante, A.Z.; Da Silva, M.A.A.P. Aroma Compounds derived from the Thermal Degradation of Carotenoids in a Cashew Apple Juice Model. Food Res. Int. 2014, 56, 108–114. [Google Scholar] [CrossRef]
- Barron, L.; Redondo, Y.; Aramburu, M.; Pérez-Elortondo, F.J.; Albisu, M.; Nájera, A.I.; de Renobales, M. Variations in Volatile Compounds and Flavour in Idiazabal Cheese Manufactured from Ewe’s Milk in Farmhouse and Factory. J. Sci. Food Agric. 2005, 85, 1660–1671. [Google Scholar] [CrossRef]
- Hinge, V.R.; Patil, H.B.; Nadaf, A.B. Aroma Volatile Analyses and 2AP Characterization at Various Developmental Stages in Basmati and Non-Basmati Scented Rice (Oryza sativa L.) cultivars. Rice. 2016, 9, 38. [Google Scholar] [CrossRef]
- Dan, T.; Wang, D.; Jin, R.L.; Zhang, H.P.; Zhou, T.T.; Sun, T.S. Characterization of Volatile Compounds in Fermented Milk using Solid-Phase Microextraction Methods coupled with Gas Chromatography-Mass Spectrometry. J. Dairy Sci. 2017, 100, 2488–2500. [Google Scholar] [CrossRef]
- Dan, T.; Wang, D.; Wu, S.; Jin, R.; Ren, W.; Sun, T. Profiles of Volatile Flavor Compounds in Milk Fermented with Different Proportional Combinations of Lactobacillus delbrueckii subsp. bulgaricus and Streptococcus thermophilus. Molecules. 2017, 22, 1633. [Google Scholar]
- Pan, D.D.; Wu, Z.; Peng, T.; Zeng, X.Q.; Li, H. Volatile Organic Compounds Profile during Milk Fermentation by Lactobacillus pentosus and Correlations between Volatiles Flavor and Carbohydrate Metabolism. J. Dairy Sci. 2014, 97, 624–631. [Google Scholar] [CrossRef]
- Kesler, M.K.; González-Orozco, B.D.; Barringer, S.A.; Alvarez, V.B. Mitigation of Undesirable Volatile Aroma Compounds in Kefir by Freeze Drying and Vacuum Evaporation. J. Food Sci. 2023, 88, 3216–3227. [Google Scholar] [CrossRef]
- Aghlara, A.; Mustafa, S.; Manap, Y.A.; Mohamad, R. Characterization of Headspace Volatile Flavor Compounds formed During Kefir Production: Application of Solid Phase Microextraction. Int. J. Food Prop. 2009, 12, 808–818. [Google Scholar] [CrossRef]
- Valero, E.; Villamiel, M.; Miralles, B.; Sanz, J.; Martı́nez-Castro, I. Changes in Flavour and Volatile Components during Storage of Whole and Skimmed UHT Milk. Food Chem. 2001, 72, 51–58. [Google Scholar] [CrossRef]
- Fang, X.; Guo, L.W.; Chen, H.; Ke, W.C.; Guo, W.; Guo, X.S.; Zhang, Y. Characteristics of Volatile Flavor Components in Traditional Fermented Yak Milk Produced in Different Ecoregions of the Qinghai-Tibetan Plateau. J. Dairy Sci. 2020, 103, 191–200. [Google Scholar] [CrossRef]
- Delgado, F.J.; González-Crespo, J.; Cava, R.; Ramírez, R. Formation of the Aroma of a Raw Goat Milk Cheese during Maturation analysed by SPME–GC–MS. Food Chem. 2011, 129, 1156–1163. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, H.; Chen, W.; Zhong, Q.; Zhang, G.; Chen, W. Beneficial Effects of Tomato Juice Fermented by Lactobacillus plantarum and Lactobacillus casei: Antioxidation, Antimicrobial Effect, and Volatile Profiles. Molecules. 2018, 23, 2366. [Google Scholar] [CrossRef]
- Zareba, D.; Ziarno, M.; Obiedzinski, M. Volatile Profile of Non-Fermented Milk and Milk Fermented by Bifidobacterium animalis subsp. lactis. Int. J. Food Prop. 2012, 15, 1010–1021. [Google Scholar] [CrossRef]
- Molimard, P.; Spinnler, H.E. Review: Compounds involved in the Flavor of Surface Mold-Ripened Cheeses: Origins and Properties. J. Dairy Sci. 1996, 79, 169–184. [Google Scholar] [CrossRef]
- Arslan, S.; Mutlu, C.; Candal, C.; Erbaş, M. Microbiological and Chemical Properties of Wet Tarhana produced by Different Dairy Products. J. Food Sci. Technol. 2018, 55, 4770–4781. [Google Scholar] [CrossRef]
- Tu, C.; Azi, F.; Huang, J.; Xu, X.; Xing, G.; Dong, M. Quality and Metagenomics Evaluation of a Novel Functional Beverage produced from Soy Whey using Water Kefir Grains. LWT-Food Sci. Technol. 2019, 113, 108258. [Google Scholar] [CrossRef]
- Rotsatchakul, P.; Visesanguan, W.; Smitinont, T.; Chaiseri, S. Changes in Volatile Compounds during Fermentation of Nham (Thai Fermented Sausage). Int. Food Res. J. 2009, 16, 391–414. [Google Scholar]
- Chen, Y.; Xu, H.; Ding, S.; Zhou, H.; Qin, D.; Deng, F.; Wang, R. Changes in Volatile Compounds of Fermented Minced Pepper during Natural and Inoculated Fermentation Process based on Headspace–Gas Chromatography–Ion Mobility Spectrometry. Food Sci. Nut. 2020, 8, 3362–3379. [Google Scholar] [CrossRef]
- Meng, Y.; Wang, X. , Li, Y.; Chen, J.; Chen, X. Microbial Interactions and Dynamic Changes of Volatile Flavor Compounds during the Fermentation of Traditional Kombucha. Food Chem. 2024, 430, 137060. [Google Scholar] [CrossRef]
- Liang, J.; Yoo, M.J.Y.; Seale, B.; Grazioli, G. Nutritional and Volatile Characterisation of Milk Inoculated with Thermo-Tolerant Lactobacillus bulgaricus through Adaptive Laboratory Evolution. Foods 2021, 10, 2944. [Google Scholar] [CrossRef]
- Buran, I.; Akal, C.; Ozturkoglu-Budak, S.; Yetisemiyen, A. Rheological, Sensorial and Volatile Profiles of Synbiotic Kefirs produced from Cow and Goat Milk Containing Varied Probiotics in Combination with Fructooligosaccharide. LWT-Food Sci. Technol. 2021, 148, 111591. [Google Scholar] [CrossRef]
- Machado, T.A.D.G.; de Oliveria, M.E.G.; Campos, M.I.F.; de Assis, P.O.A.; de Souza, E.L.B.; Madruga, M.S.; Pacheco, M.T.B.; Pintado, M.M.E.-, Queiroga, R.C.R.E. Impact of Honey on Quality Characteristics of Goat Yogurt Containing Probiotic Lactobacillus acidophilus. LWT-Food Sci. Technol. 2017, 80, 221–229. [CrossRef]
- Irigoyen, A.; Arana, I.; Castiella, M.; Torre, P.; Ibañez, F.C. Microbiological, Physicochemical, and Sensory Characteristics of Kefir during Storage. Food Chem. 2005, 90, 613–620. [Google Scholar] [CrossRef]
- Perez, R.; Alonso-Gonzalez, E.; Mejuto, J.C.; Guerra, N. Production of a Potentially Product for Animal Feed and Evaluation of Some of its Probiotic Properties. Int. J. Mol. Sci. 2021, 22, 10004. [Google Scholar] [CrossRef]
Figure 1.
Chemical Composition (as Means ± Standard Deviation of Triplicate Samples and Two Analytical Replications Each) of the UHT Whole Milk (UHT-WM) and Milk Kefir Beverages Obtained from the first (Sub-I), second (Sub-II), third (Sub-III), and fourth (Sub-IV) 24-h Batch Subcultures. L, TN, Pr, LA, AA, EtOH, and GOH are the concentrations of lactose, total nitrogen, proteins, lactic acid, acetic acid, ethanol, and glycerol, respectively. TAA and KGW are the total antibacterial activity and drained kefir grain weight, respectively. Similar letters above the bars for each variable indicate that there is no statistical difference between their values (P > 0.05).
Figure 1.
Chemical Composition (as Means ± Standard Deviation of Triplicate Samples and Two Analytical Replications Each) of the UHT Whole Milk (UHT-WM) and Milk Kefir Beverages Obtained from the first (Sub-I), second (Sub-II), third (Sub-III), and fourth (Sub-IV) 24-h Batch Subcultures. L, TN, Pr, LA, AA, EtOH, and GOH are the concentrations of lactose, total nitrogen, proteins, lactic acid, acetic acid, ethanol, and glycerol, respectively. TAA and KGW are the total antibacterial activity and drained kefir grain weight, respectively. Similar letters above the bars for each variable indicate that there is no statistical difference between their values (P > 0.05).
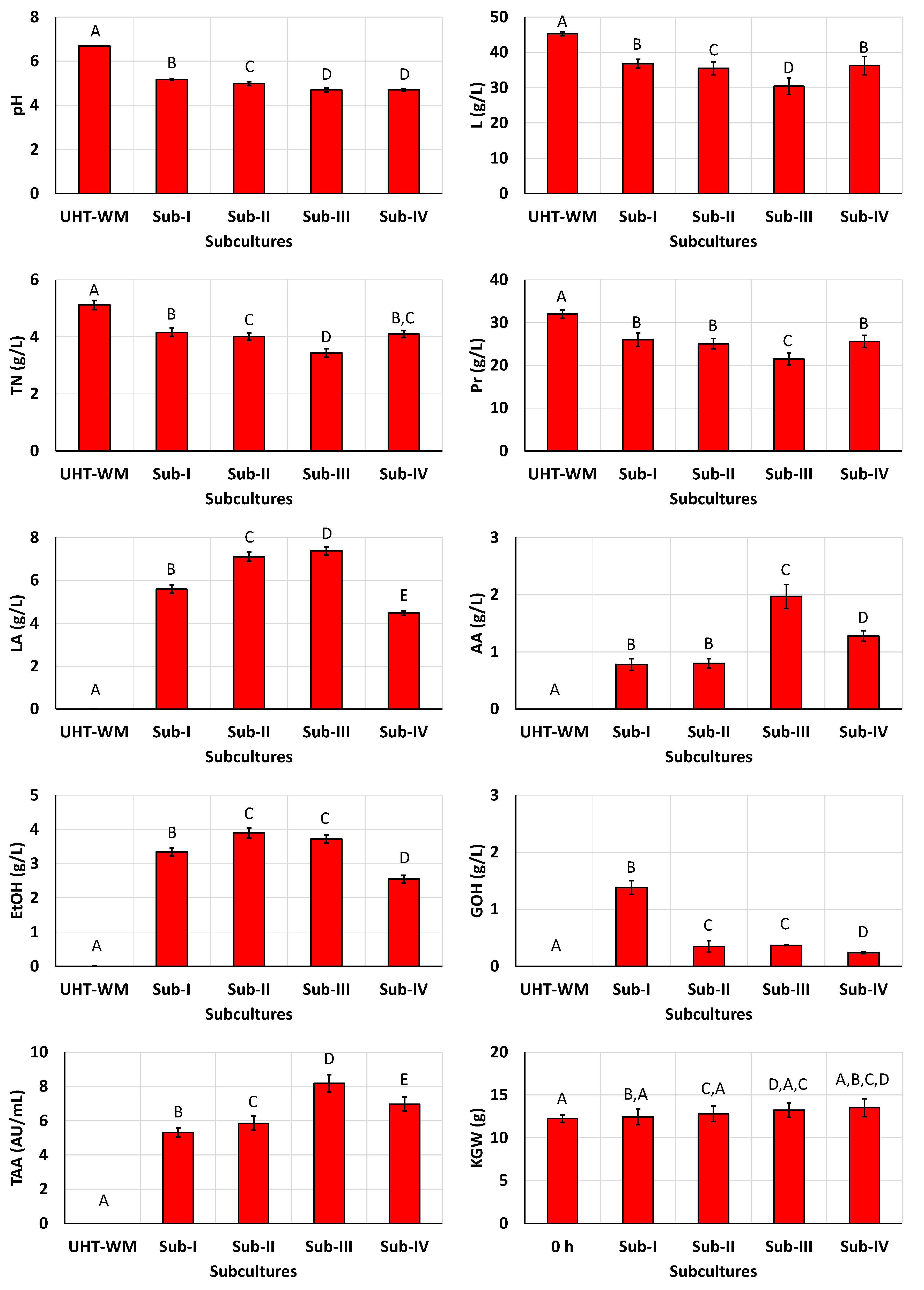
Figure 2.
Chemical Composition (as Means ± Standard Deviation of Triplicate Samples and Two Analytical Replications Each) of the UHT Whole Milk (UHT-WM) and Milk Kefir Beverages Obtained from the first (Sub-I), second (Sub-II), third (Sub-III), and fourth (Sub-IV) 48-h Batch Subcultures. L, TN, Pr, LA, AA, EtOH, and GOH are the concentrations of lactose, total nitrogen, proteins, lactic acid, acetic acid, ethanol, and glycerol, respectively. TAA and KGW are the total antibacterial activity and drained kefir grain weight, respectively. Similar letters above the bars for each variable indicate that there is no statistical difference between their values (P > 0.05).
Figure 2.
Chemical Composition (as Means ± Standard Deviation of Triplicate Samples and Two Analytical Replications Each) of the UHT Whole Milk (UHT-WM) and Milk Kefir Beverages Obtained from the first (Sub-I), second (Sub-II), third (Sub-III), and fourth (Sub-IV) 48-h Batch Subcultures. L, TN, Pr, LA, AA, EtOH, and GOH are the concentrations of lactose, total nitrogen, proteins, lactic acid, acetic acid, ethanol, and glycerol, respectively. TAA and KGW are the total antibacterial activity and drained kefir grain weight, respectively. Similar letters above the bars for each variable indicate that there is no statistical difference between their values (P > 0.05).
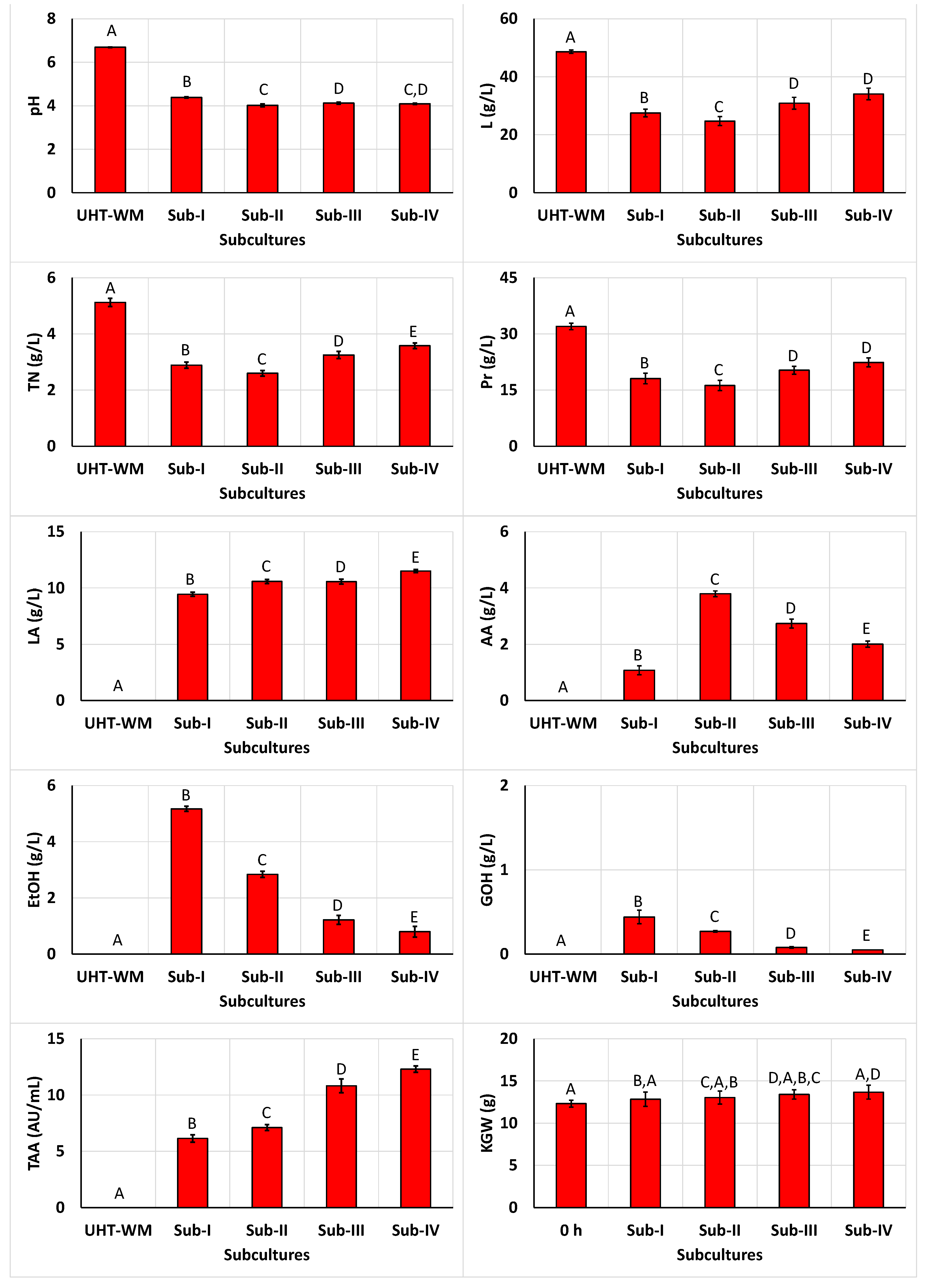
Figure 3.
Visual appearance of milk kefir beverages at 24 (A), 26 (B), 28 (C), 30 (D) and 48 h (E) of fermentation under static conditions.
Figure 3.
Visual appearance of milk kefir beverages at 24 (A), 26 (B), 28 (C), 30 (D) and 48 h (E) of fermentation under static conditions.

Figure 4.
Radar map of the total OAVs of the odor descriptors of UHT whole milk and kefir beverages from the four consecutive static subcultures of fermentation I (I−24 h, I−48 h, I−72 h and I−96 h). .
Figure 4.
Radar map of the total OAVs of the odor descriptors of UHT whole milk and kefir beverages from the four consecutive static subcultures of fermentation I (I−24 h, I−48 h, I−72 h and I−96 h). .
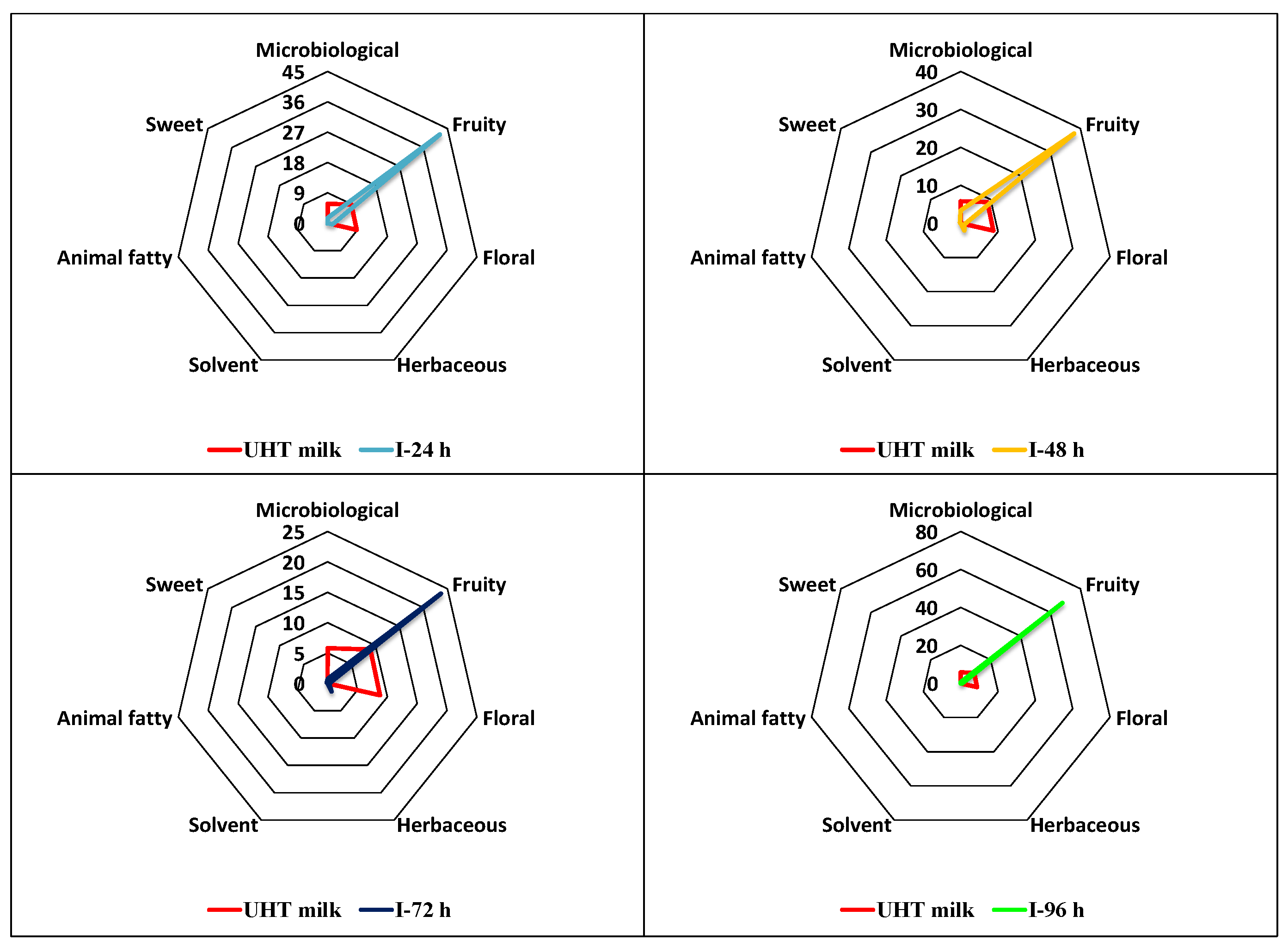
Figure 5.
Radar map of the total OAVs of the odor descriptors of UHT whole milk and kefir beverages from the four consecutive static subcultures of fermentation II (II−48 h, II−96 h, II−148 h and II−192 h).
Figure 5.
Radar map of the total OAVs of the odor descriptors of UHT whole milk and kefir beverages from the four consecutive static subcultures of fermentation II (II−48 h, II−96 h, II−148 h and II−192 h).
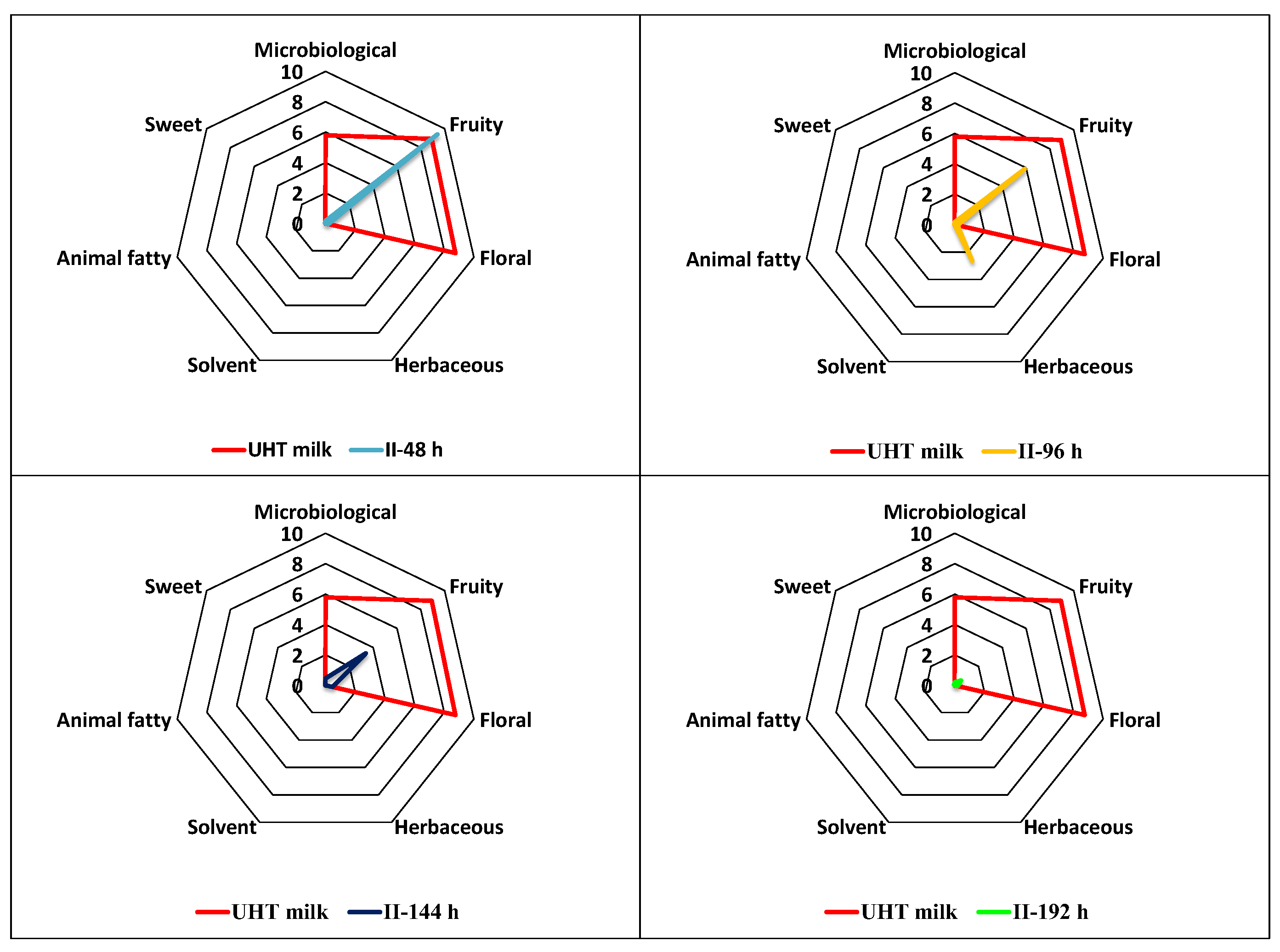
Figure 6.
Principal component analysis based on the differences between the pH values and contents in lactose (L, in g/L), total nitrogen (TN, in g/L), proteins (Pr, in g/L), lactic acid (LA, in g/L), acetic acid (AA, in g/L), ethanol (EtOH, in g/L), glycerol (GOH, in g/L), and total antibacterial activity (TAA, in AU/mL) in the unfermented UHT milk and fermented beverages from fermentation I (I−24 h, I−48 h, I−72 h, and I−96 h) and II (II−48 h, II−96 h, II−148 h and II−192 h). .
Figure 6.
Principal component analysis based on the differences between the pH values and contents in lactose (L, in g/L), total nitrogen (TN, in g/L), proteins (Pr, in g/L), lactic acid (LA, in g/L), acetic acid (AA, in g/L), ethanol (EtOH, in g/L), glycerol (GOH, in g/L), and total antibacterial activity (TAA, in AU/mL) in the unfermented UHT milk and fermented beverages from fermentation I (I−24 h, I−48 h, I−72 h, and I−96 h) and II (II−48 h, II−96 h, II−148 h and II−192 h). .
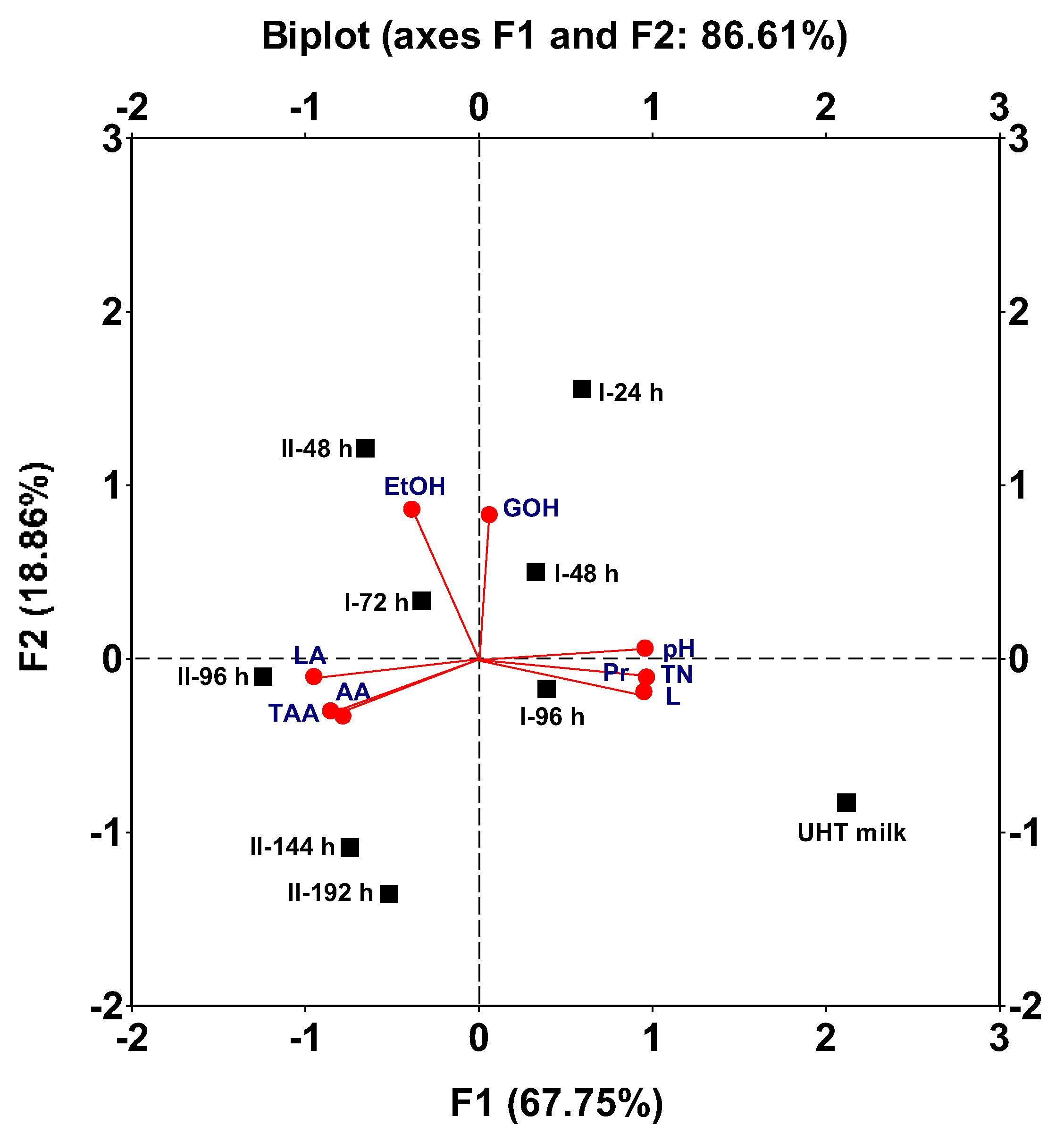
Figure 7.
Principal component analysis based on the differences between the total concentrations of volatile compounds (acids, alcohols, aldehydes, ketones, esters, hydrocarbons and other compounds) in the unfermented UHT milk and fermented beverages from fermentation I (I−24 h, I−48 h, I−72 h, and I−96 h) and II (II−48 h, II−96 h, II−148 h, and II−196 h).
Figure 7.
Principal component analysis based on the differences between the total concentrations of volatile compounds (acids, alcohols, aldehydes, ketones, esters, hydrocarbons and other compounds) in the unfermented UHT milk and fermented beverages from fermentation I (I−24 h, I−48 h, I−72 h, and I−96 h) and II (II−48 h, II−96 h, II−148 h, and II−196 h).
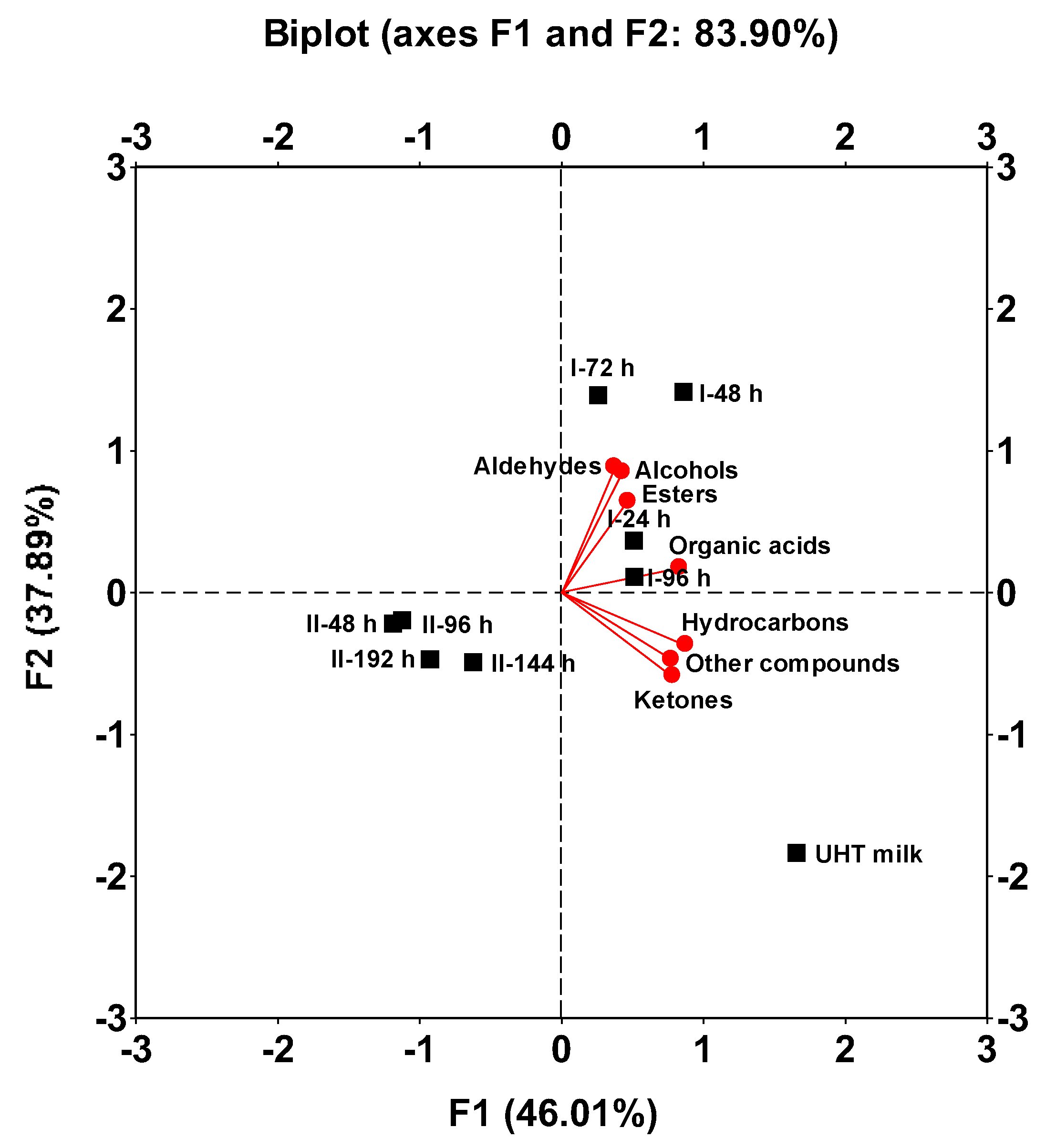
Figure 8.
Evolution of the lactic acid bacteria (AAB), acetic acid bacteria (AAB) and yeasts in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
Figure 8.
Evolution of the lactic acid bacteria (AAB), acetic acid bacteria (AAB) and yeasts in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
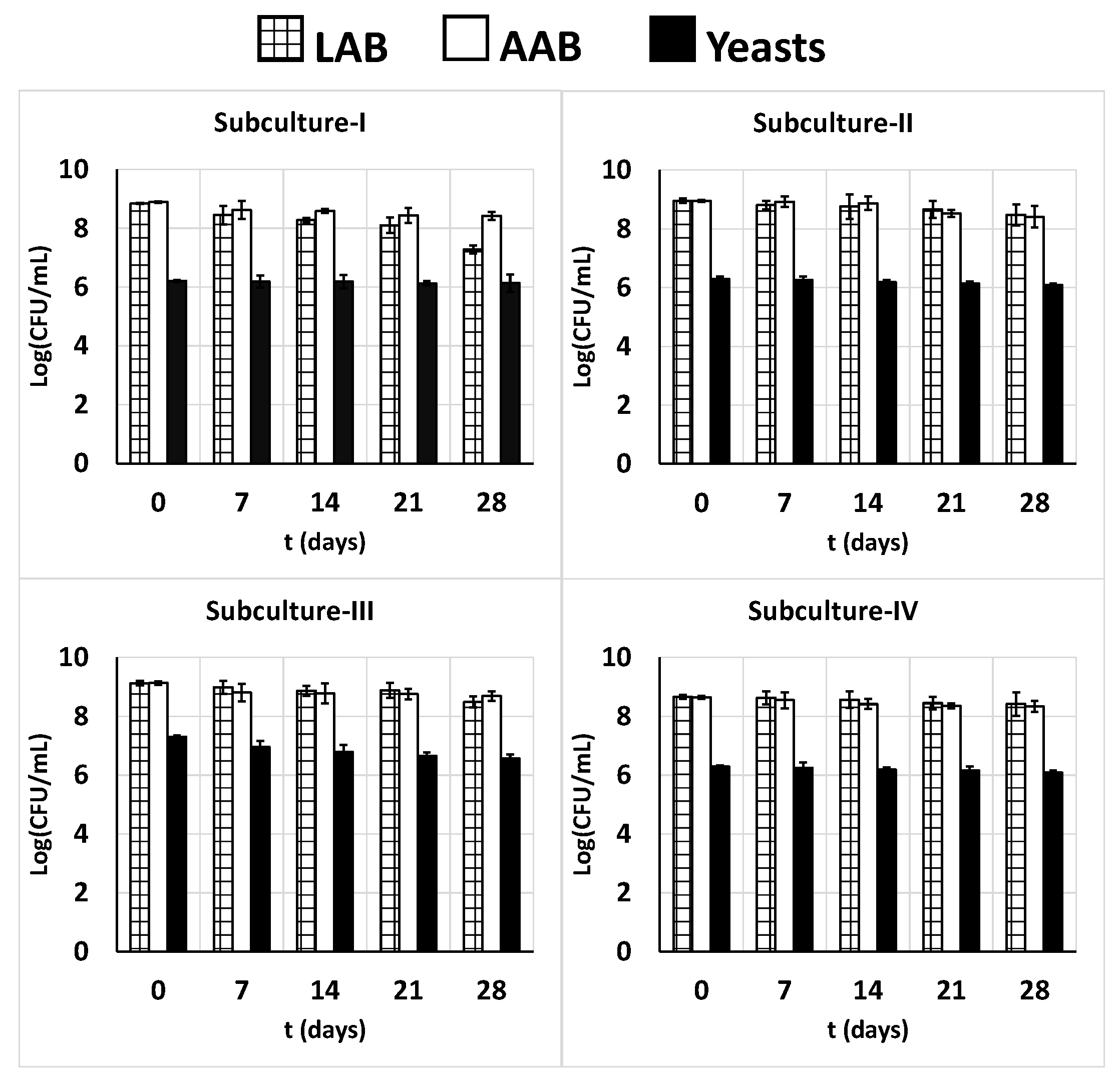
Figure 9.
Evolution of pH and nutrients (L: lactose and Pr: proteins) in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
Figure 9.
Evolution of pH and nutrients (L: lactose and Pr: proteins) in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
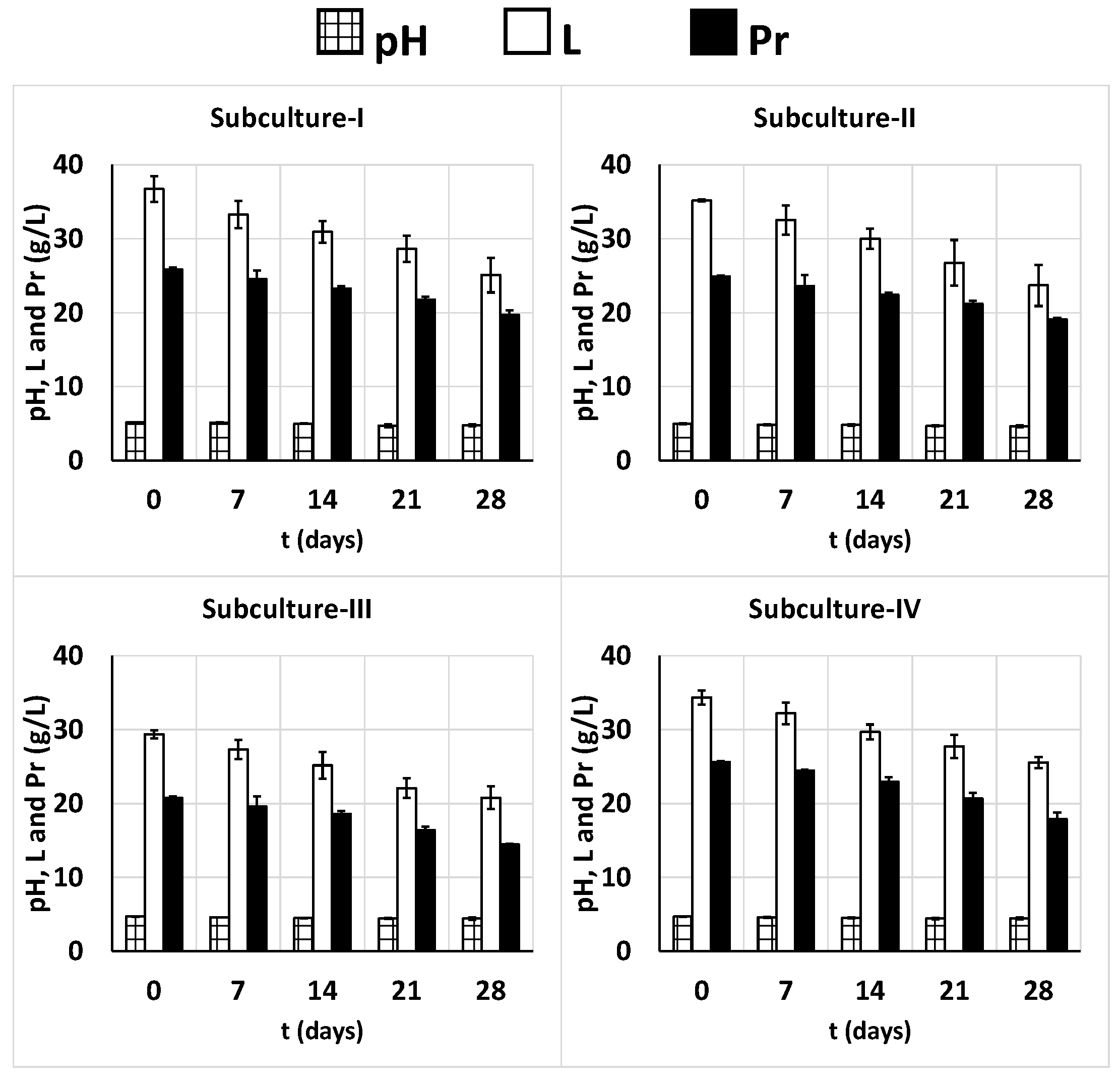
Figure 10.
Evolution of fermentation metabolites (LA: lactic acid, AA: acetic acid, EtOH: ethanol, and GOH: glycerol) in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
Figure 10.
Evolution of fermentation metabolites (LA: lactic acid, AA: acetic acid, EtOH: ethanol, and GOH: glycerol) in fermented beverages from the four subcultures of fermentation I during refrigerated storage at 4 °C.
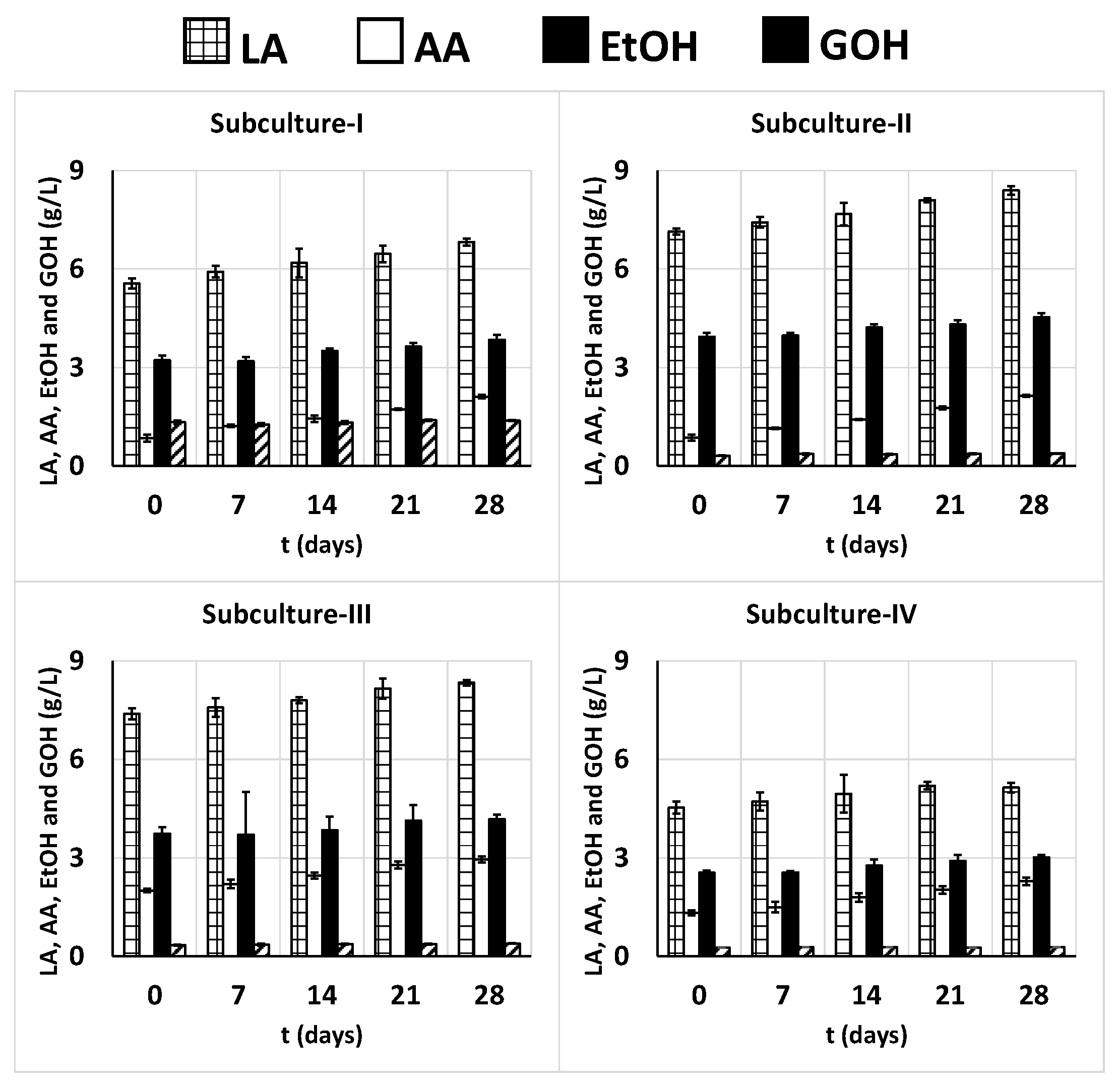

Table 1.
Concentration (µg/L) of Volatile Organic Compounds (Means ± Standard Deviations of Three Experiments and Two Analytical Replications Each) identified in the Unfermented (UWM) and Fermented UHT Whole Milk with Kefir Grains in the Four Consecutive Subcultures in Fermentations I and II.
Table 1.
Concentration (µg/L) of Volatile Organic Compounds (Means ± Standard Deviations of Three Experiments and Two Analytical Replications Each) identified in the Unfermented (UWM) and Fermented UHT Whole Milk with Kefir Grains in the Four Consecutive Subcultures in Fermentations I and II.
| Fermentation I | Fermentation II | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| No. | Compound | UWM | 0-24 h | 24-48 h | 48-72 h | 72-96 h | 0-48 h | 48-96 h | 96-144 h | 144-192 h |
| Organic Acids | ||||||||||
| 1 | Acetic acid | N.d. | N.d. | 1.90 ± 0.00 | 2.69 ± 0.83 | 2.35 ± 1.48 | 0.61 ± 0.06 | 3.07 ± 0.53 | 3.38 ± 0.23 | 2.30 ± 1.10 |
| 2 | Butanoic acid | N.d. | 0.68 ± 0.00 | N.d. | 0.80 ± 0.00 | N.d. | 0.21 ± 0.00 | 0.20 ± 0.00 | 0.34 ± 0.01 | 0.31 ± 0.08 |
| 3 | Pentanoic acid | N.d. | N.d. | N.d. | 0.76 ± 0.00 | 0.87 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 4 | Hexanoic acid | N.d. | 7.63 ± 5.98 | 23.45 ± 9.71 | 13.53 ± 0.62 | 11.08 ± 2.80 | 3.85 ± 0.17 | 4.44 ± 0.73 | 7.81 ± 0.07 | 7.82 ± 2.82 |
| 5 | 2-Ethyl hexanoic acid | N.d. | 0.52 ± 0.00 | 0.78 ± 0.45 | 1.01 ± 0.26 | 0.99 ± 0.96 | 0.30 ± 0.20 | 1.58 ± 0.03 | 2.61 ± 0.63 | 1.31 ± 0.37 |
| 6 | 2-Methyl-propanoic acid | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.64 ± 0.01 | 2.02 ± 0.19 | 1.32 ± 0.24 |
| 7 | Heptanoic acid | N.d. | N.d. | 0.92 ± 0.25 | 0.87 ± 0.00 | 0.78 ± 0.00 | 0.20 ± 0.10 | 0.23 ± 0.00 | 0.31 ± 0.01 | N.d. |
| 8 | Octanoic acid | N.d. | N.d. | N.d. | N.d. | 1.69 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 9 | Nonanoic acid | 4.89 ± 0.00 | N.d. | 0.72 ± 0.05 | 1.89 ± 0.00 | N.d. | N.d. | 0.41 ± 0.21 | 0.66 ± 0.03 | 0.26 ± 0.07 |
| 10 | Ethyl boronic acid | N.d. | N.d. | N.d. | 0.59 ± 0.14 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 11 | 8-Methyl-6-nonenoic acid | N.d. | N.d. | N.d. | 0.71 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 12 | Cyclopentane-undecanoic acid | 7.55 ± 0.19 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 13 | Cis-11-eicosenoic acid | 28.38 ± 1.43 | 8.02 ± 0.00 | 11.46 ± 1.96 | 14.39 ± 1.43 | 11.07 ± 6.88 | 2.97 ± 1.10 | 4.37 ± 0.56 | 4.42 ± 0.18 | 3.20 ± 0.83 |
| 14 | Trans-2-undecenoic acid | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 1.54 ± 0.00 | N.d. | N.d. |
| Number of Organic Acids | 3 | 4 | 6 | 10 | 7 | 6 | 9 | 8 | 7 | |
| Total concentration | 40.83 ± 0.01A | 16.85 ± 5.98B | 39.24 ± 11.27C,A | 37.25 ± 2.63D,C | 28.84 ± 10.86E,D | 8.14 ± 1.07F | 16.48 ± 1.21G,B,E | 21.55 ± 0.63B,E | 16.52 ± 2.53B,G | |
| Alcohols | ||||||||||
| 15 | 1-Pentanol | N.d. | 44.08 ± 1.22 | 40.86 ± 3.86 | 36.06 ± 11.08 | 9.78 ± 0.00 | 10.28 ± 5.49 | 4.16 ± 0.14 | 2.67 ± 0.00 | 1.68 ± 0.81 |
| 16 | 2-Phenylethanol | 8.86 ± 0.00 | 12.36 ± 5.54 | 23.09 ± 9.51 | 22.90 ± 3.73 | 11.41 ± 1.74 | 10.70 ± 4.84 | 9.06 ± 0.39 | 5.34 ± 1.00 | 3.20 ± 0.87 |
| 17 | 2-Methyl-1-propanol | N.d. | N.d. | N.d. | 0.90 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 18 | 1,3-Octanediol | N.d. | 0.88 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 19 | 1,5-Hexanediol | N.d. | 0.42 ± 0.00 | 1.59 ± 0.16 | N.d. | 0.65 ± 0.37 | 0.62 ± 0.26 | 0.57 ± 0.04 | 0.52 ± 0.17 | N.d. |
| 20 | 1-Heptanol | N.d. | N.d. | N.d. | 1.54 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 21 | 2,10-Dimethyl-9-undecen-1-ol | N.d. | N.d. | 5.83 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 22 | 2,4-Dimethyl-3-pentanol | N.d. | N.d. | N.d. | N.d. | 0.82 ± 0.00 | 0.16 ± 0.00 | N.d. | 0.27 ± 0.00 | N.d. |
| 23 | 2,7-Dimethyl-1-octanol | N.d. | N.d. | 9.60 ± 0.00 | 0.39 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 24 | 2-Butyl-1-octanol | N.d. | N.d. | N.d. | 3.47 ± 0.00 | 0.93 ± 0.00 | 0.56 ± 0.00 | N.d. | 0.60 ± 0.00 | 0.25 ± 0.00 |
| 25 | 2-Butyl-2,7-octadien-1-ol | N.d. | N.d. | 2.64 ± 0.19 | 3.07 ± 0.69 | N.d. | 0.15 ± 0.00 | 0.85 ± 0.00 | 0.68 ± 0.02 | 0.56 ± 0.15 |
| 26 | 2-Ethyl-hexanol | 6.27 ± 0.00 | 12.00 ± 0.59 | 19.23 ± 3.27 | 17.59 ± 0.83 | 8.03 ± 7.34 | 2.43 ± 0.90 | 3.20 ± 0.03 | 3.37 ± 0.18 | 3.05 ± 0.13 |
| 27 | 2-Heptanol | N.d. | 0.64 ± 0.00 | 0.80 ± 0.00 | N.d. | N.d. | 0.31 ± 0.19 | 0.18 ± 0.00 | 0.37 ± 0.00 | N.d. |
| 28 | 2-Methyl-5-(1- methylethenyl)-cyclohexanol | N.d. | N.d. | N.d. | 2.84 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 29 | 3,6-Dimethyl-3-heptanol | N.d. | N.d. | N.d. | 0.68 ± 0.12 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 30 | 3-Methyl-1,5-pentanediol | N.d. | 0.50 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 31 | 3-Methyl-1-pentanol | N.d. | 0.78 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 32 | 5,9-Dimethyl-1-decanol | N.d. | N.d. | 1.16 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 33 | 5-Nonadecen-1-ol | N.d. | 11.03 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 34 | Z-10-Pentadecen-1-ol | N.d. | N.d. | 4.08 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 35 | Butane-1,3-diol | N.d. | N.d. | N.d. | N.d. | N.d. | 0.20 ± 0.00 | N.d. | N.d. | N.d. |
| 36 | 1,3,5-Pentanotriol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.60 ± 0.00 |
| 37 | 2-Furanmethanol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.60 ± 0.00 |
| 38 | (E)-2-Tridecen-1-ol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 1.13 ± 0.76 | N.d. | N.d. |
| 39 | 1-Eicosanol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.21 ± 0.00 | N.d. | N.d. |
| 40 | 2-Hexyl-1-octanol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.33 ± 0.00 |
| 41 | 2-Nonen-1-ol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.48 ± 0.00 | N.d. | N.d. |
| 42 | 3-Tetradecyn-1-ol | N.d. | N.d. | N.d. | N.d. | N.d. | 0.21 ± 0.00 | 0.68 ± 0.00 | N.d. | N.d. |
| 43 | 4-Methyl-2-pentanol | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.15 ± 0.00 |
| Number of Alcohols | 2 | 9 | 10 | 10 | 6 | 10 | 10 | 8 | 9 | |
| Total concentration | 15.13 ± 0.00A | 82.71 ± 5.84B | 108.89 ± 16.56C | 89.47 ± 13.96B | 31.63 ± 8.45D | 25.62 ± 6.80E,D | 20.52 ± 0.46E | 13.82 ± 0.86F | 10.42 ± 0.86G | |
| Aldehydes | ||||||||||
| 44 | (E)-2-Nonenal | N.d. | N.d. | 0.84 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 45 | (Z)-14-methyl-8-hexadecenal | N.d. | N.d. | 1.66 ± 0.06 | 2.35 ± 0.49 | 2.61 ± 0.00 | N.d. | N.d. | 0.58 ± 0.01 | 0.67 ± 0.09 |
| 46 | 10-Undecenal | N.d. | 0.83 ± 0.13 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 47 | 2,6-Dimethyl benzaldehyde | N.d. | 0.36 ± 0.11 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 48 | 2-Ethyl hexanal | N.d. | N.d. | 1.21 ± 0.64 | 0.62 ± 0.25 | N.d. | 0.17 ± 0.00 | 0.19 ± 0.11 | 0.24 ± 0.09 | 0.15 ± 0.06 |
| 49 | 2-Methyl undecanal | N.d. | N.d. | 0.81 ± 0.06 | 0.94 ± 0.00 | N.d. | N.d. | 0.14 ± 0.00 | 0.17 ± 0.00 | N.d. |
| 50 | Benzaldehyde | N.d. | 2.26 ± 0.00 | 3.77 ± 0.91 | 2.60 ± 0.37 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 51 | Dodecanal | N.d. | N.d. | N.d. | 0.79 ± 0.00 | 0.51 ± 0.29 | N.d. | N.d. | N.d. | N.d. |
| 52 | Phenylacetaldehyde | N.d. | N.d. | 1.18 ± 0.20 | 0.94 ± 0.25 | N.d. | 0.15 ± 0.05 | 0.17 ± 0.02 | 0.19 ± 0.01 | 0.35 ± 0.15 |
| 53 | Nonanal | N.d. | N.d. | N.d. | 1.61 ± 0.81 | N.d. | N.d. | N.d. | N.d. | N.d. |
| Number of Aldehydes | 0 | 3 | 6 | 7 | 2 | 2 | 3 | 4 | 3 | |
| Total concentration | 0.00 ± 0.00A | 3.45 ± 0.03B | 9.47 ± 0.87C | 9.84 ± 1.40C | 3.12 ± 0.29D | 0.32 ± 0.05E | 0.50 ± 0.18F | 1.18 ± 0.10G | 1.17 ± 0.18G | |
| Ketones | ||||||||||
| 54 | 2,6-Dimethyl-4-heptanone | 2.18 ± 0.00 | 4.98 ± 0.00 | 2.23 ± 1.47 | 2.52 ± 0.00 | 3.19 ± 0.00 | 0.06 ± 0.00 | 0.18 ± 0.00 | 0.42 ± 0.27 | 0.36 ± 0.29 |
| 55 | 2-Heptanone | 28.96 ± 0.00 | 5.52 ± 0.62 | 4.05 ± 0.00 | N.d. | 2.39 ± 0.00 | 0.40 ± 0.24 | 0.59 ± 0.00 | 2.14 ± 0.12 | 0.88 ± 0.70 |
| 56 | 2-Methyl-4-octanone | N.d. | 1.06 ± 0.00 | N.d. | 1.30 ± 0.00 | 1.92 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 57 | 2-Nonanone | 14.26 ± 0.00 | 0.40 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 58 | 8-Hydroxy-2-octanone | N.d. | N.d. | N.d. | N.d. | 2.06 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 59 | 5-Methyl-3-heptanone | N.d. | N.d. | N.d. | N.d. | N.d. | 0.16 ± 0.00 | N.d. | N.d. | N.d. |
| Number of Ketones | 3 | 4 | 2 | 2 | 4 | 3 | 2 | 2 | 2 | |
| Total concentration | 45.40 ± 0.00A | 11.95 ± 0.62B | 6.28 ± 1.47C | 3.82 ± 0.01D | 9.55 ± 0.01E | 0.62 ± 0.24F | 0.77 ± 0.00G,F | 2.56 ± 0.33H | 1.24 ± 0.76F,G | |
| Esters | ||||||||||
| 60 | 2-Phenylethyl acetate | 1.19 ± 0.00 | 3.27 ± 0.98 | 4.31 ± 2.00 | 4.38 ± 0.92 | 2.70 ± 2.21 | 2.90 ± 1.75 | 1.45 ± 0.40 | 0.75 ± 0.11 | 0.37 ± 0.03 |
| 61 | 2-Methylpentyl acetate | N.d. | N.d. | 0.80 ± 0.20 | N.d. | N.d. | N.d. | N.d. | N.d. | 0.32 ± 0.13 |
| 62 | Ethyl acetate | N.d. | 8.07 ± 0.00 | 5.95 ± 0.94 | 6.19 ± 0.94 | 4.11 ± 0.00 | 2.11 ± 1.11 | 4.62 ± 2.09 | 2.82 ± 0.25 | 1.72 ± 0.53 |
| 63 | Isoamyl acetate | N.d. | 2.58 ± 0.72 | N.d. | 2.02 ± 1.50 | 3.79 ± 2.50 | 0.65 ± 0.00 | 0.81 ± 0.26 | 0.39 ± 0.00 | N.d. |
| 64 | Pentyl acetate | N.d. | N.d. | 2.60 ± 2.11 | N.d. | N.d. | 0.32 ± 0.00 | 2.10 ± 0.00 | 0.21 ± 0.00 | 0.29 ± 0.00 |
| 65 | β-Terpinyl acetate | N.d. | N.d. | 0.98 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 66 | 1-Methylundecyl acrylate | N.d. | 2.12 ± 0.00 | 0.79 ± 0.00 | 1.58 ± 0.32 | N.d. | N.d. | 0.11 ± 0.00 | N.d. | N.d. |
| 67 | 3-Methylbutyl chloroacetate | N.d. | N.d. | N.d. | N.d. | 0.44 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 68 | Ethyl 5-methyl nonanoate | N.d. | N.d. | N.d. | N.d. | 0.72 ± 0.00 | N.d. | 0.51 ± 0.00 | N.d. | N.d. |
| 69 | 3-Tridecyl methoxyacetate | N.d. | 1.81 ± 0.00 | N.d. | 1.26 ± 0.00 | N.d. | N.d. | 0.34 ± 0.07 | N.d. | N.d. |
| 70 | 9-Dodecenyl methoxyacetate | N.d. | N.d. | N.d. | N.d. | 0.33 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 71 | Allyl pentadecyl oxalate | 2.67 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 72 | Pentyl isohexyl oxalate | N.d. | 0.35 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 73 | Benzyl oleate | N.d. | 1.24 ± 0.00 | N.d. | N.d. | 1.50 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 74 | Decyl butanoate | N.d. | 6.16 ± 4.68 | 4.93 ± 1.50 | 2.69 ± 0.00 | 3.53 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 75 | Dimethyl ethylboronate | N.d. | N.d. | N.d. | 1.75 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 76 | Ethyl phenylacetate | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.19 ± 0.00 | 0.14 ± 0.03 | N.d. |
| 77 | Ethyl hexanoate | N.d. | 36.47 ± 0.61 | 34.07 ± 1.86 | 19.74 ± 15.87 | 64.05 ± 0.00 | 8.10 ± 3.25 | 2.36 ± 1.75 | 2.52 ± 0.54 | 0.18 ± 0.00 |
| 78 | Ethyl heptanoate | N.d. | 1.43 ± 0.00 | 1.38 ± 0.28 | 0.56 ± 0.08 | 0.31 ± 0.00 | 0.28 ± 0.07 | 0.25 ± 0.00 | N.d. | N.d. |
| 79 | Ethyl octanoate | 3.25 ± 0.00 | 30.70 ± 17.87 | 30.12 ± 2.35 | 12.80 ± 1.49 | 18.92 ± 6.47 | 11.15 ± 3.32 | 2.33 ± 1.46 | 1.92 ± 0.03 | 0.63 ± 0.21 |
| 80 | Ethyl nonanoate | N.d | N.d | N.d | N.d | N.d | 0.12 ± 0.00 | N.d | N.d | N.d |
| 81 | Ethyl decanoate | 2.18 ± 0.01 | 5.53 ± 3.64 | 5.14 ± 1.35 | 2.26 ± 0.01 | 3.63 ± 0.25 | 1.51 ± 0.43 | 0.99 ± 0.00 | 0.61 ± 0.07 | 0.40 ± 0.00 |
| 82 | Ethyl dodecanoate | N.d. | 0.75 ± 0.00 | N.d. | N.d. | N.d. | 0.28 ± 0.10 | N.d. | N.d. | N.d. |
| 83 | Ethyl isobutanoate | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.26 ± 0.00 | N.d. | N.d. |
| 84 | Isobutyl 2-methylaminobenzoate | N.d. | 1.43 ± 0.00 | N.d. | N.d. | 0.91 ± 0.00 | 0.27 ± 0.00 | 0.35 ± 0.00 | N.d. | N.d. |
| 85 | Methyl hippurate | 3.67 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 86 | Methyl L-valinate | N.d. | N.d. | N.d. | 2.18 ± 0.00 | 1.75 ± 1.25 | N.d. | 1.97 ± 0.69 | 3.10 ± 1.41 | 2.42 ± 1.47 |
| 87 | Pentyl pentanoate | N.d. | N.d. | N.d. | N.d. | 0.75 ± 0.00 | 0.17 ± 0.05 | N.d. | N.d. | N.d. |
| 88 | Octyl propanoate | N.d. | N.d. | N.d. | 1.65 ± 0.32 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 89 | Octyl pentanoate | N.d. | N.d. | N.d. | N.d. | N.d. | 0.15 ± 0.09 | N.d. | N.d. | N.d. |
| 90 | Octyl butanoate | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.45 ± 0.00 | 0.35 ± 0.00 | N.d. |
| 91 | Hexyl propanoate | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.28 ± 0.00 |
| 92 | 2-Hidroxy-4-methyl 4-methylpentanoate |
N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 0.76 ± 0.00 | N.d. |
| Number of Esters | 5 | 14 | 11 | 13 | 15 | 13 | 16 | 11 | 9 | |
| Total concentration | 12.96 ± 0.01A | 101.91 ± 19.08B | 91.07 ± 4.22B | 59.06 ± 18.48C | 107.45 ± 8.31B | 28.01 ± 3.02D | 19.09 ± 3.61E | 13.57 ± 1.15A | 6.61 ± 1.16F | |
| Hydrocarbons | ||||||||||
| 93 | (Z)-3-Dodecene | N.d. | N.d. | 1.41 ± 0.00 | 0.63 ± 0.00 | 0.86 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 94 | 1,2,4-Trimethylbenzene | N.d. | 23.78 ± 0.00 | N.d. | 7.49 ± 0.00 | 21.30 ± 0.00 | N.d. | N.d. | 5.51 ± 0.00 | 2.56 ± 2.02 |
| 95 | 1,3-Bis(1,1-dimethylethyl)benzene | N.d. | N.d. | 3.64 ± 0.80 | 2.70 ± 0.39 | N.d. | N.d. | N.d. | N.d. | N.d. |
| 96 | 3-Hexadecyne | 27.29 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 1.31 ± 0.00 | N.d. |
| 97 | 5-Butyl-4-nonene | 4.45 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 98 | 9-Eicosyne | N.d. | N.d. | N.d. | N.d. | 7.95 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 99 | Decane | 13.37 ± 0.00 | 5.58 ± 4.70 | 3.75 ± 1.90 | 3.72 ± 0.51 | 3.63 ± 0.00 | 0.31 ± 0.11 | 0.50 ± 0.00 | N.d. | N.d. |
| 100 | Dodecane | N.d. | N.d. | N.d. | N.d. | 6.72 ± 0.00 | N.d. | N.d. | N.d. | N.d. |
| 101 | n-Hexane | 11.37 ± 0.00 | 2.70 ± 0.00 | 3.15 ± 2.36 | 2.29 ± 0.30 | 2.05 ± 0.39 | N.d. | N.d. | N.d. | 3.48 ± 2.03 |
| 102 | Tetradecane | 12.47 ± 0.00 | 2.45 ± 0.00 | 4.02 ± 1.02 | 2.44 ± 0.00 | 2.99 ± 1.54 | 0.52 ± 0.13 | 0.83 ± 0.48 | N.d. | 0.71 ± 0.00 |
| Number of Hydrocarbons | 5 | 4 | 5 | 6 | 7 | 2 | 2 | 2 | 3 | |
| Total concentration | 68.95 ± 0.01A | 34.51 ± 4.70B | 15.97 ± 6.01C | 19.27 ± 1.19C | 45.50 ± 1.78D | 0.83 ± 0.10E | 1.33 ± 0.48F | 6.82 ± 0.00G | 6.75 ± 2.31G | |
| Other Compounds | ||||||||||
| 103 | Octyl oxirane | 4.54 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 104 | o-Decyl hydroxylamine | N.d. | 1.04 ± 0.26 | 1.42 ± 0.46 | N.d. | 1.64 ± 0.33 | 0.37 ± 0.08 | N.d. | 0.41 ± 0.03 | N.d. |
| 105 | 3,3,4,4-Tetramethylbutyrolactone | N.d. | 1.68 ± 0.00 | 0.94 ± 0.00 | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. |
| 106 | 2,4,5-Trimethyl-1,3-dioxolane | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | N.d. | 1.52 ± 0.18 | 1.11 ± 0.20 |
| 107 | 3-Trifluoroacetoxy dodecane | N.d. | N.d. | N.d. | N.d. | N.d. | 0.13 ± 0.00 | N.d. | N.d. | N.d. |
| Number of Other Compounds | 1 | 2 | 2 | 0 | 1 | 2 | 0 | 2 | 1 | |
| Total concentration | 4.54 ± 0.00A | 2.72 ± 0.26B | 2.36 ± 0.46C,B | N.d. | 1.64 ± 0.33D | 0.50 ± 0.08E | N.d. | 1.93 ± 0.18C,D | 1.11 ± 0.20F | |
| Total number of volatile compounds | 19 | 40 | 42 | 48 | 42 | 38 | 42 | 37 | 34 | |
| Total concentration of volatile compounds | 187.80 ± 0.01A | 254.11 ± 13.60B | 273.28 ± 29.50C,B | 218.71 ± 24.10D | 227.73 ± 14.73B,D | 64.04 ± 6.63E | 58.69 ± 3.45F,E | 61.43 ± 1.34G,E | 43.82 ± 4.66H | |
* N.d.: not detected; A-I: Mean values in columns with different letter are significantly different at P < 0.05.
Table 2.
Olfactory perception threshold (OPT, µg/L), odor descriptors and aroma series for the volatile compounds identified in unfermented and fermented UHT milk with kefir grains in the four consecutive 24- or 48-h subcultures.
Table 2.
Olfactory perception threshold (OPT, µg/L), odor descriptors and aroma series for the volatile compounds identified in unfermented and fermented UHT milk with kefir grains in the four consecutive 24- or 48-h subcultures.
| Volatile compound | OPT (µg/L) | Odor descriptors | Series |
|---|---|---|---|
| Organic acids | |||
| Acetic acid | 99000.00 [42] | Vinegar, sour [43,44] | 1 |
| Butanoic acid | 1400.00 [42] | Sweaty, butter, cheese, acid, fermented odor [45] | 1 |
| Pentanoic acid | 3000.00 [42] | Green fruit, vegetables, sweat [14,46] | 1, 4 |
| Hexanoic acid | 3000.00 [42] | Goat cheese, fatty acids, vegetable oil, sweaty, sharp, acidic, green, gammy, dairy sour, dairy, stale, butter, sour, fruity and pungent [43,45,47,48,49] | 1, 2, 4 |
| 2-Ethyl-hexanoic acid | 27000.00 [42] | Taste similar to paint [50] | 5 |
| 2-Methyl-propanoic acid | 6550.50 [42] | Fuits [14] | 2 |
| Heptanoic acid | 640.00 [42] | Cheese [47] | 1 |
| Octanoic acid | 3000.00 [42] | Sweat, cheese, rancid, fatty acids, vegetable oil [47,51] | 1 |
| Nonanoic acid | 8800.00 [42] | Fatty, soapy, waxy, green, goat [52] | 1, 4 |
| Alcohols | |||
| 1-Pentanol | 120.00 [42] | Green fruit, fermented, bread and yeast [47,53,54] | 1, 2 |
| 2-Phenylethanol | 564.20 [42] | Floral, pink, flowery, like honey [47,51,55,56] | 3, 7 |
| 2-Methyl-1-propanol | 550.00 [42] | Wine, solvent, bitter, malty [57,58,59] | 5, 6 |
| 1-Heptanol | 5.40 [42] | Fat [60] | 1 |
| 2-Heptanol | 65.23 [42] | Green [14,47] | 4 |
| 2,4-Dimethyl-3-pentanol | 82.00 [42] | Bitter almond [61] | 2 |
| 2-Ethyl-hexanol | 830.00 [42] | Leather, goat, stable, cow and sheep, cardboard [45,52] | 5, 6 |
| 3-Methyl-1-pentanol | 7.50 [42] | Unpleasant chemical fusel [62] | 5 |
| 2-Furanmethanol | 4500.00 [42] | Floral, sweet [63] |
6 |
| Butane-1,3-diol | 10000.00 [42] | Greasy, floral [64] | 1, 3 |
| Aldehydes | |||
| (E)-2-Nonenal | 0.40 [42] | Fatty, green [59] | 1, 4 |
| 2-Ethyl hexanal | 41.00 [42] | Legume smell [65] | 1 |
| Benzaldehyde | 750.00 [42] | Almond, nut like bitter [47,65] | 2 |
| Dodecanal | 10.00 [42] | Floral, waxy [66] | 1, 3 |
| Phenylacetaldehyde | 6.30 [42] | Honey [47] | 7 |
| Nonanal | 1.10 [42] | Green, fresh, citrus-like, soapy, fruity odor [47,51,67] | 2, 4 |
| Ketones | |||
| 2,6-Dimethyl-4-heptanone | 110.00 [68] | Fruity, sweet [49] | 2 |
| 2-Heptanone | 5.00 [69] | Banana, fruity, floral and musty, fresh cream flavor [47,53,67] | 1, 2, 3 |
| 2-Nonanone | 5.00 [69] | Sweet, fruity, floral, musty [47,67] | 2, 3 |
| 5-Methyl-3-heptanone | 41.00 [42] | Fruit [70] | 2 |
| Esters | |||
| 2-Phenethyl acetate | 249.59 [42] | Floral, rose, honey [71] | 3, 7 |
| Ethyl acetate | 5000.00 [71] | Pineapple, fruit [15,47,72] | 2 |
| Isoamyl acetate | 2.00 [42] | Banana, fruity, sweet [47,73] | 2 |
| Pentyl acetate | 43.00 [42] | Fruit [71] | 2 |
| Decyl butanoate | 250.00 [74] | Fruity, waxy, fatty and slightly citrus [75] | 1, 2 |
| Ethyl decanoate | 23.00 [42] | Apple, floral, fruity and musty [15,71,76] | 2, 3 |
| Ethyl dodecanoate | 5900.00 [42] | Fruity, floral [71] | 2, 3 |
| Ethyl heptanoate | 1.90 [42] | Fruit, overripe [47,71] | 2 |
| Ethyl hexanoate | 1.00 [42] | Banana, apple [71,77] | 2 |
| Ethyl octanoate | 19.30 [42] | Fruity, apple, banana [71,77] | 2 |
| Ethyl phenylacetate | 250.00 [75] | Sweet fragrance of honey [47] | 7 |
| Ethyl nonanoate | 377.00 [42] | Floral, fruit [66,71] | 2 |
| Ethyl isobutanoate | 0.10 [42] | Unripe fruit [78] | 2, 4 |
| Octyl butanoate | 250.00 [42] | Herb, orange, flower [79] | 2, 3, 4 |
| Hexyl propanoate | 8.00 [42] | Apple [80] | 2 |
| Hydrocarbons | |||
| 1,2,4-Trimethyl-benzene | 260.00 [42] | Fruity, citrus, sweet [81] | 2 |
| Decane | 10000.00 [42] | Solvent, sweet-etheric, diffusive [82] | 5 |
| Dodecane | 10000.00 [42] | Alkanes [53] | 5 |
| Tetradecane | 1000.00 [42] | Gasoil [83] | 5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Copyright: This open access article is published under a Creative Commons CC BY 4.0 license, which permit the free download, distribution, and reuse, provided that the author and preprint are cited in any reuse.



